Embed Size (px)
Citation preview

The Biostratigraphy and Taxonomy of Graptoloids from the Ordovician and Silurian
of Britain
Thesis submitted for the degree of
Doctor of Philosophy
at the University of Leicester
by
Lindsey Taylor BSc. (Manchester)
Department of Geology
University of Leicester
September 1998

UMI Number: U115829
All rights reserved
INFORMATION TO ALL USERS The quality of this reproduction is dependent upon the quality of the copy submitted.
In the unlikely event that the author did not send a complete manuscript and there are missing pages, these will be noted. Also, if material had to be removed,
a note will indicate the deletion.
Dissertation Publishing
UMI U115829Published by ProQuest LLC 2013. Copyright in the Dissertation held by the Author.
Microform Edition © ProQuest LLC.All rights reserved. This work is protected against
unauthorized copying under Title 17, United States Code.
ProQuest LLC 789 East Eisenhower Parkway
P.O. Box 1346 Ann Arbor, Ml 48106-1346

The Biostratigraphy and Taxonomy of Graptoloids from the Ordovicianand Silurian of Britain
By Lindsey Taylor
ABSTRACT
This thesis presents new graptoloid biostratigraphic range charts for the Ordovician and
Silurian of Britain. The Ordovician graptoloid succession is represented by two separate biostratigraphic frameworks; one for England and Wales, another for Scotland, due to
significant stratigraphic disparity between the graptoloid faunal assemblages of the two
regions. The British Silurian sequence is represented by a single graptoloid
biostratigraphic scheme reflecting more uniform graptoloid assemblages.The reliability of the taxonomic foundation upon which the range charts are based
is tested using the dominant Ludlow Series (Silurian) graptoloid subfamily Saetograptinae
Urbanek. The results suggest that intraspecific variation has led to over-elaborate taxonomic subdivision of this group at the species and subspecies level. The poor preservation of some of the type material has resulted in problems of misidentification and
reinforces the need for a full taxonomic review of the saetograptids. Despite this, three basic theca/spine relationships can be recognised which may serve as a basis for future
species identification: a chimaera-type morphology whereby thecal spines project from the middle of the apertural margin; a leintwardinensis-type structure, in which the spine extends from the free ventral wall of the proceeding theca; and a colonus-type relationship whereby the lower part of the apertural margin is prolonged into a hook-like lip.
Graptoloid biodiversity data extracted from the new range charts highlights several periods of biotic crisis during the Ordovician and Silurian of the UK. These
'events' correlate with globally recognised intervals of major graptoloid extinctions, which in turn correspond with large-scale palaeoenvironmental fluctuations, such as
eustatic sea-level and climate change. Patterns of morphological change within UK graptoloids are also linked to these biodiversity trends and consequently, to environmental controls.

CONTENTS
Abstract
Contents i
List of figures ii
Acknowledgements iii
Introduction page 1.1
A taxonomic reassessment of the saetograptine genus Saetograptus page 2.1
Pribyl, 1942.
A global stratigraphy of British Ordovician and Silurian graptoloids. page 3.1
Patterns of diversity and morphological change in British Ordovician and page 4.1
Silurian graptoloids
Conclusions page 5.1
References page ref. 1
APPENDIX 1
Ordovician and Silurian graptoloid species diversity per biozone (data).
APPENDIX 2
Representative illustrations of British Silurian graptoloid species.
APPENDIX 3
Rhabdosome dimensions of Silurian graptoloid species from illustrations in
Appendix 2 (data).
i

LIST OF FIGURES
Introduction
Figure 1.1 Map showing the distribution of Ordovician and Silurian age strata in the
British Isles with particular reference to that of the Ludlow Series.
A taxonomic reassessment of the saetograptine genus Saetograptus Pribyl, 1942
Figure 2.1
Figure 2.2
Figure 2.3
Figure 2.4
Figure 2.5
Figure 2.6
Table 2.1
Table 2.2
A schematic diagram of a typical saetograptid graptolite.
Saetograptid type specimens, a. S. chimaera chimaera (L19973) x6.5. b. S. c.
semispinosus (BU1509) x5.7. c. S. c. salweyi (A24699) approx. x6.5. d. S.
leintwardinensis leintwardinensis (BU1526) x6.7. e. S. I. incipiens (BU1529) x5.
f. S. fritschi fritschi (L17764) x7. g. S. clunensis (72024 & 72027) x5. h. S.
wandalensis (A52619) xlO. i. S. colonus colonus (L19977) x8. j. S. varians
varians x8. k. S. colonus compactus (BU1498) x8. 1. S. roemeri (L17761) x6.7.
m. S. varians pumilis x5 (topotype).
Published illustrations of Saetograptus chimaera chimaera. a-c. After Boucek
(1936) all x5. d-f. After Pribyl (1942) all x5. g. After Barrande (1850) approx.
x5. h-i. After Elies & Wood (1911) all x5. j-k. After Berry (1964) all x2.5.1-m.
After Pemer (1899) x4 and x5 respectively.
Drawings of plates of isolated saetograptids from Urbanek, 1958. (a-d)
Saetograptus chimaera chimaera and (e-f) Saetograptus chimaera salweyi, lateral
and ventral view respectively, a, x l5; b & d, x l2; c, xlO; e & f, x l7 .
Published illustrations of Saetograptus leintwardinensis primus, a-e. After
Boucek (1936) all x5. f-h. After Pribyl (1942) all x5. Saetograptus fritschi
linearis, y \ . After Boucek (1936) all x5.
Published illustrations of Saetograptus colonus colonus. a-c. After Urbanek
(1958), a & c xlO, b xl3. d-h. After Boucek (1936) all x5. i-j. After Barrande
(1850). k. After Perner (1899) x7. m. After Elies & Wood (1911) x 5 .1. After
Wood (1900) x9.
Available rhabdosome dimensions (in mm) for the type material of the subgenus
Saetograptus (Saetograptus) Pribyl, 1942.
Available rhabdosome dimensions (in mm) for the type material of the subgenus
Saetograptus (Colonograptus) Pribyl, 1942.
A global biostratigraphy of British Ordovician and Silurian graptoloids
following
page 1.2
page 2.2
page 2.3
page 2.4
page 2.4
page 2.15
page 2.19
page 2.3
page 2.19
Figure 3.1 Proposed Ordovician chronostratigraphy, after Fortey et al. 1995, and page 3.1

comparison of Ordovician graptoloid biostratigraphy for England and Wales
and Scotland.
Figure 3.2 Silurian chronostratigraphy, after Holland et al. 1989, and accompanying
graptoloid biostratigraphy for the UK, this study.
Figure 3.3 Comparison of Silurian graptoloid biostratigraphic schemes for the UK,
Rickards (1976) and this study.
Figures 3.4.1- Ordovician graptoloid biostratigraphic range charts for England and Wales.
3.4.5
Figures 3.5.1- Ordovician graptoloid biostratigraphic range charts for Scotland.
3.5.3
Figures 3.6.1- Silurian graptoloid biostratigraphic range charts for the UK.
3.6.8
Patterns of diversity and morphological change in British Ordovician and Silurian graptoloids
Figure 4.1 UK Silurian graptoloid species diversity (with zonal boundary values).
Figure 4.2 UK Silurian graptoloid species diversity (without zonal boundary values),
showing main biodiversity crises.
Figure 4.3 Ordovician graptoloid species diversity for England and Wales (without zonal
boundary values), showing main biodiversity crises.
Figure 4.4 Ordovician graptoloid species diversity for Scotland (without zonal
boundary values), showing main biodiversity crises.
Figure 4.5 Total species diversity/biozone for la & b. The Ordovician of England and
Wales. 2a & b. The Ordovician of Scotland. 3a & b. The Silurian of the UK.
Figure 4.6 UK Ordovician and Silurian graptoloid species diversity (the Ordovician portion
of the chart combines data from England and Wales and Scotland).
Figure 4.7.1 Comparison of Ordovician Anglo-Welsh graptoloid diversity and number of
first appearance of species.
Figure 4.7.2 Comparison of Ordovician Anglo-Welsh graptoloid diversity and number of
last occurrence of species.
Figure 4.7.3 Ratio of graptoloid species diversity (appearance/disappearance) for the
Ordovician of England and Wales.
Figure 4.7.4 Comparison of % species disappearance and actual number of disappearances
for graptoloids of the Anglo-Welsh Ordovician.
Figure 4.8.1 Comparison of Ordovician Scottish graptoloid diversity and number of first
appearance of species.
Figure 4.8.2 Comparison of Ordovician Scottish graptoloid diversity and number of last
occurrence of species.
Figure 4.8.3 Ratio of graptoloid species diversity (appearance/disappearance) for the
Ordovician of Scotland.
page 3.1
page 3.9
page 3.9
page 3.9
page 3.23
page 4.3
page 4.3
page 4.3
page 4.5
page 4.6
page 4.3
page 4.3
page 4.3
page 4.4
page 4.4
page 4.3
page 4.3
page 4.6
ii

Figure 4.8.4 Comparison of % species disappearance and actual number of species
disappearances for graptoloids of the Scottish Ordovician.
Figure 4.9.1 Comparison of British Silurian graptoloid diversity and number of first
appearance of species.
Figure 4.9.2 Comparison of British Silurian graptoloid diversity and number of last
occurrence of species.
Figure 4.9.3 Ratio of graptoloid species diversity (appearance/disappearance) for the
Silurian of the UK.
Figure 4.9.4 Comparison of % species disappearance and actual number of species
disappearances for UK Silurian graptoloids.
Figure 4.10 Comparison of eustatic sea-level, after Loydell 1998, and
UK graptoloid species biodiversity, this study, for the Silurian.
Figure 4.11 Area diagram showing percentage total biozone fauna occupied by each
Silurian graptoloid family.
Figure 4.12 Area diagram showing the actual number of species per graptoloid family
present in each Silurian biozone.
Figure 4.13 Percentage of total fauna per biozone of selected graptoloid genera.
page 4.6
page 4.7
page 4.7
page 4.8
page 4.8
page 4.13
page 4.16
page 4.16
page 4.16
ii

Acknowledgements
This project was funded by a NERC studentship.
I would like to thank my supervisors Drs. Jan Zalasiewicz and David Siveter for all their advice
and support. Special thanks to Drs. Adrian Rushton, Mark Williams and Steve Tunnicliff
(BGS) for loads of help and useful suggestions, as well as posh coffee, choccy biccies and
interesting conversation. Also Drs. Petr Storch (Czech Academy of Sciences) and Petr Budil
(Czech Geological Survey) for being really nice and hospitable during my time in Prague. Many
thanks to Dr. Barrie Rickards (Cambridge) for perusing my range charts, as well as Drs. Roy
Clements (Leicester), Mike Dorling (Sedgwick Museum, Cambridge), John Clattworthy and
Dr. Paul Smith (Lapworth Museum, Birmingham) for allowing me to borrow the specimens I
needed.
I would never have completed this PhD if not for the support and friendship of the folks at
Leicester University Geology Department past and present. First and foremost, Annette, a
tireless, fearless friend and Tri’Sarah’tops (the Witches of Eastwick). Also, Duncan, Marion,
Roz, Matt, Trev, Loz, Jane, Kim, Stef, Craig & Rich & Alan the B, Sam, Karine, Barney
(poetic genius), Phil, Branney, Andy Saunders, Dick Aldridge, Norry, Dougy, Rod, Bill, etc.
Special thanks to the unfortunates who had to share an office with me, Tanya and Tiff (tops!).
Biggest thanks of all though to Jan for his relaxed and philosophical attitude to life - I ’ve got a
lotto learn!

Introduction
The graptoloids are a group of extinct hemichordates which are believed to have formed
planktonic colonies composed of interconnected zooids. They existed from the early Ordovician
until the middle Devonian and as fossils they are widespread, occurring globally, often in
abundance. These factors, combined with their apparently rapid rate of evolution make
graptoloids useful biostratigraphic indicators, worldwide, from the beginning of the Ordovician
to the mid-Devonian. In the U.K. however, the stratigraphic range of the group is restricted
from the early Ordovician to the late Silurian.
Project Background
Largely as a result of their biostratigraphic suitability, graptoloids have undergone a great deal
of palaeontological study; from their taxonomic classification and palaeobiology to their
taphonomy, palaeoecology, evolutionary patterns and biodiversity, graptoloid research has
advanced greatly in the last 40 years (Bulman, 1964, Bulman 1970, Rickards 1975, Crowther
& Rickards 1977, Rickards et al. 1977, Rickards & Dumican 1984, Dilly 1986, Kirk 1969,
Fortey & Cooper 1986, Briggs et al. 1995). Recently, many British graptoloid studies have
tended to focus on Ordovician and Silurian biostratigraphy and the refinement of the
biostratigraphical framework (Zalasiewicz 1992, 1994, 1996, Zalasiewicz & Tunnicliff 1994,
Zalasiewicz et al. 1995, Davies et al. 1994, Williams 1982, 1987, 1988, 1994, Stone &
Rushton 1983, Loydell & Cave 1993, Loydell 1993, Williams & Zalasiewicz in press).
Complete UK graptoloid biostratigraphical range charts have not been published since Rickards
(1976) for the Silurian, or since Elies and Wood (1901-18) for the Ordovician. For this reason,
revised and updated charts were required to take into account new stratigraphic and taxonomic
work.
Project Aims
The principal aim of this PhD has been the compilation and assessment of a graptoloid database,
containing full nomenclature and stratigraphic range information for all British graptoloid
species and subspecies from the Ordovician and Silurian. From this database, new British
Page 1.1

graptoloid biostratigraphical range charts could be produced for the Lower Palaeozoic.
Complementary to this part of the study has been an investigation of the often problematical
taxonomic basis of graptoloid biostratigraphy. This has been based on a re-examination of the
type material and literature of the dominant Ludlow Series graptoloid subfamily Saetograptinae.
A further project aim has been an examination of biodiversity and morphological change within
UK graptoloids and their relationship to global and local palaeoenvironmental controls. For this
purpose, the new graptoloid range charts provided a useful data source.
Geological Setting
British Ordovician and Silurian strata outcrop in the Southern Uplands of Scotland, the Lake
District of northern England and Wales and the Welsh Borderland (Fig. 1.1). These areas were
important centres of sedimentary deposition on the margins of the Lower Palaeozoic Iapetus
Ocean. The Ludlow Series forms part of the Upper Silurian (Fig. 3.2), and it is strata of this
age from which Wood (1900) obtained much of the type saetograptid material discussed later in
this study (Fig. 1.1).
Chrono- and Biostratigraphic Framework for the British Ordovician and Silurian
Biostratigraphic schemes are employed to enhance the fine-scale correlative value of the
chronostratigraphic column by utilising detailed changes in the composition of faunal
assemblages. Several fossil groups are useful biostratigraphically, for example, the conodonts,
acritarchs and ammonites. However, for the Lower Palaeozoic, graptoloids are the most widely
used for global correlation.
The Ordovician System is divided into five series, the Tremadoc, Arenig, Llanvirn,
Caradoc and Ashgill (Fortey et al. 1995). However, the British Isles was divided into two
provinces at this time separating Scotland from England and Wales for much of the Ordovician
(Fortey & Cocks 1986). As a result, the graptoloid faunas of both regions differ significantly in
strata representing this period and precise correlation is often difficult. Therefore, two different
biostratigraphic schemes are used. For the early to middle Ordovician of Scotland, graptoloids
closely resemble those of Australasia and hence the Australasian biostratigraphic framework has
been adopted for this part of the Scottish sequence (Stone & Rushton 1983). The Anglo-Welsh
Page 1.2

Ordovician succession is currently subdivided into 17 graptoloid biozones, while the succession
in Scotland contains 13 biozones (Fig. 3.1).
The base of the Tremadoc Series coincides with the base of the Ordovician and the series
is subdivided into two stages the younger of which, the Migneintian, possesses the earliest
recorded British graptoloids and approximately correlates with the murrayi graptoloid Biozone
of England and Wales (Fig. 3.1, 3.4.1). Graptolitic strata of this age are not recorded from
Scotland. The Arenig Series is divided into three stages and six graptoloid biozones in England
and Wales. The earliest Scottish graptoloid faunas occur in the lower Arenig, but they are
sparse and occur in isolated patches of strata (Stone & Strachan 1981, Stone & Rushton 1983).
The stratigraphy of the Llanvim Series has recently been reassessed and now includes the lower
part of the former Llandeilo Series (Fortey et al. 1995). It comprises two stages, the
Abereiddian and the Llandeilian, which are divided into three graptoloid biozones. The start of
the Caradoc Series corresponds with an increase in similarity between the graptoloid faunas of
Scotland and those of England and Wales, and the Scottish graptoloid sequence is continuous.
Consequently, a better biostratigraphic correlation between these regions can be achieved,
although for the remainder of the Ordovician the Scottish succession can be more finely
subdivided (Fig. 3.1).
By the base of the Silurian graptoloid faunas throughout the UK can be sufficiently well
correlated to allow a single biostratigraphic framework. Currently, the British Silurian sequence
is divided into 32 graptoloid biozones with a further 10 sub-biozones (Fig. 3.2), most of which
occur in the Llandovery Series. Chronostratigraphically, the system consists of four series; the
Llandovery, Wenlock, Ludlow and Pridolf, which are in turn split into stages. Strata
representing these series outcrop in the UK, but the Pridolf is non-graptolitic as the graptoloids
became extinct in Britain following the Bohemograptus proliferation Biozone in the early
Ludfordian Stage of the Ludlow Series (Fig. 3.2).
Thesis Structure
The thesis is structured to represent the three main stages of biostratigraphical work. Firstly, the
taxonomic assessment and identification of species and subspecies, which are essential for all
accurate biostratigraphic studies. Secondly, the production of faunal range charts which form
Page 1.3

the basis of local and global biostratigraphical correlation of sedimentary sequences. Finally,
applied biostratigraphy; that is the use of faunal ranges to elucidate patterns of diversity and
morphological change within a group of organisms, and how these may be affected by broader
palaeoenvironmental factors.
This thesis is written in the style of the Journal of the Geological Society, London.
Page 1.4

Clwydian RangeDenbighMoors
Llangollen
Long Mountain
LudlowftAbberleys
^M alvem s
BuilthWell
LEGEND
t i l l Ludlow
LEGEND
Silurian
Ordovician
Fig. 1.1. Map showing the distribution of Ordovician and Silurian age strata in the British Isles with
particular reference to the Ludlow Series.
Fig. 1.1

A taxonomic assessment of the saetograptine genus Saetograptus
Pribyl, 1942
A sound classification of any group of organisms is necessary if the group in question is to be
of any biostratigraphical use. Ideally, the type specimen of a taxon should be consulted before it
can be correctly identified elsewhere, though this is generally not the case in practice. This
situation, therefore, increases the need for good, up-to-date illustrations and descriptions of
type material, to ensure that ‘taxonomic drift’, or modification of the species concept in the
literature by successive incorrect identifications, does not occur. In the UK, parts of the
graptoloid stratigraphic succession have undergone little modem biostratigraphic study; for
example, the graptoloids of the Ludlow Series have been relatively little studied in comparison
with those from the Llandovery and Wenlock series (Loydell 1991, 1993, Loydell & Cave
1993, 1996, Zalasiewicz 1994, 1996, Williams & Zalasiewicz in press). Consequently, many
of the species and subspecies of graptoloid found in Ludlow age strata require taxonomic
revision. The purpose of this study is to illustrate and re-describe the type material belonging to
the subfamily Saetograptinae, a major Ludlow graptoloid group, in order to highlight any
taxonomic problems that may exist, and to constrain the morphological concept of each taxon.
This taxonomic reassessment also provides an insight into the level of reliability of the species
names appearing within the range charts assembled in the course of this thesis (see chapter 3).
The genus Saetograptus dominates graptolite-bearing strata in the Ludlow Series
worldwide, both in abundance and species diversity (Lapworth 1880, Wood 1900, Perner
1899, Boucek 1936, Urbanek 1958, Tomczyk 1956, Jaeger 1959, Krandievsky et al. 1968,
Lenz 1972, 1992, Berry 1964, Ross 1962, Cuerda 1965). Saetograptus is believed (Hutt 1969)
to have originated through a lineage which included Pristiograptus ludensis. In the UK,
saetograptid species range from the base of nilssoni Biozone at the Wenlock-Ludlow boundary,
to the leintwardinensis Biozone at the base of the Ludfordian Stage, upper Ludlow Series. The
type species of the genus Saetograptus, Graptolithus chimaera, was originally described from
Bohemia by Barrande (1850). The current taxonomic definition of the genus Saetograptus was
established by Pribyl (1942), who designated two subgenera distinguished by differences in
their thecal apertural processes: Saetograptus (Colonograptus) is biform, with hook-like,
Page 2.1

possibly retroverted thecae proximally (first 2-3 thecae), and simple tube-like thecae distally;
Saetograptus (Saetograptus) possesses true paired thecal spines, either throughout the
rhabdosome, or is semispinose (Fig. 2.1). Hutt (1969) emended the diagnosis of Saetograptus,
stating that ‘the thecae are straight and cylindrical with no ventral curvature, having round or
elliptical apertures with paired lateral processess. These processes take the form of lappets,
lappets and spines, or spines of enrolled periderm.’ This definition effectively renders obsolete
the use of the concept of the subgenus Colonograptus, although it still remains in use by
taxonomists. Based on this re-examination of saetograptid type material, I believe that there is
sufficient evidence for a subgeneric division of the group, with a clear distinction between those
forms with lappet-like processes and fully developed spines and those with proximal lappet-like
processes followed by simple thecae. Figure 2.1 is a sketch drawing of a typical saetograptid
and is intended as a guide to the terminology used in this chapter, it is not an accurate
representation of graptoloid anatomy.
Materials, methods and repositories
The type material for all available saetograptid species and subspecies has been re-drawn and
the morphological concepts of these species and subspecies analysed.
Specimens figured here with a catalogue number prefixed SM are housed in the Sedgwick
Museum, Cambridge, those with BU at Birmingham University, those with NMP at the
National Museum, Prague, and those without a prefix are housed at the British Geological
Survey, Keyworth, Nottingham.
Systematic Palaeontology
Order GRAPTOLOIDEA Lapworth, 1873
Suborder MONOGRAPTINA Lapworth, 1880
Family MONOGRAPTINAE Lapworth, 1873
Subfamily SAETOGRAPTINAE Urbanek, 1958
Genus SAETOGRAPTUS Pribyl, 1942, emend. Hutt, 1969
Subgenus SAETOGRAPTUS (SAETOGRAPTUS) Pribyl, 1942, emend. Hutt, 1969
Page 2.2

Type species. Original designation; Graptolithus chimaera Barrande, 1850.
Saetograptus (Saetograptus) chimaera chimaera (Barrande, 1850)
Text-figure 2.2a
1850 Graptolithus chimaera Barrande, p. 52, pi. 4, fig. 34.
1899 Monograptus chimaera', Pemer, p. 39, pi. 17, fig. 15a-b.
1900 Monograptus chimaera’, Wood, p. 471, pi. 25, fig. 18a-d.
1911 Monograptus chimaera; Elies & Wood, p. 398, pi. 39, fig. 3a-d,
text-fig. 266a, b.
1936 Monograptus chimaera; Boucek, p. 17, text-fig. 3a-c.
1942 Pristiograptus (Saetograptus) chimaera chimaera; Pribyl, p. 177, pi. 3,
figs. 1-2.
1955 Monograptus chimaera; Kiihne, p. 372-379, fig. 6.
1956 Pristiograptus (Saetograptus) chimaera chimaera; Tomczyk, p. 55-56, pi.
8, 2, text-fig. 16b.
1958 Saetograptus chimaera; Urbanek, p. 53, text-figs. 26-31, pi. 2, figs. 1-
4, pi. 3, figs. 1-3, text-pl. 2, 3.
1964 Monograptus chimaera; Berry, p. 591, text-fig. 3A, B.
Holotype
NMP L19973, figured Barrande (1850, pl.4, fig.34), refigured Pemer (1899, pi. 17, fig. 15a-
b) from the nilssoni biozone of Zadnf Kopanina, Czech Republic.
Description
The holotype specimen of S. c. chimaera shows no relief and is preserved as compressed
carbonaceous periderm. It is mostly complete, but the sicula is damaged (Fig. 2.2a). The
rhabdosome is straight and relatively large, reaching a length of 12.44 mm, and widening most
significantly over the first five thecae to a maximum width of 1.63 mm at th 10 (see Table 1).
The sicula apex reaches halfway up theca 2. Although their preservation is poor, the first 14
thecae possess paired apertural spines which are discrete and slender, extending from the
Page 2.3

middle to the distal part of the lateral apertural margin. The spines are up to 0.37mm in length.
The 'spine' on theca 1 appears to be a thickened elongation of the whole lateral apertural
margins, rather than a ‘true’ spine, as seen in subsequent thecae. The four most distal thecae do
not appear to possess any spines or processes. Thecal spacing approximately 2TRD 0.9 mm at
thl-3, increasing to 1.6 mm for thlO-12. Full rhabdosome dimensions given in Table 2.1.
Discussion
S. c. chimaera was originally described and figured by Barrande (1850), although he appears to
have used the name for a variety of forms (Pemer 1899). For this reason, Pemer (1899)
emended the diagnosis and designated a type specimen (Fig. 2.3). In his description, Pemer
remarks that the rhabdosome is straight, proximally narrow and up to 2.0 mm wide, with a
thecal spacing of 13 in 10 mm. Thecal morphology displays a free upper theca prolonged into a
convex, pointed spine at about the middle part of the thecal aperture. Below this the aperture is
indented in a ‘half-circle’ shape with an obliquely rounded lower lip which forms a narrow
indentation in the lower free thecal wall. Pemer (1899) distinguished between these and the two
proximal thecae, which he described as hook-like in appearance. He also noted that some distal
thecae had processes extending from the upper part of the thecal aperture. Pemer’s description
of S. c. chimaera corresponds with the one presented in this study, the only exception being the
lack of thecal processes on the five most distal thecae shown here; this might result from
preservational factors, but on balance appears to be an original feature (Fig. 2.3).
There appear to be distinct geographical differences in rhabdosome dimensions between
populations of S. c. chimaera . Other Czech material is like that of the holotype specimen
(Pribyl 1942), while British specimens are generally longer, up to 4.0mm (Wood 1900, Elies
& Wood 1911). Berry (1964) also commented on this variation and noted that North American
specimens were of both varieties, and suggested that longer specimens could result from
preservational factors. Most authors agree with the subtle variations in the thecal morphology of
S. c.chimaera and describe or illustrate its characteristically saetograptid biform thecae (Fig.
2.3) (Wood 1900, Elies & Wood 1911, Boucek 1936, Pribyl 1942, Tomczyk 1956, Kuhne
1955). Berry (1964) recognised a detailed three-fold division of thecal types in this species; he
described, in addition to the standard colonus-type proximal thecae, th 3-6 with spines curving
Page 2.4

outwards and down from a point where the apertural margin and the free ventral wall join, on
the upper part of the aperture. The most distal thecae had shorter spines projecting from a point
close to where the apertures join with the free thecal walls or midway between this join and the
apertural lip.
Urbanek (1958), studied isolated material from Poland (Fig. 2.4). He noted that the
proximal thecae were similar to those of C olonograptus, suggesting that the ontogeny is
analogous to that subgenus, and that the shape of the processes changes during astogeny.
Urbanek also differentiated three types of chimaera thecal morphology. Distal thecae have
shorter, straighter processes, and th 8-10 possess small, blunt, postero-laterally directed
processes. These three thecal forms cannot be distinguished in the type material of S.
c. chimaera. With flattened material, there is a danger that compression may slightly alter the
relationship between thecal spine and apertural margin (Wood 1900, Elies & Wood 1911, this
study). Taking into account compressional factors and possible variation in local populations of
S. c. chimaera, in general, it can be said that the thecae are biform; with proximal having
paired, broad-based extensions of the lateral thecal aperture, which in many cases are prolonged
into a short, sharp point. The change to distal thecal type can be either sudden or gradual,
resulting in a 'step' in the lower apertural margin, with a pair of longer, finer ‘true’ spines,
projecting from the middle to upper part of the apertural margin. In some cases, the most distal
thecae may show a reduction or total loss of this spine.
Accurate sicula measurements for the holotype of S. c. chimaera were not
obtainable as this part of the specimen was badly damaged. However, most authors give a
sicula length from 1.6-2.0 mm (Wood 1900, Berry 1964, Boucek 1936, Pribyl 1942), and a
width from 0.2-0.34 mm (Wood 1900, Berry 1964). The slight proximal rhabdosome
curvature described by several authors (Elies & Wood 1911, Pribyl 1942, Tomczyk 1956,
Urbanek 1958, Berry 1964), appears to be the result of rapid width expansion in the proximal
portion of the rhabdosome. It is worth noting that some of Pribyl's (1942) and Boucek's
(1936) figured specimens may be Saetograptus chimaera semispinosus.
Page 2.5

Saetograptus (Saetograptus) chimaera salweyi (Lapworth, 1880)
Text-figure 2.2c
1880 Monograptus salweyi Lapworth, p. 150, pi. 4, figs. 2a, b.
1900 Monograptus chimaera salweyi (Hopkinson MS); Wood, text-fig. 18, pi.
XXV, figs. 19a-b.
1911 Monograptus chimaera var. salweyi (Hopkinson MS); Elies & Wood, p. 400,
pi. XXXIX, figs. 5a-d, tfs. 267a, b.
1942 Pristiograptus (Saetograptus) chimaera salweyi (Hopkinson MS); Pribyl, p.
175, pi. 1, fig. 1, pi. 3, figs. 5-8.
1956 Pristiograptus (Saetograptus) chimaera salweyi (Hopkinson MS); Tomczyk, p.
117, figs. 16c, d, pi. VIII, fig. 1.
Holotype
SM A24699, refigured Elies & Wood (1911, pi. XXXIX, fig. 5b), from Stormer Hall, near
Leintwardine, Shropshire, England (no longer accessible).
Description
The holotype specimen of Saetograptus chimaera salweyi shows no relief and is preserved as
compressed carbonaceous periderm which is very patchy. The specimen represents only a distal
fragment of the rhabdosome (Fig. 2.2c). As width varies very little and all thecae appear to be
well formed, it is likely that a considerable portion of the rhabdosome is missing, and it is
therefore impossible to take accurate rhabdosome dimensions for this subspecies. With the
exception of a few poorly preserved thecae, most thecae have apertural spines, which extend
from the upper part of the apertural margin, close to the free ventral wall of the proceeding
theca.
Discussion
S. c. salweyi was originally collected and described by Hopkinson (1873); however, it was
Lapworth (1880) who first figured it. Unfortunately, the holotype is only a distal fragment, but
Page 2.6

there are several apparent accounts of this subspecies in the literature. Lapworth's (1880)
original figure displays a biform thecal morphology similar in character to that of S. c.
chimaera, although many authors record the thecal spines as more slender and generally longer
at about 1.0mm (Wood 1900, Elies & Wood 1911, Pribyl 1942, Tomczyk 1956, Urbanek
1958), and measurements obtained from the re-illustration of the holotype show distal thecae
with significantly longer thecal spines (0.73mm compared with 0.4mm for S. c.chimaera).
Wood (1900) described the taxon as ‘a well marked variety of M. chimaera, with the exception
of a shorter, narrower rhabdosome, up to 12.7mm long and a maximum width of 1.6mm,
apparently from topotype material. The smaller rhabdosome dimensions of S. c. salweyi in
comparison with S. c. chimaera can generally be confirmed by other records (Pribyl 1942,
Tomczyk 1956, Urbanek 1958), however, there does appear to be a continuum of
measurements ranging between the two forms, especially with regards rhabdosome length.
This fact has been noted by several authors (Wood 1900, Urbanek, 1958). Indeed,
rhabdosome widths for the holotype specimen of S. c. chimaera (0.88-1.63mm, this study) are
comparable with measurements quoted for supposed material of S. c. salweyi (Wood 1900,
Elies & Wood 1911, Pribyl 1942, Tomczyk 1956). It is possible that the holotype of S. c.
salweyi is in fact an example of S. c. chimaera, but without a complete type specimen this is
impossible to determine. Urbanek's (1958) examination of isolated material (Fig. 2.4) he
assigned to S. c. salweyi, revealed a narrower rhabdosome and longer thecal spines. However,
Urbanek acknowledged that there was a strong case for considerable intraspecific variation, and
possible transitional forms between the chimaera subspecies.
Of Pribyl's (1942) four figured specimens of S. c. salweyi, only one shows particularly
elongate spines, the others differ from his figures of S. c. chimaera only in that the
rhabdosomes are narrower. These specimens may represent part of a continuum of forms
between S. c. chimaera and S. c. salweyi, or local variation.
No sicula dimensions could be obtained from the holotype of S. c. salweyi', however,
Pribyl (1942) records that it is narrow, reaching up to 1.5mm in length with considerable distal
prolongation of the virgula.
Page 2.7

Saetograptus (Saetograptus) chimaera semispinosus (Elies & Wood, 1911)
Text-figure 2.2b
1900 Monograptus chimaera var. a Wood, p. 473, pi. 25, fig. 20.
1911 Monograptus chimaera var. semispinosus Elies & Wood, p. 401, pi. XXXIX,
figs. 4a-c.
1936 Monograptus pseudochimaera’, Boucek, p. 16, pi. 17, fig. 3d-f.
1942 Pristiograptus (Saetograptus) chimaera semispinosus’, Pribyl, p. 177, pi. 3,
figs. 9-10.
1956 Pristiograptus (Saetograptus) chimaera semispinosus', Tomczyk, p. 117, fig.
16a, pi. VII, fig. 4 & pi. VIII, fig. 3.
Holotype
BU 1509, figured Elies & Wood (1911, pl.XXXIX, figs. 4c), from Elton-Ludlow Road,
Shropshire, England.
Description
The holotype specimen displays no relief and is preserved as compressed carbonaceous
periderm which is mostly intact, although the sicula is incomplete (Fig. 2.2b). The rhabdosome
is largely straight, although there is a hint of ventral curvature at the proximal end. Widening of
the rhabdosome occurs most significantly in the first five thecae, measurements for rhabdosome
widths and 2TRD are given in Table 2.1. 25 thecae are visible, although thecal processes are
only a regular feature of the first nine thecae. The spines are about 0.2-0.35mm in length, and
their exact relationship to the apertural margin is difficult to deduce given the overall state of
preservation. In general, spines appear to be projecting from the lower 'lip' of the thecal
aperture, however, th3 and 4 appear to show the standard chimaera-type theca, that is, a spine
projecting from the middle part of the apertural margin. Th 1-2 display a more typical colonus-
type hook-like form, without 'true' spines. The distal, non-spinose thecae are simple, tubular
'pristiograptid-type' thecae.
Page 2.8

Discussion
This subspecies of S. c. chimaera was originally figured and described by Wood (1900), who
noted its unusual resemblance to Saetograptus colonus colonus. She believed that it was
distinguished from S. c. chimaera by a broader rhabdosome (up to 2.5mm) and that only the
proximal thecae possessed spines. These distinctions between the subspecies are also noted by
other authors (Elies & Wood 1911, Pribyl 1942, Tomczyk 1956). Pribyl (1942) recognised the
basic chimaera-type thecae on Czech specimens of S. c. semispinosus, and appears to have
reused Boucek's (1936) figures for Monograptus pseudochimaera.
Rhabdosome widths taken from the holotype specimen of S. c. semispinosus for the
purposes of this study are greater than those of the other S. chimaera subspecies, but not
significantly greater than those of S. c. chimaera (Table 2.1). This, coupled with the
semispinose nature of the rhabdosome of the S. c. chimaera holotype, leaves open the
possibility that it is a senior synonym of S. c. semispinosus. Unfortunately, sicula
measurements for this taxon are not obtainable from the holotype.
Saetograptus (Saetograptus) clunensis (Earp, 1944)
Text-figure 2.2g
1944 Monograptus clunensis Earp, p. 184, text-fig. la-d.
Syntype Specimens
72024 (narrow form) & 72027 (short form ), figured Earp (1944, text-figs. lc & d), from Clun
Forest (Drefor Dingle & Pool Hill), Welsh Borderlands.
Description
Four specimens were figured by Earp (1944) as syntypes. His figures la-b represented the
standard morphological concept of the species, while figures lc-d represented narrow and small
varieties respectively. Unfortunately, the 'standard' syntypes of Saetograptus clunensis have
been lost, leaving only the atypical forms for study (Fig. 2.2g). However, I was able to study
about 25 topotype specimens of S. clunensis from the British Geological Survey collections, so
Page 2.9

a range of values for this population can be given. These specimens display no relief and are
preserved as compressed carbonaceous periderm which is mostly intact but patchy in areas.
Rhabdosome width at th 1 usually varies between 0.57 mm and 0.78 mm, with the largest
recorded width of 0.9 mm. The rhabdosome displays a slight dorsal curvature which usually
occurs around th 1-5. Most thecal spines extend from just above the middle or from the upper
part of the aperture. The spines are slender and relatively long, reaching up to 0.84 mm in
length. Many specimens of S. clunensis show typical biform saetograptid thecae, with a
colonus type th 1, as in the co-type 'short-form' specimen; however, this is not always the
case. Several specimens display chimaera-type thecae from as early as th 1, which may be the
result of compression, or real variations within this local population. This latter postulation may
be correct, as a few specimens show an apparently intermediate form with a colonus-type th 1,
while th 2 is clearly of the chimaera-typQ. Few of the S. clunensis specimens however, have the
often marked curvature of the thecal margin below the protruding spine. The bases of spines
seen in this population are not thickened to the same degree as those observed in the holotype
and recorded population of S. c. chimaera. In most cases, the specimens of S. clunensis appear
to be semispinose. Sicula length varies from 0.9-1.75 mm in this population, (see Table 2.1).
Discussion
S. clunensis has not been described since it was discovered by Earp (1944), who noted the
wide morphological variation within this species, drawing particular attention to changes in
rhabdosome dimensions, the degree of rhabdosome curvature, and thecal morphology. Such
variations correspond with those observed in the population recorded here, suggesting a high
degree of intraspecific variation. Earp related S. clunensis most closely with S. chimaera, but
distinguished S. clunensis by its curved proximal end and the increased length of thecal spines.
Jaeger (1959) suggested that S. clunensis was the saetograptid form most similar to
Saetograptus fritschi linearis, while noting that the sicula of S. clunensis is normal.
Page 2.10

Saetograptus (Saetograptus) wandalensis (Watney & Welch, 1911)
Text-figure 2.2h
1911 Monograptus wandalensis Watney & Welch, p. 235, tf. 4A-C.
1967 Monograptus wandalensis', Rickards, p. 145, tf. 13a, b.
Lectotype
SM A52619, proposed and figured by Rickards (1967, text-fig. 3b), from the nilssoni Biozone
of Wandale Hill, Howgill Fells, England.
Description
The lectotype specimen appears to be complete and is preserved in relief as a pyritic internal
mould (Fig. 2.2h). This specimen appears to be relatively narrow with little widening of the
rhabdosome throughout its length, and reaches 0.85 mm in width at th 10. Full rhabdosome
measurements are given in Table 2.1. The rhabdosome shows slight dorsal curvature which is
most pronounced distally. The first two thecae and one distal theca possess long, slender spines
reaching up to about 0.6 mm in length. The mode of preservation makes it difficult to accurately
assess the spine-aperture relationship, but the thecae appear to become markedly narrower
towards the apertural margin, and the spines seem to extend from the free ventral wall of the
theca. The sicula length is 0.87 mm long and 0.18 mm wide at its base.
Discussion
The description of the lectotype of S. wandalensis given here is consistent overall with the
original outline of the morphological concept by Watney & Welch (1911), and that given by
Rickards (1967), when he proposed the lectotype specimen. However, the original description
did not include the recognition of thecal spines, which were later identified by Rickards (1967).
Both sets of authors record variation in the nature of the dorsal curvature, and this concurs with
my examination of some topotype material. The maximum rhabdosome width and spine length
given here for the lectotype fall somewhat short of figures quoted by other authors (Watney &
Welch 1911, Rickards 1967), however, topotype material confirms that width ranges up to
Page 2.11

about 1.2 mm, and in one specimen, a thecal spine length of 1.0 mm was obtained. As with
previously described saetograptid taxa, there is significant morphological variation between
individuals, although the three-dimensional nature of preservation in S. wandalensis may be
responsible to some degree for such differences.
Saetograptus (Saetograptus) leintwardinensis leintwardinensis (Lapworth, 1880)
Text-figure 2.2d
1873 Monograptus leintwardinensis Hopkinson, p. 520 (nom. nud.)
1880 Monograptus leintwardinensis (Hopkinson MS); Lapworth, p. 149, pi. 4,
fig.la-d.
1900 Monograptus leintwardinensis; Wood, p. 474, text-fig. 19, pi. XXV, fig.
21A,B.
1911 Monograptus leintwardinensis', Elies & Wood, p. 401, pi. XXXIX, fig. 8a-f,
tfs. 268a-c.
1942 Pristiograptus (Saetograptus) leintwardinensis leintwardinensis', Pribyl, p.
178, pi. 3, figs. 11-12.
1956 Pristiograptus (Saetograptus) leintwardinensis leintwardinensis', Tomczyk, p.
118, fig. 17a-c, pi. VIII, fig. 4, 6a, b.
Lectotype
BU 1526, refigured Elies & Wood (1911, pi. XXXIX, fig. 8b), from Church Hill Quarry,
near Leintwardine, Shropshire, England.
Description
The lectotype specimen displays no relief and is preserved as compressed carbonaceous
periderm, which is largely intact (Fig. 2.2d). The rhabdosome is straight with slight proximal
tapering and is relatively short (8.6 mm in length), reaching a maximum width of 1.14 mm at th
10. All thecae are spinose with the exception of the most distal theca, and this may be the result
of preservation. Thecal spines appear to be formed from the free ventral wall of the subsequent
Page 2.12

theca. On theca 1 particularly, the spine appears to be extended from the mid to lower part of
the thecal aperture, however, this is not a colonus-type process, it is a true saetograptid spine.
The sicula is 2.3 mm long, with a basal width of 0.35 mm. (see Table 2.1).
Discussion
S. I leintwardinensis was originally collected by Hopkinson (1873), but was first fully
described and figured by Lapworth (1880). The species has been reported many times since,
and most authors agree on the overall morphological concept of the taxon.
Lapworth (1880) described the spine-aperture structure of S. I. leintwardinensis as
consisting of a deep ‘excavation’ in the free ventral wall, which is prolonged into a horizontal
spine. However, in the figured specimen, the position of the interthecal septum gives the
appearance of the spine extending from the lower edge of the aperture with a ridge in the middle
of the aperture. This thecal structure does not agree with specimens of this species figured by
other authors (Wood 1900, Elies & Wood 1911, Pribyl 1942, Tomczyk 1956, this study).
While an approximate colonus-tyipe th 1 can be seen on some specimens, the process seen in
this species is a spine and not a hook. Some examples show an intermediate chimaera type
thecal stage between the initial theca and the characteristic leintwardinensis-type thecae that
occur distally. One of Wood's (1900) S. I leintwardinensis figures clearly displays paired
spines on some thecae. Pribyl (1942) goes into little detail on thecal structure, giving his
description as after Elies & Wood (1911). Tomczyk (1956) describes leintwardinensis thecae as
‘Monoclimacis-type’, which is clearly a misinterpretation, and may be the result of poor
preservation, although his descriptions are consistent with the information given here.
The sicula characteristics are agreed upon by most authors and concur with those of the
lectotype specimen (Wood 1900, Elies & Wood 1911, Pribyl 1942, this study); however,
Tomczyk’s (1956) Polish material shows slightly shorter siculae (1.8-2.0 mm).
Page 2.13

Saetograptus (Saetograptus) leintwardinensis incipiens (Wood, 1900)
Text-figure 2.2e
1900 Monograptus leintwardinensis var. incipiens Wood, p. 475, pi. 25, figs. 22A,
B.
1911 Monograptus leintwardinensis var. incipiens; Elies & Wood, p. 402, pi.
XXXIX, figs. 9a, b.
1942 Pristiograptus (Saetograptus) leintwardinensis incipiens; Pribyl, p. 179, pi. 3,
fig. 13.
71956 Pristiograptus (Saetograptus) leintwardinensis incipiens; Tomczyk, p. 118,
fig. 17d, e, pi. VIII, fig. 5a, b.
1965 Monograptus leintwardinensis var. incipiens', Cuerda, p. 175, fig. 3.
Lectotype
BU 1529, figured Wood (1900, pi. 25, figs. 22A), from Montgomery Road, Shropshire,
England.
Description
The lectotype specimen displays no relief and is preserved as flattened carbonaceous periderm,
which is largely intact (Fig. 2.2e). The rhabdosome shows significant widening from 0.7 mm
at th 1 up to 1.95 mm at th 15. This subspecies is semispinose, with only th 1-3 possessing
thecal spines, which all appear to of standard leintwardinensis-type morphology. Th 8 may
have an apertural process, the exact nature of which is difficult to distinguish. Many of the
distal thecae have 'up-turned', horizontal apertures. Sicula length is 1.8 mm. (see Table 2.1).
Discussion
Wood (1900) stated that S. leintwardinensis incipiens was distinct from S. I. leintwardinensis
principally by its wider, longer rhabdosome and semispinose thecae, but was clearly a
subspecies due to its spine-aperture relationship and thecal spacing. The description of the
lectotype and rhabdosome dimensions given in this study bear out Wood's original diagnosis.
Page 2.14

Wood (1900) also noted that the two forms are not found together, and suggested that S. I.
incipiens may be a forerunner of S. I. leintwardinensis . Subsequent authors' work concurs
with the findings already mentioned here (Elies & Wood 1911, Pribyl 1942). It is also worth
noting that none of the specimens figured by these authors display colonus-type thecae
proximally, and that in some cases, thecal spines do not appear to originate from the free ventral
wall, but from the upper part of the aperture, which may result from compression. Pribyl
(1942) used Elies & Woods’ figured material, which shows distal thecal apertures almost at
right angles to the rhabdosome.
The specimen figured by Tomczyk (1956) as this species is a questionable identification
of S. /. incipiens. Th 1-2 show colonus-type thecae followed by leintwardinensis-type thecae
which continue up to th 12 before simple, non-spinose thecae occur. It may be possible that this
specimen is a transitional form between S. I. incipiens and S. I. leintwardinensis.
Saetograptus (Saetograptus) leintwardinensis primus (Boucek, 1936)
Text-figure 2.5a-h
1936 Monograptus leintwardinensis var. primus, Boucek, p. 18-19, text-fig. 3g-k,
pi. II, fig. 3.
1942 Pristiograptus leintwardinensis primus', Munch, p. 255, pi. IV, fig. 11-12, pi.
VII, figs. 8-9.
1942 Pristiograptus (Saetograptus) leintwardinensis primus', Pribyl, p. 179, pi. 3,
figs. 19-21.
Holotype
The specimen is missing from the National Museum, Prague. Figured, Boucek (1936, Fig. 3 g
& h) from the nilssoni and scanicus biozones of Reporyje (Muslovka), Czech Republic.
Discussion
This species is not widely recognised. It was first described and illustrated by Boucek (1936),
who stated that S. leintwardinensis primus was distinguishable as a subspecies of S. I.
Page 2.15

leintwardinensis due to its larger rhabdosome dimensions (Fig.2.5). Boucek (1936) described a
long, distally straight rhabdosome, with characteristic dorsal curvature of the sicula region. He
noted a gradual width increase of up to 1.6-1.8 mm. The thecae are long and slightly flexed,
with spines about 0.6-0.8 mm in length. The apertural spines are of standard leintwardinensis-
type. The sicula is about 1.0mm long. This subspecies also appears to be semispinose. Pribyl's
(1942) description of S. I. primus agrees with that of Boucek (1936), as they appear to have
used the same figures. However, Pribyl (1942) remarked that proximal thecae have a weak s-
shaped curvature with spines slopping diagonally downwards.
All of the figures published by Boucek (1936) and Pribyl (1942) are poor; only one
specimen actually shows the marked sicula curvature of the description. In most other respects,
the material referred to by these authors is very similar to specimens of S. I incipiens. What
appear to be chimaera-type thecae can be seen proximally in the figured specimens but this is
not clear from the illustrations. It is not possible to assess the morphological concept of S. I
primus without studying the holotype.
Saetograptus (Saetograptus) fritschi fritschi (Pemer, 1899)
Text-figure 2.2f
1899 Monograptus clavulus Pemer, p. 24, pi. 17, fig. 15.
1936 Monograptus fritschi', Boucek, p. 19, text-fig. 31-n, pi. II, fig. 1.
1942 Pristiograptus (Saetograptus) fritschi fritschi', Pribyl, p. 180, pi. 3, fig. 14-16.
Holotype
PNM L17764, figured Pemer (1899, pi. 17, fig. 15), from Zadnf Kopanina, Czech Republic.
Description
The holotype specimen shows no relief and is preserved as compressed carbonaceous
periderm, which is patchy (Fig. 2.2f). The rhabdosome displays strong dorsal curvature,
particularly in the proximal portion. Rhabdosome width increases gradually from 0.83 mm up
to 1.15 mm at th 10. Some of the thecae are damaged; otherwise, all thecae are spinose, with
Page 2.16

the spine projecting from approximately the middle part of the apertural margin. The spines
vary from 0.35-0.6 mm in length. Th 1 seems to be of colonus-type form, with slight extension
of the lower part of the apertural margin. The base of the sicula is damaged so accurate
dimensions cannot be obtained from the holotype. (see Table 2.1).
Discussion
Although Boucek (1936) figured S. fritschi fritschi, he did not describe it, instead he referred
the reader to Pemer's (1899) illustration and description of M. clavulus, which he regarded as
representing S. fritschi fritschi. Pribyl's (1942) account of Czech specimens of this species
concurs with the description and rhabdosome dimensions given here for the holotype specimen.
Saetograptus (Saetograptus) fritschi linearis (Boucek, 1936)
Text-figure 2.5
1936 Monograptus fritschi mut. linearis Boucek, p. 20, fig. 3u.
1942 Pristiograptus (Saetograptus) fritschi linearis', Pribyl, pi.3, fig. 17-18.
1992 Saetograptus fritschi linearis', Lenz, p. 207, fig. 3s-u.
Type Specimen
The specimen of S. fritschi linearis illustated by Boucek (1936, Fig. 3u) as typical of the
subspecies is missing from the National Museum, Prague. It was obtained from Reporyje,
Czech Republic.
Discussion
Boucek (1936) did not give a full description of this subspecies, although he did provide
illustrations of material (Fig. 2.5). Pribyl (1942) figured two of Boucek's specimens, including
the 'typical form1. By way of a description, he merely remarked that the morphological concept
of S. fritschi linearis was the same as that of S. fritschi fritschi, except that the rhabdosome of
S. f linearis was shorter. Study of both sets of illustrations shows this description to be
incorrect. These drawings show no marked dorsal curvature of the rhabdosome, and in some
Page 2.17

cases, the thecae have a leintwardinensis-typ& appearance. However, isolated specimens
identified by Lenz (1992) from Cornwallis Island in Arctic Canada as S. f linearis, show
considerable morphological variations in the sicular area and of the spines, from S. fritschi
fritschi. Such differences were not described by Boucek (1936) or Pribyl (1942). Lenz (1992)
recorded a characteristic flaring sicular aperture and occasional bifurcation of thecal spines, (see
Table 2.1).
Subgenus SAETOGRAPTUS (COLONOGRAPTUS) Pribyl, 1942
Type species. Original designation; Graptolithus colonus Barrande, 1850.
Saetograptus (Colonograptus) colonus colonus (Barrande, 1850)
Text-figure 2.2i
1850 Graptolithus colonus Barrande, p. 42, pi. 2, figs. 2, 3.
1883 Monograptus colonus; Tullberg, pi. I, figs. 22-23.
1897 Pristiograptus colonus; Freeh, p. 655-656, fig. 209.
1899 Monograptus colonus', Pemer, p. 18-19, pi. XIV, figs. 3, 12, text-fig. 12.
1900 Monograptus colonus', Wood, p. 463, pi. XXV, fig. 10A-D, text-fig. 14.
1911 Monograptus colonus; Elies & Wood, p. 391, pi. XXXVIII, fig. 8a-d, tfs.
260a-c.
1920 Monograptus colonus, Gortani; p. 33-34, pi. II, figs. 28-29.
1936 Monograptus colonus; Boucek, p. 13, fig. 2a-e.
1942 Pristiograptus (Colonograptus) colonus colonus; Pribyl, p. 166, pi. 2, figs.l-
3.
1955 Monograptus colonus; Kiihne, p. 370-372, fig. 5A-B.
1958 Colonograptus colonus; Urbanek, p. 50, text-figs. 23-25, pi. I, figs. 4, 5.
1962 Monograptus colonus;Ross, p. 71, text-figs. 3C, H, J; 5A, B, F.
Page 2.18

Lectotype
PNM L19977, figured Barrande (1850, pi. 2, figs. 1-5), refigured Pemer (1899, pi. XIV,
figs. 3, 12, text-fig. 12), from Butovice, Czech Republic.
Description
The lectotype specimen is well preserved as a carbonaceous film, it displays partial relief, and
the periderm is intact (Fig. 2.2i). The rhabdosome is long and straight, widening very gradually
throughout much of its length, from 0.85 mm at th 1 to 1.5 mm at th 30. The most proximal
thecae (th 1-5) display typical Colonograptus-type apertural processes, that is, lip/hook-like
extensions of the lower part of the apertural margin, but not a tue Saetograptus-typQ spine. In
some thecae there is a curvature of the free ventral wall, but otherwise, the thecae without
processes are of simple, pristiograptid-type form. The sicula is complete, 1.43 mm in length
and 0.28 mm wide at the base, with prominent sicular spines. The lectotype has a total of 44
thecae. (see Table 2.2).
Discussion
According to Pemer (1899), Barrande used the name M. colonus for several entirely different
species; therefore, on the basis of his and others' work, as well as having studied Barrande’s
original specimens, he emended the diagnosis and designated a type specimen. All subsequent
workers have based their studies upon this species concept (Fig. 2.6). Perner's (1899)
description agrees with that presented in this re-examination of the lectotype specimen, the
principal exception being the maximum width values, which Pemer quoted as up to 2.5 mm,
which in the lectotype is 1.5 mm; however, Pemer took a range of values from topotype
material. The illustrations and descriptions of other authors (Lapworth 1880, Wood 1900, Elies
& Wood 1911, Ross 1962, Urbanek 1958), concur with Perner's (1899) diagnosis and the
description of the lectotype presented here (Fig. 2.6).
Page 2.19

Saetograptus (Colonograptus) colonus compactus (Wood, 1900)
Text-figure 2.2k
1900 Monograptus colonus var. compactus Wood, p. 466, pi. 25, fig. 12.
1911 Monograptus colonus var. compactus; Elies & Wood, p. 393, pi. XXXVIII,
fig. lOa-c, tf. 261.
1942 Pristiograptus (Colonograptus) colonus compactus', Pribyl, p. 167.
1964 Monograptus colonus var. compactus', Berry, p. 592, tfs. 2C, D.
1972 Saetograptus colonus compactus', Lenz, p. 1153, fig. 2M, N, O, P.
Holotype
BU 1498, figured Wood (1900, pi. 25, fig. 12), from Elton-Evenhay Lane, Shropshire,
England.
Description
The holotype specimen shows no relief and is preserved as compressed carbonaceous
periderm, which is patchy (Fig. 2.2k). The proximal end is very badly damaged. The
rhabdosome appears to be straight, but tapers at both ends with proximal tapering most marked.
The most proximal portion of the rhabdosome is missing; however, the first visible theca is 0.8
mm wide, with the maximum width of 2.05 mm at about thlO. The initial thecae are missing
from the holotype so it is not possible to comment on the existence of any apertural processes,
the distal thecae are of a simple, tube-like form. No sicular dimensions were obtainable, (see
Table 2.2).
Discussion
Wood (1900) stated that S. colonus compactus could be differentiated from S. c. colonus by
its curved rhabdosome, more closely spaced thecae and by tapering at both ends. These
differences were reiterated by Elies & Wood (1911), who added that the rhabdosome of S.
colonus compactus was shorter. However, with the exception of proximal and distal tapering,
none of these distinguishing features can be seen in the holotype specimen, whose rhabdosome
Page 2.20

dimensions appear to be consistent with those recorded for S. c. colonus here and elsewhere. It
is uncertain that the holotype of S. colonus compactus belongs within the subgenus
Colonograptus, due to the lack of proximal thecae. Subsequent identifications of S. colonus
compactus (Berry 1964, Lenz 1972), outlining features and rhabdosome dimensions consistent
with those given by Elies & Wood (1911), are similar to rhabdosome dimensions for the
holotype of S. colonus compactus and the lectotype of S. c. colonus. It is clear that the type
specimen of S. c. compactus is of little use in defining the morphological concept of this
subspecies.
Saetograptus (Colonograptus) roemeri (Barrande, 1850)
Text-figure 2.21
1850 Graptolithus Roemeri Barrande, p.41, pi. 2, figs. 9-11.
1899 Monograptus roemeri; Pemer, p. 16-17, pi. XIV, figs. 1, 10, 18, 24, tfs. 11.
1900 Monograptus roemeri’, Wood, p.470-471, tfs. 17, pi. XXV, fig. 13A-B.
1911 Monograptus Roemeri', Elies & Wood, p. 397, pi. XXXIX, fig. 2a-d, tfs.
265a, b.
1936 Monograptus roemeri’, Boucek, p. 15, pi. II, figs. 4-6, tfs. 2j, k.
1942 Pristiograptus (Colonograptus) roemeri’, Pribyl, p. 170, pi. 2, figs. 12-13; pi.
1, fig- 2.
1962 Monograptus roemeri’, Ross, p. 65, text-fig. 3K.
Paralectotype
PNM L17761, figured Barrande (1850, pi. 2, fig. 9), from Butovice, Czech Republic.
Description
The paralectotype is well preserved as a carbonaceous film, showing partial relief with much
periderm intact (Fig. 2.21). The rhabdosome is long with a total of 57 thecae, and has a slight
ventral curvature at the proximal end, which becomes subtly dorsal distally. Maximum width of
Page 2.21

the specimen is achieved at th30 (3.1 mm), proximal width is 0.75 mm. The rhabdosome
expands rapidly initially. The sicula is incomplete and a large portion appears to be missing,
making measurements impossible to obtain. No thecal spines or processes can be seen in this
specimen, although in some cases, the free ventral walls of the thecae are curved, and the thecae
appear to be simple tubes, (see Table 2.2).
Discussion
It is not possible to identify this specimen as a saetograptid from the paralectotype (Fig. 2.21).
However, with the exception of the proximal details, the descriptions and rhabdosomal
dimensions quoted by other authors (Barrande 1850, Wood 1900, Elies & Wood 1911,
Boucek 1936, Pribyl 1942, Ross 1962) are compatible with those given here for the
paralectotype.
Saetograptus (Colonograptus) varians varians (Wood, 1900)
Text-figure 2.2j
1900 Monograptus varians Wood, p. 467, pi. 25, fig. 14A, B, tf. 15.
1911 Monograptus varians', Elies & Wood, p. 395, pi. XXXIX, fig. 6a-e, tf. 264a-
c.
1942 Pristiograptus (Colonograptus) varians varians', Pribyl, p. 268, pi. 2, figs.
10- 11.
1962 Monograptus varians', Ross, p. 67, text-figs. 3B, G; 4A.
1964 Monograptus varians', Berry, p. 597, text-fig. 2F.
1969 Saetograptus varians', Hutt, p. 363, figs. 1-4.
1972 Saetograptus varians’, Lenz, p. 1158, fig. 3F, L.
1986 Saetograptus varians, Palmer, p.257, fig. 6.
Lectotype
BU 1511, Elies & Wood (1911, pi. XXXIX, Fig. 6a) from Old Dingle Mill, Long Mountain,
Welsh Borderlands. The lectotype specimen is missing.
Page 2.22

Discussion
S. varians varians is a widely recorded subspecies. Wood's (1900) original description of the
taxon outlined it as related to S. c. colonus by its typical Colonograptus-type th 1, but that it
differed by having a shorter, narrower rhabdosome, a smaller thecal overlap, an excavation at
the base of the free thecal wall and distal prolongation of the virgula. She also noted that th 2-3
had distinct apertural spines. However, examination of Palmer's (1986) illustration of the
lectotype clearly shows no spine on th 3, although th 1-2 do possess apertural processes; distal
thecae are simple and tubular. The 'excavation' in the base of the free thecal wall is visible from
the lectotype, but is not seen in all thecae, and this may be the result of preservation, for
although it can be seen in the part specimen, it is not visible from the counterpart. The
rhabdosome dimensions given by Wood (1900) for S. v. varians are equivalent to those of the
lectotype of S. c. colonus, although the lectotype of S. v. varians is indeed shorter than that of
S. c. colonus. Several topotype specimens of S. v. varians illustrated by Palmer (1986) show
th 2 with a distinct Saetograptus-type spine (Fig. 2.2j). Hutt (1969), in her emended diagnosis
of the genus Saetograptus, which was based primarily on her study of S. v. varians, extended
the definition to include both subgenera, Saetograptus and Colonograptus, and indeed, S. v.
varians appears to be an intermediary form, (see Table 2.2).
Saetograptus (Colonograptus) varians pumilus (Wood, 1900)
Text-figure 2.2m
1900 Monograptus varians var. pumilus Wood, p. 469, pi. 25, fig. 17A, B.
1911 Monograptus varians var. pumilus', Elies & Wood, p. 396, pi. XXXIX, fig. 6a.
1942 Pristiograptus (Colonograptus) varians pumilus', Pribyl, p. 170, pi. II, fig. 9.
Lectotype
Figured, Elies & Wood (1911, pi. XXXIX, Fig. 7a), from Elton Lane, near Ludlow,
Shropshire, England. The lectotype specimen is missing.
Page 2.23

Discussion
Although Wood (1900) and Elies & Wood (1911) recorded local variations in populations of S.
v. varians, they described a distinct subspecies on the basis of its smaller rhabdosome (Fig.
2.2m), slight proximal curvature, longer sicula and more closely spaced thecae, named S.
varians pumilis. Unfortunately, without viewing the lectotype specimen it is impossible to
provide a detailed comparison of the type material of the two subspecies here, (see Table 2.2).
General Taxonomic Discussion
The subgeneric holotype specimens, Saetograptus chimaera chimaera and Saetograptus
(Colonograptus) colonus colonus, were first describeded by Barrande in 1850, and the latest
major revision of the group as a whole was carried out by Pribyl (1942), when he designated
the genus Saetograptus and divided it into two subgenera, Colonograptus and Saetograptus.
The generic diagnosis was subsequently emended by Hutt (1969), but this was based on her
work on S. varians only. The purpose of this study has been to assess the taxonomic validity
and usefulness of the type material of the species and subspecies of the genus Saetograptus,
and to compare these findings with identifications and descriptions of populations given by
other authors, in an attempt to analyse how the morphological concepts of these taxa have come
about.
Many saetograptid species and subspecies have been widely recorded, but it is generally
accepted that considerable intraspecific variation occurs within the group as a whole (Wood
1900, Elies & Wood 1911, Boucek 1936, Berry 1964, Urbanek 1958, this study). Such
variation creates difficulty in defining clear morphological outlines for accurate taxonomic
identification. This fact is not ameliorated by the often dubious status of saetograptid type
material. The type specimen of Saetograptus chimaera chimaera is of great importance (Fig.
2.2a), as it is also the type for the sub-genus Saetograptus (Pribyl 1942). As a species it is
defined by its distal thecae which have slightly concave apertures, with spines extending from
the middle to upper part of the apertural margins, the initial thecae being of Colonograptus-type.
There are slight variations in rhabdosome dimensions from one population to another, as
shown by other authors (Wood 1900, Berry 1964, Boucek 1936). However, maximum width
is a diagnostic feature, and this varies widely between holotype and recorded accounts, 1.63
Page 2.24

mm for the holotype, and approx. 2.0 mm for recorded specimens. These greater widths
approach those of the broader sub-species S. chimaera semispinosus (Fig. 2.2b) from the UK
Elies & Wood 1911). Such differences, may indicate continuous variation between these two
morphologies, placing doubt on the validity of the sub-species. This is an example of
‘taxonomic-drift’, where workers have relied on the descriptions of others in order to identify
species, rather than the type specimen, with species characteristics becoming distorted in the
literature. When comparing S. chimaera chimaera to another British sub-species, S chimaera
salweyi, another problem in graptolite taxonomy is highlighted, poor preservation of type
specimens (Fig. 2.2c). The type of salweyi is only a distal fragment, though this sub-species is
traditionally distinguished from S. chimaera chimaera by a narrower rhabdosome and longer
spines. Distal measurements taken in this study show that for the type specimens, S. c. salweyi
is not narrower than S. c. chimaera, nor does it have significantly longer spines. In the absence
of a proximal end, it is possible that the type specimen for S. chimaera salweyi is in fact a
specimen of S. chimaera chimaera, making it a junior synonym of that taxon. If further work
shows this to be the case, the S. c. salweyi 'concept' will need redefinition, assuming that
systematic differences remain between 'S. c. salweyi' and S. c. chimaera. Unfortunately,
collecting topotype material of S. c. salweyi is no longer possible, as the original locality has
been destroyed due to road widening. On present evidence, the suspicion is that S. chimaera
salweyi might be synonymous with S. chimaera chimaera.
S. fritschi fritschi has characteristic chimaera -type thecae, but is narrower (maximum
width 1.2 mm) and generally believed to be shorter than S. c. chimaera. However, given that
such width and length differences are used to define subspecies within the chimaera group, and
that it is thecal variation that denotes saetograptid species, it appears that the only significant
distinguishing feature of S. fritschi fritschi is its marked rhabdosome dorsal curvature (Fig.
2.2f). It is uncertain whether this is sufficient difference to justify S. fritschi fritschi being a
separate species. However, temporal variation between these forms may be enough to justify
their classification. Another designated British species, Saetograptus clunensis, is also very
similar to S. fritschi fritschi in morphology, but is less curved; these differences might be
ascribed to intraspecific variation. Unfortunately, the type specimen of Saetograptus fritschi
Page 2.25

linearis is missing and so these two subspecies cannot be compared directly using the type
material.
S. colonus and S. roemeri are examples of the subgenus Colonograptus. The type
specimens show S. roemeri to be wider and slightly longer than S. colonus, but it is impossible
to see the proximal thecae of S. roemeri clearly due to poor preservation (Fig. 2.2i, 2.21).
Without evidence of colonograptid initial thecae, distinguishing this specimen as a saetograptid
rather than a pristiograptid is optimistic.
The single most important distinction between species of saetograptids is the nature of
the theca/apertural spine relationship; this creates a narrow margin of distinction, especially in a
group of graptoloids displaying intraspecific variation. However, analysis of type material and
figured illustrations clearly shows that there are three gross morphological differences in
theca/spine structure within the group as a whole: (i) The chimaera-type theca, in which the
thecal spine extends from the middle or upper part of the thecal aperture (Fig. 2.2a). (ii) The
leintwardinensis-type theca, where the thecal spine projects from the upper part of the aperture
and free ventral wall of the following theca (Fig. 2.2d). (iii) The colonus-type theca, in which
the lower part of the apertural margin is extended into a lip-like process rather than a true spine
(Fig. 2.2i). In general, the group is biform, with all species possessing proximal colonus-type
thecae and distal chimaera-typdleintwardinensis-type thecae, or simple thecae without any
spines or processes. However, there appears to be a continuum of forms between even these
morphologies. For example, isolated material of S. v. varians figured by Hutt (1969), shows
some specimens with no colonus-type thecae at all, indicating a possible link between the two
subgenera. Also, some figured specimens of leintwardinensis subspecies possess all three
types of theca/spine morphology. Given this considerable variation between basic
morphologies and the problems encountered with the type material, there is a strong case to
suggest that the saetograptids have been 'over split' taxonomically. This appears to be
particularly prevalent at the subspecies level, where subtle differences in rhabdosome
dimensions and degree of rhabdosome curvature have been the deciding factors between taxa.
On the basis of this taxonomic reassessment, I would make the following conclusions:
Page 2.26

1) S. chimaera semispinosus and S. chimaera salweyi cannot be clearly differentiated as
subspecies of S. chimaera chimaera and must, therefore, be included within it.
2) S. wandalensis and the subspecies of S. leintwardinensis are taxonomically valid.
3) S. fritschi fritschi is a subspecies of S. chimaera chimaera and also includes S. clunensis.
4) S. colonus compactus cannot be clearly differentiated as a subspecies of S. colonus colonus
and must be included within it.
5) S. roemeri, S. varians varians and S. v. pumilis are taxonomically valid.
6) I have been unable to recognise S. fritschi linearis.
Page 2.27

Lateral view
Distal End
Nema
Interthecal Septum
Thecal Spines
Free Ventral Wall
Thecal Aperture
Sicula Theca
VirgellaProximal End
Ventral view
B
Fig. 2.1. A sketch diagram of a typical saetograptid graptolite.
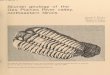
Fig. 2.1

h
m1
Fig. 2.2. Saetograptid type specimens, a. S. chimaera chimaera (L19973) x6.5. b. S. c. semispinosus (BU1509) x5.7. c. S. c. salweyi (A24699) approx. x6.5. d. S. leintwardinensis leintwardinensis (BU1526) x6.7. e. S. I. incipiens (BU1529) x5. f. S. fritschi fritschi (L17764) x7. g. S. clunensis (72024 & 72027) x5. h. S. wandalensis (A52619) xlO. i. S. colonus colonus (L19977) proximal portion x8. j. S. varians varians (type missing) x8. k. S. colonus compactus (BU1498) x 8 .1. S. roemeri (L17761) x6.7. m. S. varians pumilis x5 (topotype).
Fig. 2.2

Fig. 2.3
Fig. 2.3. Published illustrations of Saetograptus chimaera chimaera. a-c. After Boucek (1936) all x5. d-f. After Pribyl (1942) all x5. g. After Barrande (1850) approx. x5. h-i. After Elies & Wood (1911) all x5. j-k. After Berry (1964) all x2.5.1-m. After Pemer (1899) x4 and x5 resepectively.

3 Fig. 2.4. Drawings of plates of isolated Saetograptids from Urbanek, 1958. (a-d) Saetograptus chimaera chimaera andto (e-f) Saetograptus chimaera salweyi, lateral and ventral view respectively, a, xl5; b & d, x l2 , c, xlO, e & f, x l7 .4^

Fig. 2.5
c
Fig. 2.5. Published illustrations of Saetograptus leintwardinensis primus, a-e. After Boucek (1936) all x5. f-h. After Pribyl (1942) all x5. Saetograptus fritschi linearis, j-1. After Boucek (1936) all x5.

Fig. 2.6
Fig. 2.6. Published illustrations of Saetograptus colonus colonus. a-c. After Urbanek (1958) a & c. xlO; b. xl3 . d-h. After Boucek (1936) all x5. i-j. After Barrande (1850). k. After Pemer (1899) x7. m. After Elies & Wood (1911) x 5 .1. After Wood (1900) x9.

Species/Subspecies TypeMaterial
SiculaLength
SiculaWidth(base)
ProximalWidth
Width Theca 3
Width Theca 5
Width Theca 10
Width Theca 20
2TRD1-3
2TRD3-5
2TRD5-7
2TRD10-12
ProcessType
ProcessLength
No. of Processes
S. chimaera chimaera Holotype ~ 1.4 ? 0.88 1.03 1.35 1.63 ----- 0.9 1.0 1.05 1.6
Spines extend from middle/ upper aperture
thl 0.19 thlO 0.45
13
S. chimaera salweyiHolotype(distalfragment)
----- ----- ----- ----- ------ ------DistalWidth1.8
------ ------ -----Distal2TRD1.7
Spines extend from middle/ upper aperture
Distal th 0.6 6 visible
S. chimaera semispinosus Holotype ? 7 0.83 1.25 1.35 1.75 1.9 1.05 1.0 1.35 1.25Spines extend from lower aperture
thl 0.45 th4 0.45 8
S. clunensis TopotypeSpecimen 1.3 0.38 0.7 0.83 1.0 1.13 1.1 0.9 1.1 1.25 1.38
Spines extend from middle of aperture
thl 0.36 th6 0.52 14
S. wandalensis Lectotype 0.87 0.18 0.57 0.63 0.6 0.85 ----- 0.89 0.98 0.87 -----
Spines Possibly extend from lower aperture
thl 0.24 th9 0.38 3 visible
S. leintwardinensisleintwardinensis Lectotype 2.3 0.35 0.77 0.9 1.22 1.14 ----- 1.02 1.13 1.08 -----
Spines extend from upperaperture/free ventral wall
thl 0.3 th5 0.54 9
S. leintwardinensis incipiens Lectotype 1.8 0.7 1.2 1.65 ----- 1.0
Spines extend from upperaperture/free ventral wall
Possibly 3 visible
S. leintwardinensis primus HolotypeMissing
S. fritschi fritschi Holotype - 1.1 -0.25 0.83 0.85 0.95 1.15 ----- 1.05 1.4 1.1 1.2Spines extend from middle of aperture
th3 0.2 th8 0.35 11
S. fritschi linearis HolotypeMissing
Table 2.1. Available rhabdosome dimensions (in mm) for the type material of the subgenus Saetograptus (Saetograptus) Pribyl, 1942.

Species/Subspecies TypeMaterial
SiculaLength
SiculaWidth(base)
ProximalWidth
Width Theca 3
Width Theca 5
Width Theca 10
Width Theca 20
2TRD1-3
2TRD3-5
2TRD5-7
2TRD10-12
ProcessType
ProcessLength
No. of Processes
Hooked
S. colonus colonus Lectotype 1.43 0.28 0.85 1.05 1.3 1.5 1.5(th30)
1.1 1.18 1.35 1.6extension of lower thecal aperture
0.15 5
S. colonus compactus Holotype ? ? 0.8 1.2 1.3 2.05 1.85
2.45
1.0 1.05 1.35 1.4
1.45
----- ----- 0
S. roemeri Paralectotype ? ? 0.75 1.2 1.5 2.05 th303.1
1.2 1.25 1.45 th20-221.55
0
S. varians variansLectotypeMissing
S. varians pumilis LectotypeMissing
Table 2.2. Available rhabdosome dimensions (in mm) for the type material of the subgenus Saetograptus (Colonograptus) Pribyl, 1942.

A global biostratigraphy of British Ordovician and Silurian
graptoloids
Outcrops of British Lower Palaeozoic (Ordovician and Silurian) strata principally occur in
regions considered to represent closing sedimentary basins on the margins of the Palaeozoic
Iapetus Ocean: the Southern Uplands of Scotland, the Lake District of northern England and
Wales and the Welsh Borderland (Fig. 1.1). The rocks are commonly highly graptolitic, and
this abundance, accompanied by a rapid rate of evolution among the graptoloids, is the key to
Ordovician and Silurian biostratigraphy, which is fundamentally based on the stratigraphic
ranges of the component taxa. This is an important aid, on the fine scale, to the
chronostratigraphic schemes of these geological systems, and has shed light on the complex
sedimentological and structural histories of these regions (Figs. 3.1, 3.2). This study reviews
UK graptoloid biostratigraphy, taking into account the large amount of recently published work,
and produces new biostratigraphic range charts, based on the newly revised British graptoloid
biostratigraphic scheme.
Biostratigraphic Method
The earliest studies of graptoloids were concerned with the description, naming and eventual
classification of individual taxa within the order. As more taxa were identified it was quickly
recognised that they had stratigraphic potential due to their diversity and abundance. With the
recognition that certain graptoloids occur at particular levels within the stratigraphic record, it
became possible to date one sequence of rocks relative to another on a fine scale. This led to a
series of publications detailing the occurrence of graptoloid species relative to particular
stratigraphic horizons, which allowed detailed biozonations and the regional and global
correlation of sedimentary sections. However, such biostratigraphy is dependent on the
accuracy of palaeontological identification and the existence of reliable reference sections.
Taxonomic Procedure
Every biological species has a type specimen or specimens which bears the species name and
upon which the morphological characteristics defining the species are based. Reliable
Page 3.1

identifications to species level need to be made essentially by reference to the type material.
Successive descriptions of specimens or populations assigned to a particular taxon, which are
made without reference to type material, may eventually, if unintentionally, modify the species
concept, introducing inaccuracy into both taxonomic and biostratigraphic studies. This may also
result from the failure to illustrate a specimen adequately. Such ‘taxonomic drift’ is exacerbated
by inadequate type material. For example, during their work on the Wenlock biostratigraphy of
Builth Wells, Williams & Zalasiewicz (in press) discovered that the type specimens of
Cyrtograptus ellesae, the zone fossil for the ellesae Biozone, were incomplete; collection of
more complete specimens showed that the pre-cladial (pre-branching) part of this graptolite is
approximately twice as long as previously supposed. The old, mistaken concept indicates little
difference between Cyrtograptus ellesae and Cyrtograptus perneri, whereas the revised concept
shows clear, striking differences between the two taxa. Similarly, they recognised that the type
specimen of Cyrtograptus linnarssoni, another zone fossil, could be a deformed example of
Cyrtograptus rigidus. As these cyrtograptids were crucial in defining Wenlock biozones, the
resolution of such problems resulted in a significant revision of Wenlock biostratigraphy
(Williams & Zalasiewicz in press).
Stratigraphic Perspective
Biostratigraphy is a tool used in the development and employment of the chronostratigraphic
framework, and the fundamental basis of biostratigraphy is the biozone. According to the
‘International subcommission on stratigraphic terminology’ (Hedberg et al. 1961), the formal
definition of a biostratigraphic zone is, “a body of strata characterised by the occurrence of a
taxonomic form or forms from one or more of which it receives its name”. They go on to
recognise that, “the simple term zone is not very significant as a formal biostratigraphic unit
since it is used indiscriminately for the concepts of assemblage and range and does not permit
distinction between the two”. Strictly then, biozone is an informal term, and when possible, a
more exact terminology is advisable (Hedberg et al. 1961, Harland et al. 1972). Consequently,
the term biozone encompasses a variety of zonal definitions, but in graptolite biostratigraphy, is
commonly taken to mean a zone defined by its faunal assemblage, the base of which is
delineated arbitrarily by the incoming of one or more species (Rickards 1994). As Rickards
Page 3.2

(1994) notes, they are often referred to by graptolite biostratigraphers as assemblage zones,
while combining the more subtle definitions of assemblage zone (defined purely by faunal
assemblage) and Oppel zone (defined by faunal assemblage and incoming species), although in
practice these two are virtually the same. This zonal concept is perhaps the one most frequently
used in biostratigraphic studies. In contrast to this view, some workers do define and use
graptolite biozones more precisely, arguing that a more accurate correlation may result. In
compiling his range charts for the Llandovery and Wenlock of Bohemia, Storch (1994) for
example, uses a variety of different types of biozones with subtle differences in definition.
These include the taxon-range zone, defined by the complete vertical and geographical range of
the zonal indicator species, and partial-range zone, representing the period between the
incoming of the zonal indicator species and that of the following zone (Harland et al. 1972).
The interval zone (Storch 1994) appears to fulfil a similar function to the interregnum (Rickards
1994), that is an interval occurring between the disappearance of one indicator species and the
appearance of the next one; it is usually characterised by the presence of long-ranging species,
and the upper and lower limits are necessarily defined by the recognition of the previous and
subsequent biozones. Storch uses what he describes as a couple-range zone to define his
acuminatus-ascensus Zone at the base of the Silurian, that is, the joining of two taxon-range
zones if the overlap between index species ranges is significant. Using biozones based on
slightly different criteria may be of benefit to local biostratigraphic problems. However, on a
larger scale, it may be preferable to take a more uniform approach to the definition and use of
biostratigraphic zones. Any attempt to compare the biostratigraphic sequence in one area with
that in another region when the respective datasets have been analysed in different ways,
introduces inaccuracy into the correlation. Given that geographic variations occur in the
composition of graptolite assemblages, and that graptolites may have different ranges in
different areas, a system of broad-scale graptolite zones (standard reference zones) has been
suggested for global correlation (Koren 1984). Rickards (1994) notes that such combined zones
do not aid the refinement of graptolite biostratigraphic correlation, though they may help
establish chronostratigraphic frameworks.
Although anchored to reference sections, biozones are not represented by formal basal
stratotype sections, in contrast to chronostratigraphic units. Nevertheless, many biozones have
Page 3.3

informal ‘type’ sections where they were initially described. As with basal stratotypes, biozonal
reference sections should ideally be well exposed, tectonically undisturbed, and be in
continuous and conformable graptolitic successions, with an abundant and diverse fossil
component. Often this is not the case. For example, the refinement of the Spirograptus
turriculatus Biozone into six subzones, as outlined by Loydell (1991), was based on sections in
western mid-Wales, an area where strata are often poorly exposed, strongly tectonised, and
intermittently unfossiliferous. Despite this, the subdivision was found to be largely effective in
practice (Davies etal. 1997).
In the course of their work, graptolite biostratigraphers examine a number of sections
within a given area, noting the occurrence of taxa within lithostratigraphical successions, from
which they may produce detailed faunal range charts, assigning particular assemblages of
graptolites observed to their corresponding biozones. Lateral variation is common, especially in
larger-scale correlations (Rickards 1994). Not all correlations are effected using biozones. For
example, a fine-scale global correlation using graptolites was achieved for the early Ordovician
by Cooper & Lindholm (1990), who used the ranges of individual species and species groups,
largely taken from measured sections, to define and correlate the number of 'bioevents', that is
first and last appearance of taxa. These 'bioevents' were statistically tested for diachroneity, and
show a highly refined global graptolite biostratigraphic correlation. The authors (Cooper &
Lindholm 1990) reinforce the need for accurate taxonomy in graptolite biostratigraphy, citing
dubious taxonomy as one of the main problems affecting the reliability of their study.
British Graptoloid Biostratigraphy: Previous Work
Lapworth (1878) first applied the concept of graptolite faunal zones in Britain, during his work
on the ‘Moffat Series’ in the Southern Uplands of Scotland. Having examined a number of
sections, he was able to subdivide the Hartfell and Birkhill Shales into eleven units based on
distinct lithological differences, and found that these corresponded with differences in their
accompanying graptolite assemblages which he called 'zones'. Lapworth noted that the
lithological boundaries between the Glenkiln, Hartfell and Birkhill Shales were abrupt, and that
each had a distinctive graptolite fauna. Although the palaeontological differences between the
Glenkiln and Hartfell Shales were gradational, if one took a typical sample from each, they
Page 3.4

were completely different. The faunal variation between the Hartfell and Birkhill Shales he
described as marked, with only two species in common. Lapworth referred to the underlying
Glenkiln Shales as poorly exposed and faulted and, although able to describe a distinct
graptolite fauna, he did not attempt to divide it into faunal zones. The paper gives detailed
lithological logs of the best sections, as well as a chart outlining the vertical ranges of all
observed taxa from the sections studied. This was to be the model of procedure for all future
biostratigraphic work involving graptolites. In the following year, Lapworth (1879-80)
extended his study of graptolite faunal zones to include all British and international material,
identifying twenty zones from the Upper Cambrian to the Silurian. He provided range charts for
existing graptolite genera and species, and referred to patterns observed within the vertical
ranges of the genera, noting that at specific level, the restriction in vertical ranges could provide
a detailed temporal subdivision of the Lower Palaeozoic. He also suggested that longer ranging
graptolites show a wider variety of morphologies than short ranging ones.
These zones were recognisable in other parts of the UK, as later papers indicated,
notably in the Stockdale Shales (Marr & Nicholson 1888) and the Skiddaw Slates (Marr 1894)
of the Lake District. Key biostratigraphic work in Wales and the Welsh Borderland increased
the number of known British graptoloid forms and identifiable graptolite faunal zones. Elies’
(1900) study of the Wenlock Shales of the Welsh Borderland, W ood’s (1900) on the Lower
Ludlow Formation of the Welsh Borderland, and H. Lapworth’s (1900) work on the Silurian
sequence at Rhayader in mid-Wales, all provide detailed lithostratigraphic accounts of numerous
sections in these areas together with descriptions of new taxa. All three authors give range
charts outlining the vertical distribution of taxa, comparing each district covered, adding new
zones and altering existing ones according to distinct local variations in the composition of its
graptolite fauna. Wood (1900) attempted to subdivide part of the Ludlow sequence at Long
Mountain into sub-zones. It is important to note that these workers also compared their Welsh
Basin range charts and faunal lists to information available from the rest of Britain and abroad,
particularly from Sweden.
‘A monograph of British graptolites’ (Elies & Wood 1901-18) is a complete account of
all the British graptolite genera, species and subspecies known at that time, providing
descriptions and figures of each taxon. Based on all available information, full stratigraphic
Page 3.5

ranges for each taxon were given, corresponding to 36 recognisable graptolite zones and
subzones covering the whole of the Lower Palaeozoic in the UK. Elies and Wood (1913)
compared these new zonal ranges to those of Lapworth (1879-80) and demonstrated how they
fitted into his biostratigraphic framework, while taking into account variations in the Ordovician
chronostratigraphy of the Lake District (Marr 1905). This was the first time that one study on
British graptolites had unified the taxonomy, biostratigraphy and chronostratigraphic
importance of the group to such an extent. The monograph continues to provide the basis for
current UK graptolite biostratigraphy and its effect on global correlation remains significant,
although many of the individual taxon descriptions and ranges have been revised in numerous
subsequent studies.
Graptolites were now established as a fundamental part of British Lower Palaeozoic
stratigraphy. Their continued use in geologic studies in the UK and abroad led to a greater
understanding of the development, palaeobiology and palaeoecology of the group, as well as a
continued recognition of new taxa and reassessment of old ones. Enhanced resolution of British
graptolite biozones, coupled with the refinement of UK Lower Palaeozoic stratigraphy, was the
result of many authors’ attempts to relate the assemblages they saw in the field to the existing
sequence of graptolite faunal zones (e.g. Jones 1921, 1947, Davies 1926, Jones & Pugh
1935). These, together with a number of large, detailed works in classic areas of Lower
Palaeozoic stratigraphy indicated the need for the overall British graptolite biostratigraphic range
charts of Elies and Wood (1913) to be revised. Sudbury (1958) published an account of the
faunas comprising the subzones of the gregarius zone from the Lower Llandovery of the
Rheidol Gorge in Wales, detailing the taxonomy and evolution of the triangulate monograptids
found in the area. Toghill (1968) updated the graptolite faunas and all the zones of the Lower
Silurian Birkhill Shales unit, the first time these had been reassessed since Lapworth’s original
study at Dob’s Linn (1878). Extensive studies on the Silurian strata of Northern England, with
a paper on the Wenlock and Ludlow Series of the Howgill Fells (Rickards 1967) and
monographs of Llandovery material from the same region (Rickards 1970), and from the Lake
District (Hutt 1974-75) provided a detailed graptolite taxonomy and stratigraphy of their
respective areas and new faunal range charts. All significant work on British Silurian graptolite
biostratigraphy since Elies and Wood’s monograph (1901-18) was collated by Rickards (1976).
Page 3.6

Together with a detailed description of each zone and subzone, he produced new graptolite
range charts for the Silurian of the UK. However, there has not been an updating of Elies and
Wood’s Ordovician charts.
The past twenty years have seen British graptolite biostratigraphers expanding
knowledge of the strata and graptolite faunas of the Ordovician, as well as carrying out more
detailed work on the Ordovician-Silurian boundary and in the Llandovery. W illiams’
descriptions of the graptoloid faunas from classic sections at Dob’s Linn (1982b, 1988) and in
the Moffat and Girvan districts (1982a, 1987, 1994) have provided revised Upper Ordovician
range charts and correlation with key sequences in Australia and North America. The section at
Dob’s Linn was designated the international basal boundary stratotype for the Ordovician-
Silurian boundary (Williams 1988). Recent geological mapping and concurrent biostratigraphic
work by the British Geological Survey (BGS) in areas of Lower Palaeozoic stratigraphy have
resulted in the acquisition of large amounts of data. Graptoloids of various Arenig ages have
been found in the Ballantrae ophiolite complex (Stone & Rushton 1983) including those
correlating with the La 3 Australasian graptolite zone (Upper Lancefieldian). Initially, Arenig
graptolites from the type area in North Wales proved of less correlative value due to their low
diversity (Zalasiewicz 1984a, 1986), though more diverse and better preserved faunas were
found in the Carmarthen area of South Wales (Fortey & Owens 1987). These studies,
especially Rushton’s (in preparation) reorganisation of Arenig biostratigraphy in the Skiddaw
Group, together with studies in higher Ordovician strata (Zalasiewicz et al. 1995, Cave &
Rushton 1996) helped clarify some of the problems traditionally associated with Ordovician
chronostratigraphy, namely the ability to correlate type British sections accurately with those
abroad.
Work associated with the remapping of central Wales by BGS led to refinement of
Llandovery and Wenlock graptolite biostratigraphy. Studies of early to middle Llandovery
(Rhuddanian-Aeronian) sections and graptoloid faunas produced some biostratigraphic
refinement (Zalasiewicz 1992, 1996, Zalasiewicz & Tunnicliff 1994). More spectacular
progress was seen in the late Llandovery (Telychian) (Loydell & Cave 1993, 1996, Loydell
1993a, Zalasiewicz 1994, Zalasiewicz et al. 1995, Davies et al. 1994) where, inter alia, the
former Spirograptus turriculatus zone has been split into two separate zones, each with several
Page 3.7

component subzones. Current work on Elies’ (1900) original Wenlock biozonal localities near
Builth Wells (Williams & Zalasiewicz in press) has resulted in a reorganisation of graptolite
biozones for this part of the stratigraphic column. Nevertheless, large sections of British Lower
Palaeozoic stratigraphy remain relatively neglected, for example the Ludlow Series, and even
classic sections and faunas require revision if they are to continue being of use for global
correlation.
Range Charts of Ordovician and Silurian graptoloids of the UK
Graptoloid faunal range charts provide a detailed index of the composition of the faunal
assemblage biozones that represent specific temporal horizons within the stratigraphic column,
the duration of which may be as little as 0.45 Ma in parts of the Silurian of the UK (Zalasiewicz
1990, Hughes 1995). The chronological and correlative potential of such information is
invaluable to geologists in general, and stratigraphers in particular, who work in areas
containing graptolite-bearing strata.
Until the present study, Elies and Wood (1901-18) provided the only full British
Ordovician and Silurian graptoloid biostratigraphic range charts, although the Silurian ranges
have been updated by Rickards (1976). It was obvious that the Ordovician was in need of
updating, given the time elapsed since Elies and Wood's study. However, refinement of Lower
Palaeozoic graptoloid biostratigraphy in the UK generally has been rapid in recent years. As a
result, the Geological Society of London commissioned a Special Report on current UK
graptoloid biostratigraphy for the Lower Palaeozoic, to contain new Ordovician and Silurian
biozonal range charts. The compilation of the comprehensive graptoloid database needed for
these charts is central to this paper.
Ensuring that the database is as complete a geographical and temporal coverage of
Ordovician and Silurian graptoloid biostratigraphy for the UK as possible was paramount.
Therefore, using the Elies and Wood (1901-18) and Rickards (1976) range charts as a starting
point for the database, the three main objectives of the current study were: (i) List all graptoloid
taxa. (ii) Apply taxonomic revisions, such as generic reassignments and synonymisation of
species where appropriate, (iii) Adjust the stratigraphic ranges of all taxa in accordance with all
Page 3.8

information now available in the data. Ranges for Ireland are not included, given that
constraints of time and space did not allow the inclusion of Irish data.
For the British Ordovician two separate graptoloid range charts were produced, one
representing England and Wales (Figs. 3.4.1-3.4.5) and the other Scotland (Figs. 3.5.1-
3.5.3), as significant regional differences in graptolite faunas exist. The separation of Scotland
from England and Wales by the Iapetus Ocean resulted in faunal provincialism during the
Ordovician; and for graptolites, this remained the case even upon the closure of Iapetus at the
end of the system, probably as a result of environmental factors (Zalasiewicz et al. 1995). Few
such problems exist in the Silurian, and a single sequence of graptolite biozones can be
recognised throughout the UK.
Biostratigraphy of UK Graptoloids
Complete UK graptoloid range charts have not been published since Rickards (1976) for the
Silurian (Fig. 3.3), or since Elies and Wood (1901-18) for the Ordovician. Taking all available
new data into account, the total number of known graptoloid taxa for both systems has
increased significantly. Many zones have been renamed and re-evaluated as a result of more
detailed study, while the biostratigraphy of some parts of the stratigraphic column can now be
refined and subdivided to a much greater degree of resolution.
Over the years, there have been generic reassignments for a number of Lower
Palaeozoic graptoloids. Unless directly discussing another authors’ work, I will use the most
recent standard reference for generic names (Strachan 1996-1997), consistent with those used in
the updated range charts.
Ordovician
Araneograptus murrayi Biozone
This biozone was first used in the UK by Cooper et al. (1995), in reference to the Skiddaw
Group from the English Lake District. The A. murrayi Biozone is currently the earliest
recognisable graptoloid biostratigraphic interval in the British sequence, representing the Upper
Tremadoc Series (Fig. 3.1). The base is taken on the appearance of the zone fossil, as well as
Acrograptusl cf. sinensis and Didymograptus sp. (declined) of Molyneux & Rushton (Rushton
Page 3.9

et al. 1997, Molyneux & Rushton 1988). These species are the only taxa as yet identified at this
level (Fig. 3.4.1).
Tetragraptus phyllograptoides Biozone
Cooper et al. (1995) initiated the use of T. phyllograptoides Biozone, an established
Scandinavian interval (Lindholm 1991), based on the homotaxy between the Skiddaw Group
faunas and those characteristic of the upper part of this biozone in Scandinavia (Maletz et al.
1989). The zone fossil is absent from the Lake District, while this interval has not been recorded
elsewhere in the UK (Cooper et al. 1995, Rushton in preparation). Species diversity is still
relatively low in the phyllograptoides Biozone, although significantly larger than that of the
previous biozone, including the first tetragraptids and temnograptid (Fig. 3.4.1). Confirmed
new taxa are A c ro g ra p tu s l p ro to b a lticu s , Tem nograptus m ultip lex, T e tra g ra p tu s
quadribrachiatus, Acrograptusl rigoletto and Tetragraptus (Pendeograptus) cf. fruticosus.
In Scotland, graptoloid faunas from the lower to middle part of the Ordovician sequence
more closely resemble those in Australasia than the traditional English and Welsh faunal
assemblages. This prompted Stone & Rushton (1983) to use the Australasian graptolite
biostratigraphic scheme (VandenBerg & Cooper 1992) during their work on the Ballantrae
ophiolite complex graptolite faunas. This sequence of biozones has been incorporated into the
Scottish graptoloid biostratigraphic range charts of the current study. Therefore, the earliest
graptoloid interval, at the base of the Arenig Series in Scotland, is called Lancefieldian 3. This
fauna consists of Tetragraptus approximatus and Tetragraptus quadribrachiatus (Fig. 3.5.1).
Corymbograptus varicosus Biozone
The C. varicosus Biozone takes the place of the traditional Corymbograptus deflexus Biozone
(Jackson 1978, Fortey & Owens 1990), following the recent revision of Skiddaw Group
graptoloid biostratigraphy (Cooper et al. 1995). Cooper et al. (1995) found C. deflexus to be
rare at this interval, whereas C. varicosus is abundant. There is no clear definition for the base
of the varicosus Biozone, due to the lack of suitable fossiliferous strata in the Lake District
(Rushton in preparation). This faunal assemblage displays a marked increase in species and
generic diversity. New taxa include Baltograptus vacillans attenuatus, Didymograptus aff.
Page 3.10

balticus, Tetragraptus reclinatus, Didymograptus cf. decens and Acrograptus filiformis, as well
as a distinct upper zone fauna consisting of: C. deflexus, Pseudophyllograptus angustifolius,
Dichograptus octobrachiatus sedgwickii, Schizograptus tardifurcatus and Expansograptus cf.
similis (Fig. 3.4.1).
The Scottish faunas at this approximate level contain elements consistent with those of
the lower Bendigonian Stage (Be 1-2) of Australasia (Stone & Rushton 1983). The most
distinctive taxa are Tetragraptus approximatus and Tetragraptus fruticosus (Fig. 3.5.1), placing
this assemblage lower in the Ordovician sequence than the varicosus fauna of England and
Wales, which equates approximately with the upper Bendigonian Stage (Be 3-4) of Australia
(Cooper & Fortey 1982). Other forms from Scotland include Acrograptus cf. filiform is,
Paradelograptus sp. A of Rushton et al. and Tetragraptus cf. decipiens (Rushton et al. 1986).
Didymograptus simulans Biozone
The former Didymograptus nitidus Biozone was renamed on the basis that the old zone fossil
does not occur at this interval in the Skiddaw Group (Cooper et al. 1995). The base of the
biozone is defined on the incoming of D. simulans and Acrograptus infrequens (Cooper et al.
1995, Rushton et al. 1997). A significant rise in graptoloid diversity can be seen at this horizon,
with about 33 taxa recorded (Fig. 3.4.1). Several new forms are confined to this biozone,
including Didymograptus minutus, Tetragraptus postlethwaitei, Adelograptus divergens,
Corymbograptus v-fractus and Tetragraptus pendens. Other noteworthy appearances are
Azygograptus eivionicus, Azygograptus lapworthi, Pseudobryograptus cumbrensis,
Acrograptus kurcki, Loganograptus logani and Tetragraptus crucifer. Cooper et al. (1995)
suggest that the presence of Isograptus victoriae cf. prim ulus at this interval indicates a
correlation with the Chewtonian Stage (Ch 1-2).
The contemporaneous Scottish taxa differ markedly from the fauna of England and
Wales, with the exception of Tetragraptus serra and Tetragraptus reclinatus which occur in both
successions (Fig. 3.5.1). Stone & Rushton's (1983) work at Ballantrae revealed a graptoloid
assemblage containing D idym ograptus cf. protom urchisoni, Tetragraptus bigsbyi cf.
askerensis, E xpansograp tusl aff. geom etricus, D idym ograptus cf. proto indentus and
Tetragraptus cf. kindlei, which they found consistent with a Chewtonian age.
Page 3.11

Isograptus victoriae Biozone
The recognition of distinct assemblages within the old Isograptus gibberulus Biozone of the
Skiddaw Group, led to the proposal of a lower I. victoriae Biozone and an upper I. gibberulus
Biozone (Rushton et al. 1997). The I. victoriae Biozone is characterised by the appearance of
the zone fossil, which is confined to this interval. However, Rushton et al. (1997) record that
the index species is rare in the Lake District, leading to difficulties in identifying the zonal
boundaries. These authors note the importance of Expansograptus hirundo in the recognition of
these two new biozones. Other forms limited to this zone are Pseudophyllograptus densus and
Isograptus victoriae cf. m a xim u s, and notable incoming taxa are E. hirundo, and
Expansograptus cf. nitidus (Figs. 3.4.1 & 3.4.2), recorded for the first time in the Shelve area
of Shropshire (Strachan 1986).
A graptoloid fauna corresponding to this interval has not been reported from Scotland
(Fig. 3.5.1).
Isograptus gibberulus Biozone
This biozone, as restricted by Rushton et al. (1997) in the Skiddaw Group, English Lake
District, is delineated by the appearance of /. caduceus gibberulus. As in the previous zone,
many of the taxa comprising the zonal assemblage are not new appearances; nevertheless,
several strati graphically significant forms do originate in the gibberulus interval (Fig. 3.4.2).
Cooper et al. (1995) suggested that the former extended I. gibberulus Biozone equated with the
Australasian Castlemainian Stage (Ca 1-4). The newly restricted I. gibberulus Biozone is
representative of the upper part of the Castlemainian (Ca 3-4). The identification of
Pseudisograptus dumosus at this horizon from the Aberdaron area of Wales, and Isograptus
caduceus cf. imitatus from Grisedale Pike in the Lake District (Jenkins 1982) would seem to
confirm this. Other noteworthy appearances include Pseudisograptus angel, Corymbograptus v-
fractus volucer, Tetragraptus bigsbyi, C orym bograptus? uniform is and Xiphograptus
svalbardensis (Fig. 3.4.2).
A Scottish faunal assemblage obtained from a borehole core taken from the Ballantrae
ophiolite complex, contained elements comparable to those for this interval in England and
Wales (Stone & Strachan 1981), and is herein referred to the late Castlemainian (VandenBerg &
Page 3.12

Cooper 1992), although the exact stratigraphic horizon has not been identified. A full list of taxa
is given in the range chart (Fig. 3.5.1). As yet, no graptoloid assemblages have been reported
from Scotland at a higher stratigraphic level, until the advent of the Nemagraptus gracilis
Biozone at the base of the Caradoc Series (Fig. 3.5.1).
Aulograptus cucullus Biozone
Having revised the underlying victoriae and gibberulus biozones, Rushton et al. (1997) rejected
the hirundo Biozone on account of the fact that E. hirundo originates in the earlier victoriae
Biozone (Fortey et al. 1990), and that it is indeed rare at this interval in the Skiddaw Group.
The upper zone was renamed the A. cucullus Biozone and the base is characterised by the
incoming of the name fossil. Many taxa originate in the biozone, among them the first
cryptograptids, Oelandograptus australodentatus anglicus, Acrograptus nicholsoni planus,
Didymograptus protobifidus and Glyptograptus shelvensis, the first glyptograptid in the British
Ordovician/Silurian sequence (Fig. 3.4.2). A. cucullus Biozone faunas have also been recorded
from the Shelve district of Shropshire (Strachan 1986). The range chart shows a few taxa
indicative of the upper part of this zone (Fig. 3.4.2), these are Acrograptus acutidens, and
possibly Amplexograptus confertus and Cryptograptus tricornis schaeferi.
Didymograptus artus Biozone
The D. artus Biozone corresponds with the base of the Llanvirn Series, and the name was
established by Fortey & Owens (1987) to replace that of the traditional Didymograptus bifidus
Biozone, which was taxonomically incorrect. This interval displays a highly diverse and
distinctive graptoloid fauna (Figs. 3.4.2, 3.4.3), with the majority of taxa appearing at the base
of the biozone, which is characterised by the zone fossil and Didymograptus spinulosus
(Fortey & Owens 1987, Fortey et al. 1990, Rushton et al. 1997). Approximately 31 species
originate in the biozone, 21 of which have ranges restricted to this level. Several genera are
recorded for the first time, notably Holmograptus lentus and including the glossograptids, the
climacograptids and the diplograptids. The artus Biozone has good potential for further
biostratigraphic subdivision. While most elements of the assemblage span the entire zone, there
is a distinguishable middle to upper biozonal fauna containing Pseudoclim acograptus
Page 3.13

scharenbergi, Pseudoclimacograptus angulatus, Trichograptus fragilis, D iplograptus
hollingworthi and Nicholsonograptus fasciculatus (Fig. 3.4.3). A few taxa help differentiate the
lower part of the biozone, such as Par agio ssograp tus sp., and D. protobifidus and
Expansograptus sparsus which originate in the cucullus Biozone but disappear in the early part
of this interval. The D. artus Biozone has been widely recognised in northern England and
Wales (Elies 1940, Strachan 1986, Fortey & Owens 1987, Fortey et al. 1990, Cooper et a l
1995, Rushton in preparation).
Didymograptus murchisoni Biozone
Graptoloid assemblages characteristic of this biozone have been recorded from the Abereiddi
and Builth-Llandrindod Wells districts of central Wales (Elies 1940, Hughes 1989, Davies et al.
1997), the Shelve area of Shropshire (Strachan 1986, Hughes 1989) and the Skiddaw Group
strata of the Tarn Moor Tunnel in the Lake District (Wadge et al. 1972). The base of the
m urchisoni Biozone sees the incoming of the type fossil, which is restricted to this
biostratigraphic interval (Fig. 3.4.3). Other new species include Diplograptus foliaceus,
Cryptograptus tricornis, Pseudoclimacograptus angulatus magnus, Pseudoclimacograptus
angulatus micidusl and Didymograptus speciosusl (Fig. 3.4.3). Diplograptus decoratus,
Amplexograptus caelatus and Climacograptus brevis appear in the middle to upper part of the
biozone, while Pterograptus elegansl, Lasiograptus retusus and Didymograptus nanus are only
reported from the lower portion of the zone: this may suggest possible refinement of the
biostratigraphy.
Glyptograptus teretiusculus Biozone
The base of this biozone is delineated by the disappearance of D. murchisoni (Hughes 1989), as
the zone fossil, Glyptograptus teretiusculus teretiusculus, originates in the middle part of the
interval (Fig. 3.4.3). The only taxa to appear at the basal boundary are D icellograptus
divaricatus divaricatus, Climacograptus antiquus and possibly Glyptograptus teretiusculus
euglyphus. Approximately half the total species comprising this biostratigraphic level are
initiated in the middle to upper part of the zone. These data provide a strong case for a
biostratigraphical review of the graptoloid faunas found at this interval. However, Hughes
Page 3.14

(1989) remarks that Elies' (1940) attempt to subdivide the teretiusculus and gracilis biozones in
the Builth area was incorrect, as her refinement was based on taxonomically unsound species
and poorly preserved material. In its present state, the teretiusculus Biozone has been recorded
from Wales and Shropshire (Elies 1940, Strachan 1986, Hughes 1989, Davies et al. 1997).
Nemagraptus gracilis Biozone
This biostratigraphic interval was first described by Lapworth (1878) and defined on the
incoming of the dicellograptids and nemagraptids. Subsequent work led to a revised definition
of the base of the biozone, which is now taken at the incoming of N. gracilis (Finney &
Bergstrom 1986). Faunas representing this biozone have been recorded from the Builth region
of Wales and the Shelve area of the Welsh Borderland (Elies 1940, Strachan 1986, Hughes
1989, Davies et al. 1997), however, the position of the base of the gracilis Biozone at Builth
Wells is unclear (Hughes 1989, Davies et al. 1997). The Anglo-Welsh N. gracilis fauna is
diverse, containing a large number of previously unrecorded species, approximately 26 taxa, as
well as many longer-ranging forms from underlying zones (Figs. 3.4.3, 3.4.4). Several species
appear in the upper part of the biozone, and although only one of these, Lasiograptus pusillus,
appears limited to this horizon, the appearances may be biostratigraphically useful in the future.
Forms coming in in the upper part of the zone include Dicranograptus nicholsoni nicholsoni,
Climacograptus bicornis bicornis, Amplexograptus perexcavatus and Glossograptus hincksii
hincksii.
Graptoloids reappear dramatically in the Ordovician sequence of Scotland at this level.
Following the sparse, low-diversity assemblages characteristic of the Scottish lower to middle
Arenig Series, and the absence of the upper Arenig and Llanvirn faunas, a varied and abundant
graptoloid fauna (approx. 45 taxa) of N. gracilis age has been recorded from southern Scotland
(Stone 1995, BGS unpublished biostratigraphic reports, Rushton et al. 1996). From this
biozone upwards, the Scottish graptoloid succession is more directly comparable with that of
England and Wales, although the two biostratigraphic schemes still reflect significant
assemblage differences (Fig. 3.1), making precise correlation difficult (Zalasiewicz et al. 1995).
In Scotland, the gracilis Biozone is similarly defined as in England and Wales, although Stone
(1995) comments on the importance of Expansograptusl superstes and Dicellograptus intortus
Page 3.15

to the characteristic gracilis fauna (Figs. 3.5.1, 3.5.2). D. intortus appears lower in the Anglo-
Welsh sequence and the exact position of the base of the gracilis interval cannot be clearly
recognised in Scotland (Rushton 1990).
Diplograptus multidens Biozone
The D. multidens Biozone is an Anglo-Welsh biostratigraphic unit, traditionally employed to
describe the graptoloid faunas found between those of the underlying gracilis Biozone, and the
Dicranograptus clingani Biozone above (Fig. 3.1). However, the taxonomic validity of D.
multidens has been questioned (Hughes 1989, Rushton 1990), and much of Elies & Wood's
(1901-18) and Elies' (1940) earlier work on this biozone from the Shelve inlier and the type
sections of the Caradoc area of Wales must be treated with caution, as many of the original taxa
have not been found following recent studies (Hughes 1989). Nonetheless, work carried out by
BGS in central Wales has confirmed the existence of a distinct and stratigraphically useful
multidens Biozone fauna (Fig. 3.4.4) with several newly occurring taxa (Davies et al. 1997,
BGS unpublished biostratigraphical reports). Rushton (1990) suggests that following full
revision the name of this interval be changed and a new name fossil appointed. An upper
biozonal assemblage with biostratigraphic potential can be identified (Fig. 3.4.4), including
Dicranograptus nicholsoni minor, Dicellograptus patulosus, Climacograptus sp. of Rickards
and Glyptograptus sp. of Rickards.
In Scotland, this biostratigraphic interval is subdivided into two biozones, a lower
Climacograptus peltifer Biozone and an upper Climacograptus wilsoni Biozone. There are
problems with the recognition of the C. peltifer Biozone, as the index fossil is taxonomically
invalid (Strachan 1971, Rushton 1990), being considered an astogenic variant of C. bicornis
(Riva 1976), and the original diagnostic fauna (Elies 1925) is virtually identical to that of the
underlying N. gracilis Biozone (Rushton 1990). Work carried out by BGS resulted in new
species being identified, and the possibility of improved zonal recognition; but this is not a
complete solution to the problem (Stone 1995, BGS unpublished biostratigraphical report).
Recent identifications include Pseudoclimacograptus cf. isknos, Orthograptus calcaratus s.l. and
possibly Thamnograptus sp. (Fig. 3.5.2).
Page 3.16

Lapworth (1878) established the C. wilsoni Zone during his work on the Moffat Series
of the Southern Uplands. It continued to be an integral part of the Scottish graptoloid
biostratigraphy despite no further significant biostratigraphic study of the interval, or
recognition of the diagnostic fauna elsewhere in the UK (Elies & Wood 1901-18, Elies 1925,
Elies 1937). A recent taxonomic and biostratigraphic review of the wilsoni Biozone by Williams
(1994), has confirmed the stratigraphical value of this graptoloid assemblage in southern
Scotland where it has now been more widely recognised (Stone 1995, Williams 1994),
resulting in the designation of a type section for the interval at Hartfell Spa (Williams 1994).
The wilsoni fauna has fewer taxa than the peltifer assemblage, and many of these continue from
the lower biozone (Fig. 3.5.2). The base of the C. wilsoni Biozone is defined by the appearance
of the index species; other incoming taxa are Orthograptus calcaratus vulgatus, Coryoides
curtus, Corynoides calicularis, Dicellograptus angulatus and Orthograptus ex. gr. calcaratus.
Dicranograptus clingani Biozone
The long established D. clingani Biozone has been recognised in Wales and Scotland (Rushton
1990, Williams 1982a, Stone 1995, Davies et al. 1997). Work at Hartfell Score in the Southern
Uplands of Scotland and Whitland in south Wales led Zalasiewicz et al. (1995) to erect two sub
biozones within the clingani interval, clearly defined by distinct graptoloid assemblages and
recognisable in both regions (Fig. 3.1), although there is some variation in faunal composition
(Zalasiewicz et al. 1995). The potential for subdivision of the zone in the Southern Uplands had
previously been remarked on by Williams (1982).
Climacograptus caudatus Subzone
The base of this subzone is defined by the incoming of the type species at Hartfell Score with a
large accompanying fauna of new forms (Zalasiewicz et al. 1995). D. clingani is restricted to the
caudatus Subzone in both Wales and Scotland (Zalasiewicz et al. 1995, Davies et al. 1997,
Figs. 3.4.4, 3.5.3). In south Wales, the fauna contains similar elements to the Southern
Uplands, such as Orthograptus quadrimucronatus, and Climacograptus spiniferus. However,
C. caudatus itself had not been recognised in this region until Davies et al. (1997) reported it
Page 3.17

from the Llanilar-Rhayader area. Overall, the caudatus fauna of Wales is less diverse than that
of Scotland, about 28 and 46 taxa present respectively.
Dicellograptus morrisi Subzone
In Wales and Scotland, the D. morrisi Subzone has a large and diverse graptoloid fauna, but
fewer incoming species than the underlying C. caudatus Subzone (Figs. 3.5.2, 3.5.3). At
Hartfell Score, Zalasiewicz et al. (1995) describe this interval as a partial-range subzone,
defined at the base by the appearance of D. morrisi and at the top, by the incoming of the
linearis Biozone assemblage. At Whitland, the base of the subzone is delineated by the incoming
of Climacograptus miserabilis and possibly D. morrisi, the range of which is limited to this
biostratigraphic horizon (Zalasiewicz et al. 1995). Glyptograptus daviesi is the only new species
definitely reported from the base of the morrisi Subzone in Wales (Fig. 3.4.4). In Scotland,
several incoming taxa, excluding the zone fossil, occur, including Climacograptus minimus,
Dicello graptus caduceus and Leptograptus flaccidus spinifer (Fig. 3.5.3). Davies et al. (1997)
did not identify a discrete upper clingani Biozone assemblage from central Wales.
Pleurograptus linearis Biozone
The original work on the graptoloid faunal assemblage of this interval was carried out by
Lapworth (1878) on the upper part of the Lower Hartfell Shales in the Southern Uplands.
Williams (1982a) undertook the only comprehensive study of the P. linearis Biozone in this area
since Elies & Wood's (1901-18) review of British graptolites. He was able to delineate the base
of the biozone for the first time in the North Cliff section of Dob's Linn, and designated it as the
type section for the interval base (Williams 1982a), although clearer definitions have been given
by subsequent workers (Zalasiewicz et al. 1995, Rushton et al. 1996). Basal diagnostic taxa
include the zone fossil, with Climacograptus tubuliferus, possibly Climacograptus styloideus
and Leptograptus flaccidus macilentus; however, several forms appear mid-biozone including
Amphigraptus divergens divergens, Leptograptus capillaris and Dicello graptus elegans elegans
(Fig. 3.5.3). The overall assemblage is quite diverse in Scotland, although most species
continue from underlying biozones. Graptoloids characteristic of this biostratigraphic interval
Page 3.18

have been reported in the Rhins of Galloway, south-west Scotland (Stone 1995) and by
Zalasiewicz et al. (1995) at Hartfell Score.
At Whitland in south Wales, a low diversity graptoloid faunal assemblage, possibly
equating with the P. linearis Biozone, has been recognised by Zalasiewicz et al. (1995), the
base of which marks the appearance of large normalograptids. They refer to this biozone as the
'Normalograptus proliferation interval' (Fig. 3.4.4). Davies et al. (1997) note that they did not
record any linearis Biozone faunas from the Llanilar-Rhayader area. Exact correlation between
the base of the Ashgill Series and the UK graptoloid biostratigraphic sequence cannot as yet be
made as the Welsh record is poor, and the Lower Ashgill of Scotland is rarely fossiliferous
(Rushton 1990). The linearis Biozone is currently thought to span the Caradoc/Ashgill
boundary, as the lower part of the zone is believed to correlate with the Onnian Substage
(uppermost Caradoc, see Fig. 3.1) (Ingham & Wright 1970, Ingham 1979).
Dicello graptus complanatus Biozone
The D. complanatus Biozone is another graptoloid assemblage zone initally recognised in the
Southern Uplands of Scotland (Dob's Linn) by Lapworth (1880), but largely neglected by
biostratigraphers since its inception. The work of Williams (1987) on strata representing this
interval from the Moffat and Girvan areas has resulted in a greater understanding of the
complanatus faunal assemblage, although the upper and lower limits of the biozone cannot be
precisely delineated, as they occur in strata lacking graptolites. The low diversity characteristic
fauna consists of mainly new species confined to this level, including the zone fossil and
Dicello graptus alector, Orthoretiolites pulcherrimus, Dicellograptus minor and Glyptograptus
occidentalis. Longer-ranging species occur, such as C. miserabilis and C. tubuliferus (Fig.
3.5.3).
The complanatus Biozone has not been conclusively proved in Wales (Rushton 1990).
However, unpublished BGS biostratigraphical reports suggest the possible existence of strata
representing this interval, although no species diagnostic of the complanatus Biozone have been
recovered.
Page 3.19

Dicello graptus anceps Biozone
Lapworth originally described the D. anceps Zone and its fauna from the Moffat area of the
Southern Uplands of Scotland (1878). A review of this interval was undertaken by Williams
(1982b), who subdivided the biozone into a lower Dicello graptus complexus Subzone and an
upper Paraorthograptus pacificus Subzone. The base of the D. complexus interval is marked by
the incoming of D. complexus and D. anceps. The subzone possesses a relatively small but
distinct graptoloid fauna with only Dicello graptus minor continuing from the previous biozone.
Other new appearances include Orthograptus fastigatus, C lim acograptus supernus,
Climacograptus normalis, Pleurograptus lui and Orthograptus abbreviatus (Fig. 3.5.3). The
upper part of the complexus Subzone is characterised by the incoming of Climacograptus latus
and Orthoretiograptus denticulatus. The overlying P. pacificus Subzone has a slightly more
diverse fauna, consisting of the same taxa from the underlying complexus Subzone with the
addition of, at the base, P. pacificus, Nymphograptus velatus and Plegmatograptus lautus (Fig.
3.5.3). The upper part of the subzone sees the incoming of Climacograptus? extraordinarius and
Glyptograptus? pseudovenustus cf. pseudovenustus. Stone (1995) notes anceps Biozone
assemblages in south-west Scotland.
Graptoloids of probable D. anceps Biozone age have been recorded in Wales, although
no potential for subdivision has been reported (Davies et al. 1997, BGS unpublished
biostratigraphical reports). Taxa include the zone fossil and C. supernus, C. normalis and O.
abbreviatus (Fig. 3.4.4).
Glyptograptus persculptus Biozone
This biozonal assemblage was originally subdivided from the lower part of the acuminatus
Biozone by Jones (1909) from Pont Erwyd, central Wales. The interval has since been
recognised throughout the UK (Davies 1929, Toghill 1968, Rickards 1970, 1976, Hutt 1974-
75, Williams 1988). The exact position of the G. persculptus Biozone within the British
chronostratigraphic framework has been long discussed. Rickards (1976) placed the interval as
the first graptolite biozone of the Silurian, acknowledging that the biostratigraphic range of the
index fossil could be extended lower, into the Ordovician succession, due to the prevalence of
unfossiliferous strata at its base. The stratotype section for the Ordovician-Silurian boundary
Page 3.20

was designated at Dob's Linn (Williams 1988) at the base of the acuminatus Biozone in order to
place the persculptus Biozone in the Ordovician, thereby providing a graptoloid interval below
this major boundary. Thus, the persculptus Biozone is now accepted as the highest graptoloid
assemblage biozone of the Ordovician in the UK biostratigraphic scheme. Indeed, recent work
on the synonomy of G. persculptus and 'Glyptograptus ' bohemicus, led Storch & Loydell
(1996) to recommend that the base of the persculptus Biozone could be further extended to
include a extraordinarius Subzone. The base of the interval is taken at the appearance of G.
persculptus’, other incoming species include Glyptograptus? avitus, Atavograptus ceryx,
Climacograptus parvulus, Climacograptus medius and Climacograptus tuberculatus (Figs.
3.4.4, 3.4.5, 3.5.3). Climacograptus scalaris indivisus appears in the middle of the biozone;
there are also questionable reports of Akidograptus ascensus and Paraclimacograptus innotatus
innotatus.
In Scotland (at Dob's Linn), the p e rsc u lp tu s Biozone is underlain by the
Climacograptusl extraor dinar ius Band (Ingham 1979, Williams 1983, 1988). However, the
graptoloid fauna is small and indistinct, based on the occurrence of the type fossil, which
originates in the preceeding biozone (Fig. 3.5.3), and C lim acograptus sp. indet. and
Glyptograptusl sp. indet. (Williams 1983).
Silurian
Parakidograptus acuminatus Biozone
This biozone is defined as the basal Silurian graptolite biozone of the global sequence. It was
originally recognised as such, and referred to as the D. acuminatus Zone by Lapworth (1878)
from the lower Birkhill Shale at Dob’s Linn. In his paper, Lapworth described a low diversity
fauna, where Diplograptus acuminatus and Diplograptus vesiculosus are common, and
Climacograptus scalaris and Dimorphograptus elongatus are rare. However, his vertical
distribution charts do not place D. vesiculosus in this zone, and indicate C. scalaris normalis as
very common. Elies & Wood (1913) also defined a ‘Cephalograptus’ acuminatus Biozone at the
base of the Silurian.
The base of the biozone has been defined by the incoming of Parakidograptus
acuminatus acuminatus and Akidograptus ascensus (Toghill 1968, Hutt 1974), in association
Page 3.21

with Parakidograptus acuminatus praematurus (Rickards 1976). Some workers recognise that
the acuminatus Biozone can be informally split into two subzones (Rickards 1976, Zalasiewicz
& Tunnicliff 1994, Davies et al. 1997). A. ascensus is generally considered more abundant in
the lower part of the biozone, with P. a. acuminatus more common in the upper part (Toghill
1968, Hutt 1974). Rickards (1976) discussed the possible subdivision on the appearance of
Climacograptus trifilis, Climacograptus rectangularis and Cystograptus vesiculosus in the
middle of the biozone. Hutt (1974) and Toghill (1968) also record this boundary higher in the
zone. Most authors place the appearance of C. rectangularis in the atavus Biozone (Baker 1981,
Rushton pers. comm., Zalasiewicz & Tunnicliff 1994). Records of typically Ordovician taxa,
most notably those of orthograptids of the truncatus group (Hutt 1974, Rickards 1976) have not
been substantiated by further work. Hutt’s (1974, Text-fig. 8, fig. 9) record is probably a
misidentification (Zalasiewicz pers. comm.).
Most authors agree on the other essential features of the biozone. This includes
significant numbers of climacograptids and diplograptids. The most frequently recorded of these
forms are Climacograptus normalis, Climacograptus medius, Climacograptus miserabilis,
Diplograptus modestus m odestus and D. modestus diminutus. Other taxa include
Climacograptus rectangularis, Climacograptus angustus (Zalasiewicz & Tunnicliff 1994),
Climacograptus trifilis (Toghill 1968, Rickards 1976), and Diplograptus modestus parvulus
(Elies & Wood 1913, Zalasiewicz & Tunnicliff 1994). Atavograptus ceryx is recorded from this
interval (Hutt 1974, Rickards 1976), as is Glyptograptus persculptus which continues into the
upper part of the biozone (Davies 1929, Zalasiewicz & Tunnicliff 1994, Davies et al. 1997).
Elies & Wood (1913) also recorded Metaclimacograptus hughesi and Climacograptus innotatus
innotatus, forms not recognised until higher in the British sequence by most workers.
The graptoloid range charts show that the criteria for identifying the base of the
Parakidograptus acuminatus Biozone remain unchanged (Fig. 3.6.1). They also reinforce the
possibility of informal subdivision of the biozone, with Glyptograptus persculptus and
Atavograptus ceryx having been recorded only in the lower part (though presumably there must
be some link between A. ceryx and later atavograptids which appear in the succeeding atavus
Biozone). P. acuminatus praematurus appears at the base of the biozone and is confined to the
lower part, while, as many workers worldwide recognise, A. ascensus is also more abundant
Page 3.22

than P. a. acuminatus in the lower part of the interval. The upper part of the zone contains
abundant P. a. acuminatus and sees the appearance of D. modestus, Cystograptus vesiculosus,
and in Central Wales, Orthograptus cabanensis (Zalasiewicz & Tunnicliff 1994). On current
evidence, D. modestus diminutus seems to be the only species originating in this biozone to
definitely span its entire length.
Atavograptus atavus Biozone
H. Lapworth (1900) originally referred to this unit as the 'Monograptus tenuis Zone', from the
Rhayader District of Central Wales. The fauna contained 'M. tenuis', Climacograptus scalaris
normalis and C. rectangularis. However, Lapworth misidentified the nominal species, and the
zone was subsequently renamed the atavus Zone (Jones 1909).
Species diversity increases in the atavus Biozone, from 18 taxa in the acuminatus
Biozone, to approximately 30 taxa (Fig. 3.6.1). Some taxa continue from the acuminatus
biozone, including numerous ‘climacograptids’. Rickards (1970) and Hutt (1974) record the
addition of Paraclimacograptus innotatus and Paraclimacograptus innotatus exquisitus
(Rickards 1970). However, it is possible that P. innotatus originates in the Ordovician, and has
been recorded sensu lato in the acuminatus Biozone (Rushton pers. comm.). The essential
feature of this biozone is the appearance at the base of significant numbers of species and
subspecies from the families Monograptidae and Dimorphograptidae: Atavograptus atavus,
Atavograptus gracilis, Raphidograptus extenuatus, Dimorphograptus confertus confertus,
Dimorpho graptus erectus erectus, Dimorpho graptus erectus nicholsoni, Dimorpho graptus
decu ssa tu s, D imorpho graptus decussatus partiliter, D im orpho graptus elongatus,
Dimorpho graptus longissimus and Dimorpho graptus epilongissimus (Elies & Wood 1913,
Rickards 1970, 1976, Hutt 1974, Zalasiewicz & Tunnicliff 1994). Rhaphidograptus toernquisti
originates in the upper part of the biozone (Hutt 1974, Rickards 1976). It is worth noting that
Dimorpho graptus confertus, Glyptograptus tamariscus and Cystograptus vesiculosus are not
recorded from this biozone in central Wales, but appear in the upper part of the acinaces
Biozone (Zalasiewicz & Tunnicliff 1994). Other forms appearing in the upper part of the zone
are Diplograptus? rarus (Rickards 1970), Coronograptus cyphus praematurus (Toghill 1968)
and Pribylograptus cf. incommodus (Rickards 1976). This suggests the possibility of informal
Page 3.23

subdivision of the biozone, as noted by Rickards (1976). Toghill (1968) did not follow the
zonal scheme adopted by most graptolite workers for this part of the Silurian. He described a
vesiculosus Biozone fauna from Dob’s Linn which equates with that of the atavus Biozone
recorded elsewhere (Rickards 1976), although Rickards (1976) noted that Toghill's (1968)
vesiculosus Biozone also included part of the acinaces Biozone. Elies & Wood (1913) describe
a zone of Orthograptus vesiculosus!Mesograptus modestus to account for the fauna between the
acuminatus and cyphus zones, and many of the taxa they recorded seem to indicate a clear
Atavograptus atavus Biozone fauna.
Lagarograptus acinaces Biozone
Jones’ (1909) initial description of the Monograptus rheidolensis Biozone fauna from the Pont
Erwyd District, Central Wales, was based on the appearance of M. rheidolensis (=
Lagarograptus acinaces), Pribylograptus sandersoni, Dimorpho graptus confertus swanstoni,
‘ Metaclimaco graptus hughesV (= Metaclimacograptus slalom) and Glyptograptus tamariscus.
Many authors describe the incoming of particular taxa, but few agree on which taxa define the
base of this zone. Recognition of the acinaces Biozone seems to be assemblage based, and
Rickards (1976) believes that the basal boundary cannot be clearly distinguished.
Although Toghill (1968) recorded L. acinaces and G. tamariscus s.I. from his
vesiculosus Biozone (= atavus and lower acinaces biozones), both Rickards (1970, 1976) and
Hutt (1974) place these as key taxa defining the base of the acinaces Biozone, along with ‘M.
hughesi.’ (= M. slalom). It may be the case that these forms appear earlier in the succession in
the Southern Uplands than in other parts of the U.K. In Central Wales, Metaclimaco graptus
slalom and G. tamariscus are not recorded until the upper part of acinaces Biozone. Only L.
acinaces comes in at the base here, and this disparity has allowed the biozone to be informally
subdivided into lower and upper assemblages (Zalasiewicz & Tunnicliff 1994, Davies et al.
1997). In this region, the zone fossil and associated climacograptids represent the lower part,
with a more diverse fauna in the upper part, with the incoming of several species, including
Metaclimaco graptus slalom, Glyptograptus tamariscus and many of the Monograptidae,
Dimorphograptidae and Diplograptidae commonly recorded in stratigraphically lower levels in
other areas. Toghill (1968) did not recognise an acinaces fauna in the Birkhill Shale at Dobb’s
Page 3.24

Linn. In his zonal scheme the cyphus Biozone follows the vesiculosus Biozone. Rickards
(1976) notes that the climacograptid and Dimorphograptidae dominated fauna from the lower
part of Toghill’s cyphus Biozone probably represents that of the acinaces Biozone, a view
supported by Hutt (1974). Stone (1995) records acinaces Biozone species from Gala Group
strata in the Rhins of Galloway area.
Jones’ (1909) original definition of the zone adequately described the core
acinaces Biozone assemblage, and the range charts herein are consistent with this (Fig. 3.6.1).
Notable additions include: Rhaphidograptus toernquisti (Rickards 1970, Zalasiewicz &
Tunnicliff 1994, Toghill 1968), Prihylograptus incommodus (Baker 1981, Zalasiewicz &
Tunnicliff 1994) in the upper part of the biozone, Atavograptus strachani (Rickards 1970, Baker
1981) and Pristiograptus fragilis pristinus (Zalasiewicz & Tunicliff 1994, BGS Davies et al.
1997), and the earlier Rhuddanian climacograptids, especially in the lower part of the zone.
Dimorpho graptus physophora and D. longissimus have been recorded at Dobb’s Linn (Toghill
1968); indeed, many of the Dimorphograptidae that appeared in the atavus Biozone occur at this
level. Taxa confined to this zone are Glyptograptus cf. nickoleyevi and 'Climacograptus' sp. of
Rickards (1970); M etaclimaco graptus slalom probably ranges higher. Apart from G. t.
tamariscus, there are possible recordings of other glyptograptids, for example, G. aff. incertus,
G. attenuatus and G. sinuatus sinuatus, the occurrence latter two being confirmed in the cyphus
Biozone.
Coronograptus cyphus Biozone
H. Lapworth’s (1900) original cyphus Biozone fauna, from the Wye Valley sequence of
Rhayader, consisted of Coronograptus c. cyphus, Monograptus revolutusl, Monograptus
attenuatus, Prihylograptus sandersonCl, Climacograptus rectangularis and C. normalis. As C. c.
cyphus has been recorded in the underlying acinaces Biozone (Zalasiewicz & Tunnicliff 1994),
its occurrence cannot define the base of the cyphus biozone. The revolutus/austerus group of
graptolites provides a more useful delineation, and their appearance at the base of the zone is
noted by most authors (H. Lapworth 1900, Jones 1909, Elies & Wood 1913, Rickards 1970,
1976, Hutt 1974, Baker 1981, Zalasiewicz & Tunnicliff 1994, Davies et al. 1997). They
include Monograptus austerus austerus, M. austerus bicornis, Monograptus sudburiae and M.
Page 3.25

revolutus (Zalasiewicz & Tunnicliff 1994). In addition, other new species are Coronograptus
gregarius gregarius (this species possibly originates in the upper acinaces Biozone),
Prihylograptus argutus argutus and Metaclimaco graptus undulatus. Hutt (1974) records
Prihylograptus incommodus as a key appearance at the base of cyphus Biozone in the Lake
District, but it has been observed earlier elsewhere (Toghill 1968, Baker 1981, Zalasiewicz &
Tunnicliff 1994, Davies et al. 1997), although it increases in abundance, along with C. c.
cyphus, in the cyphus Biozone (Toghill 1968, Rickards 1976). Some taxa continue from the
previous biozone, notably Atavograptus atavus, Lagarograptus acinaces, Climacograptus
m edius, C. rectangularis, Rhaphidograptus toernquisti, cf. Orthograptus mutahilis,
Cystograptus vesiculosus and Metaclimacograptus slalom, but they are mostly in decline and are
not recorded from all areas.
M. revolutus and C. g. gregarius appear in the middle of Toghill’s (1968) cyphus
Biozone at Dobb’s Linn. This, combined with an increase in numbers of C. c. cyphus at about
the same level, suggests that Rickards’ (1976) and Hutt’s (1974) belief, that only the upper part
of Toghill’s biozone represents the cyphus Zone seen in other parts of Britain, is correct. Elies
& Wood (1913) include a cyphus Zone in their zonal scheme, containing C. c. cyphus and C.
g. gregarius. Both these forms have been recorded from the underlying acinaces Biozone
(Rickards 1976, Zalasiewicz & Tunnicliff 1994), and the appearance of Rhaphido graptus
toernquisti, Glyptograptus tamariscus, Lagarograptus acinaces and Prihylograptus incommodus
in their cyphus Biozone suggests that at least the lower part of it is equivalent to the acinaces
Biozone. The recognition of C. g. gregarius by Elies & Wood (1913) in this fauna, probably
defines part of it as a true cyphus Biozone assemblage.
A distinct middle to upper part to this zone can be recognised (Fig. 3.6.2), mainly based
on the appearance of several glyptograptids, notably Glyptograptus cuneatus, G. tamariscus
distans, G. t. varians, G. t. linearis, G. t. angulatus and 1G. attenuatus. C lino climacograptus
retroversus is also a key upper biozone species. Further evidence for this is seen in the
appearance of Monograptus revolutus and Coronograptus gregarius, with greater numbers of
Coronograptus cyphus in the middle of the zone (Toghill 1968).
Page 3.26

Monograptus triangulatus Biozone
The M onograptus triangulatus Biozone represents the lowest part of a broad interval
traditionally known as the ‘Monograptus’ gregarius Biozone (Lapworth 1878, Elies & Wood
1913). The gregarius Biozone is still used internationally, and was standard in the U.K. until
comparitively recently (Toghill 1968). The possibility of a more refined subdivision of the
biozone was noted by Elies & Wood (1913), who set up three subzones. The lowest of these,
the fim briatus subzone, equates with the current triangulatus Biozone. Sudbury’s (1958)
detailed work on triangulate monograptids from the Rheidol Gorge in central Wales contributed
greatly to the concept of the existing triangulatus Biozone. Despite using the gregarius Biozone
herself, she also acknowledged a possible three-fold subdivision based on her reassessment of
the morphologies and ranges of the triangulates, a stratigraphically important group. This
enabled Rickards (1976) to outline the distinctive assemblages comprising the triangulatus,
magnus and leptotheca biozones.
The base of the triangulatus Biozone is marked by the appearance of several triangulate
monograptids (Fig. 3.6.2) including Monograptus triangulatus triangulatus, M. t. separatus and
M. t. praedecipiens. Diversity is high at this stratigraphic level, with 61 species and subspecies
currently recognised. Locally, however, biostratigraphically barren strata are found at the base
of the biozone. For example, in the Lake District, (Hutt 1974), central Wales (Zalasiewicz &
Tunnicliff 1994, Davies et al. 1997), and at Llanystumdwy, North Wales (Baker 1981), taxa
indicating the middle of the biozone appear above the cyphus Biozone. In Scotland, Toghill
(1968) also described a gregarius Biozone from Dob’s Linn. This section can be similarly
subdivided, with his horizons 33-31 containing a characteristic triangulatus fauna.
Many forms continue from the previous biozone. New appearances include
Orthograptus bellulus and IPristiograptus concinnus, Monograptus communis communis and
M. c. rostratus, in the upper part of the zone. A number of new taxa from the revolutus/austerus
group appear in this biozone; M. revolutus bicornis, and in the middle to upper part, M. r.
sequens and M. r. praecursor. The middle to upper parts of the biozone are marked by the
incoming of Petalolithus minor, Rastrites peregrinus, Glyptograptus tamariscus acutus, G.
alternis, 'Clim acograptus ' tamariscoides and Pseudo glyptograptus rhayaderensis. The
possibility exists of subdivision within the triangulatus Biozone. Monograptus triangulatus
Page 3.27

predecipiens, M. t. extremus? and Lagarograptus acinaces are confined to the lower part of the
biozone. Significant numbers of taxa appear in the middle to upper part of the biozone; these
mainly include monograptids and glyptograptids such as Monograptus triangulatus major, M.
communis communis, Monograptus communis rostratus, Glyptograptus elegans and G.
tamariscus acutus, as well as Neodiplograptus magnusl and Pribylograptus leptothecal Taxa
indicative of this biozone have been recovered from the Gala Group of south-west Scotland
(Stone 1995).
Diplograptus magnus Biozone
Jones (1909) originally recognised a ‘magnus band’ from the Rheidol Gorge section. Later
studies revealed a magnus Biozone fauna from the Machynlleth area, Wales (Jones & Pugh
1916). The biozone is mainly identified by the appearance of Diplograptus magnus (Hutt 1974,
Rickards 1976, Baker 1981, Zalasiewicz & Tunnicliff 1994, Davies et al. 1997), accompanied
by new appearances (Fig. 3.6.3) including Monograptus pseudoplanus (Rickards 1976, Hutt
1974, Baker 1981), P seudoglyptograptus vas (Hutt 1974), Pseudo glyptograptus barriei
(Zalasiewicz & Tunnicliff 1994, Davies et al. 1997) and Monograptus chrysalis (Zalasiewicz &
Tunnicliff 1994). Previous studies (Hutt 1974) have placed the incoming of
Clinoclimacograptus retroversus in this biozone; however, it is now thought to originate in the
upper cyphus Biozone (Rickards 1976). Monograptus triangulatus fimbriatus and M. t. major
are common associates, while M. t. triangulatus may persist into the lower part of the biozone
(Baker 1981).
Many taxa continue from previous biozones, but it is interesting to note that the rastritids
and petalolithids, that appeared in the triangulatus Biozone elsewhere in the U.K., initially occur
in this zone at Llanystumdwy (Baker 1981). This biozone represents the M. triangulatus
subzone (‘M .’ gregarius Biozone) of Elies & Wood (1913). However, the boundaries they
drew between the three *M. ’ gregarius subzones do not appear as reliable as those defined by
other authors (Sudbury 1958, Hutt 1974, Rickards 1976). Elies & Wood recorded the incoming
of 'Mesograptus ’ magnus from their underlying M. fimbriatus subzone, and subsequent work
suggests that several taxa they assigned to this level are indicative of the overlying ‘M. ’
argenteus subzone (= Pribylograptus leptotheca Biozone). These forms include Monograptus
Page 3.28

millepeda, M. lobiferus and M. clingani. The upper part of the magnus Biozone is poorly
represented in the U.K., commonly being represented by barren strata (Baker 1981,
Zalasiewicz 1990). Stone (1995) recognises the magnus Biozone in the Rhins of Galloway
district, southern Scotland.
Pribylograptus leptotheca Biozone
This biozone was initially identified in Northern England, where it was called the argenteus
Biozone (Nicholson & Marr 1888). Later, a similar fauna from this interval in Wales (at
Machynlleth), was referred to as the leptotheca Biozone (Jones & Pugh 1916), and this has
been generally adopted as the name for the biozone. Both species have been recorded in all areas
of Britain (Hutt 1974, Baker 1981, Davies et al. 1997), and although there are variations in
relative abundance, the presence of Pribylograptus leptotheca and/or Monograptus argenteus
characterise the base of the zone. New taxa noted in this biozone include Pristiograptus regularis
(Toghill 1968), and Pristiograptus jaculum (Baker 1981); Hutt (1974) identified the incoming of
the monoclimacids at this level in the Lake District.
A major feature of the leptotheca Biozone is the marked increase in Monograptus s.s.,
that is hooked monograptids (Rickards 1976). The following forms have been recorded: M.
millepeda Hutt 1974, Baker 1981), M. lobiferus (Rickards 1970, Baker 1981, Davies et al.
1997) and M. aff. undulatus (Rickards 1970). Several species continue from the previous
biozone (Figs. 3.6.2, 3.6.3), and ‘Monograptus’ gregarius gregarius is singled out by Hutt
(1974) as being very common. The fauna from Toghill’s (1968) horizon 29 represents this
biozone. Among the forms appearing in this biozone are P. folium, Glyptograptus serratus and
Monograptus cerastus. This biozone is also equivalent to the Monograptus argenteus subzone of
Elies & Wood (1913). Although they describe the incoming of Pribylograptus leptotheca,
Monograptus argenteus, Pristiograptus regularis and Pristiograptus jaculum at this level, their
list of species appearances seems to contain a mixture of forms recorded from both higher and
lower levels by other authors.
At many localities, for example, central Wales (Zalasiewicz 1990, Davies et al. 1997),
leptotheca Biozone assemblages occur in isolated, fossiliferous strata within a generally barren
Page 3.29

sequence. This makes subdivision difficult, although more continuously fossiliferous sequences
are found outside the U.K.
Monograptus convolutus Biozone
This biozone was initially described at Skelgill by Nicholson & Marr (1888); however, the
broad features of the zone have been agreed upon by most recent authors (Rickards 1976, Hutt
1974, Toghill 1968, Baker 1981). A number of taxa appear at or around the base of the
biozone, including Monograptus clingani, M. decipiens decipiens, Cephalograptus cometa
cometa, Cephalograptus tubulariformis and M. convolutus. Glyptograptus incertus s.s. appears
at this level. Zalasiewicz (1996) adds Torquigraptus urceolinus to the list of appearances
defining the base of the biozone, although this species is rare and at present has only been
recorded from the lower part of the biozone.
At Dob's Linn, this biozone represents Lapworth's (1878) original clingani band (lower
part of convolutus Biozone) and cometa Zone (upper part of convolutus Biozone). Baker (1981)
records Lapworthograptus knockensis from this biozone at Llanstumdwy.
Current data show that other incoming taxa include Monoclimacis crenularis, Rastrites
phleoides, Rastrites approximatus geinitzi, ?Pribylograptus jonesi and Monograptus urceolus.
Many species and subspecies continue from previous biozones (Figs. 3.6.2, 3.6.3). Several
workers (Toghill 1968, Rickards 1976, Hutt 1974, Davies et al. 1997) record the incoming of
Cephalograptus cometa extrema as a reliable indicator of the upper part of the convolutus
Biozone. A general subdivision of the biozone has been suggested (Zalasiewicz 1996, Davies et
al. 1997), with a possible lower convolutus Biozone containing Orthograptus bellulus, Rastrites
spina sensu Rickards 1970, Torquigraptus urceolinus, Glyptograptus tamariscus aff. angulatus
and Monograptus lobiferus aff. harpago. Elies & Wood (1913) adopted a convolutus Biozone
in their Silurian range charts, which broadly agrees in faunal content with subsequent studies,
though their species list is unreliable in detail.
Stimulograptus sedgwickii Biozone
This biozone faunal assemblage was originally described by Lapworth (1878), as the M.
spinigerus Zone (= M. sedgwickii) from the upper Birkhill Shale of Dob’s Linn. He recorded a
Page 3.30

characteristically diverse fauna, and distinguished an upper and lower subdivision within the
biozone. The basic sedgwickii Biozone fauna includes Stimulograptus sedgwickii, Rastrites
hybridus, Glyptograptus tamariscus, Petalolithus folium and Pseudoclimacograptus hughesi.
The lower zone fauna is distinguished by the presence of Monograptus clingani, Monograptus
runcinatus, Monograptus leptotheca and Climacograptus scalaris. The upper zone fauna is
distinguished by the presence of Monograptus proteus, Monograptus hisingeri, Monograptus
attenuatus, Monograptus intermedius, Monograptus becki, Diplograptus sinuatus and Retiolites
perlatus (Figs. 3.6.3, 3.6.4).
Many authors note the abundance of Stimulograptus sedgwickii and the appearance of
Lagarograptus tenuis, as characteristic of the base of this biozone (Hutt 1974, Rickards 1976,
Davies et al. 1997). Other new taxa include, Glyptograptus sinuatus crateriformis,
Pribylograptus argutus sequens, Glyptograptus packhami, Rastrites fugax, Pristiograptus
variabilis, Petalolithus kurcki and Pristiograptus nudus (Rickards 1970, 1976, Hutt 1974).
Several forms seem indicative of the middle part of the biozone: Monograptus communis
obtusus, 'Climacograptus' simplex, Rastrites sp. A of Rickards 1970 and Glyptograptus spp.
of Rickards 1970 (Rickards 1970, 1976).
Hutt (1974) refers to the characteristic abundance of Glyptograptus incertus at the base
of this biozone. It has been remarked that L. tenuis may be locally restricted to the lower part of
the biozone (Rickards 1976, Davies et al. 1997). This is the case with the Rhayader Mudstones
of central Wales (Davies et al. 1997), where L. tenuis is replaced in the upper part of the
Stimulograptus sedgwickii Biozone by Stimulograptus halli and associated species, indicating
the Stimulograptus halli Biozone. Rickards (1976) notes the appearance of M. knockensis s.I.
and M. ?spiralis as typical of this biozone. However, Bjerreskov (1975) attributed Rickards' M.
knockensis s.I. specimen to M. lobiferus harpago, while M. spiralis has been widely
misidentified in the past (J. A. Zalasiewicz pers. comm.).
Stimulograptus halli Biozone
Work subsequent to Lapworth’s studies at Dob’s Linn (1878), in the Machynlleth-Llyfnant area
of central Wales (Jones & Pugh 1916), led to the recognition of the Stimulograptus halli
Biozone. This biozone encompassed the upper part of the sedgwickii Biozone. The halli
Page 3.31

Biozone was disregarded by Rickards (1976). He did not believe the sedgwickii Biozone fauna
sufficiently distinctive to warrant subdivision and, owing to difficulties in identification between
Stimulograptus sedgwickii and Stimulograptus halli, recommended that it be removed from the
U.K. biostratigraphic scheme. Subsequently, Loydell (1991a) was able to firmly differentiate
halli Biozone faunas in the Cwmsymlog Formation of western mid-Wales, and reinstated the
biozone. Loydell (1992-93) records that species defining the base of the biozone are
Stimulograptus halli, Pristiograptus pristinus, Rastrites linnaei and M etaclimacograptus
undulatus (Fig. 3.6.4). However, Davies et al. (1997) only distinguished the halli Biozone in
the Llanilar-Rhayader area at a few localities, due to identification difficulties. As well as the
basal indicator species outlined by Loydell (1992-93), the updated range charts show that the
halli Biozone contains a well defined fauna, with several taxa originating at the base of the zone,
many of which are confined to it: Rastrites schaueri, Monograptus capillaris sensu Loydell, M.
admirabilis and Petalolithus praecedens. A number of relatively long-ranging species also occur:
these include Monograptus capis, Torquigraptus involutus, Stimulograptus sedgwickii and
Oktavites contortus.
Spirograptus guerichi Biozone
Formerly, the Rastrites maximus Subzone represented the lower part of the turriculatus
Biozone, first being recognised at Dob’s Linn (Lapworth 1878). However, Rickards (1976)
remarked that the maximus Subzone does not occur at the type locality of the turriculatus
Biozone at Browgill, Lake District (Nicholson & Marr, 1888). Rickards (1970, 1976) identified
the maximus Subzone in the Howgill Fells, and believed it to be an important part of the
turriculatus Biozone, although the subzone itself was defined only on the presence of the
nominal species. Toghill (1968) described a maximus Subzone fauna from the upper Birkhill
Shales, and Hutt (1974-75) recorded one locality containing Ra. maximus in the Lake District.
Loydell (1991a & b) did not use the maximus Subzone for the Anglo-Welsh sequence, although
he did record the species from Scotland; he divided the turriculatus Biozone into six subzones
based on his work in western mid-Wales. This biostratigraphic subdivision was further
enhanced when he divided the former turriculatus Biozone into two, creating a lower
Spirograptus guerichi Biozone followed by an upper Spirograptus turriculatus Biozone (Loydell
Page 3.32

1993-4). His six subzones remained in place, with the biozonal boundary occurring mid utilis
Subzone. Loydell (1993-4) defined the guerichi Biozone as ‘the interval from the first
appearance of Spirograptus guerichi to that of Spirograptus turriculatus,' having observed that
Sp. turriculatus itself does not appear until the middle of the old turriculatus Biozone. Davies et
al. (1997) remark that not all of Loydell's subzones could be consistently recognised in the
Llanilar/Rhayader area. As expected with a refined biostratigraphic subdivision, Loydell's
(1992-93) guerichi and turriculatus zones and component subzones are defined by the
appearance or relative abundance of the index species (Figs. 3.6.4, 3.6.5).
Paradiversograptus runcinatus Subzone
The base is delineated by the appearance of Pa. runcinatus and Sp. guerichi, with the nominal
species being abundant (Loydell, 1992-93). Loydell states that the numbers of Pa. runcinatus
have been enhanced by current sorting, but that this does not invalidate the subzone, as it can be
recognised in Scandinavia. Davies et al. (1997) could not separately differentiate the subzone in
the Llanilar/Rhayader district, and assigned some localities to a runcinatus to renaudi interval.
As the range charts show, Monograptus gemmatus, Pristiograptus renaudi, Streptograptus
plumosus, Streptograptus pseudoruncinatus and Petalolithus altissimus all appear at the base of
this subzone (Fig. 3.6.4).
‘Monograptus ’ gemmatus Subzone
Loydell (1992-93) described a diverse, distinctive fauna including S trep tograp tus
pseudoruncinatus, Spirograptus guerichi, Rastrites fugax, Glyptograptus fastigatus, G.
tamariscus, and M onograptus ' gemmatus. In addition to these, several species appear at the
base of the subzone and are confined to it, for example, the petalolithids, Pe. elongatus, Pe.
globosus and Pe. regius, with Glyptograptus auritus (Fig. 3.6.4). This subzone has been
separately recognised by Davies et al. 1997.
Pristiograptus renaudi Subzone
This is characterised by abundant Pristiograptus renaudi and Streptograptus strachani (Loydell,
1993), though neither of these taxa originate at the base of this subzone. However, Loydell
Page 3.33

records that Torquigraptus planus and T. cavei appear here, and are useful indicators of the
renaudi to utilis subzone interval. Other new species include M onograptus marri and
Petalolithus hispanicus (Figs. 3.6.4, 3.6.5). Both Loydell (1991) and Davies et al. (1997) point
out that the sequence from the gemmatus Subzone into this subzone has not yet been recognised
in a continuous section, and only one faunal assemblage from the Llanilar/Rhayader district has
been referred to this interval (Davies et al. 1997).
Spirograptus turriculatus Biozone
In its current, more biostratigraphically refined usage, the base of the turriculatus Biozone is
defined by 'the first appearance of Spirograptus turriculatus to that of 'Monograptus' crispus,
or other species indicative of the crispus Biozone' (Loydell, 1992-93). In this context,
Spirograptus turriculatus is taxonomically distinct from Spirograptus guerichi (Loydell et al.
1993), which characterises the previous biozone. As a result, older definitions for the base of
the turriculatus biozone (Rickards 1970, 1976, Hutt 1974-75) are no longer applicable, since
these now refer to the guerichi Biozone. Loydell's (1992-93) definition of the base of
turriculatus Biozone places it in the middle of the Stimulograptus utilis Subzone (Fig. 3.6.4).
Stimulograptus utilis Subzone
Loydell (1991) erected this subzone as the fourth subdivision of the turriculatus Biozone.
Subsequently (Loydell 1992-93), he found it possible to divide the utilis Subzone into two
parts, marking the boundary between the guerichi and turriculatus Biozones. This was based on
the incoming of Petalolithus tenuis, Spirograptus turriculatus and Streptograptus johnsonae,
and an abundance of Monograptus marri and S. plumosus in the middle of the subzone. The
range chart also indicates that several species seem to disappear mid-subzone, including
Petalolithus kurcki, Rastrites linnaei, Paradiversograptus runcinatus, Petalolithus conicus and
Pseudo streptograptus williamsi (Fig. 3.6.4). Davies et al. (1997) recorded the utilis subzone
widely in the Llanilar/Rhayader district.
Page 3.34

Streptograptus johnsonae Subzone
This subzone is defined on the abundance of Streptograptus johnsonae and common
Monograptus bjerreskovae and M. marri (Loydell 1991, 1992-93), and has been recognised in
the Llanilar/Rhayader area (Davies et al. 1997). Appearances at the base of the subzone are rare,
including only Petalolithus schaueri and Pristiograptus schucherti (Loydell 1992-93). As the
range charts demonstrate (Figs. 3.6.4, 3.6.5), a significant proportion of the johnsonae
Subzone fauna is composed of relatively long-ranging species.
Torquigraptus proteus Subzone
Torquigraptus proteus appears at the base of this subzone, and its presence and abundance
characterises this interval (Loydell 1991, 1992-93). As other authors record (Zalasiewicz 1994,
Davies et al. 1997), appearances also include Monograptus rickardsi and Monoclimacis?
galaensis, as well as Pristiograptus huttae (Loydell 1992-93). The bulk of this relatively small
fauna contains taxa from previous biozones and subzones (Figs. 3.6.4, 3.6.5). Notably,
Loydell (1992-93) records Monograptus priodon from the proteus Subzone in Wales.
Torquigraptus camicus Subzone
Zalasiewicz (1994) recognised a further faunal subzonation in central Wales, representing the
highest part of the turriculatus Biozone sequence. He distinguishes the base by the appearance
of T. camicus, Streptograptus whitei, Streptograptus exiguus and Monoclimacis? galaensis
(Fig. 3.6.5), although Davies et al. (1997) record Monoclimacis? galaensis in the proteus
Subzone of the same area. Streptograptus storchi is noted as abundant in the camicus Subzone
(Zalasiewicz 1994, Davies et al. 1997). The range charts show that much of this fauna consists
of elements from previous intervals. Zalasiewicz (1994) reported the origination of
Monograptus clintonensis and the rare occurrence of Monograptus aff. crispus here.
Monograptus crispus Biozone
Initially described from the Lake District (Marr & Nicholson 1888), this biozone is generally
recognised by the incoming of M. crispus and M. discus, together with associated forms,
although Zalasiewicz (1994) records M. aff. crispus in the cam icus Subzone (turriculatus
Page 3.35

Biozone). Rickards (1976) characterises the base of the biozone on the recognition of
Monograptus crispus and the zonal faunas above and below it (Fig. 3.6.5). He also remarks on
the appearance of M. discus, Retiolites geinitzianus geinitzianus and Retiolites g. angustidens.
Hutt (1974-75) added Streptograptus exiguus exiguus and Monograptus sp. et subsp. nov. C to
her Lake District list of diagnostic taxa. However, in central Wales, the boundary between the
crispus Biozone and the underlying turriculatus Biozone is not easily resolved (Zalasiewicz
1994, Davies et al. 1997), as both characteristic species are rare at the base of the biozone.
Despite this problem, Zalasiewicz (1994) was able to divide the crispus Biozone into three
subzones in the Llanilar/Rhayader district, a scheme taken up by Davies et al. (1997) in their
survey of the area. Loydell (1992-93) assigned only one locality in the Aberystwyth Grits
Formation to the crispus Biozone.
Monoclimacis? galaensis Subzone
Zalasiewicz (1994) refers to this interval as a partial-range subzone, delineated at the base by the
overlap of Monoclimacis? galaensis and Torquigraptus camicus with Monograptus crispus and
Monograptus discus. This description does not precisely follow the definition of a partial-range
zone (Harland et al. 1972); however, it does recognise the appearance of the eponymous
species and M onograptus discus as characteristic of the base of this subzone, and
Monoclimacis? galaensis and Torquigraptus camicus do disappear towards the top (Fig. 3.6.5).
Zalasiewicz (1994) also notes Monograptus clintonensis and Streptograptus exiguus at this level
in central Wales. These species relationships are clearly seen in the range charts.
Monograptus crispus Subzone
Zalasiewicz (1994) also describes this subzone as a partial-range subzone, defined by the
overlap of Monograptus crispus and Streptograptus loydelli. Again, the usage of partial-range
zone is imprecise, although Monograptus crispus is abundant in this subzone, disappearing at
the top of the interval, and the incoming of Streptograptus loydelli delineates the base (Fig.
3.6.5). This subzone has been recorded at Cwm Ystwyth in the Llanilar/Rhayader area by
Davies et al. (1997).
Page 3.36

Streptograptus sartorius Subzone
The base of this subzone is represented by the incoming of Streptograptus sartorius with
Torquigraptus pragensis pragensis and Torquigraptus pragensis ruzickai?, and the
disappearance of Monograptus crispus (Zalasiewicz 1994). This distinction is clearly observed
in the range charts (Fig. 3.6.5). Davies et al. (1997) also record this subzone at Cwm Ystwyth
and define the interval as an interregnum. This is not strictly correct, as the base of the subzone
is defined by the appearance of the index species and associated taxa, not solely on the
disappearance of Monograptus crispus .
Monoclimacis griestoniensis Biozone
First recognised by Wood (1906) in the Talerddig Grits of Tarannon, Wales, this biozone has a
characteristic fauna, containing many new species. It is recognised by the occurrence of the
zone fossil, Mel. griestoniensis of Elies & Wood (Fig. 3.6.5), which Rickards (1976) remarks
as abundant. He also notes the first definite record of Monograptus priodon at this level. Both
Davies et al. (1997) and Zalasiewicz (1994) recognise the base of this biozone in the
Llanilar/Rhayader district, on the appearance of Monoclimacis griestoniensis s.s. and Mcl.
cf .griestoniensis sensu Elies & Wood. They also suggest that the griestoniensis Biozone can be
informally subdivided on the basis that Mcl. cf .griestoniensis of Elies & Wood is more
abundant in the lower part of the zone, while Monoclimacis griestoniensis s.s. dominates the
upper part (Zalasiewicz 1990). As the range charts demonstrate, this biozone contains a number
of previously undescribed taxa including, M onoclim acis d irecta , M o n o g ra p tu s
pseudo com m unis and Streptograptus aff. loydelli of Zalasiewicz (Zalasiewicz 1994,
Zalasiewicz et al. 1995), which are confined to this level, as well as Torquigraptus pergracilis.
Monoclimacis crenulata Biozone
Initially referred to as the Monograptus crenulatus Biozone by Wood (1906) from Tarannon,
she defined the zone on the appearance and abundance of the zone fossil (=Monoclimacis
crenulata), associated with a generally low diversity, undistinctive fauna. Subsequent studies
have confirmed this (Fig. 3.6.5), with the assemblage consisting mainly of forms from
previous biozones (Davies et al. 1997, Zalasiewicz 1994), including Monograptus discus and
Page 3.37

Torquigraptus tullbergi. However, the incoming of Monoclimacis vomerina vomerina is a
diagnostic indicator of the base of this biozone (Rickards 1976, Zalasiewicz 1994, Davies et al.
1997). The Monoclimacis crenulata Biozone formerly included all UK strata from the top of the
Monoclimacis griestoniensis Biozone to the base of the Wenlock Series. Rickards (1976) noted
that this zone was rarely found abroad, where a more refined biostratigraphic subdivision could
be recognised (Boucek 1953, Bjerreskov 1975). Since then, work by Loydell & Cave (1993,
1996) has led to the recognition of more biozones within this interval, and the current crenulata
Biozone now represents the lowest part the original zone.
Oktavites spiralis Biozone
Graptoloid faunas representing the Oktavites spiralis Biozone were not recognised in the UK
until relatively recently (Loydell & Cave 1993,1996). Initial biostratigraphic work at Buttington
Brick Pit, central Wales (Loydell & Cave 1993) and subsequent studies in eastern mid-Wales
(Loydell & Cave 1996) resulted in division of all but the lowest part of the long-established
crenulata Biozone, and allowed direct comparison with the graptolite biozonation of Bohemia
(Boucek 1953). Oktavites spiralis Biozone has a distinctive faunal assemblage (Figs. 3.6.5,
3.6.6), and Loydell & Cave (1993, 1996) and Zalasiewicz (1994) record the introduction of
several taxa at the base. These include the zone fossil with: M onoclimacis hemmanni,
Streptograptus? anguinus, Monoclimacis geinitzi, 'Monograptus' nodifer, Barrandeograptus?
bomholmensis, Retiolites venosus and Monograptus vesiculosus. Other notable appearances
are Monograptus parapriodon and Monoclimacis linnarssoni. Zalasiewicz (1994) and Davies et
al. (1997) indentified this biozone in the Llanilar/Rhayader area of central Wales, noting that
Oktavites spiralis itself is rare.
Cyrtograptus lapworthi Biozone
The Cyrtograptus lapworthi Biozone was originally recorded by Loydell & Cave (1996) from
Banwy River, central Wales. It represents the strata between that of the underlying spiralis
Biozone and the Cyrtograptus insectus Biozone above, and roughly equates with the
Stromatograptus grandis Biozone of the Czech Republic. The authors point out that these
biozones are not directly correlatable as the British lapworthi Biozone also includes the top of
Page 3.38

the Czech spiralis Biozone. As yet, Loydell & Cave's work is the only published report of this
zone in the UK; however, they describe a relatively broad and characteristic faunal assemblage
(Figs. 3.6.5, 3.6.6). Several new species at the base of the biozone, excluding the zone fossil,
are confined to its lower and middle parts, allowing for potential subdivision; these include
M onoclim acis sublinnarssoni, M onograptus speciosus, Streptograptus wimani and
Pristiograptus largus. A number of taxa continue from the previous zone, and a few originate
mid-biozone, notably Monoclimacis vomerina basilica and Barrandeograptus pulchellus. C.
lapworthi itself marks the first appearance of the cyrtograptids. In the Llanilar/Rhayader district
of central Wales, Davies et al. (1997) assign strata overlying the spiralis Biozone to a
'spiralis/'centrijugus interregnum'. The fauna here contains only long-ranging graptoloids such
as the vomerinids, priodon-type monograptids and Retiolites geinitzianus.
Cyrtograptus insectus Biozone
The biostratigraphic succession outlined by Loydell & Cave (1996) from Banwy River included
the first, and so far only, recorded occurrence of the Cyrtograptus insectus Biozone in the UK.
As these authors indicated (see Figs. 3.6.5, 3.6.6), the biozone contains a number of taxa
whose ranges are limited to the lower-middle or middle-upper parts of this interval, suggesting
the possibility of further biostratigraphic subdivision. This biozone is notable for the
introduction of the mediograptids, and Mediograptus flittoni appears at the base together with
Monoclimacis shottoni, C. insectus and Cyrtograptus sp. of Loydell & Cave. Monoclimacis
linnarssoni disappears in the lower part of the zone. Species characterising the middle and upper
parts of the biozone include Mediograptus morleyae, Monograptus pseudo cult ellus and
Mediograptus sp. aff. inconspicuus of Loydell & Cave.
Cyrtograptus centrijugus Biozone
In Britain, the C. centrijugus Biozone was first identified in the Howgill Fells, northern
England (Rickards 1967). Rickards (1967, 1976) describes a fauna containing large numbers of
robust cyrtograptids, including C. centrijugus, also monoclimacids and restricted
monograptids. The base of this biozone is now widely assumed to be coincident with the base
of the Wenlock Series in the UK, although as Loydell & Cave (1996) point out, it cannot be
Page 3.39

correlated with the basal boundary stratotype section. The incoming of the zone fossil defines
the biozonal base, though several other species also appear for the first time, some of which are
confined to this interval (Rickards 1976, Williams & Zalasiewicz in press, Davies et al. 1997).
Additions include Mediograptus danbyi, Mediograptus cautleyensis, Monoclimacis kettneri,
Monograptus simulatus, Monoclimacis? sp. 1 of Williams & Zalasiewicz and Monoclimacis
adunca. The range charts show (Figs. 3.6.5, 3.6.6) that M ediograptus inconspicuus and
Monograptus flexuosus appear in the middle to upper part of the biozone. A significant
proportion of the faunal assemblage of this biozone consists of relatively long-lived taxa
ranging up from previous zones (Loydell & Cave 1996, Williams & Zalasiewicz in press,
Davies et a l 1997).
Cyrtograptus murchisoni Biozone
In many areas of Britain, the murchisoni Biozone faunal assemblage is often indistinguishable
from the underlying centrijugus Biozone when the eponym is absent (Rickards 1967, 1976,
Williams & Zalasiewicz in press, Davies et al. 1997). The biozone was initially recognised by
Lapworth (1880) from Builth Wells, and contains several cyrtograptid species, including the
type species, Monograptus priodon and Monoclimacis vomerina. Many graptolite workers
remark that the base of this biozone is recognised principally on the appearance of C.
murchisoni (Williams & Zalasiewicz in press, Davies et al. 1997, Loydell & Cave 1996).
Recent studies in central Wales (Loydell & Cave 1996, Williams & Zalasiewicz in press) have
led, though, to the recognition of several newly recognised taxa which range from the base of
the biozone, a number of which are confined to the lower part of the biozone, suggesting the
potential for finer biostratigraphic subdivision. Incoming species are Monoclimacis vikensis,
Pristiograptus praedubius, Monograptus radotinensis radotinensis, Cyrtograptus murchisoni
bohemicus, IPseudoplegmatograptus wenlockianus and Monograptus firmus s.l. (Fig. 3.6.6).
Loydell & Cave (1996) note that species diversity drops markedly by the upper part of the
biozone, this is clearly shown in the range chart (see Fig. 3.6.6).
Page 3.40

Monograptus firrnus Biozone
The rapid decline in taxa diversity and abundance in the upper part of the murchisoni Biozone,
coupled with the incoming of M. firmus in the the Banwy River section, central Wales, resulted
in Loydell & Cave (1996) recognising the firm us Biozone for the first time in the UK.
Approximately seven species, most of which have long biostratigraphical ranges, including M.
priodon, M. vomerina vomerina and possibly M. v. basilica, are found at this level (Figs.
3.6.5, 3.6.6). Loydell & Cave (1996) identified Pristiograptus dubius dubius from the biozone
and notably, the disappearance of IP. wenlockianus in the lower half of the biozone. This
interval represents the lower part of the traditional Monograptus riccartonensis Biozone.
Although as yet, only Loydell & Cave (1996) have confirmed the existence of the firm us
Biozone in the British Isles, other authors observe significantly lower species diversity and
abundance at the base of the riccartonensis Biozone (Rickards 1976, Williams & Zalasiewicz in
press). Williams & Zalasiewicz (in press) reported the appearance of rare M. firmus s.l. from
strata which they assigned to the upper part of the murchisoni Biozone at Builth Wells in Wales,
and which may correlate with the firmus Biozone in the Banwy River.
Monograptus riccartonensis Biozone
The riccartonensis Biozone was first recognised in the UK by Elies (1900) at Builth Wells,
south-central Wales. It has since been recognised in many areas of Britain, and notable studies
include Rickards' (1967, 1969) work in northern England and the Davies et a l (1997) survey
of the Llanilar/Rhayader district of central Wales. Williams & Zalasiewicz (in press) have
recently re-examined the Builth Wells sequence. The base of the biozone is generally taken at
the incoming of Monograptus riccartonensis, the biostratigraphic range of which delineates the
biozone (Fig. 3.6.6). Other taxa first appearances include Monograptus radotinensis inclinatus
and Pristiograptus latus, the latter being limited to the lower part of the biozone. The middle to
upper part of this biozone is represented by the incoming of several species, such as
Monograptus firm us sedberghensis, M onograptus antennularis, M onograptus flexilis
belophorus, Pristiograptus meneghinii meneghinii and possibly Monoclimacis flumendosae
flumendosae. Rickards (1976) notes the possibility of further subdivision of the riccartonensis
Biozone, and in an earlier work (Rickards 1967) in the Howgill Fells, he describes a local,
Page 3.41

three-fold subdivision, representing, in general, increasing species diversity and abundance
through the biozone.
Pristiograptus dubius Biozone
Williams & Zalasiewicz (in press) recognise the dubius Biozone in the UK for the first time at
Builth Wells, south-central Wales. They refer to this interval as an interregnum, typified by
long-lived taxa from the Pristiograptus dubius group, as well as forms of Monograptus priodon
and Monograptus flemingii, and Monoclimacis 'flumendosae'. However, a number of species
do originate in this biozone, as shown in the range charts (Fig. 3.6.6) they include:
Monograptus retroflexus, Monograptus priodon-flemingii intermediate forms, and in the middle
of the zone, Pristiograptus dubius pseudolatus. Recognition of this horizon is largely reliant on
recognition of the biozones directly above and below it (Williams & Zalasiewicz in press). A
dubius Biozone fauna had already been defined in the Czech Republic (Boucek 1960), and
Rickards (1967, 1969) found similar faunal evidence for what he called the Monograptus
antennularis Zone in northern England, roughly comparable to Boucek's (1960) interval, and
including the upper part of the former riccartonensis Biozone. Rickards did not however,
incorporate this biozone formally into his review of British graptolite biostratigraphy (Rickards
1976). Davies et al. (1997) describe a faunal assemblage at the top of their riccartonensis
Biozone in the Llanilar/Rhayader district, central Wales, similar to that outlined by Williams &
Zalasiewicz (in press), but make no subdivision of the zone.
Cyrtograptus rigidus Biozone
The base of the rigidus Biozone is defined by the appearance of the nominal fossil, which is
limited to this biozone (Rickards 1976, Williams & Zalasiewicz in press) (Fig. 3.6.7).
Formerly taken to equate with Elies' (1900) symmetricus Zone from her Builth Wells section
(Rickards 1976), recent work on this section has shown that Elies' linnarssoni Zone can also be
encompassed in the rigidus Biozone as defined by Williams & Zalasiewicz (in press).
At Builth Wells, the rigidus Biozone is overlain by the Cyrtograptus lundgreni Biozone,
and the intervening Cyrtograptus linnarssoni and Cyrtograptus ellesae biozones cannot be
identified (Williams & Zalasiewicz in press). Reports elsewhere in the UK of the linnarssoni
Page 3.42

{=flexilis) Biozone were dependent upon recognition of C. linnarssoni, rarely documented and
commonly mis-identified (M. Williams pers. comm.), and sporadic Monograptus flexilis
(Rickards 1976). However, new work (Williams & Zalasiewicz in press, cf. Rickards 1976)
has revealed that M. flexilis can be shown to originate in the dubius Biozone of their
terminology, and that C. linnarssoni, which is an ill-defined taxon, forms part of a more
distinctive rigidus faunal assemblage together with the zone species. Williams & Zalasiewicz (in
press) did not distinguish any rigidus subspecies in the Builth Wells area; therefore, the
definition of their rigidus Biozone does not exactly match that of Rickards (1976), which
includes the appearance of Cyrtograptus rigidus cautleyensis (see range chart). Davies et a l
(1997) describe a lower linnarssoni (-flexilis) Biozone fauna containing C. r. cautleyensis from
Mynydd-Llys, central Wales, although this can be included in the revised rigidus fauna of
Williams & Zalasiewicz (in press).
Cyrtograptus lundgreni Biozone
In the Builth Wells area, where the Wenlock graptoloid biozonal scheme was first defined, the
lundgreni Biozone overlies the rigidus Biozone without the intervening linnarssoni or ellesae
Biozones (Williams & Zalasiewicz in press). These authors could not recognise a distinct
ellesae zonal assemblage from Elies' (1900) original sections at Builth Wells, and found the
zonal index C. ellesae to occur above C. lundgreni. As identification of the ellesae Biozone was
largely dependent on the recognition of the type species, it could not be wholly relied upon to
represent a discrete biozone, although as the range chart shows, the ranges of C. ellesae and C.
lundgreni do overlap (Warren 1971, Rickards 1976, Bassett et al. 1975). Davies et a l (1997)
were not able to distinguish an ellesae Biozone fauna in the Llanilar/Rhayader area, nor were
White et a l (1992) in the Southern Uplands.
The lundgreni Biozone fauna is distinctive, containing several incoming taxa which
form the bulk of the assemblage, including the zone fossil, which seems to be restricted to this
horizon (Fig. 3.6.7). Other additions are Cyrtograptus cf. urbanekii, Cyrtograptus ellesae,
Monoclimacis flumendosae kingi, Cyrtograptus lundgreni gracilis, Testograptus testis and
Paraplectograptus eiseli. This interval also has a discrete middle to upper zone fauna suggesting
the potential for finer subdivision, including Cyrtograptus ramosus, Gothograptus nassa,
Page 3.43

Pristiograptus aff. ja eg eri of Holland et al., Pristiograptus sp. 1 of Holland et al. and
Cyrtograptus cf. hamatus. Davies et a l (1997) recorded this biozone from central Wales, as did
Bassett et al. (1975). It has also been recognised in north Wales (Warren 1971) and Scotland
(White et al. 1992).
Gothograptus nassa Biozone
The nassa Biozone was first adopted for use regarding British strata by Warren (1971) in north
Wales, and demonstrates a dramatic decline in graptoloid species diversity from the preceding
lundgreni Biozone (Rickards 1976, Williams & Zalasiewicz in press). Indeed, only five taxa
have been recorded (Figs. 3.6.6, 3.6.7), one of which is a new occurrence. The whole fauna
consists of P. dubius dubius, abundant G. nassa (although very rare at Builth Wells), P. aff.
jaegeri of Holland et al. 1969, and appearing in the upper part of the biozone, Pristiograptus
jaegeri. Davies et al. (1997) found this faunal assemblage in the Llanilar/Rhayader district,
marking the youngest graptolitic strata in the area. Williams & Zalasiewicz (in press) refer to a
nassa-ludensis Biozone representing the uppermost interval of Wenlock strata at Builth Wells.
Both indicator species are rare in this area and their biostratigraphic ranges cannot be clearly
delineated.
Monograptus ludensis Biozone
The base of the ludensis Biozone is taken at the incoming of the zonal species (Fig. 3.6.7)
which co-occurs in the lower part of the interval with taxa from the preceding G. nassa
Biozone. This zonal assemblage indicates the possibility of a more refined biostratigraphic
subdivision (see Rickards 1976), as all other species first occurrences range from the middle to
upper part of the biozone. These include Holoretiolites (.Balticograptus) lawsoni, Pristiograptus
auctus, Monograptus deubeli and M onograptus aff. uncinatus orbatus. The loss of P. aff.
jaegeri of Holland et al. 1969, mid way through the biozone, reinforces the distinction between
a lower and an upper fauna within the ludensis interval.
Page 3.44

Neodiversograptus nilssoni Biozone
In recent years, little biostratigraphic work has been carried out on British Ludlow Series strata.
As a result, the biozonal criteria for the series have not changed significantly since Rickards'
review of Silurian graptolite bio stratigraphy (1976). The base of the N. nilssoni Biozone
coincides approximately with the base of the Ludlow Series, and faunas of this type were first
recognised by Barrande (1850) from Bohemia. Many Lower Ludlow taxa had been identified in
the UK, notably from the Lake District and Ludlow itself (Nicholson 1868, Hopkinson 1873),
before Lapworth (1880), working in central Wales and the Welsh Borderlands, designated the
'zone of Monograptus nilssoni', which included the whole Lower Ludlow. Further studies in
Wales, the Welsh Borderlands and northern England, most importantly that of Wood (1900),
led to subdivision of this broad biostratigraphic unit and a refined faunal definition for the
nilssoni Biozone (Rickards 1967, 1969, Warren 1971). As the range chart clearly shows (Fig.
3.6.7), this nilssoni interval displays a much greater species diversity than the preceding
Wenlock biozones, with approximately 27 taxa, compared to 9 found in the P. ludensis
Biozone. The base of the nilssoni Biozone is delineated by the appearance of the zone fossil,
with true Monograptus uncinatus orbatus, Plectograptus macilentus, Spinograptus spinosus,
the first saetograptids, Saetograptus colonus colonus and Saetograptus varians various, and
Bohemograptus bohemicus s.l. Of all the species in this biozone, only the latter three taxa occur
throughout the biozone. The other forms in this list, together with G. nassa, M. ludensis and P.
auctus which disappear mid-nilssoni Biozone, comprise a distinct lower to middle zone fauna.
This subdivision is strongly reinforced by a highly distinguishable and diverse faunal
assemblage which appears in the middle to upper part of the biozone and contains many new
genera and species of Monograptidae, notably Cucullograptus progenitor, Cucullograptus
scanicus, M onoclim acis m icropom a, M onoclim acis haupti, Pristiograptus vicinus,
Critinograptus critinus and Lobograptus simplex. The saetograptids diversify rapidly in this
upper zone interval with the incoming of Saetograptus roemeri, Saetograptus fritschi fritschi,
Saetograptus chimaera salweyi, Saetograptus leintwardinensis incipiens, Saetograptus
wandalensis, Saetograptus chimaera chimaera and Saetograptus chimaera semispinosus.
M ost recent British authors have recognised the potential for biostratigraphic
subdivision within the nilssoni Biozone (Warren 1971, Rickards 1976), as has been formally
Page 3.45

acknowledged in Poland and the Czech Republic, where a lower nilssoni Biozone is succeeded
by an upper progenitor Biozone (Urbanek 1966, Teller 1969, Pribyl 1983). Although not as yet
formally proposed for the UK, a C. progenitor Biozone could promise to be a valuable
biostratigraphic horizon, particularly in Wales where separate faunas can be clearly
distinguished (BGS unpublished memoir, Montgomery). Rickards (1967, 1976) notes that the
lower nilssoni Biozone fauna has not been identified from northern England. The occurrence of
so many new taxa in the n ilsson i Biozone suggests a graptoloid evolutionary 'high1
representing the base of the Ludlow Series.
Cucullograptus (Lobograptus) scanicus Biozone
Wood (1900) established the scanicus Biozone in her paper on the Lower Ludlow Formation of
Wales and the Welsh Borderland. Few new species appear at this interval; those that do occur
mid-biozone (Fig. 3.6.7), so this interval is characterised by the abundance of C. scanicus and
S. c. chimaera (Wood 1900, Rickards 1976). Most taxa present in scanicus Biozone originated
in the underlying N. nilssoni Biozone. The incoming of Pristiograptus tumescens tumescens,
Saetograptus clunensis, Pristiograptus welchae and Bohemograptus bohemicus tenuis in the
middle part of the scanicus interval distinguishes the upper part of the biozone. In his review of
British graptolite biostratigraphy, Rickards (1976) notes that prior to this study, the nilssoni and
scanicus biozones were often amalgamated due to difficulties in the recognition of a distinct
scanicus Biozone faunal assamblage; however, he expected this 'lumping' to decrease with
continued recognition of the biozone. Most recent graptoloid biostratigraphic work for the
Ludlow Series has been carried out by BGS as part of their remapping program in Wales (BGS
unpub.). Although some distinction between the nilssoni and scanicus faunas has been possible
at a number of localities, a significant proportion of BGS registered localities have been
assigned a nilssoni - scanicus biozonal age, indicating that, despite extensive work, there are
still problems with clear identification of the base of the scanicus Biozone.
Saetograptus incipiens/Pristiograptus tumescens Biozone
Wood (1900) also initiated this graptolite biostratigraphic interval following her extensive
studies in Wales and the Welsh Borderland. The biozonal name varies locally according to the
Page 3.46

relative abundance and/or the presence or absence of Saetograptus incipiens and Pristiograptus
tumescens, both of which originate in earlier biozones. Graptoloid species diversity is greatly
reduced in the incipiens/tumescens Biozone compared with previous Ludlow zones, with
approximately 10 confirmed taxa (Fig. 3.6.7). Only one form appears in this biozone,
Saetograptus? aff. incipiens (Wood 1900). A number of species disappear in the lower to
middle part of the interval, indicating the possibility for local biostratigraphic subdivision, these
include M. haupti, S. clunensis, P. vicinus and S. v. varians.
Saetograptus leintwardinensis Biozone
This biozone was initially defined by Marr (1892) in the Lake District and has been widely
recognised in the UK (Wood 1900, Holland et al. 1963, Rickards 1967), largely on the
incoming of the type species (Fig. 3.6.7). Only two other confirmed taxa occur in this biozone,
they are S a e to g ra p tu s l aff. in c ip ien s , which continues from the previous zone, and
Saetograptus leintwardinensis primus, which is confined to the lower-middle part of the
biozone (Shergold & Shirley 1968). Such a low diversity fauna reflects an evolutionary crisis
amongst British graptoloids at this time.
Bohemograptus proliferation Biozone
This biozone was originally described from the UK by Holland and Palmer (1974), when they
identified Bohemograptus bohemicus tenuis from strata overlying the leintwardinensis Biozone
in the Welsh Borderlands. This taxon is the youngest graptoloid in the UK biostratigraphic
sequence, and the sole representative of this Biozone (Fig. 3.6.7). Despite appearing in the
scanicus Biozone, B. b. tenuis has not been positively reported from the leintwardinensis
Biozone (Rickards 1976).
Page 3.47

Chronostratigraphy(Proposed Standard, after Fortey e t al. 1995)
Ordovician Graptoloid Biostratigraphy of the U.K.
Syst
emSe
ries
Stage/Sub-StageEngland & Wales
Sub-ZoneScotland
Graptoloid Biozone Graptoloid Biozone
Ord
ovic
ian
Ash
gill
Himantian G lyptograptus persculptus G lyptograptus persculptus Clim acog ? extraordinarius
Rawtheyan
Cautleyan
Pusgillian
D icellograptus anceps
Paraorthog. pacificus t
Dicellograptuscomplexus
D icellograptus anceps
( D icello graptus com planatus)
D icellograptuscomplanatus
(P leurograptus linearis) Pleurograptus linearis
Car
adoc
StreffordianOnnian
ActonianD icranograptus clingani
Dicellograptusmorrisi
Climacograptuscaudatus
- D icrano graptus clinganiCheneyan
MarshbrookianWoolstonian
Burrellian Soudleyan D iplograptus multidensC lim acograptus wilsoni
Hamagian Clim acograptus peltifer
AurelucianCostonian
Velfreyan N em agraptus gracilis N em agraptus gracilis
Lla
nvim
Llandeilian G lyptograptusteretiusculus
(Darriwilian 3)
(Darriwilian 1 - 2)
Abereiddian
D idym ograptusm urchisoni
D idym ograptus artus
Are
nig
FennianA ulograptus cucullus
Isograptus gibberulus(Yapeenian 1-2)
WhitlandianCastlemainian 3
Isograptus victoriae (Castlemainian 1-2)
Moridunian
Corymbograptus simulans Chewtonian 1 -2Corymbograptus varicosus (Bendigonian 3 - 4 )
Bendigonian 1-2
Tetragraptusphyllograpto ides Lancefieldian 3
Trem
adoc
Migneintian A raneograptus m urrayi (Lancefieldian 2)
Cressagian
Fig. 3.1. Proposed Ordovician chronostratigraphy, after Fortey et al. 1995, and comparison of Ordovician
graptoloid biostratigraphy for England & Wales and Scotland. Biozones in brackets have not been
conclusively identified in these respective areas.Fig. 3.1

Chronostratigraphy(Global Standard, after Holland et al. 1989)
Biostratigraphy
System Series Stage Graptoloid Biozone Sub-ZoneP
rido
li
U.K. sequence undivided No later biozones
recorded in the U.K.
£o
Ludfordian Bohemograptus proliferation Zone
Saetograptus leintwardinensis
3hJ
Saetograptus incipiens
Gorstian Lobograptus scanicus
Neodiversograptus nilssoni
c03• t—HU3
Pristiograptus ludensis
Homerian Gothograptus nassa• CO Mo Cyrtograptus lundgreni
"c Cyrtograptus rigidusPristiograptus dubius
SheinwoodianMonograptus riccartonensisMonograptus firmusCyrtograptus murchisoniCyrtograptus centrijugusCyrtograptus insectus Cyrtograptus lapworthiOktavites spiralis 1Streptograptus sartorius
Telychian Monoclimacis crenulata Monograptus crispus
Monoclimacis griestoniensis Monograptus crispus Spirograptus turriculatus
1 Monoclimacis? galaensis
e*<D/ Torquigraptus camicus
Torquigraptus proteus Streptograptus johnsonae
Spirograptus guerichi \> Stimulograptus utilisOStimulograptus halli \ Pristiograptus renaudi
c3 Stimulograptus sedgwickii \ Monograptus gemmatus
Aeronian Monograptus convolutus \ Paradiversog. runcinatus
Pribylograptus leptothecaDiplograptus magnusMonograptus triangulatus
Coronograptus cyphus
Rhuddanian Lagarograptus acinaces Atavograptus atavusParakidograptus acuminatus
Fig. 3.2. Silurian chronostratigraphy, after Holland et al. 1989, and accompanying graptoloid biostratigraphy
for the UK, this study.
Fig. 3.2

Silurian Graptoloid Biostratigraphy of the U.K.Rickards, 1976
Sub-Zone
Rastrites maximus
Graptoloid Biozone
No later biozones recorded in the U.K.
Bohemograptus proliferation Zone
Saetograptus leintwardinensis
Saetograptus incipiens
Lobograptus scanicus
Neodiversograptus nilssoni
Monograptus ludensis
Gothograptus nassaCyrtograptus lundgreni Cyrtograptus ellesaeCyrtograptus linnarssoni Cyrtograptus rigidus
Monograptus riccartonensis
Cyrtograptus murchisoni Cyrtograptus centrijugus
Monoclimacis crenulata
Monoclimacis griestoniensis Monograptus crispus
Monograptus turriculatus
Monograptus sedgwickii
Monograptus convolutus Pribylograptus leptotheca Diplograptus magnusMonograptus triangulatus
Coronograptus cyphusLagarograptus acinaces Atavograptus atavusOrthograptus acuminatus
Glytograptus persculptus
<uDC
31/5
J£O,eou.£?A-w</3Oe2jsU
§
£T33
oO
§'SBoa
e2-3oo£
'34300
g£
ccd• fHeo<d<
g§T3343P4
This Study
Graptoloid Biozone
No later biozones recorded in the U.K.
Bohemograptus proliferation Zone
Saetograptus leintwardinensis
Saetograptus incipiens
Lobograptus scanicus
Neodiver so graptus nilssoni
Pristiograptus ludensis
Gothograptus nassa
Cyrtograptus lundgreni
Cyrtograptus rigidus Pristiograptus dubius Monograptus riccartonensis Monograptus firmus Cyrtograptus murchisoni Cyrtograptus centrijugusCyrtograptus insectus Cyrtograptus lapworthi Oktavites spiralisMonoclimacis crenulata Monoclimacis griestoniensis Monograptus crispus Spirograptus turriculatusSpirograptus guerichiStimulograptus halli Stimulograptus sedgwickii Monograptus convolutusPribylograptus leptotheca Diplograptus magnus Monograptus triangulatus
Coronograptus cyphus Lagarograptus acinaces Atavograptus atavusParakidograptus acuminatus
ORDOVICIAN
Sub-Zone
Streptograptus sartoriusMonograptus crispus Monoclimacis? galaensisTorquigraptus camicusTorquigraptus proteus Streptograptus johnsonaeStimulograptus utilisPristiograptus renaudi Monograptus gemmatus Paradiversog. runcinatus
Fig. 3.3. Comparison of Silurian graptoloid biostratigraphic schemes for the UK, Rickards (1976) and this
study.
Fig. 3.3

SYMBOLS : X ranges through whole biozone
L found in lower part of biozone
M found in middle part of biozone
U found in upper part of biozone
A species is abundant
? doubtful identification of taxon
§ stratigraphic horizon in which taxon occurs is doubtful
AremgTremadoc lanvim
Monduman /Whitlandiaiy Fennian / Abereiddian
1 Araneograplus murrayi (Hall, 1865)/ V. q.
Xv , A O , V
7, 102 Acrograptus? cf. sinensis (Lee & Chen, 1962) • X , 7, 223 Didymograptus sp. (declined) o f M olyneux & Rushton, 1988 . X , ? 7,224 Acrograptus? protobalticus Monsen, 1937 X 75 Temnograptus multiplex (Nicholson, 1868) ' X ? 76 Acrograptus? rigoletto (Maletz et al., 1991) * X 7,217 Tetragraptus quadribrachiatus (Hall, 1858) , x X x , x , x X , 78 Tetragraptus (Pendeograptus) fruticosus (Hall, 1858) cf. X X 79 Tetragraptus amii Elies & Wood, 1902 X? u? X 7 X X 7
1 0 Tetragraptus pseudobigsbyi Skevington, 1965 7 ? ; X X X ' X 711 Baltograptus vacillans attenuatus (Monsen, 1937) ' X 71 2 Didymograptus aff. balticus s.l. Tullberg, 1881 X 7,2513 Trochograptus diffusus Holm, 1881 X § 1414 Acrograptus fdiform is Tullberg, 1880 X X ' 715 Corymbograptus varicosus Wang, 1968 • i X ' X 716 Didymograptus cf. decens Tornquist, 1889 L , L 7,2517 Tetragraptus reclinatus Elies & Wood, 1902 X X X 7 7,3318 Corymbograptus cf. kunmingensis Ni, 1979 u 719 Dichograptus octobrachiatus sedgwickii Salter, 1863 u 1 X ? X 1 ? 72 0 Corymbograptus deflexus Elies & Wood, 1901 u X L X , cf. 7, 28,332 1 Schizograptus tardifurcatus Elies, 1898 u ; X , ? X , u? , 72 2 Dichograptus separatus Elies, 1898 u? X X 723 Pseudophyllograptus angustifolius (Hall, 1858) u ' x 1 X X ' X X 7,2824 Expansograptus similis Hall, 1865 cf. 1 § 725 Adelograptus divergens (Elies & Wood, 1902) , X 726 Isograptus cf. primulus Harris, 1933 X 727 Didymograptus (D idym ograptellus) minutus Tornquist, 1879 X 728 Tetragraptus postlethwaitei Elies, 1898 1 X 1429 Tetragraptus pendens Elies, 1898 X 1 ■ 730 Acrograptus gracilis (Tornquist, 1891) , X , , 7,25,2831 Corymbograptus v-fractus (Salter, 1863) § 1432 Holograptus deani Elies & Wood, 1902 § 1433 Azygograptus eivionicus Elies, 1922 ' 1 1 X 1 L 2,3,3334 Acrograptus infrequens Kraft, 1973 X L 7,2535 Pseudobryograptus cumbrensis Elies, 1898 X L 8,2636 Acrograptus kurcki (Tornquist, 1901) X L 7,2537 Didymograptus simulans Elies & Wood, 1901 X ' L ’ cf. ' cf. 10,28, 3338 Tetragraptus crucifer (Hall, 1858) X X 1 739 Expansograptus cf. praenuntius Tornquist, 1901 i ; X X , § 3,7,28, 3340 Azygograptus lapworthi N icholson, 1875 § § § 2
41 Pseudotrigonograptus ensiformis (Hall, 1858) X ' X ' X ' 7 742 Loganograptus logani (Hall, 1858) X 1 X X ' X 743 Dichograptus octobrachiatus (Hall, 1858) X , 7 , X , X 744 Tetragraptus serra (Brongniart, 1828) X X X x , ? 7,2445 Pseudophyllograptus densus (Tornquist, 1879) X 7,2546 Isograptus victoriae victoriae Harris, 1933 1 ’ X 10
47 Isograptus victoriae cf. maximus Harris, 1933 X ■ 1948 Corymbograptus? uniformis lepidus Ni, (1979) x x 3, 1649 Expansograptus extensus linearis Monsen, 1937 X X 3,7, 10/1650 Tetragraptus headi (Hall, 1858) X X 751 Pseudotrigonograptus minor (Mu & Lee, 1958) X ' X ' ? 752 Expansograptus goldschm idti (Monsen sensu Kraft, 1977) X X , ? 15, 10/1653 Expansograptus hirundo (Salter, 1863) X X , ? 3, 10/16, 28, 37
Fig. 3.4.1. Ordovician graptoloid biostratigraphic range charts for England & Wales.
Fig. 3.4.1

Series Arenig Llanvirn jC aradoc
Stage W hitlandian/ Fennian / Abereiddian / / Aurelucian
Sr' i
-S’4"$, . &g, $<
' r & *? <? •*? O'64 Isograptus caduceus gibberulus (Nicholson, 1875) X 3,7 , 10,1955 Isograptus caduceus ssp. nov. ° X - 756 Isograptus caduceus imitatus Harris, 1933 : X , , 7, 1957 Expansograptus distinctus (Harris & Thomas, 1935) X 3,7 , 1658 Pseudisograptus n.sp. A o f Jenkins, 1982 X 759 Corymbograptus? uniformis Elies & Wood, 1901 X ' 7, 1660 Pseudisograptus angel Jenkins, 1982 ! X . 1 3,761 Pseudisograptus dumosus (Harris, 1933) : 5 , s 362 Xiphograptus svalbardensis (Fortey & Archer, 1974) X ? 763 Expansograptus nitidus (Hall, 1858) ? ! X ' X 7,2864 Corymbograptus v-fractus volucer (H.O. Nicholson, 1890) ‘ X 1 X 1 7, 10/1665 Expansograptus suecicus (Tullberg, 1880) : Cf. , cf. , 286 6 Corymbograptus cf. inflexus (Chen & Xia, 1979) cf. cf. 2867 Tetragraptus bigsbyi (Hall, 1865) X X 7 7 ' 7 ,2 8 ,3 26 8 Tham nograptus’ doveri Nicholson, 1875 ! u 1 769 Eoglyptograptus dentatus (Brongniart, 1828) X X L? ' 7, 16,26,2870 Acrograptus nicholsoni planus Elies & Wood, 1901 X 7, 1671 Undulograptus sinensis (Mu & Lee, 1958) X 7,2572 Bergstroemograptus sp. X 7,2573 Cryptograptus hopkinsoni (Nicholson, 1869) X 1674 Glyptograptus shelvensis Bulman, 1963 X Cf. : 16, 2875 Didymograptus protobifidus Elies, 1933 X L , 1276 Expansograptus sparsus (Hopkinson, 1875) X L 16, 2877 Aulograptus cucullus (Bulman, 1932) X X 16,2678 Pseudoclimacograptus cumbrensis Bulman, 1963 X X 1 16,2679 Cryptograptus antennarius (Hall, 1865) X X : , 16,2480 Acrograptus affinis (Nicholson, 1869) X X 7, 16,2681 'Didymograptus' sp. A o f Skevington, X X 15, 1682 Oelandograptus austrodentatus anglicus (Bulman, 1963) X X 1 1 7,2683 Acrograptus acutidens Elies & Wood, 1901) u X : 16,26,2884 Amplexograptus confertus (Lapworth, 1875) u? LM , L , 3, 27 ,28,3285 Cryptograptus tricornis schaeferi Lapworth, 1880/tricomis s.l. o f Hughes, 1989 u? X L X 7 3 ,1 7 ,2 6 ,2 8 ,3 28 6 Paraglossograptus sp. L : 7,2587 Expansograptus robustus (Ekstrom, 1937) X 268 8 Climacograptus angustatus Ekstrom, 1937 X , 2589 Didymograptus acutus Ekstrom, 1937 X 2690 D iplograptus ellesi Bulman, 1963 X 3,2691 Didymograptus cf. dubitatus Harris, 1935 X 1 (92 Pseudophyllograptus glossograptoides (Ekstrom, 1937) X : 26,2893 Glossograptus armatus (Nicholson, 1869) X 7,2894 Azygograptus coelebs Lapworth, 1880 X 295 Didymograptus spinulosus Perner, 1895 X * 3 ,496 Climacograptus tailbertensis Skevington, 1970 X ‘ 1 7 ,1 6 ,2 697 Holmograptus lentus (Tornquist, 1911) X ; 7,25, 2698 Glossograptus acanthus Elies & Wood, 1908 X , 7,2899 Didymograptus stabilis Elies & Wood, 1901 X 4,26 ,28
1 0 0 Glossograptus fim briatus (Hopkinson, 1872) x : X ' X X 7,17 ,281 0 1 Didymograptus p luto Jenkins, 1963 X 20?, 281 0 2 Didymograptus nicholsoni nicholsoni Lapworth, 1875 x , , 3 ,7 ,25103 Didymograptus 'bifidus'Hall, 1865 X ? 26, 32104 Didymograptus geminus (Hisinger, 1840) X ? 3,5105 D idymograptus pakrianus Jaanusson, 1960 Xcf. 1 7 26, 32106 Pseudophyllograptusl nobilis (Harris & Keble, 1932) ! ? ? 25
\ t<£\
Fig. 3.4.2. Ordovician graptoloid biostratigraphic range charts for England & Wales.
Fig. 3.4.2

Series
Stage
Arenig Llanvirn Caradoc
Whitlandian / Fennian / Abereiddiann / y / / / / // M / 4 / ‘S / D -c lingan i
■r-v & Mi..if, '/sM .M,J r it. M/f/107 D idymograptus artus Elies & Wood, 1901 ------ r f
1 X L? ’ 28108 Expansograptus euodus Lapworth, 1875 1 X L ! X? 15,26,32109 Didym ograptus cf. miserabilis Bulman, 1931 , X X , 7 ,26 ,28 , 291 1 0 Lonchograptus sp. of Rushton, 1993 M 4, 16,281 1 1 Trichograptus fragilis (Nicholson, 1869) M 31 1 2 D iplograptus hollingworthi Skevington, 1970 1 M 26, 38113 Pseudoclimacograptus angulatus (Bulman, 1953) ■ M 7,26114 Pseudoclimacograptus scharenbergi (Lapworth, 1876) , MU X , X X X 7115 Phyllograptus cf. typus Hall, 1858 ’ § ’ ? MU L? 28,31 ,32 , 36116 Nicholsonograptus fasciculatus (Nicholson, 1869) ‘ U 37117 Didymograptus nanus Lapworth, 1875 ■ u L ■ 7, 25, 26, 27118 Pterograptus elegansl Holm, 1881 L : 7119 Pseudoclimacograptus angulatus magnus (Berry 1964) X 321 2 0 Pseudoclimacograptus angulatus m icidusl (Berry, 1964) X 321 2 1 Didymograptus murchisoni (Beck, 1839) X 321 2 2 Didymograptus speciosus Ekstrom, 1937 X? ‘ 5, 6 ,20 ,28 , 32123 Orthograptus calcaratus priscus Elies & Wood, 1907 : 1 x , L? 13, 32124 Lasiograptus retusus Lapworth, 1880 LM L 7125 Pseudoclimacograptus angulatus sebyensis Jaanusson, 1960 X L 5126 Cryptograptus tricornis (Carruthers, 1859) X [ X X L 7, 17127 Diplograptus foliaceus (Murchison, 1839) 1 X . X X X 7,28,31128 Diplograptus decoratus (Harris & Thomas, 1935) MU , LM 5,7 , 17,28 ,31 ,34129 Amplexograptus caelatus (Lapworth, 1875) M X 7, 17130 Climacograptus brevis Elies & Wood, 1906 U ' X X X cf. 7,29131 Glyptograptus teretiusculus euglyphus Lapworth, 1880 X? 5,7 , 17, 28,29, 34, 36132 D icellograptus divaricatus divaricatus (Hall, 1859) X M 31133 Climacograptus antiquus Lapworth, 1873 X X X X 7, 17,28,29134 D icellograptus geniculatus Bulman, 1932 MU 5,7 , 17 ,28 ,29 ,31 ,36135 Dicranograptus irregularis Hadding, 1913 MU L 7,17136 D icellograptus intortus Lapworth, 1880 MU X L 7,17 ,28137 Glyptograptus teretiusculus (Hisinger, 1840) MU L LM 7,17 ,2 8 ,2 9138 Climacograptus sheldoni (Hughes, 1989) U L 7, 17,28,29,31139 Nemagraptus cf. subtilis Hadding, 1913 U X 7,17140 Pseudoclimacograptus modestus (Ruedemann, 1908) U X L 7,17141 Dicellograptus divaricatus salopiensis Elies & Wood, 1904 U X LM 5 ,7 ,1 7 ,2 8 ,3 4 , 36142 Dicellograptus cambriensis Hughes, 1989 U LM LM 7 ,17 ,28 ,29 , 34143 Dicranograptus brevicaulis Elies & Wood, 1904 U LU X 7,17 ,28144 Dicranograptus rectus Hopkinson, 1872 U X X 7, 17,28,29145 Nemagraptus gracilis (Hall, 1847) X 7, 28,29146 Leptograptus validus validus Elies & Wood, 1903 X 7, 17,28,29147 Orthograptus uplandicus (Wiman, 1895) X 7, 17,28,29148 Expansograptus? superstes (Lapworth, 1876) X 28149 Orthograptus whitfieldi (Hall, 1859) X 7,29150 Dicranograptus furcatus minimus Lapworth, 1876 X 7151 Dicranograptus ramosus (Hall, 1847) X 7152 Hallograptus mucronatus (Hall, 1847) X ? 7153 Dicellograpraptus sextans exilis Elies & Wood, 1904 X L 7154 D iplograptus leptotheca Bulman, 1946 X LM 7,29155 Dicellograptus sextans sextans (Hall, 1843) X X 5,7 ,2 8156 Lasiograptus harknessi costatus (Lapworth, 1873) X X X 29157 Orthograptus apiculatus (Elies & Wood, 1907) LU? X Laff. 5 ,17 ,28 ,29 , 36158 G lossograptus hincksii hincksii (Hopkinson, 1872) MU LM 29,36159 *Corynoides curtus Lapworth, 1876 MU X X 5, 17, 28, 34, 36
v v - * v \ % W V ^\ t
* The biostratigraphic range o f Corynoides curtus continues up to the morrisi Subzone.
Fig. 3.4.3. Ordovician graptoloid biostratigraphic range charts for England & Wales.
Fig. 3.4.3

Series Caradoc / Ashgill
Stage A u r e lu c ia iy / y ^ h e n e y a y 1 S t r e ff o r d ia n f ^
/ £? / Dicranograptus / / & / ^ / clingani / / ■% /
/ £ / 4/ /
C / £ / 4 f
/ /$ / &
/ /
/AA *
. ?
■ H - t
/ i , j s t 4? • <?
A ? , , A , , A A A / • & ' £
: i / ^i i / S
i t / 3
& i /160 Lasiograptus pusillus Ruedemann, 1947 u '
,7, 34
161 Amplexograptus perexcavatus (Lapworth, 1876) U ' X 1 5 ,7 ,2 9162 Amplexograptus arctus Elies & Wood, 1907 U : X , 5,29163 D iplograptus molestus Thorsland, 1948 u X 5,34164 Dicranograptus ramosus spinifer Elies & Wood, 1904 u X 5,7 , 17, 29, 34165 Orthograptus calcaratus acutus Elies & Wood, 1907 u ! X 1 7,29166 Climacograptus bicornis bicornis (Hall, 1847) u ■ X i 7, 17, 28 ,29 ,36167 Dicranograptus nicholsoni nicholsoni Hopkinson, 1870 u , x , X 7,29, 36168 Climacograptus cf. bekkeri (Opik, 1927) 7 ? 7169 Climacograptus cf. putillus (Hall, 1865) 7 7 1 7170 Glossograptus cf. ciliatus Emmons, 1855 ? 1 ? , i 7171 Pseudoclimacograptus isknos Zalasiewicz, 1992 X , , 7,34172 Dicranograptus ziczac Lapworth, 1876 X , 29173 Normalograptus sp. (bulbous virgella) o f Davies et al., 1997 X 5174 Glyptograptus plurithecatus Obut & Sobolevskaya, 1964 X ! 1 ! ! : : 34175 Climacograptus meridionalis (Ruedemann, 1947) ? , i , > 34176 Orthograptus amplexicaulis (Hall, 1847) X , X 1 x , L? , 7, 17,36177 Orthograptus ex. gr. calcaratus (Lapworth, 1876) X X X LM? 5,7 ,29 , 36178 Lasiograptus harknessi (Nicholson, 1867) X X X ? 5 ,7 ,2 9 , 34, 36179 Climacograptus miserabilis Elies & Wood, 1906 X ‘ Xcf. ' Xcf. ‘ X ’ ? ? ? 7, 36180 Diplograptus sp. o f Rickards, 1970 , M U ? i 5181 Orthograptus calcaratus tenuicornis Elies & Wood, 1907 M U 29,36182 Orthograptus calcaratus vulgatus Elies & Wood, 1907 MU X X 29, 36183 D icellograptus patulosus Lapworth, 1880 u 1 29184 Climacograptus sp. of Rickards, 1970 U ' 5185 Glyptograptus sp. o f Rickards, 1988 u , 5186 Dicranograptus nicholsoni minor Bulman, 1945 u 6,34187 Orthograptus calcaratus basilicus Elies & Wood, 1907 u ' MU X ’ Lcf. 7 ,2 9 ,3 6188 Amplexograptus compactus Elies & Wood, 1907 ! U 1 X ! X 7 ,29 ,3 6189 Climacograptus aff. mohawkensis Ruedemann, 1912 7 | cf. i ? . 5 ,7190 Dicranograptus clingani resicis W illiams, 1994 , 7 7 7191 Orthograptus cf. quadrimucronatus spinigerus (Lapworth, 1876) L 5192 Climacograptus aff. antiquus (broad) Lapworth, 1873 L 5193 Dicranograptus clingani Carruthers, 1868 X 5 ,7 ,2 9 ,3 6194 Climacograptus cf. caudatus Lapworth, 1876 X i 5195 Dicellograptus morrisi Hopkinson, 1871 L? X L 7 ,29 ,3 6196 D icellograptus flexuosus Lapworth, 1876 L? X ’ X? 7 ,29 ,36197 Neurograptus margaritatus (Lapworth, 1876) 1 Xsp L? 7, 36198 Climacograptus spiniferus Ruedemann, 1912 X L . 5,7 , 36199 Orthograptus pageanus micracanthus Elies & Wood, 1907 X x 72 0 0 Orthograptus truncatus pauperatus Elies & Wood, 1907 X X 72 0 1 Orthograptus quadrimucronatus quadrimucronatus (Hall, 1865) X X ’ cf.? ’ 5 ,7 ,2 9 ,3 62 0 2 Climacograptus dorotheus Riva, 1976 u X 7, 36203 G lyptograptus daviesi W illiams, 1982 X , X? , 7,36204 Corynoides ultimus Ruedemann, 1925 u , L 7,36205 Normalograptus (broad form) o f Zalasiewicz et al., 1995 LM ' 7,36206 Dicellograptus anceps (Nicholson, 1867) 1 ? X ? 7207 Orthograptus truncatus abbreviatus Elies & Wood, 1907 i i ■ ? A 7 7208 Climacograptus supem us Elies & Wood, 1906 , ? , U , ? 7209 Climacograptus normalis Lapworth, 1877 ? X 7 1,7, 18,23,25, 30, 352 1 0 Climacograptus tuberculatus Nicholson, 1869 L 142 1 1 Climacograptus scalaris indivisus D avies, 1929 M 112 1 2 G lyptograptus? avitus D avies, 1929 X 11.25. 35
$ \\ \ *
\ \
\ *U'
Fig. 3.4.4. Ordovician graptoloid biostratigraphic range charts for England & Wales.
Fig. 3.4.4

Series Ashgill
Stage / // / „ / $ - /£ / 4 / / £ ' / £ ■& / / -S 'S . , / s ’•V M l / */ / /
213214215216217218
Glyptograptus persculptus Elies & Wood, 1907 X Atavograptus ceryx (Rickards & Hutt, 1970) 1 X Climacograptus parvulus (H. Lapworth, 1900) X 1 Climacograptus medius Tornquist, 1897 X Paraclim acograptus innotatus innotatus Nicholson, 1869 ? Akidograptus ascensus Davies, 1929 ?
3518351, 18,25,30,35 3018,30, 35
\ \ \ X \ \ % \%%- \ \ \ \
\ \ \ <1\
Fig. 3.4.5. Ordovician graptoloid biostratigraphic range charts for England & Wales.
Fig. 3.4.5

Series
Stage
Arenig Llanvim
tlandi
’ ^ ’f ' # ' .A V #^ s ^ Q * £a f & Y T V / C ?^ 4. 4, o d c j # < f , < f 45 4?)
1 Tetragraptus approximatus N icholson, 1873 § : X ‘ ------!------ 1— 42 Tetragraptus cf. decipiens T.S. Hall, 1899 i X , i i 2
3 Tetragraptus quadribrachiatus (Hall, 1858) § . X . 2,44 Tetragraptus (Pendeograptus) fruticosus (Hall, 1858) X 45 Acrograptus c i.fd iform is Tullberg, 1880 ! x : 2
6 Paradelograptus sp. A o f Rushton et al., 1986 X 1 1 2
7 Expansograptus"! aff. geom etricus Tornquist, 1901 , x , 48 Didymograptus cf. protomurchisoni Decker, 1944 X 1 49 Tetragraptus bigsbyi cf. askerensis Monsen, 1937 X 1 41 0 Sigmagraptus praecursor Ruedemann, 1904 ’ X ‘ 411 Tetragraptus cf. kindlei Ruedemann, 1947 ■ X 1 1 41 2 Expansograptus extensus (Hall, 1858) , X , i cf.i , 4,513 Tetragraptus serra (Brongniart, 1828) cf. X aff.§ 2 ,4 ,514 Paradelograptus sp. B o f Rushton et al., 1986 ’ X ‘ 2 ,415 Tetragraptus reclinatus Elies & Wood, 1902 1 X 1 ; cf.§ ; 4,516 Didymograptus cf. protoindentus Monsen, 1937 , X : , 417 Isograptus caduceus australis Cooper, 1973 • V 518 Isograptus caduceus imitatus Harris, 1933 § 519 Isograptus victoriae lunatus Harris, 1933 1 i § . 52 0 Pseudoisograptus dumosus (Harris, 1933) ‘ § 1 52 1 Pseudophyllograptus angustifolius (Hall, 1858) , , , 1 , 52 2 Corymbograptus cf. dejlexus (Elies & Wood, 1901) § 523 Didymograptus cf. protobifidus Elies, 1901 § 524 Tetragraptus amii Elies & Wood, 1902 ; § , 525 Tetragraptus pseudobigsbyi Skevington, 1965 , ■ § 1 526 Pseudoclimacograptus modestus (Ruedemann, 1908) . , X 3,627 Glyptograptus teretiusculus (Hisinger, 1840) L 1,328 Acrograptus? serratulus (Hall, 1847) i x ,
6
29 Nemagraptus explanatus pertenuis (Lapworth, 1876) ■ ■ X i 630 Amplexograptus compactus (Elies & Wood, 1907) , , , : Cf. , 6
31 Corynoides? pristinus Ruedemann, 1947 cf. 7 10
32 Thamnograptus scoticus Lapworth, 1876 ? 6
33 Dicellograptus patulosus Lapworth, 1880 X ' 3 ,6 , 1034 Dicellograptus alabamensis Ruedemann, 1947 i i L i 335 Hallograptus mucronatus (Hall, 1847) X 6
36 D icellograptus salopiensis Elies & Wood, 1904 X 3,637 Hallograptus bimucronatus (N icholson, 1869) : X ' 638 Thamnograptus capillaris Hall, 1859 ’ X ' 3,639 D icellograptus sextans (Hall, 1843) , , X § 640 Nemagraptus gracilis (Hall, 1847) X 3,641 Orthograptus calcaratus acutus Elies & Wood, 1907 X 7 6
42 Dicranograptus furcatus (Hall, 1847) : X 1 7 643 D icellograptus divaricatus s.l. (Hall, 1959) 1 f X . X 644 D icellograptus exilis Elies & Wood, 1904 , , X , X 6
45 Dicranograptus rectus Hopkinson, 1872 X X 6
46 Expansograptus? superstes (Lapworth, 1876)i x ,
X 3,647 Dicranograptus ramosus spinifer E lies & Wood, 1904 1 ■ X • X 1 , 6
48 Glyptograptus euglyphus Lapworth, 1880 ■ x , X 3,649 Dicranograptus furcatus minimus Lapworth, 1876 X X 6
50 Dicellograptus divaricatus rigidus Lapworth, 1880 X § 6
51 Dicranograptus brevicaulis Elies & Wood, 1904 ‘ X ’ § 3,652 Reteograptus geinitzianus Hall, 1859 ; i X , X 3,6
\ t
<£ Qt Q ' G ' «£' O ' <> i r z ai- YVVV V v \ ■ Y \ ' \\\V 4h: YYYVvWV'V vY V - V < /% ■ » v - . V
Fig. 3.5.1. Ordovician graptoloid biostratigraphic range charts for Scotland.
Fig. 3.5.1

Series Caradoc Ashgill
Stage Aureluciat / Burrellian/ Cheneyan / Streffordian /
usgillianD. clingani
/ /
4
/ // / ^5 //i/ s r/ h
/ * i
7W is" A /
7f t ;
.7 7 /4 • />/ < * / ■ $ / /
/0
53 Orthograptus whitfieldi (Hall, 1859) X cf.? 654 D iplograptus foliaceus (Murchison, 1839) 7 1 655 Dicellograptus intortus Lapworth, 1880 X 7 Lcf. . 3 ,6 , 1056 G lossograptus hincksii (Hopkinson, 1872) X 7 L 6, 1057 Pseudoclimacograptus scharenbergi (Lapworth, 1876) X X LM7U 3, 1058 Climacograptus bicornis (Hall, 1847) X X LM§ ’ ? 1,6, 1059 Climacograptus antiquus Lapworth, 1873 X § § 1L 1,6, 1060 Climacograptus brevis Elies & Wood, 1906 X § § X 1,6, 1061 Lasiograptus costatus Lapworth, 1873 X X X X 6,1062 Dicranograptus nicholsoni nicholsoni Hopkinson, 1870 X cf. X X cf. 6, 1063 Cryptograptus tricornis (Carruthers, 1858) X X : L 1 X X 1,3 ,6 , 1064 Lasiograptus harknessi (N icholson, 1867) X X ! X? X X 1,665 Dicranograptus ramosus (Hall, 1847) X X X , X X 6,76 6 Orthograptus amplexicaulis (Hall, 1847) X X X X X X 1,6 ,7 , 1067 Orthograptus calcaratus (group) X X X 1 X X X 66 8 Thamnograptus sp. 7 1069 Pseudoclimacograptus isknos Zalasiewicz, 1992 cf. 170 Amplexograptus perexcavatus (Lapworth, 1876) 7 § 7 671 Diplograptus leptotheca Bulman, 1946 7 X L 6, 1072 Dicranograptus ziczac Lapworth, 1876 1 7 X X X 6, 1073 Corynoides calicularis N icholson, 1867 X . X X 6, 1074 D icellograptus angulatus Elies & Wood, 1904 x X X 675 Leptograptus flaccidus (Hall, 1865) 7 7 ? X 676 Orthograptus calcaratus s.l. Lapworth, 1876 1 ; X X X X X 1,677 Climacograptus wilsoni Lapworth, 1876 X 6,1078 Orthograptus calcaratus vulgatus Elies & Wood, 1907 , X , X LM 679 Corynoides curtus Lapworth, 1876 X X X ? 680 Orthograptus ex gr. calcaratus X UM X LM 7,1081 Orthograptus truncatus intermedius Elies & Wood, 1907 X X X? 682 *Climacograptus m iserabilis Elies & Wood, 1906 X X X 6 ,7 ,983 D icellograptus flexuosus Lapworth, 1876 7 X X 7 1 ,6 ,7 , 1084 Diplograptus? pilatus W illiams, 1982 L 785 Climacograptus antiquus lineatus Elies & Wood, 1907 X 108 6 Coryoides incurvus Hadding, 1915 X 187 Orthograptus calcaratus tenuicom is Elies & Wood, 1907 X 18 8 D icellograptus moffatensis (Carruthers, 1858) X X? 6,789 Climacograptus caudatus Lapworth, 1876 X 1,6, 1090 Dicranograptus clingani Carruthers, 1868 MU : 791 Dicranograptus ramosus longicaulis Elies & Wood, 1904 X X 6
92 G lyptograptus daviesi W illiams, 1982 , X X 793 Orthograptus pageanus Elies & Wood, 1873 X X 7 6
94 Neurograptus fibratus (Lapworth, 1876) X X 7 6
95 D icellograptus carruthersi Toghill, 1970 X X X 6,796 Climacograptus spiniferus Ruedemann, 1912 , X LM X 6, 10
97 Amphigraptus divergens radiatus Lapworth, 1876 X X X 698 D icellograptus pumilus Lapworth, 1876 X X X 1,6 ,799 Orthograptus quadrimucronatus (Hall, 1865) X X X 1 ,6 ,7 ,1 0
1 0 0 Neurograptus margaritatus (Lapworth, 1876) ■ X X X 6,71 0 1 Climacograptus dorotheus Riva, 1976 X X X 6,71 0 2 Plegmatograptus nebula Elies & Wood, 1908 7 X LM 6,7103 Leptograptus flaccidus m acer Elies & Wood, 1903 7 X LMcf. 1,7104 Orthograptus pauperatus Elies & Wood, 1907 1 ? X X 6,7105 Climacograptus mohawkensis (Ruedemann, 1906) , ? X X 6,7
%\ > \ \ % \ \
\AV *
% - A - A \ ■,A \ & \A '
The biostratigraphic range o f Climacograptus miserabilis continues up into the Silurian.
Fig. 3.5.2. Ordovician graptoloid biostratigraphic range charts for Scotland.
Fig. 3.5.2

Series Caradoc
Cheneyan yZreffordi:
D. clingani /
Ashgill
Stage£ .
i 4f- .o V <-V > ^
/, iv .
106 Orthograptus calcaratus basilicus Elies & Wood, 1907 MU X L 6107 Orthograptus quadrimucronatus spinigerus Lapworth, 1876b ‘ U 1 X ; X ■ 1 ,6 ,7108 D icellograptus caduceus Lapworth, 1876 , X , 10109 Leptograptus flaccidus spinifer Elies & Wood, 1903 X 7 11 1 0 Climacograptus minimus (Carruthers, 1868), non Elies & Wood X Lcf. 11 1 1 D icellograptus morrisi Hopkinson, 1871 1 X : X 1 1 ,6 ,71 1 2 Climacograptus styloideus Elies & Wood, 1906 ■ U? i X 1 1 6113 Pleurograptus linearis linearis (Carruthers, 1858) ? X , 1 ,6 ,7114 Climacograptus tubuliferus Lapworth, 1876 aff. X X 1 ,6 ,7 , 8115 D icellograptus elegans rigens Elies & Wood, 1904 7 1 11116 D icellograptus elegans elegans (Carruthers, 1867) M ' 1 1 7117 Amphigraptus divergens divergens (Hall, 1859) M , , 7118 Leptograptus capillaris (Carruthers, 1868) M 7119 Leptograptus flaccidus macilentus Elies & Wood, 1903 X 111 2 0 Orthograptus socialis (Lapworth, 1880) ?M ' L 1 8
1 2 1 Dicellograptus complanatus Lapworth, 1880 L 1 ‘ 81 2 2 Orthoretiolites pulcherrimus (Keble & Harris, 1934) L , 1 8123 Dicellograptus alector Carter, 1972 L 8124 Glyptograptus occidentalis Ruedemann, 1947 L 8125 Dicellograptus minor Toghill, 1970 1 L X X 7 ’ 8,9126 Orthograptus fastigatus Davies, 1929 X L : 7 ,9127 Dicellograptus complexus D avies, 1929 X , LM , 7 ,9128 Dicellograptus aff. complexus X LM 7 ,9129 D icellograptus anceps (Nicholson, 1867) X X ? 9130 Pleurograptus lui Mu, 1950 1 X X 1 7 i 7 ,9131 Climacograptus supem us Elies & Wood, 1906 , X X . 7 , 7 ,9132 Orthograptus abbreviatus Elies & Wood, 1907 X X 7 7 ,9133 Plegmatograptus? craticulus Williams, 1982 X X 7 7 ,9134 Climacograptus normalis Lapworth, 1877 X ! X X ’ X 7 ,9135 Climacograptus latus Elies & Wood, 1906 U : X 7 i 7 ,9136 Orthoretiograptus denticulatus Wang et al., 1977 u , x , 7 7 ,9137 Plegmatograptus? lautus Koren & Tzaj, 1980 ? L 7,9138 Nymphograptus velatus Elies & Wood, 1908 L 1 7 ,9139 Paraorthograptus pacificus (Ruedemann, 1947) ' X 7 7 ,9140 Dicellograptus ornatus Elies & Wood, 1904 MU ; 7 . 9141 Climacograptus? extraordinarius (Sobolevskaya, 1974) u LM 7,9142 Glyptograptus? pseudovenustus cf. pseudovenustus (Legrand, 1976) LM 6,9143 Glyptograptus? avitus Davies, 1929 1 ’ MU 6,9144 G lyptograptus cf. persculptus o f Elies & Wood, 1907
V r\' -Si' .^A r-Y aT' MU
r\'9
\
Fig. 3.5.3. Ordovician graptoloid biostratigraphic range charts for Scotland.
Fig. 3.5.3

S i l u r i a n G r a p t o l o i d B i o s t r a t i g r a p h y o f t h e 2 , tg - B ' S' B
' 3CL' &
« 1 <« 1 3 1 « 3 s , 3 i 9- , 2 P.S a. 2 fl &ca tv o cd
3 ._3
U . K . ( R i c k a r d s , 1 9 7 6 )&o
o acum
int
Ata
vogr
atav
us
Lag
arog
acin
aces
Cor
onoj
cyph
us
Mon
ogr
tria
ngul
Dip
logr
mag
nus
Prib
yloj
lept
othe
Mon
ogr
conv
olu
Mon
ogr
sedg
wic
Series Llandovery
Stage Rhuddanian Aeronian
/ /
? /
/ / , > / £ / £ . # / $ /< $ . / /
i "Climacograptus" tuberculatus N icholson, 1869 X 1 f ! ■ 4, 152 Glyptograptus avitus Davies, 1929 X 1 3, 15, 183 G lyptograptus persculptus (Elies &Wood, 1907) L , , , 184 Atavograptus ceryx (Rickards & Hutt, 1970) L 65 Diplograptus modestus parvulus (H. Lapworth, 1900) X 186 Climacograptus scalaris m iserabilis Elies &Wood, 1906 X > X X 6, 13, 15, 167 Climacograptus medius (Tornquist, 1897) X i k X X ! 1,6, 15,16,188 Climacograptus normalis (Lapworth, 1877) A , i X X , x? , L? , 1,6, 13,15, 16, 189 "Climacograptus" indivisus D avies, 1929 X 31 0 "Paraclimacograptus" innotatus innotatus Nicholson, 1869 s.l. X L ' 15, 1611 Parakidograptus acuminatus praematurus (Davies, 1929) : L 6, 131 2 Climacograptus trifdis Manck, 1923 : LM , 15, 1613 Parakidograptus acuminatus acuminatus (Nicholson, 1867) X(UA). 6,13, 15, 16, 1814 Akidograptus ascensus Davies, 1929 ’X(LA) ’ 6, 15, 16, 1815 D iplograptus modestus diminutus Elies &Wood, 1907 X ' X 1 ' 6,13, 14, 1516 Diplograptus modestus modestus Lapworth, 1876 ■ MU : X L ? ! i 6, 13, 15, 1617 Cystograptus vesiculosus (Nicholson, 1868) , MU X(I.A) , X X , 1,6, 13, 15, 1618 "Orthograptus" cabanensis Zalasiewicz & Tunnicliff, 1994 U 1819 Coronograptus cf. gregarius minusculus Obut & Sobolevskaya, 1968 Xs 152 0 Pristiograptus fragilis pristinus (Hutt, 1975) ! ?s 6, 152 1 Diplograptus modestus tenuis Rickards, 1970 : L 132 2 Atavograptus sp. 1 (of Hutt & Rickards, 1970) LM 723 Dimorphograptus elongatus Lapworth, 1876 L L 6, 15, 1624 Rhaphidograptus extenuatus (Elies &Wood, 1908) cf. 1 L ! 6,15,1625 Coronograptus cyphus praematurus (Toghill, 1968a) ' L? ! L? ' 15, 1626 Diplograptus sp. of Rickards, 1970 , Ji 1327 Atavograptus gracilis Hutt, 1975 X 6,15, 1828 Diplograptus? rarus Rickards, 1970 X X 13,1529 Climacograptus rectangularis (McCoy, 1850) ' X ; X X X 1 1,6, 13, 15,16, 1830 Atavograptus atavus (Jones, 1909) ■X(UA). X X , X , L? ' 1,6, 13, 15, 16, 1831 D iplograptus elongatus Churkin & Carter, 1970 M aff. , , X 6,1532 Dimorphograptus decussatus Elies & Wood, 1908 M 6,1633 Pribylograptus aff. incommodus sensu Hutt, 1975 : u : 634 Diplograptus? sp. (Hutt, 1974) ' U : , 635 Dimorphograptus erectus erectus Elies &Wood, 1908 : MU , X , LM , 6,14, 15, 1636 Dimorphograptus epilongissimus Rickards, 1970 u X X 13, 1437 Dimorphograptus longissimus (Kurck, 1882) U X ' X 6, 14,1538 Dimorphograptus physophora (N icholson, 1868) ‘ ?U 1 X ' X ; ! 1 ■ 6, 14,15, 1639 Dimorphograptus confertus confertus (Nicholson, 1868) . u . A X , L? , 1, 6, 13, 14, 15, 16, 1840 Rhaphidograptus toernquisti (Elies &Wood, 1906) , U" , x , x , X , X , X , X , L” 1,6,13,14, 15, 16, 1841 'Climacograptus' sp. o f Rickards, 1970 X 1342 M etaclimacograptus slalom Zalasiewicz, 1996 X ' 2143 G lyptograptus cf. nickoleyevi (Obut, 1965) X ' 6, 1444 Dimorphograptus confertus swanstoni Lapworth, 1876 X , L i i 1, 14, 1645 Orthograptus aff. mutabilis sensu Hutt, 1974 X X 646 Coronograptus cyphus cyphus (Lapworth, 1876) X X 1,6, 13, 14, 15, 16, 1847 Orthograptus mutabilis Elies &Wood, 1907 U ’ X : 1, 15, 16,1848 Pribylograptus sandersoni (Lapworth, 1876) u?x x , L? , 1, 13, 14,15,16, 1849 Lagarograptus acinaces (Tornquist, 1899) A , X , L , : I , 1,6, 15, 16,1850 Pribylograptus incommodus (Tornquist, 1899) U X X L 1,15, 16, 1851 Atavograptus strachani (Hutt & Rickards, 1970) 7 X X ? ? L? ! 1,6, 13, 14,1552 Pristiograptus fragilis frag ilis (Rickards, 1970) U? 1 X ' X ‘ X X X 1,6, 13, 14, 1553 Coronograptus gregarius gregarius (Lapworth, 1876) U? , X i X : X X X LM? 6. 13. 14, 15. 16. 18.2
X%\ 4 % \ *
C O^ ^\ % gk Os’®. ^ o \ &
\ \ X \ X
Fig. 3.6.1. Silurian graptoloid biostratigraphic range charts for the UK.
Fig. 3.6.1

Silurian Graptoloid Biostratigraphy of the U.K. (Rickards, 1976) & | 60 O
3 'S
I&O V,
U u
t ip :o Sog 9 , S 'E
; aCL 3 „ 3 ._
9 -3 1 9 a3 = J2 fja s i 1 £ £
&>•§ > o oa £° s • S g
fch-g2 s 8 60 O T3s s
3 «. 9-H
Series LlandoveryTelychianAeronianRhuddaruan Spirograptus euerichi
54 G lyptograptus tamariscus tamariscus (N icholson, 1868) X X X X X X X 7 7 1,6, 12, 14, 15, 16, 1855 *Glyptograptus incertus (Elies &Wood, 1907) ! aff. : aff. aff. aff. ■ 7 A ■ X ’ X X 6, 10,14, 15, 1656 G lyptograptus attenuatus (Rickards, 1970) ? : M , i 1,13,14, 1657 Glyptograptus sinuatus sinuatus (N icholson, 1869) ? X X X X 1,6, 13, 14, 16,2158 Cystograptus penna (Hopkinson, 1869) LM 1459 M onograptus revolutus vulgaris (Hutt, 1974) X 6, 1460 Coronograptus cf. leei ! X 1561 Coronograptus hipposideros (Toghill, 1968b) , X 1662 Coronograptus cirrus (Hutt, 1975) X 1563 'Climacograptus' wyensis (Zalasiewicz & Tunnicliff, 1994) ! XM? 1864 M onograptus austerus austerus Tornquist, 1899 ■ ? 1 X X 1,6, 14, 1565 M onograptus revolutus revolutus Kurck, 1882 , X X 1,6, 14, 16, 186 6 Monograptus austerus sudburiae Hutt, 1974 7 X X 1,6 ,1 4 ,1 867 G lyptograptus enodis enodis Packham, 1962 7 X X 6, 12, 14, 186 8 G lyptograptus cuneatus Rickards, 1970 : M ' 13, 1469 "Climacograptus" sp. 1 o f Hutt, 1974 ; M 6, 1470 M onograptus difformis Tornquist, 1899 , MU X , , 1,6, 13, 14, 15, 1671 Glyptograptus tamariscus distans Packham, 1962 MU X X aff. 1,6, 12, 14, 1572 G lyptograptus tamariscus varians Packham, 1962 aff'MU aff?X X aff. 12, 13, 14, 15, 1873 G lyptograptus tamariscus linearis (Pemer, 1897) > MU X X X A : 12, 14,1574 M etaclimacograptus undulatus Elies &Wood, 1913 : MU? 9 9 X X : X : 6, 13, 14, 15, 18,2175 Orthograptus sp. nov. B (of Cocks & Toghill, 1973) , U 276 Glyptograptus tamariscus angulatus Packham, 1962 U" X X X 12, 1477 Clinoclimacograptus retroversus Bulman & Rickards, 1968 ■■ u X X X ’ A ’ X 1 ,6 ,1 3 ,1 4 ,1 578 M onograptus triangulatus triangulatus (Rickards, 1970) ! u? X X ? 1,6, 13, 14, 15, 1879 Orthograptus cyperoides (Tornquist, 1897) iCf.U? X x , x , X i x i 1,6 ,13 , 14, 15, 1680 Pristiograptus concinnus (Lapworth, 1876) 7 X X X A X 7 1,6 ,13 , 14, 15,16, 1881 Pribylograptus argutus argutus (Lapworth, 1876) ? X X A A L? 1,6 ,13 , 14, 15, 16, 1882 M etaclimacograptus undulatus Kurck, 1882 ? X X ! X X X ! X u 1 ,6 ,13 ,14 , 15,1883 M onograptus triangulatus predecipiens Sudbury, 1958 L 1,1484 M onograptus triangulatus extremus Sudbury, 1958 LM? 14, 15, 1885 M onograptus triangulatus sim ilis Elies &Wood, 1913 7 148 6 Petalolithus sp. (of Sudbury, 1958) ? 1 ' 1487 M onograptus revolutus bicornis Hutt, 1974 X X ■ ■ 6, 148 8 M onograptus triangulatus separatus Sudbury, 1958 X x , 1,6. 13, 1489 Petalolithus palm eus latus (Barrande, 1850) ? X 1, 14, 15, 1690 M onograptus triangulatus fim briatus (N icholson, 1868) ? A ? 1,6, 14, 15, 16, 1891 Petalolithus ovatoelongatus (Kurck, 1882) X X ! A ; A ! ? 6, 13, 14, 15, 16, 18,2192 Rastrites spina sensu Rickards, 1970 7 X : X : A : 1,6, 13,14,2193 Orthograptus bellulus (Tornquist, 1890) 7 X ; A ; X ; ? 6, 14,15,16,2194 Rastrites longispinus Perner, 1897 7 X X A ? 1,6 ,13 , 14, 15, 16,2195 M onograptus revolutus sequens Hutt, 1974 M 696 Rhaphidograptus toernquisti sensu Sudbury, 1958 M 1497 Petalolithus minor (Elies, 1897) MU x 1 A , X i i 1,14, 15, 1698 Rastrites peregrinus Barrande, 1850 MUaff. X Xaff. X 1 ,6 ,14 ,1 5 ,1 6 ,2 199 M onograptus delicatulus Elies &Wood, 1913 MU X X A X ?L 1,6 ,141 0 0 G lyptograptus tamariscus acutus Packham, 1962 U 12, 141 0 1 G lyptograptus alternis Packham, 1962 U 12,14, 181 0 2 "Climacograptus" tamariscoides Packham, 1962 U 12, 14103 Pseudoglyptograptus rhayaderensis (Rickards & Koren, 1972) U 14104 M onograptus revolutus praecursor Elies &Wood, 1911 U L ! 6, 14105 M onograptus pseudoplanus Sudbury, 1958 U? L? ; 1 1,6, 14, 15, 18106 M onograptus triangulatus m ajor Elies &Wood, 1913
\U X ,
,/A , n' - V 1.6. 13. 14. 15
v
* The biostratigraphic range o f Glyptograpus incertus continues up to the renaudi Subzone.
Fig. 3.6.2. Silurian graptoloid biostratigraphic range charts for the UK.
Fig. 3.6.2

Silurian Graptoloid Biostratigraphy of the U.K. (Rickards, 1976)
Ata
vogr
aptu
sat
avus
Lag
arog
rapt
usac
inac
esC
oron
ogra
ptus
cyph
us
Mon
ogra
ptus
tria
ngul
atus
Dip
logr
aptu
sm
agnu
sPr
ibyl
ogra
ptus
lept
othe
caM
onog
rapt
usco
nvol
utus
Mon
ogra
ptus
sedg
wic
kii B M
IS§is i
Series Llandovery
Stage RhuHHsmian Acrnnian TelychianSpirograptus guerichi jSp. turriculati
/ // / ^
^ /A
/ i n
- ............................. ... ' ‘g ' *fg 'g '<? ’<? 'g '4f 'g '4? g ‘g /
i f /
¥107 Pseudoglyptograptus vas (Bulman & Rickards, 1968)
f-------1 u ? 1 X
r 1 ! ! 1 6 ,14 ,18108 D iplograptus magnus H. Lapworth, 1900 1 ■ u ? 1 A ‘ ? ! 1 1 1,6,13, 14, 15,16109 M onograptus communis rostratus Elies &Wood, 1913 . , u i X X i L? 1 ' 1,6, 14,18110 Monograptus communis communis Lapworth, 1876 u X A A 7 1, 13, 14, 15, 16, 18111 Pribylograptus leptotheca (Lapworth, 1876) 1 u? X A X X 1 ,6 ,1 3 ,1 4 ,1 5 ,1 6 ,1 8 ,::112 Monograptus intermedius (Carruthers, 1868) 1 u ' X X 1 X ' ? ! ? 1 1 14, 16113 Glyptograptus elegans Packham, 1962 , , u , X X , A , A ; X , , X 12, 14, 15114 Glyptograptus sinuatus crateriform is Rickards, 1970 X 13, 14115 Monograptus chrysalis (Zalasiewicz, 1992) X 17116 Monograptus cf? tenuissimus (Obut & Sobolevskaya, 1968) 1 1 X ! 1 < 6, 14117 Monograptus changyangensis Sun, 1933 .X? = 1 ' , 10118 Torquigraptus denticulatus (Tornquist, 1899) , , X A , x 1 ' 1 ,6 ,14 ,15 ,16 ,21119 Orthograptus insectiformis (N icholson, 1869) X A A 6, 14, 15, 16,21120 Monograptus capis Hutt, 1975 X X X X X ' X 1,6, 14,15121 Torquigraptus involutus (Lapworth, 1876a) 1 1 ? X 1 X ' X ! X 1 L ' 1,6, 14, 16,21122 Pseudoglyptograptus sp. 2 (of Rickards, 1972) i , M ! i ' 14123 Monograptus sp. (of Rickards, 1974) ( M ! i » 14124 'Diplograptus' thuringiaceus (Eisel, 1919) M 14
125 M onograptus millipeda (McCoy, 1850) ' X L? : i 1 1,6, 14, 16,21126 M onograptus cerastus (Hutt, 1974) X ' L? ! i ' 1,6, 14, 15127 M etaclimacograptus hughesi (N icholson, 1869) , X , L = i i 1,6, 13, 14, 15, 16,21128 M onograptus im ago (Zalasiewicz, 1992) X , L 17,21129 Petalolithus folium (Hisinger, 1837) X A 14, 16130 M onograptus argenteus (Nicholson, 1869) A ' X i • 1,6, 14, 15, 16,21131 M onograptus convolutus (Hisinger, 1837) 1 cf. > X ' L? i < 1 ,6 ,14 ,15 , 16,21132 Monograptus lobiferus (McCoy, 1850) , X , A ( ? < i 1 1,6 ,13 ,14 , 15, 16,21133 Glyptograptus serratus (Elies &Wood, 1907) X X L 14, 15,16134 Monograptus limatulus (Tornquist, 1892) X A X 1,6, 13, 14, 15, 16,21135 Pristiograptus jaculum (Lapworth, 1876) ' A 1 A 1 X : i i 1, 14, 15, 16,21136 Climacograptus scalaris (Hisinger, 1837) < ? < A . X S 1 1 1, 13, 14, 15, 16,21137 Pristiograptus regularis (Tornquist, 1899) , U , A , A 0 1,6, 13, 14, 15,16,21138 Torquigraptus urceolinus (Stein, 1965) L 21139 Cephalograptus tubuliformis (N icholson, 1867) 1 L? 5 i 1 14, 15, 16140 M onograptus urceolus Richter (Elies & Wood, 1913) 1 1 L? i > 1141 Monoclimacis crenularis (Lapworth, 1880) , x : i » 1,6, 14, 15, 16,21142 M onograptus clingani (Carruthers, 1867) X 1,6, 14, 15, 16,21143 Pribylograptus jon esi (Rickards, 1970) 7 13144 Rastrites phleoides Tornquist, 1887 1 1 X : i ' 6,14145 Rastrites approximatus geinitzi Tornquist, 1907 . X , 7 i l 1 14, 16146 Cephalograptus cometa cometa (Geinitz, 1852) , X 7 : 1 1 6,14 ,1 5 ,1 6147 Monograptus decipiens decipiens Tornquist, 1899 X X 1,6, 14, 15, 16148 Pseudoretiolites perlatus (Nicholson, 1868) ' X X s 6, 10,14,21149 Rastrites hybridus hybridus Lapworth, 1876 ' ' MU t ' 15,21150 Cephalograptus cometa extrema Boucek & Pribyl, 1942 , U ,Laff. ! J 1 6,13, 14,21151 M onograptus distans (Portlock, 1843) u L 14152 Stimulograptus sedgwickii (Portlock, 1843) u X X 1 X ' X X ' X 6, 13, 14, 15, 16153 Oktavites contortus (Pemer, 1897) ' 1 u ? 1 ? X > X 1 X j X 10154 Pribylograptus argutus sequens (Rickards, 1970) , L 5 l » 13, 14155 Monograptus angustus Rickards, 1970 , i L? : 1 ' 13,14156 Glyptograptus packhami Rickards, 1970 LM 13, 14157 Lagarograptus tenuis (Portlock, 1843) ' X 6, 14, 15158 Rastrites fugax Barrande, 1850 ' ‘ A ; X i X ‘ A 10.15159 Pristiograptus variabilis (Pemer, 1897) , i A A . X , X _i------ 6. 10. 14.15
\ \ %X \% \ ' V
Fig. 3.6.3. Silurian graptoloid biostratigraphic range charts for the UK.
Fig. 3.6.3

S i l u r i a n G r a p t o l o i d B i o s t r a t i g r a p h y o f t h e a ’ 1 ‘ a - 1 3 .„8*23
3 « 81s
3Cl
U.K. ( R i c k a r d s , 1976) o i ’ £ ! ' o f ■ .& !■ . - s i , § i ,o ! £ £ S 8
o iC 60o *oS Si
bra§ 1 o ES i
I I
i'S
Series Llandovery
Stage A p . r n n i a nTelychian
Spirograptus guerichi | Spirog. turriculatus Monog. crispus
/ / / i ? / / • 4^ ■<? > 4 f ^'ev / ■£? / ' s /
&
,C5i .
/
160 Petalolithus kurcki (Rickards, 1970)—1 ---- 1 " T
L X X X L! 1 : 1
10, 14, 15161 * Pristiograptus nudus (Lapworth, 1880) • X ! X 1 X ! X 1 X X 1 X - 1 1,6, 13, 14, 15, 16162 M onograptus communis obtusus Rickards, 1970 i i M : 1 13, 14163 Rastrites sp. A o f Rickards, 1970 : M 13, 14164 "Climacograptus " simplex Rickards, 1970 M 13, 14165 G lyptograptus spp. o f Rickards, 1970 ' M : 13, 14166 Rastrites gracilis Pribyl, 1943b 1 : cf? ’ X 1 10, 15167 Petalolithus clavatus Boucek & Pribyl, 1941a ; ? , 10168 Petalolithus sp. o f Loydell, 1993 X 10169 Petalolithus ovatus (Barrande, 1850) X ? 10, 15170 Petalolithus praecedens Boucek & Pribyl, 1941a , x , 10171 Pristiograptus sp. 1 o f Loydell, 1993 > : X 1 10172 M onograptus adm irabilis (Pribyl & Munch, 1942) . = X , 10173 M onograptus capillaris sensu Loydell, 1993 X 10,21174 Rastrites schaueri Storch & Loydell, 1992 1 , x > 10175 M onograptus pulcherrimus (Manck, 1928) 1 , x , 10176 Streptograptus ansulosus (Tornquist, 1892) , i X , X ; 10177 M onograptus yichangensis Ni, 1978 . x , X 10178 Petalolithus conveniens (Koren', 1967) X X X 10179 Rastrites carnicus Seelmeier, 1936 : X ' ! X ‘ X ! 1 ! ! 1 10180 Rastrites linnaei Barrande, 1850 ' , X ' X i X 1 X ’ L 1 I : t 10181 Stimulograptus halli (Barrande, 1850) , , ? , X , X , X , X , X 1 1 = 1 1, 10, 13, 15, 16182 Pristiograptus pristinus Pribyl, 1940a X X X X X X X 10, 15183 Pristiograptus sp. 2 o f Loydell, 1993 X ‘ 10184 Pristiograptus sp. 3 o f Loydell, 1993 ' X : 1 i 10185 M onograptus gemmatus (Barrande, 1850) , X : A I : 10186 Stimulograptus glanfredensis Loydell, 1993 X X 10187 Pristiograptus renaudi (Philippot, 1950) X X A 10188 Streptograptus strachani Loydell, 1993 X : X 1 X ! 10189 Spirograptus guerichi Loydell, et al. 1993 1 X i X ■ X ‘ X 10190 Paradiversograptus runcinatus (Lapworth, 1876a) A , ? , X , L 10,15191 Streptograptus plumosus (Baily, 1871) X X X X 10, 15192 Streptograptus pseudoruncinatus (Bjerreskov, 1975) X : X X X 10193 M onograptus bjerreskovae Loydell, 1993b ' X : X 1 X ! X 1 X ' X ! X 1 x 10, 19194 G lyptograptus auritus Bjerreskov, 1975 i X 1 1 i 1 10195 G lyptograptus supem us Fu, 1986 , ? , X I 1 1 i 10, 15196 Petalolithus elongatus Boucek & Pribyl, 1941a X 10197 Petalolithus globosus Chen, 1984a X 10198 Petalolithus regius (Hundt, 1957) ' X 1 1 ! i 10199 Streptograptus? sp. nov. Loydell, 1993b , , x 1 f 1 102 0 0 Streptograptus sp. o f Loydell, 1993 X 102 0 1 G lyptograptus fastigatus Haberfelner, 1931a ? ! A X 10, 152 0 2 Streptograptus? sp. nov. o f Loydell, 1993 1 : ? 1 A? : l i l t 10203 Rastrites maximus Carruthers, 1867b > ? : X . X , i l l ! 8,10 ,15204 Petalolithus conicus (Boucek, 1932) , , X , X , L 1 i : i 10205 Petalolithus palm eus (Barrande, 1850) X 7 X 10, 14, 15206 Petalolithus altissimus (Elies & Wood, 1908) x ! X 1 X ’ X ‘ X ' X ’ X 1 X 10, 14207 Streptograptus fdiform is Chen, 1984a ? : X , X ■ X 1 ? ! X 1 , 10, 15208 Rastrites sp. aff. maximus o f Loydell, 1993 X 7 7
x 1 x 10209 Pseudoplegmatograptus obesus obesus (Lapworth, 1877) ? ' X X X X X X X X 6, 10, 142 1 0 Petalolithus giganteus Boucek & Pribyl, 1941a 1 x ! ? I 102 1 1 Petalolithus hispanicus (Haberfelner, 1931b) I X 1 102 1 2 Pseudostreptograptus william si Loydell, 1991a 7 . x , L U ... * J, 10, 5
1V \ ^ \ \
. V W\ O-t/>
s ' ' ' ' ' V i
* The biostratigraphic range o f Pristiograptus nudus continues up to the spiralis Biozone.
Fig. 3.6.4. Silurian graptoloid biostratigraphic range charts for the UK.
Fig. 3.6.4

S i lu r ia n G r a p t o lo id B io s t r a t ig r a p h y o f th e3 v.8*3
i i-------
3 u ’s 'c a a 2
1
•if B3 55,3-= 1 Q.®
U .K . ( R ic k a r d s , 1 9 7 6 ) o fc§>3 ' 8 2 'a c l a ft
1 1 s'& '
8 |§ i
feB.felja, a o o -e o f i
S B s £ u 1 S -g
S e r i e s L l a n d o v e r y W e n l o c k
S t a g esi
T e l y c h i a nQVi/ain\i/rvr\Hi a ni
. guericlp Sp. tumculatus Monog. crispus |
/ / / ■ / / / £
^/
r . £ & A -Ac, ,^ & £ A $
S r-S / &
f *
213 Torquigraptus cavei Loydell, 1993 ' X X ' "T ] ---- T10
214 Pristiograptus huttae Loydell, 1993 ' ' ' X 1 1 1 0215 Glyptograptus(?) nebula Toghill & Strachan, 1970 , , , X X , X 7 i ? . X ; 1 1 19216 Torquigraptus planus (Barrande, 1850) x X 10
217 M onograptus m arri Pemer, 1897 X X X X X X cf. cf. cf. 10, 19218 Streptograptus barrandei (Suess, 1851) ' 1 X ' 1 0
219 Stimulograptus utilis Loydell, 1991 > i A > 10
220 Pristiograptus bjerringus (Bjerreskov, 1975) ( , X , X 10
221 Rastrites equidistans spengillensis Rickards, 1970 L X X 1 0
222 Streptograptus storchi Loydell, 1991 ' x X X A 10, 19223 Streptograptus johnsonae Loydell, 1991 1 1 U 1 A X 1 i 1 0
224 Spirograptus turriculatus {Barrande, 1850) , , U > X X 1 1 1 0
225 Petalolithus tenuis (Barrande, 1850) : u 7 X X X , 10, 15, 19226 Streptograptus tenuis Loydell, 1993 X 1 0
227 Petalolithus schaueri Loydell, 1991 ' ' ' X X1 1
1 0
228 Pristiograptus sc.hucherti Bjerreskov, 1981 1 ' 1 X X 1 1 1 0
229 Stimulograptus becki (Barrande, 1850) , , , X 1 , : : t t I 1 0
230 Torquigraptus proteus (Barrande, 1850) X X 10, 19231 M onograptus rickardsi (Hutt, 1975) X x x ' X 1 7 19232 M onoclim acisl galaensis (Lapworth, 1876) ' 1 1 7 X X ' X 1 I i 19233 Streptograptus whitei Zalasiewicz, 1994 , , i X : ! : : : ! 1 ( 19234 Torquigraptus cam icus (Gortani, 1923) . , t x , x X , 1 : : 1 19235 Monograptus crispus Lapworth, 1876 7 X X 19236 Streptograptus exiguus (Nicholsoni, 1868) ' ' ' x : X x 1 L ..................................... 19237 Stimulograptus d in tonensis (sensu Hall, 1852) ' 1 ? : X X ' L . 19238 Streptograptus loydelli Zalasiewicz, 1994 , , , A , ? 1 1 1 ! 19239 Torquigraptus pragensis pragensis (Pribyl, 1943) X ? L 19240 M onograptus discus Tornquist, 1883 X X ' X X L 19241 Retiolites geinitzianus geinitzianus (Barrande, 1850) ' 1 ' X X 1 X X X X X X X ' L 11,14, 19,22242 Monograptus pragensis ruzickai (Pribyl, 1943) i i X 1 1 i i 19243 Streptograptus aff. sartorius o f Zalasiewicz, 1994 ( x , t i 19244 Streptograptus sartorius (Tornquist, 1881) X L 19245 Torquigraptus tullbergi spiraloides (Pribyl, 1945) 1 1 7 ' X ' '
i t 14246 Monograptus pseudocommunis Zalasiewicz, 1994 1 1 ' L • i i 19247 M onoclimacis directa Zalasiewicz et al, 1995 , , : A , ! I 19248 M onoclimacis griestoniensis (Nichol, 1850) A
19249 Streptograptus aff. loydelli o f Zalasiewicz, 1994 X 19250 Torquigraptus pergracilis (Boucek, 1931) ' ' X : 11, 19251 Torquigraptus tullbergi tullbergi (Boucek, 1931) * 1 1 X : X : 14,19252 Pristiograptus initialis (Kirste, 1919) , i • , x 19253 Monograptus priodon (Bronn, 1835) 7 7 ? X X X LM X X X X X 11, 14, 19,22254 Torquigraptus aff. pragensis'} o f Zalasiewicz, 1994 1 1 ' u ' 19255 Monoclimacis crenulata (Tornquist, 1881) ' 1 ' X ' L 1 i 1 11, 19256 Lapworthograptus grayae Lapworth, 1867a , , , x , x . t 1 23257 Monoclimacis vomerina vomerina (N icholson, 1872) X . 7 7 7 ■ X I X X : L 11, 19,22258 Monoclimacis hemmanni (Pribyl, 1941c) L 11
259 Monoclimacis n. sp. 1 o f Loydell & Cave, 1996 ' 1 L ! 11
260 Monograptus vesiculosus Pemer, 1899 > i 1 i L : 1 11
261 Streptograptus anguinus (Pribyl, 1941) , , : L , , 11262 Retiolites venosus (Hall, 1852) X 11263 M onograptus parapriodon (Boucek, 1931) ! X ! ' 11, 19264 Pristiograptus aff. initialis (broad form) (Kirste, 1919) ■ 1 X 19265 'Monograptus' nodifer Tornquist, 1881 , , , X , , ' 1 6 , 11, 14
° 4%
% \
Fig. 3.6.5. Silurian graptoloid biostratigraphic range charts for the UK.
Fig. 3.6.5

Silurian Graptoloid Biostratigraphy of the U.K. (Rickards, 1976)
Mon
oclim
acis
cren
ulat
a
Cyr
togr
aptu
sce
ntri
fugu
s.
..
.C
yrto
grap
tus
mur
chis
oni
Mon
ogra
ptus
ricc
arto
nens
is
Cyr
tpgr
aptu
sri
gidu
s
Cyr
tpgr
aptu
slin
nars
soni
Cyr
tpg.
elle
sae
Cyr
tpg.
lun
dgre
ni
Got
hogr
aptu
sna
ssa
Series Llandovery Wenlock
Stage Telychian Sheinwoodian Homerian
/ / / . / ■ & ^ 4 • / ■ 4 n/ / ■ £ / / /
/ / / f y - 4 4 ' / / ' 4 ' 4 ' 4 4266 Diversograptus ramosus Manck, 1923 X L
t------------ I 1 -r ' 11
267 Oktavites spiralis (Geinitz, 1842) X ’ L M * 11, 19268 M onoclimacis linnarssoni (Tullberg, 1883) X X L , i 11, 19269 Barrandeograptus? bom holm ensis (Laursen, 1940) X X , X X L? , 11270 Retiolites geinitzianus angustidens Elies & Wood, 1908 X X X X L 11
271 M onoclimacis geinitzi (Boucek, 1932) X 1 X : X X ! X 1 11
272 Pristiograptus prantli (Pribyl, 1940) M U ’ 1 i 1 , ■ 11273 M onoclimacis sublinnarssoni (Tullberg, 1883) , LM , i 11274 M onograptus speciosus Tullberg, 1883 L M 11275 Streptograptus wimani (Boucek, 1932) L M 11, 14276 Pristiograptus largus Pemer, 1899 ‘ LM ’ 11
277 Cyrtograptus lapworthi Tullberg, 1883 , X . 11
278 M onoclimacis vomerina ssp. 1 o f Loydell & Cave, 1996 M 11
279 Barrandeograptus pulchellus (Tullberg, 1883) M 11
280 M onoclimacis vomerina basilica (Lapworth, 1880) 1 M ! X X 1 X : 7 : X ! : 1 1 , 2 2
281 M ediograptus flitton i Loydell & Cave, 1996 L 1 11
282 M onoclimacis shottoni Rickards, 1965 L M X , ( , 11
283 Cyrtograptus insect us Boucek, 1931 X ? 1 1 , 2 2
284 Cyrtograptus sp. of Loydell & Cave, 1996 X LM 11
285 M onograptus pseudocultellus Boucek, 1932 M U X 11
286 M ediograptus morleyae Loydell & Cave, 1996 U , 11
287 M ediograptus sp. aff. inconspicuus o f Loydell & Cave, 1996 U X ( , 11
288 M onoclimacis? sp. 1 o f Williams & Zalasiewicz, in press X 22
289 M onograptus danbyi Rickards, 1965 X ' = 2 2
290 Barrandeograptus sp. of Loydell & Cave, 1996 X , X : X i L ! ' 22
291 M ediograptus cautleyensis (Rickards, 1965) X , 14
292 M onograptus simulatus Rickards, 1965 X 14
293 Cyrtograptus centrifugus Boucek, 1931b X ‘ L? 1 1 , 2 2
294 M ediograptus inconspicuus (Boucek, 1931) M U L M ‘ ' 11
295 Plectograptus spp. o f Williams & Zalasiewicz, in press X i 22
296 Cyrtograptus spp. o f Williams & Zalasiewicz, in press X X 2 2
297 M onoclimacis kettneri Boucek, 1931 L M ? ’ LM ? 1 1 , 2 2
298 M onoclimacis adunca (Boucek, 1931) L U 1 L?aff. : 1 1 , 2 2
299 M ediograptus flexuosus (Tullberg, 1883) U ’ ' 11
300 M onoclimacis vikensis Bassett & Rickards, 1971 . L , , , 11
301 M onoclimacis n. sp. 2 o f Loydell & Cave, 1996 L 11
302 Pristiograptus praedubius (Boucek, 1931) 1 L 11
303 M onograptus radotinensis radotinensis Boucek, 1931b L 11
304 M onograptus radotinensis o f Loydell & Cave, 1996 , L , 11
305 Cyrtograptus murchisoni bohemicus Boucek, 1931 L M 11
306 Cyrtograptus murchisoni Carruthers, 1867 X 1 1 , 2 2
307 IPseudoplegm atograptus wenlockianus Storch, 1992 ’ X : L 11
308 M onograptus ftrm us firm us Boucek, 1931 . X i X i X 1 11, 1 4 , 2 2
309 Pristiograptus latus Boucek, 1932 L | , 11
310 * Pristiograptus dubius dubius (Suess, 1851) X X X X X 11, 1 4 , 2 2
311 M onograptus riccartonensis Lapworth, 1876 : X 11, 1 4 , 2 2
312 M onograptus radotinensis inclinatus Rickards, 1965 = X 1 : 1 1 4 , 2 2
313 M onograptus firm us sedberghensis Rickards, 1965 , M U 14
314 M onograptus Jlexilis belophorus (M eneghini, 1857) M U X 14
315 Monograptus instrenuus Lenz & M elchin, 1991 7 22
316 Cyrtograptus cf. urbanekii Teller, 1976 L ’ 22
317 M onograptus priodon-flem ingii intermediates 1M U
. X 14
318 M onograptus antennularius (Meneghini, 1857) , , X , L M 14
V \\ \
%% \ •A ‘‘ o f i
$ 8H
-8V t% 'A \V 4V / i \ %
' \
* The biostratigraphic range o f Pristiograptus dubius dubius continues up into the Ludlow Series.
Fig. 3.6.6. Silurian graptoloid biostratigraphic range charts for the UK.
Fig. 3.6.6

Silurian Graptoloid Biostratigraphy of the U.K. (Rickards, 1976)
Cyr
tpgr
aptu
slin
nars
soni
Cyr
tpg.
elle
sae
Cyr
tpg.
lun
dgre
ni
Got
hogr
aptu
sna
ssa
Pris
tiogr
aptu
slu
dens
is
Neo
dive
rsog
rapt
usni
isso
ni
Lob
pgra
ptus
scan
icus
Saet
ogra
ptus
inci
pien
s
Saet
ogra
ptus
lein
twar
dine
nsis
Boh
emog
rapt
usbo
hem
icus
Series Wenlock Ludlow
Stage Sheinwoodian Homerian Gorstian Ludfordian
j* / /§"$'i t 'S' : S ' $
/ N
^ ^ /■ ? ' £ ■ & ■ & / /
/ cP
319 Pristiograptus meneghinii meneghinii (Gortani, 1923) U 14320 M onoclimacis flum endosae flum endosae (Gortani, 1923) U? X i X ’ X ; . 14,22321 M onograptus retroflexus Tullberg, 1883 , X , X , XL? : i 14,22322 M onograptus flem ingii flem ingii (Salter, 1852) X X X 14,22323 M onograptus Jlexilis flexilis Elies, 1900 MU LM 14,22324 Pristiograptus dubius pseudolatus Rickards, 1965 1 MU ‘ X L? i 14,22325 Cyrtograptus rigidus rigidus Tullberg, 1883 • X 1 14,22326 Plectograptus? bouceki Rickards, 1967 X 14327 Cyrtograptus rigidus cautleyensis Rickards, 1967 X 14328 Cyrtograptus linnarssoni Lapworth, 1880 : X?M ' , ; 14,22329 Cyrtograptus perneri Boucek, 1933 X X . 14,22330 Cyrtograptus ellesae Gortani, 1923 , X ; 14,22331 M onoclimacis flum endosae kingi Rickards, 1965 X 14332 Cyrtograptus lundgreni Tullberg, 1883 ‘ X ’ ' ’ ’ 14,22333 Cyrtograptus lundgreni gracilis Boucek, 1933 ' X : l 14334 Paraplectograptus eiseli (Manck 1918) , x , 14335 Testograptus testis (Barrande, 1850) X 14336 Cyrtograptus ramosus Boucek, 1931 : MU? 14, 22337 Cyrtograptus hamatus (Baily, 1862) 1 ' XUcf. ’ i 14, 22338 Pristiograptus sp. 1 o f Holland et al., 1969 ■ u ■ 14339 Pristiograptus aff. ja egeri Holland et al., 1969 U , X LM 14340 Gothograptus nassa Holm, 1890 U A X L 14,22341 Pristiograptus jaegeri Holland et al., 1969 i u X ’ 14342 Monograptus ludensis Murchison, 1839 ■ MU ' X ' L ■ 14,22343 Holoretiolites (Balticograptus) lawsoni Holland et al., 1969 i M , 14344 Pristiograptus auctus Rickards, 1965 MU LM 14345 Monograptus deubeli Jaeger, 1959 U 14346 Monograptus uncinatus orbatus Wood, 1900 1 aff. 1 LM : 14347 Plectograptus macilentus (Tornquist, 1887) . LM i 14348 N eodiversograptus nilssoni (Barrande, 1850) i , LM 14349 Spinograptus spinosus (Wood, 1900) LM 14350 Saetograptus (Colonograptus) colonus colonus (Barrande, 1850) ' X ; LM ' 14351 Saetograptus (Colonograptus) varians varians (Wood, 1900) X X L 14352 Bohemograptus bohemicus s.l. (Barrande, 1850) , X : A , X , 14353 Spinograptus clathrospinosus (Eisenack, 1951) MU 14354 Saetograptus (Colonograptus) roemeri (Barrande, 1850) MU 14355 Saetograptus (?Saetograptus) wandalensis (Watney & Welch, 1911) 1 1 MU ’ 14356 Saetograptus fritsch i fritsch i s.l. (Boucek, 1936) i MU i 14357 Cucullograptus (Lobograptus) progen itor Urbanek, 1966 . , MU . 14358 Saetograptus (Colonograptus) colonus com pactus (Wood, 1900) MU LM 14359 Lobograptus simplex Urbanek, 1966 MU LM 14360 Cucullograptus (Lobograptus) scanicus (Tullberg, 1883) ' MU • / 1 1 14361 Lobograptus critinus (Wood, 1900) i MU , X , 14362 Saetograptus (Saetograptus) chimaera salw eyi (Lapworth, 1880) MU X 14363 Monoclimacis m icropoma (Jaekel, 1889) MUA > 14364 Pristiograptus vicinus (Perner, 1899) ' ' MU ’ >: ' l ' 14365 Monoclimacis haupti (Kuhne, 1955) MU X LM 14366 Saetograptus (Saetograptus) leintwardinensis incipiens (Wood, 1900) , MU , X , A , ? , 14367 Saetograptus (Saetograptus) chimaera chim aera (Barrande, 1850) U / 14368 Saetograptus (Saetograptus) chimaera sem ispinosus (Elies & Wood, 1911) ' U ' > ' X 14369 Pristiograptus welchae Rickards, 1965 M 14370 Saetograptus clunensis (Earp, 1944) ! , MU ! LM 1 14
371 Pristiograptus tumescens tumescens (Wood, 1900) , MU , A ? 14372 Bohemograptus bohemicus tenuis (Boucek, 1936) MU X L 14
S
% \ \ ' ' '■ ' \ ^ \
Fig. 3.6.7. Silurian graptoloid biostratigraphic range charts for the UK.
Fig. 3.6.7

Series Ludlow
Gorstian LudfordianStage
T7T374375
Saetograptus? aff. incipiens (Wood, 1900)Saetograptus (Saetograptus) leintwardinensis prim us (Boucek, 1936)Saetograptus (Saetograptus) leintwardinenis leintwardinensis (Lapworth, 1880) ,
LM
No later biozones recorded in the U. K.
Fig. 3.6.8. Silurian graptoloid biostratigraphic range charts for the UK.
Fig. 3.6.8

Patterns of diversity and morphological change in British
Ordovician and Silurian graptoloids
The main purpose of biostratigraphy is to aid in the development of the chronostratigraphic
framework by enhancing the correlative value of fossiliferous sedimentary sequences. In
addition, biostratigraphical data provides detailed information on the biodiversity and
morphological changes that occur within a fossil group over a given geologic time period. This
study demonstrates diversity trends in Ordovician and Silurian graptoloids from the UK. The
data are derived from the revised biostratigraphical range charts (see chapter 3). Broad patterns
of morphological change within British Silurian graptoloids are also analysed using these range
charts, in conjunction with data obtained from representative illustrations of each Silurian
graptoloid species featured in the range charts (see APPENDIX 1). Several species biodiversity
crises can be identified within the Ordovician and Silurian graptolitic succession of Britain.
These biodiversity trends are often associated with patterns of graptoloid morphological change
and, in turn, reflect broader palaeoenvironmental controls, such as fluctuations in sea-level,
climate, ocean circulation and chemistry (Spjeldnaes 1961, 1978, Fortey 1984, Brenchley et al.
1994, Loydell 1998).
Previous Studies
Work on graptoloid biodiversity has mainly been carried out for the Silurian and taken many
forms, from the global to the local scale. All such studies attempt to outline periods of biotic
crisis and recovery within the group, detailing temporal extinction and speciation patterns in
order to understand how these relate to broader palaeoenvironmental changes (e.g. Koren 1987,
Kaljo & Marss 1991, Urbanek 1993, Storch 1995, Melchin et a l 1998). A principal feature of
these studies has been the identification of ‘bioevents’- most often intervals of significant
species extinctions.
Koren’s (1987) research into Silurian and Devonian graptolite dynamics attempted
to correlate graptoloid biostratigraphic data from several regions, and to combine these to
produce global patterns of graptoloid biodiversity. She outlined three distinct mass extinction
‘events’, all related to major palaeoenvironmental changes. Urbanek (1970, 1993) recognised
Page 4.1

four such events or ‘biotic crises’ during the upper Silurian, two of which (his C \ and C2)
correspond with the end -lundgreni and leintwardinensis events of Koren (1987). Indeed, the
lundgreni graptoloid extinction event has been widely recognised by many other workers (e.g.
Storch 1995, Melchin et al. 1998, Jaeger 1991, Koren 1991). The most recent global study for
Silurian graptoloids identified 10 separate graptolitic extinction events based on diversity and
survivorship analysis (Melchin et al. 1998), and incorporated some of the more recently
available biostratigraphic data (Cave & Loydell 1996, Storch 1994, Melchin 1994, Zalasiewicz
& Tunnicliff 1994, Gutierrez-Marco et al. 1996, Gutierrez-Marco & Storch 1998). Earlier
‘events’ correspond with those of previous global studies, and despite the necessity of using a
broader generalised biostratigraphic zonal scheme for correlative purposes, also largely coincide
with ‘events’ recognised in regional diversity work (Storch 1995, Loydell 1994, Koren 1988,
1991).
Studies of morphological change are often carried out locally or regionally and take
an evolutionary approach, focusing on the development of graptoloid lineages (e. g. Rickards et
al. 1976, Koren & Urbanek 1994). All these studies are based in some measure on graptoloid
biostratigraphic data.
The present study is a detailed regional diversity study of all Ordovician and Silurian
graptoloid species from the UK, and a study of the major morphological trends in British
Silurian graptoloids.
Diversity change
Biostratigraphic data are collected for the purposes of investigating how a given fossiliferous
section relates to an existing biostratigraphical scheme, and how it may be correlated with other
sections. When using such information for graptoloid diversity studies it must be remembered
that published biostratigraphic data have already been subject to the collection and processing
biases and requirements of the biostratigrapher. For example, a biostratigrapher may make a
generalisation regarding the appearance or termination of a taxon depending on knowledge and
experience of faunal ranges in a given area, in order to produce a useful range chart. As a result,
species ranges, particularly at zonal boundaries, may give a false impression of the biotic
structure in a section. A very different graptoloid species diversity curve results when the
Page 4.2

number of taxa recorded at biozonal boundaries is included (Figs. 4.1, 4.2). The species
diversity curve containing boundary information shows significant population extinctions in the
upper part of most biozones, followed by population increases in the lower part of most
succeeding biozones (Fig. 4.1). This might be (i) an artefact of the biostratigraphic data, and
diversity changes across biozonal boundaries may in reality represent a continuum of species
decline and recovery. It may equally be the case that (ii) small scale boundary events reflect real
episodes of faunal flux which conveniently define zonal boundaries. Without very detailed
reassessment of the zonal boundaries at several key biostratigraphic sections though, it is
difficult to say with certainty which hypothesis is more accurate. As a result, I prefer to be
cautious and leave the boundary values out, as did Storch (1995) in his study of the species
diversity of Bohemian Silurian graptoloids.
It is important to distinguish between a 'species diversity event' and a 'species extinction
event'. Extinctions need not be catastrophic to cause an overall diversity crisis. Similarly, a
diversity crisis can be as much the result of a lack of, or decline in incoming species as of a high
number of extinctions.
Ordovician (England & Wales)
Ordovician graptoloid biostratigraphic data for the UK is not as comprehensive as it is for the
Silurian; there are simply fewer taxa. During the early and middle Ordovician (Fortey & Cocks
1986) the British Isles was divided into two faunal provinces: England and Wales, and
Scotland. For this reason, the British Ordovician is dealt with in two, regional parts. However,
Ihave attempted to combine data from the two areas in order to examine graptoloid biodiversity
changes at the Ordovician/Silurian boundary (Fig. 4.6). Despite these stratigraphic and
taxonomic problems, significant fluctuations in overall graptoloid diversity, in the form of first
appearance and last occurrence of species, can be observed in both areas (Figs. 4.7.1, 4.7.2,
4.8.1, 4.8.2). Such changes are manifested here as a series of biodiversity 'events' which often
take place over two or more biozones but which are initiated at a specific horizon.
Page 4.3

The late Corymbograptus simulans Biozone-early Isograptus victoriae Biozone event
The earliest stages of the Ordovician in the Anglo-Welsh area see species diversity increase in a
'step-like' pattern into the upper part of the varicosus Biozone (Fig. 4.3), at which time a
rapidly increasing number of incoming species, coupled with a relatively low number of
extinctions led to a dramatic rise in graptoloid biodiversity up to the early simulans Biozone,
after which species diversity trends were stable until the upper part of the simulans Biozone
(Figs. 4.7.1, 4.7.2). The ratio of species appearances to disappearances initially declines in the
middle of the phyllograptoides Biozone and continues to decline throughout this diversity crisis
(Fig. 4.7.3). The end of the sim ulans Biozone saw the onset of the first major decline in
graptoloid species diversity in the Ordovician of England and Wales. The decline is abrupt,
terminating in the lower part of the victoriae Biozone. This event marks the largest increase in
species extinctions so far in the Anglo-Welsh Ordovician, and the effect is made more notable
by the lack of any incoming taxa at its initiation.
Following this extinction episode, the graptoloids recover much of their former species
diversity levels, and this appears to have been relatively sudden (Figs. 4.7.1. 4.7.4). This rapid
rise in diversity at the end of the victoriae Biozone to the early part of the gibberulus Biozone, is
followed by a diversity plateau through to the middle part of the cucullus Biozone, during which
the ratio of species appearances to disappearances declines (4.7.3). Diversity is seen to increase
through to the middle of the artus Biozone, attaining its highest value for the Anglo-Welsh
sequence although the species diversity ratio is seen to increase slightly (Figs. 4.7.1, 4.7.3).
A large number of extinctions at the start of the artus Biozone (about 24 species), does
not affect the diversity curve significantly, as it is balanced by a rapid rise in the number of
incoming taxa, which peak at a maximum for the Ordovician of England and Wales shortly
afterwards. However, the subsequent dramatic decline in speciation from the middle to late part
of the artus Biozone, together with an equally dramatic rise in the number of extinctions in the
late artus Biozone, result in a second biodiversity crisis for graptoloids (Figs. 4.7.1, 4.7.2).
The late Didymograptus artus Biozone-early Glyptograptus teretiusculus Biozone event
This second event, initiated in the upper part of the artus Biozone, sees a rapid decline in the
number of graptoloid species and subspecies, reflected in both the total zonal diversity and
Page 4.4

actual diversity plots (Figs. 4.5, 4.7.1, 4.7.2). The biodiversity decrease continues into the
lower murchisoni Biozone, and from here, declines more gradually until the lower teretiusculus
Biozone, where graptoloid diversity reaches its lowest point since the early Ordovician. The %
species extinction decreases markedly from the gibberulus to the murchisoni biozones, although
the initial decline is gradual (Fig. 4.7.4).
Following the latter event, graptoloid biodiversity increases markedly into the upper part
of the multidens Biozone, due to the combined effect of relatively low extinction levels and a
consistent increase in the number of incoming species. Overall, graptoloid biodiversity almost
recovers to the high levels of the middle part of the artus Biozone. The murchisoni to
teretiusculus biozones show an increase in the species appearance to disappearance ratio jjust
preceeding this artus Biozone biodiversity increase (Fig. 4.7.3).
The late Diplograptus multidens Biozone- late Pleurograptus linearis Biozone event
The dramatic recovery of Ordovician Anglo-Welsh graptoloid species was succeeded by another
graptoloid biotic crisis. The onset of this crisis occurs in the latter part of the multidens Biozone
and is marked by an increase in the % of species extinctions up to about 70% (Fig. 4.7.4). An
initially swift, then more gradual decline in diversity can be seen through to the late linearis
Biozone, although the % species extinction indicates a second peak, reaching up to 90% in the
linearis Biozone (Fig. 4.7.4). A rapid rise in the number of extinctions, coupled with a
relatively low, though slowly increasing number of incoming species resulted in a sharp species
diversity decline as the rate of species appearances fell well below the number of species
disappearances (Figs. 4.7.1, 4.7.2). The graptoloids never fully recovered from this
biodiversity event, although a moderate pulse of incoming taxa from the late anceps Biozone
into the persculptus Biozone resulted in a diversity increase at the Ordovician/Silurian boundary.
Ordovician (Scotland)
The assessment of diversity changes of the Scottish Ordovician graptoloids is difficult given the
scarcity of complete data, especially for the early Ordovician (Fig. 4.4). Pre-Caradoc graptolitic
sections are sporadic, yielding poor biodiversity information. As a result, this part of the
sequence has been excluded from the biodiversity analysis of Ordovician graptoloids from
Page 4.5

Scotland. Graptoloid species found in this part of the Scottish sequence bear closer similarity to
A ustralasian forms than their A nglo-W elsh counterparts, and so the Australasian
biostratigraphic scheme (VandenBerg & Cooper 1990) has been adopted for the early
Ordovician Scottish succession.
The late Climacograptus peltifer Biozone- late Climacograptus wilsoni Biozone event
The lower part of the gracilis Biozone sees the first discernible major diversification of
Ordovician graptoloids in Scotland. Through the gracilis and peltifer biozones the % of species
disappearing rises steadily, and the ratio of species appearances to disappearances falls
markedly (Figs. 4.8.4, 4.8.3). Incoming and outgoing species appear to remain in equilibrium
until the upper part of the peltifer Biozone (Figs. 4.8.1, 4.8.2). From this point a rapidly
increasing number of outgoing species is coincident with a low number of incoming taxa,
producing the first marked decline in species diversity in the Scottish sequence. This
approximately corresponds with the middle to upper multidens Biozone of England and Wales,
just preceding the late Diplograptus multidens- late Pleurograptus linearis biodiversity crisis of
the Anglo-Welsh succession (Fig. 4.7.1). The species diversity curve displays a very gradual
decline after this, until the end of the wilsoni Biozone (Fig. 4.8.1). From this interval onwards,
there is a marked rise in graptoloid biodiversity, reaching its highest level for the Scottish
Ordovician in the middle part of the clingani Biozone. Indeed, the total and actual diversity of
species in the clingani Biozone in Scotland is significantly greater than that of England and
Wales regardless of whether the total forms within the biozone, or one of the component
subzones is considered (Figs. 4.5, 4.7.1, 4.8.1). This peak is preceded by a rapid rise in
incoming species, coupled with a relatively low number of taxa becominging extinct.
The middle Dicranograptus clingani Biozone- late Dicellograptus complanatus Biozone event
Following the early clingani Biozone diversity high, a decline and sustained low number of new
species mirrored by a steady, but significant number of species disappearances combined to
produce a second major species biodiversity crisis (Figs. 4.8.1, 4.8.2). This event correlates
with the upper part of the Diplograptus multidens Biozone- late Pleurograptus linearis Biozone
biodiversity crisis of the Anglo-Welsh succession (Fig. 4.7.1), and continues more gradually
Page 4.6

up into the latter part of the complanatus Biozone. This relatively protracted biodiversity event is
mediated by a slight increase in species in the middle of the linearis Biozone, at a point where
the number of incoming species rose briefly (Fig. 4.8.1). This diversity event correlates with a
marked and sustained peak in the % of species extinctions (Fig. 4.8.4) and an initially gradual,
followed by a more rapid increase in the species appearance to disappearance ratio (Fig. 4.8.3).
The overall pattern of decline is much more dramatic than during any prior or subsequent biotic
crises; actual species diversity drops from approximately 87 taxa in the mid clingani Biozone to
fewer than 5 species in the middle of the com planatus Biozone (Fig. 4.8.1). When the
biostratigraphic subzonation of the clingani Biozone is taken into account, the number of species
lost during this period is fewer, but still dramatic. This biodiversity event could almost be
considered a 'double-event', with two consecutive and largely sustained extinction peaks
coupled with a relatively low and sustained speciation (Figs. 4.8.1, 4.8.2).
The middle to upper Dicellograptus anceps Biozone event?
Graptoloid biodiversity recovers slightly following the previous biotic crisis, peaking at
approximately 30 species in the middle anceps Biozone, but never fully recovers to the levels of
speciation seen earlier in the Ordovician (Fig. 4.8.1). The subsequent decline of incoming taxa
creates another possible diversity event, taking place from the middle anceps to upper anceps
Biozone. This may not be an actual 'event' as the resulting fall in biodiversity is relatively small;
however, it does appear to be sudden, even though changes in appearance and extinction of
species are not high (Figs. 4.8.1, 4.8.2). From here, biotic diversity plateaus through the
extraordinarius Biozone, gradually declining through the persculptus Biozone, resulting in
critically low numbers of graptoloid species at the Ordovician-Silurian boundary, certainly
fewer than those observed in English and Welsh sequences for this time.
In reference to the combined Ordovician graptoloid diversity data (Fig. 4.6), overall
diversity reaches a 'low' at the Ordovician/Silurian boundary; however, this diversity level is
not as critical as those seen during the complanatus Biozone or the Wenlock Series (Fig. 4.6).
Page 4.7

Silurian
Graptoloid biodiversity in the early Llandovery of Britain appears to have increased steadily
culminating in a Silurian diversity peak of approximately 53 taxa in the upper triangulatus
Biozone (Fig. 4.9.1). This rise corresponds with a global diversity high as regards graptoloid
species (Melchin et al. 1998). The late Parakidograptus acuminatus global extinction event of
Melchin et al. (1998) does not seem to be represented by the the species diversity curve
presented here (Fig. 4.9.1). Although a small decrease in species diversity is observed in the
upper acuminatus Biozone, it seems not significant, and despite the rate of incoming species
being high in the upper acuminatus to acinaces Biozone interval, it is not matched by the number
of species extinctions recorded (Figs. 4.9.1, 4.9.2). Thus, the possibility of high speciation
masking high extinction at this interval, as suggested by Melchin et al. (1998), does not seem to
hold true for the British graptoloid sequence. It is worth noting that the highest number of
incoming species takes place in the lower acinaces Biozone, but extinctions at this level are low.
Further work on these faunas may shed light on this ‘extinction event’ in the UK. Interestingly,
there is a large extinction peak representing the interval from the late cyphus Biozone to the early
triangulatus Biozone, during which time there were also large numbers of incoming taxa, the
sum effect on the species diversity curve being minimal (Figs. 4.9.1, 4.9.2). The peak values
for Silurian graptoloid biodiversity occur in the upper triangulatus Biozone, where there is a
correlation between significant numbers of incoming taxa, but also a relatively high number of
extinctions.
Graptoloid species diversity declines gradually from the upper triangulatus Biozone,
across the magnus Biozone. This is mirrored by a fall in new taxa, coupled with an abrupt
increase in species extinctions at the end of the magnus Biozone.
The late Monograptus convolutus Biozone- middle Stimulograptus halli Biozone event
The first significant graptoloid extinction event in the British Silurian sequence is initiated at the
end of the convolutus Biozone and takes place across the sedgwickii Biozone continuing to a
lesser degree well into the halli Biozone (Fig. 4.9.1). This corresponds well with the ‘early
Stimulograptus sedgwickii event’ of Melchin et al. (1998) and relates to the Bohemian
‘convolutus event’ of Storch (1995). In Britain, the onset of this event coincides with a dramatic
Page 4.8

increase in the number of species extinctions, coupled with a relatively low number of incoming
species (Figs. 4.9.1, 4.9.2). This crisis coincides with declines in both the % of species
extinctions and the ratio of appearances to disappearances (Figs. 4.9.3, 4.9.4). Melchin et a l
(1998) note that diversity is particularly reduced in their upper sedgwickii Biozone, equal to the
British halli Biozone, which they describe as a biozone of low species diversity. Although they
are correct in their observation that species diversity in the halli Biozone is lower than that of the
sedgwickii and guerichi biozones, the halli Biozone, which contains more than 30 species, is
not a low diversity interval in the UK.
Overall diversity increases again in the middle of the guerichi Biozone, as the number of
incoming species exceeds the number of extinctions, although it should be noted that the
extinction rate is still relatively high (Figs. 4.9.1, 4.9.2). The subsequent increase, together
with a declining speciation rate leads to a second extinction event.
The late Spirograptus guerichi Biozone- middle Monoclimacis crenulata Biozone event
Initiated in the upper guerichi Biozone, this biotic crisis is more devastating to overall graptoloid
biodiversity than the preceeding event, and one that the graptoloids never fully recover from
(Fig. 4.9.1). The diversity drop continues steeply into the middle turriculatus Biozone, and
from there decreases gradually into the upper crenulata Biozone. This corresponds with both the
local and global utilis events of Storch (1995), Kaljo et a l (1995) and Melchin et a l (1998),
although much of the decline takes place across the turriculatus Biozone. The % of species
extinctions reaches a peak of about 75% in the crispus Biozone, this number gradually
decreases before stabilising in the crenulata Biozone. This decline is mirrored by a decrease in
the species appearance to disappearance ratio, although this is seen to increase slightly in the
griestoniensis Biozone (Fig. 4.9.3, 4.9.4).
The late Spirograptus guerichi- middle Monoclimacis crenulata event is followed by a
rapid diversity increase from the upper crenulata Biozone to the lower spiralis Biozone. This
rise is preceeded by an extinction decrease, and corresponds with a dramatic rise in the number
of incoming species (Figs. 4.9.1, 4.9.2). The subsequent biodiversity decline in the spiralis
Biozone is not significant, and diversity remains relatively constant until the middle of the
lapworthi Biozone. The significant spiralis extinction event observed by Storch (1995) in
Bohemia is not seen in the UK. Species diversity drops quite sharply in the middle of the
Page 4.9

lapworthi Biozone, however, and the decline here is comparable to the decline occurring across
the magnus Biozone, as is the number of extinctions, although the number of incoming taxa is
less in the middle part of the lapworthi Biozone (Figs. 4.9.1, 4.9.2). This situation highlights
some important considerations to be taken into account when defining a species biodiversity
event, namely, even though the magnitude of an extinction may not be significant, does the
abruptness of the resulting species diversity change constitute an 'event'? This diversity
decrease, and its accompanying extinction peak, correspond well with the C. lapworthi
extinction event of Melchin et al. (1998), which they believe equates with the Bohemian spiralis
event (Storch 1995), but it seems not sufficiently well-marked in the UK to be referred to as a
discrete species biodiversity event.
Following the latter diversity decrease, speciation increases, developing a ‘step-like’
pattern and reaches its highest level since the late guerichi Biozone in the lower part of the
centrifugus Biozone (Fig. 4.9.1). Graptoloid biodiversity is stable through to the lower part of
the murchisoni Biozone.
The early Cyrtograptus murchisoni Biozone- middle Monograptus firmus Biozone event
A sudden increase in species extinction and a relatively low, though rising, number of incoming
species in the lower murchisoni Biozone led to another Silurian biodiversity crisis (Figs. 4.9.1,
4.9.2). Diversity decreases dramatically early in the m urchisoni Biozone and continues
decreasing steadily through the biozone and into the firmus Biozone. This depletion of taxa is
dramatic and results in the lowest overall graptoloid species diversity thus far for the British
Silurian (Fig. 4.9.2). From this interval onwards, graptoloid diversity in the UK fluctuates
rapidly, while generally increasing into the upper part of the lundgreni Biozone. Although this
biodiversity crisis correlates with a decline in the % of species extinctions (Fig. 4.9.4), the ratio
of species appearances to disappearances is seen to fall markedly (Fig. 4.9.3). This murchisoni
event is recognised in other regions as well as globally (Kaljo et al. 1995, Melchin et al. 1998,
Storch 1995).
Page 4.10

The late Cyrtograptus lundgreni Biozone- early Gothograptus nassa Biozone event
The late lundgreni to early nassa biozones display a further extinction event, and one which is
probably the last in the British graptoloid sequence. It is preceeded by a low, but relatively
significant number of disappearances of taxa, during a period of low species diversity (Fig.
4.9.2). Graptoloid species diversity reaches its lowest point in the Silurian so far, and this is a
globally recognised extinction event (Jaeger 1991, Koren 1991, Kaljo et a l 1995, Storch 1995,
Urbanek 1993, Melchin et al. 1998). This event coincides with a decline in the % of species
extinctions and the ratio of species appearances to disappearances (Figs. 4.9.3, 4.9.4).
From this level onwards, speciation remains very low, with the exception of an abrupt
but short-lived increase in the nilssoni to scanicus biozones (Fig. 4.9.1). Although extinction
rate is never high, it is consistent: as a result, from the mid-scanicus Biozone onwards,
graptoloid diversity gradually declines until the whole group disappears from the British
stratigraphic sequence at the end of the Bohemograptus proliferation Biozone (Figs. 4.9.1,
4.9.2). Globally graptoloids continue, but the leintwardinensis event and subsequent upper
Silurian extinction events described by other authors cannot be indentified in Britain (Urbanek
1993, Storch 1995, Melchin et a l 1998).
Discussion
Having outlined patterns of graptoloid biodiversity change through the Ordovician and Silurian
of the UK, it is important to examine how these changes may have been generated, taking into
account global palaeoenvironmental trends and possible local variations. Many authors have
noted the relationship between graptoloid diversity and environmental fluctuation (Koren &
Rickards 1980, Koren 1987, Berry & Wilde 1990, Loydell 1994, 1998, Storch 1995, Melchin
et a l 1998), invoking a complex combination of biotic and abiotic factors to account for
diversifications and extinctions occurring within the group. Eustatic sea-level change has long
been accepted by palaeontologists as the driving mechanism for major changes in the biotic
realm (Newell 1967, Hallam 1992, Fortey 1984, Koren 1987), with the general belief that mass
extinctions correlate with large-scale regressions, while mass speciation is linked to overall
transgressions (Hallam 1992, Storch 1995). Other important palaeoenvironmental factors
include climate and oceanic circulation, temperature and chemistry, and attempts have been
made to understand palaeo-oceanic and atmospheric changes, largely based on palaeogeographic
Page 4.11

reconstructions, isotope geochemistry, and modem oceanographic studies (Spjeldnaes 1961,
1978, Berry & Wilde 1990, Wilde et al. 1991, Brenchley et al. 1994, Zalasiewicz et al. 1995).
However, any suggestion that faunal changes are the result of one, or a dominant environmental
factor are likely to be incorrect. The often subtle, complex interplay of all these environmental
elements, as observed in the modem realm, and their relationship to the biota, have important
implications for the way in which we view palaeontological information, and how it may be
integrated with other geological evidence to reconstruct palaeoenvironmental processess. The
study of ancient environmental systems is further complicated by the potential loss of
information through geological time: however, it is still possible to assess the validity of such
data in relation to other studies.
Eustatic sea-level change has been widely quoted as a primary cause for major changes
in faunal diversity and many attempts have been made to correlate between patterns of
graptoloid biodiversity and those of global sea-level fluctuations (Newell 1967, Hallam 1992,
Fortey 1984, Koren 1987, Melchin et al. 1998). However, there are problems associated with
the production of eustatic sea-level curves. Most sea-level curves have been produced by a
combination of lithostratigraphic information from key sections and biostratigraphy and water
depth analysis of benthic organisms (Johnson et al. 1991, Ross & Ross 1996, Johnson 1987).
However, it has previously been argued that the distribution of benthic organisms is too
susceptible to a variety of environmental factors, many of which may be unrelated to sea-level
change, to be relied upon as eustatic indicators (Aldridge et al. 1993, Jeppsson 1990, Loydell
1998), also, the biostratigraphy of benthic groups is not clearly constrained. As a result, the
sea-level curves produced from such studies are highly generalised and lack the refinement
required to a direct and meaningful comparison with more detailed diversity studies for
individual groups of fossils. However, there are examples of possible correlation between the
Ordovician sea-level curve produced by Fortey (1984), largely from lithological evidence, and
the graptoloid biodiversity curve generated in the present study, although this is not always the
case. For example, there is a global sea-level low at the base of the Arenig Series, followed by a
steady transgression through this interval which corresponds with the overall trend of gradually
increasing graptoloid species biodiversity in the Arenig of England and Wales (Fig. 4.3).
However, the generalised sea-level curve (Fortey 1984) does not provide any correlation with
Page 4.12

the late Corymbograptus simulans-early Isograptus victoriae graptoloid diversity crisis (middle
Arenig Series) seen in the Anglo-Welsh sequence. The period of maximum sea-level low
recognised by Fortey (1984) at the Arenig-Llanvim boundary, is not recorded by Leggett et al.
(1981), and is not reflected by a decline in UK graptoloid biodiversity (Fig. 4.3). Indeed, as the
diversity curve shows (Fig. 4.3), this represents a period of high graptoloid biodiversity,
culminating in the highest recorded levels for the Ordovician of England and Wales during the
late artus Biozone. However, this eustatic low does correlate with a significant number of
species extinctions, but the effect on overall biodiversity is masked by an equally large number
of species appearances at this time (Figs. 4.7.1, 4.7.2).
In an attempt to produce a eustatic sea-level curve for the Silurian, Loydell (1998) used
the well constrained and highly refined graptoloid biostratigraphy and accompaying facies
changes as the basis of his work. The result was a more detailed pattern of sea-level change than
those previously published, showing more rapid eustatic fluctuations which, not suprisingly,
correspond closely to patterns of graptoloid biodiversity change outlined here, with most of the
major UK graptoloid diversity crises and recoveries being coincident with eustatic sea-level fall
and rise respectively (Fig. 4.10). However, the graptoloids are no less susceptible to several
environmental elements than benthic organisms, particularly if water mass specificity is invoked
as the principal factor in graptoloid distribution, which Loydell (1998) supports. Indeed the
reliance on any single group of fossils as indication of global sea-level change is fraught with
possible inconstancies and not until there is a much greater correlative integration of a number of
biostratigraphically useful groups will it be possible to produce a more meaningful and
hopefully more accurate eustatic sea-level curve, a fact acknowledged by Loydell (1998).
Climate is a major factor affecting oceanic environments and consequently, faunal
diversity (Holland 1971, Boucot 1975, Frakes et al. 1992, Spjeldnaes 1961, 1978, 1981,
Jeppsson 1990, Wilde & Berry 1986, 1991). Frakes et al. (1992) were able to produce a broad-
scale picture of global climate changes through the Ordovician and Silurian based largely on the
work of previous authors with regard to lithostratigraphic, palaeontological and tectonic
evidence. However, as with many of the global sea-level curves produced for these intervals,
this general pattern of climate change cannot provide an explanation for many of the more
detailed changes in graptoloid biodiversity. Frakes et al. (1992) describe the period from the
Page 4.13

Tremadoc to the Llanvim stages as being generally warm and containing well developed faunal
provinces. Certainly in England and Wales at this time the overall biodiversity pattern of
graptoloids was one of general increase, reaching peak species diversity in the early part of the
Llanvim Stage (artus Biozone) (Fig. 4.3). The late Corymbograptus simulans-ezr\y Isograptus
victoriae graptoloid diversity crisis (middle Arenig Series) and the late Didymograptus artus-
early Glyptograptus teretiusculus biozone graptoloid diversity crisis both occur in this interval,
although Frakes et al. (1992) do not note any climatic fluctuation at these times (Fig. 4.3,4.4).
Spjeldnaes (1961, 1978), however, on some of whose work Frakes et al. (1992) base their
assumptions, recorded a period of marked global climatic temperature decrease coincident with
the Arenig-Llanvim boundary. Also, Fortey (1984) reported that the 'Llandeilo Series' (now
incorporated into the late Llanvim/early Caradoc series: Fortey et al. 1995), and the Caradoc
Series were marked by regressive eustatic events; with the Llandeilo being characterised by a
regression and ensuing transgression, based largely on the findings of Wilcox & Lockley’s
(1981) work in the type area. This combined evidence would be coincident with the onset and
continuation of the late Didymograptus artus-Qarly Glyptograptus teretiusculus biozone
graptoloid species diversity crisis in England and Wales, and the subsequent diversity rise
through to the early Caradoc (Fig. 4.3). According to Frakes et al. (1992), the remainder of the
Ordovician (Caradoc and Ashgill series) and the early Silurian (Llandovery Series) was a glacial
period, with the late Caradoc representing the early phase during which faunal diversity began
to fluctuate, and the Ashgill/Llandovery representing the later stages of glaciation during which
there was considerable decline in faunal diversity. Certainly, in both Scotland, and England and
Wales, the late Caradoc/early Ashgill represents an interval of marked decline in graptoloid
species diversity, although the subsequent recovery is seen to occur very early in the
Llandovery, suggesting the influence of another environmental factor at this interval, most
probably the increase in eustatic sea-level (Fig. 4.3. 4.4). This hypothesis of a rather protracted
glacial period is at odds with the views of Brenchley et al. (1994), who, on the basis of oxygen
isotope data, believe that the early-middle Himantian glacial episode was very restricted in
duration. The UK graptoloid species biodiversity evidence suggests the possibility of a more
prolonged onset of glaciation, as outlined by Frakes et al. (1992), with a more rapid
biodiversity recovery in response to rising temperatures as suggested by Brenchley et al.
Page 4.14

(1994), although these changes in graptoloid biodiversity also correspond with significant
changes in eustatic sea-level.
The Frakes et al. (1992) conclusions agree broadly with those of Spjeldnaes (1961,
1978, 1981), how ever, all these workers warn against the heavy reliance on one
palaeoenvironmental factor as an explanation for faunal diversity trends, and stress the need for
integration between data for palaeoclimatic and eustatic sea-level change. On the basis of the
graptoloid species biodiversity evidence presented here, it cannot be conclusively noted that
periods of crisis or diversification correlate with recorded changes in temperature as they are
seen to occur during periods of warming and cooling. However, it is undisputed that the major
climatic changes of the Ashgill Series (the height of the late Ordovician glaciation) do coincide
with a catastrophic decline in graptoloid species, although this may be a result of the speed and
intensity of the climatic change. Also, the late Wenlock graptoloid species decline coincides with
a regression.
The comparison of the UK graptoloid biodiversity curve presented here, together with
other published global sea-level curves and palaeoclimatic data (Leggett et al. 1981, Johnson et
al. 1991), provides some correlation on a very broad scale. However, there are considerable
differences when a close comparison is made; this is probably due to the differences of scale
between the two types of study. Loydell (1998) has pointed out that the P and S model cycles of
Jeppsson (1990), relating to periods of specific combined environmental factors including
plankton productivity, are not always correlatable with observed changes in graptoloid diversity
and their corresponding lithologies. It may occasionally, though, be possible to link the findings
proposed for this model with certain changes in graptoloid species biodiversity. For example,
the Jong Primo episode indicating high planktic production and believed to correlate, in part,
with the convolutus Biozone (Aldridge et al. 1993), would correspond with an interval of peak
graptoloid biodiversity in the UK (Fig. 4.2).
Page 4.15

Morphological change in Silurian graptoloids
Familial- and Generic-level composition
Fundamental morphological differences can be observed between organisms at the Family level.
During the Silurian five major graptoloid families occur, each with distinctive rhabdosomal
variations. The Diplograptidae Lapworth, 1873 were biserial; the Monograptidae Lapworth,
1873 were uniserial; the Dimorphograptidae Lapworth, 1876 had a biform rhabdosome, that is a
uniserial proximal end, but were biserial distally. The Retiolitidae Lapworth, 1873 were also
biserial; however, their rhabdosome was composed of a highly characteristic mesh-like
structure. The Cyrtograptidae Boucek, 1933 had a monograptid-type uniserial rhabdosome, but
were also the main Silurian group to possess thecal cladia. Using the new biostratigraphical
range charts (see chapter 3), I was able to calculate the percentage of the total graptoloid fauna
per biozone occupied by each family, as well as obtaining the actual numbers of species
representing each family, making it possible to trace the pattern of change seen in basic
graptoloid morphology through the Silurian (Fig. 4.11, 4.12).
The base of the Silurian shows a graptoloid fauna dominated by species from the family
Diplograptidae; there were also significant numbers of Dimorphograptidae and rarer
Monograptidae which formed less than 10% of the total assemblage (Figs. 4.11, 4.12). The
increase in numbers of monograptid species is dramatic in the atavus Biozone (Fig. 4.9.1); at
the start of the Aeronian most graptoloids possessed a monograptid-type rhabdosome although
more than 40% of the total fauna was still diplograptid. This correlates with the peak UK
Silurian graptoloid species diversity during the tr ia n g u la tu s Biozone. Numbers of
dimorphograptids decreased markedly by the triangulatus Biozone and continued to do so
through the remainder of the Aeronian stage. By the convolutus Biozone, a new family of
graptoloids, the Retiolitidae, had appeared, but these formed a very small proportion of the total
assemblage. The Diplograptidae continued to decline steadily, and their apparent increased
fortune in the sedgwickii Biozone is presumably a reflection of a more significant drop in the
number of monograptids during the late convolutus-middle halli biotic crisis (Fig. 4.9.2). This
graptoloid extinction event also saw the disappearance of the Dimorphograptidae (Fig. 4.11,
4.12). The ensuing species diversity crisis, the late guerichi-middle crenulata event, effected the
demise and loss of the Diplograptidae by the griestoniensis Biozone. The subsequent speciation
Page 4.16

event in the crenulata Biozone coincides with the initiation of another graptoloid family, the
Cyrtograptidae (Figs. 4.11, 4.12, 4.9.1). The cyrtograptids comprised a significant percentage
of the total graptoloid fauna up to the murchisoni Biozone, and their rise is mirrored by the
retiolitids; despite this the Monograptidae remain dominant.
The early murchisoni-middle firmus biodiversity event saw a decline in the proportion of
the graptoloid fauna occupied by retiolitid and cyrtograptid species (Fig. 4.11, 4.12); indeed,
the Retiolitidae disappear following this crisis, but reappear at the end of the Sheinwoodian.
Whether this is a true extinction and re-emergence, or an artefact of palaeontological sampling
methods is unclear, especially given the relatively short absence interval, and the fact that overall
numbers of retiolitid taxa are few. The same arguments may be applied to the absence of the
Cyrtograptidae during the dubius Biozone (Fig. 4.11, 4.12).
Despite a general increase in biodiversity up to the lundgreni Biozone, total numbers of
graptoloid species remain relatively low (Fig. 4.9.1). This generates a marked change in
composition during the lundgreni Biozone, with monograptids occupying approximately 50%
of the assemblage, their lowest percentage since the early Silurian. The remainder of the
assemblage is composed principally of cyrtograptids and retiolitids. The late lundgreni Biozone
represents the onset of another graptoloid biotic crisis in the UK (Fig. 4.9.2), the aftermath of
which saw the decline and loss of both the Cyrtograptidae and Retiolitidae from the British
graptoloid sequence. UK graptoloids, from the scanicus Biozone until their total extinction in
the Ludfordian Stage, remain exclusively of monograptid-type morphology (Fig. 4.11, 4.12).
M ore subtle morphology variations through the Silurian are observed when the
proportions of the total graptoloid fauna are analysed at generic level. For this part of the study,
10 relatively taxonomically uncontentious genera, representing most of the Silurian graptoloid
families covering the whole system were selected for analysis (Fig. 4.13).
What is immediately clear from the faunal percentage per biozone plots (Fig. 4.13), is
that most of these genera are biostratigraphically restricted (< 10 biozones). The most notable
exception to this is the genus Pristiograptus, which ranges through virtually the entire Silurian,
although it is absent for much of the early Wenlock. With the exception of two genera,
Pristiograptus and Streptograptus, all of these graptoloid groups have their acme during periods
of peak/rising numbers of incoming taxa (Figs. 4.13, 4.9.1). The pristiograptids reached peak
Page 4.17

percentage of the overall graptoloid assemblage (approx. 60%) in the nassa Biozone, which
corresponds with an interval when the number of graptoloid species was greatly reduced; thus
this long-ranging genus formed the bulk of the total assemblage. The streptograptids obtained
their highest percentage during a period when the number of new species was still significantly
high but falling (Figs. 4.13, 4.9.2). Most genera become extinct or become a significantly less
important proportion of the assemblage either during, or shortly after one of the major Silurian
graptoloid biotic crises; of those remaining, the pristiograptids and saetograptids disappear
coincident with the total UK graptoloid extinction in the Ludlow Series (Fig. 4.13).
Interestingly, few of the genera chosen for this study have their total percentage fauna peak
during the same biozone as any other genus. This also seems largely to be the case with
Bulman’s (1970) analysis of the stratigraphic distribution of graptoloid genera, with the larger
peaks being relatively short lived, although he did not quantify his distributions, and these
diversity peaks are not as pronounced as those presented here (Fig. 4.13). This is probably the
result of more recent and more highly constrained generic taxonomy. Bulman (1970) did,
however, analyse a larger number of genera than is presented here, and those genera with
smaller, generally prolonged peak diversity, do tend to overlap stratigraphically. This trend is
also seen in a similar plot by Rickards et al. (1977). The only anomalous groups are the
dimorphograptids/rhaphidograptids, which obtain their greatest percentage of the assemblage in
the atavus Biozone, along with the atavograptids.
Page 4.18

AgeChrono- strati graphic Biostratigraphic
Bohemograptus
Saetograptusleintwardinensis
SaetograptusincipiensLudlow
Lobograptus
N eodiversograptusnilssoni
Pristiograptusludensis
Gothograptus
Cyrtograptuslundgreni
Cyrtograptusrigidus
Wenlock Pristiograptusdubius
M onograptusriccartonensis
M onograptusfirm us
Cyrtograptusmurchisoni
Cyrtograptuscentrifugus
Cyrtograptusinsectus
Cyrtograptuslapworthi
O ktavitesspiralis
M onoclim aciscrenulata
M onoclim acisgriestoniensis
M onograptuscrispus
Spirograptusturriculatus
Spirograptusguerichi
Llandovery Stimulograptushalli
Stimulograptussedgwickii
M onograptusconvolutus
Pribylograptusleptotheca
Diplograptusmagnus
M onograptustriangulatus
Coronograptuscyphus
Lagarograptus
A tavograptusatavus
Parakidograptusacuminatus ~~
O t— t o U> L /i OsO O O O O O
Number of Graptoloid Species
Fig. 4.1. UK Silurian graptoloid species diversity (with zonal boundary values).
Fig. 4.1

AgeChrono-stratigraphic Biostratigraphic
Bohemograptus
Saetograptusleintwardinensis
Ludlow Saetograptusincipiens
Lobograptus
N eodiversograptusnilssoni
Pristiograptusludensis
G othograptus
Cyrtograptuslundgreni
Cyrtograptusrigidus
Wenlock Pristiograptusdubius
M onograptusriccartonensis
M onograptus_________ firm usCyrtograptus
murchisoni
C yrtograptuscentrifugus
Cyrtograptusinsectus
Cyrtograptuslapworthi
O ktavitesspiralis
M onoclim aciscrenulata 7 N ~
M onoclim acisgriestoniensis
M onograptuscrispus
Spirograptusturriculatus
Spirograptusguerichi
Llandovery Stimulograptushalli
Stimulograptus _______ sedgwickii
M onograptusconvolutus
Pribylograptusleptotheca
D iplograptusmagnus
M onograptustriangulatus
Coronograptuscyphus
Lagarograptus
A tavograptusatavus
Parakidograptusacuminatus ~~
U>OO
Number o f Graptoloid Species
Fig. 4.2. UK Silurian graptoloid species diversity (without zonal boundary values), showing mainbiodiversity crises.
Fig. 4.2

AgeChrono-stratigraphic Biostratigraphic
Glyptograptuspersculptus
Dicello graptus ancepsAshgill
Dicellograptuscomplanatus
Pleurograptuslinearis
Dicrano graptus clingani
_Caradoc Diplograptusmultidens
Nemagraptusgracilis
Glyptograptusteretiusculus
DidymograptusmurchisoniLlanvirn
Didymograptusartus
Aulo graptus cucullus
Isograptus gibberulus
Isograptusvictoriae
ArenigCorymbograptussimulans
Corymbograptusvaricosus
Tetragraptusphyllograptoides
AraneograptusmurrayiTremadoc
oI s )O to u>o u>UiOlo o
Number of Graptoloid Species
Fig. 4.3. Ordovician graptoloid species diversity for England & Wales (without zonal boundary values),showing main biodiversity crises.
Fig. 4.3

AgeChrono-stratigraphic Biostratigraphic
Glyptograptuspersculptus
Climacograptus ? extraordinarius
AshgillDicello graptus _ anceps
Dicellograptuscomplanatus
Pleurograptuslinearis
Dicranograptus __ clingani
Clim acograptus.. wilsoniCaradoc
Climacograptuspeltifer
Nemagraptusgracilis
Castlemainian 3 - -
Chewtonian 1-2 --
Arenig
Bendigonian 1-2"
Lancefieldian 3 0
-jo4o O nO VOOt oo u>o ooOo o
Number of Graptoloid Species
Fig. 4.4. Ordovician graptoloid species diversity for Scotland (without zonal boundary values), showingmain biodiversity crises.
Fig. 4.4

Age (biostratigraphic)
Age (biostratigraphic)
Age (biostratigraphic)
clingani
persculptus
anceps
cumplanatus
linearis
morrisi
caudatus
multidens
gracilis
teretiusculus
murchisoni
artus
cucullus
gibberulus
victoriae
Simula ns
varicosus
phyllograptoides
murrayi
persculpms
clingani i * |
i multidens
> gracilis
O teretiusculus
------------------------------------------ U ] murchisoni
amis
'• ■ 1 cucullus
gibberulus ^
victoriae
1 simuUns $ ]
V X " .....J
varicosus *
phyllograptoides
murrayi
g K g
Total Species Diversity
5 S K S
T o ta l S p e c ie s D iv e rs ity
anceps
clingani
persculptus
extraordinanus
padncus
complexus
complanatus
: ■. •
______________ Wm Wm 1111 1
Castlcinainiun 3
Chewtoman 1-2
Bendigoman 1-2
Lancefieldian 3
g a sTotal Species Diversity
persculptus
extraordinanus
anceps
complanatus
linearis
clingani
wilsoni
peltifer
gracilis
Castlemainian 3
Chewlonian 1-2
Bcndigonian 1-2
Lancefieldian 3
>eotoo s 00oo o o
T o ta l S p e c ie s D iv e rs ity
guerichi
mograptus leintwardinensis
incipiens
niissotu
nccan
creijulata gnestoniensis
sarlorius cnspu
galaensis camicus proteu
johnsonae
renaudigcmniatu .......
Bohemograptus leintwardinensis
incipiens scanicus nilssoni ludcnsis
nassa lundgreni
rigidus dubius
riccartonensis firmus
murchisoni centrifugus
insechis lapworlhi
spiralis crenulata
griestoniensis g r crispus£■ turriculatus
guerichi halli
sedgwickii convolutus leptolheca
magnus triangulatus
cyphus acinaces
atavus acuminatus
g &T o ta l S p e c ie s D iv e r s i ty
s aT o ta l S p e c ie s D iv e rs ity
Fig. 4.5. Total species diversity/biozone for la & b. The Ordovician of England & Wales. 2a & b. The Ordovician of Scotland. 3a & b. The Silurian of the UK.
F ig . 4 .5

Graptoloid Biozone
Bohemog raptus leintwardinensis
incipiens scanicus nilssoni ludensis
nassa lundgreni
rigidus dubius
riccartonensis firmus
murchisoni centrifugus
insectus lapworthi
spiralis crenulata
griestoniensis crispus
turriculatus guerichi
halli sedgwickii convolutus leptotheca
magnus triangulatus
cyphus acinaces
atavus acuminatus
p e r s c u l p t u s ! extraordinanus & persculptusanceps
complanatuslinearis
clinganimultidens!peltifer & wilsoni
gracilis teretiusculus
murchisoni artus
cucullus gibberulus
victoriae simulans
varicosus phyllograptoides
murrayi
SILURIAN
ORDOVICIAN
0 a 00 H H H
Fig. 4.6. UK Ordovician and Silurian graptoloid species diversity (the Ordovician portion of the chart combines data from England and Wales and Scotland).
Fig. 4.6

AgeChrono-stratigraphic Biostratigraphic
Glyptograptuspersculptus
DicellograptusancepsAshgill
Dicellograptuscomplanatus
Pleurograptuslinearis
Dicrano graptus clingani
Caradoc Diplograptusmultidens
Nemagraptusgracilis
Glyptograptusteretiusculus
DidymograptusmurchisoniLlanvim
Didymog rapt us artus
Aulograptus cucullus
Isograptusgibberulus
Isograptusvictoriae
ArenigCorymbograptussimulans
Corymbograptusvaricosus
Tetragraptusphyllograptoides
AraneograptusmurrayiTremadoc
to U-> UJ 4 4^0o <-/» o ui o ui o cm
O o
No. of recorded graptoloid species No. of first appearances of graptoloids
Fig. 4.7.1. Comparison of Ordovician Anglo-Welsh graptoloid diversity and number of first appearance ofspecies.
Fig. 4.7.1

AgeChrono-stratigraphic Biostratigraphic
Glyptograptuspersculptus
DicellograptusancepsAshgill
Dicellograptuscomplanatus
Pleurograptuslinearis
Die rano graptus clingani
CaradocDiplograptusmultidens
Nemagraptusgracilis
Glyptograptusteretiusculus
Didymograptusmurchisoni
Llanvirn
Didymograptusartus
Aulograptuscucullus
Isograptusgibberulus
Isograptusvictoriae
ArenigCorymbograptussimulans
Corymbograptusvaricosus
Tetragraptusphyllograptoides
Araneo graptus murrayiTremadoc
-fkOO CM o CM O CM o CM
toCM
COOO o
No. of recorded graptoloid species No. of last occurrences of graptoloids
Fig. 4.7.2. Comparison of Ordovician Anglo-Welsh graptoloid diversity and number of last occurrence ofspecies.
Fig. 4.7.2

persculptus
anceps
complanatus
linearis
clingani
multidens
gracilis
oo'51wo'No3O
teretiusculus
murchisoni
artus
cucullus
gibberulus
victoriae
simulans
varicosus
phyllograptoides
murrayio u> U)
La
Species ratio (appearance/disappearance)
Fig. 4.7.3. Ratio of graptoloid species diversity (appearance/disappearance) for the Ordovician of England and Wales.
Fig. 4.7.3

Graptoloid
Biozon
e
persculptus
anceps
complanatus
linearis
clingani
multidens
gracilis
teretiusculus
murchisoni
artus
cucullus
gibberulus
victoriae
simulans
varicosus
phyllograptoides
murrayitoo
toUio o o o o o o o oo oo H- to
o o o
% sp e c ie s d isap p earin g A ctu a l no . o f sp ec ie s d isappearing
Fig. 4.7.4. Comparison of % species disappearance and actual number of species disappearance forgraptoloids of the Anglo-Welsh Ordovician.
Fig. 4.7.4

AgeChrono-stratigraphic Biostratigraphic
Glyptograptuspersculptus
Climacograptus ? extraordinanus
AshgillDicellograptusanceps
Dicellograptuscomplanatus
Pleurograptuslinearis
Dicrano graptus clingani
ClimacograptuswilsoniCaradoc
Climacograptuspeltifer
Nemagraptusgracilis
Castlemainian 3
Chewtonian 1-2
Arenig
Bendigonian 1-2
Lancefieldian 3
U> 4 ^ U i O n 0 0 v o Oo o o o o o o
o
No. of recorded graptoloid species No. of first appearances of graptoloids
Fig. 4.8.1. Comparison of Ordovician Scottish graptoloid diversity and number of first appearance ofspecies.
Fig. 4.8.1

AgeChrono- strati graphic Biostratigraphic
Glyptograptuspersculptus
Climacograptus ? extraordinanus
AshgillDicellograptusanceps
Dicellograptuscomplanatus
Pleuro graptus linearis
Dicrano graptus clingani
Climacograptuswilsoni
Caradoc
Climacograptuspeltifer
Nemagraptusgracilis
Castlemainian 3
Chewtonian 1-2
Arenig
Bendigonian 1-2
Lancefieldian 3
o I— to W ^ U s i 00 VO O o o o o o o o o o
to to u>O ot o
No. of recorded graptoloid species No. of last occurrences of graptoloids
Fig. 4.8.2. Comparison of Ordovician Scottish graptoloid diversity and number of last occurrence ofspecies.
Fig. 4.8.2

Graptoloid
Biozone
persculptus
extraordinanus
anceps
complanatus
linearis
clingani
wilsoni
peltifer
gracilis
Castlemainian 3
Chewtonian 1-2
Bendigonian 1-2
Lancefieldian 3ONo t o U)
Species ratio (appearance/disappearance)
Fig. 4.8.3. Ratio of graptoloid species diversity (appearance/disappearance) for the Ordovician of Scotland.
Fig. 4.8.3

persculptus
extraordinanus
anceps
complanatus
linearis
clingani
wilsoni
peltifer
gracilis
Castlemainian 3
Chewtonian 1-2
Bendigonian 1-2
Lancefieldian 3O h N ) W £ I A O \ s) O O ' 0 h O K) w w
0 UiHHu
% species disappearing Actual no. of species disappearing
Fig. 4.8.4. Comparison of % species disappearance and actual number of species disappearances forgraptoloids of the Scottish Ordovician.
Fig. 4.8.4

AgeChrono-stratigraphic Biostratigraphic
Bohemograptus
Saetograptusleintwardinensis
SaetograptusincipiensLudlow
Lobograptus
N eodiversograptusnilssoni
Pristiograptusludensis
Gothograptus
Cyrtograptuslundgreni
Cyrtograptusrigidus
PristiograptusdubiusWenlock
M onograptusriccartonensis
M onograptusfirm us
Cyrtograptusmurchisoni
Cyrtograptuscentrifugus
Cyrtograptusinsectus
Cyrtograptuslapworthi
O ktavitesspiralis
M onoclim aciscrenulata
M onoclimacisgriestoniensis
M onograptuscrispus
Spirograptusturriculatus
Spirograptusguerichi
Stimulograptushalli
LlandoveryStimulograptus
sedgwickii
M onograptusconvolutus
Pribylograptusleptotheca
D iplograptusmagnus
M onograptustriangulatus
Coronograptuscyphus
Lagarograptus
A tavograptusatavus
Parakidograptusacuminatus
Os O 4 ^O oo©No. of recorded graptoloid species No. of first appearances of graptoloids
Fig. 4.9.1. Comparison of British Silurian graptoloid diversity and number of first appearance of species.
Fig. 4.9.1

AgeChrono-stratigraphic Biostratigraphic
Bohemograptus
Saetograptusleintwardinensis
Ludlow Saetograptusincipiens
Lobograptus
N eodiversograptusnilssoni
Pristiograptusludensis
Gothograptus
Cyrtograptuslundgreni
Cyrtograptusrigidus
PristiograptusdubiusWenlock
M onograptusriccartonensis
M onograptusfirm us
Cyrtograptusmurchisoni
Cyrtograptuscentrifugus
Cyrtograptusinsectus
Cyrtograptuslapworthi
Oktavitesspiralis
M onoclim aciscrenulata
M onoclimacisgriestoniensis
M onograptuscrispus
Spirograptusturriculatus
Spirograptusguerichi
StimulograptushalliLlandovery
Stimulograptussedgwickii
M onograptusconvolutus
Pribylograptusleptotheca
D iplograptusmagnus
M onograptustriangulatus
Coronograptuscyphus
Lagarograptus
A tavograptusatavus
Parakidograptus- 1a s o to -Pxo
acuminatus
LnOO oo aso o
No. of recorded graptoloid species No. of last occurrences of graptoloids
Fig. 4.9.2. Comparison of British Silurian graptoloid diversity and number of last occurrence of species.
Fig. 4.9.2

Graptoloid
Biozone
Bohemograptus leintwardinensis
incipiens
scanicus
nilssoni ludensis
nassa lundgreni
rigidus
dubius
riccartonensis
firmus
murchisoni
centrijugus
insectus
lapworthi
spiralis
crenulata
griestoniensis
crispus
turriculatus
guerichi
halli sedgwickii
convolutus leptotheca
magnus
triangulatus
cyphus acinaces
atavus
acuminatus© I— to w ^ Ui ON
Species ratio (appearance/disappearance)
Fig. 4.9.3. Ratio of graptoloid species diversity (appearance/disappearance) for the Silurian of the UK.
Fig. 4.9.3

Graptoloid
Biozone
Bohemograptus
leintwardinensisincipiensscanicusnilssoniludensis
nassalundgreni
rigidusdubius
riccartonensisfirmus
murchisonicentrifugus
insectuslapworthi
spiraliscrenulata
griestoniensiscrispus
turriculatusguerichi
hallisedgwickiiconvolutusleptotheca
magnustriangulatus
cyphusacinaces
atavusacuminatus
in o\ n]0 0 0
K) W £0 0 0
H K) N) W W0 M
% species disappearing Actual no. of species disappearing
Fig. 4.9.4. Comparison of % species disappearance and actual number of species disappearances for UKSilurian graptoloids.
Fig. 4.9.4

AgeChrono-stratigraphic Biostratigraphic
Bohemograptus
Saetograptusleintwardinensis
SaetograptusincipiensLudlow
Lobograptus
N eodiversograptusnilssoni
Pristiograptusludensis HIGHLOW
Gothograptus
Cyrtograptuslundgreni
Cyrtograptusrigidus
PristiograptusdubiusWenlock
M onograptusriccartonensis
M onograptusfirm us
Cyrtograptusmurchisoni
Cyrtograptuscentrifugus
Cyrtograptusinsectus
Cyrtograptuslapworthi
O ktavitesspiralis
M onoclim aciscrenulata
M onoclim acisgriestoniensis
M onograptuscrispus
Spirograptusturriculatus
Spirograptusguerichi
Stimulograptushalli
LlandoveryStimulograptus
sedgwickii
M onograptusconvolutus
Pribylograptusleptotheca
D iplograptusmagnus
M onograptustriangulatus
Coronograptuscyphus
Lagarograplus
Atavograptusatavus
Parakidograptusacuminatus
O OsOu>OoGlobal sea-level curve,
No. of recorded graptoloid species after Loydell, 1998
Fig. 4.10. Comparison of eustatic sea-level, after Loydell 1998, and UK graptoloid species biodiversity, this study, for the Silurian.
Fig. 4.10

B oh em ograp tu s
le in tw ard in en sis
in cip iens
scan icu s
n ilsson i
ludensis
n assa
lundgreni
rig idu s —
dubius
riccarton en sis
firm u s
m urchison i
cen trifugus —
insectus
lapw orth i
sp ira lis
crenu la ta
g rieston ien sis
crispu s
tu rricu la tus
gu erich i
halli
sed g w ick ii
convolu tus
lep to th eca ~
m agnus
tr ian gu la tu s
cyphus
a cin a cespSH V
a tavu s -]
acum inatus
| Diplograptidae
] Monograptidae
| | Dimorphograptidae
g j Retiolitidae
] Cyrtograptidae
*
1
50 60 70 80 90 100
% of total fauna
F ig . 4 .1 1 . Area diagram showing percentage total biozone fauna occupied by each Silurian graptoloid family.

Graptoloid
Biozone
B ohem ogrciptus
le in tw ard in en sis
a c in aces
a tavu s
acu m inatus
| Diplograptidae
Monograptidae
[U Dimorphograptidae
[~~~| Retiolitidae
] Cyrtograptidae
incip ien s --
scan icus
n ilsson i
ludensis - 3
nassa
lundgreni
rig idus
dubius
riccarton en sis
firm u s
m urcliison i
cen trifugus
insectus
la pw orth i
sp ira lis —
crenulata
g rie s to m en sis
crispu s
tu rricu la tus
gu erich i
sed g w ick ii
con volu tu s
lep to th eca
m agnus
tr ian gu la tu s
cyphus
No. of species
Fig. 4.12. Area diagram showing the actual number of species per graptoloid family present in each Silurian biozone.
F ig . 4 .1 2

acuminatus
Graptoloid Biozone
S.G. o cro 5 i< c
_ O in n o n-■aso3 5 < «£a S . O S
(TO — “ ‘
3.5 v> x . c ^ *T C o ^ 3 - ET±2 3 t S< o S 3 £ S o £-^3 5 ELp a> c p c o e Jf. e- p- c ff 2. rj cr.C/5 C/5 C/5 C/5 C / 5 P C / 5 . —. — . — . C/5 C/5 C/5 P C/5
- « 5/5 P5 - U 9.
“ c/T 3 T3
— o S* S’c/5 £ . 3 C PCO. tn M
1— I— I— I— I— (— I— I— I— I— I— I— I— I— I— I— t
2 CD
B S* « 2 E2 3 H. o S-'B3 u2 . 3c/5 c/s
Petalolithus/Cephalograptus
Monoclimacis
Pristiograptus
Saetograptus
Dimorphograptus/Rhaphidograptus
Torquigraptus
Cyrtograptus
Streptograptus
XFJ
Mediograptus
o
Atavograptus
F ig . 4 .1 3
Fig.
4.1
3. P
erce
ntag
e of
tota
l fa
una
per
bioz
one
of se
lecte
d gr
apto
loid
ge
nera
.

Conclusions
Re-examination o f Saetograptid Type Material
The Ludlow Series graptoloid subfamily Saetograptinae Urbanek 1958 requires a full
taxonomic review of all species and subspecies within the group, in light of the re-examination
of the type material presented here. The type specimens and their original descriptions, along
with literature by a number of authors regarding subsequent identifications of these taxa,
provide strong evidence that the saetograptids have been 'over split' taxonomically, largely as a
result of intraspecific variation within the group. Fortunately, the holotypes of S. chimaera
chimaera and S. colonus colonus, the designated type specimens of the two saetograptid
subgenera Saetograptus and Colonograptus, are in a reasonable state of preservation and their
morphological concepts have not 'drifted' significantly in the literature. However, several other
species and subspecies are not as taxonomically well constrained. The status of much of the
type material is questionable, being either poorly preserved, missing or misidentified. Despite
these problems, three basic saetograptid morphologies can be determined based on the
theca/spine relationship; a chimaera-type theca (Fig. 2.2a), a leintwardinensis-type theca (Fig.
2.2d) and a colonus-type theca (Fig. 2.2i). In general, the group is biform and all species and
subspecies possess proximal colonus-type thecae and distal chimaera-typdleintwardinensis-type
thecae, or simple thecae without any spines or processes. However, there is a continuum of
forms between these gross morphologies.
British Graptoloid Biostratigraphy
New graptoloid biostratigraphic range charts are presented for the Ordovician and Silurian of
Britain. These include two separate biostratigraphic frameworks for the Ordovician, one for
England and Wales and another for Scotland, taking into account the considerable differences of
graptoloid faunal assemblages and correlation between these regions (Fig. 3.4.1-5, 3.5.1-3).
There is a uniform graptoloid biostratigraphic framework for the British Silurian (Fig. 3.6.1-8).
The updated schemes show 17 graptoloid biozones for the Anglo-Welsh Ordovician sequence
and 13 for that of Scotland. The Silurian is subdivided into 32 biozones and 10 sub-biozones.
Page 5.1

The charts provide stratigraphic ranges for all UK graptoloid species and subspecies together
with full supporting taxonomic and biostratigraphic reference information.
Graptoloid Biodiversity and Morphological Change
Using the new graptoloid range charts as a data source, several significant biodiversity crises
can be identified within British graptoloids through the Ordovician and Silurian. Many
environmental factors affect the biodiversity of the oceans, such as climate, oceanic circulations,
temperatures and chemistry; but on the basis of existing geological evidence, the diversity
events displayed by UK graptoloids most closely correlate with changes in eustatic sea-level
(Fortey 1984, Loydell 1998). Commonly, periods of high graptoloid diversity coincide with
intervals of relatively high eustatic sea-level, and low diversity correlate with a global
regression. Four biodiversity events occur in the Anglo-Welsh Ordovician and three in the
Scottish Ordovician, which in general, correspond with each other. The British Silurian
graptoloid sequence shows four biodiversity crises, which correlate well with other localised
and global graptoloid biodiversity trends (Koren 1987, Melchin et al. 1998, Storch 1995).
Tentative links between high levels of graptoloid extinction in the UK and generalised patterns
of global temperature change during the Ordovician can be made (Spjeldnaes (1961, 1978),
although these may not necessarily affect overall graptoloid biodiversity (Fig. 4.7.1, 4.7.2).
Changes in the morphology of British graptoloids at the family level display a marked
correlation with the periods of biodiversity crisis and speciation observed in this study for the
Ordovician and Silurian sequence (Fig. 4.11, 4.12), which are in turn associated with broader
scale environmental controls. Considerable biostratigraphic restriction of genus level graptoloid
morphology occurs among many UK graptoloids (Fig. 4.13), with little peak diversity overlap
between genera. This implies that even on a stratigraphically and morphologically fine scale the
dominance of a particular graptoloid form was being exerted.
Further Work
A complete taxonomic reassessment of the graptoloid subfamily Saetograptinae Urbanek 1958,
involving the collection and examination of large numbers of topotype specimens, is required to
test the legitimacy of the morphological concept behind many of the alleged species and
Page 5.2

subspecies of the group. This will prove difficult for taxa such as S. chimaera salweyi, for
which Wood's (1900) original locality is no longer accessible, and the holotype specimen is
merely a dubiously identified distal fragment. In such cases, it may be necessary, where a
persistent if informal morphological concept has been recognised in the literature, to designate a
new subspecies name and type specimen.
The taxonomic problems outlined above regarding the saetograptids are by no means an
isolated occurrence among graptoloids (Zalasiewicz pers. comm.). As a result, future research
on graptoloid taxonomy, as well as further biostratigraphical work in the UK, will ultimately
lead to progressive refinement and reassessment of the graptoloid range charts presented here.
As a comprehensive database on Ordovician and Silurian graptoloids from Britain, the
information contained in the new range charts can be utilised from a number of perspectives.
Further to this study however, the work on morphological variations within British Silurian
graptoloids could be applied to those of the British Ordovician. It may be possible to carry out
similar studies from other regions based on biostratigraphic data to produce a broad picture of
global graptoloid morphological change.
Page 5.3

REFERENCES
ALDRIDGE, R. J., JEPPSSON, L. & DORNING, K. J. 1993. Early Silurian oceanic episodes and events.
Journal o f the Geological Society, 150, 501-513.
* BAKER, S. J. 1981. The graptolite biostratigraphy of a Llandovery outlier near Llanystumdwy,
Gwynedd, North Wales. Geological Magazine, 118, 355-365. (1, 1)
BARNES, C. R. 1984. Early Ordovician eustatic events in Canada. In: BRUTON, D. L. (ed.) Aspects of the
Ordovician System, 51-63. Oslo Universitetsforlaget.
BARRANDE, J., 1850. Graptolites de Boheme. Systeme Silurien du Centre de la Boheme, Prague.
BECKLY, A. J. 1988. The stratigraphy of the Arenig Series in the Aberdaron to Sam areas, western Llyn,
north Wales. Geological Journal, 23, 321-337.
* BECKLY, A. J. & MALETZ, J. 1991. The Ordovician graptolites Azygograptus and Jishougraptus in
Scandinavia and Britain. Palaeontology, 34, 887-925. (2)
BERRY, W. B. N., 1964. Early Ludlow graptolites from the Presque Isle Quadrangle, Maine. Journal of
Paleontology, 38, 587-599.
BERRY, W. B. N. & WILDE, P. 1990. In: MCKERROW, W. S. & SCOTESE, C. R. (eds.) Palaeozoic
Palaeogeography and Biogeography, Geological Society Memoir, 12, 129-137.
* BGS (Unpublished Biostratigraphical Reports). (7 ,1)
BOUCEK, B., 1936. Graptolitova fauna ceskeho spondniho ludlowu. Rozpravy ceske Akademie, 46, 1-26.
BRENCHLEY, P. J., MARSHALL, J. D„ CARDEN, G. A. F., ROBERTSON, D. B. R. & LONG, D. G. F.
1994. Bathymetric and isotopic evidence for a short-lived Late Ordovician glaciation in a greenhouse period.
Geology, 22, 295-298.
BRIGGS, D. E. G., KEAR, A. J., BAAS, M., DE LEEUW, J. W. & RIGBY, S. 1995. Decay and composition
of the hemichordate Rhabdopleura: implications for the taphonomy of graptolites. Lethaia, 28, 15-23.
* BULMAN, O. M. B. 1963. On Glyptograptus dentatus (Brongniart) and some allied species. Palaeontology,
6 , 665-689. (8)
BULMAN, O. M. B. 1964. The Lower Palaeozoic Plankton. Quarterly Journal o f the Geological Society of
London, 120, 455-476.
BULMAN, O. M. B. 1970. Graptolithina with sections on Enteropneusta and Pterobranchia. In: TEICHERT, C.
(ed.) Treatise in Invertebrate Palaeontology, Part V (2nd Edition). Geological Society of America and
ref 1

University of Kansas Press.
CAMERON, I. B., STONE, P. & SMELLIE, J. L. 1986. Geology of the country around Girvan.
Explanation o f 1:50 000, British Geological Survey, Sheet 7 (Scotland).
* COCKS, L. R. M. & TOGHILL, P. 1973. The biostratigraphy of the Silurian rocks of the Girvan District,
Scotland. Journal o f the Geological Society4 London, 129, 209-243. (2)
* COOPER, A. H., RUSHTON, A. W. A., MOLYNEUX, S. G., MOORE, R. M. & WEBB, B. C. 1995.
The stratigraphy, correlation, provenance and palaeogeography of the Skiddaw Group (Ordovician) in the
English Lake District. Geological Magazine, 132, 185-211. (10)
COOPER, R. A. & FORTEY, R. A. 1982. The Ordovician graptolites of Spitsbergen. Bulletin o f the British
Museum (Natural History), Geology, 36, 157-302.
COOPER, R. A. & LINDHOLM, K. 1990. A precise worldwide correlation of early Ordovician graptolite
sequences. Geological Magazine, 127, 497-525.
CROWTHER, R. R. & RICKARDS, R. B. 1977. Cortical bandages and the graptolite zooid. Geologica and
Palaeontologica, 11, 9-46.
CUERDA, A. J., 1965. Monograptus leintwardinensis var. incipiens Wood en el Silurico de la Precordillera.
Ameghiniana, 4, 171-178.
* DAVIES, K. A. 1929. Notes on the graptolite faunas of the Upper Ordovician and Lower Silurian.
Geological Magazine, 66, 1-27. (11, 3)
* DAVIES, J. R., FLETCHER, C. J. N„ WATERS, R. A., WILSON, D., WOODHALL, D. G. &
ZALASIEWICZ, J. A. 1997. Geology of the county around Llanilar and Rhayader. Memoir of the British
Geological Survey, Sheets 178 and 179 (England and Wales). (5)
DILLY, P. N. 1986. Modern Pterobranchs: Observations on their behaviour and tube building. In: HUGHES, C.
P., RICKARDS, R. B. & CHAPMAN, A. J. (eds.) Palaeoecology and Biostratigraphy of graptolites.
Geological Society Special Publication, 20, 261-269.
EARP, J. R., 1944. Observations on Upper Silurian graptolites. Geological Magazine, 81, 181-185.
ELLES, G. L. 1900. The zonal classification of the Wenlock Series of the Welsh Borderland. Quarterly Journal
of the Geological SocietyA London, 56, 370-414.
ELLES, G. L. 1904. Graptolite zones in the Arenig rocks of Wales. Geological Magazine, 1, 199-211.
ELLES, G. L. 1925. The characteristic assemblages of the graptolite zones of the British Isles. Geological
M agazine, 62, 337-347.
ref 2

* ELLES, G. L. 1933. The Lower Ordovician graptolite faunas with special reference to the Skiddaw Slates.
Summary o f Progress. Geological Survey o f Great Britain (for 1932), 94-111. (12)
ELLES, G. L. 1937. The classification of the Ordovician rocks. Geological Magazine, 7 4 , 481-495.
* ELLES, G. L. 1940. The stratigraphy and faunal succession in the Ordovician rocks of the Builth-
Llandrindod inlier, Radnorshire. Quarterly Journal of the Geological Society of London, 95, 383-445. (13)
* ELLES, G. L. & WOOD E. M. R. 1901-18. A monograph o f British graptolites. The Palaeontographical
Society, London. (14, 4)
FINNEY, S. C. & BERGSTROM, S. M. 1986. Biostratigraphy of the Ordovician Nemagraptus gracilis Zone.
In: HUGHES, C. P. & RICKARDS, R. B. (eds.) Palaeoecology and Biostratigraphy o f Graptolites.
Geological Society Special Publication, 20 ,47-59.
FORTEY, R. A. 1984. Global earlier Ordovician transgressions and regressions and their biological
implications. In: BRUTON, D. L. (ed.) Aspects o f the Ordovician System, 37-50. Oslo
Universitetsforlaget.
FORTEY, R. A. & COCKS, L. R. M. 1986. Marginal faunal belts and their structural implications, with
examples from the Lower Palaeozoic. Journal o f the Geological Society o f London, 1 4 3 , 151-160.
* FORTEY, R. A., BECKLY, A. J. & RUSHTON, A. W. A. 1990. International correlation of the base of the
Llanvirn Series, Ordovician System. Newsletters on Stratigraphy, 22, 119-142. (16)
FORTEY, R. A., HARPER, D. A. T., INGHAM, J. K., OWEN, A. W. & RUSHTON, A. W. A. 1995. A
revision of Ordovician series and stages from the historical type area. Geological Magazine, 132, 15-30.
FORTEY, R. A., HARPER D. A. T., INGHAM, J. K., OWEN, A. W. & RUSHTON, A. W. A. 1996.
Discussion on a revision of Ordovician Series and Stage divisions from the historical type area-Reply.
Geological Magazine, 133, 770-772.
FORTEY, R. A. & COOPER, 1986. A phylogenetic classification of the graptoloids. Palaeontology, 29, 631-
654.
* FORTEY, R. A. & OWENS, R. M. 1987. The Arenig Series in South Wales. Bulletin o f the British
Museum (Natural History) Geology, 4 1 , 69-307. (15)
FORTEY, R. A. & OWENS, R. M. 1990. Arenig biostratigraphy and correlation in the Welsh Basin. Journal of
the Geological Society, London 1 4 7 , 607-610.
FRAKES, L. A., FRANCIS, J. E. & SYKTUS, J. I. 1992. Climate modes o f the Phanerozoic. Cambridge
University Press.
ref 3

FRECH, F., 1897. Lethaea geognostica. Vol. 1, Stuttgart.
GORTANI, M., 1923. Contributioni alio studio del paleozoico caraico. Faune a Graptoliti. Paleontographica
Italica, 26 (for 1920), 1-56.
GUTIERREZ-MARCO, J. C., LENZ, A. C., ROBARDET, M. & PigARRA, J. M. 1996. Wenlock-Ludlow
graptolite biostratigraphy and extinction: a reassessment from the south-western Iberian Peninsula (Spain
and Portugal). Canadian Journal of Earth Sciences, 33, 656-663.
GUTIERREZ-MARCO, J. C. & STORCH, P. 1998. Graptolite biostratigraphy of the Lower Silurian
(Llandovery) shelf deposits of the Western Iberian Cordillera, Spain. Geological Magazine, 135,71-92.
HALLAM, A. 1992. Phanerozoic sea-level changes. Perspectives in Paleobiology and Earth History Series,
Columbia University Press.
HARLAND, W. B., AGER D. V., BALL, H. W., BISHOP, W. W., BLOW, W. H„ CURRY, D., DEER,
W. A., GEORGE, T. N., HOLLAND, C. H., HOMES, S. C. A., HUGHES, N. F., KENT, P. E.,
PITCHER, W. S., RAMSBOTTOM, W. H. C., STUBBLEFIELD, C. J., WALLACE, P. &
WOODLAND, A. W. 1972. A concise guide to stratigraphical procedure. Journal o f the Geological
Society, London 128, 295-305.
* HARRIS, J. H. 1987. The geology of the Wenlock Shales around Builth Wells. Unpublished Ph.D. Thesis,
University of Cambridge. (5)
HOLLAND, C. H. & PALMER, D. C. 1974. Bohemograptus, the youngest graptoloid known from the British
Silurian sequence. In\ RICKARDS, R. B., JACKSON, D. E. & HUGHES, C. P. (eds.) Graptolite studies
in honour o f O. M. B. Bulman. Special Papers in Palaeontology, The Palaeontological Association,
London, 13, 215-236.
HOLLAND, C. H. & M. G. BASSETT, (eds.) 1989. A global standard fo r the Silurian System. National
Museum of Wales Geological Series.
HOLLAND, C. H., LAWSON, J. D. & WALMSLEY, V. G. 1963. The Silurian rocks of the Ludlow District,
Shropshire. Bulletin o f the British Museum (Natural History) Geology, 8, 95-171.
HOPKINSON, J. 1873. On the Occurrence of Numerous Species of Graptolites in the Ludlow Rocks of
Shropshire. Geological Magazine, 10, 519-520.
* HUGHES, R. A. 1989. Llandeilo and Caradoc graptolites o f the Builth and Shelve Inliers. The
Palaeontographical Society, London. (17)
HUGHES, R. A., 1995. The durations of Silurian graptolite zones. Geological Magazine, 132, 113-115.
ref 4

* HUTT, J. E. 1974-75. The Llandovery graptolites o f the English Lake District. The Palaeontographical
Society, London. (6)
HUTT, J., 1969. The development of the Ludlovian graptolite Saetograptus varians. Lethaia, 2, 361-368.
*HUTT, J. E. & RICKARDS, R. B. 1970. The evolution of the earliest Llandovery monograptids.
Geological Magazine, 107, 67-77. {18, 7)
INGHAM, J. K. 1979. The Moffat area. In: BASSETT, M. G., COCKS, L. R. M., HOLLAND, C. H.,
INGHAM, J. K., LAWSON, J. D., RICKARDS, R. B. & TEMPLE, J. T. Guidebook to field meeting,
Great Britain, March 30-April 11, 1979, Subcommission on Silurian Stratigraphy, Ordovician-Silurian
Boundary Working Groupp, I. U. G. S., 42-46.
INGHAM, J. K. & WRIGHT, A. D. 1970. A revised classification of the Ashgill Series. Lethaia, 3, 233-242.
JACKSON, D. E. 1978. The Skiddaw Group. In: MOSELEY, F. (ed.) The geology o f the Lake District.
Yorkshire Geological Society, Occasional Publication, 3, 79-98.
JAEGER, H., 1959. Graptolithen und Stratigraphie des jiingsten Thiiringer Silurs. Abhandlungen der deutschen
Akademie der Wissenschaften zu Berlin, 2, 1-197.
JAEGER, H. 1991. Neue Standard-Graptolithenzonenfolge nach der "Grossen Krise" an der Wenlock/Ludlow
Grenze (Silurs). Neues Jarhbuch fur Geologie und Palaontologie, Abhandlungen, 182, 303-354.
* JENKINS, C. J. 1982. Isograptus gibberulus (Nicholson) and the isograptids of the Arenig Series
(Ordovician) of England and Wales. Proceedings o f the Yorkshire Geological Society, 44, 219-248. (19)
JEPPSSON, L. 1990. An oceanic model for lithological and faunal changes tested on the Silurian record. Jornal
o f the Geological Society, London, 147, 663-674.
JONES, O. T. 1909. The Hartfell-Valentian succession in the district around Plynlimon and Pont Erwyd (North
Cardiganshire). Quarterly Journal o f the Geological Society of London, 65, 463-537.
JOHNSON, M. E., K A U O D. L. & RONG, J.-Y. 1991. Silurian eustasy. In: BASSETT, M. G., LANE, P. D.
& EDWARDS, D. (eds.) The Murchison Symposium, Special Papers in Palaeontology, 44, 145-163.
KALJO, D. & MARSS, T. 1991. Pattern of some Silurian bioevents. Historical Biology, 5, 145-152.
KALJO, D., BOUCOT, A. J., CORFIELD, R. M., LE HERISSE A., KOREN, T. N., KRIZ, J., MANNIK,
P., MARSS, T., NESTOR, V., SHAVER, R. H., SIVETER, D. J. & VIIRA, V. 1995. Silurian bio
events. In: WALLISER, O. H. (ed.) Global events and event stratigraphy in the Phanerozoic, 173-224.
Springer-Verlag, Berlin.
KIRK, N. 1969. Some thoughts on the ecology, mode of life, and evolution of the Graptolithina. Proceedings of
ref 5

the Geological Society o f London, 1659, 273-292.
KOREN, T. N., 1987. Graptolite dynamics in Silurian and Devonian time. Bulletin o f the Geological Society of
Denmark, 35, 149-159.
KOREN, T. N. 1988. Evolutionary crisis ofAshgill graptolites. In: WILLIAMS, S. H. & BARNES, C. R.
(eds.) Programs and Abstracts for Fifth International Symposium on the Ordovician System.
KOREN, T. N. 1991. The lundgreni extinction event in Central Asia and its bearing on graptolite biochronology
within the Homerian. Proceedings o f the Estonian Academy o f Science, Tallinn, 40, 74-78.
KOREN, T. N. & RICKARDS, R. B. 1979. Extinction of the graptolites. In: HARRIS, A. L., HOLLAND, C.
H. & LEAKE, B. E. (eds.) The Caledonides o f the British Isles-Reviewed. Geological Society of London,
457-466.
KOREN, T. N. & URBANEK, A. 1994. Adaptive radiation of monograptids after the late Wenlock crisis. Acta
Palaeontologica Polonica, 39, 137-167.
KRANDIEVSKY, V. S., ISHCHENKO, T. A. & KIRYANOV, V. V. 1968. Palaeontology and stratigraphy of
the Volhyno-Podolian Lower Palaeozoic. Naukova Dumka, Kiev, [in Russian], 1-124.
KUHNE, W. G., 1955. Unterludlow-Graptolithen aus Berliner Geschieben. Neues Jahrbuch fur Geologie und
Palaontologie, Abhandlungen, 100, 350-401.
LAPWORTH, C. 1873. On an improved classification of the Rhabdophora. Geological Magazine, 10, 500-504,
555-560.
LAPWORTH, C. 1878. The Moffat Series. Quarterly Journal o f the Geological Society, London, 34.
LAPWORTH, C. 1880. On new British graptoites. Annals and Magazine o f Natural History, London, 5, 149-
177.
LAPWORTH, C. 1880. The geological distribution of the Rhabdophora. Annals and Magazine of Natural
History, London, 6, 201.
LAPWORTH, H. 1900. The Silurian sequence of Rhayader. Quarterly Journal of the Geological Society,
London, 56, 67-137.
LEGGETT, J. K., McKERROW, W. S., COCKS, L. R. M. & RICKARDS, R. B. 1981. Periodicity in the
early Palaeozoic marine realm. Journal o f the Geological Society, London, 138, 167-176.
LENZ, A. C., 1972. Graptolites from the Laforce and St. Leon Formations, Northern Gaspe, Quebec. Canadian
Journal o f Earth Sciences, 9, 1148-1162.
LENZ, A. C. 1982. Ordovician to Devonian sea-level changes in western and northern Canada. Canadian Journal
ref 6

o f Earth Sciences, 1 9 , 1919-1932.
LENZ, A. C. 1992. Some late Wenlock and Ludlow monograptid graptolites, Cornwallis Island, Arctic Canada.
Canadian Journal o f Earth Sciences, 2 9 , 200-207.
* LOYDELL, D. K. 1991. The biostratigraphy and formational relationships of the upper Aeronian and lower
Telychian (Llandovery, Silurian) formations of western mid-Wales. Geological Journal, 2 6 , 209-244. (8)
LOYDELL, D. K. 1991. Dob's Linn - the type locality of the Telychian (Upper Llandovery) Rastrites maximus
Biozone? Newsletters in Stratigraphy, 2 5 , 155-161.
* LOYDELL, D. K. 1992-93. Upper Aeronian and Lower Telychian (Llandovery) graptolites from western
mid-Wales. The Palaeontographical Society, London. (10)
LOYDELL, D. K. 1998. Early Silurian sea-level changes. Geological Magazine, 135, 447-471.
* LOYDELL, D. K. & CAVE, R. 1993. The Telychian (Upper Llandovery) stratigraphy of Buttington Brick
Pit, Wales. Newsletters in Stratigraphy, 29, 91-103. (9)
* LOYDELL, D. K. & CAVE, R. 1996. The Llandovery-Wenlock boundary and related stratigraphy in eastern
mid-Wales with special reference to the Banwy River section. Newsletters in Stratigraphy, 3 4 , 39-64. (11)
MELCHIN, M. J. 1994. Graptolite extinction at the Llandovery-Wenlock boundary. Lethaia, 27, 285-290.
MELCHIN, M. J., KOREN, T. N. & STORCH, P. 1998. Global diversity and survivorship patterns of Silurian
graptoloids. In: LANDING, E. & JOHNSON, M. E. (eds.) Silurian cycles: linkages of dynamic
stratigraphy with atmospheric, oceanic and tectonic changes. New York State Museum Bulletin, 4 9 1 , 165-
182.
MIHAJLOVIC, M., 1974. Silurski graptoliti instocne Srbije (njihov stratigrafski polozaj). Bulletin du museum
d'histoire naturelle, Belgrade (A), 2 9 , 123-247.
* MOLYNEUX, S. G. & RUSHTON, A. W. A. 1988. The age of the Watch Hill Grits, English Lake District:
structural and palaeogeographical implications. Transactions of the Royal Society of Edinburgh, Earth
Sciences, 7 9 , 43-69. (22)
MUNCH, A., 1942. Die Graptolithen des unteren Ludlow von Ronneburg und Umgebung, Beitrage zur Geologie
von Thuringen. Jena.
NICHOLSON, H. A. 1868. On the graptolites of the Coniston Flags. Quarterly Journal of the Geological
Society, London, 2 4 , ?
* PACKHAM, G. H. 1962. Some Diplograptids from the British Lower Silurian. Palaeontology, 5 , 498-526.
(12)
ref 7

PALMER, D. C., 1971. The Lulow graptolites Neodiver so graptus nilssoni and Cucullograptus (Lobograptus)
progenitor. Lethaia, 4, 357-384.
PALMER, D. C., 1986. The monotypic 'population' accompanying the lectotype of Saetograptus varians (Wood
1900). In\ HUGHES, C. P., & RICKARDS, R. B. (eds.), Palaeoecology and Biostratigraphy of
Graptolites. Geological Society of London Special Publication, Blackwell Scientific Publications, 249-
259.
PERNER, J., 1899. Etudes sur les graptolites de Boheme. Paleont. Bohemiae, IIIB, Prague.
PRIBYL, A., 1942. Revision der Pristiograpten aus den Untergattungen Colonograptus n. subg. und
Saetograptus n. subg. , 52, 163-184.
PRIBYL, A. 1983. Graptolite biozones of the Kopanina and Pridoli Formations in the Upper Silurian of central
Bohemia. Cas. Mineral. Geol., 28, 149-167.
RICKARDS, R. B. 1967. The Wenlock and Ludlow succession in the Howgill Fells (north-west Yorkshire
and Westmorland. Quarterly Journal o f the Geological Society, London 123, 215-251.
RICKARDS, R. B. 1969. Wenlock graptolite zones in the English Lake District. Proceedings of the
Geological Society, London 1654, 61-65.
* RICKARDS, R. B. 1970. The Llandovery (Silurian) graptolites o f the Howgill Fells, Northern England.
The Palaeontographical Society, London. (23, 13)
RICKARDS, R. B. 1975. Palaeoecology of the Graptolithina, an extinct class of the Phylum Hemichordata.
Biological Reviews, 50, 397-436.
* RICKARDS, R. B. 1976. The sequence of Silurian graptolite zones in the British Isles. Geological Journal,
11, 153-188. (14)s
RICKARDS, R. B. 1995. Utility and precision of Silurian graptolite biozones. Lethaia, 28, 129-137.
RICKARDS, R. B., HUTT, J. E. & BERRY, W. B. N. 1976. Evolution of the Silurian and Devonian
graptoloids. Bulletin o f the British Museum (Natural History), 28, 1-120.
RICKARDS, R. B. & DUMICAN, L. W. 1984. The fibrillar component of the graptolite periderm. Irish
Journal o f Earth Sciences, 6, 175-203.
* RUSHTON, A. W. A. (pers. comm./unpublished). (25, 15)
RUSHTON, A. W. A. 1990. Ordovician graptolite biostratigraphy in the Welsh Basin: a review. Journal o f
the Geological Society, London, 147, 611-614.
RIVA, J. 1976. Climacograptus bicornis bicornis (Hall), its ancestor and likely descendents. In: BASSETT,
ref 8

M. G. (ed.) The Ordovician System: Proceedings o f a Palaeontological Association symposium,
Birmingham, September 1974. Cardiff.
ROSS, C. A., 1962. Silurian monograptids from Illinois. Palaeontology, 5, 59-72.
ROSS, C. A. & ROSS, R. P. 1996. Silurian sea-level fluctuations. In: WITZKE, B. J., LUDVIGSON, G. A.
& DAY, J. (eds.) Paleozoic sequence stratigraphy: views from the North American craton. Geological
Society of America Special Paper no. 306.
* RUSHTON, A. W. A. & MOLYNEUX, S. G. 1989. The biostratigraphic age of the Ordovician Skiddaw
Group in the Black Combe inlier, English Lake District. Proceedings of the Yorkshire Geological Society,
4 7 , 267-276. (24)
* RUSHTON, A. W. A., STONE, P., SMELLIE, J. L. & TUNNICLIFF, S. P. 1986. An early Arenig age for
the Pinbain sequence of the Ballantrae Complex. Scottish Journal o f Geology, 22, 41-54. (2)
* RUSHTON, A. W. A., TUNNICLIFF, S. P. & TRIPP, S. P. 1996. The faunas of the Albany Group in
the Girvan area, and their palaeogeographical implications. Scottish Journal of Geology, 3 2 , 23-32. (3)
RUSHTON, A. W. A. In: STONE, P. (ed.) The geology of the Skiddaw Group. Memoir of the British
Geological Survey (in preparation).
SHERGOLD, J. H. & SHIRLEY, J. 1968. The faunal-stratigraphy of the Ludlovian rocks between Craven
Arms and Bourton near Much Wenlock, Shropshire. Geological Journal, 6, 119-?
* SKEVINGTON, D. 1970. A Lower Llanvirn graptolite fauna from the Skiddaw Slates, Westmorland.
Proceedings o f the Yorkshire Geological Society, 3 7 , 395-444. (27)
* SKEVINGTON, D. 1970. The significance of the Hendre Shales graptolite fauna described by Toghill (1970).
Proceedings o f the Geological Society, London, 1 6 6 4 , 199-203. (26)
SPJELDN4BS, N. 1961. Ordovician climatic zones. Norsk geol. Tidsskr., 4 1 , 45-77.
SPJELDN/ES, N. 1978. Faunal provinces and the Proto-Atlantic. In: BOWES, D. R. & LEAKE, B. E. (eds.)
Crustal evolution in northwestern Britain and adjacent regions. Geological Journal Special Issue, 10, 139-
150.
* STONE, P. 1995. Geology of the Rhins of Galloway district. Memoir o f the British Geological Survey,
Sheets 1 and 3 (Scotland). (6)
* STONE, P. & RUSHTON A. W. A. 1983. Graptolite faunas from the Ballantrae ophiolite complex and
their structural implications. Scottish Journal o f Geology, 19, 297-310. (4)
* STONE, P. & STRACHAN, I. 1981. A fossiliferous borehole section within the Ballantrae ophiolite. Nature,
ref 9

293, 455-457. (5)
STORCH, P. 1994. Graptolite biostratigraphy of the Lower Silurian (Llandovery and Wenlock) of Bohemia.
Geological Journal, 29, 137-165.
STORCH, P. 1995. Biotic crises and post-crisis recoveries recorded by Silurian planktonic graptolite faunas of
the Barrandian area (Czech Republic). Geolines, 3, 59-70.
STORCH, P. & LOYDELL, D. K. 1996. The Hirnantian graptolites Normalograptus persculptus and
'Glyptograptus' bohemicus: strati graphical consequences of their synonomy. Palaeontology, 39, 869-
881.
STRACHAN, I. 1971. A synoptic supplement to "A monograph o f British Graptolites by Miss G. L. Elies and
Miss E. M. R. Wood. The Palaeontographical Society, London.
* STRACHAN, I. 1986. The Ordovician graptolites of the Shelve District, Shropshire. Bulletin o f the British
Museum (Natural History), 40, 1-58. (28)
* STRAHAN, A. & JONES, O. T. 1914. The geology of the South Wales Coalfield. Part XI. The country
around Haverfordwest. Memoir o f the Geological Survey of Great Britain, Sheet 228. (29)
TELLER, L. 1969. The Silurian biostratigraphy of Poland based on graptolites. Acta geologica Polonica, 19,
393-?
* TOGHILL, P. 1968. The graptolite assemblages and zones of the Birkhill Shales (lower Silurian) at Dobb's
Linn. Palaeontology, 11, 654-668. (30, 16)
* TOGHILL, P. 1970. A fauna from the Hendre Shales (Llandeilo) of the Mydrim area, Carmarthenshire.
Proceedings o f the Geological Society , London, 1663, 121-129. (31)
TOMCZYK, H., 1956. Wenlok i Ludlow w synklinie Kielckiej Gor Swietokrzyskich. Prace Instytut
geologiczny, 16, 1-77 [in Polish], 79-104 [in Russian], 105-129 [in English].
TULLBERG, S. A., 1883. Skanes Graptoliter II. Grapolifaunarna i Cardiolaskiffern och Cyrtograptusskiffern.
Sveriges Geologiska Undersokning, Afhandlingar, Ser. C, 55, 1-43.
URBANEK, A., 1958. Monograptidae from erratic boulders of Poland. Palaeontologia Polonica, 9, 1-105.
URBANEK, A. 1966. On the morphology and evolution of the Cucullograptus (Monograptidae, Graptolithina).
Acta geologica Polonica, 11, 291-?
URBANEK, A. 1970. Neocucullograptinae n. subfam. (Graptolithina) - their evolutionary and stratigraphic
bearing. Acta Palaeontologica Polonica, 15, 163-388.
URBANEK, A., 1993. Biotic crises in the history of Upper Silurian graptoloids: a palaeobiological model.
ref 10

Historical Biology, 7, 29-50.
VANDENBERG, A. H. M. & COOPER, R. A. 1992. The Ordovician graptolite sequence of Australasia.
Alcheringa, 16, 33-85.
* WADGE, A. J., M. J. C. NUTT & SKEVINGTON, D. 1972. Geology of the Tarn Moor Tunnel in the
Lake District. Bulletin o f the Geological Survey o f Great Britain, 41, 55-73. (32)
WALKER, M., 1953. The development of Monograptus dubius and Monograptus chimaera. Geological
Magazine, 90, 362-373.
WARREN, P. T. 1971. The sequence and correlation of graptolite faunas from the Wenlock-Ludlow rocks of
north Wales. Bureau Recherches Geologique et Minieres Memoir, 73, 451-460.
WATNEY, G. R., & WELCH, E. G., 1911. The zonal classification of the Salopian rocks of Cautley and
Ravenstonedale. Quarterly Journal of the Geological Society o f London, 67, 215-267.
WHITE, D. E., BARRON, H. F., BARNES, R. P. & LINTERN, B. C. 1992. Biostratigraphy of late
Llandovery (Telychian) and Wenlock turbiditic sequences in the SW Southern Uplands, Scotland.
Transactions o f the Royal Society o f Edinburgh, Earth Sciences, 82, 297-322.
WILDE, P., BERRY, W. B. N. & QUINBY-HUNT, M. S. 1991. Silurian oceanic and atmospheric circulation
and chemistry. In: BASSETT, M. G., LANE, P. D. & EDWARDS, D. (eds.) The Murchison Symposium,
Special Papers in Palaeontology, 44, 123-143.
* WILLIAMS, M. AND J. ZALASIEWICZ (in press). Graptolite biozonation of the Wenlock (Silurian) rocks
of the Builth Wells district, central Wales. Geological Magazine. (22)
* WILLIAMS, S. H. 1982a. Upper Ordovician graptolites from the top Lower Hartfell Shale Formation (D.
clingani and P. linearis zones) near Moffat, southern Scotland. Transactions o f the Royal Society of
Edinburgh: Earth Sciences, 72, 229-255. (7a)
* WILLIAMS, S. H. 1982b. The late Ordovician graptolite fauna of the Anceps Bands at Dob's Linn, southern
Scotland. Geologica et Palaeontologica, 16, 29-56. (7b)
WILLIAMS, S. H. 1983. The Ordovician-Silurian boundary graptolite fauna of Dob's Linn, southern Scotland.
Palaeontology, 26, 605-639.
WILLIAMS, S. H. 1986. Top Ordovician and lowest Silurian of Dob's Linn. In C. P. HUGHES &
RICKARDS, R. B. (eds.) Palaeoecology and Biostratigraphy of Graptolites. The Geological
Society/Blackwell, London, 20, 165-171.
* WILLIAMS, S. H. 1987. Upper Ordovician graptolites from the D. complanatus Zone of the Moffat and
ref 11

Girvan districts and their significance for correlation. Scottish Journal o f Geology, 23, 65-92. (8)
* WILLIAMS, S. H. 1988. Dob's Linn - the Ordovician-Silurian boundary stratotype. Bulletin o f the British
Museum o f Natural History (Geology), 4 3 , 17-30. (9 )
* WILLIAMS, S. H. 1994. Revision and definition of the C. wilsoni graptolite Zone (middle Ordovician) of
southern Scotland. Transactions o f the Royal Society o f Edinburgh: Earth Sciences, 8 5 , 143-157. (1 0 )
WOOD, E. M. R. 1900. The Lower Ludlow Formation and its Graptolite-Fauna. Quarterly Journal of the
Geological Society, London 5 6 , 415-492.
WOOD, E. M. R. 1904. The graptolites from the Lower Ludlow Shales. Proceedings of the Geologists'
Association, 1 8 , 446-451.
WOOD, E. M. R. 1906. The Tarannon Series of Tarannon. Quarterly Journal o f the Geological Society, London
6 2 , 644-701.
ZALASIEWICZ, J. A. 1984. A re-examination of the type Arenig Series. Geological Journal, 19, 105-124.
* ZALASIEWICZ, J. A. 1986. Graptolites from the type Arenig Series. Geological Magazine, 123, 537-544.
(33)
* ZALASIEWICZ, J. A. 1992. Graptolite dating of Ordovician vulcanicity in the Arenig area, North Wales.
Geological Journal, 2 7 , 379-389. (34)
* ZALASIEWICZ, J. A. 1992. Two new graptolites from the early Silurian (Llandovery: Aeronian) of central
Wales: an origin for monoclimacid thecal morphology. Geological Magazine, 1 2 6 , 779-785. (17)
* ZALASIEWICZ, J. A. 1994. Middle to late Telychian (Silurian: Llandovery) graptolite assemblages of central
Wales. Palaeontology, 37, 375-396. (19)
* ZALASIEWICZ, J. 1996. Aeronian (Silurian: Landovery) graptolites from central Wales. Geologica et
Palaeontologica, 30, 1-14. (21)
* ZALASIEWICZ, J. A., RUSHTON, A. W. A. & OWEN, A. W. 1995. Late Caradoc graptolitic faunal
gradients across the Iapetus Ocean. Geological Magazine, 1 3 2 , 611-617. (36, 11)
* ZALASIEWICZ, J. A., LOYDELL, D. K. & STORCH, P. 1995. A taxonomic revision of three mid-
Telychian Monoclimacids. Journal of Paleontology , 6 9 , 961-967. (20)
* ZALASIEWICZ, J. A. & TUNNICLIFF, S. 1994. Uppermost Ordovician to Lower Silurian graptolite
biostratigraphy of the Wye Valley, Central Wales. Palaeontology, 3 7 , 695-720. (55, 18)
References prefixed * are data sources used in the compilation of the graptoloid biostratigraphic range charts.
ref 12

Bracketed numbers at end of reference relate to the chart for which the reference was used;
Italics = Ordovician (England & Wales)
Bold = Ordovician (Scotland)
Plain = Silurian
ref 13

A PPEN D IX 1
BRITISH ORDOVICIAN AND SILURIAN
GRAPTOLOID SPECIES DIVERSITY PER BIOZONE

ENGLAND & WALES: ORDOVICIANBIOZONE TOTAL LOWER ZONE MIDDLE ZONE UPPER ZONE X BOUNDARY SUBZONE TOTAL LOWER MIDDLE UPPER X BOUNDARY
SPECIES SPECIES SPECIES SPECIES SPECIES SPECIES SUBZONE SUBZONE SUBZONE SPECIESSPECIES SPECIES SPECIES
murrayi 3 3 3 3 1phyllograptoides 8 8 8 8 5varicosus 19 12 11 18 13simulans 33 33 32 32 18victoriae 20 20 24 24 22gibberulus 38 37 37 38 23cucullus 41 37 37 41 17artus 48 40 43 40 15murchisoni 28 25 19 18 10teretiusculus 24 13 14 20 18gracilis 46 32 32 41 30multidens 50 39 38 40 16clingani 47 26 25 18 10 caudatus 28 25 21 23 18
morrisi 19 18 16 17 10linearis 11 11 7 5 1complanatus 5 5 5 5 5anceps 5 4 4 5 5persculptus 13 12 12 11
SCOTLAND: ORDOVICIAISfBIOZONE TOTAL LOWER ZONE MIDDLEZONE UPPER ZONE X BOUNDARY SUBZONE TOTAL LOWER MIDDLE UPPER X BOUNDARY
SPECIES SPECIES SPECIES SPECIES SPECIES SPECIES SUBZONE SUBZONE SUBZONE SPECIESSPECIES SPECIES SPECIES
Lancefieldian 3 2 2 2 2 2Bendigonian 1-2 7 7 7 7 0Chewtonian 1-2 10 10 10 10 0Castlemainian 3 12 12 12 12 0gracilis 44 44 42 42 37peltifer 42 42 42 42 16wilsoni 23 23 19 18 17clingani 88 46 85 42 30 caudatus 46 42 43 44 35
morrisi 42 41 41 40 30linearis 37 33 35 28 3complanatus 8 8 2 2 1anceps 29 10 27 14 12 complexus 12 10 10 12 12
pacificus 17 15 13 12 12extraordinarius 12 12 12 11 1persculptus 4 2 4 3 2

U.K: SILURIANBIOZONE TOTAL LOWER ZONE MIDDLEZONE UPPER ZONE X BOUNDARY SUBZONE TOTAL LOWER MIDDLE UPPER X BOUNDARY
SPECIES SPECIES SPECIES SPECIES SPECIES SPECIES SUBZONE SUBZONE SUBZONE SPECIESSPECIES SPECIES SPECIES
acuminatus 19 16 15 15 8atavus 30 20 19 23 15acinaces 36 32 29 33 29cyphus 54 41 46 46 30triangulatus 61 42 43 54 44magnus 54 51 49 46 35leptotheca 48 47 46 47 41convolutus 58 53 46 50 34sedgwickii 49 45 40 34 18halli 33 33 32 32 17
guerichi 58 33 38 35 20 runcinatus 33 32 31 32 25gemmatus 36 36 36 36 24renaudi 30 30 30 30 21
turriculatus 36 26 19 16 12 utilis 29 26 20 23 15johnsonae 16 16 16 16 15proteus 20 20 20 20 10camicus 16 16 16 16 12
crispus 20 15 12 12 6 galaensis 15 15 15 15 12crispus 13 13 13 13 7sartorius 12 12 10 10 6
griestoniensis 14 13 11 12 6crenulata 8 8 6 6 4spiralis 19 18 14 14 9lapworthi 17 14 16 7 8insectus 15 12 11 12 12centrifugus 24 22 23 22 13murchisoni 22 22 12 9 6firmus 6 6 5 5 5riccartonensis 15 10 10 12 5dubius 9 7 9 9 7rigidus 12 12 12 10 6lundgreni 18 14 13 16 13nassa 5 3 4 5 5ludensis 9 6 8 7 5nilssoni 27 11 23 20 14scanicus 18 14 18 14 9incipiens 10 10 8 6 3leintwardinensis 5 5 5 3 0Bohemograptus 1 1 0 0 0

A PPEN D IX 2
REPRESENTATIVE ILLUSTRATIONS OF
BRITISH SILURIAN GRAPTOLOID SPECIES


18
r$\ma
13
14
20

24






90




122





171
172
173
174
177
179
180
182


195
196198
199
201
200
203
197



232
231
230227
235
234
233




267

277279
282281 283
285

2 8 7


307

323318
319
320
322
324
325
326

\\
T\
\\ v
\\
330
\ 328
&i
&
332-V y
329 331

LV

345



3 67

APPENDIX 2 Magnifications and references
1. xlO (Elies & Wood, 1901-18).
2. xlO (Zalasiewicz & Tunnicliff, 1994)
3. xlO (Zalasiewicz & Tunnicliff, 1994)
4. x l5 (Rickards & Hutt, 1970)
5. xlO (Zalasiewicz & Tunnicliff, 1994)
6. xlO (Zalasiewicz & Tunnicliff, 1994)
7. x9 (Davies, 1929)
8. xlO (Zalasiewicz & Tunnicliff, 1994)
9. x9 (Davies, 1929)
10. xlO (Zalasiewicz & Tunnicliff, 1994)
11. x7.5
12. x5 (Rickards, 1988)
13. x7.5
14. xlO
15. xlO
16. x5
17. xlO
18. xlO
19. xlO
20. xlO
21. xlO (Rickards, 1970)
22. x5 (Hutt & Rick, 1970)
23. x7.5
24. xlO (Hutt, 1974-75)
25. x4 (Toghill, 1968)
26. xlO (Rickards, 1970)
27. xlO
28. x5
29. x5
30. x5
61. x7 (Toghill, 1968)
62. xlO (Hutt, 1974-75)
63. xlO (Zalasiewicz & Tunnicliff, 1994)
64. x7.5 (Hutt, 1974)
65. x2.5 (Sudbury, 1958)
66. x7.5 (Hutt, 1974-75)
67. x7.5 (Packham, 1962)
31. x7.5 (Hutt, 1974-75)
32. xlO
33. x5
34. x7.5 (Hutt, 1974-75)
35. xlO
36. x5
37. xlO (Elies & Wood, 1901-18).
38. xlO (Elies & Wood, 1901-18).
39. xlO (Zalasiewicz, 1996)
40. xlO (Zalasiewicz, 1996)
41. xlO (Rickards, 1970)
42. xlO (Zalasiewicz, 1996)
43. xlO(Zalasiewicz, unpublished)
44. xlO (Elies & Wood, 1901-18)
45. xlO (Hutt, 1974-75)
46. x7.5 (Rickards, 1970)
47. xlO (Elies & Wood, 1901-18)
48. x7.5 (Rickards, 1970)
49. xlO (Hutt, 1974-75)
50. xlO (Hutt, 1974-75)
51. xlO (Hutt, 1974-75)
52. xlO (Hutt, 1974-75)
53. xlO (Hutt, 1974-75)
54. x5 (Loydell, 1993-94)
55. x5 (Loydell, 1993-94)
56. x7.5 (Rickards, 1970)
57. xlO (Davies et al., 1997)
58. x9 (Rickards, 1967)
59. x7.5 (Hutt, 1974-75)
60.
91. xlO (Elies & Wood, 1901-18)
92. xlO (Hutt, 1974-75)
93. xlO
94. xlO
95. x7.5 (Hutt, 1974-75)
96. x5 (Atlas)
97. xlO (Elies & Wood, 1901-18)

68. xlO (Rickards, 1970)
69. xlO (Hutt, 1974-75)
70. xlO (Hutt, 1974-75)
71. x7.5 (Packham, 1962)
72. x7.5 (Rickards, 1970)
73. x7.5 (Packham, 1962)
74. x7.5 (Rickards, 1970)
75. x3 (Cocks & Toghill, 1973)
76. x7.5 (Packham, 1962)
77. x7.5 (Rickards, 1970)
78. x5 (Rickards, 1970)
79. x7.5 (Rickards, 1970)
80. xlO (Zalasiewicz, unpublished)
81. xlO (Hutt, 1974-75)
82. x5 (Loydell, 1993-94)
83. x l2.5 (Sudbury, 1958)
84. x l2 .5 (Sudbury, 1958)
85. xlO (Elies & Wood, 1901-18)
86 .
87. x7.5 (Hutt, 1974-75)
88. x7.5 (Rickards, 1970)
89. xlO (Elies & Wood, 1901-18)
90. xlO (Hutt, 1974-75)
121. x5 & xlO (Hutt, 1974-75)
122. xlO (Rickards, 1972)
123. x25 (Rickards, 1973)
124. x5 (Cocks & Toghill, 1973)
125. xlO (Hutt, 1974-75)
126. x 10 (Hutt, 1974-75)
127. xlO (Zalasiewicz, 1996)
128. x25 (Zalasiewicz, 1992)
129. xlO (Elies & Wood, 1901-18)
130. xlO (Hutt, 1974-75)
131. x 10 (Hutt, 1974-75)
132. xlO & x20 (Hutt, 1974-75)
133. xlO (Elies & Wood, 1901-18)
134. xlO (Hutt, 1974-75)
135. xlO (Elies & Wood, 1901-18)
136. xlO (Elies & Wood, 1901-18)
98. xlO (Elies & Wood, 1901-18)
99. x5 (Hutt, 1974-75)
100. x7.5 (Packham, 1962)
101. x 10 (Zalasiewicz & Tunnicliff, 1994)
102. x7.5 (Packham, 1962)
103. x 10 (Rickards & Koren, 1974)
104. x2.5 (Sudbury, 1958)
105. x 10 (Hutt, 1974-75)
106. x5 (Rickards, 1970)
107. xlO (Hutt, 1974-75)
108. xlO (Zalasiewicz & Tunnicliff, 1994)
109. xlO (Hutt, 1974-75)
110. x7.5 (Rickards, 1970)
111. x5 (Rickards, 1970)
112. Atlas (halved)
113. x5 (Loydell, 1993-94)
114. xlO (Rickards, 1970)
115. xlO (Zalasiewicz, 1992)
116. xlO (Hutt, 1974-75)
117. x7.5 (Rickards, 1970)
118. x7.5 (Rickards, 1970)
119. xlO (Hutt, 1974-75)
120. x20 (Hutt, 1974-75)
151. xlO (Elies & Wood, 1901-18)
152. xlO
153. x5 (Loydell, 1993-94)
154. x7.5 (Rickards, 1970)
155. xlO (Rickards, 1970)
156. xlO (Rickards, 1970)
157. xlO (Davies et al., 1997)
158. x5 (Rickards, 1970)
159. x5 (Loydell, 1993-94)
160. x7.5 (Rickards, 1970)
161. x5 (Atlas)
162. x5 (Rickards, 1970)
163. x30 (Rickards, 1970)
164. xlO (Rickards, 1970)
165.
166. x5 (Loydell, 1993-94)

137. x5 (Rickards, 1970)
138. xlO (Zalasiewicz, 1996)
139. xlO (Zalasiewicz, 1996)
140. xlO (Zalasiewicz, unpublished)
141. x 10 (Zalasiewicz, 1996)
142. xlO (Hutt, 1974-75)
143. x7.5 (Rickards, 1970)
144. x5 (Hutt, 1974-75)
145. xlO (Elies & Wood, 1901-18)
146. xlO
147. xlO
148. xlO
149. xlO (Davies et al., 1997)
150. xlO
181. x5 (Loydell, 1993-94)
182. x5 (Loydell, 1993-94)
183. x5 (Loydell, 1993-94)
184. x5 (Loydell, 1993-94)
185. x5 (Loydell, 1993-94)
186. x5 (Loydell, 1993-94)
187. x5 (Loydell, 1993-94)
188. x5 (Loydell, 1993-94)
189. x5 (Loydell, 1993-94)
190. x5 (Loydell, 1993-94)
191. x5 (Loydell, 1993-94)
192. x5 (Loydell, 1993-94)
193. x5 (Loydell, 1993-94)
194. xlO (Loydell, 1993-94)
195. x5 (Loydell, 1993-94)
196. x5 (Loydell, 1993-94)
197. x5 (Loydell, 1993-94)
198. x5 (Loydell, 1993-94)
199.
200.x5 (Rickards, 1970)
201. x5 (Loydell, 1993-94)
202. x5 (Loydell, 1993-94)
203. x5 (Loydell, 1993-94)
204. x5 (Loydell, 1993-94)
205. x5 (Loydell, 1993-94)
167. x4.7 (Boucek & Pribyl, 1941)
168. x5 (Loydell, 1993-94)
169. x5 (Loydell, 1993-94)
170. x5 (Loydell, 1993-94)
171. x5 (Loydell, 1993-94)
172. x5 (Loydell, 1993-94)
173. x5 (Loydell, 1993-94)
174. x5 (Loydell, 1993-94)
175. x5 (Loydell, 1993-94)
176. x5 (Loydell, 1993-94)
177. x5 (Loydell, 1993-94)
178. x5 (Rickards, 1970)
179. x5 (Rickards, 1970)
180. x5 (Loydell, 1993-94)
211. x5 (Loydell, 1993-94)
212. xlO (Hutt, 1974-75)
213. x5 (Loydell, 1993-94)
214. x7.5 (Rickards, 1970)
215. x5 (Loydell, 1993-94)
216. x5 (Loydell, 1993-94)
217. x5 (Loydell, 1993-94)
218. x5 (Loydell, 1993-94)
219. x5 (Loydell, 1993-94)
220. x5 (Loydell, 1993-94)
221. x5 (Loydell, 1993-94)
222. x5 (Loydell, 1993-94)
223. x5 (Loydell, 1993-94)
224. x5 (Loydell, 1993-94)
225. x5 (Loydell, 1993-94)
226. xlO (Hutt, 1974-75)
227. x 10 (Rickards, 1968)
228. xlO (Zalasiewicz, 1994)
229. xlO
230. xlO (Zalasiewicz, 1994)
231. xlO
232. xlO
233. xlO
234.x5
235. xlO

206. x5 (Hutt, 1974-75)
207. x5 (Loydell, 1993-94)
208. x5 (Loydell, 1993-94)
209. x5 (Loydell, 1993-94)
210. x5 (Loydell, 1993-94)
241. xlO
242. xlO (Zalasiewicz, 1995)
243. xlO
244. xlO (Zalasiewicz, 1995)
245. xlO
246. xlO
247. xlO
248. xlO
249. xlO (Zalasiewicz, 1994)
250. x5 (Zalasiewicz, 1995)
251. x5 (Williams & Zalasiewicz, in press)
252. x5 (Loydell & Cave, 1996)
253. x5 (Loydell & Cave, 1996)
254. x5 (Loydell & Cave, 1996)
255. x5 (Loydell & Cave, 1996)
256.
257. x5 (Loydell & Cave, 1996)
258. xlO (Zalasiewicz, 1995)
259. x5 (Loydell & Cave, 1996)
260. x5 (Loydell & Cave, 1996)
261. x5 (Loydell & Cave, 1996)
262. x5 (Loydell & Cave, 1996)
263. x5 (Loydell & Cave, 1996)
264. xlO (Elies & Wood, 1901-18)
265. x5 (Loydell & Cave, 1996)
266. x5 (Loydell & Cave, 1996)
267. x5 (Loydell & Cave, 1996)
268. x5 (Loydell & Cave, 1996)
269. x5 (Loydell & Cave, 1996)
270. x5 (Loydell & Cave, 1996)
301. x5 (Loydell & Cave, 1996)
302. x5 (Williams & Zalasiewicz, in press)
303. x5 (Loydell & Cave, 1996)
304. x5 (Loydell & Cave, 1996)
236. xlO (Elies & Wood, 1901-18)
237. xlO
238. x20
239. x20
240. xlO
271. x5 (Loydell & Cave, 1996)
272.
273. x5 (Williams & Zalasiewicz, in press)
274. x5 (Williams & Zalasiewicz, in press)
275. xlO (Loydell & Cave, 1996)
276. x5 (Loydell & Cave, 1996)
277. x5 (Loydell & Cave, 1996)
278. x5 (Loydell & Cave, 1996)
279. x5 (Loydell & Cave, 1996)
280. xlO (Loydell & Cave, 1996)
281. x5 (Loydell & Cave, 1996)
282. x5 (Williams & Zalasiewicz, in press)
283. x5 (Loydell & Cave, 1996)
284. x5 (Loydell & Cave, 1996)
285. x5 (Loydell & Cave, 1996)
286. x5 (R ickards, 1965)
287. x5 (Loydell & Cave, 1996)
288. x5 (Loydell & Cave, 1996)
289.
290.
291. x5 (Williams & Zalasiewicz, in press)
292. x5 (Loydell & Cave, 1996)
293. x5 (Loydell & Cave, 1996)
294. x5 (Loydell & Cave, 1996)
295. x5 (Loydell & Cave, 1996)
296. x5 (Loydell & Cave, 1996)
297. x5 (Loydell & Cave, 1996)
298. x5 (Loydell & Cave, 1996)
299. x5 (Loydell & Cave, 1996)
300. x5 (Loydell & Cave, 1996)
331. x5 (Williams & Zalasiewicz, in press)
332. x l5 (Holland et al., 1969)
333.
334. xlO (Elies & Wood, 1901-18)

305. x5 (Williams & Zalasiewicz, in press)
306. x5 (Williams & Zalasiewicz, in press)
307. Atlas (halved)
308. x5 (Rickards, 1965)
309. x5 (Williams & Zalasiewicz, in press)
310. x5 (Williams & Zalasiewicz, in press)
311.
312. x4 (Storch, 1994)
313. x5 (Rickards, 1965)
314. x5 (Williams & Zalasiewicz, in press)
315. x5 (Williams & Zalasiewicz, in press)
316. x5 (Davies et al., 1997)
317. x5 (Williams & Zalasiewicz, in press)
318. x5 (Williams & Zalasiewicz, in press)
319. x5 (Williams & Zalasiewicz, in press)
320. xlO (Davies et al., 1997)
321. x5 (Davies et al., 1997)
322. x5 (Williams & Zalasiewicz, in press)
323. x5 (Williams & Zalasiewicz, in press)
324. x5 (Williams & Zalasiewicz, in press)
325. xlO (Rickards, 1965)
326. x5 (Williams & Zalasiewicz, in press)
327. x0.75 (Boucek, 1933)
328. (Manck, 1918)
329. x4 (Storch, 1994)
330. x4.5 (Boucek, 1933)
361. x 10 (Taylor, this study)
362. xlO (Taylor, this study)
363. x5 (Rickards 1965)
364. (Taylor, this study)
365. x5 (Atlas)
366. x5 (Boucek, 1936)
367.
368. x5 (Boucek, 1936)
369. (Taylor, this study)
370.
335. xlO (Holland et al., 1969)
336. x5 (Williams & Zalasiewicz, in press)
337. xlO (Holland et al., 1969)
338. x5 (Rickards, 1965)
339. x4 (Storch, 1994)
340. x5 (Atlas)
341. xlO (Elies & Wood, 1901-18)
342. x6 (Palmer, 1971)
343. x 10 (Elies & Wood, 1901-18)
344. (Taylor, this study, halved)
345. x 10 (Hutt, 1969)
346. x5 (Boucek, 1936)
347.
348. xlO (Elies & Wood, 1901-18)
349. xlO (Taylor, this study)
350. (Taylor, this study)
351. x6 (Palmer, 1971)
352. (Taylor, this study)
353. x27 (Urbanek, 1966)
354. xlO (Elies & Wood, 1901-18)
355. xlO (Atlas)
356. xlO (Elies & Wood, 1901-18)
357. x l 8 (Urbanek, 1958)
358. x5 (Boucek, 1936)
359. x5 (Rickards, 1965)
360. x5 (Atlas)