Embed Size (px)
Citation preview

2665Journal of Cell Science 111, 2665-2679 (1998)Printed in Great Britain © The Company of Biologists Limited 1998JCS9826
Participation of GRP94-related protein in secretion of pancreatic bile salt-
dependent lipase and in its internalization by the intestinal epithelium
Nadine Bruneau 1, Dominique Lombardo 2 and Moïse Bendayan 1,*1Département de Pathologie et Biologie Cellulaire, Faculté de Médecine, Université de Montréal, Montréal, Quebec, H3C 3J7Canada 2INSERM U-260, Unité de Recherche de Physiopathologie des Régulations Hormono-Nutritionnelles, Marseilles, France*Author for correspondence (e-mail: [email protected])
Accepted 20 June; published on WWW 13 August 1998
d
In previous studies on the AR4-2J cell line, we have shownthat secretion of bile salt-dependent lipase (BSDL) involvesa multiprotein complex, including a protein of 94 kDa (p94)that is immunologically related to the chaperone Grp94,which seems to play essential roles in the folding process ofBSDL. Combined biochemical and immunocytochemicalinvestigations were carried out to study the secretion ofBSDL by normal pancreatic cells and its transport to thesmall intestine where this enzyme is thought to exert itsphysiological function. Both BSDL and Grp94 antigenicsites were localized and found to be associated all along thepancreatic acinar cell secretory pathway. Grp94 and BSDLremain associated from leaving the pancreas until arrivingat the intestinal lumen. In pancreatic juice, both proteins
appear as a complex of high molecular mass (180 kDa)containing at least one each of p94 and BSDL molecules,interacting by hydrophobic forces. At the intestinal level,associated Grp94 and BSDL were detected on microvilliand in the endosomal compartment of enterocytes. TheBSDL mRNA, however, was not expressed by the intestinalmucosa. The pancreatic Grp94-BSDL complex wasinternalized through the endosomal compartment ofenterocytes. Finally, the two proteins dissociated in thiscompartment and BSDL, but not Grp94, was transferredto the basolateral membrane.
Key words: Pancreatic bile salt-dependent lipase, Glucose-regulateprotein 94, Immunocytochemistry
SUMMARY
goces,),
l.,eds,L,
dare
inlls rate-es
ss
ndn
INTRODUCTION
The bile salt-dependent lipase (BSDL, EC.3.1.1.-), also cacholesterol esterase, is a lipolytic enzyme, which upactivation by the primary bile salts catalyzes the hydrolysisa wide range of substrates (Lombardo et al., 1980; Lombaand Guy, 1980). The actual function of BSDL remaicontroversial. Several data suggest, however, that physiological roles of BSDL are the facilitation of intestinhydrolysis of cholesterol esters into free cholesterol and faacids (Howles, 1996) and participation in the intestinal frecholesterol absorption as a cholesterol transfer protein (MyPayne et al., 1995). This enzyme is found in pancreatsecretions of all examined species from fish to human andbeen cloned in several species (Hernell and Blackberg, 19The main differences between species are in the numberstructure of the C-terminal tandem-repeated sequences, ware highly O-glycosylated (Wang and Hartsuck, 199Landberg et al., 1997). However, over 70% homology existsthe N-terminal region preceding the repeated sequences.
Deletion of the C-terminal region of BSDL does not affethe activity of the recombinant enzyme, the truncated BShaving the same dependence on bile salts as the native(Hanson et al., 1993). However, compared to the full-len
lledon ofrdo
nsthe
alttye-
ers-ic has94). andhich3; in
ctDL onegth
enzyme, a truncated variant lacking the C terminus startinfrom Leu-519, showed an increased susceptibility tproteolysis, suggesting that the C-terminal repeated sequenmay regulate proteolytic degradation of BSDL (Loomes1995). Each repeated sequence is also rich in proline (Pglutamic acid (E), serine (S) and threonine (T) (Han et a1987; Reue et al., 1991). Proteins that are rapidly degradwithin eukaryotic cells frequently contain such sequencereferred to as PEST regions (Rogers et al., 1986). On BSDPEST regions coexist with the C-terminal cluster of O-linkeoligosaccharides (Han et al., 1987; Reue et al., 1991) and absent in sequences of other known pancreatic enzymes.
We have shown that the transfection of CHO-ldlD cells withthe full-length cDNA of rat BSDL (CHO ldlD-6B clone), whengrown under conditions that prevent O-glycosylation, results a protein that is rapidly degraded intracellularly; only the smafraction of BSDL that was able to be O-glycosylated wasecreted (Bruneau et al., 1997). These data suggest that theof secretion of BSDL depends upon the cell’s ability to Oglycosylate at the level of the C-terminal repeated sequencand that O-glycosylation regulates its secretory proce(Bruneau et al., 1997).
We have also shown that human (Bruneau et al., 1995) arat (Bruneau and Lombardo, 1995) pancreatic BSDL, i

2666
ly
cora,g
andal.,hebyse
ia,gGd
od
ofg
i-e Mased
0.byiesleditse
d toderin-
r,
eMndS-
edand
nc,C-
,H
d
C-
N. Bruneau, D. Lombardo and M. Bendayan
contrast to other secretory pancreatic enzymes, are assocwith intracellular membranes during their secretory proceThis association with intracellular membranes involves multimeric folding complex, including a chaperone molecuimmunologically related to the glucose-regulated protein of kDa (Grp94). These data, together with previous evidence other pancreatic secretory proteins (Le-Gall and Benday1996; Vélez-Granell et al., 1994), demonstrate that the foldand secretion of pancreatic enzymes could be under the conof chaperones.
The Grp94 chaperone is a member of the glucose-regulaprotein family and shares 50% amino acid identity with the kDa heat shock protein (Hsp90) (Little et al., 1994). Grp9also known as Erp99 or endoplasmin, is a 94 kDa glycoproteGrp94 was first identified as a gene product whose synthesregulated upon glucose starvation and in response to varexperimental treatments that induce the accumulation unfolded proteins in the endoplasmic reticulum (ER) (Little al., 1994). It has been shown to contribute to protein traffickiand appears to associate with folding intermediates onumber of oligomeric proteins. In the endoplasmic reticulu(ER), Grp78 and Grp94 act sequentially to fold intermediaof the newly synthesized immunoglobulin chains (Melnick al., 1992, 1994). The Grp94 chaperone, as protein disulfiisomerase (PDI) and Grp78, is located in the ER lumenvarious cells. However, despite possessing the ER retensignal KDEL (Lys-Asp-Glu-Leu) at their C termini, Grp94 awell PDI and Grp78 have also been localized in the Goapparatus and secretory granules of pancreatic cells (Takemet al., 1992).
Since the biosynthesis of pancreatic BSDL seems to dependent upon the presence of the Grp94-related pro(Bruneau and Lombardo, 1995), we attempted to determineimmunocytochemical and biochemical approaches whetherGrp94-related protein was associated with BSDL in normpancreatic acinar cells and secreted. The present studyshown that Grp94 and BSDL antigenic sites are indeassociated all along the secretory pathway and, upon secreremain associated in the pancreatic juice when transpofrom the pancreas to the intestinal lumen. We have furthshown that the Grp94-related protein-BSDL complex internalized by enterocytes. Finally, both proteins dissociatethe late endosomal compartment and the BSDL, but not Grp94, is transferred to the basolateral membrane.
MATERIALS AND METHODS
Biological materialPancreatic and duodenal tissues were sampled from normal 10Sprague-Dawley male rats and processed for biochemical morphological studies.
Pancreatic juice was collected upon cannulation of the pancreduct from various anaesthetized rats for 6 hour periods.
cDNA probescDNA probes specific for BSDL and for Grp94 were obtained bchemical synthesis (Life Technologies, Burlington, ON, Canada). Oprobe corresponded to a sequence complementary to nucleo1041-1071 of pancreatic BSDL mRNA (Kissel et al., 1989), and tother to a sequence complementary to nucleotides 1798-1828murine Grp94 mRNA (Mazzarella and Green, 1987). cDNA prob
iatedss.a
le94foran,ingtrol
ted904,in.
is isiousof
etngf amtesetde
oftionslgioto
betein by
theal
hasedtion,rteder
is inthe
0 gand
atic
yne
tideshe ofes
were labeled with digoxigenin (Dig-) as described previous(Gingras and Bendayan, 1995).
AntibodiesAffinity-purified rabbit polyclonal antibodies against rat pancreatiBSDL (Abouakil et al., 1988) and monoclonal antibodies specific fGrp94 (clone 9G10) from StressGen Biotechnology Corp. (VictoriBC, Canada) were used for immunocytochemistry, immunoblottinand immunoprecipitation. These antibodies are well characterized have previously been used in various experiments (Nauseef et 1995; Kuznetsov et al., 1994; Bruneau and Lombardo, 1995). Tmonoclonal antibody directed against the Grp94 was prepared immunization of rats. Rat spleen cells were then fused with mouplasmacytoma cell lines (StressGen Biotechnology Corp., VictorBC, Canada). Thus secondary antibodies to rat or mouse Imolecules were used for the immunochemical animmunocytochemical techniques.
Protein assaysProteins were routinely assayed by the bicinchoninic acid meth(Pierce, Rockford, IL) using bovine serum albumin as a standard.
Polyacrylamide gel electrophoresis and western blottingGel electrophoresis (SDS-PAGE) was performed in slab gels polyacrylamide (7.5%) and sodium dodecyl sulfate (1%) in reducinconditions according to Laemmli (1970), using a BioRad minprotean II apparatus. After SDS-PAGE, proteins werelectrophoretically transferred to a nitrocellulose membrane in 0.2Tris/HCl buffer pH 9.2 (10% methanol), at 150 mA for 3 hours in cold room (Burnette, 1981). Completeness of transfer was assesby staining polyacrylamide gels with Coomassie Blue R25Destaining was performed in ethanol/acetic acid/water (2/3/35 volume). Western blots were performed using the specific antibodfor BSDL and Grp94. The antigen-antibody complexes were reveaeither by the BM chemiluminescence Western-blotting k(Boehringer-Mannheim, Germany) or by the alkaline-phosphatatechnique.
ImmunoprecipitationPancreatic juice proteins were incubated overnight at 4°C with 10 µgof antibodies. Pre-washed Protein A-Sepharose (10 mg) was addeantibody-antigen complexes and incubated for 4 hours at 4°C unagitation. At the end of the incubation, the antigen-antibody-ProteA complex was recovered by centrifugation (10,000 g, 20 minutes).The pellet was then washed twice with the 10 mM Tris/HCl buffepH 7.4, 25 mM EDTA and 1% Triton X-100 (washing buffer), twicewith the washing buffer supplemented with 1 M NaCl, twice with thlast buffer supplemented with 0.1% SDS and twice again with 10 mTris/HCl buffer, pH 7.4, containing 5 mM EDTA. The pellet was thetransferred into Laemmli’s sample buffer for the SDS-PAGE, warmefor 5 minutes at 95°C, centrifuged and electrophoresed on SDPAGE.
In situ hybridizationRat tissues were fixed with Bouin’s solution for 24-48 hours, washin water, dehydrated through increasing concentrations of ethanol embedded in paraffin. 5 µm thick sections were mounted in diethylpyrocarbonate (DEPC)-treated water (RNAse-free water) oSuperfrost/Plus slides (Fisher Scientifique, Montreal, QuebeCanada). The sections were deparaffinized and rinsed in DEPtreated water. They were then washed twice in 2× SSC (1× SSC = 150mM sodium chloride, 15 mM sodium citrate) for 10 minutes at 60°Cdipped in DEPC-treated water, then in 0.05 M Tris/HCl buffer, p7.6, at room temperature incubated with Proteinase K (5 µg/ml in 50mM Tris/HCl, buffer pH 7.6) for 5 minutes at 37°C, and finally rinsewith 50 mM Tris/HCl buffer, pH 7.6. The sections were fixed in 0.4%paraformaldehyde in PBS at 4°C for 20 minutes, rinsed in DEP

2667Pancreatic bile salt-dependent lipase secretion and internalization
y
L
ofofn,c.,d
chhedssehe
ofhenethe
dn,s.yo
forolLr
,inalslet
y
Lry
th).
n).rs
ldard
treated water and incubated for 1 hour at 37°C in the hybridizatbuffer (R&D systems, Minneapolis, MN, USA). Dig-labeled probewere diluted to the required concentration in the hybridization buffThe hybridization with the specific probe was then performed inhumid chamber at 37°C overnight. After hybridization, the sectiowere washed at 37°C twice for 5 minutes successively with: ×SSC/30% formamide, 2× SSC/30% formamide and 0.2× SSC/30%formamide. They were incubated in 50 mM Tris/HCl buffer, pH 7.150 mM NaCl, 2mM MgCl2 (TBS) containing 0.1% bovine serumalbumin and the Dig-labeled hybrids were detected with the anti-Dalkaline phosphatase antibodies diluted 1/500 in TBS containing 0BSA. The sections were then incubated twice for 5 minutes in revelation buffer (100 mM Tris/HCl, pH 9.5, 150 mM NaCl, 50 mMMgCl2) and finally incubated with the revelation buffer containinNBT and X-phosphate (Boehringer-Mannheim, Germany). They wmounted with 50% glycerol in PBS and examined with a LeOrthoplan microscope (Montreal, Quebec, Canada). Contexperiments were performed following exactly the same protocol using the Dig-labeled sense probes.
ImmunocytochemistryImmunofluorescenceRat pancreatic and duodenal tissues were fixed in Bouin’s fixative embedded in paraffin. Tissue sections were deparaffinized and watwice in 10 mM phosphate buffer, pH 7.4, 150 mM NaCl (PBS) aincubated with the anti-BSDL (dilution 1/10, 2 hours) or the anGrp94 (dilution 1/20, overnight) antibodies. For revealing BSDantigenic sites, the section were washed twice in the PBS and incubated with the FITC-goat anti-rabbit IgG antibody (SigmChemicals, St Louis, MO, USA) for 1 hour. For revealing Grp9antigenic sites, the sections were incubated successively wbiotinylated anti-mouse IgG antibody (1/200, 1 hour), streptavidHRP (1/500, 30 minutes) and FITC-tyramide (1/50, 10 minutes) in ×amplification diluent (NEN Life Science products, Boston, MA, USAas described previously (Mayer and Bendayan, 1997). The sectwere observed on a Leitz DM RB microscope using regular and immersion PL Fluotar objectives.
Immunoelectron microscopyRat pancreatic and duodenal tissues were fixed by immersion with(v/v) glutaraldehyde in 0.1 M sodium phosphate buffer, pH 7.4, fohours at 4°C. The tissue samples were washed in the phosphate bdehydrated by a series of graded ethanol solutions and embeddUnicryl or Lowicryl (British Biocell, Cardiff, UK) at −30°C asdescribed previously (Bendayan, 1995). Thin sections were mounon Parlodion and carbon-coated nickel grids. The grids wsuccessively incubated by floating them on a drop of 1% ovalbumin PBS for 30 minutes at room temperature, a drop of the primantibody (anti-BSDL at 1/200 dilution for 3 hours at roomtemperature or anti-Grp94 at 1/20 dilution at 4°C overnight), arinsed with PBS. They were then transferred to 1% ovalbumsolution and incubated for 30 minutes on a drop of a goat anti-raIgG-gold complex for the BSDL or a goat anti-mouse IgG-gocomplex for the Grp94. The grids were then thoroughly washed wPBS and distilled water. The thin sections were stained with uraacetate and examined with a Philips 410 electron microscope.
For sequential double labeling, the tissue sections, mountednickel grids, were incubated with the monoclonal antibody specfor Grp94 (dilution 1/20) at 4°C overnight and the goat anti-mouIgG-gold complex (10 nm in size) for 30 minutes. Upon rinsing wiPBS and distilled water, the same protocol was repeated with polyclonal antibody specific for BSDL (dilution 1/200) and the goanti-rabbit IgG-gold complex (15 nm in size).
Control experimentsControl experiments, for light and electron microscop
ionser. ans4
6,
ig-.1%the
gereitzrolbut
andshedndti-Lthena4ith
in-1)
ionsoil-
1%r 2uffer,ed in
tedere
inary
ndin
bbitldithnyl
onificseththe
at
y
immunochemistry, were performed by omitting the primary antibodand by using the antigen-adsorbed-antibody complexes.
Quantitative evaluation and determination of percentage ofassociationThe labeling intensities obtained by electron microscopy for BSDand Grp94, defined as the number of gold particles per µm2, wereestablished over different cellular compartments on thin sections pancreatic and duodenal tissues. Direct planimetry and counting gold particles were carried out as described previously (Bendaya1995) using a Videoplan 2 image processing system (Carl Zeiss InToronto, Ontario, Canada). 30 micrographs were recorded ananalyzed for each protein in each cellular compartment and in eatissue. The percentage of association, BSDL-Grp94 revealed by tdouble-labeling experiments was evaluated as follows: only golparticles revealing Grp94 (10 nm gold) that were separated by lethan 20 nm from a gold particle representing BSDL (15 nm gold) wertaken into account and both particles were considered associated. Tpercentage of association was calculated by dividing the number associated particles by the total number of Grp94 particles. For tevaluation of the density of labeling along the basolateral membraof the enterocyte, we measured the length of the membrane and density was defined as the number of gold particles per µm.
RESULTS
Intracellular localization of BSDL in rat pancreasIn order to determine the precise localization of BSDL anGrp94 in the rat pancreatic acinar cells and their associatiowe used different morphological and biochemical approacheIn situ hybridization was performed on rat pancreatic tissue busing Dig-tagged synthetic oligonucleotides complementary tBSDL cDNA. This led to a black positive reaction restrictedto the basolateral region of the acinar cells (Fig. 1A). Cells othe islet of Langerhans were negative, as were all nuclei. Nsignal was found when sections were processed under contconditions, i.e. using synthetic sense oligonucleotides to BSDcDNA (result not illustrated). Therefore, the acinar cells appeato be the only pancreatic cells expressing BSDL mRNA.
Immunofluorescence for detecting BSDL antigenic sitesperformed on pancreatic tissue, revealed a positive signal acinar cells. The staining was particularly intense in the apicregion of the cells (Fig. 1B), known to contain large amountof secretory granules. No specific labeling was observed in isendocrine cells.
At the electron microscope level, the labeling obtained busing specific antibodies to BSDL in conjunction with theimmunogold approach, indicated the presence of BSDantigenic sites in the various compartments along the secretoroute, rough endoplasmic reticulum (RER)→Golgiapparatus→condensing vacuoles→zymogen granules (Fig.1C). These data agreed with previous results obtained wihuman pancreas (Lechêne de La Porte et al., 1987Quantitation of the labeling showed that BSDL waspreferentially present in those compartments, with aincreasing gradient from ER to zymogen granules (Table 1This reflects the process of protein concentration that occualong the RER→Golgi→granules secretory pathway(Bendayan et al., 1980). Immunofluorescence and immunogocontrol experiments resulted in absence of labeling. The acincell is thus able to express the specific mRNA for BSDL an
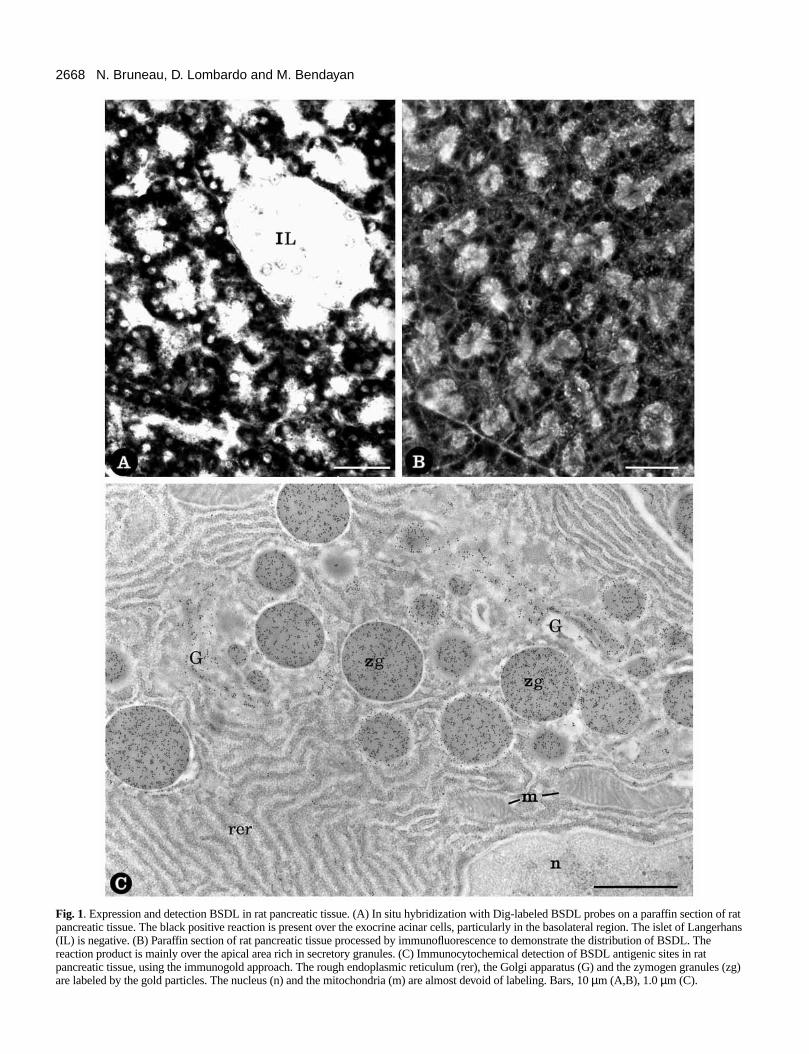
2668 N. Bruneau, D. Lombardo and M. Bendayan
Fig. 1. Expression and detection BSDL in rat pancreatic tissue. (A) In situ hybridization with Dig-labeled BSDL probes on a paraffin section of ratpancreatic tissue. The black positive reaction is present over the exocrine acinar cells, particularly in the basolateral region. The islet of Langerhans(IL) is negative. (B) Paraffin section of rat pancreatic tissue processed by immunofluorescence to demonstrate the distribution of BSDL. Thereaction product is mainly over the apical area rich in secretory granules. (C) Immunocytochemical detection of BSDL antigenic sites in ratpancreatic tissue, using the immunogold approach. The rough endoplasmic reticulum (rer), the Golgi apparatus (G) and the zymogen granules (zg)are labeled by the gold particles. The nucleus (n) and the mitochondria (m) are almost devoid of labeling. Bars, 10 µm (A,B), 1.0 µm (C).

2669Pancreatic bile salt-dependent lipase secretion and internalization
ofheic
hatn
ellshehedenticesey.d,Lof
renseonsE,llyatinthlarisertwotic a
sng).
ted4r).aesesnsce
bye
ally,edatane
Table 1. Densities of labeling obtained with the anti-BSDLand the anti-Grp94 over different cellular compartments
of the rat pancreatic acinar cellsCellular compartments BSDL* Grp94* Association (%)
RER 7.5±0.4 2.9±0.2 28.8±1.4Golgi apparatus 84.8±2.8 14.5±0.9 37.0±1.4Condensing vacuoles 239.5±7.4 36.4±2.6 55.5±1.6Zymogen granules 235.8±3.7 31.8±1.8 61.9±1.6Acinar lumen 135.9±7.8 19.5±1.3 58.7±1.8Mitochondria 2.9±0.4 0.9±0.2
% Data show the percentage of association of Grp94 with BSDL. *Gold particles/µm2 (mean values ± s.e.m.).
to translate it. In its fundamental attributes, the secretory roof BSDL was similar to that of other pancreatic secretoproteins and it followed a regulated pathway of secretion.
Intracellular localization of Grp94 in the rat pancreasThe expression of Grp94 was assessed in pancreatic tissusitu hybridization was carried out by using Dig-tagged syntheoligonucleotides complementary to Grp94 cDNA. A blacpositive staining was detected in the basolateral area of the accells while a weak staining was present in the islet cells (F2A). No signal was found under control conditions, i.e. usisynthetic sense oligonucleotides to Grp94 cDNA (data nshown). This indicates that the mRNA of Grp94 is transcribby exocrine and, to a lesser extent, by endocrine pancreatic c
Immunofluorescence for Grp94 antigenic sites revealedstrong positive signal in pancreatic acinar cells, whereas signal was much lower in endocrine cells. The staining acinar cells was localized around the nuclei and in the apregion of the cells. Nuclei were not labeled (Fig. 2B).
At the electron microscope level, specifiimmunocytochemical labeling for Grp94 was found ovdifferent cellular compartments of the acinar cells (Fig. 2CIndeed, the presence of Grp94 antigenic sites was detectethe RER, the Golgi apparatus, the condensing vacuoles andzymogen granules. Quantitation of the labeling was perform(Table 1) and confirmed that the Grp94 is expressed by acinar cells, this chaperone being present in all tcompartments of the regulated secretory pathway. Conexperiments confirmed the specificity of the results.
Co-distribution of BSDL and GRP94 in the pancreasIn pancreatic acinar cells, BSDL and Grp94 appear to folloa regulated pathway of secretion. Previous studies hdemonstrated that a multiprotein complex, including thGrp94-related protein, plays an essential role in the foldingBSDL (Bruneau and Lombardo, 1995).
To demonstrate the association of BSDL with Grp9double-labeling experiments were performed with antibodagainst BSDL and Grp94 combined to different-sized goparticles. Results indicate that the association of BSDL wGrp94 takes place all along the secretory pathway of acicell, from the RER to the zymogen granules (Fig. 3). Indea high number of small (10 nm) and large (15 nm) goparticles, representative of Grp94 and BSDL antigenic sirespectively, were found within less than 20 nm distanceeach other, suggesting that they are associated. Howeverall gold particles revealing BSDL or Grp94 were found to b
utery
e. Intickinarig.
ngot
edells. athein
ical
cer).d in theedthehetrol
wavee
of
4,iesldithnared,ldtes of, note
associated; a significant number remained free. The degreeassociation between both proteins varied according to tcellular compartment and increased from the endoplasmreticulum to the zymogen granules (Table 1). This suggests tthe Grp94-related protein is associated mainly with BSDL ithe late compartment of the secretory process.
Secretion of BSDL and Grp94 by pancreatic acinarcells Based on these results, we predicted that pancreatic acinar cwould secrete both proteins. Therefore, we checked for tpresence of BSDL and Grp94 in pancreatic secretion. At telectron microscope level, the immunogold labeling obtaineby using the specific antibodies to BSDL indicated thpresence of BSDL antigenic sites in discharging zymogegranules and in the content of the acinar lumen (pancreajuice) (Fig. 4A). The proteins of the pancreatic juice werseparated by SDS-PAGE, electrotransferred to a nitrocellulomembrane and finally developed with the anti-BSDL antibodAs shown in Fig. 4A(inset), a single band was recovereindicating that one protein reacts with the antibody to rat BSDwith a molecular mass of 74 kDa, compatible with the mass mature BSDL in rat pancreatic juice (Abouakil et al., 1993).
For the Grp94, gold particles revealing antigenic sites wealso found in discharging zymogen granules and over the deflocculent material present in the acinar lumen (Fig. 4B). Tconfirm the secretion of this Grp94-related protein, the proteiof the pancreatic juice were separated by SDS-PAGelectrotransferred to a nitrocellulose membrane and finadeveloped with the monoclonal antibody to Grp94 and a goanti-rat IgG antibody conjugated with peroxidase. As shown Fig. 4B(inset), antibodies to Grp94 revealed two proteins, wimolecular masses of 94 kDa and 180 kDa. This high molecumass for Grp94-related protein, which is normally 94 kDa, compatible with either an homodimer of p94 or a heterodimcontaining at least one p94 protein. These data suggest that forms of Grp94-related protein could be present in pancreasecretion, one being in a monomeric form and the other incomplex form.
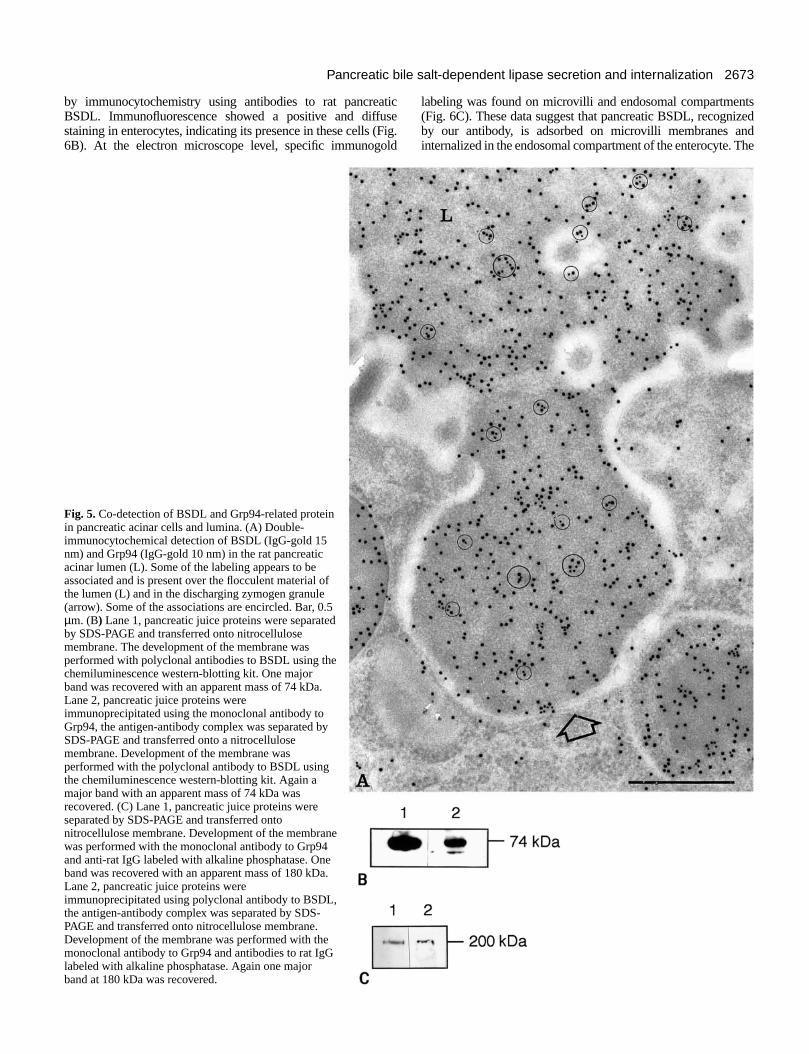
Association between BSDL and Grp94 in pancreaticjuice Double-immunogold labeling revealed the simultaneoupresence of BSDL and Grp94 antigenic sites in dischargigranules and in the content of the acinar lumen (Fig. 5AMoreover, we have found that the Grp94 remains associawith the BSDL in both compartments. The amount of Grp9labeling associated with that of BSDL in acinar lumen is similato that found for the zymogen granules (about 60%; Table 1
We have shown (Fig. 4B,inset) that Grp94 displays molecular mass compatible with the formation of a complex. Wthus attempted to determine whether this complex encompasboth the BSDL and the Grp94. The pancreatic juice proteiwere subjected to immunoprecipitation with antibodies specififor Grp94 on the one hand and for BSDL on the other. Thimmunoprecipitated material with anti-Grp94 was separated SDS-PAGE and electrotransferred onto a nitrocellulosmembrane. The Ponceau Red staining indicated that, essentinot just one but several proteins were co-immunoprecipitatwith the antibody to Grp94 (data not shown). This indicates thGrp94 is associated to a few pancreatic proteins. This membr
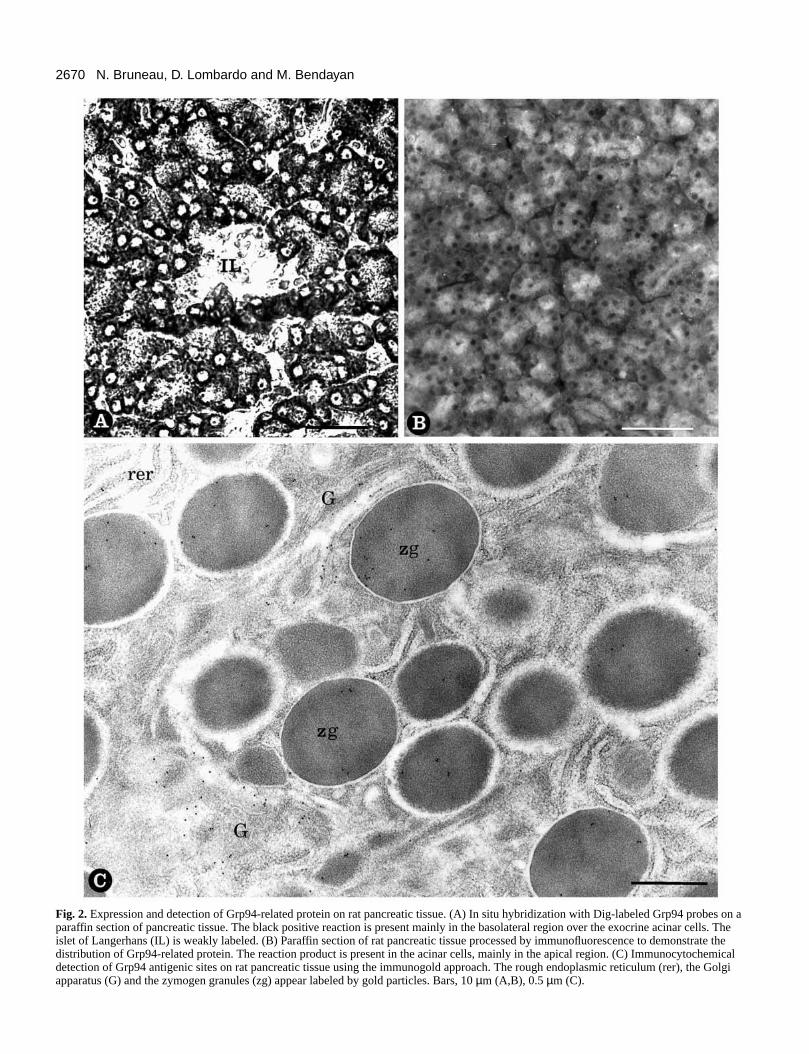
2670 N. Bruneau, D. Lombardo and M. Bendayan
Fig. 2.Expression and detection of Grp94-related protein on rat pancreatic tissue. (A) In situ hybridization with Dig-labeled Grp94 probes on aparaffin section of pancreatic tissue. The black positive reaction is present mainly in the basolateral region over the exocrine acinar cells. Theislet of Langerhans (IL) is weakly labeled. (B) Paraffin section of rat pancreatic tissue processed by immunofluorescence to demonstrate thedistribution of Grp94-related protein. The reaction product is present in the acinar cells, mainly in the apical region. (C) Immunocytochemicaldetection of Grp94 antigenic sites on rat pancreatic tissue using the immunogold approach. The rough endoplasmic reticulum (rer), the Golgiapparatus (G) and the zymogen granules (zg) appear labeled by gold particles. Bars, 10 µm (A,B), 0.5 µm (C).
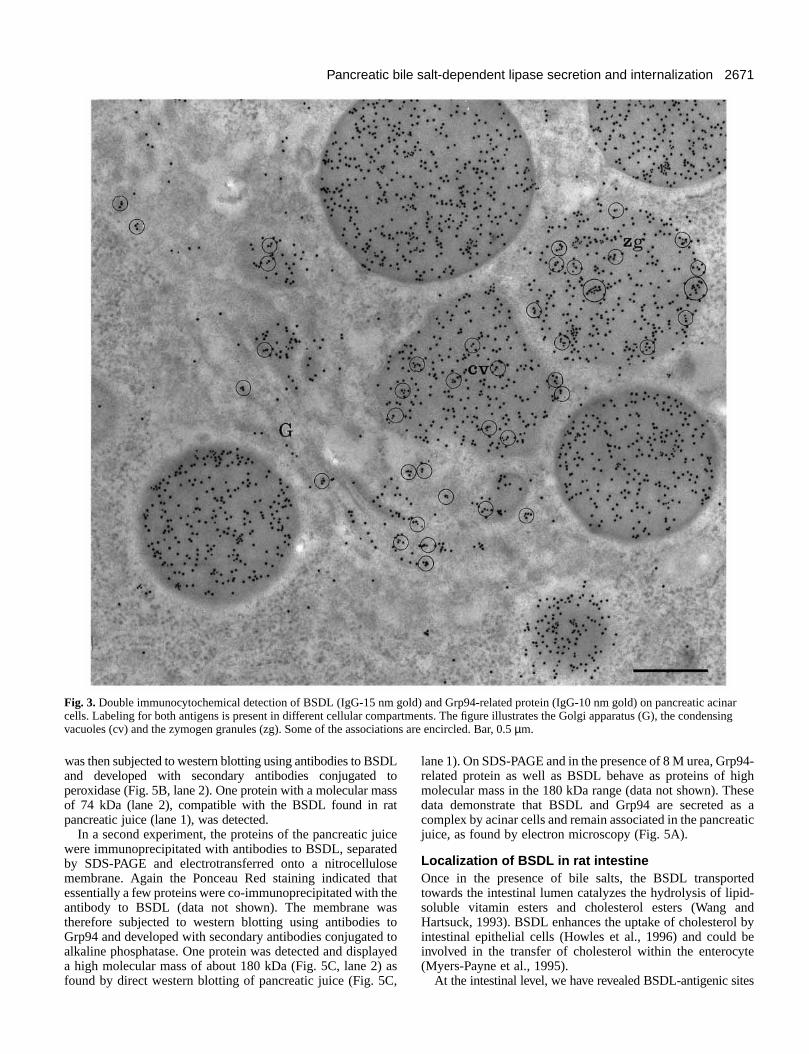
2671Pancreatic bile salt-dependent lipase secretion and internalization
4-ghse
s aatic
ed-
ndbyee
s
Fig. 3.Double immunocytochemical detection of BSDL (IgG-15 nm gold) and Grp94-related protein (IgG-10 nm gold) on pancreatic acinarcells. Labeling for both antigens is present in different cellular compartments. The figure illustrates the Golgi apparatus (G), the condensingvacuoles (cv) and the zymogen granules (zg). Some of the associations are encircled. Bar, 0.5 µm.
was then subjected to western blotting using antibodies to BSand developed with secondary antibodies conjugated peroxidase (Fig. 5B, lane 2). One protein with a molecular mof 74 kDa (lane 2), compatible with the BSDL found in rapancreatic juice (lane 1), was detected.
In a second experiment, the proteins of the pancreatic juwere immunoprecipitated with antibodies to BSDL, separatby SDS-PAGE and electrotransferred onto a nitrocellulomembrane. Again the Ponceau Red staining indicated tessentially a few proteins were co-immunoprecipitated with tantibody to BSDL (data not shown). The membrane wtherefore subjected to western blotting using antibodies Grp94 and developed with secondary antibodies conjugatedalkaline phosphatase. One protein was detected and displaa high molecular mass of about 180 kDa (Fig. 5C, lane 2)found by direct western blotting of pancreatic juice (Fig. 5C
DLto
asst
iceedsehatheasto toyed
as,
lane 1). On SDS-PAGE and in the presence of 8 M urea, Grp9related protein as well as BSDL behave as proteins of himolecular mass in the 180 kDa range (data not shown). Thedata demonstrate that BSDL and Grp94 are secreted acomplex by acinar cells and remain associated in the pancrejuice, as found by electron microscopy (Fig. 5A).
Localization of BSDL in rat intestineOnce in the presence of bile salts, the BSDL transporttowards the intestinal lumen catalyzes the hydrolysis of lipidsoluble vitamin esters and cholesterol esters (Wang aHartsuck, 1993). BSDL enhances the uptake of cholesterol intestinal epithelial cells (Howles et al., 1996) and could binvolved in the transfer of cholesterol within the enterocyt(Myers-Payne et al., 1995).
At the intestinal level, we have revealed BSDL-antigenic site
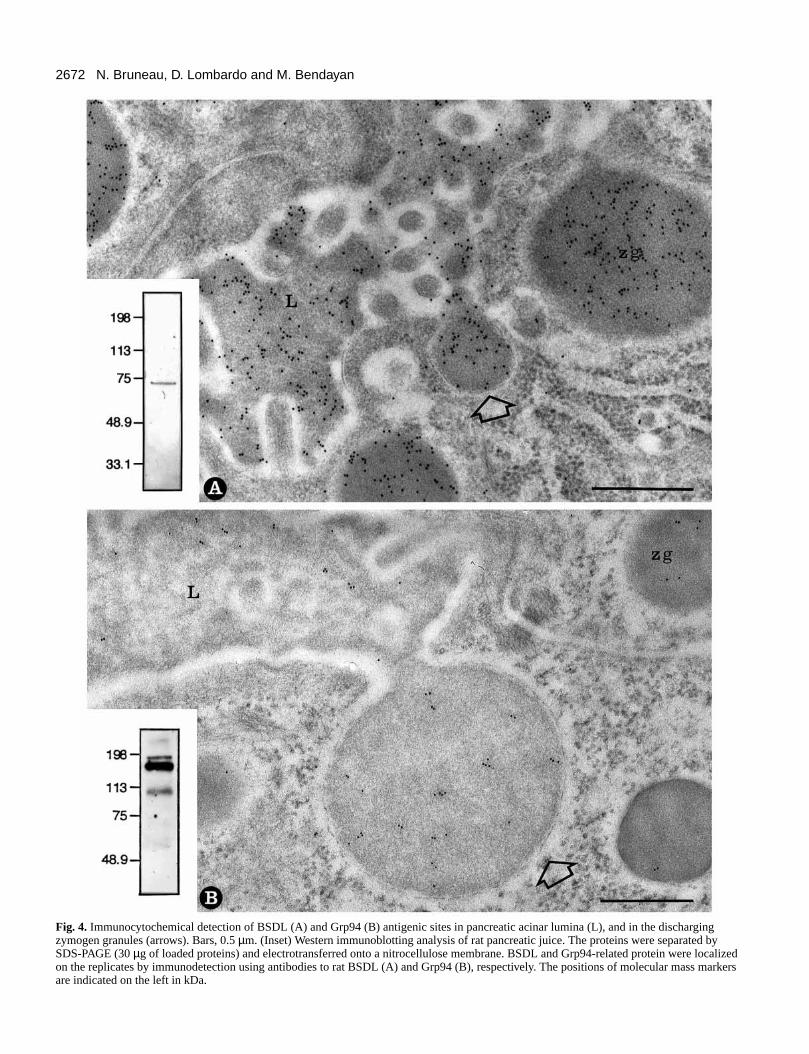
2672 N. Bruneau, D. Lombardo and M. Bendayan
Fig. 4. Immunocytochemical detection of BSDL (A) and Grp94 (B) antigenic sites in pancreatic acinar lumina (L), and in the dischargingzymogen granules (arrows). Bars, 0.5 µm. (Inset) Western immunoblotting analysis of rat pancreatic juice. The proteins were separated bySDS-PAGE (30 µg of loaded proteins) and electrotransferred onto a nitrocellulose membrane. BSDL and Grp94-related protein were localizedon the replicates by immunodetection using antibodies to rat BSDL (A) and Grp94 (B), respectively. The positions of molecular mass markersare indicated on the left in kDa.
2673Pancreatic bile salt-dependent lipase secretion and internalization
tseddhe
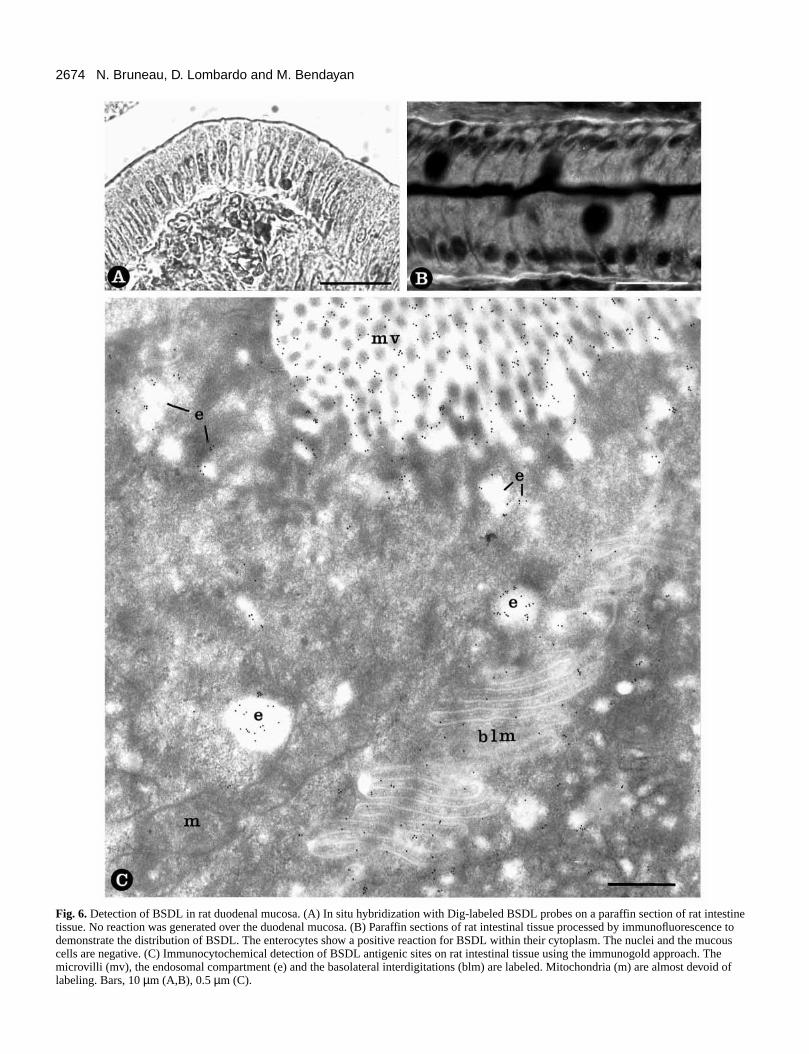
by immunocytochemistry using antibodies to rat pancreaBSDL. Immunofluorescence showed a positive and diffustaining in enterocytes, indicating its presence in these cells (6B). At the electron microscope level, specific immunogo
Fig. 5.Co-detection of BSDL and Grp94-related proteinin pancreatic acinar cells and lumina. (A) Double-immunocytochemical detection of BSDL (IgG-gold 15nm) and Grp94 (IgG-gold 10 nm) in the rat pancreaticacinar lumen (L). Some of the labeling appears to beassociated and is present over the flocculent material ofthe lumen (L) and in the discharging zymogen granule(arrow). Some of the associations are encircled. Bar, 0.5µm. (B) Lane 1, pancreatic juice proteins were separatedby SDS-PAGE and transferred onto nitrocellulosemembrane. The development of the membrane wasperformed with polyclonal antibodies to BSDL using thechemiluminescence western-blotting kit. One majorband was recovered with an apparent mass of 74 kDa.Lane 2, pancreatic juice proteins wereimmunoprecipitated using the monoclonal antibody toGrp94, the antigen-antibody complex was separated bySDS-PAGE and transferred onto a nitrocellulosemembrane. Development of the membrane wasperformed with the polyclonal antibody to BSDL usingthe chemiluminescence western-blotting kit. Again amajor band with an apparent mass of 74 kDa wasrecovered. (C) Lane 1, pancreatic juice proteins wereseparated by SDS-PAGE and transferred ontonitrocellulose membrane. Development of the membranewas performed with the monoclonal antibody to Grp94and anti-rat IgG labeled with alkaline phosphatase. Oneband was recovered with an apparent mass of 180 kDa.Lane 2, pancreatic juice proteins wereimmunoprecipitated using polyclonal antibody to BSDL,the antigen-antibody complex was separated by SDS-PAGE and transferred onto nitrocellulose membrane.Development of the membrane was performed with themonoclonal antibody to Grp94 and antibodies to rat IgGlabeled with alkaline phosphatase. Again one majorband at 180 kDa was recovered.
ticseFig.ld
labeling was found on microvilli and endosomal compartmen(Fig. 6C). These data suggest that pancreatic BSDL, recognizby our antibody, is adsorbed on microvilli membranes aninternalized in the endosomal compartment of the enterocyte. T
2674 N. Bruneau, D. Lombardo and M. Bendayan
Fig. 6.Detection of BSDL in rat duodenal mucosa. (A) In situ hybridization with Dig-labeled BSDL probes on a paraffin section of rat intestinetissue. No reaction was generated over the duodenal mucosa. (B) Paraffin sections of rat intestinal tissue processed by immunofluorescence todemonstrate the distribution of BSDL. The enterocytes show a positive reaction for BSDL within their cytoplasm. The nuclei and the mucouscells are negative. (C) Immunocytochemical detection of BSDL antigenic sites on rat intestinal tissue using the immunogold approach. Themicrovilli (mv), the endosomal compartment (e) and the basolateral interdigitations (blm) are labeled. Mitochondria (m) are almost devoid oflabeling. Bars, 10 µm (A,B), 0.5 µm (C).
2675Pancreatic bile salt-dependent lipase secretion and internalization
al.,
ls.
its
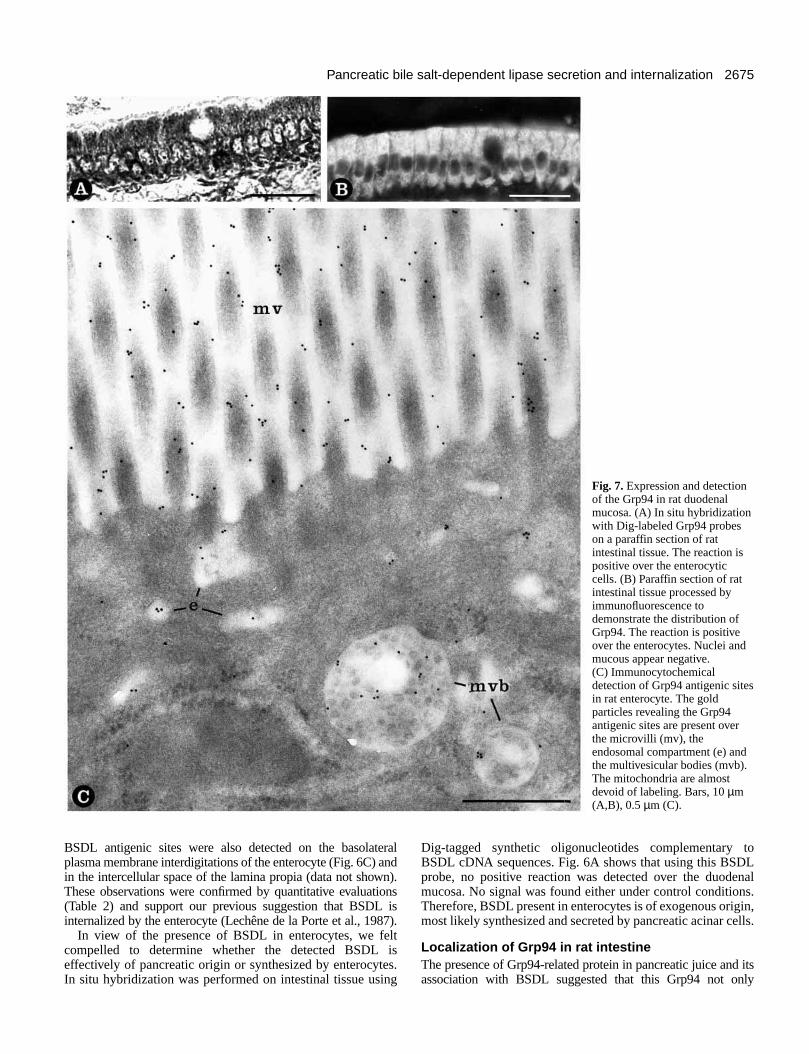
Fig. 7.Expression and detectionof the Grp94 in rat duodenalmucosa. (A) In situ hybridizationwith Dig-labeled Grp94 probeson a paraffin section of ratintestinal tissue. The reaction ispositive over the enterocyticcells. (B) Paraffin section of ratintestinal tissue processed byimmunofluorescence todemonstrate the distribution ofGrp94. The reaction is positiveover the enterocytes. Nuclei andmucous appear negative.(C) Immunocytochemicaldetection of Grp94 antigenic sitesin rat enterocyte. The goldparticles revealing the Grp94antigenic sites are present overthe microvilli (mv), theendosomal compartment (e) andthe multivesicular bodies (mvb).The mitochondria are almostdevoid of labeling. Bars, 10 µm(A,B), 0.5 µm (C).
BSDL antigenic sites were also detected on the basolatplasma membrane interdigitations of the enterocyte (Fig. 6C) in the intercellular space of the lamina propia (data not showThese observations were confirmed by quantitative evaluat(Table 2) and support our previous suggestion that BSDLinternalized by the enterocyte (Lechêne de la Porte et al., 19
In view of the presence of BSDL in enterocytes, we fcompelled to determine whether the detected BSDL effectively of pancreatic origin or synthesized by enterocytIn situ hybridization was performed on intestinal tissue us
eralandn).
ions is87). elt
ises.ing
Dig-tagged synthetic oligonucleotides complementary toBSDL cDNA sequences. Fig. 6A shows that using this BSDLprobe, no positive reaction was detected over the duodenmucosa. No signal was found either under control conditionsTherefore, BSDL present in enterocytes is of exogenous originmost likely synthesized and secreted by pancreatic acinar cel
Localization of Grp94 in rat intestineThe presence of Grp94-related protein in pancreatic juice and association with BSDL suggested that this Grp94 not only
2676
t
eals.
.
N. Bruneau, D. Lombardo and M. Bendayan
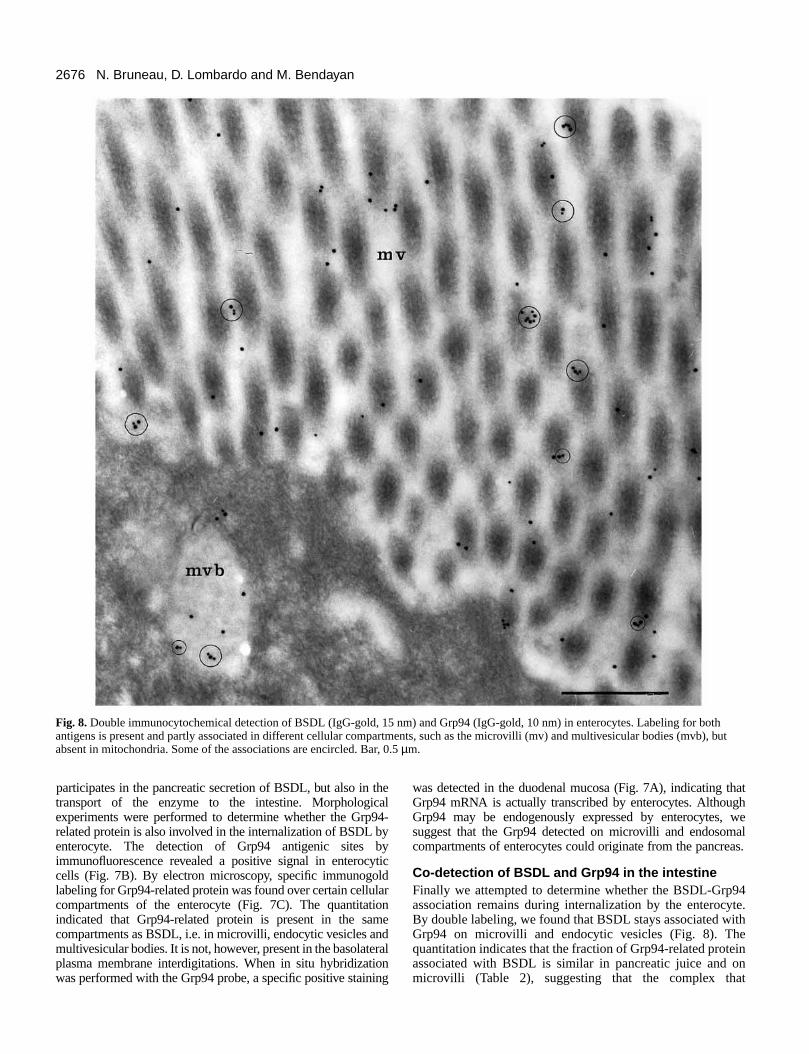
Fig. 8.Double immunocytochemical detection of BSDL (IgG-gold, 15 nm) and Grp94 (IgG-gold, 10 nm) in enterocytes. Labeling for bothantigens is present and partly associated in different cellular compartments, such as the microvilli (mv) and multivesicular bodies (mvb), butabsent in mitochondria. Some of the associations are encircled. Bar, 0.5 µm.
participates in the pancreatic secretion of BSDL, but also in transport of the enzyme to the intestine. Morphologicexperiments were performed to determine whether the Grprelated protein is also involved in the internalization of BSDL enterocyte. The detection of Grp94 antigenic sites immunofluorescence revealed a positive signal in enteroccells (Fig. 7B). By electron microscopy, specific immunogolabeling for Grp94-related protein was found over certain cellucompartments of the enterocyte (Fig. 7C). The quantitatindicated that Grp94-related protein is present in the sacompartments as BSDL, i.e. in microvilli, endocytic vesicles amultivesicular bodies. It is not, however, present in the basolatplasma membrane interdigitations. When in situ hybridizatiwas performed with the Grp94 probe, a specific positive stain
theal94-bybyyticldlarionmenderaloning
was detected in the duodenal mucosa (Fig. 7A), indicating thaGrp94 mRNA is actually transcribed by enterocytes. AlthoughGrp94 may be endogenously expressed by enterocytes, wsuggest that the Grp94 detected on microvilli and endosomcompartments of enterocytes could originate from the pancrea
Co-detection of BSDL and Grp94 in the intestineFinally we attempted to determine whether the BSDL-Grp94association remains during internalization by the enterocyteBy double labeling, we found that BSDL stays associated withGrp94 on microvilli and endocytic vesicles (Fig. 8). Thequantitation indicates that the fraction of Grp94-related proteinassociated with BSDL is similar in pancreatic juice and onmicrovilli (Table 2), suggesting that the complex that

2677Pancreatic bile salt-dependent lipase secretion and internalization
esed,kesle,en
achtoryin
d
thees
). toe
ed94iss-
ofiteseshe’n
ldheld byar
beng
th+ionebeell
ataticoalrentain.alasnd,uiceasce
eshe19,nd
o
Table 2. Densities of labeling obtained with the anti-BSDLand the anti-Grp94 over different cellular compartments
of the enterocyte cellsCellular compartments BSDL Grp94 Association (%)
Microvilli 12.8±1.0* 5.2±0.4* 67.8±4.0Endocytic vesicles 104.4±16.4* 43.8±9.3* 42.0±14.7Multivesicular bodies 25.9±3.5* 4.4±1.2* n.d.Basolateral membrane 3.0±0.3‡ 0.3±0.03‡ n.d.
interdigitationsMitochondria 0.9±0.08* 0.4±0.1* n.d.
% Data show the percentage of association of Grp94 with BSDL.*Gold particles/µm2 (mean values ± s.e.m.).‡Gold particles/µm (mean values ± s.e.m.). n.d., not determined.
encompasses BSDL and Grp94-related protein in the juicepreferentially internalized by the intestinal mucosa and boproteins dissociate in the late endosomal compartment.
DISCUSSION
Previous and present results have established that, as witdigestive enzymes in rat pancreas, the BSDL follows tregulated secretory pathway (RER→Golgi→zymogen granules)originally defined by Palade (1975). Our recent data on AR4pancreatic acinar cells line allowed us to propose a modelthe specific transport of BSDL along the secretory pathw(Abouakil et al., 1993; Bruneau et al., 1995). A particular featuin BSDL secretion by AR4-2J cells is its association wiintracellular membranes. The membrane-associated BSDL isand N-glycosylated and consequently the enzyme shouldassociated with membranes until the trans-Golgi compartment.We have shown that this association involves a multiprotcomplex, including the chaperone Grp94-related protein, whis linked directly to BSDL (Bruneau and Lombardo, 1995). Tchaperone Grp94-related protein seems to play an essentialin the folding and transport of BSDL. Multiple reasons may advocated to explain the particular secretory process of BSThe most likely would reflect the fact that BSDL is a uniqusecretory pancreatic protein with mucin-like, tandem-repeasequences on which O-linked oligosaccharides are clusteRecently, we have shown that the rate of secretion of BSdepends upon the ability of the cell to O-glycosylate the terminal repeats of the protein and that only O-glycosylatBSDL is secreted (Bruneau et al., 1997).
In this study, using morpho-biochemical techniques, whave defined the secretory pathway of pancreatic BSDL,association with Grp94-related protein and its internalizatiby duodenal enterocytes. Morphological data have revealedpresence of both BSDL and the chaperone Grp94-relaprotein, together with a BSDL-Grp94-related protein complein different compartments along the RER→Golgi→zymogengranules secretory pathway (Figs 1-3). However not all tproteins revealed appear to be associated. This could reeither the existence of associated and free BSDL molecuwithin the cells and the pancreatic juice, or an underestimatof the amount of association due to technical limitations of tmorphological approach. These morphological data, togetwith previous results, suggested that post-translatio
isth
h allhe
-2J forayre
th O- be
einichhe rolebeDL.e
tedred.DLC-ed
e itson thetedx,
heflectlesionhehernal
modifications of BSDL are required for the release of thmultiprotein complex from the membrane but not for itdissociation from the chaperone Grp94-related protein. Indewe have shown that the association between both proteins taplace in the RER up to the Golgi and the secretory granuand remains in the pancreatic juice present in the acinar lum(Figs 3, 5). This association appears to increase and to reits highest percentage in the last compartments of the secrepathway (Table 1), suggesting that the Grp94-related protewithin the multiprotein complex is required for the qualitycontrol of BSDL folding in the RER and Golgi apparatus, anmust intervene in the transport of BSDL until the lumen.
Several chaperones have been shown to participate in process of the synthesis and secretion of pancreatic enzym(Le-Gall and Bendayan, 1996, Vélez-Granell et al., 1994Furthermore, some of them were found to be associatedpancreatic secretory proteins, to follow them along thRER→Golgi→granules secretory pathway and to be secretinto the pancreatic juice (Le-Gall and Bendayan, 1996). Grpis a glycoprotein that belongs to the Hsp90 family, which implicated in the transport of important biological moleculewithin the cell (Koyasu et al., 1986) and carries the KDEL Cterminal sequence for the retention of proteins in the lumenthe endoplasmic reticulum (Sorger and Pelham, 1987). Despits KDEL sequence, the protein is secreted. Several hypothecan be put forward to account for the secretion of Grp94 in tpancreatic juice. (1) Secretion could result from an ‘overflowof the KDEL-bearing proteins from the ER cisternae, as showby Hensel et al. (1994) for other chaperones. (2) It woureflect a defect in the sorting mechanism taking place in tER of pancreatic cells (Takemoto et al., 1992. (3) We cousuggest the existence of a homologous p94 protein detectedthe antibody directed against Grp94, and which would not bethe KDEL sequence, or in which this sequence would not accessible to its receptor, allowing this protein to proceed alothe secretory pathway.
Another role ascribed to Grp94 is its ability to associate wia population of immunogenic peptides and to elicit CD8cytotoxic T-cell responses. It has been observed that a fractof Grp94 is localized to the cell surface (Nicchitta, 1998). Thmechanistic basis of this unusual localization remains to determined and the pancreatic secretion of Grp94 may wresult from similarly unusual processing.
By immunolabeling and immunoblotting, we have shown ththe pancreatic Grp94-related protein is secreted in the pancrejuice. One peculiarity of this Grp94-related protein is its twforms: one form of 94 kDa, corresponding to its conventionmolecular mass, and a second displaying an unusual appamass of approximately 180 kDa, compatible with either homodimer of p94 or a dimer containing at least one p94 proteWe have further shown by morphological and biochemictechniques that the main fraction of Grp94-related protein wassociated with the BSDL in the acinar lumen (see Table 1) aas a consequence, could remain as a dimer in pancreatic j(Fig. 5B, lane 2). Grp94-related protein and BSDL behave proteins of high molecular mass on SDS-PAGE in the presenof 8 M urea, which suggests that the formation of dimers involvhydrophobic forces. Hydrodynamic studies of Grp94 allowed tidentification of a discrete domain, defined by residues 676-7which mediates the dimeric assembly of the protein (Wearsch aNichitta, 1996). The hydrophobicity profile of BSDL has als

2678
s or
enetedn
thedes.ionfndiale
sh adstoot
at)heed.
hetehethes.to ishe;de).
dether G.lyutris,as
of
d
lls:ion
N. Bruneau, D. Lombardo and M. Bendayan
allowed for the characterization of a hydrophobic domain locabetween residues 98 and 121 (Sbarra et al., 1998). It is posthat these hydrophobic domains of Grp94 and BSDL are involvin forming the complex, which contains at least one Grp9related protein and one BSDL molecule. Analysis of Grpstructure by hydrodynamic and electron microscopic approacindicated that the molecule is a tri-nodular rod comprised of tsubunits oriented in a direct, antiparallel array (Nichitta, 199The orientation is conferred by a C-terminal oligomerizatiodomain that maintains the N termini at opposing ends of molecule (Nichitta, 1998). The antibody directed against tGrp78, which recognized the KDEL sequence in both the Grpand the Grp94, was used in western blots on rat pancreatic jand no protein was detected (our unpublished observatioTherefore, the Grp94 KDEL sequence is either absent or accessible. The BSDL-Grp94 complexes, which are ndissociated in ER, are thus secreted via the regulated secrepathway. Furthermore, it has recently been demonstrated tonce bound, the peptide-Grp94 complex is SDS-resist(Nichitta, 1998). Our electrophoresis data show that in tpancreatic juice, the BSDL-Grp94 complex is also SDS-resistand involves the hydrophobic interactions.
By a morphological approach, we have shown (Figs 5 andthat the Grp94-related protein remains associated with the BSuntil reaching the microvilli at the level of duodenal enterocyteIt has been shown that biological activities of pancreatic enzymdecrease during transit from the proximal to the distal gut, rates of degradation differing, however, from one enzyme to other. As to the colipase-dependent lipase, its activity decrearapidly in contrast to proteases and amylase, which are relativmore stable. The mechanism by which the colipase-dependlipase activity is destroyed reflects chymotrypsin-catalyzproteolysis. Less than 1% of lipolytic activity secreted into thduodenum reaches the ileum, as compared to 22% for the tryactivity (Holtmann et al., 1997). As with the colipase-dependelipase, the BSDL may be extremely sensitive to proteolysis abe degraded by chymotrypsin. In such a case, the Grp94-relprotein could, by its association with BSDL, protect this enzymfrom degradation in the duodenal lumen and allow for its bindiand internalization by enterocytes.
Once in the vicinity of microvilli and upon binding to thecell surface, the complex is internalized (see Fig. 8 and Ta2) via endocytic vesicles by the enterocytes. This is agreement with previous immunolocalization studies human intestine (Lechêne de la Porte et al., 1987), showingpresence of BSDL in enterocyte. In addition, our in sihybridization has established that the enterocyte is devoidBSDL mRNA and thus unable to synthesize this proteTherefore any BSDL detected in enterocytes is of exogenorigin and must derive from the pancreas.
In the intestinal lumen, the role of BSDL is to catalyze thhydrolysis of cholesterol esters, which cannot be absorbwithout prior hydrolysis to free cholesterol, and to increase tfree-cholesterol concentration gradient between the micellar plasma membrane pools as well as enhancing the passive upof free cholesterol (Huang and Hui, 1990; Shamir et al., 19Howles et al., 1996). Whereas its role within the enterocyremains to be established, BSDL could act by functioning eitas a carrier for cholesterol (Lopez-Candales et al., 1993) oan intracellular esterifying enzyme (Gallo et al., 1984). Howevrecent in vitro studies seem to invalidate this hypothesis (Sha
tedsibleed4-
94heswo8).n
thehe78uicen).notottoryhat,antheant
8)DLs.es
thetheseselyent
ede
psinntnd
atede
ng
blein
on thetu ofin.ous
eedheandtake
95;te
herr aser,mir
et al., 1995). In addition, BSDL taken up by CaCo-2 cells wafound to have a short residence time and is either degradedrapidly secreted (Huang and Hui, 1990).
Recently, it has been shown that both phospholipase A2 andBSDL are required for efficient cholesterol uptake by thintestinal mucosa (Mackay et al., 1997). We suggest that oother secreted pancreatic protein, namely the Grp94-relaprotein, is important in promoting the enterocytic absorptioof BSDL.
Finally one question remains unanswered and concerns mechanism by which BSDL interacts with the membrane anis internalized by the intestinal cells. Both proteins could brecognized by corresponding receptors or binding siteIndeed, Bosner et al. (1988) have demonstrated the interactof BSDL with heparin-like molecules at the surface ointestinal cells. Using a heterologous system (CaCo-2 cells aporcine enzyme), Huang and Hui (1990) showed that the initinteraction of BSDL with these cells was mediated by thbinding of BSDL to low-affinity and high-capacity bindingsites on the cell surface. Heparin, a component of the bruborder membrane, binds pancreatic triglyceride lipase inreceptor-like manner to promote the absorption of fatty aci(Bosner et al., 1989). By analogy, BSDL has been shown possess a functional heparin binding site, which does ninterfere with the BSDL-Grp94 complex (Bruneau andLombardo, 1995). On the other hand, the presence of Grp94the plasma membrane of dentritic cells (Nicchitta, 1998suggests that a receptor to this protein may also exist. Tnature of these sites and/or receptors remains to be determin
In summary, we have demonstrated for the first time that ttransport of BSDL all along the pancreatic secretory rouoccurs via an association with a Grp94-related protein. Tcomplex is secreted in the pancreatic juice and reaches duodenal lumen, where it is internalized by enterocyteFinally, the dissociation of the Grp94-BSDL complex seems take place in the endosomal compartment and the BSDLtransferred to the basolateral membrane of the enterocyte. Tfunction of BSDL within the enterocyte was not elucidatedhowever, we propose that BSDL could act in a concertemechanism with acyl-coA cholesterol acyltransferase in thformation of chylomicrons, as suggested by Gallo et al. (1984
The authors address their special thanks to Dr E Levy (Centrerecherche, Hôpital Sainte Justine, Montréal, Canada) for providing pancreatic juice and Dr I Londoño (Université de Montréal) foconstructive advice. The technical assistance of D. Gingras andMayer as well as the photographic work of J. Léveillé are highappreciated. Dr N. Bruneau is a recipient of a fellowship from InstitNational de la Santé et de la Recherche Médicale (INSERM, PaFrance) and the Medical Research Council of Canada. This work wsupported by grant MT 7284 from the Medical Research Council Canada to M. Bendayan.
REFERENCES
Abouakil, N., Rogalska, E., Bonicel, J. and Lombardo, D.(1988).Purification of pancreatic carboxylic-ester hydrolase by immunoaffinity anits application to the human bile-salt-stimulated lipase. Biochim. Biophys.Acta. 961, 299-308.
Abouakil, N., Mas, E, Bruneau, N., Benajiba, A. and Lombardo, D.(1993).Bile salt-dependent lipase biosynthesis in rat pancreatic AR 4-2J ceessential requirement of linked oligosaccharide for secretion and expressof a fully active enzyme. J. Biol. Chem. 268, 25755-25763.

2679Pancreatic bile salt-dependent lipase secretion and internalization
nd
nt
I.
ol
e:
l
ain
ic
d
e.
.
a
f.
he
ls
10, of
Bendayan, M.(1995). Colloidal gold post-embedding immunocytochemistrProg. Histochem. Cytochem. 29, 1-163.
Bendayan, M., Roth, J., Perrelet, A. and Orci, L.(1980). Quantitativeimmunocytochimical localization of pancreatic secretory proteins subcellular compartments of the rat acinar cell. J. Histochem. Cytochem. 28,149-160.
Bosner, M. S., Gulick, T., Riley, D. J. S., Spilburg, C. A. and Lange, L. G.(1988). Receptor-like function of heparin in the binding and uptake neutral lipids. Proc. Nat. Acad. Sci. USA 85, 7438-7442.
Bosner, M. S., Gulick, T., Riley, D. J. S., Spilburg, C. A. and Lange, L. G.(1989). Heparin-modulated binding of pancreatic lipase and uptakehydrolysed triglycerides in the intestine. J. Biol. Chem. 264, 20261-20264.
Bruneau, N. and Lombardo, D. (1995). Chaperon function of a Grp 94-related protein for folding and transport of the pancreatic bile salt-dependlipase. J. Biol. Chem. 270, 13524-13533.
Bruneau, N., Lechene de la Porte, P., Sbarra, V. and Lombardo, D.(1995).Association of bile salt-dependent lipase to membrane of human pancremicrosomes. Eur. J. Biochem. 233, 209-218.
Bruneau, N., Nganga, A., Fisher, E. and Lombardo, D.(1997). O-Glycosylation of C-terminal tandem repeated sequences regulatessecretion of rat pancreatic bile salt-dependent lipase. J. Biol. Chem. 272,27353-27361.
Burnette, W. N. (1981). ‘Western Blotting’: Electrophoretic transfer oproteins from sodium dodecyl sulfate-polyacrylamide gels to unmodifinitrocellulose an radiographic detection with antibody and radiodinaprotein A. Anal. Biochem. 112, 195-203.
Gallo, L. L. S., Bennett-Clark, S., Myers, S. and Vahouny, G. V.(1984).Cholesterol absorption in rat intestine: role of cholesterol esterase and acoenzyme A: cholesterol acyltransferase. J. Lipid Res. 25, 604-612.
Gingras, D. and Bendayan, M.(1995). Colloidal gold electron microscopicin situ hybridation: combination with immunocytochemistry for the studof insulin and amylase secretion. Cell vision 2, 218-224.
Han, J. H., Stratowa, C. and Rutter, W. J.(1987). Isolation of full-lengthputative rat lysophospholipase cDNA using improved methods for mRNisolation and cDNA cloning. Biochemistry26, 1617-1625.
Hanson, L., Bläckberg, L., Edlund, M., Lundberg, L., Strömqvist, M. andHernell, O. (1993). Recombinant human milk bile salt-stimulated lipase.J.Biol. Chem. 268, 26692-26698.
Hensel, G., Aßmann, and Kern, H. F.(1994). Hormonal regulation of proteindisulfide isomerase and chaperone synthesis in the rat exocrine pancEur. J. Cell Biol. 63, 208-218.
Hernell, O. and Blackberg, L. (1994). Human milk bile salt-stimulatedlipase: Functional and molecular aspects. J. Pediatrics 125, S56-S61
Holtmann, G., Kelly, D. G., Sternby, B. and Dimagno, E.(1997). Survivalof human pancreatic enzymes during small bowel transit: effect of nutriebile acids, and enzymes. Am. J. Physiol. 273, G553-G558.
Howles, P., Carter, C. and Hui, D. Y.(1996). Dietary free and esterifiedcholesterol absorption in cholesterol esterase (bile salt-stimulated lipagene targeted mice. J. Biol. Chem. 271, 7196-7202
Huang, Y. and Hui, D. Y. (1990). Metabolic fate of pancreas derivedcholesterol esterase in intestine: an in vitro study using Caco-2 cells. J. Lipid.Res. 31, 2029-2037.
Kissel, J. A., Fontaine, R. N., Turck, C. W., Brockman, H. L. and Hui, D.Y. (1989). Molecular cloning and expression of cDNA for rat pancreacholesterol esterase. Biochim. Biophys. Acta1006, 227-236.
Koyasu, S., Nishida, E., Kadowaki, T., Matsuzaki, F., Iida, K., Harada, F.,Kasuga, M., Sakai, H. and Yahara, I.(1986). Two mammalian heat shockproteins, HSP90 and HSP100, are actin-binding proteins. Proc. Nat. Acad.Sci. USA 83, 8054-8058
Kuznetsov, G., Chen, L. B. and Nigam, S. K.(1994). Several endoplasmicreticululm stress proteins, including Erp72, interact with thyroglobulduring its maturatoin. J. Biol. Chem. 267, 22990-22995.
Laemmli, U. K. (1970). Cleavage of structural proteins during the assemof the head of bacteriophage T4. Nature 227, 680-685.
Landberg, E., Pahlsson, P., Krotkiewski, H., Stromqvist, M., Hansson, L.and Lundblad, A. (1997). Glycosylation of bile-salt-stimulated lipase fromhuman milk: comparison of native and recombinant forms. Arch. Biochem.Biophys. 344, 94-102.
Lechêne de la Porte, P., Abouakil, N., Lafont, H. and Lombardo, D.(1987).Subcellular localization of cholesterol ester hydrolase in the humintestine. Biochim. Biophys. Acta. 920, 237-246.
Le-Gall, I. and Bendayan, M.(1996). Possible association of chaperonin 6with secretory proteins in pancreatic acinar cells. J. Histochem. Cytochem.44, 743-749.
y.
in
of
of
ent
atic
the
fedted
cyl-
y
A
reas.
nts,
se)
tic
in
bly
an
0
Little, E., Ramakrishman, M., Roy, B., Gazit, G. and Lee, A. S.(1994).The Glucose regulated proteins (Grp78 and Grp94): functions, gene, aapplications. Crit. Rev. Eukaryotic Gene Expression4, 1-19.
Loomes, K. M. (1995). Structural organization of human bile-salt-activatedlipase probed by limited proteolysis and expression of a recombinatruncated variant. Eur. J. Biochem. 230, 607-613.
Lombardo, D., Fauvel, J. and Guy, O.(1980). Studies on the substratespecificity of a carboxyl ester hydrolase from human pancreatic juice. Action on carboxyl esters, glycerides and phospholipids. Biochim. Biophys.Acta 611, 136-146.
Lombardo, D. and Guy, O. (1980). Studies on the substrate specificity of acarboxyl ester hydrolase from human pancreatic juice. II. Action on cholesteresters and lipid-soluble vitamin esters. Biochim. Biophys. Acta 611, 147-155.
Lopez-Candales, A., Bosner, M. S., Spilburg, C. A. and Lange, L. G.(1993). Cholesterol transport function of pancreatic cholesterol esterasdirected sterol uptake and esterification in enterocytes. Biochemistry32,12085-12089
Mackay, K., Starr, J. R., Lawn, R. M. and Ellsworth, J. L. (1997).Phosphatidylcholine hydrolysis is required for pancreatic cholesteroesterase and phospholipase A2 facilitated cholesterol uptake into intestinalCaco-2 cells. J. Biol. Chem. 272, 13380-13389.
Mayer, G. and Bendayan, M.(1997). Biotinyl-tyramide: a novel approachfor electron microscopic immunocytochemistry. J. Histochem. Cytochem.45, 1449-1454.
Mazzerella, R. A. and Green, M.(1987). ERP99, an abundant, conservedglycoprotein of the endoplasmic reticulum, is homologous to the 90 kDheat shock protein (hsp90) and the 94 kDa glucose regulated prote(Grp94). J. Biol. Chem. 262, 8875-8883.
Melnick, J., Dui, J. L. and Argon, Y. (1994). Sequential interation of thechaperones BiP and Grp94 with immunoglobulin chains in the endoplasmreticulum. Nature370, 373-375.
Melnick, J., Ariel, S. and Argon, Y.(1992). The endoplasmic reticulum stressprotein Grp94 in addition to BiP, associates with unassembleimmunoglobulin chains. J. Biol. Chem. 267, 21303-21306.
Myers-Payne, S. C., Hui, D. Y., Brockman, H. L. and Schroeder, F.(1995).Cholesterol esterase: a cholesterol transfer protein. Biochemistry 34, 3942-3947.
Nauseef, W. N., McCormick, S. J. and Clark, R. A.(1995). Calreticulinfunction as a molecular chaperone in the biosynthesis of myeloperoxidasJ. Biol. Chem. 270, 4741-4747.
Nicchitta, C. V. (1998). Biochemical, cell biological and immunological issuessurrounding the endoplasmic reticulum chaperone Grp94 / gp96. Curr. Opin.Immunol. 10, 103-109
Palade G. E.(1975). Intracellular aspects of the process of protein secretionScience. 189, 347-358.
Reue, K., Zambaux, J., Wong, H., Lee, G., Leete, T. H., Ronk, M., Shively,J. E., Sternby, B., Borgström, B., Ameis, D. and Schotz, M. C.(1991).cDNA cloning of carboxyl ester lipase from human pancreas reveals unique proline-rich repeat unit. J. Lipid Res. 32, 267-276.
Rogers, S., Wells, R. and Rechsteiner, M.(1986). Amino acid sequencescommon to rapidly degraded proteins: the PEST hypothesis. Science 234,364-369.
Sbarra, V., Bruneau, N., Mas, E., Hamosh, M., Lombardo, D. andHamosh, P.(1998). Molecular cloning of the bile salt-dependent lipase oferret lactating mammary gland : an overview of functional residuesBiochim. Biophys. Acta(in press).
Shamir, R., Johnson, W. J., Zolfaghari, R., Lee, H. S. and Fisher, E. A.(1995). The role of bile salt-dependent cholesteryl ester hydrolase in tuptake micellar cholesterol by intestinal cells. Biochemistry 34, 6351-6358.
Sorger, P. K. and Pelham, H. R. B.(1987). The glucose regulated proteinGrp94 is related to heat shock protein Hsp90. J. Mol. Biol. 194, 341-344.
Takemoto, H., Yoshimoro, T., Yamamoto, A., Miyata, Y., Yahara, I., Inoue,K. and Tashiro, Y. (1992). Heavy chain binding protein and endoplasmin areexported from the endoplasmic reticulum in rat exocrine pancreatic celsimilar to protein disulfide-isomerase. Arch. Biochem. Biophys. 296, 129-136.
Vélez-Granell, C. S., Arias, A. E., Torres-Ruiz, J. A. and Bendayan, M.(1994). Molecular chaperones in pancreatic tissue: the presence of cpncpn60 and hsp70 in distinct compartments along the secretory pathwaythe acinar cells. J. Cell Sci. 107, 539-549.
Wang, C. S. and Hartsuck, J. A.(1993). Bile salt-activated lipase. A multiplefunction lipolytic enzyme. Biochim. Biophys. Acta. 1166, 1-19.
Wearsch, P. A. and Nicchitta, C. V.(1996). Purification and partial molecularcharacterization of Grp94, an ER resident chaperone. Protein ExpressionPurification 7, 114-121.





























