Embed Size (px)
Citation preview

Peptoid-containing collagen mimetics with cellbinding activity
Graham Johnson,1,2 Michelle Jenkins,1,2 Keith M. McLean,1,3 Hans J. Griesser,1,3 Juliann Kwak,4
Murray Goodman,4 John G. Steele1,2
1Cooperative Research Centre for Eye Research and Technology, P.O. Box 184, North Ryde, New South Wale 16702CSIRO Molecular Science, P.O. Box 184, North Ryde, New South Wales 16703CSIRO Molecular Science, Private Bag 10, Clayton, Victoria 3169, Australia4Department of Chemistry and Biochemistry, University of California at San Diego, La Jolla, California 92093-0343
Received 7 April 1999; revised 21 December 1999; accepted 27 January 2000
Abstract: Collagen mimetic peptides containing the pep-toid residue Nleu (Goodman Bhumralkar, Jefferson, Kwak,Locardi. Biopolymers 1998;47:127–142) were tested for inter-actions with epithelial cells and fibroblasts. Molecules con-taining the sequence Gly-Pro-Nleu with a minimum of ninerepeats showed cell binding activity. The activity of thesemolecules appeared to be conformationally sensitive, withthe triple-helical form being preferred. When immobilizedon a surface, the (Gly-Pro-Nleu)10-Gly-Pro-NH2 sequencestimulated the attachment and growth of corneal epithelialcells and fibroblasts and the migration of epithelial tissue.The peptide sequence KDGEA inhibited cell attachment to
the (Gly-Pro-Nleu)10-Gly-Pro-NH2 sequence, suggestingthat cell binding to this collagen mimetic involves the a2b1heterodimer integrin receptor. Interestingly, peptides con-taining the sequence (GlyNleu-Pro-)10-NH2 did not have cellbinding activity. The discovery that triple-helical peptidescontaining the Gly-Pro-Nleu sequences interact with cellsopens up new opportunities in the design of collagen mi-metic biomaterials.© 2000 John Wiley & Sons, Inc. J Biomed Mater Res, 51,612–624, 2000.Key words: collagen mimetics; peptoids; cell binding pep-tides; corneal epithelial cells; a2b1 integrin receptor
INTRODUCTION
Collagen and collagen-based materials have exten-sive applications in biomedical devices and tissue en-gineering.1 They are used, for example, in the fabrica-tion of cardiac and aortal valves2,3 and vascular grafts4
and have been proposed for use in ophthalmic im-plants (see Thompson, 1991, for review5,6 abdominalrepair fabrics,7 and ligaments.8 Chemical stabilizationof collagen is often used to reduce the potential ofimmune and inflammatory reactions and to stabilize itagainst proteolytic degradation. Such collagen andcollagen-based materials seem to have an inherent bio-compatibility that is difficult to mimic in syntheticpolymers. However, the biological origin of collagens,with the consequential risk of disease transmission,can complicate their manufacture and can be an issue
during the regulatory approval process. Furthermore,there can be ethical considerations for their use inwhich the collagen is derived from human tissue. Thedesign of mimics of collagen may facilitate the devel-opment of wholely synthetic “collagen-based” bioma-terials, which have the biocompatibility of native col-lagens while having the manufacturing and regula-tory characteristics of a synthetic polymer.
For implant applications in which tissue integrationis desired, novel biomaterials have been designedfrom the combination of synthetic polymers and ex-tracellular matrix components and/or defined mo-lecular fragments of these molecules (see Hubbell,1995, for review.9,10 Previously, cell attachment motifs,such as the Arg-Gly-Asp (RGD) sequence, have beenincorporated within the polymer itself or applied tothe surface of synthetic polymers as a means of elicit-ing tissue attachment or migration.11 Collagen-basedpeptides and intact collagen molecules have also beenused as surface coatings in such applications as bonerepair matrices12 and corneal implants.13,14 This reportdescribes an investigation of the potential of a series ofsynthetic collagen-mimetic structures for use at bio-material surfaces where epithelial tissue adhesion andmigration are required.
The synthetic collagen mimetics examined in this
Correspondence to: G. Johnson; email: [email protected]
The benefits accruing to the author or authors from a com-mercial or industrial party will be applied to a research fund,nonprofit institution or other organization with which theauthor(s) are associated.
Contract grant sponsors: CSIRO, the Cooperative Re-search Centre for Eye Research and Technology
Contract grant sponsor: CIBA VISION Corporation, At-lanta, GA
© 2000 John Wiley & Sons, Inc.
study were primarily composed of repeat sequences ofglycine (Gly), proline (Pro), and N-isobutylglycine(Nleu), an N-substituted glycine or peptoid residue.15
The propensity of such novel structures to form stabletriple-helices analogous to the collagen structure hasrecently been described.16–20 In this study, the interac-tions of these synthetic mimetics with cells was exam-ined, not only to test for any potential cytotoxicity ofthese molecules but also to determine whether theysupport corneal epithelial cell adhesion, proliferation,and tissue migration. In screening for cell attachmentactivity in these collagen-mimetic structures, the strat-egy used was similar to that used previously in thecase of RGDS.21–24 Molecules in free solution that werefound to inhibit cell attachment were then immobi-lized onto a surface for evaluation of their ability todirectly support cell attachment and growth. The in-teraction of these collagen-mimetic molecules withcorneal epithelial tissue was tested in parallel, as partof our objective of designing surfaces that support thecoverage of corneal implants with a functioning epi-thelium. Although not the subject of this study, it isrelevant to note that the presence of the peptoid resi-dues may mean that these collagen-like structureswould have enhanced resistance to peptidase activ-ity.15
MATERIALS AND METHODS
Peptide/peptoid synthesis
Single chain collagen analogues, including analogues as-sembled on a Kemp triacid (KTA) (1,3,5-trimethyl cyclohex-
ane-1,3,5-tricarboxylic acid) template, were synthesized andcharacterized as described previously.18 Most of the collagenanalogues contained the peptoid residue N-isobutylglycine,Nleu (Nl). Terminally acetylated collagen-mimetic struc-tures were used in some experiments in which free solutionswere required. A summary of the compounds used with theabbreviations for those used in this report is shown in TableI. Commercially available (Gly-Pro-Nleu)10-Gly-Pro-NH2
[(GPNl)10GP-NH2] and (Gly-Nleu-Pro)10-NH2 [(GNlP)10-NH2] were obtained from BioResearch, San Diego, CA. Thepeptides Tyr-Ile-Gly-Ser-Arg (YIGSR), Arg-Gly-Glu-Ser(RGES), Lys-Asp-Gly-Glu-Ala (KDGEA), and Lys-Asp-Gly-Ala-Ala (KDGAA) were purchased from Auspep Pty. Ltd.,Melbourne, Australia, and the Gly-Arg-Gly-As-Ser-Pro-Lys(GRGDSPK) peptide was obtained from American PeptideCompany, Sunnyvale, CA. Collagen type I (Vit, Vitrogen100) was purchased from Collagen Biomaterials, Palo Alto,CA, and bovine plasma fibronectin from Sigma-Aldrich,Castle Hill, NSW, Australia.
Melting studies for (GPNl)10GP-NH2collagen-mimetic sequences
The peptides were synthesized on solid phase, cleavedfrom the resin, and purified by RP-HPLC. The pure peptidewas lyophilized and dried with P2O5, over refluxing ac-etone. Before the melting studies, the collagen mimeticswere stored in a 10 mg/mL solution of H2O or ethyleneglycol (EG)/H2O (v/v, 2:1) for 7 days at 4°C, diluted to 0.2mg/mL, and equilibrated for 24 h at 4°C. Optical rotationswere measured by a Perkin-Elmer 241 Polarimeter equippedwith a model 900 isotemp circulator (Fisher Scientific) anddata were collected at 365 nm (Hg).
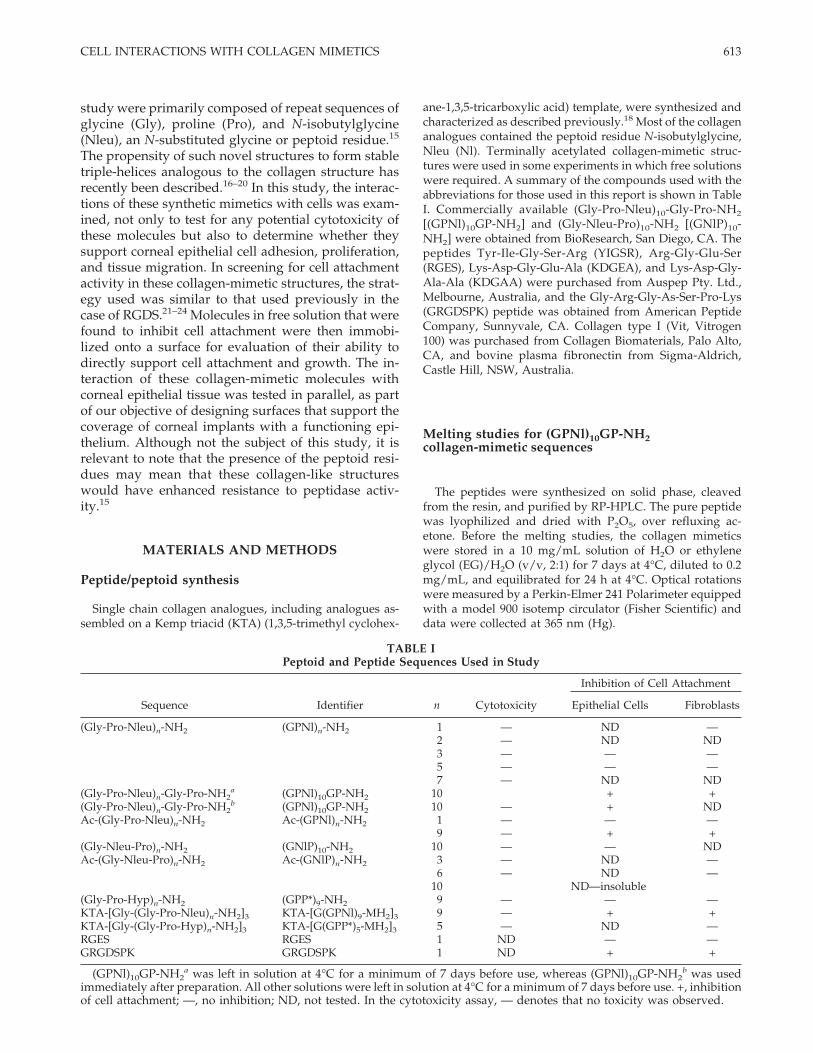
TABLE IPeptoid and Peptide Sequences Used in Study
Sequence Identifier n Cytotoxicity
Inhibition of Cell Attachment
Epithelial Cells Fibroblasts
(Gly-Pro-Nleu)n-NH2 (GPNl)n-NH2 1 — ND —2 — ND ND3 — — —5 — — —7 — ND ND
(Gly-Pro-Nleu)n-Gly-Pro-NH2a (GPNl)10GP-NH2 10 + +
(Gly-Pro-Nleu)n-Gly-Pro-NH2b (GPNl)10GP-NH2 10 — + ND
Ac-(Gly-Pro-Nleu)n-NH2 Ac-(GPNl)n-NH2 1 — — —9 — + +
(Gly-Nleu-Pro)n-NH2 (GNlP)10-NH2 10 — — NDAc-(Gly-Nleu-Pro)n-NH2 Ac-(GNlP)n-NH2 3 — ND —
6 — ND —10 ND—insoluble
(Gly-Pro-Hyp)n-NH2 (GPP*)9-NH2 9 — — —KTA-[Gly-(Gly-Pro-Nleu)n-NH2]3 KTA-[G(GPNl)9-MH2]3 9 — + +KTA-[Gly-(Gly-Pro-Hyp)n-NH2]3 KTA-[G(GPP*)5-MH2]3 5 — ND —RGES RGES 1 ND — —GRGDSPK GRGDSPK 1 ND + +
(GPNl)10GP-NH2a was left in solution at 4°C for a minimum of 7 days before use, whereas (GPNl)10GP-NH2
b was usedimmediately after preparation. All other solutions were left in solution at 4°C for a minimum of 7 days before use. +, inhibitionof cell attachment; —, no inhibition; ND, not tested. In the cytotoxicity assay, — denotes that no toxicity was observed.
613CELL INTERACTIONS WITH COLLAGEN MIMETICS

Preparation of solutions of collagenmimetic molecules
For those experiments in which the peptoid-containingmolecules were to be used in solution, samples were madeup as either a 2 mg/mL, or a 4 mg/mL stock in sterilebuffered Dulbecco’s Modified Eagle’s/Ham’s F12, pH 7.2[SFM, 50:50 (w/w), ICN Biomedicals, Sydney, Australia].These solutions were stored at 4°C for at least 1 week beforeuse, to allow those collagen-mimetic solutions capable offorming a triple-helical structure to reach equilibrium.19
Freshly made solutions were used in all other instances.
Immobilization of peptides/peptoids
The strategies used for the immobilization of peptoid-containing collagen mimetics are described in detail else-where, and involve coupling onto radiofrequency-plasmamodified fluorinated ethylene propylene copolymer.25
Briefly, fluorinated ethylene propylene copolymer film wasfunctionalized by heptylamine plasma polymerization togive an amine rich surface. The amines were then used tocovalently attach a custom-synthesized carboxymethyldex-tran spacer molecule (molecular weight, approximately70,000; the degree of carboxylization of which was deter-mined to be 1 carboxy group per 2 sugar units), onto whichwas then coupled the molecule of interest.25 For the colla-gen-mimetic molecules, samples were incubated at 4°C forat least 48 h to allow time for the formation of self-assembledtriple-helix structures on the surface, after which sampleswere washed in Milli-Q–purified water. Collagens, glyco-proteins, and control peptides were immobilized onto fluo-rinated ethylene propylene copolymer film by the samemethods as used for the peptoid-containing collagen ana-logues. X-ray photoelectron spectroscopy and Surface-MALDI-MS26 were used to confirm the presence of the coat-ings.
Samples of fluorinated ethylene propylene copolymerfilm containing immobilized molecules were cut intosquares of either 1 cm × 1 cm, for use in the cell attachmentand growth assays, or 2 cm × 2 cm for tissue outgrowthassays. Each sample was soaked overnight at 4°C in a solu-tion of phosphate buffered saline (pH 7.2) containing 120mg/mL penicillin and 200 mg/mL streptomycin (both fromCSL, Melbourne, Australia) before use.
Cell culture
Corneal epithelial cells and stromal fibroblasts were iso-lated from bovine corneas as described previously27,28 andused for the various assays between passages 1–2. Cellswere maintained in a culture medium composed of Dulbec-co’s Modified Eagle’s/Ham’s F12 [DMEM/F12, 50:50 (v/v);ICN Biomedicals] supplemented with fetal bovine serum(PA Biologicals, Sydney, Australia) at 20% (v/v) for the cor-neal epithelial cells and 10% (v/v) for the corneal fibroblasts.
All culture media contained 60 mg/mL penicillin and 100mg/mL streptomycin. All cells were incubated at 37°C in ahumidified atmosphere of 5% CO2 in air. Culture mediawere replenished every second day. In experiments in whichthe culture medium did not contain serum, 5 mg/mL insulin,5 mg/mL transferrin, and 5 ng/mL selenious acid (ITS™Premix; Collaborative Biomedical Products, Bedford, MA)were added to the medium.
Cytotoxicity assay
Corneal epithelial cells were seeded onto 96-well tissueculture polystyrene (TCPS) culture trays (Nunclon™ DeltaTC Microwell plates, cat. no. 167008, Nunc, Denmark) at 1 ×104 cells/well in DMEM/F12 culture medium supple-mented with 20% (v/v) fetal bovine serum and incubatedfor 24 h. This medium was then replaced with 100 mL ali-quots of solution of collagen-mimetic molecules, serially di-luted in serum-free DMEM/F12 in a range from 1 mg/mLdown to zero. Each dilution point was assayed in triplicate.The cells were incubated for a further 24 h and final numbersof attached cells measured by a tetrazolium dye-based col-orimetric assay (MTT, Sigma Chemical Co., Sydney, Austra-lia) read at 595 nm on a plate reader. The mean (±SD) num-ber of cells found in the wells was expressed as a percentageof that detected in the wells in which no collagen-mimeticmolecule was present.
Short-term assay for inhibition of cell attachment
Two types of attachment inhibition assays were per-formed:
1. Identification of peptoid-containing sequences capable of in-hibiting the initial attachment of cells to TCPS. Cornealepithelial cells were seeded onto 96-well TCPS culturetrays at a concentration of 1 × 104 cells/well in thepresence of peptide/peptoid solutions serially dilutedin serum-free DMEM/F12 ranging from 1 mg/mLdown to zero. Each dilution point was assayed in trip-licate. The cells were incubated for 24 h and the numberof cells that were attached at this time was measuredand expressed in a similar manner to that described forthe cytotoxicity assay.
2. Identification of peptide sequences capable of inhibiting cellattachment to immobilized (GPN1)10GP-NH2. Sterilestrips (1 × 1 cm) of films containing immobilized(GPNl)10GP-NH2 were placed into the wells of a 24-well tissue-culture tray (Cell Wells™, cat. no. 25820,Corning, NY). Corneal epithelial cells were seeded at adensity of 2 × 105 cells/well in the presence of 2 mg/mL peptide solutions in serum-free DMEM/F12.Where the competing molecule solution contained apeptoid residue, the concentration of the solutions usedwas 125 mg/mL, to avoid the effects of increased vis-cosity. Each sample was assayed in triplicate, each oneof which was contained in a separate well. Cells were
614 JOHNSON ET AL.

incubated for 6 h, fixed with formol-saline (10% v/vformalin/phosphate buffered saline), then stainedwith a solution of 0.1% crystal violet in 0.02M phos-phate buffer (pH 7.0). The bound stain was solubilizedwith 10% (v/v) acetic acid and the absorbance thenread at 595 nm on a plate reader. Cell numbers wereexpressed in the same manner as described for the cy-totoxicity assay.
Cell growth assay
Cells were seeded at a density of 3 × 104 cells/well ontosterile 1-cm2 fluorinated ethylene propylene copolymerstrips containing immobilized collagen-mimetic molecules.Individual samples were contained in separate wells of a24-well tissue-culture tray in DMEM/F12 culture mediumsupplemented with 20% (v/v) fetal bovine serum that hadbeen depleted of the cell attachment glycoproteins vitronec-tin and fibronectin.29 Cells were cultured for 7 days, with amedium change every second day. Final cell numbers weremeasured and expressed in the same manner as describedfor the cytotoxicity assay. Each assay was performed in trip-licate.
Corneal epithelial tissue outgrowth assay
The preparation of the explant tissue was based on modi-fications of previously described methods.30,31 Briefly, cor-neas were excised from enucleated cows’ eyes using finescissors and removed with a thin scleral rim still attached tofacilitate handling. Forceps were used to remove the endo-thelium, Descemet’s membrane, and the bulk of the stromaso that only the epithelial layer and a small amount of as-sociated stromal tissue remained. Disks, 6 mm in diameter,were then cut from around the periphery of the remainingcorneal tissue using a sterile biopsy punch (Stiefel Labora-tories, Sydney, Australia) for use in the outgrowth assay.
Two types of tissue outgrowth assays were performed:
1. Identification of peptoid-containing sequences capable of in-hibiting the epithelial tissue outgrowth onto TCPS. For thetissue outgrowth assay, 6-mm diameter tissue buttonswere placed epithelial side up onto the surface in thecenter of a 12-well TCPS plate (Cell Wells™, cat. no.25815). Three replicates of each test condition were as-sayed, and to reduce experimental bias from possiblecornea-to-cornea variation, the tissue samples used foreach of the three replicates were cut from different cor-neas. The tissue buttons were allowed to attach to theunderlying surface for 20 min in air. In this assay inwhich collagen-mimetic molecules were tested for in-hibition, 0.5 mL of a solution made up at a concentra-tion of 1 mg of collagen mimetic/mL in serum-freeDMEM/F12 culture medium was added to each wellcontaining a “test” treatment. This volume raised thelevel of the culture medium to the top of the tissueexplant without completely immersing it. The mediumused to replenish these wells on every second day of
the 8-day assay period also contained 1 mg of collagenmimetic/mL. At day 8, the explants were washed withphosphate buffered saline, pH 7.2, and fixed in formolsaline. The fixed explants were stained with 1% (w/v)methylene blue (Gurr GI 52015) in 0.01M borate bufferpH 8.5 for 30 min, rinsed several times with boratebuffer, and tissue outgrowth measured using a LeicaCambridge Q570 image analyzer. An outgrowth index,calculated as the final outgrowth area divided by theinitial tissue area, was established for each sample andthen expressed as a mean (±SD) percentage of the out-growth measured on the control TCPS surface.
2. Identification of immobilized collagen-mimetic sequences ca-pable of stimulating tissue outgrowth. To measure cornealepithelial tissue outgrowth on immobilized peptides/peptoids, sterile test samples each 4 cm2 in size, wereplaced into the wells of a 12-well tissue culture tray. Asingle 6-mm diameter disk of corneal tissue was placed,epithelial side uppermost, on the center of each poly-mer sample. Three replicates of each polymer samplewere assayed. Tissue explant disks were allowed toattach to the surface of the polymer samples for 20 minbefore adding 0.5 mL of serum-free DMEM/F12 cul-ture medium. The explants were incubated for 8 daysand the culture medium was changed three times dur-ing that period. The final explant outgrowth index wascalculated as described above.
Statistical treatment of data
Each experiment was repeated at least twice and statisticalanalysis of data was done using Student’s t test.
RESULTS
Melting studies for (GPNl)10GP-NH2collagen-mimetic sequences
The thermal stability of (GPNl)10GP-NH2 was as-sessed by temperature-dependent optical rotationmeasurements (Fig. 1). Table II compares these datawith the results of a series of collagen mimetics con-taining (GPNl)n-NH2 sequences.
Cytotoxicity
Before conducting any other assay, the collagen-mimetic molecules were assessed for cytotoxicity. Cor-neal epithelial cells and corneal fibroblasts were al-lowed to attach and spread on TCPS for 24 h inDMEM/F12 culture medium supplemented with 20%(v/v) fetal bovine serum. They were then exposed fora further 24 h to serum-free culture media containing
615CELL INTERACTIONS WITH COLLAGEN MIMETICS
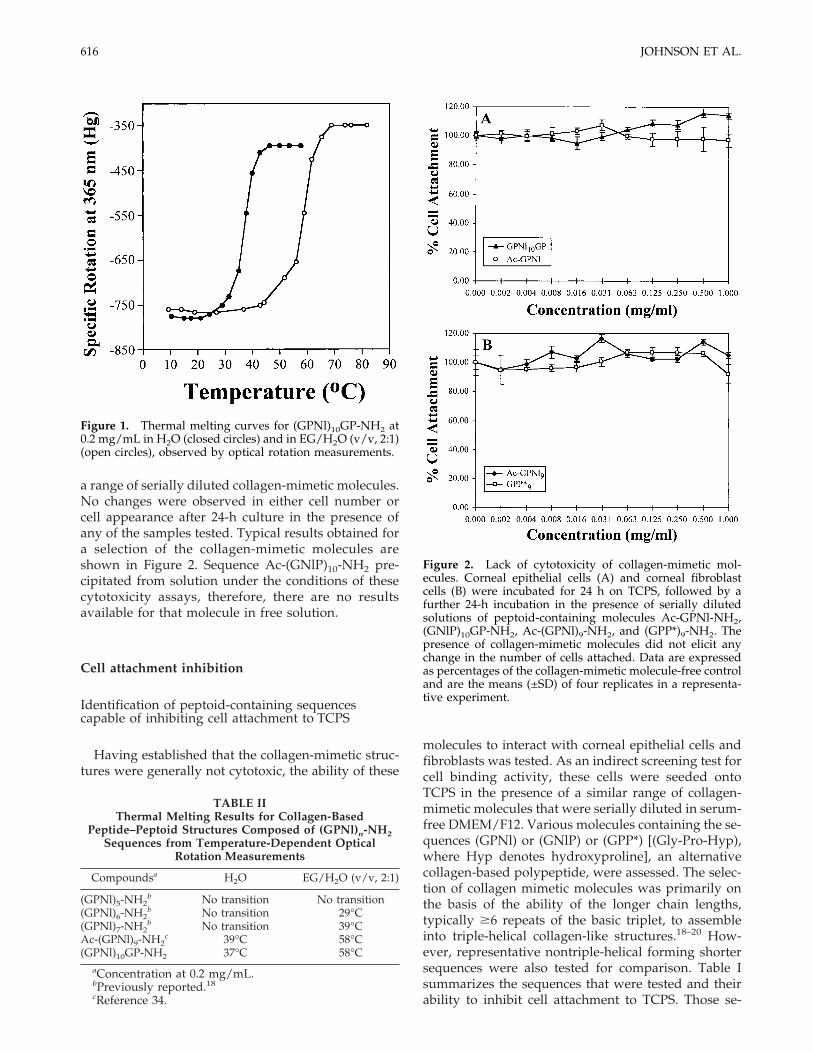
a range of serially diluted collagen-mimetic molecules.No changes were observed in either cell number orcell appearance after 24-h culture in the presence ofany of the samples tested. Typical results obtained fora selection of the collagen-mimetic molecules areshown in Figure 2. Sequence Ac-(GNlP)10-NH2 pre-cipitated from solution under the conditions of thesecytotoxicity assays, therefore, there are no resultsavailable for that molecule in free solution.
Cell attachment inhibition
Identification of peptoid-containing sequencescapable of inhibiting cell attachment to TCPS
Having established that the collagen-mimetic struc-tures were generally not cytotoxic, the ability of these
molecules to interact with corneal epithelial cells andfibroblasts was tested. As an indirect screening test forcell binding activity, these cells were seeded ontoTCPS in the presence of a similar range of collagen-mimetic molecules that were serially diluted in serum-free DMEM/F12. Various molecules containing the se-quences (GPNl) or (GNlP) or (GPP*) [(Gly-Pro-Hyp),where Hyp denotes hydroxyproline], an alternativecollagen-based polypeptide, were assessed. The selec-tion of collagen mimetic molecules was primarily onthe basis of the ability of the longer chain lengths,typically $6 repeats of the basic triplet, to assembleinto triple-helical collagen-like structures.18–20 How-ever, representative nontriple-helical forming shortersequences were also tested for comparison. Table Isummarizes the sequences that were tested and theirability to inhibit cell attachment to TCPS. Those se-
TABLE IIThermal Melting Results for Collagen-Based
Peptide–Peptoid Structures Composed of (GPNl)n-NH2Sequences from Temperature-Dependent Optical
Rotation Measurements
Compoundsa H2O EG/H2O (v/v, 2:1)
(GPNl)5-NH2b No transition No transition
(GPNl)6-NH2b No transition 29°C
(GPNl)7-NH2b No transition 39°C
Ac-(GPNl)9-NH2c 39°C 58°C
(GPNl)10GP-NH2 37°C 58°CaConcentration at 0.2 mg/mL.bPreviously reported.18
cReference 34.
Figure 1. Thermal melting curves for (GPNl)10GP-NH2 at0.2 mg/mL in H2O (closed circles) and in EG/H2O (v/v, 2:1)(open circles), observed by optical rotation measurements.
Figure 2. Lack of cytotoxicity of collagen-mimetic mol-ecules. Corneal epithelial cells (A) and corneal fibroblastcells (B) were incubated for 24 h on TCPS, followed by afurther 24-h incubation in the presence of serially dilutedsolutions of peptoid-containing molecules Ac-GPNl-NH2,(GNlP)10GP-NH2, Ac-(GPNl)9-NH2, and (GPP*)9-NH2. Thepresence of collagen-mimetic molecules did not elicit anychange in the number of cells attached. Data are expressedas percentages of the collagen-mimetic molecule-free controland are the means (±SD) of four replicates in a representa-tive experiment.
616 JOHNSON ET AL.
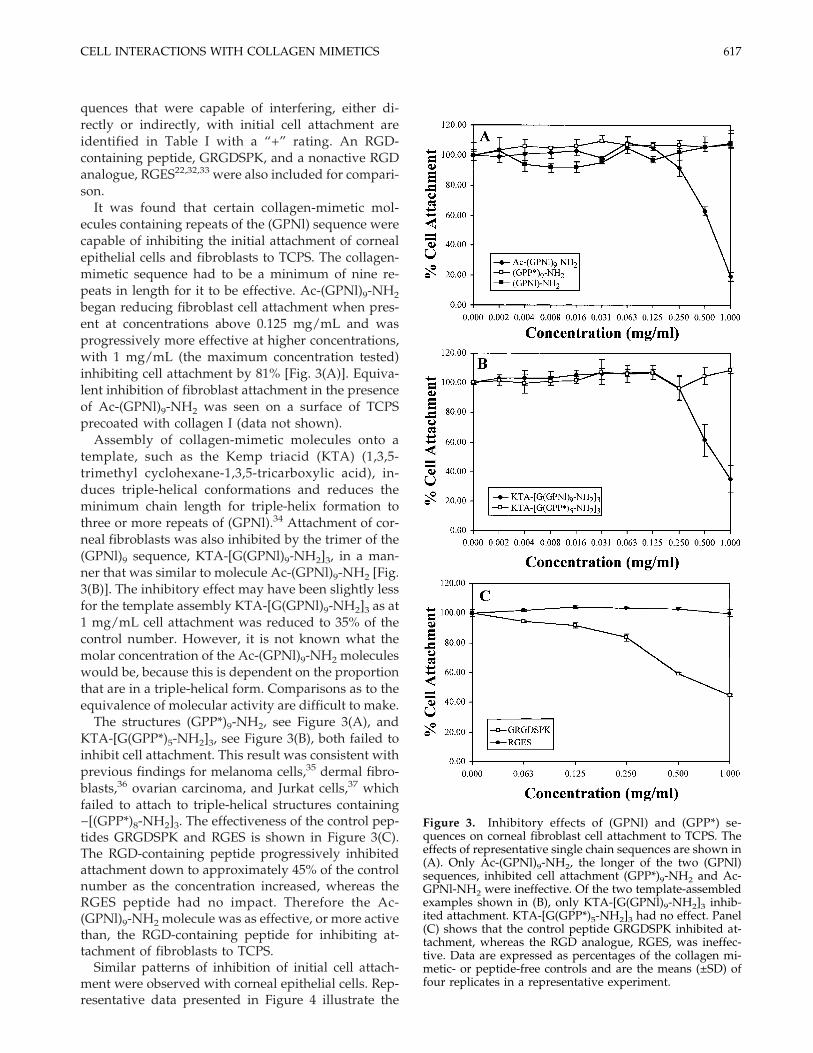
quences that were capable of interfering, either di-rectly or indirectly, with initial cell attachment areidentified in Table I with a “+” rating. An RGD-containing peptide, GRGDSPK, and a nonactive RGDanalogue, RGES22,32,33 were also included for compari-son.
It was found that certain collagen-mimetic mol-ecules containing repeats of the (GPNl) sequence werecapable of inhibiting the initial attachment of cornealepithelial cells and fibroblasts to TCPS. The collagen-mimetic sequence had to be a minimum of nine re-peats in length for it to be effective. Ac-(GPNl)9-NH2
began reducing fibroblast cell attachment when pres-ent at concentrations above 0.125 mg/mL and wasprogressively more effective at higher concentrations,with 1 mg/mL (the maximum concentration tested)inhibiting cell attachment by 81% [Fig. 3(A)]. Equiva-lent inhibition of fibroblast attachment in the presenceof Ac-(GPNl)9-NH2 was seen on a surface of TCPSprecoated with collagen I (data not shown).
Assembly of collagen-mimetic molecules onto atemplate, such as the Kemp triacid (KTA) (1,3,5-trimethyl cyclohexane-1,3,5-tricarboxylic acid), in-duces triple-helical conformations and reduces theminimum chain length for triple-helix formation tothree or more repeats of (GPNl).34 Attachment of cor-neal fibroblasts was also inhibited by the trimer of the(GPNl)9 sequence, KTA-[G(GPNl)9-NH2]3, in a man-ner that was similar to molecule Ac-(GPNl)9-NH2 [Fig.3(B)]. The inhibitory effect may have been slightly lessfor the template assembly KTA-[G(GPNl)9-NH2]3 as at1 mg/mL cell attachment was reduced to 35% of thecontrol number. However, it is not known what themolar concentration of the Ac-(GPNl)9-NH2 moleculeswould be, because this is dependent on the proportionthat are in a triple-helical form. Comparisons as to theequivalence of molecular activity are difficult to make.
The structures (GPP*)9-NH2, see Figure 3(A), andKTA-[G(GPP*)5-NH2]3, see Figure 3(B), both failed toinhibit cell attachment. This result was consistent withprevious findings for melanoma cells,35 dermal fibro-blasts,36 ovarian carcinoma, and Jurkat cells,37 whichfailed to attach to triple-helical structures containing−[(GPP*)8-NH2]3. The effectiveness of the control pep-tides GRGDSPK and RGES is shown in Figure 3(C).The RGD-containing peptide progressively inhibitedattachment down to approximately 45% of the controlnumber as the concentration increased, whereas theRGES peptide had no impact. Therefore the Ac-(GPNl)9-NH2 molecule was as effective, or more activethan, the RGD-containing peptide for inhibiting at-tachment of fibroblasts to TCPS.
Similar patterns of inhibition of initial cell attach-ment were observed with corneal epithelial cells. Rep-resentative data presented in Figure 4 illustrate the
Figure 3. Inhibitory effects of (GPNl) and (GPP*) se-quences on corneal fibroblast cell attachment to TCPS. Theeffects of representative single chain sequences are shown in(A). Only Ac-(GPNl)9-NH2, the longer of the two (GPNl)sequences, inhibited cell attachment (GPP*)9-NH2 and Ac-GPNl-NH2 were ineffective. Of the two template-assembledexamples shown in (B), only KTA-[G(GPNl)9-NH2]3 inhib-ited attachment. KTA-[G(GPP*)5-NH2]3 had no effect. Panel(C) shows that the control peptide GRGDSPK inhibited at-tachment, whereas the RGD analogue, RGES, was ineffec-tive. Data are expressed as percentages of the collagen mi-metic- or peptide-free controls and are the means (±SD) offour replicates in a representative experiment.
617CELL INTERACTIONS WITH COLLAGEN MIMETICS

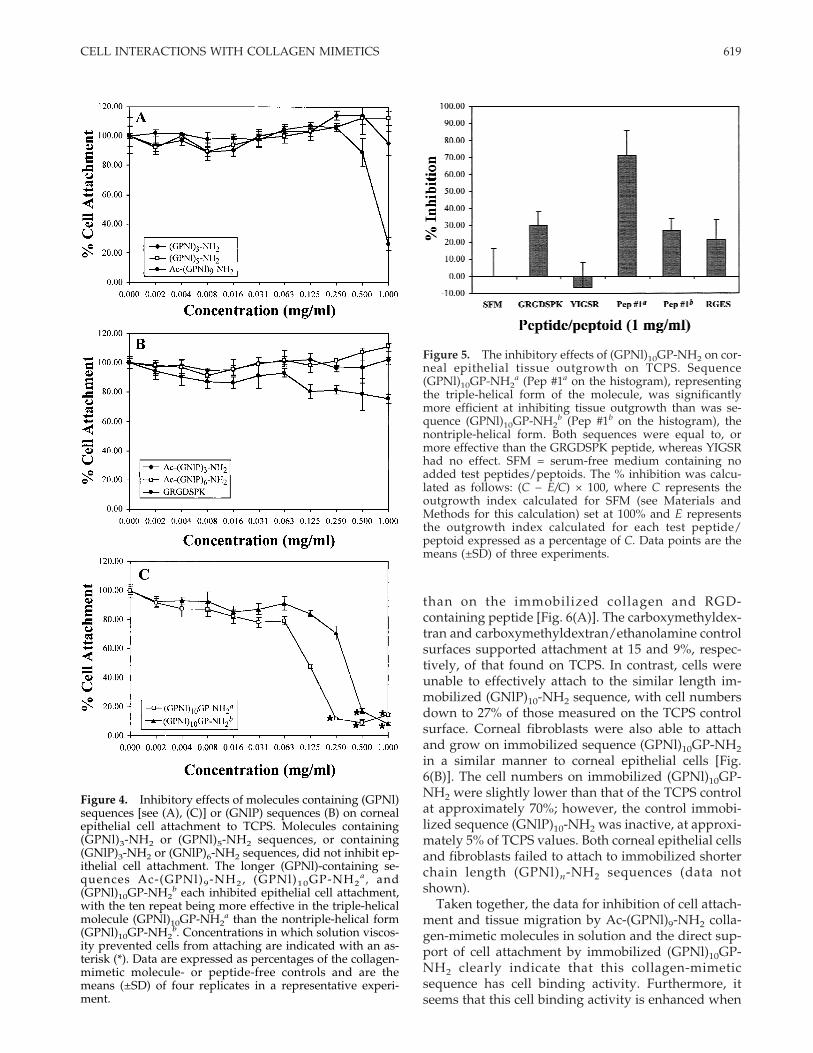
influence of sequence length, and of sequence speci-ficity. Although Ac-(GPNl)9-NH2 inhibited epithelialcell attachment to TCPS, molecules containing lessthan nine repeats of (GPNl) were ineffective. Mol-ecules with three or five repeats of the (GPNl) se-quence had little impact on the ability of corneal epi-thelial cells to attach to TCPS [Fig. 4(A)]. Ac-(GPNl)9-NH2 was equally effective at inhibiting cornealepithelial cell attachment as it was with the cornealfibroblasts. In the presence of 1 mg/mL, numbers ofattached cells were reduced to just 25% of those foundin the controls [Fig. 4(a)]. The control GRGDSPK pep-tide inhibited corneal epithelial cells to a lesser extent(inhibition of approximately 20%) than for the cornealfibroblasts. This inhibition was significant, however,when compared with the controls used in this experi-ment, the sequences Ac-(GNlP)3-NH2 and Ac-(GNlP)6-NH2 (p = 0.0002 and p < 0.0001, respectively).
The control sequence, (GNlP)n-NH2 was available inthe forms of three and six repeats and these weretested for effects on cell attachment. Both sequenceswere ineffective at inhibiting corneal epithelial cell at-tachment to TCPS [Fig. 4(B)]. The longer sequence Ac-(GNlP)10-NH2, which would be the preferred se-quence for comparison to either Ac-(GPNl)9-NH2 orAc-(GPNl)10GP-NH2, tended to precipitate out of so-lution (see Table III of Goodman et al.34) and so couldnot be tested in this solution inhibition assay format.
The final attachment inhibition data presented com-pares the efficiency of the triple-helical and singlechain conformations of the longest sequence tested,ten repeats of (GPNl). The triple-helical form(GPNl)10GP-NH2
a was prepared by storing a 2 mg/mL solution at 4°C for a minimum of 1 week. A secondsolution (GPNl)10GP-NH2
b was used when freshlymade, at which stage the sequence would be expectedto be in nontriple-helical form (see Feng, 199719). At125 mg/mL, the highest concentration at which allcells could settle onto the surface, both versions of themolecule inhibited cell attachment. The triple-helicalconformation (GPNl)10GP-NH2
a, however, was sig-nificantly (p = 0.0036) more effective at inhibiting cor-neal epithelial cell attachment than the nontriple-helical form [Fig. 4(C)]. At all other concentrations be-low 0.125 mg/mL, the triple-helical form was alsomore effective. It was discovered that both solutionsof this sequence became viscous if higher concentra-tions were used in this assay. Solution (GPNl)10GP-NH2
a became viscous above 0.125 mg/mL, while(GPNl)10GP-NH2
b became viscous above a concentra-tion of 0.250 mg/mL. The effect of viscosity abovethose concentrations was to prevent most of the cellsfrom reaching the TCPS attachment substrate duringthe assay period, and so the highest concentration thatcould be used in the assay to give an interpretableresult was 125 mg/mL.
Corneal tissue outgrowth inhibition
Because (GPNl)10GP-NH2 interferes with cell at-tachment and so appears to have cell binding activity,this collagen mimetic was also tested for ability toinhibit epithelial tissue migration. Ascertaining the ef-fect on corneal tissue outgrowth was of interest be-cause one of the objectives of this study was to identifypeptoid-containing molecules that could be used assurfaces in ophthalmic implants. Both triple-helical(GPNl)10GP-NH2
a and nontriple-helical (GPNl)10GP-NH2
b forms were tested for the ability to inhibit theoutgrowth of intact corneal epithelial tissue on TCPS.
The triple-helical form of (GPNl)10GP-NH2 stronglyinhibited corneal epithelial tissue migration. It inhib-ited tissue outgrowth on TCPS by 70%, whereas thenontriple-helical form was less effective at 27% inhi-bition (Fig. 5). By comparison, the laminin pentapep-tide YIGSR did not inhibit tissue migration at all andGRGDSPK and RGES peptides inhibited outgrowthby only 30 and 20%, respectively.
Cell attachment and growth on immobilizedpeptides and collagen-mimetic molecules
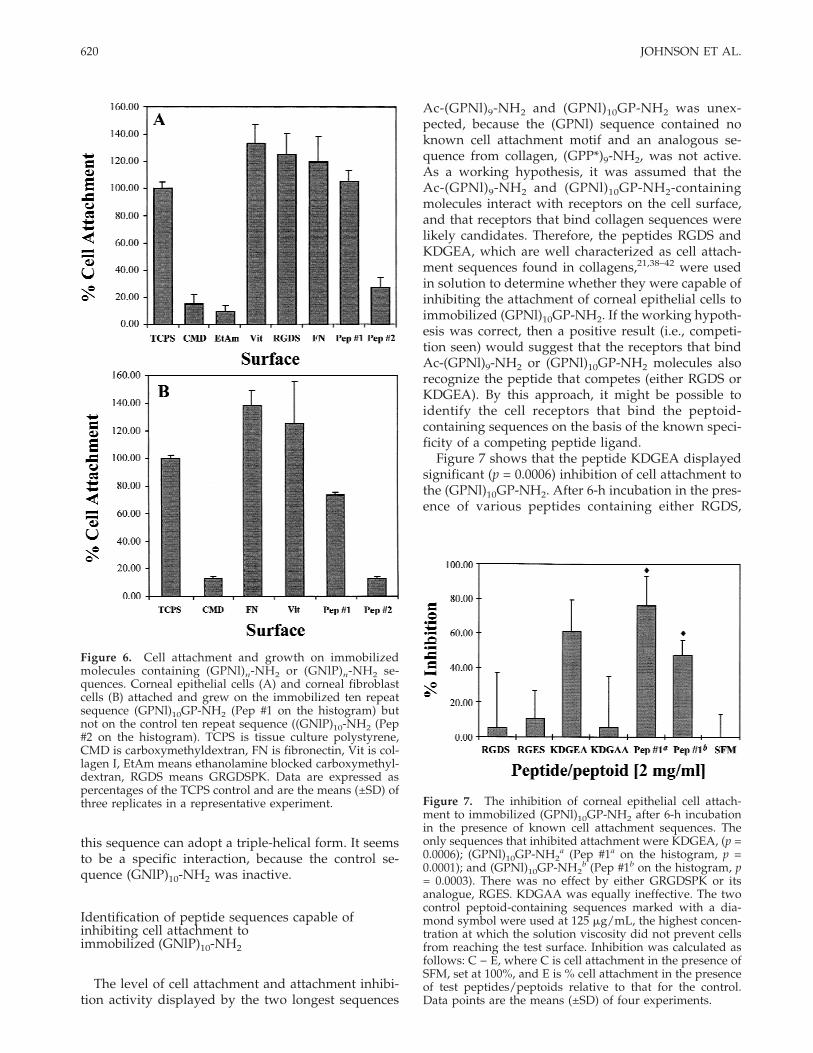
The inhibition of cell attachment and tissue migra-tion, when the collagen-mimetic molecules were pre-sented in solution as a competitor, is indirect evidencefor these molecules having cell binding activity. Toconfirm that these collagen-mimetic molecules werecapable of direct cell binding, the (GPNl)10GP-NH2sequence was immobilized onto a surface. The immo-bilization was accomplished via a carboxymethyldex-tran spacer molecule that was previously attached toplasma-modified fluorinated ethylene propylene co-polymer. Carboxymethyldextran-containing surfaces(without collagen mimetic attached) were inactive forcell attachment. The similar length control sequence(GNlP)10-NH2 was coupled in a similar manner andused as a peptoid-containing control surface. Se-quence length was also an important factor in the abil-ity of these molecules containing (GPNl) repeats tointeract with cell attachment mechanisms, so a seriesof shorter-length sequences of (GPNl)nNH2, in whichn = 2, 5, and 7 (see Table I) were immobilized. Thesewere tested for the ability to support attachment andgrowth of corneal epithelial cells and fibroblasts. Ex-amples of known cell attachment molecules such asfibronectin (FN), collagen I, and the RGD-containingpeptide, GRGDSPK, were used as positive controls.
Immobilized sequence (GPNl)10GP-NH2 supportedthe attachment and proliferation of corneal epithelialcells to similar levels as those found on the TCPS andfibronectin control surfaces, while being slightly lower
618 JOHNSON ET AL.
than on the immobilized collagen and RGD-containing peptide [Fig. 6(A)]. The carboxymethyldex-tran and carboxymethyldextran/ethanolamine controlsurfaces supported attachment at 15 and 9%, respec-tively, of that found on TCPS. In contrast, cells wereunable to effectively attach to the similar length im-mobilized (GNlP)10-NH2 sequence, with cell numbersdown to 27% of those measured on the TCPS controlsurface. Corneal fibroblasts were also able to attachand grow on immobilized sequence (GPNl)10GP-NH2in a similar manner to corneal epithelial cells [Fig.6(B)]. The cell numbers on immobilized (GPNl)10GP-NH2 were slightly lower than that of the TCPS controlat approximately 70%; however, the control immobi-lized sequence (GNlP)10-NH2 was inactive, at approxi-mately 5% of TCPS values. Both corneal epithelial cellsand fibroblasts failed to attach to immobilized shorterchain length (GPNl)n-NH2 sequences (data notshown).
Taken together, the data for inhibition of cell attach-ment and tissue migration by Ac-(GPNl)9-NH2 colla-gen-mimetic molecules in solution and the direct sup-port of cell attachment by immobilized (GPNl)10GP-NH2 clearly indicate that this collagen-mimeticsequence has cell binding activity. Furthermore, itseems that this cell binding activity is enhanced when
Figure 4. Inhibitory effects of molecules containing (GPNl)sequences [see (A), (C)] or (GNlP) sequences (B) on cornealepithelial cell attachment to TCPS. Molecules containing(GPNl)3-NH2 or (GPNl)5-NH2 sequences, or containing(GNlP)3-NH2 or (GNlP)6-NH2 sequences, did not inhibit ep-ithelial cell attachment. The longer (GPNl)-containing se-quences Ac-(GPNl)9-NH2, (GPNl)10GP-NH2
a , and(GPNl)10GP-NH2
b each inhibited epithelial cell attachment,with the ten repeat being more effective in the triple-helicalmolecule (GPNl)10GP-NH2
a than the nontriple-helical form(GPNl)10GP-NH2
b. Concentrations in which solution viscos-ity prevented cells from attaching are indicated with an as-terisk (*). Data are expressed as percentages of the collagen-mimetic molecule- or peptide-free controls and are themeans (±SD) of four replicates in a representative experi-ment.
Figure 5. The inhibitory effects of (GPNl)10GP-NH2 on cor-neal epithelial tissue outgrowth on TCPS. Sequence(GPNl)10GP-NH2
a (Pep #1a on the histogram), representingthe triple-helical form of the molecule, was significantlymore efficient at inhibiting tissue outgrowth than was se-quence (GPNl)10GP-NH2
b (Pep #1b on the histogram), thenontriple-helical form. Both sequences were equal to, ormore effective than the GRGDSPK peptide, whereas YIGSRhad no effect. SFM = serum-free medium containing noadded test peptides/peptoids. The % inhibition was calcu-lated as follows: (C − E/C) × 100, where C represents theoutgrowth index calculated for SFM (see Materials andMethods for this calculation) set at 100% and E representsthe outgrowth index calculated for each test peptide/peptoid expressed as a percentage of C. Data points are themeans (±SD) of three experiments.
619CELL INTERACTIONS WITH COLLAGEN MIMETICS
this sequence can adopt a triple-helical form. It seemsto be a specific interaction, because the control se-quence (GNlP)10-NH2 was inactive.
Identification of peptide sequences capable ofinhibiting cell attachment toimmobilized (GNlP)10-NH2
The level of cell attachment and attachment inhibi-tion activity displayed by the two longest sequences
Ac-(GPNl)9-NH2 and (GPNl)10GP-NH2 was unex-pected, because the (GPNl) sequence contained noknown cell attachment motif and an analogous se-quence from collagen, (GPP*)9-NH2, was not active.As a working hypothesis, it was assumed that theAc-(GPNl)9-NH2 and (GPNl)10GP-NH2-containingmolecules interact with receptors on the cell surface,and that receptors that bind collagen sequences werelikely candidates. Therefore, the peptides RGDS andKDGEA, which are well characterized as cell attach-ment sequences found in collagens,21,38–42 were usedin solution to determine whether they were capable ofinhibiting the attachment of corneal epithelial cells toimmobilized (GPNl)10GP-NH2. If the working hypoth-esis was correct, then a positive result (i.e., competi-tion seen) would suggest that the receptors that bindAc-(GPNl)9-NH2 or (GPNl)10GP-NH2 molecules alsorecognize the peptide that competes (either RGDS orKDGEA). By this approach, it might be possible toidentify the cell receptors that bind the peptoid-containing sequences on the basis of the known speci-ficity of a competing peptide ligand.
Figure 7 shows that the peptide KDGEA displayedsignificant (p = 0.0006) inhibition of cell attachment tothe (GPNl)10GP-NH2. After 6-h incubation in the pres-ence of various peptides containing either RGDS,
Figure 6. Cell attachment and growth on immobilizedmolecules containing (GPNl)n-NH2 or (GNlP)n-NH2 se-quences. Corneal epithelial cells (A) and corneal fibroblastcells (B) attached and grew on the immobilized ten repeatsequence (GPNl)10GP-NH2 (Pep #1 on the histogram) butnot on the control ten repeat sequence ((GNlP)10-NH2 (Pep#2 on the histogram). TCPS is tissue culture polystyrene,CMD is carboxymethyldextran, FN is fibronectin, Vit is col-lagen I, EtAm means ethanolamine blocked carboxymethyl-dextran, RGDS means GRGDSPK. Data are expressed aspercentages of the TCPS control and are the means (±SD) ofthree replicates in a representative experiment. Figure 7. The inhibition of corneal epithelial cell attach-
ment to immobilized (GPNl)10GP-NH2 after 6-h incubationin the presence of known cell attachment sequences. Theonly sequences that inhibited attachment were KDGEA, (p =0.0006); (GPNl)10GP-NH2
a (Pep #1a on the histogram, p =0.0001); and (GPNl)10GP-NH2
b (Pep #1b on the histogram, p= 0.0003). There was no effect by either GRGDSPK or itsanalogue, RGES. KDGAA was equally ineffective. The twocontrol peptoid-containing sequences marked with a dia-mond symbol were used at 125 mg/mL, the highest concen-tration at which the solution viscosity did not prevent cellsfrom reaching the test surface. Inhibition was calculated asfollows: C − E, where C is cell attachment in the presence ofSFM, set at 100%, and E is % cell attachment in the presenceof test peptides/peptoids relative to that for the control.Data points are the means (±SD) of four experiments.
620 JOHNSON ET AL.
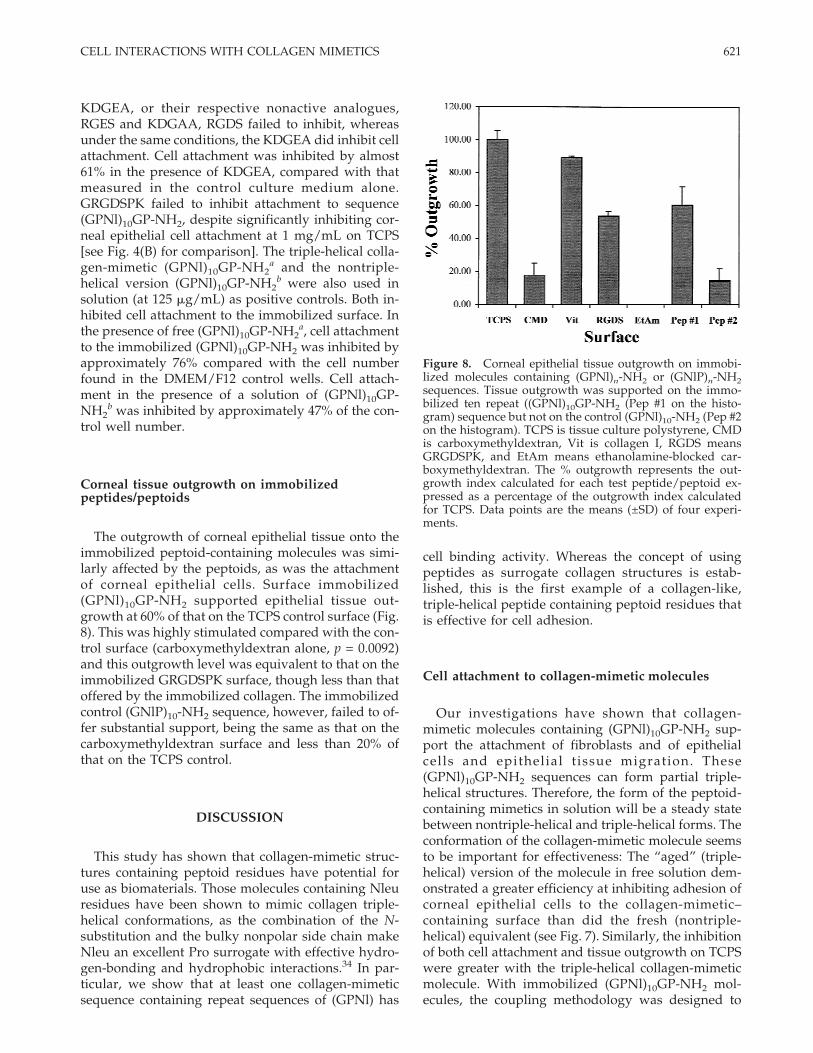
KDGEA, or their respective nonactive analogues,RGES and KDGAA, RGDS failed to inhibit, whereasunder the same conditions, the KDGEA did inhibit cellattachment. Cell attachment was inhibited by almost61% in the presence of KDGEA, compared with thatmeasured in the control culture medium alone.GRGDSPK failed to inhibit attachment to sequence(GPNl)10GP-NH2, despite significantly inhibiting cor-neal epithelial cell attachment at 1 mg/mL on TCPS[see Fig. 4(B) for comparison]. The triple-helical colla-gen-mimetic (GPNl)10GP-NH2
a and the nontriple-helical version (GPNl)10GP-NH2
b were also used insolution (at 125 mg/mL) as positive controls. Both in-hibited cell attachment to the immobilized surface. Inthe presence of free (GPNl)10GP-NH2
a, cell attachmentto the immobilized (GPNl)10GP-NH2 was inhibited byapproximately 76% compared with the cell numberfound in the DMEM/F12 control wells. Cell attach-ment in the presence of a solution of (GPNl)10GP-NH2
b was inhibited by approximately 47% of the con-trol well number.
Corneal tissue outgrowth on immobilizedpeptides/peptoids
The outgrowth of corneal epithelial tissue onto theimmobilized peptoid-containing molecules was simi-larly affected by the peptoids, as was the attachmentof corneal epithelial cells. Surface immobilized(GPNl)10GP-NH2 supported epithelial tissue out-growth at 60% of that on the TCPS control surface (Fig.8). This was highly stimulated compared with the con-trol surface (carboxymethyldextran alone, p = 0.0092)and this outgrowth level was equivalent to that on theimmobilized GRGDSPK surface, though less than thatoffered by the immobilized collagen. The immobilizedcontrol (GNlP)10-NH2 sequence, however, failed to of-fer substantial support, being the same as that on thecarboxymethyldextran surface and less than 20% ofthat on the TCPS control.
DISCUSSION
This study has shown that collagen-mimetic struc-tures containing peptoid residues have potential foruse as biomaterials. Those molecules containing Nleuresidues have been shown to mimic collagen triple-helical conformations, as the combination of the N-substitution and the bulky nonpolar side chain makeNleu an excellent Pro surrogate with effective hydro-gen-bonding and hydrophobic interactions.34 In par-ticular, we show that at least one collagen-mimeticsequence containing repeat sequences of (GPNl) has
cell binding activity. Whereas the concept of usingpeptides as surrogate collagen structures is estab-lished, this is the first example of a collagen-like,triple-helical peptide containing peptoid residues thatis effective for cell adhesion.
Cell attachment to collagen-mimetic molecules
Our investigations have shown that collagen-mimetic molecules containing (GPNl)10GP-NH2 sup-port the attachment of fibroblasts and of epithelialcells and epithelial tissue migration. These(GPNl)10GP-NH2 sequences can form partial triple-helical structures. Therefore, the form of the peptoid-containing mimetics in solution will be a steady statebetween nontriple-helical and triple-helical forms. Theconformation of the collagen-mimetic molecule seemsto be important for effectiveness: The “aged” (triple-helical) version of the molecule in free solution dem-onstrated a greater efficiency at inhibiting adhesion ofcorneal epithelial cells to the collagen-mimetic–containing surface than did the fresh (nontriple-helical) equivalent (see Fig. 7). Similarly, the inhibitionof both cell attachment and tissue outgrowth on TCPSwere greater with the triple-helical collagen-mimeticmolecule. With immobilized (GPNl)10GP-NH2 mol-ecules, the coupling methodology was designed to
Figure 8. Corneal epithelial tissue outgrowth on immobi-lized molecules containing (GPNl)n-NH2 or (GNlP)n-NH2sequences. Tissue outgrowth was supported on the immo-bilized ten repeat ((GPNl)10GP-NH2 (Pep #1 on the histo-gram) sequence but not on the control (GPNl)10-NH2 (Pep #2on the histogram). TCPS is tissue culture polystyrene, CMDis carboxymethyldextran, Vit is collagen I, RGDS meansGRGDSPK, and EtAm means ethanolamine-blocked car-boxymethyldextran. The % outgrowth represents the out-growth index calculated for each test peptide/peptoid ex-pressed as a percentage of the outgrowth index calculatedfor TCPS. Data points are the means (±SD) of four experi-ments.
621CELL INTERACTIONS WITH COLLAGEN MIMETICS

maximize the probability of triple-helical structuresbeing placed onto the surface through the use of“aged” solutions of molecules. The exact conformationof the coupled collagen-mimetic molecules is the sub-ject of another study and is yet to be determined.However, with surfaces that contained immobilized(GPNl)10GP-NH2 molecules and that were then auto-claved to denature any triple-helical formations, thecell attachment capacity was reduced by up to 80%(data not shown). The competition and direct cell ad-hesion data suggest that cell binding is dependent onthe (GPNl)n-containing collagen-mimetic structuresachieving a triple-helical conformation. The require-ment for nine or more repeats of the (GPNl) sequencefor cell binding activity correlates with the thermalstability data that indicate that seven or more repeatsare required for the stability associated with a triple-helical conformation18 to be achieved. It is most likely,therefore, that the lesser activity of molecules contain-ing less than nine repeats of the (GPNl) sequence isrelated to lack of triple helix formation, with resultantpoor receptor binding. Whether self-assembly oftriple-helical structures can occur after coupling ontothe surface is not known and we cannot exclude thepossibility that the nontriple-helical forms of collagen-mimetic molecules containing nine repeats of (GPNl)have activity, but at reduced levels. However, the al-ternative is that the response shown in the presence ofthe nontriple-helical version, the freshly (GPNl)10GP-NH2
b, might simply reflect the proportion of mol-ecules in this solution that had already begun to as-sume a triple-helical conformation. What is clear isthat triple-helicity alone, in a collagen mimetic, is notsufficient to provide an appropriate binding scaffoldfor cell attachment as demonstrated by the lack of cellbinding activity by (GPP*)n-NH2 (see also35–37). Fur-thermore, although (GNlP)n-NH2 sequences havehigher propensity for triple-helical packing than do(GPNl)n-NH2 sequences,34 (GNlP)10-NH2 was inactivefor cell binding. Study of the conformation of triple-helical and nontriple-helical conformations of colla-gen-mimetic molecules, and further comparison of theattachment of cells to these, may lead to the design ofmore conformationally appropriate collagen-mimeticmolecules.
Involvement of a2b1 cell surface receptors
We present indirect evidence that cell binding tomolecules containing the (GPNl)9-NH2 sequence oc-curs through the a2b1 cell surface receptor system.Based on a comparison of the evidence presentedherein and previous peptide competition studies, the
longer (GPNl)n-NH2 sequences seem to interact withcorneal epithelial cells through a similar mechanism tothat found in native collagens. The presence of thea2b1 heterodimer has been reported in human,43–46
rabbit,47 and rat48 corneal epithelial cells, although itwas not assayed for in the bovine corneal cells used inthis study. The basis of the tentative identification ofa2b1 integrin heterodimer as the likely binding site tolonger (GPNl)n-NH2 sequences is based on the inhibi-tion by the KDGEA peptide sequence of corneal epi-thelial cell attachment to the immobilized collagen-mimetic molecule. The data from previous studies thatidentify the tetrapeptide DGEA as a recognition sitefor the a2b1 integrin heterodimer are as follows.Staatz et al.39 identified DGEA as the a2b1 binding sitein collagen type I, although this interaction may besomewhat cell specific. In that instance, a2b1-mediated binding of breast carcinoma cells and plate-lets to collagen type I was inhibited by a KDGEA pen-tapeptide in solution. Similar findings were reportedfor the inhibition of adhesion of rabbit smooth musclecells to collagen type I,42,49 albeit after it had beenheat-denatured. Dermal fibroblasts,36 fibrosarcomacells,50 and (less effectively) cardiac fibroblasts andRat-1 cells,51 can also bind to native collagen type I viathe a2b1 complex. For most of these cases, denatur-ation of the collagen resulted in loss of the a2b1 bind-ing interaction, often to be replaced by a cryptic RGD-based interaction with a5b1.51,52 However the pres-ence of a DGEA-containing sequence is not aprerequisite for the interaction of a2b1 with most col-lagens and, indeed, second recognition sites have beendescribed recently for collagens type I and III. In col-lagen type I, this site contains an essential GER trip-let53 and in type III, the site has been mapped to thesequence GGPP*GPR54 and adjacent regions.55 DGEAhad no effect on a2b1-mediated binding of chondro-sarcoma cells to type II collagen,52 or of HT 1080 cellsto collagen type I.53 In many cases, no equivalent se-quences have been identified, although a2b1 integrinsare implicated in the binding of a variety of cells andplatelets to the triple-helical regions of collagens typeVI,33,56 type V,57 type IV,58–61 and type II.52 In all ofthese instances, the triple-helical conformation neededto be intact for effective a2b1 binding. Given thata2b1-mediated binding to collagens relies predomi-nantly on the conformation being triple-helical and theKDGEA pentapeptide significantly inhibits cornealepithelial cell attachment to the immobilized collagenmimetic, epithelial cell attachment to molecules con-taining the (GPNl)10GP-NH2 sequence may also bea2b1-dependent. Binding to the same receptor sitethat recognizes the ubiquitous RGD motif is notimplicated, as demonstrated by the inability ofGRGDSPK to inhibit cell binding to the immobilizedcollagen-mimetic molecule.
622 JOHNSON ET AL.

CONCLUSIONS
Significant progress has been made in the use ofpeptides selected for their ability to elicit specific cel-lular responses, particularly in the areas of cell attach-ment and tissue integration. These structures are con-structed primarily to mimic known peptide sequencessuch as the RGD and YIGSR cell attachment mo-tifs.11,62–65 In this study, we present a novel collagen-mimetic sequence, composed of nine or ten repeats ofthe sequence (GPNl), that has cell binding activitythrough an “RGD-independent” mechanism. Al-though we have not studied extensively the mecha-nism of cell attachment to this collagen-mimetic mol-ecule, we believe that the a2b1 integrin complex isinvolved. The primary aim of this study was to evalu-ate collagen-mimetic structures that contain peptoidresidues for potential use as biomaterials, especiallyfor use in ophthalmic implants. These collagen-mimetic molecules described herein appear to havepotential as cell and tissue support matrices.
The authors thank Drs. J. Werkmeister and P. A. Under-wood for critical review of the manuscript, and W. D. Fullerand V. V. Yalamoori from BioResearch Inc.
References
1. Ramshaw JA, Werkmeister JA, Glattauer V. Collagen-basedbiomaterials. Biotechnol Genet Eng Rev 1996;13:335–382.
2. Hilbert SL, Ferrans VJ, Jones M. Tissue-derived biomaterialsand their use in cardiovascular prosthetic devices. Med ProgTechnol 1988;14:115–163.
3. Khor E. Methods for the treatment of collagenous tissues forbioprostheses. Biomaterials 1997;18:95–105.
4. Ramshaw JA, Peters DE, Werkmeister JA, Ketharanathan V.Collagen organization in mandrel-grown vascular grafts. JBiomed Mater Res 1989;23:649–660.
5. Thompson KP, Hanna K, Waring GO III, Gipson I, Liu Y, Gaili-tis RP, Johnson–Wint B, Green K. Current status of syntheticepikeratoplasty. Refract Corneal Surg 1991;7:240–248.
6. Kirkham SM, Dangel ME. The keratoprosthesis: Improved bio-compatibility through design and surface modification. Oph-thalmic Surg 1991;22:455–461.
7. Cavallaro JF, Kemp PD, Kraus KH. Collagen fabrics as bioma-terials. Biotechnol Bioeng 1994;43:781–791.
8. Dunn MG, Avasarala PN, Zawadsky JP. Optimization of ex-truded collagen fibers for ACL reconstruction. J Biomed MaterRes 1993;27:1545–1552.
9. Hubbell JA. Biomaterials in tissue engineering. Biotechnology(NY) 1995;13:565–576.
10. Shakesheff K, Cannizzaro S, Langer R. Creating biomimeticmicro-environments with synthetic polymer-peptide hybridmolecules. J Biomater Sci Polym Ed 1998;9:507–518.
11. Pierschbacher MD, Polarek JW, Craig WS, Tschopp JF, SipesNJ, Harper JR. Manipulation of cellular interactions with bio-materials toward a therapeutic outcome: A perspective. J CellBiochem 1994;56:150–154.
12. Qian JJ, Bhatnagar RS. Enhanced cell attachment to anorganicbone mineral in the presence of a synthetic peptide related tocollagen. J Biomed Mater Res 1996;31:545–554.
13. Trinkaus–Randall V, Capecchi J, Newton A, Vadasz A,Leibowitz HM, Franzblau C. Development of a biopolymerickeratoprosthetic material. Invest Ophthalmol Vis Sci 1988;29:393–400.
14. Xie RZ, Sweeney DF, Beumer GJ, Johnson G, Griesser HJ,Steele JG. Effects of biologically modified surfaces of syntheticlenticules on corneal epithelialization in vivo. Aust N Z J Oph-thalmol 1997;25(Suppl 1):S46–49.
15. Simon RJ, Kania RS, Zuckermann RN, Huebner VD, Jewell DA,Banville S, Ng S, Wang L, Rosenberg S, Marlowe CK, Spell-meyer DC, Tan R, Frankel AD, Santi DV, Cohen FE, BartlettPA. Peptoids: A modular approach to drug discovery. ProcNatl Acad Sci USA 1992;89:9367–9371.
16. Melacini G, Feng Y, Goodman M. Collagen-based structurescontaining the peptoid residue N-isobutylglycine (Nleu): Con-formational analysis of Gly-Nleu-Pro sequences by 1H-NMRand molecular modeling. Biochemistry 1997;36:8725–8732.
17. Melacini G, Feng Y, Goodman M. Collagen-based structurescontaining the peptoid residue N-isobutylglycine (Nleu). 6.Conformational analysis of Gly-Pro-Nleu sequences by H-NMR, CD, and molecular modeling. J Am Chem Soc 1996;118:10725–10732.
18. Feng Y, Melacini G, Taulane JP, Goodman M. Collagen-basedstructures containing the peptoid residue N-isobutylglycine(Nleu): Synthesis and biophysical studies of Gly-Pro-Nleu se-quences by circular dichroism, ultraviolet absorbance, and op-tical rotation. Biopolymers 1996;39:859–872.
19. Feng Y, Melacini G, Goodman M. Collagen-based structurescontaining the peptoid residue N-isobutylglycine (Nleu): Syn-thesis and biophysical studies of Gly-Nleu-Pro sequences bycircular dichroism and optical rotation. Biochemistry 1997;36:8716–8724.
20. Goodman M, Melacini G, Feng Y. Collagen-like triple helicesincorporating peptoid residues. J Am Chem Soc 1996;118:10928–10929.
21. Pierschbacher MD, Ruoslahti E. Cell attachment activity of fi-bronectin can be duplicated by small synthetic fragments of themolecule. Nature 1984;309:30–33.
22. Nishida T, Nakagawa S, Watanabe K, Yamada KM, Otori T,Berman MB. A peptide from fibronectin cell-binding domaininhibits attachment of epithelial cells. Invest Ophthalmol VisSci 1988;29:1820–1825.
23. Hayman EG, Pierschbacher MD, Ruoslahti E. Detachment ofcells from culture substrate by soluble fibronectin peptides. JCell Biol 1985;100:1948–1954.
24. Hirano Y, Kando Y, Hayashi T, Goto K, Nakajima A. Synthesisand cell attachment activity of bioactive oligopeptides: RGD,RGDS, RGDV, and RGDT. J Biomed Mater Res 1991;25:1523–1534.
25. Griesser HJ, McLean KM, Beumer GJ, Gong X, Kingshott P,Johnson G, Steele JG. Surface immobilisation of synthetic pro-teins via plasma polymer interlayers. Conference Proceedings,25th Materials Research Society, Boston, MA; 1998. To appear.
26. Kingshott P, St. John HAW, Chatelier RC, Griesser HJ. Study ofprotein adsorption onto polysaccharide contact lens coatingsby MALDI-TOF-MS and XPS. Polym Sci Eng 1997;76:81–82.
27. Evans MDM, Steele JG. Multiple attachment mechanisms ofcorneal epithelial cells to a polymer: Cells can attach in theabsence of exogenous adhesion proteins through a mechanismthat requires microtubules. Exp Cell Res 1997;233:88–98.
28. Steele JG, Johnson G, Griesser HJ, Underwood PA. Mechanismof initial attachment of corneal epithelial cells to polymericsurfaces. Biomaterials 1997;18;1541–1551.
29. Underwood PA, Bennett FA. A comparison of the biologicalactivities of the cell-adhesive proteins vitronectin and fibronec-tin. J Cell Sci 1989;93:641–649.
30. Pettit DK, Horbett TA, Hoffman AS, Chan KY. Quantitation of
623CELL INTERACTIONS WITH COLLAGEN MIMETICS

rabbit corneal epithelial cell outgrowth on polymeric sub-strates in vitro. Invest Ophthalmol Vis Sci 1990;31:2269–2277.
31. Fitton JH, Dalton BA, Beumer G, Johnson G, Griesser HJ, SteeleJG. Surface topography can interfere with epithelial tissue mi-gration. J Biomed Mater Res 1998;42:245–257.
32. Dedhar S, Ruoslahti E, Pierschbacher MD. A cell surface re-ceptor complex for collagen type I recognizes the Arg-Gly-Aspsequence. J Cell Biol 1987;104:585–593.
33. Aumailley M, Mann K, von der Mark H, Timpl R. Cell attach-ment properties of collagen type VI and Arg-Gly-Asp depen-dent binding to its alpha 2(VI) and alpha 3(VI) chains. Exp CellRes 1989;181:463–474.
34. Goodman M, Bhumralkar M, Jefferson EA, Kwak J, Locardi E.Collagen mimetics. Biopolymers 1998;47:127–142.
35. Fields CG, Mickelson DJ, Drake SL, McCarthy JB, Fields GB.Melanoma cell adhesion and spreading activities of a synthetic124-residue triple-helical “mini-collagen.” J Biol Chem 1993;268:14153–14160.
36. Grab B, Miles AJ, Furcht LT, Fields GB. Promotion of fibroblastadhesion by triple-helical peptide models of type I collagen-derived sequences. J Biol Chem 1996;271:12234–12240.
37. Miles AJ, Skubitz AP, Furcht LT, Fields GB. Promotion of celladhesion by single-stranded and triple-helical peptide modelsof basement membrane collagen alpha 1(IV)531–543: Evidencefor conformationally dependent and conformationally inde-pendent type IV collagen cell adhesion sites. J Biol Chem 1994;269:30939–30945.
38. Yamada Y, Kleinman HK. Functional domains of cell adhesionmolecules. Curr Opin Cell Biol 1992;4:819–823.
39. Staatz WD, Fok KF, Zutter MM, Adams SP, Rodriguez BA,Santoro SA. Identification of a tetrapeptide recognition se-quence for the alpa 2 beta 1 integrin in collagen. J Biol Chem1991;266:7363–7367.
40. Santoro SA, Zutter MM, Wu JE, Staatz WD, Saelman EU, KeelyPJ. Analysis of collagen receptors. Methods Enzymol 1994;245:147–183.
41. Gullberg D, Terracio L, Borg TK, Rubin K. Identification ofintegrin-like matrix receptors with affinity for interstitial col-lagens. J Biol Chem 1989;264:12686–12694.
42. Yamamoto K, Yamamoto M. Cell adhesion receptors for nativeand denatured type I collagens and fibronectin in rabbit arte-rial smooth muscle cells in culture. Exp Cell Res 1994;214:258–263.
43. Maldonado BA, Furcht LT. Involvement of integrins with ad-hesion-promoting, heparin-binding peptides of type IV colla-gen in cultured human corneal epithelial cells. Invest Ophthal-mol Vis Sci 1995;36:364–372.
44. Lauweryns B, van den Oord JJ, Volpes R, Foets B, Missotten L.Distribution of very late activation integrins in the human cor-nea: An immunohistochemical study using monoclonal anti-bodies. Invest Ophthalmol Vis Sci 1991;32:2079–2085.
45. Tervo K, Tervo T, van Setten GB, Virtanen I. Integrins in hu-man corneal epithelium. Cornea 1991;10:461–465.
46. Virtanen I, Tervo K, Korhonen M, Paallysaho T, Tervo T. In-tegrins as receptors for extracellular matrix proteins in humancornea. Acta Ophthalmol Suppl 1992;18–21.
47. Wu XY, Cornell–Bell A, Davies TA, Simons ER, Trinkaus–Randall V. Expression of integrin and organization of F-actin inepithelial cells depends on the underlying surface. Invest Oph-thalmol Vis Sci 1994;35:878–890.
48. Stepp MA, Spurr–Micharud S, Gipson IK. Integrins in thewounded and unwounded stratified squamous epithelium ofthe cornea. Invest Ophthalmol Vis Sci 1993;34:1829–1844.
49. Yamamoto M, Yamato M, Aoyagi M, Yamamoto K. Identifica-tion of integrins involved in cell adhesion to native and dena-
tured type I collagens and the phenotypic transition of rabbitarterial smooth muscle cells. Exp Cell Res 1995;219:249–256.
50. Tuckwell D, Calderwood DA, Green LJ, Humphries MJ. Inte-grin alpha 2 I-domain is a binding site for collagens. J Cell Sci1995;108:1629–1637.
51. Gullberg D, Gehlsen KR, Turner DC, Ahlen K, Zijenah LS,Barnes MJ, Rubin K. Analysis of alpha 1 beta 1, alpha 2 beta 1and alpha 3 beta 1 integrins in cell–collagen interactions: Iden-tification of conformation dependent alpha 1 beta 1 bindingsites in collagen type I. Embo J 1992;11:3865–3873.
52. Tuckwell DS, Ayad S, Grant ME, Takigawa M, Humphries MJ.Conformation dependence of integrin-type II collagen binding:Inability of collagen peptides to support alpha 2 beta 1 binding,and mediation of adhesion to denatured collagen by a novelalpha 5 beta 1-fibronectin bridge. J Cell Sci 1994;107:993–1005.
53. Knight CG, Morton LF, Onley DJ, Peachey AR, Messent AJ,Smethurst PA, Tuckwell DS, Farndale RW, Barnes MJ. Identi-fication in collagen type I of an integrin alpha(2)beta(1)-binding site containing an essential GER sequence. J Biol Chem1998;273:33287–33294.
54. Morton LF, Peachey AR, Knight CG, Frandale RW, Barnes MJ.The platelet reactivity of synthetic peptides based on the col-lagen III fragment alphaI(III)CB4: Evidence for an integrinalpha2beta1 recognition site involving residues 522–528 of thealpha1(III) collagen chain. J Biol Chem 1997;272:11044–11048.
55. Glattauer V, Werkmeister JA, Kirkpatrick A, Ramshaw JA.Identification of the epitope for a monoclonal antibody thatblocks platelet aggregation induced by type III collagen. Bio-chem J 1997;323:45–49.
56. Pfaff M, Aumailley M, Specks U, Knolle J, Zerwes HG, TimplR. Integrin and Arg-Gly-Asp dependence of cell adhesion tothe native and unfolded triple helix of collagen type VI. ExpCell Res 1993;206:167–176.
57. Ruggiero F, Comte J, Cabanas C, Garrone R. Structural require-ments for alpha 1 beta 1 and alpha 2 beta 1 integrin mediatedcell adhesion to collagen V. J Cell Sci 1996;109:1865–1874.
58. Underwood PA, Bennett FA, Kirkpatrick A, Bean PA, MossBA. Evidence for the location of a binding sequence for thealpha 2 beta 1 integrin of endothelial cells, in the beta 1 subunitof laminin. Biochem J 1995;309:765–771.
59. Tuckwell DS, Reid KB, Barnes MJ, Humphries MJ. The A-domain of integrin alpha 2 binds specifically to a range ofcollagens but is not a general receptor for the collagenous mo-tif. Eur J Biochem 1996;241:732–739.
60. Vandenberg P, Kern A, Ries A, Luckenbill–Edds L, Mann K,Kuhn K. Characterization of a type IV collagen major cell bind-ing site with affinity to the alpha 1 beta 1 and the alpha 2 beta1 integrins. J Cell Biol 1991;113:1475–1483.
61. Kern A, Eble J, Golbik R, Kuhn K. Interaction of type IV col-lagen with the isolated integrins alpha 1 beta 1 and alpha 2 beta1. Eur J Biochem 1993;215:151–159.
62. Huber M, Heiduschka P, Kienle S, Pavlidis C, Mack J, Walk T,Jung G, Thanos S. Modification of glassy carbon surfaces withsynthetic laminin-derived peptides for nerve cell attachmentand neurite growth. J Biomed Mater Res 1998;41:278–288.
63. Glass JR, Dickerson KT, Stecker K, Polarek JW. Characteriza-tion of a hyaluronic acid-Arg-Gly-Asp peptide cell attachmentmatrix. Biomaterials 1996;17:1101–1108.
64. Grzesiak JJ, Pierschbacher MD, Amodeo MF, Malaney TI,Glass JR. Enhancement of cell interactions with collagen/glycosaminoglycan matrices by RGD derivatization. Biomate-rials 1997;18:1625–1632.
65. Drumheller PD, Hubbell JA. Polymer networks with graftedcell adhesion peptides for highly biospecific cell adhesive sub-strates. Anal Biochem 1994;222:380–388.
624 JOHNSON ET AL.





























