Embed Size (px)
Citation preview

Biological Control 62 (2012) 65–74
Contents lists available at SciVerse ScienceDirect
Biological Control
journal homepage: www.elsevier .com/locate /ybcon
Paenibacillus lentimorbus B-30488r controls early blight disease in tomato byinducing host resistance associated gene expression and inhibiting Alternaria solani
Noor Khan, Aradhana Mishra, Chandra Shekhar Nautiyal ⇑Division of Plant–Microbe Interactions, National Botanical Research Institute, Rana Pratap Marg, Lucknow 226001, India
h i g h l i g h t s
" Paenibacillus lentimorbus B-30488r
demonstrates antagonism againstAlternaria solani.
" Foliar inoculation of P. lentimorbusB-30488r significantly reduces earlyblight infection in tomato.
" Possible mechanisms involved areinduction of defense mechanisms inplants and direct lysis of fungalhyphae.
1049-9644/$ - see front matter � 2012 Elsevier Inc. Ahttp://dx.doi.org/10.1016/j.biocontrol.2012.03.010
⇑ Corresponding author. Fax: +91 522 2206651.E-mail addresses: [email protected], csn@nb
g r a p h i c a l a b s t r a c t
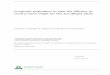
Pictorial representation demonstrating the influence of Paenibacillus lentimorbus B-30488r on biologicalinteractions with tomato plant and pathogen Alternaria solani.
a r t i c l e i n f o
Article history:Received 2 March 2011Accepted 22 March 2012Available online 2 April 2012
Keywords:Paenibacillus lentimorbus B-30488r
Alternaria solaniTomatoEarly blight
a b s t r a c t
The ability of Paenibacillus lentimorbus B-30488r (B-30488r) in reducing the early blight (EB) disease caused byAlternaria solani in tomato was assessed. Foliar application of the bacteria reduced the disease incidence by45.3% as compared to control. In vitro studies indicated that B-30488r effectively reduces the radial growthof A. solani in dual culture, and SEM studies demonstrated complete degradation of fungal hyphae on co-cul-ture with B-30488r. Both B-30488r and A. solani exhibited high similarity in the nutrient utilization of carbonsources present in tomato tissues. Changes in gene expression studied using RT-PCR analyses indicate signif-icant up-regulation of defense and growth-related genes in tomato plants treated with B-30488r foliar sprayand A. solani challenged plants pre-treated with B-30488r foliar spray. Principal component analysis resultsindicated that tomato plants treated with A. solani affected microbial community structure and populationof the rhizosphere as compared to B-30488r. Results suggest that P. lentimorbus B-30488r has a multiple modeof action for its biological control activity by inducing resistance in tomato plants, by degrading the pathogencell wall, and by competing for similar sources of nutrients on the phyllosphere. Additionally, B-30488r wouldnot affect microbial populations on the rhizosphere of the tomato plants.
� 2012 Elsevier Inc. All rights reserved.
1. Introduction
Tomato (Solanum lycopersicum Mill.) is an important vegetablecrop cultivated worldwide. In India, it is grown in an area of
ll rights reserved.
ri.res.in (C.S. Nautiyal).
0.5 million ha with an annual production of 7.4 million tonnesand productivity of 14.3 tonnes ha�1 (Shanmugasundaram, 2004;Latha et al., 2009). Tomato early blight (EB) disease caused by Alter-naria solani (Ellis and Martin) Jones and Grout is one of the mostcommon foliar disease of tomato in India, affecting the foliage,stems, and fruits of infected plants (Jones et al., 1991), and result-ing in up to 80% yield losses (Singh, 1985; Mathur and Shekhawat,

66 N. Khan et al. / Biological Control 62 (2012) 65–74
1986; Chandravanshi et al., 1994; Rotem, 1994; Patel et al., 2011).Improving the resistance of tomato cultivars seems to be a prom-ising option for control of EB, but the lack of single-gene resistanceand complex quantitative nature of inheritance of EB resistancehas resulted in the availability of no commercial tomato cultivarthat possesses adequate levels of resistance to A. solani (Nashand Gardner, 1988; Maiero and Ng, 1989; Thirthamallappa andLohithaswa, 2000). Therefore, disease control of EB is mainlyaccomplished with fungicides. However, these fungicides do notalways prevent the infestation of the fruits and severe losses canstill occur. Additionally, pesticides could have serious threat tohealth and environment in case of inappropriate application orstorage. Therefore, in order to provide protection against A. solaniin tomato, an alternative to usual chemical plant protection meth-ods is the induction of plant defenses by systemic acquired resis-tance (SAR) (Hunt and Ryals, 1996; Louws et al., 2001; Pérezet al., 2003; Vallad and Goodman, 2004). SAR is an induced systemof resistance triggered by exposing plant to biological or chemicalresistance inducers that confers long-lasting protection against abroad spectrum of disease-causing organisms (Durrant and Dong,2004; Asada et al., 2011).
The current investigation was conducted to study the effect(s)of Paenibacillus lentimorbus B-30488r application on tomato in sup-pressing the EB disease and the effect of the bacterial strain onmycelium of A. solani. Also, we investigated the capacity of antag-onist B-30488r in inducing genes associated with SAR in tomatoplants and the capacity of competition for carbon sources betweenthe pathogen and the antagonist. Moreover, the impact of B-30488r
foliar application on rhizopheric microflora of tomato plants wasassessed.
2. Materials and methods
2.1. Bacterial, fungal strains and inoculum preparation
The isolation procedure and media of cultivation of strain B-30488 was conducted as described in our previous studies (Nautiyalet al., 2006b; Khan et al., 2011, 2012). The strain B-30488 was firstidentified as Bacillus lentimorbus B-30488 through a fatty-acid anal-ysis performed by CABI Bioscience (Egham, Surrey, UK). After that,considering work of Pettersson et al. (1999), the strain was reclassi-fied as member of Paenibacillus genus. Strain B-30488 has beendeposited under the Budapest treaty into Agricultural Research Ser-vice (ARS) patent culture collection, United States Department ofAgriculture, Illinois. For monitoring the cell count of the inoculatedbacteria in the plant system, a stable spontaneous Rifr derivative ofB-30488 (B-30488r) was used as described earlier (DasGupta et al.,2006). The enumeration of the B-30488r population on tomato leaf-lets, immediately after inoculation and then every 24 h for 3 dayswas done by soaking a gram of aseptically collected fresh leaf tissuein 10 ml of sterile saline solution (0.85% [w/v] NaCl) followed by vor-texing (30 s) and plating serial 10-fold dilutions of the samples pre-pared in sterile saline up to 10�6 onto NA medium containingRifampicin (50 lg/ml). After 48 h of incubation at 28 �C, colonieswere counted and the population of B-30488r was expressed as logCFU g–1 fresh weight of tissue.
The fungus A. solani was maintained as described by Spletzerand Enyedi (1999). Briefly, the pathogen was subcultured biweeklyto modified cornmeal (MCM) agar (1.7% cornmeal agar, 0.2% glu-cose, 0.3% sucrose, and 0.1% yeast extract, w/v). Cultures displayeda gray, velvet-like appearance on MCM agar. To induce sporulation,mycelia plugs were cut from MCM plates and subcultured ontofreshly prepared MCM agar and were incubated at 25 �C in the darkfor 2 weeks. The resulting spore suspension was filtered through afine nylon mesh to remove bigger mycelial parts, quantified using ahemocytometer and adjusted to 104 conidia ml�1.
2.2. In vitro antagonistic studies
Antagonistic activity of B-30488r against A. solani was observeddirectly using the dual culture on Potato Dextrose Agar (PDA) fromHIMEDIA Laboratories, Mumbai, India). Actively growing B-30488r
was streaked on the one edge and a mycelial plug (9 mm diameter)from a 7-day-old culture of A. solani was placed on the oppositeside of a PDA plate. PDA plate with an A. solani mycelia plug inthe center served as the control. After incubation at 28 �C, the inhi-bition zone in the test plate was measured when fungus attainedfull growth in the control set (approximately 7 days). This experi-ment was conducted twice with three replicates for eachtreatment.
Microbial interactions between A. solani and B-30488r werestudied in co-culture using scanning electron microscopy (SEM)and were examined under a Phillips XL-20 (Holland) scanning elec-tron microscope as described earlier (Islam et al., 2005).
2.3. Determination of similarity in carbon source utilization by thepathogen and biocontrol agent
Similarity in carbon source utilization was quantified as the ra-tio of number of tomato carbon sources utilized in vitro by both thebiocontrol agent and the pathogen to the number of carbon sourcesutilized by the pathogen (the niche overlap index [NOI]). A total of32 carbon sources reported to be present in tomato tissues (Ji andWilson, 2002) were included in the test, and were individuallyincorporated into minimal medium A, MinA, (Miller, 1972) at aconcentration of 10 mM. Carbon sources included in the studywere: acetic acid, aconitic acid, L-alanine, ascorbic acid, asparticacid, citric acid, ethanolamine, formic acid, fumaric acid, fructose,glucose, L-glutamic acid, L-glutamine, L-glycine, glycolic acid, L-his-tidine, isoleucine, lactic acid, L-lysine, L-leucine, malic acid,L-methionine, oxalacetic acid, oxalic acid, L-phenylalanine,L-proline, succinic acid, sucrose, starch, tartaric acid, L-threonineand L-tryptophan. NOI values of >0.9 represent competencebetween species while scores of <0.9 represent occupation ofseparate niches (Wilson and Lindow, 1994a,b). The experimentwas carried out in triplicates (n = 3).
2.4. Assessment of biocontrol efficacy on A. solani (greenhouse studies)
Seeds of tomato (var. Damayanti) were surface disinfected byimmersing in 70% ethanol for 1 min, followed by three rinses withsterile distilled water (SDW). The seeds were sown in earthen trayscontaining non-sterilized soil and after the development of two trueleaves were transplanted to earthen pots (size: 0.35 m diameter,0.50 m height; volume of soil: 0.04 m3) and maintained in the green-house (27 ± 2 �C). Foliar spray with the treatments was performedon entire aboveground portion of 5 week old plants having devel-oped five fully unfolded leaves until run-off. Bacteria treated plantswere divided into two sets. In the first set, treated plants were chal-lenge inoculated with A. solani conidial suspension (104 conidiaml�1, at one day after foliar spray) and in the second set, treatedplants were not challenged with the fungal pathogen. Plants withoutprior treatment of bacteria were inoculated with the fungal patho-gen. The inoculated plants were then covered with wet plastic bagsfor 48 h to maintain high relative humidity and ensure conidia ger-mination and survival. The plants treated neither with the bacterianor challenged by the fungal pathogen served as control, and weresprayed with SDW. Thus treatments utilized in this study included:untreated control (C), B-30488r treated (B), A. solani treated (F), B + Ftreated, and a fungicide control (M = mancozeb [Akola ChemicalsLimited, India] at doses of 2.4 g l�1) with and without A. solani inoc-ulation. Bacterial treatments (B and B + F) used B-30488r suspensionof 1.8 � 109 CFU ml�1. Six replicates of each treatment, with five

N. Khan et al. / Biological Control 62 (2012) 65–74 67
plants in each pot, were maintained. The experiment was conductedusing completely a randomized design under greenhouse condi-tions. The trial was repeated once and the data presented were thepooling of two greenhouse trials. Measurements on morphologicalparameters, namely, shoot length and root length were recordedat the time of harvesting, using general methods as described earlier(Nautiyal et al., 2006a). The chlorophyll content of leaves was ac-quired by homogenizing the leaf tissue in 80% acetone, measuringthe absorbance at 663 and 645 nm (Arnon, 1949).
2.5. Disease assessment
The observations on assessment of disease severity in eachtreatment were made 10 days after inoculation by estimating thepercentage of early blight lesions in the middle third of the plantcanopy as described by Christ (1991). Ten leaves from each repli-cate were randomly collected for recording the observations, anda pot represented one replicate. The intensity of the disease was re-corded in each treatment following the score chart 0–9 scale (0:healthy; 1: 1–5%; 3: 6–10%; 5: 11–25%; 7: 26–50% and 9: >51% leafarea infected) proposed by Ramakrishnan et al. (1971). The percentdisease index (PDI) was estimated using the formula suggested byMckinney (1923). At harvest, the shoots were cut from the plantsdirectly above the soil surface and weighed to measure the freshplant biomass. As Alternaria defoliates the host plants and causeschlorosis at infected leaves resulting in reduced turgor, the freshplant biomass observation served as an alternative measure fordisease severity. The dead shedded leaves were excluded in thisestimation. Percent defoliation was estimated for each plant usingthe modified method suggested by Horsfall and Barratt (1945). Thedry weight was determined after a drying period of at least 72 h at70 �C.
2.6. RNA isolation and semi-quantitative RT-PCR analysis
For each treatment, leaves from the middle third canopy werecollected from each of the six replicates. The harvested leaves werepooled, flash frozen in liquid nitrogen, and stored at �80 �C untiluse. Total RNA was extracted from frozen leaves using the RNeasyPlant Mini RNA extraction kit (Qiagen, Valencia, CA, USA) followingthe manufacturer’s instructions. The quantity of RNA recoveredwas determined at 260 nm with a densitometer (model 8452A,Hewlett Packard Diode Array Spectrophotometer; Hewlett–Pack-ard Company, Sunnyvale, CA). Total RNA prepared was treatedwith RNase free DNase (MBI Fermantas, Germany) and used forfirst strand cDNA synthesis using RevertAid™ H minus first strandcDNA synthesis kit (MBI Fermantas, Germany) and oligo dT primeras per manufacturer’s recommendations. The cDNA obtained wasused as template for the semi-quantitative RT-PCR analyses byusing Thermo cycler (Flexi Gene-Techne, Cambridge, UK). PCR con-ditions were 3 min at 94 �C, followed by 30 cycles (94 �C, 45 s;64 �C, 1 min; 72 �C, 1 min), and 3 min at 72 �C. As a control to con-firm lack of genomic DNA contamination, 200 ng of DNase treatedRNA was used as template. The ubiquitin gene as the internal stan-dard was amplified using the gene-specific primers. For detectingthe influence of B-30488r inoculation on plants, we selected geneswell reported for plant growth and defense regulation in tomato(Gu et al., 2000; Zhou et al., 1997; Kavroulakis et al., 2006). Theprimers for genes related to plant growth [indole-3-acetic acid(IAA3), Gibberellin (GA3)]; defense regulation [PR1b (basic PR1),PR2a (acidic glucanase), PR2b (basic glucanase), Chitinase 3 (acidic),Chitinase 9 (basic), Pti4, Pto kinase] and Ubi3 (internal control) weredesigned by Primer 3.0 software (Applied Biosystems, http://fok-ker.wi.mit.edu/primer3/input.htm) based on tomato mRNA se-quences deposited in GenBank, and were synthesized byBangalore Genei (Banglore, India). The sequences of gene-specific
primer pairs used are presented in Table 1. The intensity of thebands of PCR products was quantitated using Scion Image Soft-ware, National Institute of Health (NIH), Maryland, USA; valueswere calculated in reference to internal control (Ubi3) and datawas plotted as relative percentage.
2.7. Determination of rhizospheric microbial diversity using carbonsource utilization pattern
Biolog Eco plates (Biolog, Inc., Hayward, CA, USA) were used todetermine the carbon source utilization pattern of four tomato rhi-zosphere soil samples as described earlier (Campbell et al., 1997;Khan et al., 2011). For individual treatments, tomato plants withroot-soil systems were shaken vigorously to separate soil nottightly adhering to the roots for about 1 min. The remaining thicksoil tightly attached to the root system was considered rhizospheresoil (Butler et al., 2003). These rhizosphere soil samples (10 g) wereshaken in 90 ml of sterile saline solution for 60 min and then ma-keup a final dilution 10�3. After incubation 150 ll of sample wereinoculated in each well of Biolog Eco plates and incubated at 30 �C.The rate of utilization is indicated by the reduction of tetrazolium,a redox indicator dye, which changes from colorless to purple. Datawere recorded for day 1–7 at 590 nm. Microbial activity in eachmicroplate, expressed as average well color development (AWCD)was determined as described by Garland (1996). Formulae used fordiversity and evenness indices calculations have been describedearlier (Staddon et al., 1997; Mishra and Nautiyal, 2009). Principalcomponent analysis (PCA) was performed on 5th day data dividedby the AWCD, as described by Garland and Mills (1991) usingStatistica 7.0. PCA is a statistical tool to reduce the number of vari-ables in a data matrix. The variables are projected onto new axes orprincipal components (PC) which account for a certain amount ofvariance of the sample. Since the first 2 PCs account for the highestvariance, they are generally the scores that are plotted. The PCsscore plot on first and second ordination axes (PC-I and PC-II)shows the spatial distribution of the similitudes and differencesamong the assessed treatments.
2.8. Statistical analysis
The collected data were analyzed statistically using a statisticalpackage, SPSS 16.0 (SPSS Inc., Chicago, IL, USA) and Statistica 7.0(StatSoft, Inc., OK 74104, USA). The results are displayed as means(±standard deviation). Means (±SD) of all groups were comparedand statistical variance analysis of the data was performed usinganalysis of variance (ANOVA). Furthermore, post-hoc proceduresuch as Duncan’s Multiple Range Test (DMRT) was used to identifythe pairs of groups where the means are significantly different ata = 0.05. Graphical work was carried out using Sigma Plot for Win-dows version 10.0 software packages.
3. Results
3.1. P. lentimorbus B-30488r – A. solani interaction
In vitro antagonism studies indicated that B-30488r effectivelyinhibited the radial growth of A. solani as indicated by an averageinhibition zone of 6 mm (data not shown). SEM investigation ofthe direct interactions between A. solani and B-30488r showedalteration and distortion of the hyphal cell wall. Six hours afterinoculation intense colonization of B-30488r was detected on fun-gal hyphae (Fig. 1A). By 12 h after inoculation, cells of B-30488r haddestructed the pathogen hyphae leading to expulsion of hyphalcytoplasmic content (Fig. 1B and C).

Table 1Sequences of gene-specific primers used in RT-PCR analysis.
Gene Primer sequence
PR1b For: 50-CCAAGACTATCTTGCGGTTC-30 Rev: 50-GAACCTAAGCCACGATACCA-30
PR2a For: 50-TATAGCCGTTGGAAACGAAG-30 Rev: 50-TGATACTTTGGCCTCTGGTC-30
PR2b For: 50-CAACTTGCCATCACATTCTG-30 Rev: 50-CCAAAATGCTTCTCAAGCTC-30
Chi3 For: 50-CAATTCGTTTCCAGGTTTTG-30 Rev: 50-ACTTTCCGCTGCAGTATTTG-30
Chi9 For: 50-AATTGTCAGAGCCAGTGTCC-30 Rev: 50-TCCAAAAGACCTCTGATTGC-30
Pti4 For: 50-ACGCTCTAAGCGTCGGATGG-30 Rev: 50-GTAACTCGAACCGGTTCCGG-30
Pto kinase For: 50-AGATTGAACCATGGCAGACC-30 Rev: 50-GATACTCTCACGCCGTAGCC-30
IAA3 For: 50-TTTTAGAAGCGAGCTATGTT-30 Rev: 50-AAGCCTTGAGTAATTGTGGA-30
GA3 For: 50-AGCTCGTCATCACAACATACGC-30 Rev: 50-CAACTCGCCCTTGTCGATAAAC-30
Ubi3 For: 50-CAGGACAAGGAAGGGAT-30 Rev: 50-ACCGCACTCAGCATTAG-30
68 N. Khan et al. / Biological Control 62 (2012) 65–74
3.2. Efficacy of B-30488r on EB incidence under greenhouse conditions
The efficacy of B-30488r against EB disease was tested under potculture conditions whereby plants pre-treated with B-30488r werechallenged A. solani through spray inoculation, along with theirrespective controls. Results showed that the bacterial inoculation,when used alone stimulated plant growth significantly as com-pared to un-inoculated control (a = 0.05), as evidenced by signifi-cant increase in shoot length, root length, chlorophyll content,fresh and dry plant biomass of bacteria treated plants than control(Table 2). Fig. 2 shows the effect of the control of EB in tomato by B-30488r. In addition, we also checked, if B-30488r could show similarinhibitory effects as like fungicide mancozeb against A. solani-in-duced EB. The results indicated B-30488r + A. solani treated plantsexhibited percent infected leaf area, percent defoliation and PDIof 39.9%, 16.0% and 40.2%, respectively, while mancozeb + A. solanitreatment showed 52.0%, 20.0% and 45.3% as compared to the A.solani treated plants where values were recorded as 73.0%, 32.1%and 68.0%, respectively. The disease reduction by a B-30488r
foliar spray inoculation was significantly higher (a = 0.05) and
Fig. 1. Scanning electron micrographs of Alternaria solani. Samples were collected at diffe12 h, (D) 1 h, (E) untreated control. Samples were prepared and visualized as described inof fungal cell wall leading to cytoplasmic leakage degradation of A. solani mycelia. Arrocontent.
comparable to the fungicide treatment (Fig. 3). The cell count ofB-30488r in bacteria treated plants was recorded to be1.5 � 106 CFU g�1, just after bacterial inoculation and it was main-tained at 2.1 � 105 CFU g�1 after 72 h. In bacteria treated plantssubsequently challenged by pathogen, the B-30488r cell countwas found to be 1.7 � 105 CFU g�1 after 72 h which implies thatthe presence of A. solani had no deleterious effect on the populationof B-30488r. No Rifr bacteria were observed on control plants.
3.3. B-30488r induced expression of defense and growth regulatorygenes in tomato
To study the pattern of disease resistance in tomato plants andthe influence of B-30488r, the expression profiles of the nine genesencoding defense related proteins and plant growth promoting fac-tors were determined (Fig. 4i). Level of gene expression increasedby 2- to 5-fold for the considered genes in B-30488r treated plantsas compared to control (Fig. 4ii). There was 2.1-, 3.0-, 3.61-, 2.73-,1.55-, 2.0- and 3.78-fold induction in PR1, PR2a, PR2b, Chi3, Chi9,Pti4 and Pto kinase genes, respectively, when plants treated with
rent times after incubation with Paenibacillus lentimorbus B-30488r, (A) 6 h, (B and C)Section 2. Note that enzymes and antimicrobials produced by B-30488r cause lysis
ws indicate further magnification sites and dotted arrows extrusion of cytoplasmic

Table 2Effect of P. lentimorbus B-30488r and fungicide mancozeb against Alternaria solani on plant growth parameters in tomato (var. Damayanti) under greenhouse conditions.
Treatments Plant growth variables
Shoot length (cm) Root length (cm) Total chlorophyll content (mg Chl/g FW) Fresh plant biomass (g) Dry plant biomass (g)
Control 13.7 ± 1.67a 6.66 ± 1.04a 2.99 ± 0.17a 8.47 ± 0.31a 5.23 ± 0.52a
B-30488r (B) 21.8 ± 2.61b 9.6 ± 0.67b 3.67 ± 0.34b 14.13 ± 0.50b 6.63 ± 0.72b
A. solani (F) 12.0 ± 1.32a 8.6 ± 1.02a 1.14 ± 0.05c 3.41 ± 0.08c 2.13 ± 0.52c
B-30488r + A. solani (B + F) 14.8 ± 2.87a 6.4 ± 1.16a 2.59 ± 0.18a 8.56 ± 0.11a 3.23 ± 0.21d
Mancozeb (M) 9.8 ± 1.41a 8.2 ± 1.34a 2.61 ± 0.28a 8.12 ± 0.07a 5.04 ± 0.46a
Mancozeb + A. solani (M + F) 15 ± 1.18a 12.2 ± 0.66c 2.58 ± 0.29a 8.05 ± 0.14a 5.16 ± 0.23a
Values are the mean of six replicates ± standard deviation, at the time of harvesting. Mean values followed by the same letters within each column are not significantlydifferent according to Duncan’s Multiple Range Test (a = 0.05).
N. Khan et al. / Biological Control 62 (2012) 65–74 69
B-30488r as compared to untreated control. Similar trend in resultswas obtained in B-30488r + A. solani treated plants, however onlyslight induction was observed in pathogen only treated plants.Up-regulated expression (approximately 2-fold) was also observedin B-30488r inoculated plants as compared to control for plantgrowth factors, IAA3 and Gibberellin.
3.4. B-30488r influences microbial diversity of tomato
To monitor changes in microbial activity and community com-position in the tomato rhizosphere as a result of B-30488r foliarspray inoculation or during biocontrol of A. solani with B-30488r,we utilized a rapid, community-level approach for assessing pat-terns of sole carbon source utilization by rhizospheric soil samplesusing Biolog Eco plate’s. Soil microbial community activity asmeasured by AWCD continued to increase during the whole incuba-tion and was consistently higher B-30488r (B) treatment than forother treatments, whereas the AWCD for A. solani (F) treatmentwas consistently the lowest (Fig. 5A). The PCA results indicate thatmaximum change in microbial community structure of the rhizo-sphere occurred in tomato plant treated with A. solani, while theinoculation with B-30488r caused no major difference in microbialcommunity structure as compared to the control (Fig. 5B). The plot-ted data are averages of three independent experiments. Variablesignificant differences among diversity and evenness indices werenoted in all the treatments. Inoculation of B-30488r makes the max-imum increase in the tomato rhizosphere functional diversity fol-lowed by B-30488r + A. solani treatment (Table 3).
3.5. Nutritional similarity
Carbon source utilization profiles of B-30488r and A. solani weredetermined based upon in vitro use, as sole carbon sources, of the32 carbon compounds reported to be present in tomato tissues andpresumed to be available in the tomato phyllosphere. Both organ-isms were able to utilize a wide variety of the compounds, indicat-ing that they are nutritionally versatile. The data obtained wereused to calculate a niche overlap index (NOI) for B-30488r and A.solani which was 0.94 (data not shown). This value confirms thatas the two organisms utilize nutrients in common, they wouldeffectively compete for a large proportion of the nutrients theyare capable of assimilating.
4. Discussion
This research focuses on the antagonistic potential of B-30488r
against A. solani where the high similarity between the nutrientutilization patterns of B-30488r and A. solani (NOI = 0.94) indicatedthat competition for nutrients could also play an important role inthe antagonistic effect. Such strong competition for niche availabil-ity is recognized as an important additional factor in the success ofB-30488r to antagonize the development of early blight disease in
tomato. These results reveal evidence for direct competition be-tween the antagonist and pathogen based on their respective met-abolic capabilities, and are in sync with the previous reportsestablishing correlation of the suppression of disease with thenutritional similarity of the antagonist and pathogen. Thepre-emptive utilization of nutrients in the phyllosphere by theinoculated biocontrol agent could influence nutrient availabilityand hence inhibit colonization and disease development of foliarplant pathogens (Wilson and Lindow, 1993; Ji and Wilson, 2002).
The importance of nutritional similarity in the biological controlof foliar bacterial diseases has been investigated previously withPseudomonas syringae pv. tomato, the causal agent of bacterialspeck of tomato (Ji and Wilson, 2002). Using a collection of non-pathogenic leaf associated bacteria and Tn5-generated catabolicmutants of P. syringae strain TLP2, a biological control agent of P.syringae pv. tomato (Ji and Wilson, 2002; Wilson et al., 2002) ob-served nutritional similarity between the non-pathogenic bacteriaor the catabolic mutants and P. syringae pv. tomato to be positivelycorrelated with reductions in bacterial speck severity.
Furthermore it was observed that foliar spray of B-30488r to to-mato plants subsequently infected with A. solani was highly effec-tive in reducing the severity of early blight disease and could alterdefense related gene expression. Suppression of the disease wasquantified as the percent reduction in disease severity comparedto the pathogen-only control when antagonist B-30488r was ap-plied to foliage 24 h prior to the pathogen. B-30488r was able tosurvive on the tomato leaf surface in relatively high amount (upto 105 CFU g�1 of leaf tissue), at least for the first 48 h after the A.solani challenge, the critical time probably necessary to suppressthe pathogen growth and subsequent infection on the leaves. Theresults reveal that B-30488r foliar inoculation reduced the diseaseseverity by 45.3% as compared to the fungicidal treatment (28.2%).Similar studies regarding the use of biocontrol agents for betterdisease control have been conducted by several workers. Lathaet al. (2009) reported that a combination of three biocontrolstrains, Pseudomonas fluorescens (Pf1 and Py15) and Bacillus subtilis(Bs16) along with the plant Zimmu extract providing the greatestand most effective disease control of EB and plant parameter ef-fects on tomato.
Tamietti et al. (1993) and Fuchs et al. (1997) hypothesized thatthe resistance induced in tomato plants by a protective strainmight be correlated with an increased accumulation of some PRproteins. Production of extracellular enzymes such as chitinasesand b-1,3-glucanases by biological control bacteria is a well-docu-mented phenomenon that has long been thought to be involved inthe lysis of the phytopathogenic fungal cell wall (Wessels andSietsma, 1981; Sela-Buurlage et al., 1993; Chernin et al., 1995). Inour previous study, we determined the production of chitinaseand b-1,3-glucanase by B-30488r, which has shown specificactivity in culture filtrate of 4.25 and 0.33 U mg�1, respectively(DasGupta et al., 2006). Additionally, evidences supporting the roleof chitinases and b-1,3-glucanases in defense against plant

Fig. 2. Disease severity on tomato plants under in vivo conditions 10 days post-pathogen challenge. The presented plants are control (C), B-30488r treated (B), A. solani treated(F) and B-30488r + Alternaria solani treated (B + F). Control plants were sprayed with sterile distilled water. The B + F treatment plants were pre-treated with bacterial culture,24 h prior to pathogen challenge.
0
10
20
30
40
50
60
70
80
Infected Leaf Area Defoliation Disease Index
Per
cent
age
(%)
F
B+F
M+Fc
c
b
b
b
Disease parameters
a
a
a
a
Fig. 3. Assessment of disease parameters for early blight disease of tomato undergreenhouse conditions. Control of B-30488r (B) and mancozeb (M) on Alternariasolani (F). Values represent mean of six replicates. Bars on each column correspondwith standard deviation. Mean values followed by the same letters on each bar foreach parameter are not significantly different according to Duncan’s multiple rangetest (a = 0.05).
70 N. Khan et al. / Biological Control 62 (2012) 65–74
pathogenic fungi in vivo have also been obtained. Mauch et al.(1988) reported that a mixture of chitinase and b-1,3-glucanasefrom pea tissues was significantly more effective against severalphytopathogenic fungi than either of these enzymes used individ-
ually. Transgenic plants expressing specific PR protein genesincluding PR-1a, basic chitinase, or basic b-1,3-glucanase exhibitedsignificantly increased protection against specific fungal pathogens(Broglie et al., 1991; Alexander et al., 1993; Zhu et al., 1994; Law-rence et al., 2000). Woo et al. (2006) demonstrated that chitinasesand glucanases produced by antagonist Trichoderma spp. on inter-action with fungal pathogen release oligosaccharide moleculesfrom the pathogen cell wall that elicit defense responses in plants.Pti4 is a tomato transcription factor that belongs to the ERF (ethyl-ene-responsive element binding factor) family of proteins. It inter-acts with the Pto kinase in tomato, which confers resistance to theP. syringae pv tomato pathogen that causes bacterial speck disease(Zhou et al., 1997; Gu et al., 2000). Interestingly, our study demon-strated significant up regulation of PR1, PR 2b, Chi3, Chi9, Pti4 andPto kinase genes in B-30488r treated tomato plants as comparedto the control. These results support the possibility that B-30488r
directly antagonizes the phytopathogenic fungi by producinghydrolytic enzymes that degrade the fungal cell wall and its foliarspray induces a potentiated state in the plant enabling it to bemore resistant to subsequent pathogen challenge, and that thismode of application can induce systemic responses in tomatoplants. Similar trend of up regulation in the mentioned geneswas also observed in plants treated with B-30488r followed bypathogen inoculation indicating that the presence of B-30488r re-duced the detrimental effect of A. solani on plants and may helpto overcome the pathogen infection by increasing levels of de-fense-related genes. In tomato, PR-1 family proteins have been

Fig. 4. Expression of plant growth (IAA3, indole acetic acid 3; Gibberellin) and defense (PR1; PR2a, acidic glucanase; PR2b, basic glucanase; Chi3, acidic Chitinase 3; Chi9, basicChitinase 9; Pti4, Pto kinase) regulatory genes in tomato leaves analyzed by semi-quantitative RT-PCR. Ubiquitin (Ubi3) was used as an internal control. Treatments: control(C), B-30488r treated (B), A. solani (F) and B-30488r + A. solani (B + F). Reaction products were analyzed by agarose gel electrophoresis (i). The intensities of the bands werequantitated, relative intensities calculated in reference to internal control (Ubi3); and values were plotted by considering untreated control (C) as 100% (ii).
Fig. 5. Impact of Paenibacillus lentimorbus B-30488r foliar inoculation on the microbial diversity of rhizosphere of tomato challenged with Alternaria solani. (A) Average wellcolor development (AWCD) based on substrate utilization pattern on Biolog Eco plates (Biolog Inc., Hayward, CA, USA) by tomato rhizosphere microflora in control (C); B-30488r (B); A. solani (F), and B-30488r + A. solani (B + F) during 7 days of incubation. (B) Principal component analysis (PCA) of carbon source utilization pattern on Biolog Ecoplates of the treatments: C, B, F and B + F in the tomato rhizosphere microflora after incubation for 5 days. Data on Biolog Eco plates were recorded every 24 h at 590 nm withan automated microplate reader (BioTek Instruments Inc., USA). PCA was performed using Statistica 7.0.
Table 3Diversity/evenness indices based on carbon utilization by microflora of tomato rhizosphere using Biolog Eco plates (Biolog Inc., Hayward, CA, USA) after incubation of 5 days aftertreatment with biocontrol agent Paenibacillus lentimorbus B-30488r against Alternaria solani.
Treatments Shannon diversity Shannon evenness Simpson diversity McIntosh diversity McIntosh evenness
Control 3.242 ± 0.010a 0.953 ± 0.002a 0.989 ± 0.001a 0.964 ± 0.002a 0.971 ± 0.001a
B-30488r (B) 3.315 ± 0.003b 0.971 ± 0.002b 0.995 ± 0.000b 0.985 ± 0.004b 0.982 ± 0.001b
A. solani (F) 3.240 ± 0.010a 0.950 ± 0.003a 0.988 ± 0.001a 0.964 ± 0.002a 0.966 ± 0.002a
B-30488r + A. solani (B + F) 3.261 ± 0.013a 0.972 ± 0.001b 0.994 ± 0.000b 0.982 ± 0.000b 0.982 ± 0.000b
Values are presented as mean of three replicated experiments ± standard deviation. Different letters indicate significant difference according to Waller–Duncan test (a = 0.05).Statistical analyses were performed using SPSS software (version 16.0).
N. Khan et al. / Biological Control 62 (2012) 65–74 71
demonstrated to have antifungal properties against Phytophthorainfestans and Cladosporium fulvum (Van Kan et al., 1992; Nidermanet al., 1995); however, no assay has been performed using A. solani.
By comparing foliar disease symptoms in B-30488r + A. solaniand A. solani control plants, we can conclude that B-30488r treat-ment does reduce susceptibility to A. solani, likely due, in part, to

Fig. 6. Pictorial representation demonstrating the influence of Paenibacillus lentimorbus B-30488r (B-30488r) on biological interactions with tomato plant and pathogenAlternaria solani.
72 N. Khan et al. / Biological Control 62 (2012) 65–74
the expression of the PR genes and subsequent induction of SAR.The PR-1 family of genes along with chitinase and b-1,3-glucanaseare known to be reliable markers of SAR useful in identifying bio-control agents with SAR abilities (Ward et al., 1991; Hunt andRyals, 1996; Bargabus et al., 2004), and B-30488r strongly inducedthe expression of the PR genes particularly PR1, PR2a, PR2b andChi3 in tomato. Furthermore, in B-30488r + A. solani treatment, ahigher or almost equal expression was recorded for genes PR1,PR2a, PR2b, Chi3, Chi9 and Pti4 as compared to B-30488r alonetreatment. Therefore, it can be inferred that no negative influenceof pathogen was exerted on bacteria mediated induction of hostdefense genes.
Considering that the strain B-30488r protects tomato plantagainst A. solani, it is interesting to determine whether B-30488r
foliar spray affected the microbial population in the tomato rhizo-sphere. Hence, to monitor changes in microbial activity and com-munity composition in the plant rhizosphere of varioustreatments, a rapid, community-level approach for assessing pat-terns of sole carbon source utilization by microbial samples usingBiolog plate’s was done. It shows promise as a means of assessingmicrobial community structure, which examines the functionalcapabilities of the microbial population, and the resulting datacan be analyzed using multivariate techniques to compare meta-bolic capability of communities (Garland, 1996). Biolog plates havefound application for the assessment of microbial metabolic diver-sity to differentiate microbial communities from diverse habitatsrepresenting freshwater, sea water, coastal lagoon, soil, rhizo-sphere, phyllosphere, groundwater, activated sludge reactors, andcompost (Garland, 1996; Gomez et al., 2000; Sala et al., 2008).AWCD based on substrate utilization pattern on Biolog Eco platesprovides information about the overall changes in total microbialactivity/abundance, and offers the potential to monitor changesin microbial diversity caused by environmental fluctuations, man-agement practises and pollution. Variable significant differencesamong treatments were noted for the samples collected usingthe McIntosh, Shannon, and Simpson indices. In addition, PCA re-sults of substrate source utilization pattern on Biolog Eco platesby tomato rhizosphere microflora in all the treatments showed
that B-30488r foliar application impacted microbial populationand native microbial community structure in rhizosphere as thebacterial spraying treatment exhibited a higher evenness and rich-ness diversity as compared to the other treatments. PCA plotshowed the four treatments [control (C); B-30488r (B); A. solani(F), and B-30488r + A. solani (B + F)] to be distributed separatelyon PC axes depicted by Factor 1 (81.4% variation) and Factor 2(11.3% variation). PCA thus indicated that there was distinct reso-lution of soil microbial communities in all treatments, and in gen-eral, treatments C, B and B + F were closely clustered as comparedfungus control treatment. This implies that the pathogen challengeimpacted native microbial community structure in soil such that itwas most separated from the rest of the three treatments (C, B andB + F), while the spray inoculation of B-30488r produced no majoreffect on the rhizospheric microbial population. PCA discrimina-tion among different treatments depended on the extent to whichcarbon substrates of Biolog Eco plates were oxidized. These obser-vations reflecting the impact of A. solani, B-30488r andB-30488r + A. solani on dynamics of microbial communities werein agreement with disease severity measurements and geneexpression analyses.
As an outcome of this research, we can propose a first modelexplaining the multifactorial mode of action of B-30488r, whichis illustrated in Fig. 6. Multifactorial mode of action has been re-ported in other biological control agents such as Trichodermaspp., where antibiosis, mycoprasitism, SAR induction and plantgrowth promotion have been involved in control of phytopatho-gens (Harman et al., 2004). Moreover, it is apparent thatP. lentimorbus B-30488r shows good prospect as an inoculant forplant growth promotion in tomato and biological control againstA. solani. The study undertaken provides important insight on thetripartite ecological interaction occurring among tomato – A. solaniand B-30488r which will help in devising novel strategies for thecontrol of A. solani, pathogen causing EB disease of tomato.
Conflict of interest statement
There is no conflict of interest in the present work.

N. Khan et al. / Biological Control 62 (2012) 65–74 73
Acknowledgments
The study was supported by Task force Grant NWP-006 fromCouncil of Scientific and Industrial Research (CSIR), New Delhi, In-dia. Part of the work was supported by TATA Innovation Fellowshipawarded to C.S.N. by Department of Biotechnology, Government ofIndia. The authors thank CSIR for awarding Senior Research Fellow-ship (SRF) to N.K.
References
Alexander, D., Goodman, R.M., Gut-Rella, M., Glascock, C., Weymann, K., Friedrich, L.,Maddox, D., Ahl-Goy, P., Luntz, T., Ward, E., 1993. Increased tolerance to twooomycete pathogens in transgenic tobacco expressing pathogenesis-relatedprotein 1a. Proceedings of the National Academy of Sciences, United States ofAmerica 90, 7327–7331.
Arnon, D.I., 1949. Copper enzymes in isolated chloroplasts. Polyphenol oxidase inBeta vulgaris. Plant Physiology 24, 1–15.
Asada, Y., Yamamoto, M., Tsutsui, T., Yamaguchi, J., 2011. The Arabidopsis NSL2negatively controls systemic acquired resistance via hypersensitive response.Plant Biotechnology 28, 9–15.
Bargabus, R.L., Zidack, N.K., Sherwood, J.E., Jacobsen, B.J., 2004. Screening for theidentification of potential biological control agents that induce systemicacquired resistance in sugar beet. Biological Control 30, 342–350.
Broglie, K., Chet, I., Holliday, M., Cressman, R., Biddle, P., Knowlton, S., Mauvais, C.J.,Broglie, R., 1991. Transgenic plants with enhanced resistance to the fungalpathogen Rhizoctonia solani. Science 254, 1194–1197.
Butler, J.L., Williams, M.A., Bottomley, P.J., Myrold, D.D., 2003. Microbial communitydynamics associated with rhizosphere carbon flow. Applied and EnvironmentalMicrobiology 60, 6793–6800.
Campbell, C.D., Grayston, S.J., Hirst, D.J., 1997. Use of rhizosphere carbon sources insole carbon utilization tests to discriminate soil microbial communities. Journalof Microbiological Methods 30, 33–41.
Chandravanshi, S.S., Singh, B.P., Thakur, M.P., 1994. Persistence of differentfungicides used against Alternaria alternata in tomato. Indian Phytopathology47, 241–244.
Chernin, L., Ismailov, Z., Haran, S., Chet, I., 1995. Chitinolytic Enterobacter agglomeransantagonistic to fungal plant pathogens. Applied and Environmental Microbiology61, 1720–1726.
Christ, B.J., 1991. Effect of disease assessment method on ranking potato cultivarsfor resistance to early blight. Plant Disease 75, 353–356.
DasGupta, S.M., Khan, N., Nautiyal, C.S., 2006. Biologic control ability of plantgrowth-promoting Paenibacillus lentimorbus NRRL B-30488 isolated from milk.Current Microbiology 53, 502–505.
Durrant, W.E., Dong, X., 2004. Systemic acquired resistance. Annual Review ofPhytopathology 42, 185–209.
Fuchs, J.G., Moenne-Loccoz, Y., Defago, G., 1997. Nonpathogenic Fusariumoxysporum strain Fo47 induces resistance to Fusarium wilt in tomato. PlantDisease 1997 (81), 492–496.
Garland, J.L., 1996. Analytical approaches to the characterization of samples ofmicrobial communities using patterns of potential C source utilization. SoilBiology & Biochemistry 28, 213–221.
Garland, J.L., Mills, A.L., 1991. Classification and characterization of heterotrophicmicrobial communities on the basis of patterns of community-level sole-carbon-source utilization. Applied and Environmental Microbiology 57, 2351–2359.
Gomez, E., Bisaro, V., Conti, M., 2000. Potencial C-source utilization patterns ofbacterial communities as influenced by clearing and land use in a vertic soil ofArgentina. Applied Soil Ecology 15, 273–281.
Gu, Y., Yang, C., Thara, Y.K., Zhou, J., Martin, G.B., 2000. Pti4 is induced by ethyleneand salicylic acid, and its product is phosphorylated by the Pto kinase. Plant Cell12, 771–786.
Harman, G.E., Howell, C.R., Viterbo, A., Chet, I., Lorito, M., 2004. Trichoderma speciesopportunistic, avirulent plant symbionts. Nature Reviews Microbiology 2, 43–56.
Horsfall, J.G., Barratt, R.W., 1945. An improved grading system for measuring plantdiseases. Phytopathology 35, 655.
Hunt, M.D., Ryals, J.A., 1996. Systemic acquired resistance signal transduction.Critical Reviews in Plant Sciences 15, 583–606.
Islam, M.T., Hashidoko, Y., Deora, A., Ito, T., Tahara, S., 2005. Suppression ofdamping-off disease in host plants by the rhizoplane bacterium Lysobacter sp.strain SB-K88 is linked to plant colonization and antibiosis against soilbornePeronosporomycetes. Applied and Environmental Microbiology 71 (7), 3786–3796.
Ji, P., Wilson, M., 2002. Assessment of the importance of similarity in carbon sourceutilization profiles between the biological control agent and the pathogen inbiological control of bacterial speck of tomato. Applied and EnvironmentalMicrobiology 68, 4383–4389.
Jones, J.B., Jones, J.P., Stall, R.E., Zitter, T.A., 1991. Infectious Diseases: DiseasesCaused by Fungi. Compendium of tomato diseases. The AmericanPhytopathological Society, St. Paul, MN, pp. 9–25.
Kavroulakis, N., Papadopoulou, K.K., Ntougias, S., Zervakis, G.I., Ehaliotis, C., 2006.Cytological and other aspects of pathogenesis-related gene expression intomato plants grown on a suppressive compost. Annals of Botany 98, 555–564.
Khan, N., Mishra, A., Chauhan, P.S., Nautiyal, C.S., 2011. Induction of Paenibacilluslentimorbus biofilm by sodium alginate and CaCl2 alleviates drought stress inchickpea. Annals of Applied Biology 159 (3), 372–386.
Khan, N., Mishra, A., Chauhan, P.S., Sharma, Y.K., Nautiyal, C.S., 2012. Paenibacilluslentimorbus enhances growth of chickpea (Cicer arietinum L.) in chromium-amended soil. Antonie van Leeuwenhoek 101 (2), 453–459.
Latha, P., Anand, T., Ragupathi, N., Prakasam, V., Samiyappan, R., 2009. Antimicrobialactivity of plant extracts and induction of systemic resistance in tomato plantsby mixtures of PGPR strains and Zimmu leaf extract against Alternaria solani.Biological Control 50, 85–93.
Lawrence, C.B., Singh, N.P., Qiu, J., Gardener, R.G., Tuzun, S., 2000. Constitutivehydrolytic enzymes are associated with polygenic resistance of tomato toAlternaria solani and may function as an elicitor release mechanism.Physiological and Molecular Plant Pathology 57, 211–220.
Louws, F.J., Wilson, M., Campbell, H.L., Cuppels, D.A., Jones, J.B., Shoemaker, P.B.,Sahin, F., Miller, S.A., 2001. Field control of bacterial spot and bacterial speck oftomato using a plant activator. Plant Disease 85, 481–488.
Maiero, M., Ng, T.J., 1989. Genetic analyses of early blight resistance in tomatoes.HortScience 24, 221–226.
Mathur, K., Shekhawat, K.S., 1986. Chemical control of early blight in kharif sowntomato. Indian Journal of Mycology and Plant Pathology 16 (2), 235–236.
Mauch, F., Mauch-Mani, B., Boller, T., 1988. Antifungal hydrolases in pea tissue. II.Inhibition of fungal growth by combinations of chitinase and beta-1,3-glucanase. Plant Physiology 88, 936–942.
Mckinney, H.H., 1923. A new system of grading plant diseases. Journal ofAgricultural Research 26, 195–218.
Miller, J.H., 1972. Experiments in Bacterial Genetics. Cold Spring Harbor Laboratory,Cold Spring Harbor, NY.
Mishra, A., Nautiyal, C.S., 2009. Functional diversity of the microbial community inthe rhizosphere of chickpea grown in diesel fuel-spiked soil amended withTrichoderma ressei using sole-carbon-source utilization profiles. World Journalof Microbiology and Biotechnology 25, 1175–1180.
Nash, A.F., Gardner, R.G., 1988. Heritability of tomato early blight resistance derivedfrom Lycopersicon hirsutum P.I. 126445. Journal of the American Society forHorticultural Science, 268.
Nautiyal, C.S., Mehta, S.D., Singh, H.B., Pushpangadan, P., 2006a. Synergisticbioinoculant composition comprising bacterial strains of accession Nos. NRRLB-30486, NRRL B-30487 and NRRL B-30488 and method of producing saidcomposition thereof. US Patent 7097830.
Nautiyal, C.S., Mehta, S., Singh, H.B., 2006b. Biological control and plant growth-promoting Paenibacillus strains from milk. Journal of Microbiology andBiotechnology 16, 184–192.
Niderman, T., Genetet, I., Bruyère, T., Gees, R., Stintzi, A., Legrand, M., Fritig, B.,Mösinger, E., 1995. Pathogenesis-related PR-1 proteins are antifungal. PlantPhysiology 108, 17–27.
Patel, S.J., Subramanian, R.B., Jha, Y.S., 2011. Biochemical and molecular studies ofearly blight disease in tomato. Phytoparasitica. http://dx.doi.org/10.1007/s12600-011-0156-6.
Pérez, L., Rodriguez, M.E., Rodriguez, F., Roson, C., 2003. Efficacy of acibenzolar-S-methyl, an inducer of systemic acquired resistance against tobacco blue moldcaused by Peronospora hyoscyami f. sp. tabacina. Crop Protection 22 (2), 405–413.
Pettersson, B., Rippere, K.E., Yousten, A.A., Priest, F.G., 1999. Transfer of Bacilluslentimorbus and Bacillus popilliae to the genus Paenibacillus with emendeddescriptions of Paenibacillus lentimorbus comb. nov. and Paenibacillus popilliaecomb. Nov. International Journal of Systematic Bacteriology 49, 531–540.
Ramakrishnan, L., Kamalnathan, S., Krishnamurthy, C.S., 1971. Studies on Alternarialeaf spot of tomato. Madras Agricultural Journal 58 (4), 275–280.
Rotem, J., 1994. The genus Alternaria: biology, epidemiology and pathogenicity.American Phytopathological Society Press, St. Paul, Minnesota.
Sala, M.M., Terrado, R., Lovejoy, C., Unrein, F., Pedros-Alio, C., 2008. Metabolicdiversity of heterotrophic bacterioplankton over winter and spring in thecoastal Arctic Ocean. Environmental Microbiology 10, 942–949.
Sela-Buurlage, M.B., Ponstein, A.S., Bres-Vloemans, S.A., Melchers, L.S., Van denElzen, P.J.M., Cornelissen, B.J.C., 1993. Only specific tobacco (Nicotiana tabacum)chitinases and beta-1,3-glucanases exhibit antifungal activity. Plant Physiology101, 857–863.
Shanmugasundaram, S., 2004. The Hindu Survey of Indian, Agriculture, p. 127.Singh, R.S., 1985. Diseases of Vegetable Crops. Oxford and IBH Publishing Co., New
Delhi, Bombay, Calcutta, p. 346.Spletzer, M.E., Enyedi, A.J., 1999. Salicylic acid induces resistance to Alternaria solani
in hydroponically grown tomato. Phytopathology 89, 722–727.Staddon, W.J., Duchesne, L.C., Trevors, J.T., 1997. Microbial diversity and community
structure of post disturbance forest soils as determined by sole-carbon-sourceutilization patterns. Microbial Ecology 34, 125–130.
Tamietti, G., Ferraris, L., Matta, A., Abbattista Gentile, I., 1993. Physiologicalresponses of tomato plants grown in Fusarium suppressive soil. Journal ofPhytopathology 138, 66–76.
Thirthamallappa, R.A.S., Lohithaswa, H.C., 2000. Genetics of resistance to earlyblight (Alternaria solani Sorauer) in tomato (Lycopersicon esculentum L.).Euphytica 113, 187–193.
Vallad, G.E., Goodman, R.M., 2004. Systemic acquired resistance and inducedsystemic resistance in conventional agriculture. Crop Science 44, 1920–1934.

74 N. Khan et al. / Biological Control 62 (2012) 65–74
Van Kan, J.A.L., Joosten, M.H.A.J., Wagemakers, C.A.M., van den Berg-Velthuis,G.C.M., de Wit, P.J.G.M., 1992. Differential accumulation of mRNAs encodingextracellular and intracellular PR proteins in tomato induced by virulent andavirulent races of Cladosporium fulvum. Plant Molecular Biology 20, 513–527.
Ward, E.R., Uknes, S.J., Williams, S.C., Dincher, S.S., Wiederhold, D.L., Alexander, D.C.,Ahl-Goy, P., Métraux, J.P., Ryals, J.A., 1991. Coordinate gene activity in responseto agents that induce systemic acquired resistance. Plant Cell 3, 1085–1094.
Wessels, J.G.H., Sietsma, J.H., 1981. Fungal cell walls: a survey. In: Tanner, W.,Loewus, F.A., (Eds.), Encyclopedia of Plant Physiology. New series, vol. 13B. PlantCarbohydrates. Springer-Verlag, Berlin, 352–394.
Wilson, M., Lindow, S.E., 1993. Population dynamics of Pseudomonas fluorescensstrain A506 in pear flowers following inoculation in relation to strategies ofbiological control of fire blight and frost injury. Acta Horticulturae 338, 331–332.
Wilson, M., Lindow, S.E., 1994a. Coexistence among epiphytic bacterial populationsmediated through nutritional resource partitioning. Applied and EnvironmentalMicrobiology 60, 4468–4477.
Wilson, M., Lindow, S.E., 1994b. Ecological similarity and coexistence of epiphyticice-nucleating (Ice+) Pseudomonas syringae strains and non-nucleating (Ice�)biological control agent. Applied and Environmental Microbiology 60, 3128–3137.
Wilson, M., Campbell, H.L., Ji, P., Jones, J.B., Cuppels, D.A., 2002. Biological control ofbacterial speck of tomato under field conditions at several locations in NorthAmerica. Phytopathology 92, 1284–1292.
Woo, S.L., Scala, F., Ruocco, M., Lorito, M., 2006. The molecular biology of theinteractions between Trichoderma spp., phytopathogenic fungi, and plants.Phytopathology 96, 181–185.
Zhou, J., Tang, X., Martin, G.B., 1997. The Pto kinase conferring resistance to tomatobacterial speck disease interacts with proteins that bind a cis-element ofpathogenesis-related genes. The EMBO Journal 16, 3207–3218.
Zhu, Q., Maher, E.A., Masound, S., Dixon, R.A., Lamb, C.J., 1994. Enhanced protectionagainst fungal attack by constitutive co-expression of chitinase and glucanasegenes in transgenic tobacco. Bio/Technology 12, 807–812.