Embed Size (px)
Citation preview

Research Article
Optimization of T-cell Reactivity by ExploitingTCR Chain Centricity for the Purpose of Safe andEffective Antitumor TCR Gene TherapyToshiki Ochi1, Munehide Nakatsugawa1, Kenji Chamoto1, Shinya Tanaka1,2,Yuki Yamashita1,Tingxi Guo1,3, Hiroshi Fujiwara4, Masaki Yasukawa4, Marcus O. Butler1,3,5, andNaoto Hirano1,3
Abstract
Adoptive transfer of T cells redirected by a high-affinityantitumor T-cell receptor (TCR) is a promising treatmentmodality for cancer patients. Safety and efficacy depend onthe selection of a TCR that induces minimal toxicity and elicitssufficient antitumor reactivity. Many, if not all, TCRs possesscross-reactivity to unrelated MHC molecules in addition toreactivity to target self-MHC/peptide complexes. Some TCRsdisplay chain centricity, in which recognition of MHC/peptidecomplexes is dominated by one of the TCR hemi-chains. In thisstudy, we comprehensively studied how TCR chain centricityaffects reactivity to target self-MHC/peptide complexes andalloreactivity using the TCR, clone TAK1, which is specific forhuman leukocyte antigen-A�24:02/Wilms tumor 1235–243(A24/WT1235) and cross-reactive with B�57:01 (B57). TheTAK1b, but not the TAK1a, hemi-chain possessed chain cen-
tricity. When paired with multiple clonotypic TCRa counter-chains encoding TRAV12-2, 20, 36, or 38-2, the de novo TAK1b-containing TCRs showed enhanced, weakened, or absent reac-tivity to A24/WT1235 and/or to B57. T cells reconstituted withthese TCRa genes along with TAK1b possessed a very broadrange (>3 log orders) of functional and structural avidities.These results suggest that TCR chain centricity can be exploitedto enhance desired antitumor TCR reactivity and eliminateunwanted TCR cross-reactivity. TCR reactivity to target MHC/peptide complexes and cross-reactivity to unrelated MHCmole-cules are not inextricably linked and are separable at the TCRsequence level. However, it is still mandatory to carefullymonitor for possible harmful toxicities caused by adoptivetransfer of T cells redirected by thymically unselected TCRs.Cancer Immunol Res; 3(9); 1070–81. �2015 AACR.
IntroductionOne challenge for the success of adoptive T-cell therapy is that
thymic and/or peripheral tolerance impairs our ability to preparehigh avidity antitumor T cells from all cancer patients (1, 2). Toovercome this issue, the concept of engineered T cells has beendeveloped in which T cells are redirected to specifically recognizetumors by the transfer of genes encoding high-affinity tumor-specific chimeric antigen receptors (CAR) or T-cell receptors (TCR;refs. 3–7). These redirected T cells are endowed with specificityand strong reactivity for tumor cells by transducing high-affinityantitumor CAR or TCR gene (3–7).
Selection of a well-characterized antitumor TCR is pivotal forTCR gene therapy to achieve clinically effective antitumorresponses with minimal adverse events. Obviously, TCR-trans-duced T cells should possess functional avidity sufficient torecognize tumor cells. However, in order to achieve a favorabletherapeutic window and avoid on-target off-tumor toxicity, theavidity should not be excessively high given the expression oftarget antigens in normal tissues (5, 7). In addition, the risk of off-target toxicity caused by cross-reactivity inherent to TCRs must beconsidered, because many, if not all, TCRs possess cross-reactivityto self and non-self MHC presenting other peptides (8–14). Inrecent clinical trials of TCRgene therapy, itwas reported that cross-reactivity of an antitumor TCR to a self-HLA–presented different,non–tumor-derived peptide caused unwanted off-target adverseevents in patients treatedwith redirected T cells (15, 16). Strategiesto remove unwanted cross-reactivity of TCRs while preservingantitumor reactivity have yet to be established.
Accumulating evidence has demonstrated that antigen-specificT cells in the periphery harbor identical or near-identicalsequences across unrelated individuals despite a potentially vastT-cell repertoire (17, 18). Interestingly, a subset of biased TCRaand b chains possess chain centricity and thereby can pair withmultiple clonotypic counter-chainswhile preserving antigen spec-ificity (19–22). Avidities for these TCRs that are composed ofchain-centric hemi-chains appear to be diverse (20, 23, 24). T cellstransduced with a dominant TCR hemi-chain demonstrate cross-reactivity to different peptides presented by the same self-MHCmolecules depending on the TCR counter-chain clonotypes while
1Immune Therapy Program, Campbell Family Institute for Breast Can-cer Research, Campbell Family Cancer Research Institute, PrincessMargaret Cancer Centre, University Health Network,Toronto, Ontario,Canada. 2Takara Bio, Inc., Otsu, Shiga, Japan. 3Department of Immu-nology, University of Toronto,Toronto, Ontario, Canada. 4Departmentof Hematology, Clinical Immunology and Infectious Disease, EhimeUniversity Graduate School of Medicine,Toon, Ehime, Japan. 5Depart-ment of Medicine, University of Toronto, Toronto, Ontario, Canada.
Note: Supplementary data for this article are available at Cancer ImmunologyResearch Online (http://cancerimmunolres.aacrjournals.org/).
Corresponding Author: Naoto Hirano, Princess Margaret Cancer Centre, 610University Avenue, Toronto, ON M5G 2M9, Canada. Phone: 416-946-2190; Fax:416-946-6529; E-mail: [email protected]
doi: 10.1158/2326-6066.CIR-14-0222
�2015 American Association for Cancer Research.
CancerImmunologyResearch
Cancer Immunol Res; 3(9) September 20151070
on July 7, 2018. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Published OnlineFirst May 5, 2015; DOI: 10.1158/2326-6066.CIR-14-0222

retaining their original antigen specificity (21). These observa-tions suggest that affinity and cross-reactivity of a TCR comprisinga dominant hemi-chain can be modulated by pairing it with anon–chain-centric counter-chain without perturbing antigenspecificity.
Wilms tumor 1 (WT1) is a promising tumor-associated antigen,becauseWT1 expression is abundant inmany cancers, but restrict-ed to some types of normal tissue, such as podocytes in the kidney,mesothelial cells, and hematopoietic cells (25–30). A number ofimmunotherapy clinical trials targeting WT1 have been con-ducted to date. No clinically evident treatment-related toxicitieshave been observed in these trials (31–34). Multiple clinical trialsin which patients with acute myelogenous leukemia are infusedwith autologous or allogeneic T cells redirected by HLA class I–restricted WT1-specific TCRs are currently ongoing.
The HLA-A�24:02/WT1235–243 (A24/WT1235)-specific TCR,clone TAK1, was previously isolated from an HLA-A24þ B�57:01(B57)� donor. Further characterization revealed that TAK1 TCRindeed possesses cross-reactivity for unrelated B57 molecules(35). Using the TAK1 TCR as a model, we comprehensivelystudied the effects of TCR chain centricity on reactivity to targetself-MHC/peptide complexes and alloreactivity.
Materials and MethodsCells
Blood samples were obtained from healthy donors followingInstitutional Review Board approval. HLA-A�24:02 positivity wasdetermined using a PCR-based genotyping method (36). Alldonors were determined negative for B�57:01 by high-resolutionHLA DNA typing (American Red Cross). Mononuclear cells wereobtained by density gradient centrifugation (Lymphoprep;Nycomed Pharma AS). CD34þ hematopoietic cells were purifiedfrom cord blood mononuclear cells using FITC-conjugated anti-CD34 mAb (clone AC136; Miltenyi Biotec) in combination withanti-FITCmicrobeads according to themanufacturer's instruction(Miltenyi Biotec). We directly obtained the following cell linesfrom the American Type Culture Collection: K562, which is anerythroleukemic cell line defective for HLA expression, and T2,which is a T-cell leukemia/B-cell large cell leukemia hybrid cellline. Jurkat 76, devoid of endogenous TCR and CD8 expression,was kindly providedbyDr.Heemskerk, LeidenUniversityMedicalCentre, Leiden, Netherlands (37). All cell lines were cultured asreported previously (38, 39) and passaged for less than 6months.
cDNAsCodon-optimized TAK1 TCR genes were produced by Thermo
Fisher Scientific (35). For transduction into peripheral T cells, theTAK1 TCR hemi-chain was fused with a truncated form of nervegrowth factor receptor (DNGFR) as described previously (20, 40).DNGFR gene alone was employed as a control. To clone TCRagenes, 50-rapid amplification of cDNA ends (RACE) PCR wasperformed using a SMARTer RACE cDNA amplification kit(Takara Bio). For the first round of PCR, cDNA was amplifiedusing a supplied 50-RACE primer and a 30-TCRa untranslatedregion primer (50-GGAGAGTTCCCTCTGTTTGGAGAG-30). Thesecond-round PCR was performed using a modified 50-RACEprimer (50- GTGTGGTGGTACGGGAATTCAAGCAGTGGTAT-CAACGCAGAGT-30) and a 30-TCRa primer (50-ACCACTGT-GCTGGCGGCCGCTCAGCTGGACCACAGCCGCAGCG-30). TCRgenes fused with or without DNGFR were cloned into a pMX
retrovirus vector and utilized to transduce cell lines and primaryhuman T cells. Nucleotide sequencing was performed at theCentre for Applied Genomics, The Hospital for Sick Children(Toronto, Canada). TCR gene allele names are in accordance withIMGT unique gene nomenclature (http://www.imgt.org/).
TransfectantsJurkat 76/CD8 cells stably express CD8a and bmolecules (39).
To generate TCR transfectants, Jurkat 76/CD8 cells were initiallytransduced with the TAK1b gene and then superinfected withindividual TCRa genes. These TCR genes were not tagged withDNGFR gene. All TCR transfectants were purified using FITC-conjugated anti-TRBV5.1mAb (clone IMMU157; BeckmanCoul-ter) in combination with anti-FITC microbeads according to themanufacturer's instruction (Miltenyi Biotec). TRBV5.1 is encodedby the TAK1b gene. T2 cells were transduced with HLA-A24 togenerate T2-A24. K562-based A24-artificial antigen-presentingcells (aAPC) and B57-aAPCs expressing HLA-A24 and B57 as asingle HLA allele, respectively, in association with CD80 andCD83, were established as reported elsewhere (38, 39). PG13-derived retrovirus supernatants were utilized to transduce genesinto T cells.
Expansion of TCR hemi-chain–transduced T cellsPeripheral T cells were stimulated with 50 ng/mL anti-CD3
mAb (clone OKT3) and 100 IU/mL human IL2 (Novartis) for 3days and retrovirally transduced with TAK1a or b gene fused withDNGFR or with DNGFR alone (control). CD8þ T cells werepurified using the CD8þ T Cell Isolation Kit (Miltenyi Biotec)and subsequently expanded using aAPCs as described previously(38, 39, 41). Briefly, CD8þ T cells were plated at 2� 106 cells perwell in RPMI 1640 supplemented with 10% human AB serum.Where indicated, A24-aAPCs were pulsed with 1 mg/mL A24-restricted WT1235–243 peptide (GenWay Biotech) for 6 hours atroom temperature and irradiated at 200Gybefore use. B57-aAPCswere always usedwithout any peptide pulse. Starting the next day,10 IU/mL IL2 (Novartis) and 10 ng/mL IL15 (Peprotech) wereadded to the cultures every 3 days. T cells were harvested, counted,and restimulated every week. T-cell analysis was performed 1 daybefore or on the day of restimulation.
Flow cytometry analysisCell surface molecules were stained with PC5-conjugated anti-
CD8mAb (clone B9.11; BeckmanCoulter), FITC-conjugated anti-NGFR mAb (clone ME20.4; Biolegend), APC-conjugated anti-CD107a mAb (clone H4A3; Biolegend), PE-conjugated anti-CD3mAb (clone UCHT1; Becton Dickinson), and FITC-conjugatedanti-TRBV5.1 mAb (Beckman Coulter). Stained cells were ana-lyzedwithflow cytometry (BDBiosciences), and data analysiswasperformed using FlowJo Version 7.6.4 software (TreeStar) asdescribed previously (39).
Tetramer stainingBiotinylated HLA-A24/WT1235–243 monomers and A24/Survi-
vin80–88 monomers were kindly provided by the NIH TetramerCore Facility. The monomers were multimerized using PE-con-jugated streptavidin (Thermo Fisher Scientific) and utilized asdescribed previously (38, 39, 41). A24/Survivin80 tetramer wasalways used as a control. Where indicated, data shown are gatedon DNGFRþ cells. Structural avidity was determined by stainingwith graded concentrations of A24/WT1235 tetramer (20).
TCR Reactivity and Chain Centricity
www.aacrjournals.org Cancer Immunol Res; 3(9) September 2015 1071
on July 7, 2018. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Published OnlineFirst May 5, 2015; DOI: 10.1158/2326-6066.CIR-14-0222

ELISPOT analysisCytokine ELISPOT assays were conducted as described previ-
ously (38, 39, 41). Briefly, T cells were incubated with 2� 104 perwell of indicated aAPCs, T2-A24 cells, or 3 � 104 per well ofCD34þ cells in the presence or absence of peptide for 20 to 24hours at 37�C. HLA-A24–restricted HIV env584–592 peptide wasused as a control. Functional avidity was studied using T2-A24cells pulsedwith graded concentrations of A24/WT1235 peptide asstimulators as reported previously (20, 39).
Cytotoxicity assayStandard cytotoxicity assay was conducted as described previ-
ously (38, 39). Briefly, 51Cr-labeled 5.0� 103 indicated aAPCs orT2-A24 cells loadedwithpeptidesweremixedwith gene-modifiedCD8þ T cells for 6 hours at 37�C in a 96-well round-bottomplate.A24/HIV env584 peptide was used as a control. Percent specificlysis was calculated using the following formula: [(experimentalresult – spontaneous release)/(maximum release – spontaneousrelease)] � 100%.
In silico sequence analysisThe ScanProsite tool (http://prosite.expasy.org/scanprosite/)
was used to search for human-derived peptide sequences contain-ing critical amino acid residues recognized by A24/WT1235 TCRswithin the entire UniProtKB/Swiss-Prot database (release2015_02 of 04-Feb-15 with 547,599 entries).
Statistical analysisStatistical analysis was performed using GraphPad Prism 6.0b.
To determinewhether two groupswere significantly different for agiven variable, analysis was performed using theWelch t test (two-sided). Comparative analyses between three or more differentgroups were achieved using repeated-measures ANOVA with theGreenhouse-Geisser correction, followed by Tukeymultiple com-parison test. P values < 0.05 were considered statistically signif-icant. Pearson correlation coefficients were utilized to assess thecorrelation between two independent variables. Values of r� 0.7were considered correlated.
ResultsTAK1b hemi-chain has a dominant role in A24/WT1235reactivity
To investigate whether the TAK1a (TRAV20�02/TRAJ33�01) orb (TRBV5-1�01/TRBJ2-1�01) chain has a dominant role in A24/WT1235 reactivity, peripheral T cells from four A24þ and twoA24�
donors were retrovirally transduced with TAK1a or b hemi-chainor a control gene (DNGFR alone). To mark hemi-chain–trans-duced T cells, each hemi-chain gene was fused to the DNGFR geneas stated in Materials and Methods. Following transduction andbefore stimulation, A24/WT1235 tetramer–positive cells weredetectable in TAK1b, but not TAK1a, hemi-chain–transducedCD8þ T cells in two of the four A24þ donors and one of the twoA24� donors (Supplementary Fig. S1).We previously reported onthe A24-aAPCs, which can expand HLA-A24–restricted antigen-specific T cells (42). To further confirm that the observed A24/WT1235 tetramer–positive cells were specific to A24/WT1235 pep-tide and not cross-reactive to the self-HLA complex, CD8þ T cellswere isolated and stimulated twice with A24-aAPCs loaded withA24/WT1235 peptide. In all 6 donors tested, A24/WT1235-specificTAK1b-transduced CD8þ T cells demonstrated significantlyincreased A24/WT1235 tetramer positivity compared with TAK1aor control transfectants (Fig. 1A, left and right).
TAK1b-transduced, but not TAK1a-transduced, T cells recog-nized exogenously pulsed A24/WT1235 peptide in both IFNgELISPOT (Fig. 1B, top) and standard killing assays (Fig. 1B,bottom), further confirming the A24/WT1235 specificity ofTAK1b-transduced T cells. The parental cell line of the aAPCs,K562, endogenously expresses WT1 protein. It has been demon-strated that K562 expresses normal proteasome machinery andcan naturally process and present HLA class I–restricted peptidesderived from endogenous antigens, such as WT1 (35, 41, 43).TAK1b-transduced, but not TAK1a-transduced, T cells were ableto recognize naturally processed and presented A24/WT1235 pep-tide in both IFNg ELISPOT analysis (Fig. 1C, top) and a standardcytotoxicity assay (Fig. 1C, bottom). However, the recognition ofendogenously processed and presented A24/WT1235 peptide wasnot as robust as exogenously pulsed A24/WT1235 peptide. Theseresults demonstrate that TAK1b, but not TAK1a, hemi-chain has adominant role in dictating A24/WT1235 specificity and that afraction of TAK1b-transduced T cells likely possess functionalavidity sufficient to recognize endogenously processed and pre-sented A24/WT1235 peptide.
TCRa chain repertoires reactive for A24/WT1235 andalloreactive for B57 in conjunction with TAK1b chain partiallybut incompletely overlap
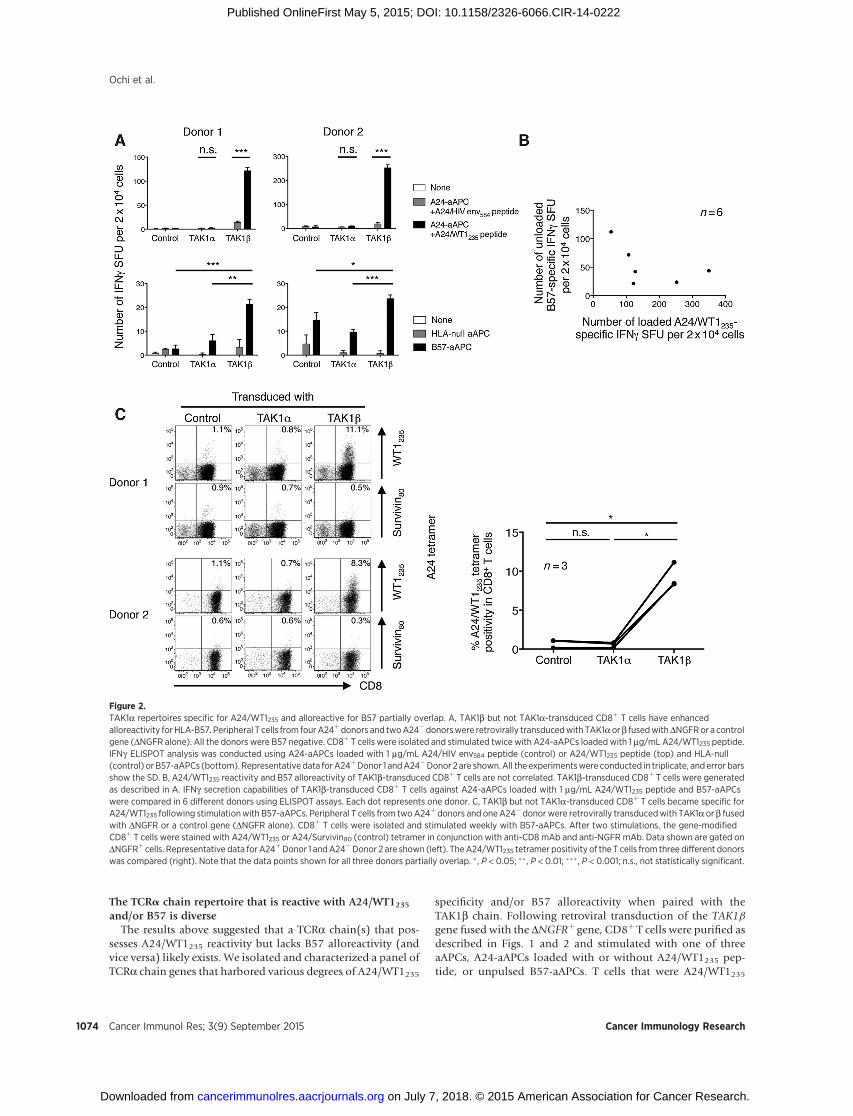
As we published previously, the TAK1 TCR cross-reacts withunrelated B57 molecules expressed in blood cells (35). We nextinvestigated whether TAK1a or b hemi-chain plays a dominantrole in B57 allo-recognition. Similar to the description abovein Fig. 1, peripheral T cells from four A24þ B57� and two A24�
B57� donors were transduced with TAK1a or b chain fused withDNGFR or a control (DNGFR alone) gene. CD8þ T cells werepurified and subsequently stimulated with A24-aAPCs pulsedwith A24/WT1235 peptide. After two stimulations, TAK1b-trans-duced, but not TAK1a-transduced, T cells from all 6 donorsdemonstrated A24/WT1235 specificity on IFNg ELISPOT anal-ysis (Fig. 2A, top). Furthermore, TAK1b transfectants consis-tently possessed enhanced B57 alloreactivity compared withendogenous B57 alloreactivity presented by control transfec-tants (Fig. 2A, bottom). Note that all the donors were B57-negative and that various levels of endogenous alloreactivity toB57 molecules were observed in the 6 donors. Because theidentity of the B57-bound peptide that is recognized by TAK1TCR is unknown, we were not able to stain the expanded T cellswith B57 tetramer. Importantly, there was no correlationbetween the observed A24/WT1235-specific and B57-specificIFNg production capabilities of the TAK1b-transduced T cellsderived from the 6 donors (Fig. 2B).
Next, peripheral T cells from two A24þ B57� donors and oneA24� B57� donor were transduced with a control (DNGFRalone) gene or with TAK1a or b chain fused with DNGFR. CD8þ
T cells were purified and stimulated with unpulsed B57-aAPCs.No A24-restricted stimulation was performed. Following twostimulations, A24/WT1235 tetramer–positive TAK1b-trans-duced, but not TAK1a-transduced, CD8þ T cells expanded inall 3 donors tested (Fig. 2C, left and right). Collectively, thesedata suggest that TAK1b, but not TAK1a, hemi-chain plays adominant role in B57 allo-recognition as well as in A24/WT1235recognition as shown in Fig. 1. Moreover, TCRa chain reper-toires reactive for A24/WT1235 and alloreactive for B57 inconjunction with TAK1b chain appear to overlap partially butnot completely.
Ochi et al.
Cancer Immunol Res; 3(9) September 2015 Cancer Immunology Research1072
on July 7, 2018. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Published OnlineFirst May 5, 2015; DOI: 10.1158/2326-6066.CIR-14-0222
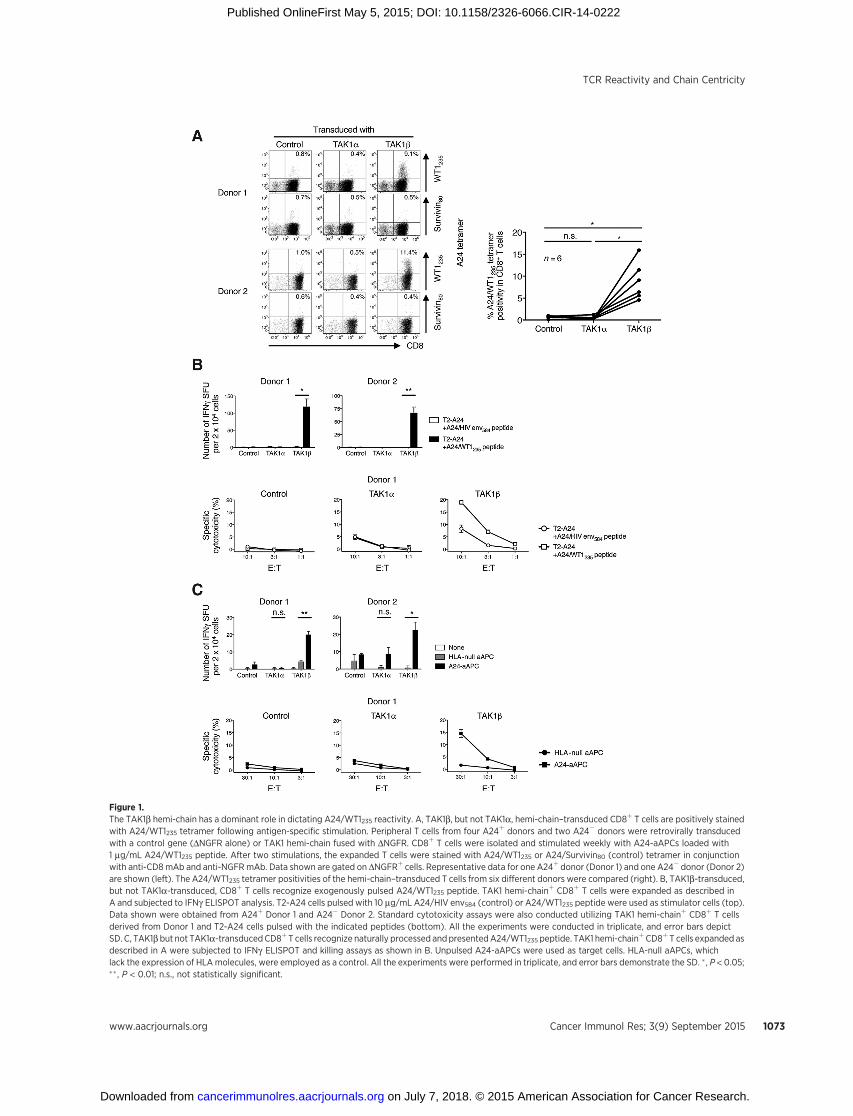
Figure 1.The TAK1b hemi-chain has a dominant role in dictating A24/WT1235 reactivity. A, TAK1b, but not TAK1a, hemi-chain–transduced CD8þ T cells are positively stainedwith A24/WT1235 tetramer following antigen-specific stimulation. Peripheral T cells from four A24þ donors and two A24� donors were retrovirally transducedwith a control gene (DNGFR alone) or TAK1 hemi-chain fused with DNGFR. CD8þ T cells were isolated and stimulated weekly with A24-aAPCs loaded with1 mg/mL A24/WT1235 peptide. After two stimulations, the expanded T cells were stained with A24/WT1235 or A24/Survivin80 (control) tetramer in conjunctionwith anti-CD8mAb and anti-NGFRmAb. Data shown are gated on DNGFRþ cells. Representative data for one A24þ donor (Donor 1) and one A24� donor (Donor 2)are shown (left). The A24/WT1235 tetramer positivities of the hemi-chain–transduced T cells from six different donors were compared (right). B, TAK1b-transduced,but not TAK1a-transduced, CD8þ T cells recognize exogenously pulsed A24/WT1235 peptide. TAK1 hemi-chainþ CD8þ T cells were expanded as described inA and subjected to IFNg ELISPOT analysis. T2-A24 cells pulsed with 10 mg/mL A24/HIV env584 (control) or A24/WT1235 peptide were used as stimulator cells (top).Data shown were obtained from A24þ Donor 1 and A24� Donor 2. Standard cytotoxicity assays were also conducted utilizing TAK1 hemi-chainþ CD8þ T cellsderived from Donor 1 and T2-A24 cells pulsed with the indicated peptides (bottom). All the experiments were conducted in triplicate, and error bars depictSD. C, TAK1b but not TAK1a-transducedCD8þT cells recognize naturally processed andpresentedA24/WT1235 peptide. TAK1 hemi-chainþCD8þ T cells expanded asdescribed in A were subjected to IFNg ELISPOT and killing assays as shown in B. Unpulsed A24-aAPCs were used as target cells. HLA-null aAPCs, whichlack the expression of HLAmolecules, were employed as a control. All the experiments were performed in triplicate, and error bars demonstrate the SD. � , P < 0.05;�� , P < 0.01; n.s., not statistically significant.
TCR Reactivity and Chain Centricity
www.aacrjournals.org Cancer Immunol Res; 3(9) September 2015 1073
on July 7, 2018. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Published OnlineFirst May 5, 2015; DOI: 10.1158/2326-6066.CIR-14-0222
The TCRa chain repertoire that is reactive with A24/WT1235and/or B57 is diverse
The results above suggested that a TCRa chain(s) that pos-sesses A24/WT1235 reactivity but lacks B57 alloreactivity (andvice versa) likely exists. We isolated and characterized a panel ofTCRa chain genes that harbored various degrees of A24/WT1235
specificity and/or B57 alloreactivity when paired with theTAK1b chain. Following retroviral transduction of the TAK1bgene fused with the DNGFRþ gene, CD8þ T cells were purified asdescribed in Figs. 1 and 2 and stimulated with one of threeaAPCs, A24-aAPCs loaded with or without A24/WT1235 pep-tide, or unpulsed B57-aAPCs. T cells that were A24/WT1235
Figure 2.TAK1a repertoires specific for A24/WT1235 and alloreactive for B57 partially overlap. A, TAK1b but not TAK1a-transduced CD8þ T cells have enhancedalloreactivity for HLA-B57. Peripheral T cells from fourA24þdonors and twoA24�donorswere retrovirally transducedwith TAK1aorb fusedwithDNGFRor a controlgene (DNGFR alone). All the donors were B57 negative. CD8þ T cells were isolated and stimulated twice with A24-aAPCs loaded with 1 mg/mL A24/WT1235 peptide.IFNg ELISPOT analysis was conducted using A24-aAPCs loaded with 1 mg/mL A24/HIV env584 peptide (control) or A24/WT1235 peptide (top) and HLA-null(control) or B57-aAPCs (bottom). Representative data forA24þDonor 1 andA24�Donor 2 are shown.All the experimentswere conducted in triplicate, and error barsshow the SD. B, A24/WT1235 reactivity and B57 alloreactivity of TAK1b-transduced CD8þ T cells are not correlated. TAK1b-transduced CD8þ T cells were generatedas described in A. IFNg secretion capabilities of TAK1b-transduced CD8þ T cells against A24-aAPCs loaded with 1 mg/mL A24/WT1235 peptide and B57-aAPCswere compared in 6 different donors using ELISPOT assays. Each dot represents one donor. C, TAK1b but not TAK1a-transduced CD8þ T cells became specific forA24/WT1235 following stimulationwith B57-aAPCs. Peripheral T cells from twoA24þ donors and one A24� donor were retrovirally transducedwith TAK1a or b fusedwith DNGFR or a control gene (DNGFR alone). CD8þ T cells were isolated and stimulated weekly with B57-aAPCs. After two stimulations, the gene-modifiedCD8þ T cells were stained with A24/WT1235 or A24/Survivin80 (control) tetramer in conjunction with anti-CD8 mAb and anti-NGFR mAb. Data shown are gated onDNGFRþ cells. Representative data for A24þDonor 1 andA24�Donor 2 are shown (left). TheA24/WT1235 tetramer positivity of the T cells from three different donorswas compared (right). Note that the data points shown for all three donors partially overlap. � , P < 0.05; �� , P < 0.01; ��� , P < 0.001; n.s., not statistically significant.
Ochi et al.
Cancer Immunol Res; 3(9) September 2015 Cancer Immunology Research1074
on July 7, 2018. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Published OnlineFirst May 5, 2015; DOI: 10.1158/2326-6066.CIR-14-0222
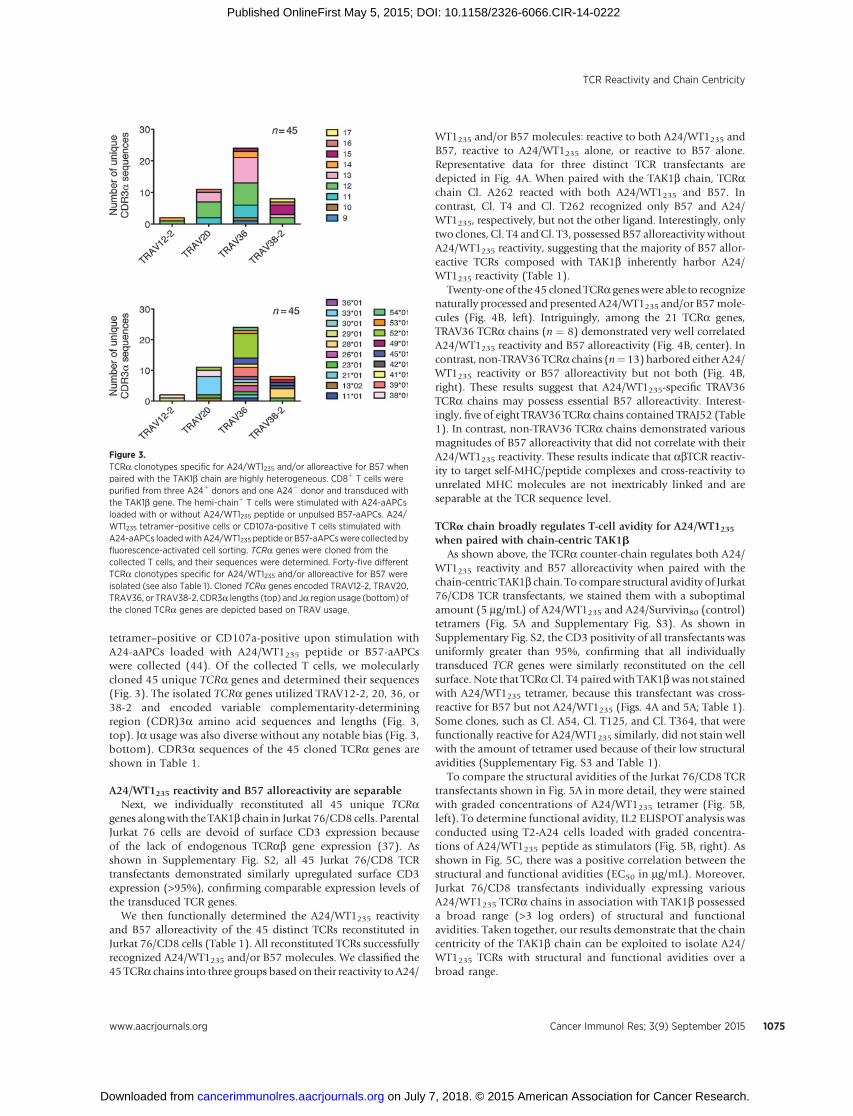
tetramer–positive or CD107a-positive upon stimulation withA24-aAPCs loaded with A24/WT1235 peptide or B57-aAPCswere collected (44). Of the collected T cells, we molecularlycloned 45 unique TCRa genes and determined their sequences(Fig. 3). The isolated TCRa genes utilized TRAV12-2, 20, 36, or38-2 and encoded variable complementarity-determiningregion (CDR)3a amino acid sequences and lengths (Fig. 3,top). Ja usage was also diverse without any notable bias (Fig. 3,bottom). CDR3a sequences of the 45 cloned TCRa genes areshown in Table 1.
A24/WT1235 reactivity and B57 alloreactivity are separableNext, we individually reconstituted all 45 unique TCRa
genes alongwith the TAK1b chain in Jurkat 76/CD8 cells. ParentalJurkat 76 cells are devoid of surface CD3 expression becauseof the lack of endogenous TCRab gene expression (37). Asshown in Supplementary Fig. S2, all 45 Jurkat 76/CD8 TCRtransfectants demonstrated similarly upregulated surface CD3expression (>95%), confirming comparable expression levels ofthe transduced TCR genes.
We then functionally determined the A24/WT1235 reactivityand B57 alloreactivity of the 45 distinct TCRs reconstituted inJurkat 76/CD8 cells (Table 1). All reconstituted TCRs successfullyrecognized A24/WT1235 and/or B57 molecules. We classified the45 TCRa chains into three groups based on their reactivity to A24/
WT1235 and/or B57 molecules: reactive to both A24/WT1235 andB57, reactive to A24/WT1235 alone, or reactive to B57 alone.Representative data for three distinct TCR transfectants aredepicted in Fig. 4A. When paired with the TAK1b chain, TCRachain Cl. A262 reacted with both A24/WT1235 and B57. Incontrast, Cl. T4 and Cl. T262 recognized only B57 and A24/WT1235, respectively, but not the other ligand. Interestingly, onlytwo clones, Cl. T4 andCl. T3, possessed B57 alloreactivity withoutA24/WT1235 reactivity, suggesting that the majority of B57 allor-eactive TCRs composed with TAK1b inherently harbor A24/WT1235 reactivity (Table 1).
Twenty-one of the 45 cloned TCRa geneswere able to recognizenaturally processed and presented A24/WT1235 and/or B57mole-cules (Fig. 4B, left). Intriguingly, among the 21 TCRa genes,TRAV36 TCRa chains (n ¼ 8) demonstrated very well correlatedA24/WT1235 reactivity and B57 alloreactivity (Fig. 4B, center). Incontrast, non-TRAV36 TCRa chains (n¼ 13) harbored either A24/WT1235 reactivity or B57 alloreactivity but not both (Fig. 4B,right). These results suggest that A24/WT1235-specific TRAV36TCRa chains may possess essential B57 alloreactivity. Interest-ingly, five of eight TRAV36 TCRa chains contained TRAJ52 (Table1). In contrast, non-TRAV36 TCRa chains demonstrated variousmagnitudes of B57 alloreactivity that did not correlate with theirA24/WT1235 reactivity. These results indicate that abTCR reactiv-ity to target self-MHC/peptide complexes and cross-reactivity tounrelated MHC molecules are not inextricably linked and areseparable at the TCR sequence level.
TCRa chain broadly regulates T-cell avidity for A24/WT1235when paired with chain-centric TAK1b
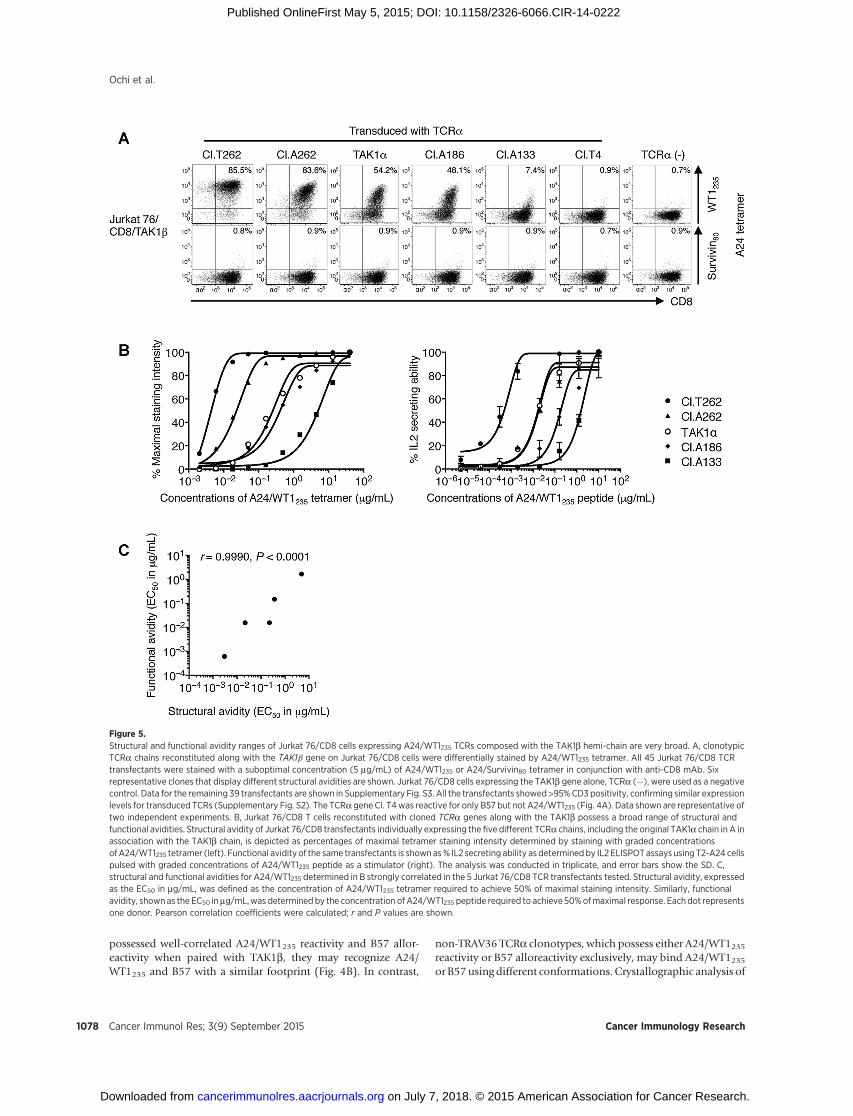
As shown above, the TCRa counter-chain regulates both A24/WT1235 reactivity and B57 alloreactivity when paired with thechain-centric TAK1b chain. To compare structural avidity of Jurkat76/CD8 TCR transfectants, we stained them with a suboptimalamount (5 mg/mL) of A24/WT1235 and A24/Survivin80 (control)tetramers (Fig. 5A and Supplementary Fig. S3). As shown inSupplementary Fig. S2, the CD3 positivity of all transfectants wasuniformly greater than 95%, confirming that all individuallytransduced TCR genes were similarly reconstituted on the cellsurface. Note that TCRaCl. T4 paired with TAK1bwas not stainedwith A24/WT1235 tetramer, because this transfectant was cross-reactive for B57 but not A24/WT1235 (Figs. 4A and 5A; Table 1).Some clones, such as Cl. A54, Cl. T125, and Cl. T364, that werefunctionally reactive for A24/WT1235 similarly, did not stain wellwith the amount of tetramer used because of their low structuralavidities (Supplementary Fig. S3 and Table 1).
To compare the structural avidities of the Jurkat 76/CD8 TCRtransfectants shown in Fig. 5A in more detail, they were stainedwith graded concentrations of A24/WT1235 tetramer (Fig. 5B,left). To determine functional avidity, IL2 ELISPOT analysis wasconducted using T2-A24 cells loaded with graded concentra-tions of A24/WT1235 peptide as stimulators (Fig. 5B, right). Asshown in Fig. 5C, there was a positive correlation between thestructural and functional avidities (EC50 in mg/mL). Moreover,Jurkat 76/CD8 transfectants individually expressing variousA24/WT1235 TCRa chains in association with TAK1b possesseda broad range (>3 log orders) of structural and functionalavidities. Taken together, our results demonstrate that the chaincentricity of the TAK1b chain can be exploited to isolate A24/WT1235 TCRs with structural and functional avidities over abroad range.
Figure 3.TCRa clonotypes specific for A24/WT1235 and/or alloreactive for B57 whenpaired with the TAK1b chain are highly heterogeneous. CD8þ T cells werepurified from three A24þ donors and one A24� donor and transduced withthe TAK1b gene. The hemi-chainþ T cells were stimulated with A24-aAPCsloaded with or without A24/WT1235 peptide or unpulsed B57-aAPCs. A24/WT1235 tetramer–positive cells or CD107a-positive T cells stimulated withA24-aAPCs loadedwithA24/WT1235 peptide orB57-aAPCswere collectedbyfluorescence-activated cell sorting. TCRa genes were cloned from thecollected T cells, and their sequences were determined. Forty-five differentTCRa clonotypes specific for A24/WT1235 and/or alloreactive for B57 wereisolated (see also Table 1). Cloned TCRa genes encoded TRAV12-2, TRAV20,TRAV36, or TRAV38-2. CDR3a lengths (top) and Ja region usage (bottom) ofthe cloned TCRa genes are depicted based on TRAV usage.
TCR Reactivity and Chain Centricity
www.aacrjournals.org Cancer Immunol Res; 3(9) September 2015 1075
on July 7, 2018. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Published OnlineFirst May 5, 2015; DOI: 10.1158/2326-6066.CIR-14-0222

Newly isolated A24/WT1235 TCRs possess potential cross-reactivity to homologous peptides but lack reactivity to CD34þ
hematopoietic cellsRecent clinical trials have demonstrated that cross-reactivity
of antitumor TCRs to other peptides presented by self-HLAmolecules can cause unwanted serious adverse events inpatients treated with TCR gene therapy (15, 16). Alaninesubstitution scanning identified that the first, second, eighth,and ninth amino acid residues of the A24/WT1235 peptideare critical for recognition by three different TCRs, Cl. T262,
Cl. A262, and Cl. T53, which have sufficient affinity to recog-nize naturally processed and presented A24/WT1235 peptide(Fig. 6A). Based on this, an in silico search was conducted toidentify protein sequences that contain the "CMXXXXXNL"sequence using the ScanProsite tool (45, 46). We synthesizedeleven human-derived peptides homologous to A24/WT1235 asshown in Supplementary Table S1. Jurkat 76/CD8 TCR trans-fectants, Cl. T262, Cl. A262, and Cl. T53, reacted to some ofthese peptides with different specificity and potency (Fig. 6B). Itshould be noted, however, that it is currently unknown whether
Table 1. Variable, joining region, and CDR3a sequences of TCRa clonotypes and their reactivities to unloaded A24-aAPC, B57-aAPC, and A24-aAPC loaded withA24/WT1235 peptide
Targets
Clone TRAV CDR3a TRAJUnloaded A24-
aAPCUnloaded B57-
aAPCA24-aAPC loaded with A24/
WT1235 peptide
TAK1a 20�02 CAVQAVDSNYQLIW 33�01 � þþ þþþTCRa chains isolated following stimulation with A24-aAPC loaded with A24/WT1235 peptideCl.B210 20�02 CAVQAWGYQKVTF 13�02 � � þþCl.A133 20�02 CAVQAYNQGGKLIF 23�01 � � þCl.A160 20�02 CAVQAHSNYQLIW 33�01 � þ þþþCl.A106 20�02 CAVQARDSNYQLIW 33�01 � þ þþþCl.A98 20�02 CAVQAGWRSNYQLIW 33�01 � � þþCl.A54 20�02 CAYVAGGTSYGKLTF 52�01 � � þþCl.A156 36/DV7�01 CAVVVHNAGNMLTF 39�01 � � þþþCl.A253 36/DV7�04 CAVIKGYNFNKFYF 21�01 � � þþCl.A128 36/DV7�04 CAAAYYNQGGKLIF 23�01 � � þþCl.A245 36/DV7�04 CAVEWDYGQNFVF 26�01 � þ þþCl.A260 36/DV7�04 CAPYSGAGSYQLTF 28�01 � � þþCl.A224 36/DV7�04 CAGVPNRDDKIIF 30�01 � � þþCl.T204 36/DV7�04 CAVVTHNAGNMLTF 39�01 � � þþCl.T216 36/DV7�04 CAQGPNNAGNMLTF 39�01 � � þþCl.A262 36/DV7�04 CAVQNAGGTSYGKLTF 52�01 þ þ þþþCl.T53 36/DV7�04 CAVITGGTSYGKLTF 52�01 þ þ þþþCl.T149 36/DV7�04 CAGNAGGTSYGKLTF 52�01 þ � þþþCl.T193 36/DV7�04 CAVTNAGGTSYGKLTF 52�01 � þ þþþCl.T168 36/DV7�04 CAVLAGGTSYGKLTF 52�01 þ � þþþCl.A217 36/DV7�04 CAVGSGGTSYGKLTF 52�01 � � þþCl.T228 36/DV7�04 CAVAAGGTSYGKLTF 52�01 � � þþCl.T154 36/DV7�04 CAVEALIQGAQKLVF 54�01 � � þCl.A186 38-2/DV8�01 CAYRSGRENSGGSNYKLTF 53�01 � þ þþþ
TCRa chains isolated following stimulation with unloaded A24-aAPCCl.T272 20�02 CAVQARRSNYQLIW 33�01 � þ þþþCl.T262 20�02 CAVQALRNNAGNNRKLIW 38�01 þþ � þþþCl.T389 20�02 CAVQAGRGNNRKLIW 38�01 � � þCl.T125 36/DV7�01 CAYSSNSGYALNF 41�01 � � þCl.T243 36/DV7�04 CAVLTQTGANNLFF 36�01 þ þ þþþCl.T258 38-2/DV8�01 CAYRSSYSGAGSYQLTF 28�01 � þ þCl.T248 38-2/DV8�01 CAYRSANYYGGSQGNLIF 42�01 � � þ
TCRa chains isolated following stimulation with unloaded B57-aAPCCl.B302 12-2�02 CAVSRRSGNTPLVF 29�01 � � þþþCl.T338 12-2�02 CAVNSRRGGNNRKLIW 38�01 � � þCl.T367 20�02 CAVQAWDSNYQLIW 33�01 � þþ þþþCl.T24 36/DV7�01 CALYSGYSTLTF 11�01 � � þCl.T16 36/DV7�01 CAVLGLRGGADGLTF 45�01 � þ þþCl.T341 36/DV7�04 CARVYGQNFVF 26�01 � � þþCl.B309 36/DV7�04 CAVAGGGADGLTF 45�01 � � þCl.T353 36/DV7�04 CAVNAGGTSYGKLTF 52�01 � � þþCl.T364 36/DV7�04 CAVPLTNSGGSNYKLTF 53�01 � � þCl.T4 38-2/DV8�01 CAYRSHYNQGGKLIF 23�01 � þ �Cl.T20 38-2/DV8�01 CAYRSAYSGAGSYQLTF 28�01 � þþ þCl.T28 38-2/DV8�01 CAYRTIYSGAGSYQLTF 28�01 � þ þþCl.T3 38-2/DV8�01 CAYRSPWGADGLTF 45�01 � þ �Cl.T1 38-2/DV8�01 CAYRRRWTGNQFYF 49�01 � þ þ
NOTE: A24/WT1235 reactivity and B57 alloreactivity of 45 Jurkat 76/CD8 TCR transfectants were measured using A24-aAPCs loaded with or without 1 mg/mL A24/WT1235 peptide andB57-aAPCs as stimulators in IL2 ELISPOTassays. All the experimentswere performed in triplicate, and themean number of IL2 SFUs per 4.0� 104
cellswas calculated.þþþ,>400SFU/4.0� 104 cells;þþ, 101–400SFU/4.0� 104 cells;þ, 10–100 SFU/4.0� 104 cells;�,<10 SFU/4.0� 104 cells. All the transfectantsproduced <5 SFU per 4.0 � 104 cells when stimulated with HLA-null aAPCs (data not shown).Abbreviation: SFU, spot-forming units.
Ochi et al.
Cancer Immunol Res; 3(9) September 2015 Cancer Immunology Research1076
on July 7, 2018. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Published OnlineFirst May 5, 2015; DOI: 10.1158/2326-6066.CIR-14-0222

these peptides are naturally processed and presented by A24 innormal tissues.
It is known that WT1 is expressed in human primary CD34þ
hematopoietic cells (27, 30). We studied the reactivity of twoJurkat 76/CD8 TCR transfectants, Cl. T262 and Cl. A262, topurified CD34þ cells. Neither TCR transfectant showed any reac-tivity to unpulsed A24þ CD34þ cells. In contrast, both transfec-tants recognized A24/WT1235 peptide exogenously pulsed intoA24þ CD34þ cells but not into A24� CD34þ cells (Fig. 6C).Collectively, our results suggest that, although newly cloned A24/WT1235 TCRs, Cl. T262 and Cl. A262, possess potential cross-reactivity to A24/WT1235-related peptides, they did not recognizehuman primary HLA-A24þ CD34þ hematopoietic cells.
DiscussionThe TAK1a chain encodes TRAV20�02 and recognizes both
the A24/WT1235 and B57 molecules when paired with TAK1bchain. Although TCRa chain Cl. T262, which has been newlycloned in this study, utilizes the same TRAV20�02, it reacts withA24/WT1235, but not with B57, molecules. This separationobviously suggests that the hypervariable CDR3a sequencesaffect the structure of the TCR, thereby regulating antigenspecificity as well as alloreactivity in a separable manner, andthat self A24-restricted WT1235 specificity and B57 cross-reac-tivity are not inextricable.
Previous crystallographic studies have demonstrated that, inaddition to the hypervariable CDR3 loop, germline-encoded
variable CDR1/2 regions can play important roles in the recog-nition of MHC/peptide complexes (47, 48). Dai and colleaguesdemonstrated that amino acid substitutions in the variableCDR1/2 regions of murine YAe62 TCRs resulted in an alterationof alloreactivity for unrelated MHC/peptide complexes (49).In line with these studies, we showed that, among the 21 TAK1achains that recognized naturally processed and presented A24/WT1235 and/or B�57:01 peptides, 8 TRAV36 TCRa counter-chains harbored well-correlated A24/WT1235 reactivity and B57alloreactivity. TRAJ52 may be involved in the correlated A24/WT1235 and B57 reactivity in association with the TRAV36 whenpaired with TAK1b. In contrast, the remaining 13 non-TRAV36TCRa chains demonstrated either exclusive A24/WT1235 reactivityor B57 alloreactivity depending upon their CDR3a sequences(Fig. 4). These findings indicate that, in concert with CDR3aloops, TCR CDR1/2a regions can affect both A24/WT1235 reac-tivity and B57 cross-reactivity when paired with the chain-centricTAK1b chain.
Although further research is required, the results obtained usingfunctional assays imply that the TAK1b hemi-chain has a dom-inant role in dictating B57 cross-reactivity (Fig. 2). It is thus highlylikely that TAK1b has a dominant role in dictating both A24/WT1235 reactivity and B57 cross-reactivity. A TCR with similarattributes has been reported previously. Crystallographic analysisof another chain-centric TCR, clone LC13 that is HLA-B8/EBVEBNA3A339-specific and B44 alloreactive, has demonstrated thatit recognizes both B8/EBV EBNA3A339 and B44 using similarconformation (10). Because isolated TRAV36 TCRa chains
Figure 4.B57 alloreactivity is separable from A24/WT1235 reactivity by changing the TCRa chains paired with the TAK1b chain. A, cloned TCRa chains demonstrateA24/WT1235 reactivity and/or B57 alloreactivity when reconstitutedwith the TAK1b chain on Jurkat 76/CD8 cells. The A24/WT1235 reactivity andB57 alloreactivity ofall 45 Jurkat 76/CD8 TCR transfectants were measured using the indicated aAPCs as stimulators in IL2 ELISPOT assays (Table 1). The A24/WT1235 reactivityand B57 alloreactivity of three representative TCRa clones, Cl. A262, Cl. T4, and Cl. T262, are shown. All the experiments were conducted in triplicate, and errorbars depict the SD. Data shown are representative of two independent experiments. B, the A24/WT1235 reactivity and B57 alloreactivity of TRAV36 butnot non-TRAV36 TCRa genes correlated. The A24/WT1235 reactivity and B57 alloreactivity of 21 TCRa chains (Table 1) that recognize naturally processed andpresentedA24/WT1235 peptide and/or B57moleculeswhen pairedwith TAK1bwere compared (left). The 21 TCRa chainswere divided into 2groups, 8 TRAV36 TCRa(center) and 13 non-TRAV36 TCRa genes (right). Each dot represents one donor. Pearson correlation coefficients were calculated; r and P values are shown.
TCR Reactivity and Chain Centricity
www.aacrjournals.org Cancer Immunol Res; 3(9) September 2015 1077
on July 7, 2018. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Published OnlineFirst May 5, 2015; DOI: 10.1158/2326-6066.CIR-14-0222
possessed well-correlated A24/WT1235 reactivity and B57 allor-eactivity when paired with TAK1b, they may recognize A24/WT1235 and B57 with a similar footprint (Fig. 4B). In contrast,
non-TRAV36 TCRa clonotypes, which possess either A24/WT1235reactivity or B57 alloreactivity exclusively, may bind A24/WT1235or B57 using different conformations. Crystallographic analysis of
Figure 5.Structural and functional avidity ranges of Jurkat 76/CD8 cells expressing A24/WT1235 TCRs composed with the TAK1b hemi-chain are very broad. A, clonotypicTCRa chains reconstituted along with the TAK1b gene on Jurkat 76/CD8 cells were differentially stained by A24/WT1235 tetramer. All 45 Jurkat 76/CD8 TCRtransfectants were stained with a suboptimal concentration (5 mg/mL) of A24/WT1235 or A24/Survivin80 tetramer in conjunction with anti-CD8 mAb. Sixrepresentative clones that display different structural avidities are shown. Jurkat 76/CD8 cells expressing the TAK1b gene alone, TCRa (�), were used as a negativecontrol. Data for the remaining 39 transfectants are shown in Supplementary Fig. S3. All the transfectants showed >95%CD3 positivity, confirming similar expressionlevels for transduced TCRs (Supplementary Fig. S2). The TCRa gene Cl. T4was reactive for only B57 but not A24/WT1235 (Fig. 4A). Data shown are representative oftwo independent experiments. B, Jurkat 76/CD8 T cells reconstituted with cloned TCRa genes along with the TAK1b possess a broad range of structural andfunctional avidities. Structural avidity of Jurkat 76/CD8 transfectants individually expressing the five different TCRa chains, including the original TAK1a chain in A inassociation with the TAK1b chain, is depicted as percentages of maximal tetramer staining intensity determined by staining with graded concentrationsofA24/WT1235 tetramer (left). Functional avidity of the same transfectants is shownas% IL2 secreting ability as determined by IL2 ELISPOTassays using T2-A24 cellspulsed with graded concentrations of A24/WT1235 peptide as a stimulator (right). The analysis was conducted in triplicate, and error bars show the SD. C,structural and functional avidities for A24/WT1235 determined in B strongly correlated in the 5 Jurkat 76/CD8 TCR transfectants tested. Structural avidity, expressedas the EC50 in mg/mL, was defined as the concentration of A24/WT1235 tetramer required to achieve 50% of maximal staining intensity. Similarly, functionalavidity, shownas the EC50 inmg/mL,was determined by the concentration ofA24/WT1235 peptide required to achieve 50%ofmaximal response. Eachdot representsone donor. Pearson correlation coefficients were calculated; r and P values are shown.
Ochi et al.
Cancer Immunol Res; 3(9) September 2015 Cancer Immunology Research1078
on July 7, 2018. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Published OnlineFirst May 5, 2015; DOI: 10.1158/2326-6066.CIR-14-0222

these distinct TCRa chains paired with the TAK1b chain iswarranted.
Multiple clonotypic TCRa counter-chains were able to recog-nize A24/WT1235 and/or B57 when paired with TAK1b, and these
TCRs possessed a very wide range (>3 log orders) of structural andfunctional avidities for A24/WT1235. Of note, TCRa hemi-chains,such as Cl. A262 and Cl. T262, were able to recognize naturallyprocessed and presented A24/WT1235 peptidewhen reconstituted
Figure 6.Newly cloned A24/WT1235 TCRs possess potential cross-reactivity to homologous peptides but lack reactivity to CD34þ hematopoietic cells. A, alanine substitutionanalysis identified A24/WT1235 peptide residues important for recognition by A24/WT1235 TCRs. Each residue in the A24/WT1235 peptide sequence was sequentiallyreplaced by an alanine residue. The reactivity of Jurkat 76/CD8 cells reconstituted with Cl. T262, Cl. A262, or Cl. T53 along with TAK1b is shown as % IL2 maximumsecretion capacity as determined by IL2 ELISPOT assays using T2-A24 cells pulsed with 50 mg/mL parental wild-type A24/WT1235 or each alanine substitutedpeptide. A24/HIV env584 was used as a negative control. Note that these three Jurkat 76/CD8 transfectants possess functional avidity sufficient to recognizeunpulsed A24-aAPCs (Table 1). The experiments were conducted in triplicate, and error bars depict the SD. � , P < 0.05; ��, P < 0.01. B, A24/WT1235 TCRs possesspotential cross-reactivity to peptides homologous to wild-type A24/WT1235. Jurkat 76/CD8 cells transfectants expressing Cl. T262, Cl. A262, or Cl. T53 werestimulated as in A. The experiments were conducted in triplicate, and error bars show the SD. Data shown are representative of two independent experiments.Sequences of 12 A24/WT1235-related peptides, including the wild-type A24/WT1235 peptide, are shown in Supplementary Table S1. C, A24/WT1235-specific TCRs didnot recognize human primary CD34þ hematopoietic cells. Two A24/WT1235-specific Jurkat 76/CD8 transfectants, Cl. T262 and Cl. A262, were studied for theirreactivity to human cord blood CD34þ cells purified from two A24þ and one A24� donors. IL2 ELISPOT assays were conducted using purified CD34þ cells asstimulator cells. CD34þ cells pulsed with 50 mg/mL A24/HIV env584 or A24/WT1235 peptide were used as a negative or positive control. The experiments wereconducted in triplicate, and error bars show the SD.
TCR Reactivity and Chain Centricity
www.aacrjournals.org Cancer Immunol Res; 3(9) September 2015 1079
on July 7, 2018. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Published OnlineFirst May 5, 2015; DOI: 10.1158/2326-6066.CIR-14-0222

with TAK1b on Jurkat 76/CD8 cells, suggesting that they possesssufficient affinity to recognize tumor cells in the presence of CD8coreceptors (Table 1). When reconstituted along with TAK1b inthe absence of CD8 molecules, however, none of the isolatedTCRa chains were stained with A24/WT1235 tetramer or recog-nized naturally processed and presented A24/WT1235 peptide(data not shown). High-affinity HLA class I–restricted TCRs thatcan recognize endogenously processed and presented peptides inthe absence of CD8 coengagement do exist andhave been isolatedfor various antigens (50). We previously reported that T cells withhigher avidities preferentially grew from A2-restricted antigen-specific CD8þ T cells that had been stimulated with aAPCsexpressing mutated HLA-A2 molecules that cannot interact withCD8 molecules (39). Stimulation of TAK1b-transduced CD8þ Tcells with aAPCs expressing mutated A24 that cannot associatewith CD8 may enrich TAK1bþ A24/WT1235 CD8þ T cells withhigher avidity and facilitate the cloning of high-affinity TCRachains that do not require CD8 coengagement to recognize targetcells (39).
The selection of TCRs with minimal on-target/off-tumor andoff-target toxicities is critical to conduct safe TCR gene therapy(5, 7). We have demonstrated that newly isolated A24/WT1235TCRs do not recognize A24þCD34þhematopoietic cells (Fig. 6C).And yet, because WT1 is expressed in other normal cells, such aspodocytes in the kidney and mesothelial cells (26, 28), potentialtoxicity to these tissues needs to be carefully monitored in anyimmunotherapy, especially adoptive T-cell therapy targetingWT1. Off-target adverse events associated with the infusion ofredirected T cells are thought to be due, at least partly, to the cross-reactivity of the transducedTCR itself or tomispaired TCRs. In fact,severe adverse events associated with the infusion of redirected Tcells that are cross-reactive for other antigens have been reported(15, 16). We have shown that it is possible to eliminate inherentalloreactivity from antigen-specific TCRs at the molecular levelby exploiting TCR chain centricity. However, newly cloned A24/WT1235 TCRs containing TAK1b chain possessed potential cross-reactivity to peptides homologous to A24/WT1235, although it isunknown whether these peptides are endogenously processedand presented by A24 in normal tissues (Fig. 6B). These findingssuggest that we cannot eliminate the possibility that TCRs lackingapparent cross-reactivity may still acquire de novo cross-reactivity
for self/non-selfHLAcomplexes.However, preparing a set of TCRswith the same antigen specificity and different cross-reactivity byexploiting TCR chain centricity would benefit cancer patients byenlarging the patient population that can be treated using TCRgene therapy.
Disclosure of Potential Conflicts of InterestS. Tanaka is an employee of Takara Bio, Inc. No potential conflicts of interest
were disclosed by the other authors.
Authors' ContributionsConception and design: T. Ochi, N. HiranoDevelopment of methodology: T. OchiAcquisition of data (provided animals, acquired and managed patients,provided facilities, etc.): T. Ochi, M. Nakatsugawa, K. Chamoto, Y. Yamashita,T. Guo, H. Fujiwara, M. YasukawaAnalysis and interpretation of data (e.g., statistical analysis, biostatistics,computational analysis): T. Ochi, N. HiranoWriting, review, and/or revision of the manuscript: T. Ochi, M. Nakatsugawa,K. Chamoto, Y. Yamashita, T. Guo, H. Fujiwara, M. Yasukawa, M.O. Butler,N. HiranoAdministrative, technical, or material support (i.e., reporting or organizingdata, constructing databases): S. Tanaka, H. Fujiwara, M. Yasukawa, M.O.ButlerStudy supervision: N. Hirano
AcknowledgmentsJurkat 76 cells were a generous gift from Dr. Heemskerk, Leiden University
Medical Centre. Tetramers were kindly provided by the NIH tetramer corefacility.
Grant SupportThis work was supported by the following: NIH grant R01 CA148673 (to N.
Hirano); Ontario Institute for Cancer Research Clinical Investigator Award IA-039 (to N. Hirano); The Princess Margaret Cancer Foundation (to M.O. ButlerandN.Hirano);Guglietti Fellowship Award (to T.Ochi); Knudson PostdoctoralFellowship (toK.Chamoto); and the Frederick Banting andCharles Best CanadaGraduate Scholarship (to T. Guo).
The costs of publication of this articlewere defrayed inpart by the payment ofpage charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Received November 23, 2014; revised March 25, 2015; accepted April 15,2015; published OnlineFirst May 5, 2015.
References1. Stritesky GL, Jameson SC, Hogquist KA. Selection of self-reactive T cells in
the thymus. Annu Rev Immunol 2012;30:95–114.2. Klein L, HinterbergerM,Wirnsberger G, Kyewski B. Antigen presentation in
the thymus for positive selection and central tolerance induction. Nat RevImmunol 2009;9:833–44.
3. Barrett DM, Singh N, Porter DL, Grupp SA, June CH. Chimeric antigenreceptor therapy for cancer. Annu Rev Med 2014;65:333–47.
4. Kershaw MH, Westwood JA, Darcy PK. Gene-engineered T cells for cancertherapy. Nat Rev Cancer 2013;13:525–41.
5. Turtle CJ, Hudecek M, Jensen MC, Riddell SR. Engineered T cells for anti-cancer therapy. Curr Opin Immunol 2012;24:633–9.
6. Stauss HJ, Morris EC. Immunotherapy with gene-modified T cells:limiting side effects provides new challenges. Gene Ther 2013;20:1029–32.
7. Hinrichs CS, Restifo NP. Reassessing target antigens for adoptive T-celltherapy. Nat Biotechnol 2013;31:999–1008.
8. Amir AL, D'Orsogna LJ, Roelen DL, van Loenen MM, Hagedoorn RS, deBoer R, et al. Allo-HLA reactivity of virus-specific memory T cells iscommon. Blood 2010;115:3146–57.
9. Burrows SR, Khanna R, Burrows JM,MossDJ. An alloresponse in humans isdominated by cytotoxic T lymphocytes (CTL) cross-reactive with a singleEpstein-Barr virus CTL epitope: implications for graft-versus-host disease. JExp Med 1994;179:1155–61.
10. MacdonaldWA, Chen Z, Gras S, Archbold JK, Tynan FE, Clements CS, et al.T cell allorecognition viamolecularmimicry. Immunity 2009;31:897–908.
11. Morice A, Charreau B,Neveu B, Brouard S, Soulillou JP, BonnevilleM, et al.Cross-reactivity of herpesvirus-specific CD8 T cell lines toward allogeneicclass I MHC molecules. PLoS One 2010;5:e12120.
12. Rist M, Smith C, Bell MJ, Burrows SR, Khanna R. Cross-recognition of HLADR4 alloantigen by virus-specific CD8þ T cells: a new paradigm for self-/nonself-recognition. Blood 2009;114:2244–53.
13. Matis LA, Longo DL, Hedrick SM, Hannum C, Margoliash E, Schwartz RH.Clonal analysis of themajor histocompatibility complex restriction and thefine specificity of antigen recognition in the T cell proliferative response tocytochrome C. J Immunol 1983;130:1527–35.
14. Suchin EJ, Langmuir PB, Palmer E, Sayegh MH, Wells AD, Turka LA.Quantifying the frequency of alloreactive T cells in vivo: new answers toan old question. J Immunol 2001;166:973–81.
Ochi et al.
Cancer Immunol Res; 3(9) September 2015 Cancer Immunology Research1080
on July 7, 2018. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Published OnlineFirst May 5, 2015; DOI: 10.1158/2326-6066.CIR-14-0222

15. Linette GP, Stadtmauer EA, Maus MV, Rapoport AP, Levine BL, EmeryL, et al. Cardiovascular toxicity and titin cross-reactivity of affinity-enhanced T cells in myeloma and melanoma. Blood 2013;122:863–71.
16. Morgan RA, Chinnasamy N, Abate-Daga D, Gros A, Robbins PF, Zheng Z,et al. Cancer regression and neurological toxicity following anti-MAGE-A3TCR gene therapy. J Immunother 2013;36:133–51.
17. Miles JJ, Douek DC, Price DA. Bias in the alphabeta T-cell repertoire:implications for disease pathogenesis and vaccination. Immunol Cell Biol2011;89:375–87.
18. Turner SJ, Doherty PC, McCluskey J, Rossjohn J. Structural determinants ofT-cell receptor bias in immunity. Nat Rev Immunol 2006;6:883–94.
19. Bouneaud C, Kourilsky P, Bousso P. Impact of negative selection on the Tcell repertoire reactive to a self-peptide: a large fraction of T cell clonesescapes clonal deletion. Immunity 2000;13:829–40.
20. Nakatsugawa M, Yamashita Y, Ochi T, Tanaka S, Chamoto K, Guo T, et al.Specific roles of each TCR hemichain in generating functional chain-centricTCR. J Immunol 2015;194:3487–500.
21. Stadinski BD, Trenh P, Smith RL, Bautista B, Huseby PG, Li G, et al. A rolefor differential variable gene pairing in creating T cell receptors specific forunique major histocompatibility ligands. Immunity 2011;35:694–704.
22. Yokosuka T, Takase K, Suzuki M, Nakagawa Y, Taki S, Takahashi H, et al.Predominant role of T cell receptor (TCR)-alpha chain in forming pre-immuneTCR repertoire revealed by clonal TCR reconstitution system. J ExpMed 2002;195:991–1001.
23. Dietrich PY, Le Gal FA, Dutoit V, Pittet MJ, Trautman L, Zippelius A, et al.Prevalent role of TCR alpha-chain in the selection of the preimmunerepertoire specific for a human tumor-associated self-antigen. J Immunol2003;170:5103–9.
24. Trautmann L, Labarriere N, Jotereau F, Karanikas V, Gervois N, ConnerotteT, et al. Dominant TCR V alpha usage by virus and tumor-reactive T cellswith wide affinity ranges for their specific antigens. Eur J Immunol2002;32:3181–90.
25. Cilloni D, Renneville A, Hermitte F, Hills RK, Daly S, Jovanovic JV, et al.Real-time quantitative polymerase chain reaction detection of minimalresidual disease by standardizedWT1 assay to enhance risk stratification inacute myeloid leukemia: a European LeukemiaNet study. J Clin Oncol2009;27:5195–201.
26. GulyasM,Dobra K,Hjerpe A. Expression of genes coding for proteoglycansandWilms' tumour susceptibility gene 1 (WT1) by variously differentiatedbenign human mesothelial cells. Differentiation 1999;65:89–96.
27. Maurer U, Brieger J, Weidmann E, Mitrou PS, Hoelzer D, Bergmann L. TheWilms' tumor gene is expressed in a subset of CD34þ progenitors anddownregulated early in the course of differentiation in vitro. Exp Hematol1997;25:945–50.
28. Mundlos S, Pelletier J, Darveau A, Bachmann M, Winterpacht A, Zabel B.Nuclear localization of the protein encoded by theWilms' tumor geneWT1in embryonic and adult tissues. Development 1993;119:1329–41.
29. Nakatsuka S, Oji Y, Horiuchi T, Kanda T, Kitagawa M, Takeuchi T, et al.Immunohistochemical detection ofWT1protein in a variety of cancer cells.Mod Pathol 2006;19:804–14.
30. Rosenfeld C, Cheever MA, Gaiger A. WT1 in acute leukemia, chronicmyelogenous leukemia andmyelodysplastic syndrome: therapeutic poten-tial of WT1 targeted therapies. Leukemia 2003;17:1301–12.
31. Chapuis AG, RagnarssonGB,NguyenHN,ChaneyCN, Pufnock JS, SchmittTM, et al. TransferredWT1-reactive CD8þ T cells canmediate antileukemicactivity and persist in post-transplant patients. Sci Transl Med 2013;5:174ra27.
32. Keilholz U, Letsch A, Busse A, Asemissen AM, Bauer S, Blau IW, et al. Aclinical and immunologic phase 2 trial of Wilms tumor gene product 1(WT1) peptide vaccination in patients with AML and MDS. Blood2009;113:6541–8.
33. Maslak PG, Dao T, Krug LM, Chanel S, Korontsvit T, Zakhaleva V, et al.Vaccinationwith synthetic analog peptides derived fromWT1 oncoproteininduces T-cell responses in patients with complete remission from acutemyeloid leukemia. Blood 2010;116:171–9.
34. Oka Y, Tsuboi A, Taguchi T, Osaki T, Kyo T, NakajimaH, et al. Induction ofWT1 (Wilms' tumor gene)-specific cytotoxic T lymphocytes by WT1 pep-tide vaccine and the resultant cancer regression. Proc Natl Acad Sci U S A2004;101:13885–90.
35. Ochi T, Fujiwara H, Okamoto S, An J, Nagai K, Shirakata T, et al. Noveladoptive T-cell immunotherapy using aWT1-specific TCR vector encodingsilencers for endogenous TCRs shows marked antileukemia reactivity andsafety. Blood 2011;118:1495–503.
36. Nakatsugawa M, Hirohashi Y, Torigoe T, Inoda S, Kiriyama K, Tamura Y,et al. Comparison of speedy PCR-ssp method and serological typing ofHLA-A24 for Japanese cancer patients. J Immunoassay Immunochem2011;32:93–102.
37. Heemskerk MH, Hoogeboom M, de Paus RA, Kester MG, van der HoornMA, Goulmy E, et al. Redirection of antileukemic reactivity of peripheral Tlymphocytes using gene transfer of minor histocompatibility antigen HA-2-specific T-cell receptor complexes expressing a conserved alpha joiningregion. Blood 2003;102:3530–40.
38. HiranoN, ButlerMO,XiaZ,Ansen S, vonBergwelt-BaildonMS,NeubergD,et al. Engagement of CD83 ligand induces prolonged expansion ofCD8þ Tcells and preferential enrichment for antigen specificity. Blood2006;107:1528–36.
39. Imataki O, Ansen S, Tanaka M, Butler MO, Berezovskaya A, Milstein MI,et al. IL-21 can supplement suboptimal Lck-independent MAPK activationin a STAT-3-dependent manner in human CD8(þ) T cells. J Immunol2012;188:1609–19.
40. Yang S, Cohen CJ, Peng PD, Zhao Y, Cassard L, Yu Z, et al. Development ofoptimal bicistronic lentiviral vectors facilitates high-level TCR gene expres-sion and robust tumor cell recognition. Gene Ther 2008;15:1411–23.
41. Hirano N, Butler MO, Xia Z, Berezovskaya A, Murray AP, Ansen S, et al.Efficient presentation of naturally processed HLA class I peptides byartificial antigen-presenting cells for the generation of effective antitumorresponses. Clin Cancer Res 2006;12:2967–75.
42. Butler MO, Hirano N. Human cell-based artificial antigen-presenting cellsfor cancer immunotherapy. Immunol Rev 2014;257:191–209.
43. Tamanaka T, Oka Y, Fujiki F, Tsuboi A, Katsuhara A, Nakajima H, et al.Recognition of a natural WT1 epitope by a modified WT1 peptide-specificT-cell receptor. Anticancer Res 2012;32:5201–9.
44. Watanabe K, Suzuki S, Kamei M, Toji S, Kawase T, Takahashi T, et al.CD137-guided isolation and expansion of antigen-specific CD8 cells forpotential use in adoptive immunotherapy. Int J Hematol 2008;88:311–20.
45. Arber C, Feng X, Abhyankar H, Romero E, Wu MF, Heslop HE, et al.Survivin-specific T cell receptor targets tumor but not T cells. J Clin Invest2015;125:157–68.
46. Cameron BJ, Gerry AB, Dukes J, Harper JV, Kannan V, Bianchi FC, et al.Identification of a Titin-derived HLA-A1-presented peptide as a cross-reactive target for engineered MAGE A3-directed T cells. Sci Transl Med2013;5:197ra03.
47. Garcia KC. Reconciling views on T cell receptor germline bias for MHC.Trends Immunol 2012;33:429–36.
48. Marrack P, Scott-Browne JP, Dai S, Gapin L, Kappler JW. Evolutionarilyconserved amino acids that control TCR-MHC interaction. Annu RevImmunol 2008;26:171–203.
49. Dai S, Huseby ES, Rubtsova K, Scott-Browne J, Crawford F,MacdonaldWA,et al. Crossreactive T Cells spotlight the germline rules for alphabeta T cell-receptor interactions with MHC molecules. Immunity 2008;28:324–34.
50. Robbins PF, Li YF, El-Gamil M, Zhao Y,Wargo JA, Zheng Z, et al. Single anddual amino acid substitutions in TCR CDRs can enhance antigen-specific Tcell functions. J Immunol 2008;180:6116–31.
www.aacrjournals.org Cancer Immunol Res; 3(9) September 2015 1081
TCR Reactivity and Chain Centricity
on July 7, 2018. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Published OnlineFirst May 5, 2015; DOI: 10.1158/2326-6066.CIR-14-0222

2015;3:1070-1081. Published OnlineFirst May 5, 2015.Cancer Immunol Res Toshiki Ochi, Munehide Nakatsugawa, Kenji Chamoto, et al.
Therapyfor the Purpose of Safe and Effective Antitumor TCR Gene Optimization of T-cell Reactivity by Exploiting TCR Chain Centricity
Updated version
10.1158/2326-6066.CIR-14-0222doi:
Access the most recent version of this article at:
Material
Supplementary
http://cancerimmunolres.aacrjournals.org/content/suppl/2015/05/05/2326-6066.CIR-14-0222.DC1
Access the most recent supplemental material at:
Cited articles
http://cancerimmunolres.aacrjournals.org/content/3/9/1070.full#ref-list-1
This article cites 50 articles, 21 of which you can access for free at:
Citing articles
http://cancerimmunolres.aacrjournals.org/content/3/9/1070.full#related-urls
This article has been cited by 2 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerimmunolres.aacrjournals.org/content/3/9/1070To request permission to re-use all or part of this article, use this link
on July 7, 2018. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Published OnlineFirst May 5, 2015; DOI: 10.1158/2326-6066.CIR-14-0222


















![IMMUNOGLOBULINE E T CELL RECEPTOR T. Strachan e A.P. … · B cell antigen receptor tetramero [ IgH 2 + IgL 2 (Ig oppure Ig )] T cell receptor (TCR) eterodimero TCR /TCR TCR /TCR](https://img.pdfslide.us/doc/110x75/5c017b5c09d3f26f1e8cc6a0/immunoglobuline-e-t-cell-receptor-t-strachan-e-ap-b-cell-antigen-receptor.jpg)










