Embed Size (px)
Citation preview

TIfE JOURNAL OF BIOWICAL Q 1994 by The American
CHEMISTRY Society for Biochemietzy and Molecular Biology, Inc.
Vol. 269, No. 37, Issue of September 16, pp. 23177-23184,1994 Printed itl U S A
GATA-4/5/6, a Subfamily of Three Transcription Factors Transcribed in Developing Heart and Gut*
(Received for publication, June 10, 1994, and in revised form, June 29, 1994)
Anne C. Lavemiere$, Colin MacNeillS, Christine MuellerO, Robert E. Poelmannll, John B. E. Burch$ll, and Todd Evan&** From the $Fox Chase Cancer Center, Philadelphia, Pennsylvania 19111, the $Department of Biological Sciences, University of Pittsburgh, Pittsburgh, Pennsylvania 15260, and the Wepartment of Anatomy and Embryology, University of Leiden, 2300 RC, Leiden, Netherlands
We describe experiments which show that: 1) two pre- viously identified members of the GATA family of tran- scription factors (both of which were originally called GATA-4) correspond to chicken cDNAs for two distinct factors (which we now refer to as cGATA-4 and eGATA-5); 2) another new member of this family corresponds to a third factor designated cGATA-6; 3) each of these mR.NAs displays a differential expression pattern. The cGATA-5 gene is initially transcribed in the cardiac crescent prior to formation of the primordial heart tube. Following formation of the primitive heart, cGATA-5 transcripts are evident in both endocardium and myocardium as well as in other lateral plate de- rivatives. The c(iA31A-5 gene is also transcribed in the primitive embryonic gut and in late stage embryos is sequentially up-regulated in distinct segments of gas- trointestinal epithelia as they undergo terminal differ- entiation. These studies thus provide novel insights into tissue-specific regulation by GATA-5, as well as into possibly overlapping regulatory functions for these three family members.
DNA regulatory elements conforming to the consensus WGATAR were first identified in studies of erythroid-specific gene transcription (1-3). An erythroid transcription factor (GATA-1) that binds specifically to GATA cis-elements was identified and shown to contain a novel DNA-binding domain consisting of two similar zinc fingers of the general form CXNCX,,CNXC (4,5). Subsequently, clones encoding two addi- tional vertebrate factors (GATA-2 and GATA-3) were isolated by cross-hybridization to GATA-1 cDNA (6) and shown to have similar DNA-binding domains. A number of proteins, some- times containing only the single COOH-terminal zinc finger, have since been identified from invertebrate sources (7-13).
The genes encoding GATA factors are transcriptionally regu- lated in a tissue-restricted manner, and the various proteins are believed to regulate distinct subsets of target genes, Al- though some differences in binding specificity have been noted (14-161, there is yet no definitive explanation regarding how
35535 ( to J. B.), CA-01689 (to C. MacNeil), CA-06927 (awarded to the * This work was supported by National Institutes of Health Grants
Fox Chase Cancer Center), 34207 (to T. E.), by an appropriation by the Commonwealth of Pennsylvania, and by NWO Grant 900-516-192 (to A. G.-deG. and R. E. P.). Parts of this work were supported by the Chicago Community Trust and the American Heart Association (to T. E.). The costs of publication of this article were defrayed in part by the payment
tisement” in accordance with 18 U.S.C. Section 1734 solely to indicate of page charges. This article must therefore be hereby marked “uduer-
this fact. // To whom correspondence should be addressed. %I.: 215-728-3696;
Fax: 215-728-3574; E-mail, [email protected]. ** Searle Scholar.
different members of the family discriminate target sites. GATA-1 is expressed in erythroid cells (4, 5), megakaryocytes and bone marrow-derived mast cells (171, eosinophils (181, and testis (19) and has been implicated in the positive regulation of terminal differentiation markers in various myeloid cells. Gene disruption experiments have demonstrated the requirement of GATA-1 for the maturation of erythroid progenitors (20) and indicate that the function of GATA factors is not redundant. GATA-2 is expressed in a number of hematopoietic cells in addition to embryonic brain and liver; GATA-3 is also expressed in hematopoietic cells (primarily T - l ~ p h ~ ~ s ) and embry- onic brain. though these three GATA factors display partially overlapping expression patterns, their relative abundance in a given cell type varies significantly (18, 21).
We recently identified inXenopus a new member of the GATA factor family which was designated xGATA-4 (22). Based on data presented below, we have renamed this factor xGATA-5. The xGATA-5 gene is highly transcribed in heart and through- out the gut, with lower levels detected in lung, gonads, liver, and gall bladder. Whole mount in situ hybridization was used to demonstrate that the xGATA-5 gene is first activated during embryonic cardiogenesis, with transcripts initially detected in endocardium. In the adult gut, xGATA-5 RNA localized to the epithelial cell layer. Coincidentally, a murine GATA factor with a very similar expression profile was identified (23). The mGATA-4 gene is transcribed in heart, intestinal epithelium, primitive (yolk sac) endoderm, and gonads. Although the two cDNAs (mGATA-4 and xGATA-5) are not well conserved out- side of the DNA-binding domain, they were both originally designated GATA-4 based on the fact that they contain related finger sequences and the observation that they display similar expression profiles. Mammalian and avian GATA-1 are also not conserved outside of the DNA-binding domain. In order to further analyze the role of GATA factors during
embryonic development, we have isolated and studied the ex- pression of chick GATA-415 homologues and have discovered, in addition, a novel related GATA factor which we have desig- nated cGATA-6. Each of the genes is transcribed with a similar but distinct tissue profile. We have used in situ hybridization experiments to analyze precisely the embryonic regulation of cGATA-5 and Northern blotting experiments to characterize the overlapping expression patterns of the chick GATA-4f516 genes. Our results indicate that this subfamily of vertebrate transcription factors is potentially involved in regulating car- diogenesis and differentiation of gut epithelium.
~ T E R ~ S AND MET~ODS I s o ~ t ~ o n and CharacteFi~ation of ~~A~~~~~~ cIXVA Clones-
A p p ~ ~ m a ~ l y 400,000 recombinant phage from a h g t l l chicken liver cDNA library (Clontech) were screened using standard hybridization conditions (24) with a random prime labeled (Amersham Corp.) frog GATA-Sa cDNA probe (previously denoted xGATA-.Pa, Ref. 22). ARer
23177

washing the filters under low stringency conditions (2 x 30 rnin in 2 x SCC, 1% SDS at 22 "C followed by 2 x 30 min in 4 x SSC, 1% SDS at 50 "C), one positive clone was identified. The EeoRI cDNA insert was subcloned into pSPORT (Life Technologies, Inc.) and sequenced using Sequenase reagents (U. S. Biochemical Corp.). The partial open reading frame revealed by this analysis was most similar to xGATA-5. This cGATA-5 cDNA was then used as a probe to analyze 400,000 recombi- nant phage from a A g t l O chicken liver cDNA library as described above except that high stringency washing conditions were used for the sec- ond set of filter washes (2 x 30 min with 0.1 x SSC, 1% SDS at 65 "C). One of the two positive clones identified in this screen proved to contain a full-length cGATA-5 cDNA insert, whereas the other cDNA clone was distinct. This novel cDNA was denoted cGATA-6.
Preliminary attempts to obtain a cDNA clone for the uncharacterized 3' end of cGATA-6 proved unsuccessful. We therefore isolated a 1.8-kb' NcoIISacI genomic fragment that we reasoned should contain the de- sired 3' end of the cGATA-6 open reading frame, since it hybridized to a probe from the 3' end of our partial cDNA clone, and because the analogous segment of the cGATA-1 gene is encoded by a single exon (25). Genomic DNA kagments of the correct size were ligated into pGEN.5ZfY (Promega). Following electroporation, cGATA-6 clones were identified by colony hybridization (26). Unexpectedly, the 3' end of the cGATA-6 open reading frame is interrupted by two introns. This was codxrmed by sequencing a 3' rapid amplification of cDNA ends RTPCR product (27) that spans the region of interest. As with the full-len~h cGATA-5 cDNA clone, the complete sequence of the cGATA-6 cDNA was deter- mined on both strands using supercoiled plasmids bearing Bal-31 deletions.
(GATA-GT2 and GATA-GT1 (2811, xGATA-5 (221, and cGATA-516 (this Based on sequence comparisons between mGATA-4 1231, rGATA-416
report), four degenerate 17-mer primers were designed to use RTlPCR 127) to screen for other members of the GATA subfamily that might be expressed in heart, stomach, or intestine: P1 (sense), 5'-GA(CpT)AT- GTT(CpT)GA(CpT)GA(CpT)TI-3' (flanking the core domain shown in Fig. lD); P2 (antisense), 5'-CC(AIG)TGNA(AIG)(CpT)'M'CAT(NG)TA-3' (amino acids 87-92); and P3 (sense), 5'-TA(CT)CA(CPT)AA(AIG)AT- GAA(CpT)GG-3' (amino acids 43-48); P4 (antisense), 5'-GC(A/GTI"M'A/ G)CANACNGG(CpT)TC-3' (amino acids 78-83).
Adult heart, intestine, and stomach cDNAs were synthesized from 1 pg of poly(A)' RNA using a dT,, adaptor primer and avian myeloblas- tosis virus reverse transcriptase (U. S. Biochemical Corp.). The cDNA template was amplified by PCR using 50 pmol each of the P1 and P2 primers in the presence of 4% dimethyl sulfoxide (40 cycles of 1 min 95 "C, 1 min 50 "C, 1.5 min 72 "C, followed by 1 cycle of 4 min at 72 "C). Samples of these PCR products were then used to program second round PCR reactions as described above except that the nested primers (P3 and P4) were used in place of P1 and P2. The PCR fragments were cloned directly into the pCRII plasmid (Invitrogen) and sequenced using SP6 and T7 primers. One of these clones was found to contain a segment of the cGATA-4 open reading kame. This insert, encoding the GATA-4 finger region, was used to generate probes for isolating clones encoding the additional sequences. ern Blot Analysis-Tissues from adult laying hens or embryos
were collected into liquid nitrogen and lysed with a Polytron homoge- nizer (adult tissues) or a mortal and pestle (embryonic tissues) in 15 volumes of lysis buffer (2% SDS, 0.2 N NaCl, 0.2 N Tris (pH 7.5), 1.5 mx MgCl,, and 0.2 mdml proteinase K). Tissue homogenates were incu- bated for 1 h at 45 "C; NaCl concentration was adjusted to 0.5 M and oligo(dT)-cellulose (type 7, Pharmacia Biotech Inc.) was added to a final concentration of 1 rndml. Samples were rocked for 1 h at 22 "C, and the oligo(dT)-cellulose was recovered by low speed (3000 xg) centrifugation. The resin was subjected to serial rounds of resuspension and pelleting: twice with lysis buffer containing 0.5 M NaCl and then three times with lysis buffer containing 0.1 M NaCI. Poly(A)' RNA was eluted from the final oligo(dT)-cellulose pellets with TE buffer (10 lll~ Tris (pH 7.51, 1 nm EDTA) and quantified by UV spectroscopy.
Samples of poly(A)+ RNA (1 pgfiane) were resolved on formaldehyde agarose gels, transferred to Nytran-Plus nylon membranes (Schleicher & Schuell), and hybridized to random prime labeled GATAfactor probes under standard conditions (24). The cGATA-4 probe was a 150-base pair cDNA fragment from the zinc finger region and identifies a unique 7-kb mRNA. The cGATA-5 probe was a cDNA fragment that encodes a seg- ment of the amino-terminal open reading frame (amino acids 44-107). The cGATA-6 probe was B cDNA fragment including the 5'-untranslated region and a segment of the amino-terminal open reading frame (amino
The abbreviations used are: kb, kilobase(s); RT, reverse transcrip- tion; PCR, polymerase chain reaction.
acids 1-1951. Similar blots (0.1 pg4ane) were hybridized to glyceralde- hyde-3-phosphate dehydrogenase (29) or 28 S rDNA (30) probes. The blots were washed under standard conditions (24) and imaged on X- Omat film (DuPont) using two Lightening Plus intensifying screens (DuPont) for various times as indicated in the figure legends. The sizes of the various GATA factor mRNAs were estimated by comparison to RNA standards (Life Technologies, Inc.). The specificity of each GATA factor cDNA probe was verified using genomic Southern blot assays (data not shown).
In Situ Hybridization to Embryo and Tissue Sections-Stage 16 (31) quail embryos were fixed in either methanovacetondacetic acid or in 4% paraformaldehyde, 0.1 M PBS and embedded in paraffin. Sections (0.5 pm) were fixed onto chromium-aluminum-coated slides. Following de- waxing, sections were treated essentially as described (32) and hybrid- ized under coverslips to antisense or control sense %-labeled RNA probes at 50 "C as described (22). An approximately 500-base pair frag- ment extending from an MscI site at nucleotide 895 (just 3' of the conserved finger domain) to the 3' end of the cDNA clone was inserted into pBluescript SKII-. RNA probes were prepared from this subclone, purified, and hydrolyzed prior to hybridization, as described (22). Sec- tions were washed as follows: 2 x 15 min at 55 "C in solution A (2 x SSC, 50% formamide, 20 m? ~-mercaptoethanol), 2 x 15 min at 55 "C in solution B (solution A plus 0.5% Triton X-1001, 2 x 2 min at room temperature in solution C (2 x SSC, 20 lll~ ~-mercaptoeth~ol). After RNase A digestion, samples were further washed 2 x 30 min at 50 "C in solution C, 2 x 30 min at 50 "C in solution A, and 2 x 5 min at room temperature in 2 x SSC. Sections were dehydrated, dipped in emulsion, and exposed for 4-7 days. Frozen sections of adult chicken small intes- tine were cut, prepared, hybridized, washed, and analyzed exactly as described (22). All sections were counterstained in hematoxylin and eosin prior to photography.
Whole Embryo in Situ Hybridization-Fertilized chicken eggs were obtained from Truslow Farms (Chesterton, MD). Stage 4 (18-19 h) to stage 14 (50-53 h) chick embryos (31) were prepared and analyzed essentially as described (33). Embryos were fured overnight a t 4 "C in 4% paraformaldehyde, rinsed in PBS, and then stored in 70% ethanol at -20 "C. Samples were rehydrated by sequential 5-min incubations at 22 "C in 70% ethanol, 50% ethanol, 30% ethanol, PBS, and (3 x) PBS, 0.3% Triton X-100. Samples were then digested at 37 "C with proteinase K (3 pg/m1 in 100 rn Tris (pH 8.0), 50 mM EDTA) for either 4 min (stages 4-7 embryos), 8 min (stages 8-10 embryos), or 12 min (stages 12-16 embryos). Samples were washed three times at 22 "C in PBS, 0.3% Triton X-100 and then prehybridized (2 h) and hybridized (24 h) at 63 "C in 50% formamide, 4 x SSC, 1 x Denhardts, 5% dextran sulfate, 500 pg/ml denatured salmon sperm DNA, 250 pgiml tRNA. The Genius RNA labeling kit (Boehringer Mannheim) was used to prepare digoxi- genin labeled cGATA-5 sense or antisense riboprobes directed against a portion of the cGATA-5 open reading frame (amino acids 44-195). Ri- boprobes were hydrolyzed to an average size of 150 nucleotides by limited alkaline hydrolysis in bicarbonate buffer at 60 "C (34). Follow- ing hybndizatjon in the presence of 0.3 pdml riboprobe, embryos were washed at 63 "C for 24 h in hybridization buffer, followed by sequential 20-min incubations at 22 "C in hybridization buffer (lacking salmon sperm DNA and tRNA) diluted (80, 60,40, and 20%) into Genius buffer 1. Finally, emhryos were washed three times with Genius Buffer 1. Embryos were incubated for 1 h at 22 "C in the same buffer supple- mented with 5% heat-inactivated sheep serum (Sigma), 0.3% Triton X-100, and then incubated for 2.5 h at 22 "C in fresh buffer containing alkaline phosphatase-conjugated anti-digoxigenin antibodies. Unbound
times in PBS, 0.3% Tween 20, twice for 20 min each in PBS, 0.5% 'been antibodies were removed by sequentially washing the embryos three
20, and finally in the Genius Buffer 3. Alkaline phosphatase activity was visualized following the addition of nitro blue tetrazolium and 5-bromo-4-chloro-3-indolyl phosphate. The color reactions were stopped after 1 h by extensive washing in TE.
RESULTS cGATA-4 f5 f 6 Represent a New Subfamily of Vertebrate
GATA Factors-To isolate avian clones related to xGATA-5 (previously designated xGATA-4), the frog cDNA was used as a probe to identify cross-hybridizing clones from a chicken liver cDNA library. One partial cDNA clone for cGATA-5 was iden- tified and subsequently used to screen a second library. This latter screen yielded a ~ 1 1 - l e n ~ h cGATA-5 cDNA clone poten- tially encoding a 391-amino acid protein with 74% identity to the deduced amino acid sequence of xGAl'A-5. Unexpectedly, a

FIG. 1. Chicken C4ATA46I6 consti- tute a subfamily of GATA factors. A, the predicted amino acid coding se- quences of cGATA-4, cGATA-5, and cGATA-6 are compared. Note that our clones appear to be lacking approximately 30 amino acids at the amino terminus of cGATA-4. B, the relationship of these pro- teins to the previously reported &ATA-1 (41, cGATA-2, and eGATA-3 (6) are illus- trated by the dendrogram, C , this dendro- gram illustrates the relationships be- tween cGATA-4/5/6 and other members of the GATA factor subfamily that are ex- pressed in heart and limited sets of endodermally derived tissues. The refer- ences for these proteins are as follows: €3 mGATA-4 (23), rGATA-4 (denoted rGATA- GT2 in (28)1, xGATA-5 (denoted xGATA-4 in (2211, and rGATA-6 (denoted rGATA- GT1 in Ref. 28). Preliminary sequence alignments between these factors indi- cated that the published mGATA-4 se- quence contains several frameshifting er- rors and that the published rGATA--6 sequence contains a frameshift error up- stream of what was suggested to be the s t a r t codon. The comparisons shown in C are based on our best estimates for the correct sequences for these two GATA fac- tors. D, residues in the respective DNA binding domains that distinguish the var- ious GATA-4./5/6 family members are highlighted by the comparison.
D
j- cGATA5
"I i
cGATA4
cGATA2
cGATA3
cGATAl
COATA4 mGATA4 rGATA4 CGATAS xGATA5 CGATA6 rOATAb
54 54 5 4 53 54 54 5 4
CGkTA4 ffiATA4 xGhTAb CGATRS
108
SATAS 107
CGATA6 ICU
rcATA6 108 108
1aa loa
partial cDNA clone for a distinct chicken GATA factor was also identified and designated cGATA-6. The 3' end of cGATA-6 was obtained using a combinat~o~ of genomic band cloning and a PCRlrapid amplification of cDNA ends protocol (see "Materials and Methods"). The predicted amino acid sequences of chick GATA-5 and GATA-6 are shown in Fig. 24.
Sequence comparison of these proteins and the three previ- ously reported chicken GATA factors indicate that cGATA-5/6 may define a GATA subfamily distinct from the GATA-1/2/3 subfamily (Fig. 1B). As illustrated in the dendrogram of Fig. lC, it seems unlikely that the clones designated mGATA-4 and xGATA-5 are homologues, but rather are distinct members of this subfamily. The cGATA-5 gene i s much more similar to xGATA-5 than it is to mGATA-4 (74% identity versus 54% iden- tity, respectively). However, mammalian cDNA clones have not yet been isolated which are more similar to XlcGATA-5 than mGATA-4. Therefore, we sought to identify a chick cDNA which would encode a homologue of mGATA-4. We used RT/PCR and degenerate oligomer primers consistent with sequences con- served in all GATA factors (within the DNA-binding domain) to
isolate such a clone. We caa therefore state unequivocally that vertebrates contain three distinct genes designated GATA-4, GATA-5, and GATA-6. The finger regions of known proteins in this family are aligned in Fig. zE1. It is clear from this align- ment that each GATA factor (4, 5, or 6) is characterized by specific sequences located within the conserved finger region. These sequences represent diagnostic amino acids that are con- served across species. While this manuscript was in prepara- tion, cDNA clones isolated from a rat stomach eDNA library were described (28); these clones clearly represent homologues of mGATA-4 and cGATA-6 and are included in the comparison.
GATA-4 I S I S Disptay ~ ~ p o r ~ l ~ and I pa ti ally O v e r l a p ~ ~ n ~ Expression ProfiEes-In order to examine transcription pat- terns for cGATA-4, -5, and -6, Northern blotting experiments were performed using poIyfAf+ RNA isolated from a variety of adult chicken tissues. As shown in Fig. 2, the transcription profiles of GATA-4 and -5 are highly conserved among verte- brates. The cGATA-4 mRNA is most abundant in heart, with lower levels detected in ovary and small intestine. A similar expression pattern has been reported for mGATA-4 (23). The

23180
GATA 4
GATA 5
GATA 6
28 S
The GATA-415 16 Dunscription Factor Subfamily
+
FIG. 2. Chicken GATA-4/5/6 RNAs are expressed in partially overlapping sets of adult tissues. Northern blots of poly(A)+ RNA from various adult tissues ( top) were hybridized to cDNA probes as indicated on the left. Blots were exposed to film for either 18 h (cGATA-5 and cGATA-6) or 5 days (cGATA-4). Each probe was confirmed to be unique and to give similar intensity signals on genomic Southern blots (data not shown). The Northern blot shown in the bottom panel was probed with a 28 S rDNA probe and was exposed for 12 h. A parallel Northern blot that was hybridized to a glyceraldehyde-3-phosphate de- hydrogenate probe gave similar signals (data not shown). Note that the chicken egg gland is synonymous to the chicken uterus.
approximately 7-kb cGATA-4 mRNA is generally less abundant than either the 2.8-kb cGATA-5 mRNA or the 3.1-kb cGATA-6 mRNA. The cGATA-5 message is most abundant in adult stom- ach and small intestine. Quantitative analysis demonstrated that 3-fold lower levels are detected in heart; 10-fold lower levels are found in RNA isolated from lung or spleen. Very low levels (30-fold less than in stomach) are found in samples from liver and ovary. These results are quite consistent with previ- ously reported studies on the xGATA-5 gene (22). The cGATA-6 gene is also highly expressed in the stomach and, to a lesser extent, intestine. Somewhat lower levels are detected in RNA from heart tissue. In contrast to cGATA-5, the cGATA-6 gene is transcribed a t higher levels in lung, liver, and ovary than in heart. Unlike cGATA-5, cGATA-6 RNAis not detected in spleen. Therefore, transcription of these related factors is restricted to distinct but overlapping sets of tissues; within a given tissue, the relative ratio of the three RNAs may vary considerably.
Expression Levels of GATA-4 1516 Correlate with the Timing of Terminal Differentiation in the Gastrointestinal System- RNA samples were isolated from chick heart, stomach, and intestine (large or small) at various times during late embry- onic development and in the first week post-hatching. These samples were analyzed by Northern blotting experiments for levels of cGATA-41516 RNA to determine how transcription of the genes varies late in development. It is clear from the North- ern blot shown in Fig. 3 that all three genes are expressed as early as embryonic day 10 in the heart. At this time, cGATA-4 RNA is barely detectable in stomach and is absent from small intestine. cGATA-5 RNA is abundant in stomach, and the gene is transcribed to a lesser extent in small intestine. cGATA-6 RNA is also detectable in both stomach and small intestine, although at a lower level relative to cGATA-5. As development proceeds, it is clear that the cGATA-5 and
cGATA-6 genes are up-regulated in concert with gut differen- tiation. By embryonic day 15, transcription of these two genes in the stomach exceeds that in the heart. The effect is amplified for cGATA-6, which is down-regulated in the heart tissue dur-
N
GATA 4 1 GATA 5 I GATA 6 I 2R S I Fa. 3. Chicken GATA-4516 display distinct development ex-
pression profiles in late stage embryos and young hatchlings. Northern blots of poly(A)+ RNA from various tissues ( top) were hybrid- ized to chicken GATA probes or to the control 28 S rDNA probe as indicated (bottom). Blots were exposed as in Fig. 2. ND indicates a time point that was not determined (large intestine is not distinguished prior to day 15).
ing late embryonic development. The levels of cGATA-516 mRNA increase around the time of hatching in the small in- testine and first become detectable in the large intestine. Soon after hatching, mRNA levels for both cGATA-516 in the small intestine become equivalent to those in stomach, and the ex- pression of these genes in large intestine equals or exceeds that seen in heart. Eventually (6 days post-hatching and data not shown) RNA transcripts decrease in large intestine to unde- tectable levels. The levels of cGATA-5 mRNA then remain high in heart, stomach, and small intestine. In adults, the levels of cGATA-6 are highest in stomach and small intestine.
Although transcribed at much lower levels than either cGATA-5 or cGATA-6, the cGATA-4 RNA also transiently in- creases in small intestine during late embryonic development. The abundance of message peaks immediately after hatching and within a week declines to nearly undetectable levels. In contrast to the other genes, very little cGATA-4 RNA is ever detected in stomach, and transcripts in large intestine are con- sistently undetectable.
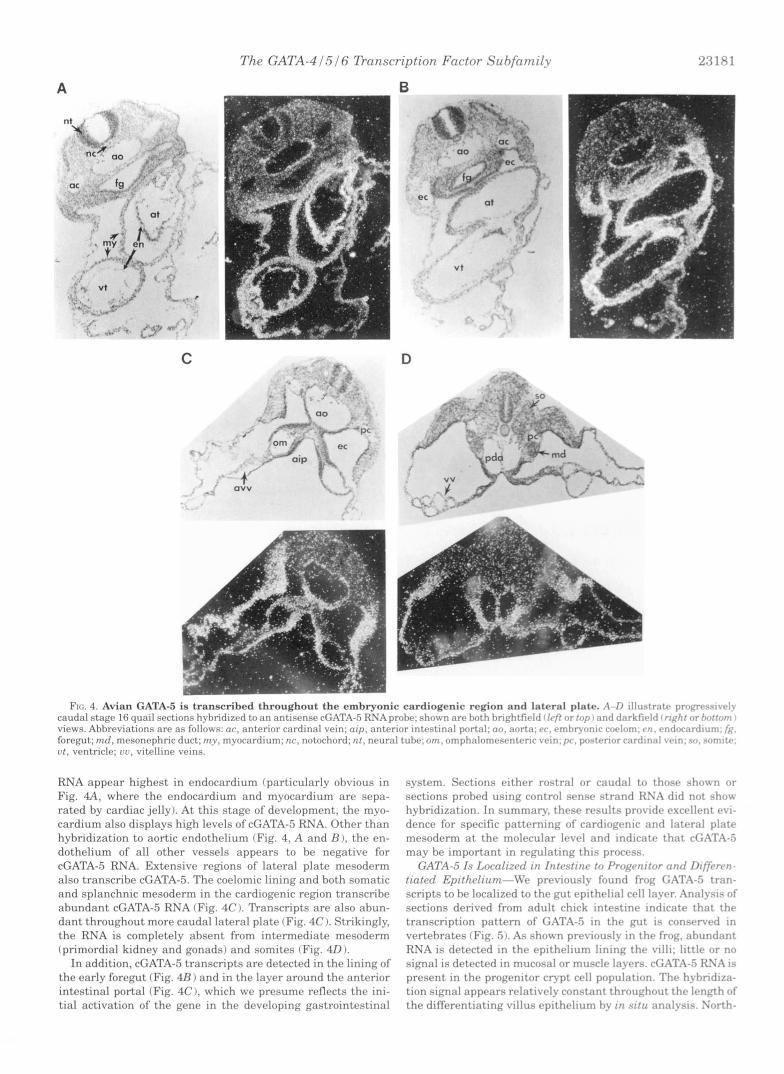
cGATA-5 Dunscripts Are Detected throughout the Cardio- genic Region-Our previous results indicated that early expres- sion of xGATA-5 might be confined to endocardium (22). The isolation of a highly conserved chick homologue has permitted us to analyze cGATA-5 expression in the relatively more acces- sible avian embryonic cardiac system. To begin to further char- acterize the localization of cGATA-5 transcripts, we analyzed sections of avian embryos for cGATA-5 RNA by in situ hybrid- ization.
Stage 16 (31) quail embryos were sectioned and hybridized to 35S-labeled antisense (or control sense strand) cGATA-5 RNA probes. The results (Fig. 4) show extensive GATA-5 transcrip- tion throughout the cardiogenic region. The levels of cGATA-5
A
C
Th.e GATA-4IIiI6 Tkanscription Factor Srchfami1.v 23181
B
RNA appear highest in endocardium (particularly obvious in Fig. 4 A , where the endocardium and myocardium are sepa- rated by cardiac jelly). At this stage of development, the myo- cardium also displays high levels of cGATA-5 RNA. Other than hybridization to aortic endothelium (Fig. 4, A and B ), the en- dothelium of all other vessels appears to be negative for cGATA-5 RNA. Extensive regions of lateral plate mesoderm also transcribe cGATA-5. The coelomic lining and both somatic and splanchnic mesoderm in the cardiogenic reg$on transcribe abundant cGATA-5 RNA (Fig. 4C I. Transcripts are also abun- dant throughout more caudal lateral plate (Fig. 4C 1. Strikingly, the RNA is completely absent from intermediate mesoderm (primordial kidney and gonads) and somites (Fig. 40).
In addition, cGATA-5 transcripts are detected in the lining of the early foregut (Fig. 4R) and in the layer around the anterior intestinal portal (Fig. 4C), which we presume reflects the ini- tial activation of the gene in the developing gastrointestinal
system. Sections either rostral or caudal to thtrsr shwvn or sections probed using control sense strand RS:I did not .show hyhridization. In summary, thew rcssults providv rxcrbllr.nt rvi- dence for specific pnttrrninp of cardiorrnir and Iatrral plate mesoderm at thc. molc,cular Irvel and indicate that cC;.AT;\-5 may he important in r rwlat ing this process.
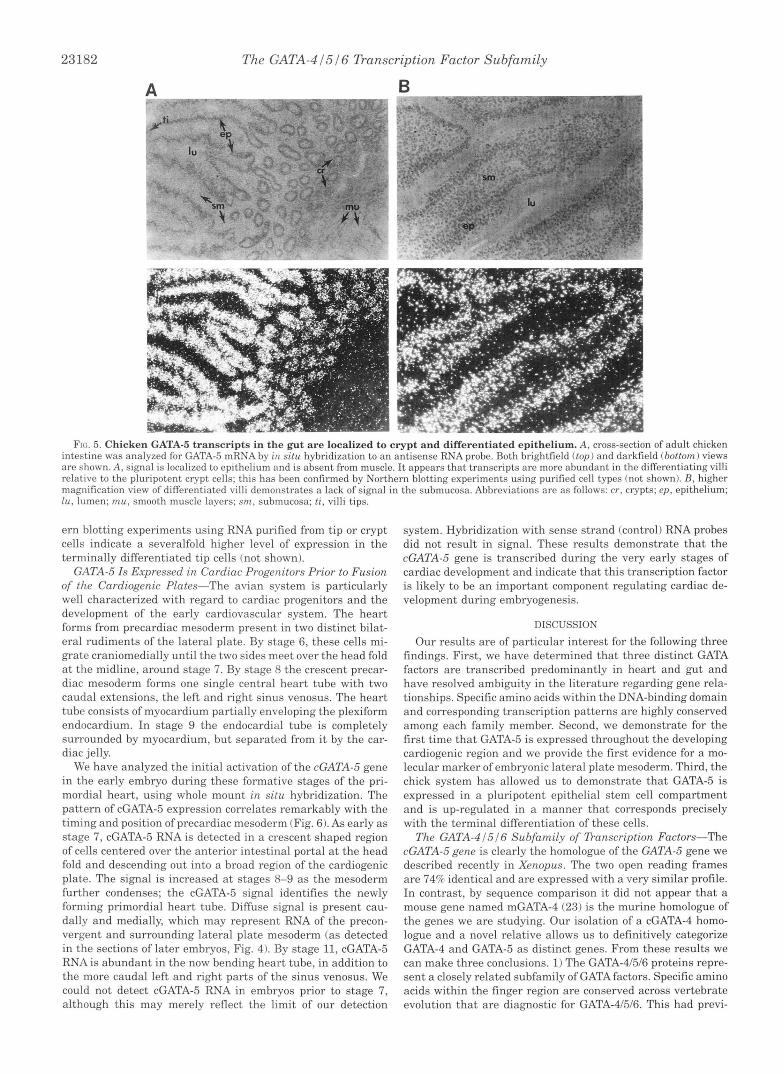
C;ATA-.5 I s Locnlizrrl r n I ~ l t c . ~ t i n ~ t o l'roqvnitnr rrnd D i f p w u - tintrd Epifhr/ium-lVr previously found froq G:ITJ\-S trnn- scripts to he localized to the gut epithclinl crll Inyrr. A n : j I y ~ i ~ of sections derived from adult chick intrstinr intlirntr that t h e transcription pattern of GATA-5 in t h r p r t is con.srn.rtl in vertebrates (Fig. 5). As shown previously in the froq. ahundnnt RNA is detected in the epithelium lining thf. villi: l i t t l r or no signal is dc>trcted in mucosal or musclr Iayrrs. di.AT.4-5 KSX is present in the progenitor c F p t cell population. Thr hyhridizn- tion sikmal appears rrlativr.lycnnstant throuchorlt thp 1f.nr-h o f the differentiating villus epithelium hy in r i t r l analysis. Xnrth-
23182 The GATA-4 I516 Dunscription Factor Subfamily
A
FIG. 5. Chicken GATA-5 transcripts in t h e gut are local ized to crypt and di f fe ren t ia ted ep i thc l ium. A, cross-section of adult chicken intestine was analyzed for GATA-5 m l K A by i u S I / U hxbridizatlon to an antisense RNA prolw. 130th Orightlic~ltl ( t o p ) and darkfield (bottom) views are shown. A, signal is localized to epithelium and is absent from muscle. It appears that transcripts arc more abundant in the differentiating villi relative to the pluripotent crypt cells; this has been confirmed by Northern blotting experiments using purified cell types (not shown). B, higher magnification view of differentiated villi demonstrates a lack of signal in the submucosa. Abbreviations are as follows: cr, crypts; ep , epithelium; lu, lumen; m u , smooth muscle layers; sm, submucosa; ti, villi tips.
ern blotting experiments using RNA purified from tip or crypt system. Hybridization with sense strand (control) RNA probes cells indicate a severalfold higher level of expression in the did not result in signal. These results demonstrate that the terminally differentiated tip cells (not shown). cGATA-5 gene is transcribed during the very early stages of
GATA-5 Is Expressed ir7 Cardiac Progenitors Prior to Fusion cardiac development and indicate that this transcription factor of the Cardiogenic Plates-The avian system is particularly is likely to be an important component regulating cardiac de- well characterized with regard to cardiac progenitors and the velopment during embryogenesis. development of the early cardiovascular system. The heart forms from precardiac mesoderm present in two distinct bilat- eral rudiments of the lateral plate. By stage 6, these cells mi- grate craniomedially until the two sides meet over the head fold at the midline, around stage 7. By stage 8 the crescent precar- diac mesoderm forms one single central heart tube with two caudal extensions, the left and right sinus venosus. The heart tube consists of myocardium partially enveloping the plexiform endocardium. In stage 9 the endocardial tube is completely
DISCUSSION
Our results are of particular interest for the following three findings. First, we have determined that three distinct GATA factors are transcribed predominantly in heart and gut and have resolved ambiguity in the literature regarding gene rela- tionships. Specific amino acids within the DNA-binding domain and corresponding transcription patterns are highly conserved among each family member. Second, we demonstrate for the
surrounded by myocardium, but separated from it by the car- first time that GATA-5 is expressed throughout the developing diac jelly. cardiogenic region and we provide the first evidence for a mo-
We have analyzed the initial activation of the cGATA-5 gene lecular marker of embryonic lateral plate mesoderm. Third, the in the early embryo during these formative stages of the pri- chick system has allowed us to demonstrate that GATA-5 is mordial heart, using whole mount in situ hybridization. The expressed in a pluripotent epithelial stem cell compartment pattern of cGATA-5 expression correlates remarkably with the and is up-regulated in a manner that corresponds precisely timing and position of precardiac mesoderm (Fig. 6). As early as stage 7, cGATA-5 RNA is detected in a crescent shaped region of cells centered over the anterior intestinal portal at the head fold and descending out into a broad region of the cardiogenic plate. The signal is increased at stages 8-9 as the mesoderm further condenses; the cGATA-5 signal identifies the newly forming primordial heart tube. Diffuse signal is present cau- dally and medially, which may represent RNA of the precon- vergent and surrounding lateral plate mesoderm (as detected in the sections of later embryos, Fig. 4). By stage 11, cGATA-5 RNA is abundant in the now bending heart tube, in addition to
with the terminal differentiation of these cells. The GATA-4 151 6 Subfamily of Danscription Factors-The
cGATA-5 gene is clearly the homologue of the GATA-5 gene we described recently in Xen,opus. The two open reading frames are 74% identical and are expressed with a very similar profile. In contrast, by sequence comparison it did not appear that a mouse gene named mGATA-4 (23) is the murine homologue of the genes we are studying. Our isolation of a cGATA-4 homo- logue and a novel relative allows us to definitively categorize GATA-4 and GATA-5 as distinct genes. From these results we can make three conclusions. 1) The GATA-4/5/6 proteins repre-
the more caudal left and right parts of the sinus venosus. We sent a closely related subfamily of GATA factors. Specific amino could not detect cGATA-5 RNA in embryos prior to stage 7, acids within the finger region are conserved across vertebrate although this may merely reflect the limit of our detection evolution that are diagnostic for GATA-4/5/6. This had previ-

The GATA-41516 Danscription Factor Subfamily 23183
FIG. 6. Chicken GATA-5 is transcribed in the prefusion cardi- ogenic precursor cells. Whole mount in situ hybridization was used to detect cGATA-5 message. Embryos from the indicated stages (7-11) were processed and hybridized to digoxigenin-labeled antisense RNA probes. The hybridized probe was detected using an anti-digoxigenin antibody conjugated to alkaline phosphatase. The stain indicates the presence of alkaline phosphatase activity. Note that cGATA-5 RNA is readily detected in the precardiac crescent (arrows in stage 7 and 8+). All are dorsal views; cranial is toward the top. Abbreviations are as follows: aip, anterior intestinal portal; hf, head fold; hn, Hensen’s node; ht, heart (bending); nf, neural fold; tu, tube of sinus venosus; s, somite.
ously been shown to be true regarding GATA-11213.2) It is clear that cGATA-5 and cGATA-6 are the most abundant GATA fac- tors expressed in differentiated heart and gut tissue, respec- tively. It is possible that cGATA-4 RNA is more predominant in early cardiac cells, although this has not been tested prior to day 10. 3) It appears unlikely that additional GATA factors (unless significantly diverged) are expressed in heart or gut. We have sequenced over 120 RTPCR products generated from stomach, intestine, or heart RNA using a variety of degenerate PCR primers and found these correspond exclusively to GATA- 41516.’ The cGATA-6 gene is clearly the homologue of the rat gene called GATA-GT1 (open reading frames are 78% identi- cal). In addition, we have isolated and characterized a homol- ogous human clone.3 Homologues of GATA-4 and GATAB have also been characterized from zebrafish,” and of GATA-41516 from Xenopus.’
Although cGATA-41516 are all expressed in heart and gut, they are not coordinately regulated. However, by day 10, RNA from each gene is detected in heart and gut. In the developing gastrointestinal system, cGATA-516 transcripts appear more abundant in proximal stomach relative to distal intestine. As the embryo develops they are activated distally, corresponding to the timing of epithelial differentiation; after hatching, the
*A. C. Laverriere and J. B. E. Burch, unpublished data. T. Evans, unpublished data. L. Zon, personal communication. ’ Y. Jiang and T. Evans, unpublished results.
gene is active throughout. Later and in adults, both cGATA-516 appear to be dramatically down-regulated in the colon, consist- ent with results found in Xenopus (22). However, this may also reflect the relatively small number of enterocytes in distal gut relative to small intestine. We never detect cGATA-4 transcrip- tion in large intestine, and RNA only accumulates transiently to a significant level in small intestine just after hatching. In adult tissues, the expression patterns for the three genes over- lap considerably. All are expressed in stomach, although cGATA-4 RNA levels are relatively low. In lung and liver, cGATA-6 transcription is high, cGATA-5 is low, and cGATA-4 is undetectable. In ovary, cGATA-6 transcription is also high, cGATA-4 is low, and cGATA-5 is undetectable. In spleen, cGATA-5 RNA is most abundant, cGATA-6 is very low, and cGATA-4 is undetectable.
GATA-5 Is an Early Marker for Committed Lateral Plate Mesoderm Including Cardiac Progenitors-We can first detect GATA-5 RNA in the cardiogenic region a t about the time (stage 7) progenitors are becoming definitively committed to form heart tissue. By this stage, the precardiac mesoderm has formed a crescent of cardiogenic material in the midregion of the cephalic portion of splanchnic mesoderm. This crescent is readily apparent in the whole mount in situ hybridization ex- periments as the domain of early cGATA-5 gene transcription. The bilateral precardiac rudiments migrate as a cohesive epi- thelium, perhaps on a fibronectin matrix, to the midline axis where they fuse and form a single heart tube at the embryonic midline. The myocardial progenitors form around the rudiment of the endocardial tube; it has been suggested that the endo- cardial progenitor cells represent a distinct cell population of the primordial cardiac forming region (35). Our data regarding cGATA-5 transcription are consistent either with activation of the gene in a common lineage or coordinate activation in two independent lineages.
The cGATA-5 gene is transcribed throughout the cardiogenic region, including myocardium, endocardium, and lateral plate derivatives. RNA appears specific to lateral plate mesoderm, as somitic and nephrogenic mesoderm do not contain cGATA-5 message. Levels of cGATA-5 RNA appear highest in endocar- dial cells. Previous results in the frog system detected primar- ily endocardial transcripts in early embryos; such analyses are often limited by the RNA detection sensitivity. In adults, xGATA-5 RNA is clearly not restricted to endocardium”
The GATA-5 gene appears to be the earliest known molecular marker for cells of the precardiogenic plate committed to form various derivatives of lateral plate mesoderm. Similar early expression (stage 7) of sarcomeric myosin has been detected histochemically (361, although this is presumably confined to myocardial precursors and was often detected only on one side of the anterior intestinal portal. Numerous structural proteins are specific to differentiating myocardium and are not acti- vated until after fusion of the anlage. The mouse homologue of the Drosophila homeobox gene tinman is also transcribed prior to myogenic differentiation and is an excellent candidate for a regulator of myogenesis (37). The early embryonic expression pattern of this gene differs from GATA-5: RNA is detected ex- clusively in mesodermal cells of progenitor myocardium, in ad- dition to prospective pharyngeal endoderm. I t will be of consid- erable interest to determine if GATA-5 is required for proper patterning of lateral plate mesoderm and whether i t is regu- lated by inductive processes presumed to function during heart development.
GATA Factor Dunscription in Gut Correlates with Terminal Differentiation of Epithelium-Differentiated epithelial cell types are derived from a small population of multipotential stem cells present in the crypt (38, 39). Cells differentiate dur-

23184
ing migration to the villus tip, prior to extrusion into the lu- men. Although cell position along the polar crypt/tip axis is clearly an important determinant, the mechanisms by which the decision to differentiate is made (and along which pathway) are completely unknown. Position along the proximalldistal digestive tract also plays a role in determining cell type; for example, the relative percentage of goblet cells is greatly in- creased in distal regions. The GATA-4 1516 genes are candidate regulatory molecules that might interpret these polarities at the level of gene expression. scriptio ion of the GATA-5 gene is highly specific to epithelium and is not detected in cells of the underlying submucosa or musculature. It appears that cGATA-415/6 transcription occurs within the large intestine only during embryogenesis and is not maintained in the adult.
Potential Targets of GATA-4 / 5 / 6 Exist in Heart and Gut-By analogy to what is known regarding the related GATA-1 tran- scription factor, we hypothesize that GATA-41516 directly acti- vate the expression of terminal differentiation markers in the heart and gut. Although progress has been made recently in identifying cardiac regulatory proteins, few have been charac- terized that are specific to cardiac relative to skeletal muscle. Using chimeric promoter constructs, a conserved motif within the promoter of the cardiac troponin T gene was found to be important for expression in skeletal muscle, whereas the pro- moter only €unctions in cardiac cells when additional upstream sequences are included (40). It is of interest that this upstream element contains a potential binding site for GATA factors. Also, an element critical for cardiac expression has been local- ized to the proximal promoter of the cardiac troponin C gene (41). It is distinct from an intronic element known to regulate the expression of this gene in embryonic skeletal muscle. Car- diac-restricted DNA binding activities were identified which interact with this region and mutations in two of these sites (CEF-1 or CEF-2) abolished transcription in cardiac myocytes. Interestingly, the sequence of the CEF sites are also compatible with binding to GATA factors.
Distinct targets for GATA-41516 are likely to reside in epithe- lial cells of the gut. Potential GATA binding sites are conserved across species in the promoter regions of two genes expressed in the adult small intestine: intestinal fatty acid-binding protein and sucrase-isomaltase. Likewise, consensus GATA binding sites are conserved in rat and human genes encoding a: and f3 subunits of the H+/K+-ATPase, a major enzyme involved in transport of protons across the apical membrane of parietal cells. As is the case for potential cardiac targets, the functional significance of these sites has not been demonstrated but they provide exceIlent candidates for GATA-41516 action. Besides markers of terminal differentiation, GATA-5 i s likely to regu- late unknown genes important in the earlier commitment step of specialized cell types, as RNA is present in prefusion pro- genitor cells of the heart and predifferentiated crypt cells of the gut epithelium. The G A T A - 4 / 5 / ~ genes therefore provide use- ful molecular tools for understanding and potentially control- ling the regulation of cardiac- and gut-specific gene expression.
son (Fox Chase) and Eric Weinberg (University of Pennsylvania) for Acknowledgments-We thank Elizabeth Pownall and Charles Emer-
help with the whole embryo in situ analysis, Monica Mentink (Leiden, Netherlands) for histology, Robert Muhlhauser (Oligonucleotide Syn-
thesis Facility, Fox Chase) for oligonucleotides, Adriana Gitknberger- deGroot (Leiden, Netherlands) for critically reading the manuscript, Leonard Zon (Harvard) €or communicating unpublished results, Doug Engel (Northwestern University) for helpful discussions, and Michelle Slook (Secretarial Services, Fox Chase) for help in preparing the manu- script.
REFERENCES 1. Orkin, S. H. (1990) Cell 63, 665-672 2. Orkin. S. H. (1992) Blood 80,575-581 3. Simon, M. C. (19931 Proc. SOC. Exp. Biol. Med. 202, 115-121 4. Evans, T., and Felsenfeld, G. (1989) Cell 68, 877-885 5. Tsai, S.-F., Martin, D. I. K, Zon, L. I., D'Andrea, A. D., Wong, G. G., and Orkin,
6. Yamamoto, M., KO, L. J., Leonard, M. W., Beug, H., Orkin, S. H., and Engef, J.
7. Cunningham, T. S., and Cooper, T. G. (1991) Mol. Cell. Biol. 11,6205 8. Fu, Y.-H., and Marzluf, 0. A. (1990) Proc. Nutl. Acud. Sei. U. S. A. 87,5331-
9. Kudla, B., Caddick, M. X., Langdon, T., Martinez-Rossi, N. M., Bennett, C . F,, 5335
Sibley, S., Davies, R. W., and Arst, H. N., Jr. (199O)EMBO J. 9, 1355-1364 10. Spieth, J. , Shim, Y. H., Lea, K., Conrad, R., and Blumeuthal, T. (1991) Mol.
Cell. Biol. 11, 4651-4669 11. Abel, T., Michelson, A. M., and Maniatis, T. (19931 Development (Cumb.) 119,
623-633 12. Ramain, P., Heitzler, P., Haenlin, M., and Simuson, P. (1993) Deuelojoment
S. H. (1989) Nature 389,446-451
D. (1990) Genes & Dev. 4,1650-1662
(Cumb.) 119,1277-1291 13. Winick, J., Ab&, T., Leonard, M. W., Michelson, A. M., C h a ~ o n - L o r i ~ ~ , I.,
Holmgren, R. A,, Maniatis, T., and Engel, J. D. (1993) D e u e ~ p ~ n t (Cumk.) 119.1055-1065
14. KO, L.'J,, and Engel, J. D. (1993) ,5402. Cell. Biol. 13, 4011-4022
16. Whyatt, D. J., deBoer, E., and Grosvdd, F. (1993) EMBO J. 12,4993-5005 15. Merika, M., and Orkin, S. H. (1993) Mol. Cell. Bioi. 13,3999-4010
17. Martin, D. 1. K., Zon, L. I., Mutter, G., and Orkin, S. H. (1990) Nature 344,
18. Zon, L. I., Yamwchi, Y., Yee, K., Albee, E. A., Kimura, A., Bennett, J. C.,
19. Ito, E., Tsutomu, T., Ishihara, H., Ohtani, H., Gu, L., Yokoyama, M., Engef, J.
20. Pevny, L., Simon, M. C., Robertson, E., Klein, W. H., %ai, S.-F., DAgati, V.,
21. Leonard, M. W., Lim, &-C., and Engel, J. D. (1993) Development (Cumb.) 119,
22. Kelley, C., Blumberg, H., Zon, L. I., and Evans, T (1993) Development (Cumk.)
23. Arceci, R. tJ., Kina, A. A. J., Simon, M. C., Orkin, S. H., and Wilson, D. B. (1993)
444-447
Orkin, S. H., and Ackeman, S. J. (1993) Blood 81,3234-3241
D., and Yamamoto, M. (1993) Nature 362,466-468
Orkin, S. H., and Costantini, F, (1991) Nature 349,257-260
519-531
118,817-827
24. Maniatis, T., Fritsch, E. F., and Sambrook, J. (1982) Molecular Cloning: A Mol. Cell. Bioc 13,2235-2246
Labomtory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, hT
25. Hannon, R., Evans, T., Felsenfeld, G., and Could, H. (1991) Proc. Nutl. Acud. Sci. U. S. A. 88,3004-3008
26. Gmnstein, M., and Hogness, D. S. 11975) Proc. Natl. Acad. Sci. U, S. A. 72, 3961-3965
27. Frohman, M. A., Dush, M. K., and Martin, G. R. (1988) Proc. Nutl. Amd. Sci. U. S. A. 89,89984002
28. Tamura, S., Wang, X.-H., Maeda, M., and Futai, M. (1993) Proc. N&l. Acad. Sci. U. S. A. 90,10876-10880
29. Dugaiczyk, A,, Haron, J. A,, Stone, E. M., Dennison, 0. E., ~ t h b l ~ , K N., and Schwartz, R. J. 11983) Biochemistry 22,1605-1613
30. Grant, C., and Deeley, R. G. (1993) Mol. Cell. Biol. 13,4186-4196 31. Hamburger, V., and Hamilton, H. L. (1951) J. Morphol. 88,49-92 32. Awgulewitsch, A,, Utset, M. F., Hart, C . P., McGinnis, W., and Ruddle, F. H.
33. Pownall, M. E., and Emerson, C. P., Jr. (1992) Dev. Biol. 151, 67-79 34. Cox, K. H., DeLeon, D. V., Angerer, L. M., and Angerer, R. C . (19841 Deu. Biol.
35. DeRuiter, M. C., Poelmann, R. E., VanderPlas-deY~es, I., Mentink, M. M. T., 101,485-502
36. Han, X, Dennis, J. E., Cohen-Gould, L., Bader, D. M., and Fisehman, D. A. and Gi t tenber~r-deGr~t , A. C . (1992) Anat. Embryol. 185,461-473
37. Lints, T. J., Parsons, L. M., Hartley, L., Lyons, I., and Harvey, R. P. (19931 (1992) Dev. Dyn. 199,257-265
38. Cheng, H., and LeBlond, C. P. (1974) Am. J. Anat. 141,537-562 Deuelopment (Camb.) 119,419-431
39. Schmidt, G. H., Wilkinson, M. M., and Ponder, B. A. J. (1985) Cell 40,425-429 40. Mar, 3. H.,Antin, P. B., Cooper, T. A,, and Ordahl, C . P. (1988) J. Cell Bid. 107,
41. Psnnaeek, M. S., Vora, A. J., Shen, T., Barr, E., Jung, E, and Leiden, J. M.
(1986) Nature 320,328435
573-585
(1992) Mol. CelZ. Biol. 12, 1967-1976




![Bis[tris(1,10-phenanthroline)nickel(II)] tris ... · Bis[tris(1,10-phenanthroline)nickel(II)] tris[dicyanidoargentate(I)] nitrate 4.2-hydrate Muhammad Monim-ul-Mehboob,a Muhammad](https://img.pdfslide.us/doc/110x75/5f74462041fcef38863090d7/bistris110-phenanthrolinenickelii-tris-bistris110-phenanthrolinenickelii.jpg)
























