Embed Size (px)
Citation preview

[CANCER RESEARCH 48, 1717-1721, April 1, 1988]
Pathobiological Effects of Acrolein in Cultured Human Bronchial Epithelial Cells1
Roland C. Grafström,2Jeannette M. Dypbukt, James C. Willey, Kristina Sundqvist, Charlotte Edman, Luigi Atzori,
and Curtis C. HarrisDepartment of Toxicology, Karolinska Institutet, S-104 01 Stockholm, Sweden fR. C. G., J. M. D., K. S., C. E., L. A.] and Laboratory of Human Carcinogenesis,National Cancer Institute, Bethesda, Maryland 20892 [J, C. W., C. C. H.]
ABSTRACT
The ability of the highly reactive aldehyde acrolein to affect growth,membrane integrity, differentiation, and thiol status and to cause DNAdamage has been studied at serum- and third-free conditions using cultured human bronchial epithelial cells. Acrolein markedly decreasescolony survival at 3 ¿JMwhereas about 10-fold higher concentrations arerequired to increase membrane permeability, measured as uptake oftrypan blue dye. Acrolein at micromolar concentrations also causesepithelial cells to undergo squamous differentiation as indicated by decreased donai growth rate, dose-dependent increased formation of cross-linked envelopes, and increased cell planar surface area. Acrolein causesa marked and dose-dependent cellular depletion of total and specific freelow-molecular-weight thiols as well as protein thiols. Exposure to acroleindid not cause oxidation of glutathione indicating that thiol depletionoccurred by direct conjugation of reduced glutathione to acrolein withoutconcomitant generation of active oxygen species. Furthermore, acroleinis genotoxic and causes both DNA single strand breaks and DNA proteincross-links in human bronchial epithelial cells. The results indicate thatacrolein causes several cytopathic effects that relate to multistage carci-
nogenesis in the human bronchial epithelium.
INTRODUCTION
The environmental occurrence, metabolism, short- and long-term effects in vitro and in vivo of acrolein (CH2 = CHCHO),have been extensively reviewed (1-3). Acrolein is present inboth tobacco smoke and automotive emissions. Acrolein isbelieved to be a decomposition product of the cellular polya-mine, spermine (4), and is a major metabolite of the widelyused chemotherapeutic drug, cyclophosphamide (5-10). Inhalation of acrolein inhibits mucociliary transport and causesdysfunction, hyperplasia, and squamous metaplasia in the respiratory tract of laboratory animals (1-3, 11).
Acrolein is highly cytotoxic toward bacteria and mammaliancells (1-3). The marked toxic potency of acrolein may in partdepend on its bifunctional high reactivity toward cellular nu-cleophiles, primarily sulfhydryls, which bind through thiol etherformation at the terminal ethylenic carbon atom (12). Subsequently, the aldehydic carbon may react with other nucleo-philes. Thus, exposure to acrolein causes depletion of the majorcellular thiol, GSH,3 in freshly isolated hepatocyte suspensions
(13, 14), cultured human leukemia cells (15), as well as in the
Received 8/7/87; revised 12/15/87; accepted 12/30/87.The costs of publication of this article were defrayed in part by the payment
of page charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
1Supported in part by grants from the Health Effects Institute, the Workers
Environment Health Fund, the Swedish National Board of Laboratory Animals,the Swedish Medical Research Council, the Swedish Cancer Society, and theSwedish Tobacco Company (R. C. G., J. M. D., K. S., C. E., and L. A.).
Research described in this article is conducted in part under contract to theHealth Effects Institute, an organization jointly funded by the United StatesEnvironmental Protection Agency (Assistance Agreement X-812059) and automotive manufacturers. The contents of this article do not necessarily reflect theviews of the Health Effects Institute, nor do they necessarily reflect the policiesof the Environmental Protection Agency or automotive manufacturers.
2To whom requests for reprints should be addressed, at Department ofToxicology, Tomtebodavâgen 30, Karolinska Institutet, S-104 01, Stockholm,Sweden.
3The abbreviations used are: GSH, reduced glutathione; LHC, Laboratory ofHuman Carcinogenesis; SDS, sodium dodecyl sulfate; GSSG, oxidized glutathi-
nasal mucosa of acrolein-exposed rats (16,17). Decreased levelsof GSH may also depend on oxidation of GSH to GSSG whichmay occur as a result of the generation of reactive oxygenspecies (18-20); such effects by acrolein have recently beenimplicated in allyl-alcohol liver toxicity, in which acrolein isbelieved to be a causative metabolite (21, 22).
In part due to its extreme toxicity, the information on thegenetic effects and the cardnogcnicity of acrolein is conflictingand limited (1-3). Incubation of DNA with a bifunctionalaldehyde such as acrolein causes formation of cyclic 1,N2-
propanodeoxyguanosine adducts (23). As measured by alkalinesucrose gradient centrifugation, acrolein causes no detectableDNA damage in yeast (8) and furthermore, no DNA proteincross-linking can be detected in the nasal mucosa of rats exposed to 2 ppm of acrolein for 6 h (16, 17). However, DNAsingle strand breaks are formed when mouse (24) or humanleukemia cells (25) are exposed to acrolein. In the Ames Salmonella assay for mutagenicity, both positive and negativeresults have been reported (7, 26, 27). Albeit at high cytotox-icity, both sister chromatid exchanges and chromosomal aberrations occur in Chinese hamster ovary cells (10). Acrolein hasbeen found not to be carcinogenic in Syrian Golden hamstersexposed to 4 ppm for 1 yr (28).
Because of the potential for human exposure and adversehealth effects in the respiratory airways, we have conducted aseries of studies on the effects of acrolein in cultured humancells (29)." In human bronchial fibroblasts, we have previouslyfound that acrolein and «,/3-unsaturated aldehydes formed bylipid peroxidation are highly cytotoxic, decrease total cellularthiols, and inhibit O6-alkylguanine-DNA alkyltransferase,which has an acrolein-sensitive thiol residue required for itscatalytic activity (29). Furthermore, we have recently found thatacrolein is mutagenic to human cells.4 To investigate if acrolein
causes effects of importance for initiation and promotion ofCarcinogenesis, we have now studied pathobiological effects ofacrolein in presumptive inhalation target cells, i.e., epithelialcells cultured from normal human bronchus (30). Cellular parameters investigated after exposure to acrolein include colonysurvival, membrane integrity, growth rate, total and specificfree thiols, protein sulfhydryl status, biochemical and morphological differentiation markers, and different types of DNAdamage.
MATERIALS AND METHODS
Cells and Cell Culture Conditions. Normal human tracheobronchialtissues were obtained from about 40 different donors at the time ofautopsy or surgery and maintained in culture according to methodsdescribed by Lechner et al. (30). Primary epithelial cell cultures wereenzymatically dissociated with calcium-free 4-(2-hydroxyethyl)-l-piper-azineethanesulfonic acid-buffered saline solution containing [ethylene-bis(oxyethylenenitrilo)]-tetraacetic acid, 0.02%; polyvinylpyrrolidone,1%; and trypsin, 0.02%, and subcultured into surface-treated (fibronec-
4Curren, R. D., Yang, L. L., Conklin, P. M., Grafström, R. C., and Harris,
C. C. Mutagenesis of Xeroderma pigmentosum fibroblasts by acrolein. MutationResearch Letters, in press, 1988.
1717
on March 16, 2020. © 1988 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

EFFECTS OF ACROLEIN IN HUMAN BRONCHIAL CELLS
tin, 10 fig/ml; type I rat tail collagen, 30 /ig/ml) IViri dishes containingLHC-8, a serum-free medium (30).
Colony-Forming Efficiency and C.'limai Growth Assays. Cells wereseeded at 250 cells/cm2 in 60-mm dishes. After 24 h in culture, cells
were exposed for indicated time periods to the various doses of acroleinin serum-free LHC basal medium (30) without thiols, i.e., cysteine.Subsequently, fresh aldehyde-free LHC-8 medium was added and after7 to 9 days of incubation, the cultures were fixed and stained. Themean colony-forming efficiency was determined from duplicate dishesand based on colonies each containing at least 16 cells. The clonalgrowth rate was determined using an IBAS-2 image analyzer (Zeiss,Stockholm, Sweden). To obtain the mean clonal growth rate (expressedas population doublings per day), the number of cells in 9 randomlyselected clones from each of 2 dishes were counted and the log? of theaverage number of cells per clone was divided by the number of days ofincubation. Variance-weighted Student's t test was used to evaluate
significant differences between control and treated groups.Trypan Blue Exclusion. Cells were inoculated at 2800 cells/cm2 in
35-mm dishes. After 24 h, cells were exposed for l h to variousconcentrations of acrolein in thiol-free LHC basal medium. Subsequently, cells were washed twice with LHC basal medium and then0.16% trypan blue dye (Sigma, St. Louis, MO) dissolved in LHC basalmedium was added for 5 min. The dye was then removed and replacedwith fresh LHC-8 medium and the percentage of viable (cells thatexclude trypan blue) and nonviable (cells that appear entirely blue orhave a blue nuclei) cells counted in four randomly selected fields perdish in a phase-contrast microscope. Percentage of viability was calculated from duplicate dishes as 100 times the ratio of number of viablecells to the sum of viable and nonviable cells. More than 95% ofunexposed control cells excluded trypan blue.
Formation of Cross-Linked Envelopes. The percentage of cells capableof forming cross-linked envelopes was measured as described by Willeyet al. (31). Cells were inoculated onto 12 well plates at 11,000 cells/cm2 in LHC-8 medium. Twenty-four h later, the number of cells per
well was calculated using a grid and the medium was replaced withmedia containing acrolein plus 0.8% agar. After a 3-h incubation, 2 mlof SDS (4%) and dithiothreitol (20 HIM)in distilled H2O were addedover the agar. After a further 4-h incubation at 37°C,the number of
SOS-insoluble, noncalcium ionophore-induced (32), cross-linked envelopes per well and percentage of cross-linked envelopes in the cellpopulation were then calculated.
Plasminoceli Activator Assay. Plasminogen activator activity wasdetermined as previously described by Willey et al. (31). Cells wereinoculated onto 24-well plates at 50,000 cells/cm2 in LHC-8 medium.
After 2 h of incubation with acrolein, 25 n\ of plasminogen (Sigma)(final concentration 0.1-0.3 unit/ml) was added and incubation wascontinued for 1 more h (a total of 3 h of incubation with acrolein).Media were removed and centrifugea in an Eppendorf microfuge for30 sec to pellet cell debris, and 90 n\ of medium were incubated with10 /¿Iof benzyloxycarbonylglycyl-L-prolyl-L-arginyl['4C]anilide (5 HIM,12.5 mCi/mmol; NEN, Boston, MA) for l h at 37°C.Proteolyticallycleaved [l4C]anilide was then extracted from the reaction mixtures with
3 x 2 ml of Econofluor II (New England Nuclear, Boston, MA) andradioactivity assayed (31).
Cell Planar Surface Area. Cells were inoculated onto 24-well platesat 5000 cells/cm2 in LHC-8 medium. Twenty-four h later, the cellswere exposed to the various concentrations of acrolein for 3 h in thiol-free LHC basal medium. For each concentration, a randomly selectedarea containing at least 30 cells was followed by phase-contrast microscopy (Olympus IM, Olympus, Stockholm), and photographed beforeand after exposure to acrolein. Change in planar cell surface area wasquantitated for each cell by the weight of print cuttings.
Assay for Free Total and Specific Nonprotein as well as ProteinThiols. The cells were inoculated onto 35-mm dishes at 6-12 x IO3/cm2 in LHC-8 medium. After 48 to 72 h, cells were exposed to acroleinfor 1 h in thiol-free LHC basal medium. After washing twice withphosphate-buffered saline, the cellular thiol status was measured byvarious methods. To measure the amounts of total free thiols, cellswere lysed and protein precipitated by addition of 0.7 ml 6.5% trichlor-acetic acid to the dish. The cellular precipitate was removed from the
dish by scraping with a rubber policeman. The resulting suspensioncontaining denatured protein and released cellular thiols was centri-fuged at 1500 x g for 8 min and 0.5 ml supernatant analyzed for thiolcontent according to Saville (33). Monobromobimane conjugation ofthiols by the method of Cotgreave and Moldéus(34) was used toanalyze the amounts of specific free low-molecular-weight thiols. Reduced cellular thiols were derivati/ed by addition of 100 /¿I8 HIMmonobromobimane (Thiolyte; Calbiochem, La .lolla, CA) in 25 mM N-ethylmorpholine buffer, pH 8.0, and 100 n\ phosphate-buffered salineonto the dish. Subsequently, the dish was sealed and allowed to stand5 min in the dark before acidification and precipitation of protein byaddition of 10 ^1 of 90% trichloroacetic acid. To analyze for thepresence of oxidized thiols a similar sample was initially treated with 5HIMdithiothreitol for 30 min, then derivati/ed with 20 mM monobromobimane, and subsequently acidified and precipitated as above. Thecellular precipitate was removed by pipetting following scraping of thedishes with a rubber policeman. The resulting suspension containingprotein and released cellular thiols were centrifuged at 3000 x g for 3min and the supernatants were analyzed for levels of GSH and totalGSH, mainly GSH plus hydrolyzed GSSG, by high-performance liquidchromatography separation of the GSH-bimane adducts (34). An estimate of the cellular content of GSSG was then calculated by subtractingthe amounts of GSH from total GSH. The derivai i/ed protein pelletswere washed 3 times with 3 ml of 5% trichloroacetic acid to removecontaminating free low-molecular-weight thiol-bimane adducts. Thewashed pellet was then dissolved in 1% SDS, protein content determined, and the solubilized protein sample was then diluted up to 50 mlwith distilled water and the fluorescence determined and standardizedagainst bovine serum albumin (34).
Alkaline Elution Assay of DNA Damage. The procedure used wasdeveloped and reviewed by Kohn et al. (35). The cells were inoculatedonto 60-mm dishes at 3-5 x 103/cm2 in LHC-8 medium. After 24 h,[2-14C]thymidine (0.02 ¿iCi/ml;52 mCi/mmol) was added for 3 daysand then removed 1-2 days prior to the experiment. Cells were exposedto acrolein for 1 h in LHC basal medium. Subsequently, the cells wereremoved from the dishes with a rubber policeman at 4"( ' in a solutionof Dulbecco's Ca2+/Mg2+-free phosphate-buffered saline containing 15
mM disodium EDTA and collected onto a 2-/¿mpolycarbonate filter(Nucleopore, Pleasanton, CA) to measure DNA single strand breaks.The cells were lysed with 5 ml of 2% SDS/0.1 M glycine/0.02 Mdisodium EDTA (pH 9.6), and 2 ml of the same solution containing0.5 mg/ml of proteinase K was then pumped through the filter at 0.04ml/min. This solution was followed by 0.02 M EDTA (acid form)/0.1%SDS plus tetrapropylammonium hydroxide (pH 12.2). Eluted fractionswere collected and assayed for radioactivity. The combination of thepolycarbonate filtration, proteinase K digestion, and the addition ofSDS in the eluting solution removed DNA protein cross-links causedby acrolein. An internal standard, [3H]thymidine-labeled 1.1210 cellsthat had received 300 rad at 4"C. was included in each assay. To
measure the extent of DNA protein cross-links, the assay was modifiedby omission of the proteinase digestion of the cell lysates and SDS wasdeleted from the eluting solution. The cells were exposed to 4 krad ofionizing radiation (to shear cellular DNA) before these cells werecollected on polyvinyl chloride filters (Millipore Corp., Bedford, MA).The frequencies of DNA single strand breaks and DNA protein crosslinks were calculated and plotted as described by Kohn et al. (35).
Source of Test Agent. Acrolein (99+, gold label) was obtained fromAldrich Chemical Company (Milwaukee, WI) or Aldrich-Chemie(Steinheim/Albuch, Federal Republic of Germany).
RESULTS
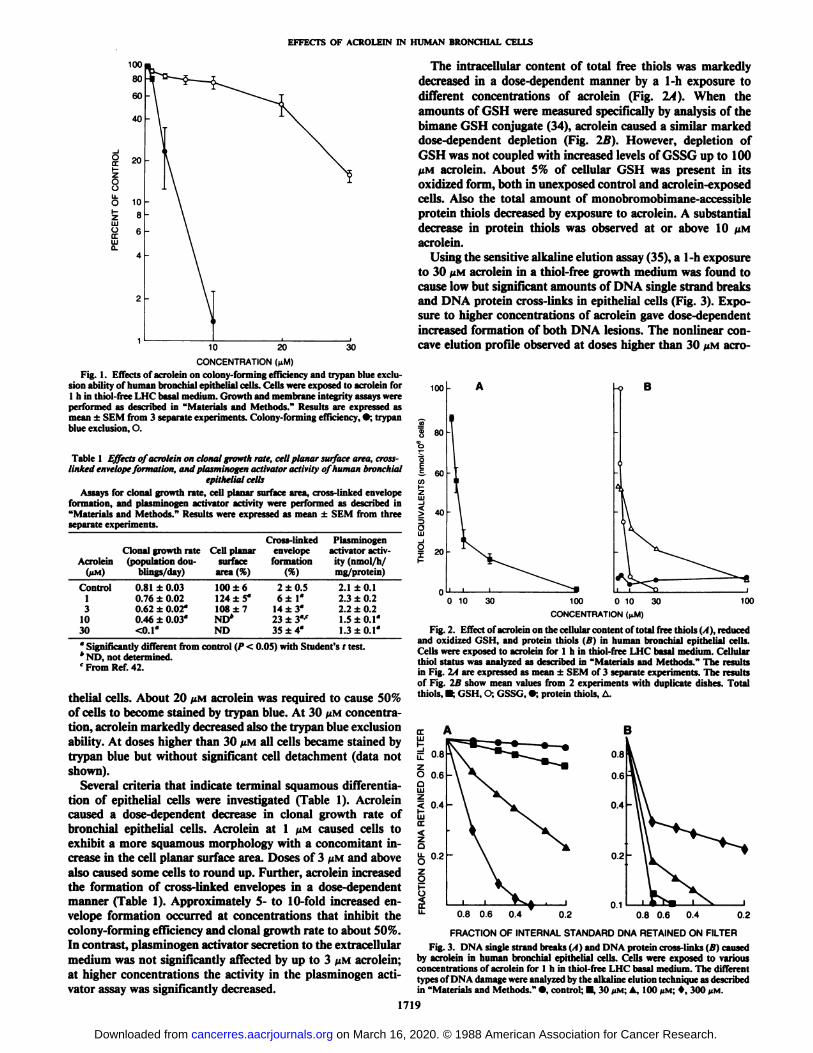
The toxicity of acrolein at defined thiol-free conditions wasinvestigated in human bronchial epithelial cells by determiningsurvival as measured by colony-forming efficiency or trypanblue exclusion (Fig. 1). A 1-h exposure to less than 3 MMacroleinwas required to decrease the colony-forming efficiency to 50%.Measurement of trypan blue exclusion, as a survival assay,could be successfully applied to cultured human bronchial epi-
1718
on March 16, 2020. © 1988 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from
EFFECTS OF ACROLEIN IN HUMAN BRONCHIAL CELLS
100
80
60
40
OerK
8
zmoce
20
2 - \10
20 30
CONCENTRATION(uM)Fig. 1. Effects of acrolein on colony-forming efficiency and trypan blue exclu
sion ability of human bronchial epithelial cells. Cells were exposed to acrolein for1 h in thiol-free LHC basal medium. Growth and membrane integrity assays wereperformed as described in "Materials and Methods." Results are expressed asmean ±SEM from 3 separate experiments. Colony-forming efficiency, •;trypanblue exclusion,O.Table
1 Effects of acrolein on clonal growth rate, cell planar surface area, cross-linked envelope formation, and plasminogen activator activity of human bronchial
epithelial cellsAssays for clonal growth rate, cell planar surface area, cross-linked envelope
formation, and plasminogen activator activity were performed as described in"Materials and Methods." Results were expressed as mean ±SEM from three
separate experiments.Cross-linked
PlasminogenClonal growth rate Cell planar envelope activator activ-
Acrolein (population dou- surface formation ity inumi li{/IM) blings/day) area (%) (%)mg/protein)Control
0.81 ±0.031 0.76 ±0.023 0.62 ±0.02"
10 0.46 ±0.03°30 <0.1°100
±6 2 ±0.5 2.1 ±0.1124 ±5" 6±1° 2.3±0.2108 ±7 14 ±3" 2.2 ±0.2ND* 23±3°'c 1.5±0.rND 35 ±4° 1.3 ±0.1°THIOL
EQUIVALENTS(nmol/106cells)25'cM "A"o8è8§8"3?^)NA protein cross-linkso higher concentrationsised formation of both E•lutionprofile observediA
I0
10 30 10CONCf
2. Effect of acrolein on the cellu
The intracellular content of total free thiols was markedlydecreased in a dose-dependent manner by a 1-h exposure to
different concentrations of acrolein (Fig. 2A). When theamounts of GSH were measured specifically by analysis of thebimane GSH conjugate (34), acrolein caused a similar markeddose-dependent depletion (Fig. 2B). However, depletion of
GSH was not coupled with increased levels of GSSG up to 100/IM acrolein. About 5% of cellular GSH was present in itsoxidized form, both in unexposed control and acrolein-exposedcells. Also the total amount of monobromobimane-accessibleprotein thiols decreased by exposure to acrolein. A substantialdecrease in protein thiols was observed at or above 10 UMacrolein.
Using the sensitive alkaline elution assay (35), a 1-h exposureto 30 UMacrolein in a thiol-free growth medium was found tocause low but significant amounts of DNA single strand breaks
B
30 100
°Significantly different from control (P < 0.05) with Student's / test.* ND, not determined.' From Ref. 42.
thelial cells. About 20 /IM acrolein was required to cause 50%of cells to become stained by trypan blue. At 30 /tM concentration, acrolein markedly decreased also the trypan blue exclusionability. At doses higher than 30 /UMall cells became stained bytrypan blue but without significant cell detachment (data notshown).
Several criteria that indicate terminal squamous differentiation of epithelial cells were investigated (Table 1). Acroleincaused a dose-dependent decrease in clonal growth rate ofbronchial epithelial cells. Acrolein at 1 /¿Mcaused cells toexhibit a more squamous morphology with a concomitant increase in the cell planar surface area. Doses of 3 /JMand abovealso caused some cells to round up. Further, acrolein increasedthe formation of cross-linked envelopes in a dose-dependentmanner (Table 1). Approximately 5- to 10-fold increased envelope formation occurred at concentrations that inhibit thecolony-forming efficiency and clonal growth rate to about 50%.In contrast, plasminogen activator secretion to the extracellularmedium was not significantly affected by up to 3 ^M acrolein;at higher concentrations the activity in the plasminogen activator assay was significantly decreased.
o 10.TION ((¿M)
ntent of total free thiols 1.1). reducedand oxidized GSH, and protein thiols (B) in human bronchial epithelial cells.Cells were exposed to acrolein for l h in thiol-free LHC basal medium. Cellularthiol status was analyzed as described in "Materials and Methods." The results
in Fig. 2A are expressed as mean ±SEM of 3 separate experiments. The resultsof Fig. 2B show mean values from 2 experiments with duplicate dishes. Totalthiols, •:GSH, O; GSSG, •;protein thiols, A.
0.6 0.4 0.2
FRACTION OF INTERNAL STANDARD DNA RETAINED ON FILTERFig. 3. DNA single strand breaks (A) and DNA protein cross-links (B) caused
by acrolein in human bronchial epithelial cells. Cells were exposed to variousconcentrations of acrolein for l h in thiol-free LHC basal medium. The differenttypes of DNA damage were analyzed by the alkaline elution technique as describedin "Materials and Methods." •,control; •30 MM;A, 100 JIM:*, 300 MM.
1719
on March 16, 2020. © 1988 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

EFFECTS OF ACROLEIN IN HUMAN BRONCHIAL CELLS
lein in the DNA single strand break assay also indicated someformation of DNA interstrand cross-links.
DISCUSSION
Acrolein significantly decreases the colony-forming efficiencyof cultured human bronchial epithelial cells already at a concentration of l /¿M.When the ability of cells to exclude trypanblue was measured, about 10- to 20-fold higher concentrationsof acrolein were required to cause membrane permeability.Thus, comparison shows that colony-forming efficiency is amore sensitive biological endpoint, although the presence ofsubstantially increased numbers of acrolein-reactive thiol targetgroups in the higher density cultures used for the trypan blueexclusion assay may contribute to its lower sensitivity. Tominimize similar interactions, serum- and thiol-free exposureconditions were used in this study because their presence in theculture medium may shift the reactivity of a bifunctional aldehyde like acrolein by unevenly scavenging effects related to themore reactive /3-carbon as compared to the aldehyde group. Forexample, recent investigations on the embryotoxicity in vitro ofacrolein support different mechanisms of action in serum-freeand serum-containing media, respectively (36).
In mammalian cells, conjugation with GSH constitutes amajor cellular defense mechanism against toxic exogenous compounds as well as many electrophilic products of metabolicactivation. Human bronchial epithelial cells contain approximately 100 nmol of total free thiols per IO6 cells and GSH
constitutes about 80% of this total amount. Acrolein is one ofthe most thiol-reactive agents known (12). We have previouslyshown that acrolein causes depletion of total free thiols infibroblasts (29). When the cellular levels of total free thiols aswell as GSH are specifically assayed in bronchial epithelialcells, acrolein causes a marked and similar depletion of bothtotal thiols and GSH at 3 /UMconcentration. When the effectof acrolein on the total amounts of free cellular protein thiolsis measured, a significant decrease is not observed below 10 fiM,indicating the protective action of GSH against acrolein alky I-ation of protein thiols. Furthermore, acrolein-induced growthinhibition occurs before significant GSH depletion, whereasloss of membrane integrity follows this effect. Acrolein is likelyto interact with specific cellular protein thiols during growthinhibition because cytotoxic concentrations are required to decrease the total amounts of protein thiols.
Exposure of bronchial epithelial cells to acrolein causedenhanced terminal squamous differentiation as indicated bydecreased clonal growth rate, increased cell surface area, andseveral-fold increased formation of cross-linked envelopes, thelatter regarded as a late event in terminal squamous differentiation (31, 32, 37). With tobacco smoke or acrolein alone asimilar response is observed in vivo, since inhalation exposureof rodents to either of these agents causes squamous metaplasiain their respiratory airways (see Ref. 3). Recent studies withcigarette smoke condensate fractions indicate that nonvolatilecomponents of smoke also can induce differentiation of normalhuman bronchial epithelial cells (38). Agents with tumor-promoting activity in the mouse skin carcinogenesis model such asaplysiatoxin, 12-O-tetradecanoylphorbol-13-acetate, or teleo-cidin B are the most potent inducers of terminal squamousdifferentiation of normal human bronchial epithelial cells currently known (31, 39). The mechanism(s) involved in acrolein-induced epithelial cell differentiation should be investigated toelucidate a possible role of acrolein in tumor promotion.
Formation of oxygen radicals has been hypothesized to play
a role in tumor promotion (18-20). Generation of radicals andsubsequent lipid peroxidation may lead to depletion of thiolsby oxidation of GSH to GSSG (19, 20). Recently, a decreasedratio of GSH to GSSH was reported in mouse epidermal cellstreated with tumor promoters (40). Moreover, acrolein hasrecently been implied as one of the causative metabolites inallyl alcohol-induced lipid peroxidation, which results in GSSGformation in mouse (21) and rat liver (22). In acrolein-exposedhuman bronchial epithelial cells, no GSSG can be detectedindicating both the absence of this type of oxidative stress andthat the marked decrease in GSH is due only to its directconjugation to acrolein. Spermine has been shown to causedepletion of GSH in cultured mammalian cells, presumably bycausing oxidative stress and subsequent GSSG formation (4).It should be noted that decomposition of spermine may lead toformation of acrolein which by conjugation will contribute tothe spermine-induced loss of GSH.
Investigation of acrolein-induced DNA damage in normalhuman cells is of importance because acrolein has recently beenfound to be mutagenic in human fibroblasts.4 From studies withDNA in vitro, acrolein was shown to cause cyclic 1,N2-propan-
odeoxyguanosine adducts (23). These in vitro studies also indicate that acrolein would be able to cross-link DNA. Whenassayed in human bronchial epithelial cells, acrolein causesboth significant DNA single strand breaks and DNA proteincross-links. Moreover, low levels of acrolein-induced inter-strand DNA cross-links are indicated by the concave curvatureof the elution profile. The observation that acrolein causesdifferent types of DNA damage should also be of importanceto fully understand the genotoxicity of the chemotherapeuticagent cyclophosphamide, from which acrolein is a primarymetabolite. Because time-dependent formation of DNA inter-strand cross-links has been shown in cyclophosphamide-ex-posed human leukemia cells (20), acrolein may have contributedto this type of DNA damage. However, the potential role of thedifferent types of acrolein-induced DNA damage in humanbronchial epithelial cells is difficult to evaluate since the concentration of acrolein, i.e., 30 fiM, that causes significant damage is also quite cytotoxic in terms of both cloning ability andmembrane integrity.
When compared to several aldehydes and peroxides previously investigated by us including formaldehyde, acetaldehyde,bcn/oy(peroxide, and hydrogen peroxide (41, 42), acrolein isthe most potent agent on a molar basis to induce pathobiolog-ical effects in bronchial epithelial cells. Previous studies withbronchial fibroblasts also show acrolein to be much more cytotoxic and potent in depleting total free cellular thiols thanother tobacco smoke or automotive exhaust-related aldehydes,e.g., formaldehyde and acetaldehyde (29).
In human cells, acrolein has now been shown to cause a broadrange of cytopathic effects that relate to both the initiation andpromotion phase of carcinogenesis. To human bronchial epithelial cells in culture, micromolar concentrations of acroleinare highly cytotoxic in terms of decreased cloning ability,growth rate, and membrane integrity, markedly affect freenonprotein and protein thiol status, induce different squamousmorphological changes, and cause several types of DNA damage. Further studies are required to explore both mechanismsunderlying these effects and their possible importance in thedevelopment of chronic obstructive pulmonary disease and lungcancer in tobacco smokers.
ACKNOWLEDGMENTS
We appreciate the helpful comments of Dr. V. Wilson. Dr. P.Moldéusand Dr. I. Cotgreave are acknowledged for providing guidance
1720
on March 16, 2020. © 1988 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

EFFECTS OF ACROLEIN IN HUMAN BRONCHIAL CELLS
with the thiol assays. The technical assistance of M. Sjöströmis alsoappreciated.
REFERENCES
1. Izard, C, and Libermann, C. Acrolein. Mutât.Res., 47: 115-138, 1978.2. IARC Monographs on the Evaluation of the Carcinogenic Risk of Chemicals
to Humans: Some Monomers, Plastics, Synthetic Elastomers, and Acrolein.Vol. 19, pp. 479-495. Geneva: World Health Organization, 1979.
3. Beauchamp, R. O., Jr., Andjelkovich, D. A., Kligerman, A. D., Morgan, K.T., and Heck, H. d'A. A critical review of the literature on acrolein toxicity.CRC Crit. Rev. Toxicol., ¡4:309-380, 1985.
4. Russo, A., Mitchell, J. B., DeGraff, W., Friedman, N., and Gamson, J.Depletion of cellular glutathione by exogenous spermim- in V79 cells: implications for spermine-induced hyperthermic sensitization. Cancer Res., 45:4910-4914, 1985.
5. Gurtoo, H. L., Hipkens, J. H., and Sharma, S. D. Role of glutathione in themetabolism-dependent toxicity and chemotherapy of cyclophosphamide.Cancer Res., 41: 3584-3591, 1981.
6. Gurtoo, H. L., Mannello, A. J., Struck, R. F., Paul, B., and Dahms, R. P.Studies on the mechanism of denaturation of cytochrome P-450 by cyclophosphamide and its metabolites. J. Biol. Chem., 256: 11691-11701, 1981.
7. Hales, B. F. Comparison of the mutagenicity and teratogenicity of cyclophosphamide and its active metabolites, 4-hydroxycyclophosphamide, phos-phoramide mustard, and acrolein. Cancer Res., 42: 3016-3021, 1982.
8. Fleer, R., and Brendel, M. Toxicity, interstrand cross-links and DNA fragmentation induced by •"activated"cyclophosphamide in yeast: comparativestudies on 4-hydroperoxy-cyclophosphamide, its mono-functional analogue,acrolein, phosphoramide mustard, and nor nitrogen mustard. Chem. Biol.Interact., 39: 1-15, 1982.
9. Wilmer, J. L., Erexson, G. L., and Kligerman, A. D. Attenuation ofcytoge-netic damage by 2-mercaptoethanesulfonate in cultured human lymphocytesexposed to cyclophosphamide and its reactive metabolites. Cancer Res., 46:203-210, 1986.
10. Au, W., Sokova, O. I., Kopnin, B., and Arrighi, F. E. Cytogenetic toxicity ofcyclophosphamide and its metabolites in vitro. Cytogenet. Cell Genet., 26:108-116, 1980.
11. Costa, D. L., Kutzman, R. S., Lehmann, J. R., and Drew, R. T. Altered lungfunction and structure in the rat after subchronic exposure to acrolein. Am.Rev. Respir. Dis., 133: 286-291, 1986.
12. Schauenstein, E., Esterbauer, H., and Zollner, H. (eds.). Aldehydes in Biological Systems: Their Natural Occurrence and Biological Activities. London:Pion Limited, 1977.
13. Ohno, Y., Ormstad, K., Ross, D., and Orrenius, S. Mechanism of allylalcohol toxicity and protective effects of low-molecular weight thiols studiedwith isolated rat hepatocytes. Toxicol. Appi. Pharmacol., 78:169-179,1985.
14. Kii. R. H., and Billings, R. E. The role of mitochondria! glutathione andcellular protein sulfhydryls in formaldehyde toxicity in glutathione-depletedrat hepatocytes. Arch. Biochem. Biophys., 247: 183-189, 1986.
15. Crook, T. R., Sou lumii. R. L., Whyman, G. D., and McLean, A. E. M.Glutathione depletion as determinant of sensitivity of human leukemia cellsto cyclophosphamide. Cancer Res., 46: 5035-5038, 1986.
16. Lam, C.-W., Casanova, M., and Heck, H. d'A. Depletion of nasal mucosalglutathione by acrolein and enhancement of formaldehyde-induced DNAprotein crosslinking by simultaneous exposure to acrolein. Arch. Toxicol.,58:67-71, 1985.
17. Heck, H. d'A., Casanova, M., Lam, C.-W., and Swenberg, J. A. The formation of DNA-protein cross-links by aldehydes present in tobacco smoke. In:D. Hoffman and C. C. Harris (eds.). Mechanisms of Tobacco Carcinogenesis,Banbury Report No. 23, pp. 215-230. Cold Spring Harbor, NY: Cold SpringHarbor Press, 1986.
18. Cerutti, P. Prooxidant states and tumor promotion. Science (Wash. DC),227:375-381, 1985.
19. Kensler, T. W., and Taffe, B. G. Free radicals in tumor promotion. Adv. FreeRad. Biol. Med., 2: 347-387, 1986.
20. Sies, H. Biochemistry of oxidative stress. Angew. Chem. Int. Ed. Engl., 25:1058-1071, 1986.
21. Jaeschke, H., Kleinwaechter, C., and Wendel, A. The role of acrolein in allylalcohol-induced lipid peroxidation and liver cell damage in mice. Biochem.Pharmacol., 36: 51-57, 1987.
22. Badr, M. Z., Belinsky, S. A., Kaufftnan, F. C., and Thurman, R. G. J.Mechanism of hepatotoxicity to periportal regions of the liver lobule due to
allyl alcohol: role of oxygen and lipid peroxidation. Pharm. Exp. I lier., 238:1138-1142,1986.
23. Chung, F.-L., Young, R., and Hecht, S. S. Formation of cyclic l,A*-propan-odeoxyguanosine adducts in DNA upon reaction with acrolein or crotonal-dehyde. Cancer Res., 44: 990-995, 1984.
24. Erickson, L. C., Ramonas, L. M., Zaharko, D. S., and Kohn, K. W. Cytotox-icity and DNA cross-linking activity of 4-sulphidocyclophosphamide derivatives in vitro. Cancer Res., 40:4216-4220, 1980.
25. Crook, T. R., Souhami, R. L., and McLean, A. E. M. Cytotoxicity, DNAcross-linking, and single strand breaks induced by activated cyclophosphamide and acrolein in human leukemia cells. Cancer Res., 46: 5029-5034,1986.
26. Lutz, D., Eder, E., Neudecker, T., and Henschler, D. Structure-mutagenicityrelationship in «,,-i-unsaturatedcarbonylic compounds and their corresponding allylic alcohols. Mutât.Res., 93:305-315, 1982.
27. Marnett, L. J., Hurd, H. K., Hollstein, M. C., Levin, D. E., Esterbauer, H.,and Ames. B. N. Naturally occurring carbonyl compounds are mutagens inSalmonella tester strain TA 104. Mutât.Res., 148: 25-34, 1985.
28. Feron, V. J., and Kruysse, A. Effects of exposure to acrolein vapor in hamsterssimultaneously treated with bcnzo(a)pyrene or diethylnitrosamine. J. Toxicol. Environ. Health., 3: 379-394, 1977.
29. Krokan, H., Grafström, R. C., Sundqvist, K., Esterbauer, H., and Harris, C.C. Cytotoxicity, thiol depletion and inhibition of O6-methylguanine-DNAmethyltransferase by various aldehydes in cultured human bronchial libroblasts. Carcinogenesis (Lond.), 6:1755-1759, 1985.
30. Lechner, J. F., Stoner, G. D., Yoakum, G. H., Willey, J. C., Grafström,R.C., Masui, T., LaVeck, M. A., and Harris, C. C. In vitro Carcinogenesisstudies with human tracheobronchial tissues and cells. In: L. J. Schiff (ed.),In vitro models of Respiratory Epithelium, pp. 143-159. Florida: CRC Press,1986.
31. Willey, J. C., Saladino, A. J., Ozanne, C., Lechner, J. F., and Harris, C. C.Acute effects of 12-O-tetradecanoylphorbol-13-acetate, teleocidin B, or2,3,7,8-tetrachlorodibenzo-p-dioxin on cultured normal human bronchialepithelial cells. Carcinogenesis (Lond.), 5: 209-215, 1984.
32. Masui, T., Wakefield, L. M., Lechner, J. F., LaVeck, M. A., Sporn, M. B.,and Harris, C. C. Type ¿¡-transforminggrowth factor is the primary differentiation-inducing serum factor for normal human bronchial epithelial cells.Proc. Nati. Acad. Sci. USA, S3: 2438-2442, 1986.
33. Saville, B. A scheme for the colorimetrie determination of microgramamounts of thiols. Analyst, 83:670-672, 1958.
34. Cotgreave, I. C., and Moldéus,P. Methodologies for the application ofmonobromobimane to the simultaneous analysis of soluble and protein thiolcomponents of biological systems. J. Biochem. Biophys. Meth., 13: 231 -249,1986.
35. Kohn, K. W., Ewig, L. C., Erikson, L. C., and Zwelling, L. A. Measurementof strand breaks by alkaline elution. ///: E. C. Friedberg and P. C. Hanawalt(eds.), DNA Repair, A Laboratory Manual of Research Procedures, pp. 379-401. New York: Marcel Dekker, 1981.
36. Slott, V. L., and Hales, B. F. The embryolethality and teratogenicity ofacrolein in cultured rat embryos. Teratology, 34: 155-163, 1986.
37. Yuspa, S. H., and Morgan, D. L. Mouse skin resistant to terminal differentiation associated with initiation of Carcinogenesis. Nature (Lond.), 293:72-74, 1981.
38. Willey, J. C., Grafström,R. C., Moser, C. E., Jr., Ozanne, C., Sundqvist, K.,and Harris, C. C. The biochemical and morphological effects of cigarettesmoke condensate and its fractions on normal human bronchial epithelialcells in vitro. Cancer Res., 47: 2045-2049,1987.
39. Harris, C. C., Willey, J. C., Saladino, A. J., and Grafström,R. C. Effects oftumor promoters, aldehydes, peroxides, and tobacco smoke condensate ongrowth and differentiation of cultured normal and transformed human bronchial cells. In: M. J. Mass et al. (eds.), Carcinogenesis, Vol. 8, pp. 159-171.New York: Raven Press, 1985.
40. Perchellet, J. P., Perchellet, E. M., Orten, D. K., and Schneider, B. A.Decreased ratio of reduced/oxidized glutathione in mouse epidermal cellstreated with tumor promoters. Carcinogenesis (Lond.), 7: 503-506, 1986.
41. Saladino, A. J., Willey, J. C., Lechner, J. F.. Grafström,R. C., LaVeck, M.,and Harris, C. C. Effects of formaldehyde, acetaldehyde, benzoyl peroxide,and hydrogen peroxide on cultured normal human bronchial epithelial cells.Cancer Res., 45:2522-2526, 1985.
42. Grafström,R. C., Willey, J. C., Sundqvist, K., and Harris, C. C. Pathobio-logical effects of tobacco smoke-related aldehydes in cultured human bronchial epithelial cells. In: D. Hoffman and C. C. Harris (eds.). Mechanisms ofTobacco Carcinogenesis, Banbury Report No. 23, pp. 273-285. Cold SpringHarbor, NY: Cold Spring Harbor Press, 1986.
1721
on March 16, 2020. © 1988 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

1988;48:1717-1721. Cancer Res Roland C. Grafström, Jeannette M. Dypbukt, James C. Willey, et al. Bronchial Epithelial CellsPathobiological Effects of Acrolein in Cultured Human
Updated version
http://cancerres.aacrjournals.org/content/48/7/1717
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/48/7/1717To request permission to re-use all or part of this article, use this link
on March 16, 2020. © 1988 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from





























