Embed Size (px)
Citation preview

Vol. 175, No. 12JOURNAL OF BACTERIOLOGY, June 1993, p. 3844-38550021-9193/93/123844-12$02.00/0Copyright X 1993, American Society for Microbiology
Vibrio parahaemolyticus Has a Homolog of the Vibrio choleraetoxRS Operon That Mediates Environmentally Induced Regulation
of the Thermostable Direct Hemolysin GeneZAW LIN,1 KEIKO KUMAGAI,1 KIYOSHI BABA,1 JOHN J. MEKALANOS,2
AND MITSUAKI NISHIBUCHI1*Department ofMicrobiology, Faculty ofMedicine, Kyoto University, Konoe-cho, Yoshida, Sakyo-ku,
Kyoto 606, Japan, 1 and Department ofMicrobiology and Molecular Genetics,Harvard Medical School, Boston, Massachusetts 021152
Received 28 December 1992/Accepted 5 April 1993
In an effort to identify the regulatory gene controlling the expression of the tdh gene, encoding thethermostable direct hemolysin of Vibrio parahaemolyticus, we examined total DNA of AQ3815 (a Kanagawaphenomenon-positive strain) for sequences homologous to that of the toxR gene of Vibrio cholerae. Theextracted DNA gave a weak hybridization signal under reduced-stringency conditions with a toxR-specific DNAprobe. Cloning and sequence analysis of the probe-positive sequence revealed an operon (Vp-toxRS) which washighly similar to the toxRS operon of V. cholerae (Vc-tarRS) (52 and 62% similarities in the two genes,respectively). The deduced amino acid sequences of the Vp-toxRS gene products (Vp-ToxRS) contained regionssimilar to the proposed transmembrane and activity domains of the Vc-toxRS gene products (Vc-ToxRS). Allclinical and environmental strains of V. parahaemolyticus examined possessed the Vp-toxRS genes. In thepresence of Vp-ToxS, Vp-ToxR promoted expression of the tdh2 gene, one of two tdh genes (tdhl and tdh2)carried by Kanagawa phenomenon-positive strains. The DNA sequence located 144 bp upstream of the tdh2coding region was shown to be important for the Vp-ToxR-stimulated expression of the tdh2 gene in anEscherichia coli background. Comparative analysis ofAQ3815 and its isogenic Vp-toxR null mutant gave thefollowing results: (i) Vp-ToxR promoted, in an AQ3815 background, expression of the tdh gene to differentdegrees in various culture media, with KP broth (2% peptone, 0.5% NaCl, 0.03 M KH2PO4, pH 6.2) beingmost effective (12-fold); (ii) the promotion oftdh gene expression in KP broth was at the level of transcription;and (iii) Vp-ToxR was essential for demonstration of enterotoxic activity of AQ3815 in the rabbit ileal loop, amodel previously used to demonstrate thermostable direct hemolysin-mediated enterotoxic activity ofAQ3815.These results demonstrate that Vp-ToxR and Vc-ToxR share a strikingly similar function, i.e., directstimulation at the transcriptional level of the gene encoding a major virulence determinant (enterotoxin) of aVibrio species.
Members of the genus Vibrio are found in aquatic envi-ronments, and some can cause diarrhea or gastroenteritis inhumans (11, 20). Diarrhea caused by Vibrio cholerae serovar01 is attributed to cholera enterotoxin (CT) produced by thisorganism. The level of in vitro CT production varies depend-ing on cultural conditions (5, 15, 48), and this modulationreflects variation in the transcription of the cax operon,which encodes CT (27). The transcription of cax depends ona transmembrane regulatory protein, ToxR, whose activityis apparently modulated by certain cultural conditions (30,33). The activity of ToxR is further enhanced by anothertransmembrane protein, ToxS, and the genes encoding ToxRand ToxS (toxR and toxS) are clustered in an operon (13, 29).In addition, several virulence-associated genes other thanthe cix operon are coordinately regulated by ToxR. Theseinclude the tcp genes, encoding toxin-coregulated pilus (32,47, 51, 55, 56); the acf genes, associated with accessorycolonization factors (46, 47); the ompU and ompT genes,encoding major outer membrane proteins (32, 56); the aldAgene, encoding aldehyde dehydrogenase (45); and severalother ToxR-activated genes of unknown function (46, 47). Ofthe ToxR-regulated genes, only the ctx operon is under the
* Corresponding author.
direct control of ToxR (30), and regulation of the other genesis mediated by a third regulator, ToxT (14, 18).
Vibrioparahaemolyticus can cause gastroenteritis, but notall strains are considered to be pathogenic. While the strainsisolated from clinical cases exhibit hemolytic activity on aspecial blood agar, Wagatsuma agar, most environmentalstrains are nonhemolytic on this agar medium (34, 49). Thus,this hemolytic activity, termed the Kanagawa phenomenon(KP), has been used as a virulence marker of V. parahae-molyticus. The KP is caused by thermostable direct hemo-lysin (TDH) produced by this organism (54). In addition tohemolytic activity, TDH was shown to have various biolog-ical activities, such as enterotoxigenicity, cytotoxicity, car-diotoxicity, lethality to small experimental animals, andability to stimulate vascular permeability (37, 54, 60). Thegene (tdh) encoding TDH was detected in all KP-positivestrains and is duplicated (19, 38, 40). Of the two tdh genecopies (tdhl and tdh2), one gene copy (tdh2) contributesmost to production of TDH (19, 40, 41). Recently, other tdhgenes (tdh3, tdh4, and tdh5) and a tdh-related hemolysingene (trh) have been cloned from rare KP-negative strains (1,40, 42).
Activation of the cax operon by ToxR in V cholerae ismodulated by various environmental factors. A low startingpH of the culture medium, for example, is one of theinducing conditions for ToxR activation of the cax operon
3844
on June 22, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

toxRS OPERON OF VIBRIO PARAHAEMOLYTICUS 3845
(47, 56). The level of TDH production by V. parahaemolyti-cus is also modulated by cultural conditions (7-10, 22), andculture media with a low starting pH stimulate high-levelTDH production (7-9). In addition, as observed with thecloned ctx operon (27), exogenous promoters on cloningvectors were required for expression of the cloned tdh genesat substantial levels in Eschenchia coli (36, 39). Theseobservations prompted us to examine the possibility that theproduction of TDH in V. parahaemolyticus may be undercontrol of a ToxR-like regulator. We report here the discov-ery of homologs of the toxRS genes in V. parahaemolyticus.V. parahaemolyticus ToxRS (Vp-ToxRS) shared expectedstructural similarities with V. cholerae ToxRS (Vc-ToxRS).We demonstrate that in an E. coli background, Vp-ToxRdirectly promotes the expression of the tdh2 gene butactivates the tdhl gene very little. In addition, examinationwith a Vp-toxR-deficient isogenic mutant showed that thedegree of the Vp-ToxR-stimulated increase in tdh geneexpression in V. parahaemolyticus was culture mediumdependent and that the tdh gene expression was stimulatedat the level of transcription. Vp-ToxR was also essential forTDH-mediated enterotoxicity in vivo. These results estab-lish that Vp-ToxR bears functional resemblances to Vc-ToxR.
MATERIALS AND METHODS
Bacterial strains, media, and plasmids. The Vp-toxRSgenes were cloned from a clinical KP-positive strain,AQ3815, of V. parahaemolyticus. V. parahaemolyticusstrains, including AQ3815, and the strains belonging to theother species of the genus Vibno used in this study werelaboratory stock strains described previously (40, 52). E. coliHB101 (39), MV1184 (59), CSR603 (3), SY327 Xpir (32), andSM10 Xpir (32) were described previously. All E. coli strainsexcept E. coli MV1184 were grown with Luria-Bertani (LB)broth or agar medium (28). YT medium (28) was employed topropagate E. coli MV1184 harboring plasmids. V. parahae-molyticus strains were grown with specified media. Whennecessary, the growth medium was supplemented with anappropriate antibiotic(s) to maintain a plasmid or a phage orboth or with 10 mM isopropyl-p-D-thiogalactoside (IPTG) toinduce the tac promoter.The plasmids employed or constructed in this study are
described in Table 1.General genetic techniques. Cellular DNA was extracted
by the method described previously (41). Digestion of DNAwith restriction enzymes, gel electrophoresis, isolation ofDNA fragments from the gel, ligation, and transformationwere performed as described previously (39).
Cloning the Vp-toxRS genes. An EcoRI digest of cellularDNA of strain AQ3815 was size fractionated by agarose gelelectrophoresis, and DNA fragments of approximately 5.6kb were isolated and ligated with EcoRI-cleaved and bacte-rial alkaline phosphatase-treated pBR322. The ligation mix-ture was used to transform E. coli HB101. The transformantsgrown on LB agar containing ampicillin were screened bythe DNA colony blot hybridization method with the Vc-toxRgene probe under reduced-stringency conditions (in solutioncontaining 35% formamide) as described previously (42).The plasmid (pKTN1007) extracted from the probe-positivecolony was confirmed to contain a 5.6-kb EcoRI insert byrestriction analysis.
Nucleotide sequence analysis. Deletions of various sizeswere introduced into the inserts of pKTN111, pKTN112,pKTN1008, pKTN1011, pKTN1012, and pKTN1018 by
the method of Henikoff (17) and transformed into E. coliMV1184, and then single-stranded DNA was prepared by themethod of Vieira and Messing (59). The DNA sequence wasdetermined by the dideoxy chain termination method ofMizusawa et al. (35). The amino acid sequence deduced fromthe nucleotide sequence of the gene was analyzed for hydro-pathic profile by the method of Kyte and Doolittle (24).
Maxicell analysis. Maxicell analysis was performed with E.coli CSR603 as the host strain as described by Boulnois andTimmis (3).DNA probes. To prepare a Vc-toxR-specific gene probe, a
630-bp EcoRI-HpaI fragment internal to the Vc-toxR gene(33) was isolated from pVM7 and used as the probe DNA. Toconstruct a Vp-toxR-specific DNA probe, a 657-bp AccI-PvuII fragment of pKTN1029 which is internal to the Vp-toxR coding region (Fig. 1) was subcloned into the SmaI siteof pUC118 by blunt-end ligation. Digestion of the resultingplasmid, pKTN118, at the sites immediately flanking thecloning sites with BamHI and EcoRI gave a 678-bp DNAfragment. This DNA fragment was isolated and used as theprobe DNA. A Vp-toxS-specific probe DNA was preparedas follows. A 1.0-kb EcoRI-PstI fragment carrying the com-plete Vp-toxS gene was isolated from pKTN111. This frag-ment was digested with AluI and HincII. A 301-bp AluI-HincII fragment internal to the Vp-toxS gene (Fig. 1) wasisolated from the digest and used as the probe DNA. Theprobe DNAs were labeled by the random priming methodwith 32P-labeled dCTP (16).DNA colony blot hybridization test. DNA colony blots
were prepared as described previously (38). The DNAcolony hybridization test with DNA probes was performedunder high-stringency conditions (in solution containing 50%formamide) or under reduced-stringency conditions (in solu-tion containing 35% formamide) as described previously(42).
Southern blot hybridization analysis. Southern blot hybrid-ization with the Vc-toxR-specific gene probe under high-stringency conditions (described above) or under reduced-stringency conditions (described above) was carried out asdescribed previously (40, 42).
Northern (RNA) blot hybridization analysis. RNA wasextracted from the test strain by the method describedpreviously (40) with a minor modification. The modificationemployed was the addition of one operation to removeresidual DNA; i.e., aggregates were removed by brief cen-trifugation at 4°C immediately after addition of ethanol forRNA precipitation. Northern blot hybridization with the tdhgene probe was carried out as described previously (40)except that 4 ,ug rather than 10 ,ug of each sample RNA wasused.
Examination of Vp-ToxR-stimulated expression of the tdhand trh genes in E. coli. E. coli HB101 harboring twocompatible plasmids, a derivative of pACYC184 bearing thetdh or trh gene and a derivative of ptac-85 bearing theVp-toxRS genes under control of the tac promoter, wasgrown to late log phase at 37°C with shaking (200 rpm) in 60ml of LB broth supplemented with chloramphenicol (20,ug/ml) and ampicillin (200 ,ug/ml). The bacterial cells werecollected by centrifugation, washed with LB broth withoutantibiotics, and suspended in 60 ml of fresh LB brothwithout antibiotics. The bacterial suspension was aliquotedinto two 30-ml suspensions, and IPTG was added (10 mMfinal concentration) to one of the suspensions to induce theexpression of the Vp-toxRS genes under control of the tacpromoter (in pKTN1015 or pKTN1031). The two cell sus-pensions in LB broth with and without added IPTG were
VOL. 175, 1993
on June 22, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

TABLE 1. Plasmids used in this study
Plasmid Description Source orPlasmidDescription reference
pBR322 2pUC118 59pUC119 59ptac-85 26pACYC184 6pAT153 58pVM7 30pCVD502 39pCVD503 39pCVD529 40pCVD530 40pCVD532 40pKTN108 3.0-kb HindIII fragment carrying the trh gene cloned from the cellular DNA of V. This study
parahaemolyticus AQ4037 (42) into the HindIII site of pACYC184pKTN111 1.0-kb PvuII fragment of pKTN1012 carrying the Vp-toxS gene cloned into the SmaI site of This study
pUC118 in the orientation opposite to that of the lacZ genepKTN112 1.0-kb PvuII fragment of pKTN1012 carrying the Vp-toxS gene cloned into the SmaI site of This study
pUC118 in the same orientation as that of the IacZ genepKTN116 3.4-kb SalI fragment of pKTN1029 carrying the Vp-taxRS genes cloned into the Sall site of This study
pCVD503pKTN118 657-bp AccI-PvuII fragment, internal to the Vp-toxR gene, of pKTN1029 cloned into the SmaI site This study
of pUC118 by blunt-end ligationpKTN250 3.0-kb NdeI-ClaI fragment of pCVD532 carrying the tdh4 gene cloned into the HinclI site of This study
pUC119 by blunt-end ligationpKTN701 41pKTN1007 5.6-kb EcoRI fragment carrying the Vp-taxRS genes cloned from the cellular DNA of V. This study
parahaemolyticus AQ3815 into the EcoRI site of pBR322pKTN1008 1.5-kb EcoRI-PvuII fragment of pKTN1007 carrying the most Vp-toxR sequence was cloned into This study
the EcoRI-SmaI sites of pUC118pKTN1011 2.8-kb EcoRI-SacI fragment of pKTN1007 carrying the Vp-toxRS genes cloned into the EcoRI-SacI This study
sites of pUC118pKTN1012 2.6-kb BglII-SacI fragment of pKTN1007 carrying the Vp-toxRS genes cloned into the BamiHI-SacI This study
sites of pUC118pKTN1013 2.7-kb HindIII fragment of pCVD529 carrying the tdh2 gene cloned into the HindIII site of This study
pACYC184 in the orientation opposite to that of the tet genepKTN1014 2.7-kb HindIll fragment of pCVD529 carrying the tdh2 gene cloned into the HindIII site of This study
pACYC184 in the same orientation as that of the tet genepKTN1015 2.6-kb BglII-SalI fragment of pKTN1011 carrying the Vp-toxRS genes cloned into the BamHI-SalI This study
sites of ptac-85pKTN1018 1,149-bp EcoRI fragment of pCVD529 carrying the tdh2 gene cloned into the EcoRI site of pUC118 This study
in the orientation opposite to that of the lacZ genepKTN1019 239-bp sequence within pKTN1018 deleted from the PstI site to the nucleotide 69 bp upstream of This study
the tdh2 coding region and religatedapKTN1020 163-bp sequence within pKTN1018 deleted from the PstI site to the nucleotide 145 bp upstream of This study
the tdh2 coding region and religatedapKTN1021 67-bp sequence within pKTN1018 deleted from the PstI site to the nucleotide 241 bp upstream of This study
the tdh2 coding region and religatedapKTN1022 39-bp sequence within pKTN1018 deleted from the PstI site to the nucleotide 269 bp upstream of This study
the tdh2 coding region and religatedapKTN1023 1,051-bp HindIII-PvuII fragment of pKTN1019 carrying the tdh2 gene cloned into the HindIlI- This study'
BamHI (blunted) sites of pACYC184pKTN1024 1,127-bp HindIII-PvuII fragment of pKTN1020 carrying the tdh2 gene cloned into the HindIII- This study
BamHI (blunted) sites of pACYC184pKTN1025 1,223-bp HindIII-PvuII fragment of pKTN1021 carrying the tdh2 gene cloned into the HindIII- This study
BamHI (blunted) sites of pACYC184pKTN1026 1,251-bp HindIII-PvuII fragment of pKTN1022 carrying the tdh2 gene cloned into the HindIII- This study
BamHI (blunted) sites of pACYC184pKTN1029 2.8-kb EcoRI-PstI fragment of pKTN1011 carrying the Vp-toxRS genes cloned into the EcoRI-PstI This study
sites of pAT153pKTN1030 360-bp PvuI-PvuII fragment internal to the Vp-toxR gene was-removed from pKTN1029 and This study
religated by blunt-end ligation (3' extension of the PvuI site removed with T4 DNA polymerase)pKTN1031 2.2-kb BglII-SalI fragment of pKTN1030 cloned into the BamiHI-SalI site of ptac-85 This studypKTN1032 2.4-kb SalI-BamiHI fragment of pCVD502 carrying the tdhl gene cloned into the SalI-BamHI sites This study
of pACYC184 in the same orientation as that of the tet gene by blunt-end ligationpKTN1033 3.0-kb HindIlI fragment of pCVD530 carrying the tdh3 gene cloned into the HindIII site of This study
pACYC184pKTN1034 3.0-kb HindIII-BamHI fragment of pKTN250 carrying the tdh4 gene cloned into the HindIII-BamHI This study
sites of pACYC184pKTN1035 3.0-kb Sall fragment of pKTN1030 carrying the Vp-taxR-S' genes cloned into the Sall site of This study
pKTN701a See "Nucleotide sequence analysis" in Materials and Methods for introduction of the deletion.
3846
on June 22, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

V tcxRS OPERON OF VIBRIO PARAHIAEMOLYTICUS 3847
El BI I
P P Sa Sc HdI I
Vp- toxR Vp-toxS
Ac PFc Au
FIG. 1. Restriction map of the 5.6-kb EcoRI DNA fragmentcloned from V. parahaemolyticus AQ3815. The restriction sites areabbreviated as follows: Ac, AccI; Au, AluI; B, BglII; EI, EcoRI;EV, EcoRV; Hd, HindIII; Hc, HinclI; P, PvuII; Sa, Sacl; Sc, ScaI;and SI, Sall. For the AccI, AluI, and HincII sites, only the sitesneeded to show the relative positions of the probe DNA fragmentsare indicated. The locations and transcriptional directions of theVp-toxRS genes revealed by nucleotide sequence determination areindicated by the arrows. The locations of the AccI-PvuII andHincII-AluI fragments used to prepare probes specific to the Vp-toxR and -toxS genes, respectively, are shown by the bars.
incubated with shaking for 3 h at 370C. A 29-ml portion ofeach culture was used to examine activity of intracellularhemolysin as described previously (39), and a 1-ml portion ofeach culture was used to determine the concentration ofintracellular TDH or TDH-related hemolysin (TRH) as fol-lows. The bacterial cells in the 1-ml portion of each culturewere harvested by centrifugation (13,000 x g), washed oncewith phosphate-buffered saline (3.3 mM NaH2PO4, 6.7 mMNa2HPO4, 0.13 M NaCl, pH 7.2), and suspended in 0.5 ml ofthe same buffer. The bacterial cells were then disrupted bysonication, and 0.5 ml of the same buffer was added to makea final volume of 1 ml. The sonicated cell suspension wascentrifuged, and the supernatant was obtained. The concen-tration of TDH or TRH in the supernatant was determinedby the bead enzyme-linked immunosorbent assay (bead-ELISA). The fold increase in the expression of the tdh or Arhgene was determined by dividing the concentration of TDHor TRH obtained with induction of Vp-toxRS gene expres-sion by that obtained without induction. The experimentswere independently performed three times, and, unlessotherwise indicated, the mean fold increase is reported.Bead-ELISA. The concentration of TDH or TRH in the
supernatant of E. coli cell lysate or in the supernatant of V.parahaemolyticus culture was determined by bead-ELISAwith polyclonal anti-TDH serum or anti-TRH serum asdescribed previously (43, 52).
Construction of an isogenic Vp-toxR mutant of V. para-haemolyficus. The Vp-taxR gene of V. parahaemolyticusAQ3815 was replaced with the mutated Vp-toxiR gene by anallelic exchange method using a suicide vector plasmid(pKTN701) as described previously (41). Briefly, the Vp-taxR-S+ genes (in-frame deletion of a 360-bp sequencewithin the Vp-toxR coding region) flanked by a 0.8-kbupstream sequence and a 1.1-kb downstream sequence ofAQ3815 origin was excised from pKTN1030 and cloned intopKTN701. The resulting plasmid (pKTN1035) carrying achloramphenicol resistance marker was obtained in E. coliSY327 X pir, transformed into E. coli SM1Q X pir, and thenmobilized into V. parahaemolyticus AQ3815 by conjugation.The transconjugant in which pKTN1035 was-integrated into
the Vp-toxRS locus of AQ3815 was selected on TCBS(thiosulfate-citrate-bile-sucrose) agar (Difco Laboratories,Detroit, Mich.) supplemented with chloramphenicol (20 jig/ml). Subsequent excision of the plasmid was detected aftergrowth in LB medium without added chloramphenicol byexamination of subclones for sensitivity to chloramphenicol.Chromosomal DNAs extracted from the chloramphenicol-sensitive subclones were examined by restriction and South-ern blot hybridization (with the Vp-toxR probe) analyses.The subclone which retained the mutated Vp-toxR genebut lost the wild-type Vp-toxR gene was designatedAQ3815AToxR.
Stability of plasmids in AQ3815AToxR. The percent loss ofthe specified plasmid in AQ3815AToxR cells after growth forthe indicated number of generations was determined by themethod described by Schmidhauser and Helinski (50).
Enterotoxicity test. The rabbit ileal loop test was per-formed essentially as described by De and Chatterje (12).Male Japanese white rabbits weighing between 1.7 and 2.0kg were subjected to 24 to 48 h of fasting (with waterprovided ad libitum) prior to the test. The test organism wasgrown in brain heart infusion broth (Difco Laboratories)supplemented with 0.5% NaCl with shaking (200 rpm) for 24h at 37°C. Chloramphenicol (20 jig/ml) was added to thegrowth medium for AQ3815AToxR(pKTN116) for this stepto prevent plasmid segregation. Five microliters of theculture was inoculated into 50 ml of the same medium andincubated with shaking at 37°C to mid-log phase. The con-centration of the bacterial cells was adjusted to 0.8 OD600(optical density at 600 nm) unit (ca. 108 viable cells per ml)with the sterile medium. Then 1 ml of the culture wasinoculated into a 10-cm ligated ileal loop, and the fluidaccumulation (FA) ratio (volume of accumulated fluid [mil-liliters] per centimeter of loops) was measured 18 h later.The loops for a negative control (sterile culture medium) anda positive control (sterile culture medium with 1 ,ug of CT[Choletox purchased from Seikagaku Kogyo Co., Ltd.,Tokyo, Japan] added) were always included. The resultsobtained with the rabbits in which a high FA ratio (>1.0) andany FA ratio (>0) were achieved by inoculation of thepositive control and AQ3815, respectively, were recorded.
Nucleotide sequence accession number. The nucleotidesequence of the Vp-taxRS genes reported in this article hasbeen submitted to GenBank; the accession number isL11929.
RESULTS
Detection of a toxRS homolog in V. parahaemolyticusAQ3815. Cellular DNA extracted from V. parahaemolyticusAQ3815 was digested with various restriction enzymes andexamined by Southern blot hybridization analysis with aVc-toxR-specific gene probe derived from an internal regionof the Vc-toxR coding sequence (33). No hybridization signalwas detected after hybridization under high-stringency con-ditions, but weak hybridization signals could be detectedwhen hybridization was performed under reduced-strin-gency conditions. A 5.6-kb EcoRI fragment exhibiting theweak hybridization signal was cloned into the EcoRI site ofpBR322, resulting in pKTN1007. Restriction and Southernblot hybridization analyses of pKTN1007 revealed that the1.6-kb EcoRI-PvuII subfragment of the cloned EcoRI frag-ment contained the probe-positive sequence (Fig. 1). Thenucleotide sequence within the probe-positive EcoRI-PvuIIregion was determined and analyzed. A sequence homolo-gous to that of the Vc-toxR gene was detected, but the
VOL. 175, 1993
on June 22, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

3848 LIN ET AL.
3'-distal sequence was truncated at the PvuII site. Thus, thenucleotide sequence within a larger probe-positive subfrag-ment (2.8-kb EcoRI-SacI fragment) was determined. Analy-sis of the determined nucleotide sequences revealed a 876-bpopen reading frame which had 52% similarity to the Vc-toxRcoding sequence (Fig. 2, positions 158 through 1033) and a513-bp open reading frame which had 62% similarity to theVc-toxS coding sequence (Fig. 2, positions 1049 through1561). These were named toxRS genes (designated Vp-toxRSgenes in this paper) because of structural and functionalsimilarities to the Vc-toxRS genes (described below).
Analysis of the Vp-toxRS genes and gene products. TheATG initiation codon in the Vc-toxR gene assigned previ-ously (Fig. 2, lower sequence, positions 122 through 124[33]) was absent at the corresponding positions of theVp-toxR gene because of a base change. Therefore, theinitiation codon of the Vp-toxR gene was assigned to theATG 36 bases downstream (Fig. 2, upper sequence, posi-tions 158 through 160). It is possible that the actual initiationcodon of the Vc-toxR gene is also the corresponding ATG atthis position. A sequence homologous to the Shine-Dalgarnosequence, a possible ribosome-binding sequence, was found5 bases upstream of the Vp-toxR coding sequence (Fig. 2,positions 145 through 150), but a sequence homologous tothe consensus sequence of the E. coli promoter (25) couldnot be found within the sequenced 147-bp region upstream ofthe coding sequence. The deduced amino acid sequences ofVp-ToxR and Vc-ToxR had 55% overall similarity (Fig. 3).The region in which the Vc-ToxR sequence had ca. 30%similarity to the amino acid sequences of other bacterialtranscriptional activators, such as OmpR, PhoM, PhoB,VirG, and SfrA (33), is indicated by the underline in Fig. 3.This region was shown to be essential for transcriptional andDNA-binding activities of Vc-ToxR (44). The Vp-ToxRsequence had a very high similarity (80%) to the Vc-ToxRsequence in this region, thereby suggesting that this regionmay be associated with transcriptional activation and DNA-binding activities of Vp-ToxR. The very hydrophobic regionof the Vc-ToxR sequence (indicated by a broken underline inFig. 3) was proposed to be a transmembrane domain ofVc-ToxR (33). This region of the Vp-ToxR sequence wasalso very hydrophobic (data not shown).The initiation codon of the Vp-toxS gene (Fig. 2, positions
1049 through 1051) started downstream of the Vp-toxR genewith an intergenic space of 11 bp, whereas the intergenicspace between the Vc-toxR and -toxS genes was 9 bp (13).As observed with the Vc-toxS gene, a putative ribosome-binding sequence (Fig. 2, positions 1038 through 1043)started 11 bases upstream of the initiation codon of theVp-toxS gene. The Vp-toxS coding sequence was followedby a 23-bp inverted repeat sequence (Fig. 2, positions 1078through 1600), a possible transcription terminator. The de-duced amino acid sequences of Vp-ToxS and Vc-ToxS had64% overall similarity (Fig. 4). The very hydrophobic regionof the N-terminal sequence of Vc-ToxS (indicated by abroken underline in Fig. 4) was presumed to be a transmem-brane domain (13). Hydropathy profiles of the Vp-ToxS andVc-ToxS sequences, including the very hydrophobic regionat the N terminus, were very similar (data not shown).The proteins encoded by the Vp-toxRS genes were ana-
lyzed by the maxicell method (Fig. 5). The Vp-toxRS-bearing sequence derived from the 2.6-kb BglII-SacI region(Fig. 1) was cloned under control of the inducible tacpromoter in an expression vector (ptac-85), resulting inpKTN1015. The profiles of proteins encoded on ptac-85grown with and without induction of the tac promoter are
identical (23). When the tac promoter was uninduced, theprofile of the proteins encoded on pKTN1015 (Fig. 5, lane 2)was identical to that of ptac-85 (Fig. 5, lane 1). When the tacpromoter was induced (Fig. 5, lane 3), however, two addi-tional proteins were detected (indicated by the arrows in Fig.5). The apparent sizes of these proteins were 36 and 22 kDaand were reasonably close to the calculated molecularweights of Vp-ToxR (32,602) and Vp-ToxS (19,200), respec-tively. The results indicated that the Vp-toxRS genes existon the same transcriptional unit and thus constitute anoperon.
Distribution of the Vp-toxRS genes in V. parahaenwlyticusand in the other vibrios. The presence or absence of theVp-toxRS genes in V. parahaemolyticus strains other thanAQ3815 and in the other species of the genus Vibrio wasexamined by the DNA colony hybridization test underhigh-stringency conditions with Vp-toxR- and Vp-toxS-specific gene probes. The results are summarized in Table 2.Both the Vp-toxR and -toxS genes were detected in allstrains of V. parahaemolyticus regardless of the origin ofisolation (clinical or environmental). On the other hand, thestrains belonging to the other Vibio species had neither ofthe two genes. Strains of V. cholerae 01 are expected tocarry Vc-toxR and -toxS genes (29, 31), but they gavenegative results because hybridization was performed underhigh-stringency conditions.
Stimulation of tdh gene expression by Vp-ToxRS in E. coli.The effect of the Vp-toxRS gene products (Vp-ToxRS) onthe expression of various tdh genes and a related hemolysingene (trh) was examined in an E. coli background.The tdh2 gene, responsible for production of most TDH in
KP-positive strains, was examined first. A 2.7-kb HindIIIfragment, cloned from V. parahaemolyticus WP1 (KP-posi-tive strain), containing the tdh2 coding sequence as wellas 1.7-kb upstream and 0.46-kb downstream sequenceswas subcloned into pACYC184, resulting in pKTN1013.pKTN1013 and a compatible plasmid, pKTN1015, bearingthe Vp-toxRS genes under control of the IPTG-inducible tacpromoter were transformed into E. coli HB101. Productionof intracellular TDH from the tdh2 gene was compared forHB101(pKTN1013, pKTN1015) grown in LB broth withoutand with IPTG induction. The lysate of HB101(pKTN1013,pKTN1015) grown with IPTG exhibited much greater hemo-lytic activity than did the culture without IPTG (Fig. 6). Toconfirm that this effect of Vp-ToxRS was due to promotionof tdh2 gene expression rather than to an increase in hemo-lytic activity of the TDH molecule by Vp-ToxRS modifica-tion, the levels of TDH in the lysates were determined by thebead-ELISA method. Three hours following IPTG induc-tion, the level of TDH produced was 5.1-fold higher for theinduced culture than for the uninduced culture. WhenpKTN1015 was replaced by pKTN1031, in which the Vp-toxR gene of pKTN1015 was inactivated by internal in-framedeletion of a 360-bp sequence, no increase in TDH produc-tion was seen for the induced culture relative to the unin-duced culture. These results indicated that Vp-ToxRS stim-ulated expression of the tdh2 gene.By replacing pKTN1013 in HB101(pKTN1013, pKTN-
1015) with an appropriate plasmid, the details of the effect ofVp-ToxR on the expression of the tdh gene were investi-gated. When the orientation of the 2.7-kb tdh2-bearingHindIII insert in pKTN1013 was reversed (pKTN1014), asimilar level (5.3-fold) of Vp-ToxR-dependent increase inTDH production was observed, indicating that stimulation oftdh2 gene expression by Vp-ToxR was mediated through thesequence within the 2.7-kb insert. It was inferred, then, from
J. BACTERIOL.
on June 22, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

VOL. 175, 1993 toxRS OPERON OF VIBRIO PARAHAEMOLYTICUS 3849
rA (.q
CDWCD -<<~~~I
O i..-~ C, Q,"-I > > C - n >4c >'c >c-3 >
=10--4.--P ,t I'M "..'00 C>CM,~-M~3>,CW;..":>0-3>C)nC, >H OC,~~~ >C, CC, C, '''0> CH 0. I H, ,C C ~0>CC >H HH:C,,. H 00 I,>1>, ~ o' ~l 0>H,.~ C,> H OH 00 H CC,1O >HO * v C,>~ OH C,>.,:.H.>.OH.. % C> ,;~ 01 >C,
OH0> ~ ~ ~ >C,. ~~~ ~ >'~~ ~~'0'. ~~ >>. oC,.0>,:'H......~,> >, >, HCC , C, )~~~ C . H 4HO. .,~ > H CH ~ H 00 ~ > O 0 O
HO..0~AH 0 . > . H. O.
0 0 > 0 >0>~OH>0 >H 0> 00' ~ ~ ~ ~-3~)~3 HO 00 >H 00 H0>D >0 0.0 H>00:> L ,0..' I 0
"J0. O >0.0~ ~O ~
0>, H > 0 0, H , , ,I->H H
,0 HO 00 0 OH-OH .a-a. H>_0 0Ha~ >~>~~~(~~~~OH~ ~~~~~~. C~~~~~H >~~~0- > 0> > )..11...
,.
.Z> --tn> mII-1.
CDH i .>HW>Z0 ~ . > 0> 0 H004 0>--
00,~a >H 0.0,:IBN.....
HO, W 0I - H> >0HO~~~~~~~~~~~~ >0~~~~~~.- .,O H. 0 . ~ > ~ r>.,"'0>""..
nj0 >0 0.,>0'0:-C 0'% 00. O H
>.~~~.~~~.>0. 00. C~~~~~~~O HO >0. .~~~~~~.0 F 0~,. HO.- HO'~;Z."' :i ,:
0.0~~~~~~~00H> ~~~~~~~~~ ~~~.0 0~~~~~~0 00 0> ~~~~~0-> 0-0> 0.HH>,~~ 0-3> ~ HO0 '0~ H 00~, HO >0 0,0
0 00 ~Pi,>*0*C-).00 HO1*-i0OH >~0
rAt.HO >H*.I HO 0>0>>000.'.. >0~Ol 0. HO... '.00.HO.nHO.t;i. '. >0. H>
-1COD OH.0.0 OH:3 00 0>:f'-. 0 0
O. ~ 0. C,(. ~* 0 0 0 >O0> HO 00 00 OH~' _a0_0 ~-3 > >O* ~ 0 > H> 0 0> ,. >000.0~~~~0 00 OH 00 00 >0 >0 00 '-~~~~~~~~~-3 ~~- >0 "40'*-, 00,.-"~:,, HO'~O0.0HO 0> 0> 0> OH 0.0 0.0. 0 OPH>.I
'~~~>HO 00 >K~~~~~~~ OH 0.0 ~~~~~3 HO >0 ~~~~~~ ~~-~~'~~3 I 0,0 0~~~~~~0 0000;.,
" 11.01CACD HO." 00 >~ HO >H-.7
OH '0 0'0 OH... OH 0.0O.
>> 0.0 >000;OH >H OH.~ ..'0I OH OH HO OH0W 00 00 O*. ~'3 0~~h) 0 ~ 3O ~00 >0 H 0 i,~O0 0 0 ' ~ >O00)~~~~~~~~)30> 00 >0 HO frf~~~~~~~~~~~~~~~~~~~~ >0 0>~~~~~~ H O O 1 >H~>H.H... Oa---a HO > 0 H
..> 0,.O~ OH0 KH;.00 -3H J-:..3.00 >0 oO: 00: jg. '..: HO, 00.,.3.>r,00,~~~~~~~~~~>>. 00, O>. ~~~~~~~~~~~~~~I-~~~; O>. H>, > HO. ~~~~~~~~~~~~~~~~~. 00.OH;~~~~~...~~~~~~~Q~~~~~~~~~~~~~Q~~~~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ .
T
on June 22, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

3850 LIN ET AL.
40
Vp-ToxR ------------ -TNL'QRNiSfAQQTVc-ToxR MFGLGHNSKEIS*SH- I)WEKK LtT IOKED
80
GN#VV - EM LLT E SESE#II QWe-V ISADgD
160
ICT*%RLSPLSSDSSSI iVEgP*SD#NDA---SAN#-#-t#ARVTV----------# MARE#EAkHDIgQP0S#Nt
TIVRP&LATODAI*EYA-44-g~--- -V--`g4FSSAT#NTj*QPA"NZVANKW;P#L
24.0
GN-R--- ItI*V$ LAV4 LL?*--S6SSFKPLTVVDQ280
EYQN*P-OT#V QCIERO V-K*HAEDS----#AVNN-#N WDLS LdVKKVNEOTGG--
*D&NH-SYSMVt" F*GQgKITLY-PVIV*"--
3312
-#N At-jVCEFIG. 3. Comparison of the Vp-ToxR and Vc-ToxR amino acid
sequences deduced from the nucleotide sequences. The nucleotidesequences are shown in Fig. 2. The amino acid sequences are
aligned for maximum homology. -, missing residue. Identical resi-dues are indicated by the dark background. The underline depictsthe region in which the Vc-ToxR sequence had ca. 30% similarity tothe amino acid sequences of the other bacterial transcriptionalactivators (33). The broken underline indicates the proposed trans-membrane domain of Vc-ToxR (33).
analogy to the activation of the cax operon by Vc-ToxR (33),that Vp-ToxR associates with (probably binds to) the up-stream sequence of the tdh2 gene for stimulation of tdh2gene expression. To examine this possibility, deletion anal-ysis was performed for the tdh2 upstream region; pKTN1013was replaced with various plasmids in which the clonedtdh2-bearing fragments consisted of upstream sequences oflimited lengths (68 to 268 bp). While DNA fragments carry-ing 268 bp (pKTN1026) or 240 bp (pKTN1025) of upstream
40
Vp-ToxS #K---*KV*SAV*AV#I*I'F*G>9 LLV0 N
*QNRH*--*MGI&LL#LtiL#S F#RFW80
QT TV1-ND-*LDDDTVVRDYI
K SL4K#NSR AM- * S 4DIT L
2 0
OAND-f" AQG-OT Tt 'K-REV SOS##S-*V XSDDN&-AMV -MFdEVDI4V
S S A 'L )VN TD#-O&Q LE MW "SW
180
*Kf L*Lf ""'SN A
FIG. 4. Comparison of the Vp-ToxS and Vc-ToxS amino acidsequences deduced from the nucleotide sequences. The nucleotidesequences are shown in Fig. 2. The amino acid sequences arealigned for maximum homology. -, missing residue. Identical resi-dues are indicated by the dark background. The broken underlineindicates the proposed transmembrane domain of Vc-ToxS (13).
1 2 3
97-
66-
45-
31-
2214-
FIG. 5. Maxicell analysis of the proteins encoded on plasmidsptac-85 and pKTN1015. Lysates of E. coli CSR603(ptac-85) grownwithout IPTG induction of the tac promoter (lane 1) and lysates of E.coli CSR603(pKTN1015) grown without (lane 2) or with (lane 3)IPTG induction of tac promoter-controlled Vp-toxRS gene expres-
sion were separated by sodium dodecyl sulfate-polyacrylamide gelelectrophoresis (10 to 20% gradient acrylamide gel). Molecularweights (103) of the marker proteins are given on the left. The arrowsindicate presumed Vp-ToxR (upper) and -ToxS (lower).
sequence gave high levels (8.3- and 8.0-fold, respectively) ofVp-ToxR-dependent activation of TDH production, a 144-bpDNA fragment (pKTN1024) yielded a significantly lower(4.7-fold) increase (P < 0.005). A 68-bp upstream DNAfragment generated by the largest deletion did not containthe -35 region of the presumed tdh2 promoter sequence (40)but did carry an artificially generated promoter sequence atthe upstream cloning site (the -10 region of the tet promotermodified at the HindIll cloning site [53]) in pKTN1023.Possibly because of this promoter, TDH production was
detected with pKTN1023 in the absence of Vp-ToxR stimu-lation. However, no increase (1.0-fold) in the level of TDHproduction was induced by Vp-ToxR stimulation of thepKTN1023 construct. This level of Vp-ToxR stimulationwas significantly lower than that of the 144-bp upstreamsequence (P < 0.005).
TABLE 2. DNA colony hybridization with DNA probes specificto the Vp-toxR and -toxS genesa
Resultb with DNAOrganism No. of probe for:
strainsVp-toxR Vp-toxS
V. parahaemolyticus, clinical strains 107 + +V. parahaemolyticus, environmental 41 + +
strainsV. cholerae, 01 6 - -V. cholerae, non-01 13 - -
Vibrio mimicus 9 - -
Vibrio fluvialis 4 - -
Vibo fumissii 3 - -
a Hybridization was performed under high-stringency conditions.b +, positive; -, negative.
J. BACTERIOL.
on June 22, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

toaxRS OPERON OF VIBRlO PARAHAEMOLYTICUS 3851
pKTN1O13 + pKTN1015 :g(tdh2 +) (Vp-toxR+ S+ ) ° _
pKTN 1013
(tdh2 ')
IPTG IPTG
FIG. 6. Results of hemolysis assay for the lysates ofHB101(pKTN1013, pKTN1015). pKTN1013 carried the tdh2 gene,
and pKTN1015 carried the tac promoter-controlled Vp-toaRSgenes. HB101(pKTN1013) served as a control. The test strains weregrown with or without IPTG induction for the tac promoter.Supernatants of the following cell lysates were placed in the wells ofthe blood agarose plate: HB101(pKTN1013, pKTN1015) grown
without IPTG (top left) or with IPTG (top right) and HB101(pKTN1013) grown without IPTG (bottom left) or with IPTG(bottom right).
The results described above indicated that the nucleotidesequence around the nucleotide 144 bp upstream of the tdh2coding region is important for stimulation of tdh2 gene
expression by Vp-ToxR. Since the tdh genes other than thetdh2 gene and the trh gene have upstream sequences homol-ogous to that of the tdh2 gene with various degrees ofsequence identity (details discussed below), stimulation ofthe expression of these genes by Vp-ToxR was examined.For this purpose, various tdh genes and the trh gene withsufficiently long upstream sequences (280 bp to 2.3 kb) werecloned into pACYC184, all in the same orientation, and wereemployed in place of pKTN1013. In these constructs, thereis no known intact promoter in the immediate upstreamregion of the cloning site (pKTN1033, pKTN1034, andpKTN108). Also, the influence of the upstream promoter(P2) in the vector (53) was shown to be negligible(pKTN1032) by changing the orientation of the insert (36).The expression levels of these cloned genes were monitoredby the intracellular concentrations of the gene products(TDH or TRH) (Table 3). Expression of the tdhl gene waspromoted very little (1.3-fold) by Vp-ToxR, but the expres-sion of the tdh3 and tdh4 genes (40) was increased by factors
TABLE 4. TDH production by AQ3815 and AQ3815AToxR invarious culture mediaa
TDH (ng/ml) produced& by: TDHMedium
AQ3815 AQ3815A&ToxR ratio
LB broth (1.0% NaCi, 150 35 4.3pH 7.0)
Modified LB brothd0.5% NaCi 350 60 5.83.0% NaCI 65 23 2.8pH 6.2 150 25 6.0pH 8.5 225 40 5.6
KP brothe 1,200 100 12
a 24-h incubation with shaking (200 rpm) at 37C.b The TDH concentration in the culture supernatant was determined by the
bead-ELISA method (43).c The TDH concentration for AQ3815 was divided by that for
AQ3815AToxR.d The NaCi concentration or pH of LB broth was modified as indicated.Composed of 2% peptone, 0.5% NaCl, and 0.03 M KH2PO4 (pH 6.2) (41).
of 3.9 and 11, respectively. However, probably because ofdifferences in the basal-level expression from their nativepromoters, the levels of TDH produced from these threeVp-ToxR-stimulated tdh genes were still lower than thatfrom the tdh2 gene without Vp-ToxR stimulation (Table 3).Vp-ToxR did not promote the expression of the trh gene.
Effect of a Vp-toxR null mutation in V. parahaemolyticusAQ3815. The 360-bp in-frame Vp-toxR deletion mutationcarried by pKTN1031 was introduced into the Vp-toxR geneof V. parahaemolyticus AQ3815 by a suicide vector-medi-ated allelic exchange method (see Materials and Methods).The resulting isogenic Vp-toxR mutant (AQ3815AToxR) andthe wild-type parent strain (AQ3815) were compared invarious assays to see the effects of this Vp-toxR null muta-tion in a V. parahaemolyticus background.
First, involvement of Vp-ToxR in regulation of tdh geneexpression was assessed in in vitro assays. The levels ofextracellular TDH produced by AQ3815 and AQ3815A&ToxRin various culture media were measured. The results werecompared only on the basis of the ratio of AQ3815 toAQ3815AToxR because of growth rate differences in variousculture media (Table 4). The results confirmed Vp-ToxRmediation of tdh regulation in V. parahaemolyticus. Com-parison of the results obtained with LB broth-based mediawith various NaCl concentrations and pHs suggests that lowand high pHs (6.2 and 8.5) and a low NaCl concentration(0.5%) induce relatively high Vp-ToxR activity (5.6- to6.0-fold difference in TDH). KP broth, which is character-
TABLE 3. Effect of Vp-ToxR on the expression of various tdh genes and the trh gene in E. coli HB101'
Concn (ng/ml) of intracellular TDH or TRHwithout (-) or with (+) induction of Vp-taxRS Increase (fold) in tdh or trh
Plasmids harbored by HB1O1" expression in pKTN1015 expression induced byVp-ToxRc
pKTN1013 (tdh2+), pKTN1015 55.3 ± 3.51 281 ± 13.6 5.07 ± 0.17pKTN1032 (tdhl+), pKTN1015 2.17 ± 0.29 2.71 ± 0.47 1.25 ± 0.05pKTN1033 (tdh3+), pKTN1015 2.43 ± 0.12 9.50 ± 0.50 3.90 ± 0.10pKTN1034 (tdh4+), pKTN1015 2.60 ± 0.36 28.5 ± 3.12 11.0 ± 0.32pKTN108 (trh+), pKTN1015 8.15 ± 0.15 7.95 ± 0.15 0.98 ± 0.00
a Values are expressed as the means + standard errors of the mean (n = 3).b The relevant characteristic of the indicated plasmid (kind of tdh gene or trh gene) is indicated in parentheses.cThe concentration of TDH or TRH obtained with induction of the Vp-toxRS genes was divided by that obtained without induction. The value was obtained
for each set of independent experiments, and the mean ± standard error of the mean was then obtained.
VOL. 175, 1993
on June 22, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
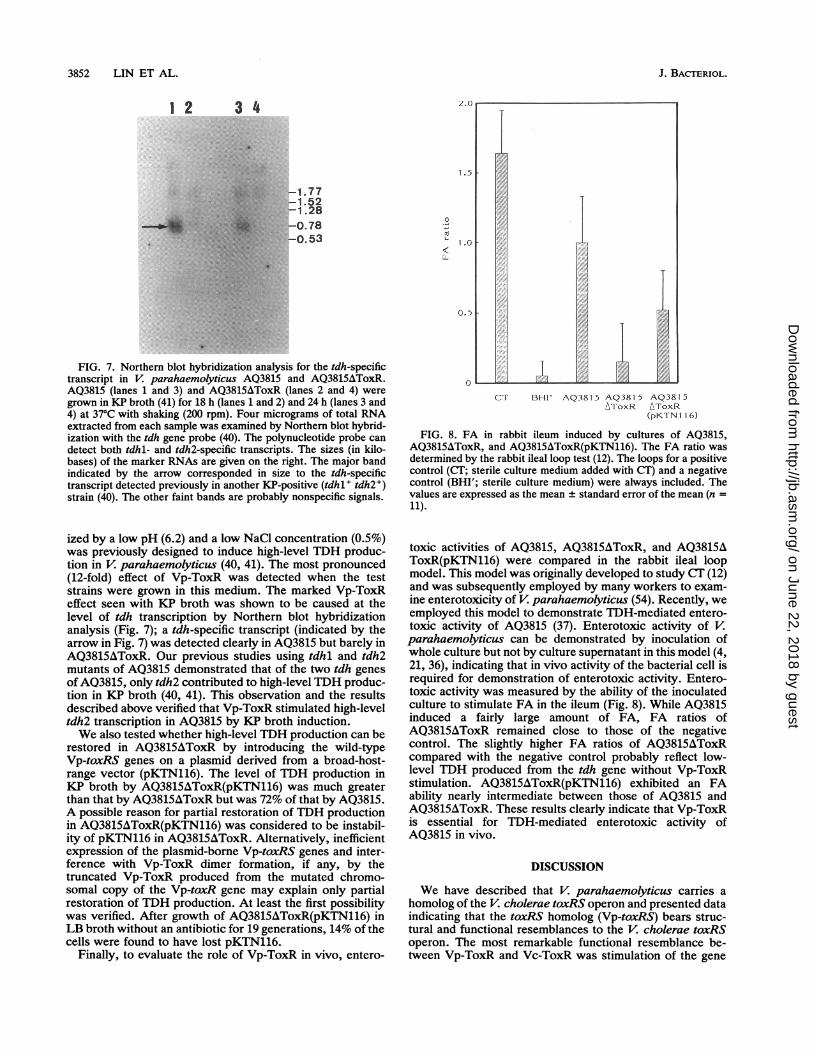
3852 LIN ET AL.
I a
-1.77-1.52-1.28-0.78-0.53
0
I
FIG. 7. Northern blot hybridization analysis for the tdh-specifictranscript in V. parahaemolyticus AQ3815 and AQ3815AToxR.AQ3815 (lanes 1 and 3) and AQ3815AToxR (lanes 2 and 4) weregrown in KP broth (41) for 18 h (lanes 1 and 2) and 24 h (lanes 3 and4) at 370C with shaking (200 rpm). Four micrograms of total RNAextracted from each sample was examined by Northern blot hybrid-ization with the tdh gene probe (40). The polynucleotide probe candetect both tdhl- and tdh2-specific transcripts. The sizes (in kilo-bases) of the marker RNAs are given on the right. The major bandindicated by the arrow corresponded in size to the tdh-specifictranscript detected previously in another KP-positive (tdhl tdh2+)strain (40). The other faint bands are probably nonspecific signals.
ized by a low pH (6.2) and a low NaCl concentration (0.5%)was previously designed to induce high-level TDH produc-tion in V. parahaemolyticus (40, 41). The most pronounced(12-fold) effect of Vp-ToxR was detected when the teststrains were grown in this medium. The marked Vp-ToxReffect seen with KP broth was shown to be caused at thelevel of tdh transcription by Northern blot hybridizationanalysis (Fig. 7); a tdh-specific transcript (indicated by thearrow in Fig. 7) was detected clearly in AQ3815 but barely inAQ3815AToxR. Our previous studies using tdhl and tdh2mutants of AQ3815 demonstrated that of the two tdh genesof AQ3815, only tdh2 contributed to high-level TDH produc-tion in KP broth (40, 41). This observation and the resultsdescribed above verified that Vp-ToxR stimulated high-leveltdh2 transcription in AQ3815 by KP broth induction.We also tested whether high-level TDH production can be
restored in AQ3815AToxR by introducing the wild-typeVp-toxRS genes on a plasmid derived from a broad-host-range vector (pKTN116). The level of TDH production inKP broth by AQ3815AToxR(pKTN116) was much greaterthan that by AQ3815AToxR but was 72% of that by AQ3815.A possible reason for partial restoration of TDH productionin AQ3815AToxR(pKTN116) was considered to be instabil-ity of pKTN116 in AQ3815AToxR. Alternatively, inefficientexpression of the plasmid-borne Vp-toxRS genes and inter-ference with Vp-ToxR dimer formation, if any, by thetruncated Vp-ToxR produced from the mutated chromo-somal copy of the Vp-toxR gene may explain only partialrestoration of TDH production. At least the first possibilitywas verified. After growth of AQ3815AToxR(pKTN116) inLB broth without an antibiotic for 19 generations, 14% of thecells were found to have lost pKTN116.
Finally, to evaluate the role of Vp-ToxR in vivo, entero-
0
CT BHI' AQ3815 AQ3815 AQ3815AToxR AToxR
(pKTN 1 16)
FIG. 8. FA in rabbit ileum induced by cultures of AQ3815,AQ3815A&ToxR, and AQ3815AToxR(pKTN116). The FA ratio wasdetermined by the rabbit ileal loop test (12). The loops for a positivecontrol (CT; sterile culture medium added with CT) and a negativecontrol (BHI'; sterile culture medium) were always included. Thevalues are expressed as the mean + standard error of the mean (n =11).
toxic activities of AQ3815, AQ3815AToxR, and AQ3815AToxR(pKTN116) were compared in the rabbit ileal loopmodel. This model was originally developed to study CT (12)and was subsequently employed by many workers to exam-ine enterotoxicity of V. parahaemolyticus (54). Recently, weemployed this model to demonstrate TDH-mediated entero-toxic activity of AQ3815 (37). Enterotoxic activity of V.parahaemolyticus can be demonstrated by inoculation ofwhole culture but not by culture supernatant in this model (4,21, 36), indicating that in vivo activity of the bacterial cell isrequired for demonstration of enterotoxic activity. Entero-toxic activity was measured by the ability of the inoculatedculture to stimulate FA in the ileum (Fig. 8). While AQ3815induced a fairly large amount of FA, FA ratios ofAQ3815AToxR remained close to those of the negativecontrol. The slightly higher FA ratios of AQ3815AToxRcompared with the negative control probably reflect low-level TDH produced from the tdh gene without Vp-ToxRstimulation. AQ3815AToxR(pKTN116) exhibited an FAability nearly intermediate between those of AQ3815 andAQ3815AToxR. These results clearly indicate that Vp-ToxRis essential for TDH-mediated enterotoxic activity ofAQ3815 in vivo.
DISCUSSION
We have described that V. parahaemolyticus carries ahomolog of the V. cholerae toxRS operon and presented dataindicating that the toxRS homolog (Vp-toxRS) bears struc-tural and functional resemblances to the V. cholerae toxRSoperon. The most remarkable functional resemblance be-tween Vp-ToxR and Vc-ToxR was stimulation of the gene
J. BACTERIOL.
on June 22, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

toxRS OPERON OF VIBRIO PARAHAEMOLYTICUS 3853
encoding a major virulence determinant. In an E. colibackground, Vp-ToxR was shown to directly promote ex-pression of the tdh2 gene. This resembles activation of thectx operon by Vc-ToxR. The Vc-ToxR activation requiredbinding of Vc-ToxR to tandem repeats of IFITI7GAT presentupstream of the cax promoter (33). The presumed promotersequence of the tdh2 gene was located within the 76-bpsequence upstream of the coding region (40). Deletion anal-ysis of the tdh2 upstream sequence indicated that the se-quence around the nucleotide 144 bp upstream of the codingregion is important for Vp-ToxR-dependent expression ofthe tdh2 gene. Previous comparative analysis of various tdhgenes cloned from KP-positive and -negative strains of V.parahaemolyticus and rare strains belonging to the otherVibrio species revealed an interesting feature of the tdhupstream sequence: the tdh genes analyzed so far had fairlyconserved upstream sequences as far as the nucleotides 134to 153 bp upstream of the coding region, but the sequencesfurther upstream lacked homology (57). This was becausethe insertion sequence-like elements (collectively namedISVs) or related sequences derived from genetic rearrange-ment of ISVs were inserted at positions 134 to 153 bpupstream in various isolates (57). Expression of the tdhl,tdh2, tdh3, and tdh4 genes was promoted by Vp-ToxR invarious degrees (1.25 to 11.3-fold), but that of the trh gene,which has only 52% overall similarity to the tdh2 upstreamsequence within the 200-bp upstream region (36, 42), was notpromoted (Table 3). The results also indicate that the Vp-ToxR stimulation of tdh gene expression is dependent onupstream sequence and that the region around the 144-bpupstream position is particularly important. From analogy toclx activation by Vc-ToxR (33), it was inferred that thisupstream region may be the Vp-ToxR-binding site. How-ever, a sequence homologous to the T'TlGAT repeats wasnot present around this region (39, 57). Given that the 48%nucleotide sequence divergence observed between the Vp-and Vc-toxR genes represents the phylogenetic distance ofthe two genes, the binding sequences of the two ToxRs maydiffer considerably. Identification of the exact Vp-ToxR-binding site would make this point clear. Attempts to dem-onstrate Vp-ToxR binding to the presumed binding region inthe presence of Vp-ToxS have been made, but successfulresults have not been obtained repeatedly. This was consid-ered to be due partly to instability of Vp-ToxRS in E. coli(36). To demonstrate binding ability of Vc-ToxR, stabiliza-tion of Vc-ToxR either by utilizing a Vc-toxR::TnphoAfusion gene or by addition of Vc-ToxS was needed (33, 44).We are currently investigating the possibility that aVp-toxR::TnphoA fusion may help stabilize Vp-ToxR in E.coli.Vc-ToxR is a global regulator coordinately regulating
virulence genes and associated genes (30, 32, 33, 45-47, 51,55, 56), and the Vc-toxRS genes are distributed in almost allstrains of V. cholerae (29, 30). Not only TDH production butalso protein profiles of AQ3815 and AQ3815AToxR weresignificantly different when the strains were grown in certainculture media, indicating that Vp-ToxRS is involved inregulation of various other genes (36). We are currentlyinvestigating what other genes are under Vp-ToxR control.This observation and the result that Vp-toxRS genes werefound in all strains of V. parahaemolyticus regardless of theorigin of isolation (Table 2) suggest that Vp-ToxR may alsobe a global regulator important for V. parahaemolyticus. TheVp-toxRS operon may phylogenetically precede the tdh genein V. parahaemolyticus. A previous study (57) presentedevidence that the tdh gene originated in an organism other
than V. parahaemolyticus and was transferred as a transpo-son-like unit flanked by the ISVs. This could explain theobservation that the tdh gene was found in only a smallproportion of the environmental strains of V. parahaemolyti-cus (38). Therefore, promotion of tdh gene expression byVp-ToxR may be a fortuitous event from a phylogeneticstandpoint. Of the duplicated tdh genes (tdhl and tdh2) inKP-positive strains, only the tdh2 gene may have acquiredthe upstream sequence containing a relatively strong pro-moter and a region suitable for interaction with Vp-ToxR(Table 3). This could have resulted in high-level TDHproduction by the tdh2 gene and thus in derivation ofKP-positive strains (40, 41).
In a previous study (37), we compared AQ3815 and itsisogenic tdhl tdh2 mutant strain and showed that productionof TDH is essential for demonstration of enterotoxicity ofAQ3815 in vivo (rabbit ileal loop model). We have shown inthis study that Vp-ToxRS is required for high-level TDHproduction by AQ3815 in KP broth (Table 4) and for dem-onstration of AQ3815 enterotoxicity in the rabbit ileal loopmodel (Fig. 8). These results establish that Vp-ToxR plays apivotal role in demonstration of enterotoxicity by mediatingTDH production in KP-positive V. parahaemolyticus. How-ever, this role of Vp-ToxR may not be simply applied toexplain virulence regulation in rare KP-negative strainsharboring the tdh or trh gene. The tdh3, tdh4, and trh geneswere present in KP-negative strains, but epidemiologicalinvestigations suggested clinical significance for these straips(23, 38, 52). In an E. coli background, Vp-ToxR promotedexpression of the tdh3 and tdh4 genes. However, probablybecause of a difference in the strength of the native pro-moter, the levels of TDH produced from these Vp-ToxR-stimulated genes were less than that from the Vp-ToxR-unstimulated tdh2 gene (Table 3). trh gene expression wasnot promoted by Vp-ToxR in an E. coli background (Table3). The seemingly less remarkable effect of Vp-ToxR on thetrh gene needs to be explained if it is true that Vp-ToxR andthe trh gene are essential for virulence. It may be possiblethat Vp-ToxR activates the trh gene best in a V. parahae-molyticus background where Vp-ToxR stimulation of thisgene may be mediated through a third regulator such asToxT (14). Alternatively, Vp-ToxR may activate the trh genein response to an unknown in vivo signal(s). It is alsopossible that the trh gene is under control of a regulator thatis related, but not identical, to Vp-ToxR. Further investiga-tion along this line is required to elucidate the association ofVp-ToxR with virulence regulation in rare KP-negativestrains possessing the tdh or trh gene.Vp-ToxR mediated environmentally regulated expression
of the tdh gene in V. parahaemolyticus; tdh2 expression wasstimulated by Vp-ToxR to various degrees in various culturemedia, with KP broth being most effective (Table 4). Theexact signal(s) required for Vp-ToxR-mediated tdh regula-tion is unclear, but our preliminary data suggest that Vp-ToxR probably mediates the response to various environ-mental signals rather than to a single signal. It is unknownwhether Vp-ToxR itself has the ability to sense environmen-tal signals. It is possible that the expression of the Vp-toxRSoperon may be regulated by environmental signals and thatregulation of the tdh gene may depend at least in part on theconcentration of Vp-ToxR. To examine this possibility andto better define environmental signals regulating Vp-ToxR-mediated tdh stimulation, we are conducting a study inwhich expression of the tdh and Vp-tocxR genes fused to areporter gene is monitored. Expression of these genes wouldbe monitored more accurately by this method than by
VOL. 175, 1993
on June 22, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

3854 LIN ET AL.
quantitation of TDH and Vp-ToxR, because these proteinsappear to be potentially unstable (36).
In conclusion, the data presented in this report stronglysuggest that the Vp- and Vc-toxRS operons were derivedfrom a common ancestor and evolved by base changes ineach genetic background but that the sequences encodingfunctional domains have been conserved in both operons. Asa result, both operons currently encode global regulators. Astriking functional similarity between the two regulators isthat they directly stimulate transcription of the gene(s)encoding a major virulence determinant (ctx or tdh2), leadingto expression of pathogenicity. Phylogenetically, the tdh2gene may have been brought under control of Vp-ToxR bychance in V. parahaemolyticus and resulted in the derivationof KP-positive, virulent strains. Identification of the othergenes under control of Vp-ToxR will help establish whetherother virulence-associated genes are coordinately regulatedby Vp-ToxR and whether coordinate regulation of thesegenes is the major role played by Vp-ToxR in the absence ofthe tdh2 gene.
ACKNOWLEDGMENTS
We are grateful to Herbert P. Schweizer for providing a plasmid.This work was supported by a Grant-in-Aid for Scientific Re-
search from the Ministry of Education, Science and Culture of Japanand by the U.S.-Japan Cooperative Medical Science Program,Cholera and Related Diarrheal Disease Panel.
REFERENCES1. Baba, K., H. Shirai, A. Terai, Y. Takeda, and M. Nishibuchi.
1991. Analysis of the tdh gene cloned from a tdh gene- and trhgene-positive strain of Vibrio parahaemolyticus. Microbiol.Immunol. 35:253-258.
2. Bolivar, F., R. L. Rodriguez, P. J. Greene, M. C. Betlach, H. L.Heyneker, and H. W. Boyer. 1977. Construction and character-ization of new cloning vehicles. II. A multipurpose cloningsystem. Gene 2:95-113.
3. Boulnois, G. J., and K. N. Timmis. 1984. Synthesis of plasmid-encoded polypeptides in maxicells, p. 204-211. In A. Puhler andK. N. Timmis (ed.), Advanced molecular genetics. Springer-Verlag, Berlin.
4. Brown, D. F., P. L. Spaulding, and R. M. Twedt. 1977. Entero-pathogenicity of Vibrio parahaemolyticus in the ligated rabbitileum. Appl. Environ. Microbiol. 33:10-14.
5. Callahan, L. T., and S. H. Richardson. 1973. Biochemistry ofVibrio cholerae virulence. III. Nutritional requirements fortoxin production and the effects of pH on toxin elaboration inchemically defined media. Infect. Immun. 7:567-572.
6. Chang, A. C. Y., and S. N. Cohen. 1978. Construction andcharacterization of amplifiable multicopy DNA cloning vehiclesderived from the piSA cryptic miniplasmid. J. Bacteriol. 134:1141-1156.
7. Cherwonogrodzky, J. W., and A. G. Clark. 1981. Effect of pHon the production of the Kanagawa hemolysin by Vibrio para-haemolyticus. Infect. Immun. 34:115-119.
8. Cherwonogrodzky, J. W., and A. G. Clark. 1982. Production ofthe Kanagawa hemolysin by Vibrio parahaemolyticus in asynthetic medium. Infect. Immun. 37:60-63.
9. Cherwonogrodzky, J. W., M. A. Skinner, and A. G. Clark. 1984.Effect of D-tryptophan on hemolysin production in Vibriopara-haemolyticus. J. Clin. Microbiol. 20:909-911.
10. Chun, D., J. K. Chung, R. Tak, and S. Y. Seol. 1975. Nature ofthe Kanagawa phenomenon of Vibrioparahaemolyticus. Infect.Immun. 12:81-87.
11. Colwell, R. R. (ed.). 1984. Vibrios in the environment. JohnWiley & Sons, Inc., New York.
12. De, S. N., and D. N. Chatterje. 1953. An experimental study ofthe mechanism of action of Vibrio cholerae on the intestinalmucous membrane. J. Pathol. Bacteriol. 66:559-562.
13. DiRita, V. J., and J. J. Mekalanos. 1991. Periplasmic interaction
between two membrane regulatory proteins, ToxR and ToxS,results in signal transduction and transcriptional activation. Cell64:29-37.
14. DiRita, V. J., C. Parsot, G. Jander, and J. J. Mekalanos. 1991.Regulatory cascade controls virulence in Vibrio cholerae. Proc.Natl. Acad. Sci. USA 88:5403-5407.
15. Evans, D. J., Jr., and S. H. Richardson. 1968. In vitro produc-tion of choleragen and vascular permeability factor by Vibriocholerae. J. Bacteriol. 96:126-130.
16. Feinberg, A. P., and B. Vogelstein. 1983. A technique forradiolabelling DNA restriction endonuclease fragments to highspecific activity. Anal. Biochem. 132:6-13.
17. Henikoff, S. 1984. Unidirectional digestion with exonuclease IIIcreates targeted breakpoints for DNA sequencing. Gene 28:351-359.
18. Higgins, D. E., E. Nazareno, and V. J. DiRita. 1992. Thevirulence gene activator ToxT from Vibno cholerae is a memberof the AraC family of transcriptional activators. J. Bacteriol.174:6974-6980.
19. lida, T., and K. Yamamoto. 1990. Cloning and expression of twogenes encoding highly homologous hemolysins from a Kana-gawa-phenomenon-positive Vibrio parahaemolyticus T4750strain. Gene 93:9-15.
20. Janda, J. M., C. Powers, R. G. Bryant, and S. L. Abbott. 1988.Current perspectives on the epidemiology and pathogenesis ofclinically significant Vibrio spp. Clin. Microbiol. Rev. 1:245-267.
21. Johnson, D. E., and F. M. Calia. 1976. False-positive rabbit ilealloop reactions attributed to Vibrio parahaemolyticus brothfiltrates. J. Infect. Dis. 133:436-440.
22. Karunasagar, I. 1981. Production of hemolysin by Vibio para-haemolyticus in a chemically defined medium. Appl. Environ.Microbiol. 41:1274-1275.
23. Kishishita, M., N. Matsuoka, K. Kumagai, S. Yamasaki, Y.Takeda, and M. Nishibuchi. 1992. Sequence variation in thethermostable direct hemolysin-related hemolysin (trh) gene ofVibrio parahaemolyticus. Appl. Environ. Microbiol. 58:2449-2457.
24. Kyte, J., and R. F. Doolittle. 1982. A simple method fordisplaying the hydropathic character of a protein. J. Mol. Biol.157:105-132.
25. Lewin, B. 1983. Genes, p. 193-195. John Wiley & Sons, Inc.,New York.
26. Marsh, P. 1986. ptac-85, an E. coli vector for expression ofnon-fusion proteins. Nucleic Acids Res. 14:3603.
27. Mekalanos, J. J., D. J. Swartz, and G. D. N. Pearson. 1983.Cholera toxin genes: nucleotide sequence, deletion analysis andvaccine development. Nature (London) 306:551-557.
28. Miller, J. H. 1972. Experiments in molecular genetics. ColdSpring Harbor Laboratory, Cold Spring Harbor, N.Y.
29. Miller, V. L., V. J. DiRita, and J. J. Mekalanos. 1989. Identifi-cation of toxS, a regulatory gene whose product enhancesToxR-mediated activation of the cholera toxin promoter. J.Bacteriol. 171:1288-1293.
30. Miller, V. L., and J. J. Mekalanos. 1984. Synthesis of choleratoxin is positively regulated at the transcriptional level by toxR.Proc. Natl. Acad. Sci. USA 81:3471-3475.
31. Miller, V. L., and J. J. Mekalanos. 1985. Genetic analysis of thecholera toxin-positive regulatory gene toxR. J. Bacteriol. 163:580-585.
32. Miller, V. L., and J. J. Mekalanos. 1988. A novel suicide vectorand its use in construction of insertion mutations: osmoregula-tion of outer membrane proteins and virulence determinants inVibio cholerae requires toxR. J. Bacteriol. 170:2575-2583.
33. Miller, V. L., R. K. Taylor, and J. J. Mekalanos. 1987. Choleratoxin transcriptional activator ToxR is a transmembrane DNAbinding protein. Cell 48:271-279.
34. Miyamoto, Y., T. Kato, Y. Obara, S. Akiyama, K. Takizawa,and S. Yamai. 1969. In vitro hemolytic characteristics of Vibrioparahaemolyticus: its close correlation with human pathogenic-ity. J. Bacteriol. 100:1147-1149.
35. Mizusawa, S., S. Nishimura, and F. Seela. 1986. Improvement ofthe dideoxy chain termination method of DNA sequencing by
J. BACTERIOL.
on June 22, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

toxRS OPERON OF VIBRIO PARAHAEMOLYTICUS 3855
use of deoxy-7-deazaguanosine triphosphate in place of dGTP.Nucleic Acids Res. 14:1319-1324.
36. Nishibuchi, M. Unpublished observation.37. Nishibuchi, M., A. Fasano, R. G. Russell, and J. B. Kaper. 1992.
Enterotoxigenicity of Vibrioparahaemolyticus with and withoutgenes encoding thermostable direct hemolysin. Infect. Immun.60:3539-3545.
38. Nishibuchi, M., M. Ishibashi, Y. Takeda, and J. B. Kaper. 1985.Detection of the thermostable direct hemolysin gene and relatedDNA sequences in DNA colony hybridization test. Infect.Immun. 49:481-486.
39. Nishibuchi, M., and J. B. Kaper. 1985. Nucleotide sequence ofthe thermostable direct hemolysin gene of Vibrio parahae-molyticus. J. Bacteriol. 162:558-564.
40. Nishibuchi, M., and J. B. Kaper. 1990. Duplication and variationof the thermostable direct haemolysin (tdh) gene in Vibrioparahaemolyticus. Mol. Microbiol. 4:87-99.
41. Nishibuchi, M., K. Kumagai, and J. B. Kaper. 1991. Contribu-tion of the tdhl gene of Kanagawa phenomenon-positive Vibrioparahaemolyticus to production of extracellular thermostabledirect hemolysin. Microb. Pathog. 11:453-460.
42. Nishibuchi, M., T. Taniguchi, T. Misawa, V. Khaeomanee-iam,T. Honda, and T. Miwatani. 1989. Cloning and nucleotidesequence of the gene (trh) encoding the hemolysin related to thethermostable direct hemolysin of Vibrio parahaemolyticus. In-fect. Immun. 57:2691-2697.
43. Oku, Y., Y. Uesaka, T. Hirayama, and Y. Takeda. 1988.Development of a highly sensitive bead-ELISA to detect bac-terial protein toxins. Microbiol. Immunol. 32:807-816.
44. Otteman, K. M., V. J. DiRita, and J. J. Mekalanos. 1992. ToxRproteins with substitutions in residues conserved with OmpRfail to activate transcription from the cholera toxin promoter. J.Bacteriol. 174:6807-6814.
45. Parsot, C., and J. J. Mekalanos. 1991. Expression of the Vibriocholerae gene encoding aldehyde dehydrogenase is under con-trol of ToxR, the cholera toxin transcriptional activator. J.Bacteriol. 173:2842-2851.
46. Parsot, C., E. Taxman, and J. J. Mekalanos. 1991. ToxRregulates the production of lipoproteins and the expression ofserum resistance in Vibrio cholerae. Proc. Natl. Acad. Sci.USA 88:1641-1645.
47. Peterson, K. M., and J. J. Mekalanos. 1988. Characterization ofthe Vibrio cholerae ToxR regulon: identification of novel genesinvolved in intestinal colonization. Infect. Immun. 56:2822-2829.
48. Richardoson, S. H. 1969. Factors influencing in vitro skin
permeability factor production by Vibno cholerae. J. Bacteriol.100:27-34.
49. Sakazaki, R., K. Tamura, T. Kato, Y. Obara, S. Yamai, and K.Hobo. 1968. Studies on the enteropathogenic, facultativelyhalophilic bacteria, Vibrioparahaemolyticus. III. Enteropatho-genicity. Jpn. J. Med. Sci. Biol. 21:325-331.
50. Schmidhauser, T. J., and D. R. Helinski. 1985. Regions ofbroad-host-range plasmid RK2 involved in replication and sta-ble maintenance in nine species of gram-negative bacteria. J.Bacteriol. 164:446-455.
51. Sharma, D. P., U. H. Stroeher, C. J. Thomas, P. A. Manning,and S. R. Attridge. 1989. The toxin-coregulated pilus (TCP) ofVibrio cholerae: molecular cloning of genes involved in pilusbiosynthesis and evaluation of TCP as a protective antigen inthe infant mouse model. Microb. Pathog. 7:437-448.
52. Shirai, H., H. Ito, T. Hirayama, Y. Nakamoto, N. Nakabayashi,K. Kumagai, Y. Takeda, and M. Nishibuchi. 1990. Molecularepidemiologic evidence for association of thermostable directhemolysin (TDH) and TDH-related hemolysin of Vibno para-haemolyticus with gastroenteritis. Infect. Immun. 58:3568-3573.
53. Stuber, D., and H. Bujard. 1981. Organization of transcriptionsignals in plasmids pBR322 and pACYC184. Proc. Natl. Acad.Sci. USA 78:167-171.
54. Takeda, Y. 1983. Thermostable direct hemolysin of Vibrioparahaemolyticus. Pharmacol. Ther. 19:123-146.
55. Taylor, R., C. Shaw, K. Peterson, P. Spears, and J. Mekalanos.1988. Safe, live Vibrio cholerae vaccines? Vaccine 6:151-154.
56. Taylor, R. K., V. L. Miller, D. B. Furlong, and J. J. Mekalanos.1987. Use ofphoA gene fusions to identify a pilus colonizationfactor coordinately regulated with cholera toxin. Proc. Natl.Acad. Sci. USA 84:2833-2837.
57. Terai, A., K. Baba, H. Shirai, 0. Yoshida, Y. Takeda, and M.Nishibuchi. 1991. Evidence for insertion sequence-mediatedspread of the thermostable direct hemolysin gene among Vibriospecies. J. Bacteriol. 173:5036-5046.
58. Twigg, A. J., and D. Sherratt. 1980. Trans-complementablecopy-number mutants of plasmid ColEl. Nature (London) 283:216-218.
59. Vieira, J., and J. Messing. 1982. The pUC plasmids, anM13mp7-derived system for insertion mutagenesis and sequenc-ing with synthetic universal primers. Gene 19:259-268.
60. Yamamoto, K., T. Honda, Y. Takeda, and T. Miwatani. 1983.Production of increased vascular permeability in rabbits bypurified thermostable direct hemolysin from Vibrio parahae-molyticus. J. Infect. Dis. 148:1129.
VOL. 175, 1993
on June 22, 2018 by guesthttp://jb.asm
.org/D
ownloaded from





























