Embed Size (px)
Citation preview

Investigative Ophthalmology & Visual Science, Vol. 30, No. 12, December 1989Copyright © Association for Research in Vision and Ophthalmology
Ocular Torsion Measured by TV- and Scanning LaserOphfhalmoscopy During Horizontal Pursuit
in Humans and MonkeysDiermar Off and Rolf Eckmiller
Ocular torsion during horizontal foveal pursuit and fixation was measured in five human subjects andtwo trained monkeys (Macaca fascicularis) by direct analysis of the ocular fundus rotation. In themonkeys the fundus images of either eye were generated with a TV-ophthalmoscope while the contra-lateral eye pursued an 8' visual target moving sinusoidally (0.3-0.9 Hz, ±5°) in the horizontal plane. Inthe humans a scanning laser ophthalmoscope (SLO) generated the fundus image of the ipsilateral eye,which pursued the visual target (same parameters as in the monkeys) mixed electronically into thelaser scan raster. The image sequences were stored on videotape and subjected to a frame-by-framerotation analysis. In both the humans and the monkeys, torsion (fundus rotation about the visual axis)sinusoidally modulated (up to 8° peak-to-peak) during foveal pursuit, approximately in phase withhorizontal eye position. Intorsion (nasal movement of the upper eye pole) or extorsion was foundduring pursuit in the temporal or nasal direction. Torsion showed considerable intra-individual fluctua-tion and interindividual variability with regard to phase and modulation depth relative to the pursuitmovements, and was interspersed with irregularly occurring rapid deflections. Torsion of the conju-gate, nonpursuing eye was similar to torsion of the pursuing eye. In contrast, torsion during fixationwas only loosely correlated with horizontal eye position. Slow torsional drifts and large, rapid deflec-tions (up to 6°) occurred in either direction at a given fixation point in the horizontal plane. Weconclude that ocular torsion during horizontal pursuit in primates is actively generated by a separate,neural oculomotor subsystem. Ocular torsion may assist figure-background discrimination duringpursuit by inducing an uneven distribution of extrafoveal slip velocities. Invest Ophthalmol Vis Sci30:2512-2520,1989
Ocular torsion (rotation of the eyes about the visualaxis) is well documented as an integral component ofeye movement in primates.1 For example, ocular tor-sion occurs as ocular countertorsion reflex23 in re-sponse to a lateral tilt of the head relative to the grav-itation vector. It also can be elicited by a static ordynamic tilt of the visual surroundings, which in-duces a constant angular deflection of the eyes in thedirection of stimulus rotation and which leads to anoptokinetic, torsional nystagmus.4'5 Large torsionaleye movements also can be executed voluntarily byhighly trained human subjects.6
From the Department of Biophysics, Division of Biocybernetics,Heinrich-Heine-Universitat Diisseldorf, D-4000 Diisseldorf, Fed-eral Republic of Germany.
Supported by the German Research Association SFB 200-B10and the Ministry for Science and Research in NRW (Eckmiller).
Submitted for publication: September 9, 1988; accepted May 18,1989.
Reprint requests: Dietmar Ott, Abteilung Biokybernetik,Heinrich-Heine-Universitat Diisseldorf, Universitatsstr. 1, D-4000Dusseldorf, F.R.G.
The ocular countertorsion reflex and visually in-duced torsion generally are explained as vestigial at-tempts of the oculomotor system to compensate ro-tatory image displacements for the benefit of (hy-pothetized) improved visual acuity.7'8 In addition,"miniature" torsional eye movements have been de-scribed to occur during fixation39"13 and presumablyalso during pursuit of moving visual targets.
Ocular torsion has proven difficult if not impossi-ble to record, and former measurement techniques(eg, orientation of after-images;14 iris photography;15
plot of the blind spot;16 attachment of optical leversto the cornea17 generally did not produce precise andtime-resolved data. The recent introduction of mod-ern, real-time video-based18 and electromagnetic3
measurement devices has considerably facilitated theaccurate detection of ocular torsion.
The interpretation of ocular torsion has been com-plicated by the existence of two different types ofocular torsion: real torsion about the visual axis, and"false" torsion.14 Real torsion is actively generated bythe oculomotor system by synergistic innervation ofthe extraocular muscles. "False" torsion, in contrast,
2512
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933145/ on 07/13/2018

No. 12 OCULAR TORSION DURING HORIZONTAL PURSUIT / Orr ond Eckmiller 2513
represents a systematic tilt of the normally verticaleye meridian in tertiary (oblique) gaze positions; thetilt is caused by the combined horizontal and verticaltranslation that occurs according to Listing's law.14
Recently, the characteristics of ocular torsion dur-ing circular pursuit and static fixation were evaluatedby Ferman et al1112 by means of a three-dimensionalscleral search coil technique for the simultaneous de-tection of horizontal, vertical, and torsional eye rota-tions. These authors described the occurrence ofspontaneous, rapid torsional eye movements (up to8°) and slow torsional drifts during fixation at variousfixed positions, but did not find a systematic differ-ence in ocular torsion at various horizontal gaze po-sitions. In contrast, ocular torsion during pursuit of atarget moving in a circular trajectory was found to besignificantly correlated with eye position.
No study of ocular torsion during pure horizontalor pure vertical pursuit is currently available. Withthese parameters, "false" torsion is absent, and con-sequently, any torsional movements would have tobe actively produced by neural control signals. Theretinal events during pursuit as generated by oculartorsion could be relevant for visual pattern recogni-tion as well as for figure-ground discrimination.19
In the current study, the time courses of oculartorsion during horizontal foveal pursuit as comparedwith those during fixation were measured directly asrotations of the ocular fundus in human subjects andtrained monkeys. The fundus images were generatedwith a conventional-television (TV) ophthalmoscopeand with the newly available scanning laser ophthal-moscope (SLO), which monitors the ocular fundusmovement and the relative movements of the retinalprojection of the visual target with high precision. Weprovide evidence for actively generated torsional eyemovements with highly different characteristics dur-ing foveal pursuit and fixation in the horizontalplane. A short summary of this paper has been pre-sented elsewhere.2021
Materials and Methods
Recording Technique in Monkeys
Two adult, female monkeys (Macaca fascicularis)were trained to pursue a small light dot (8' arc) mov-ing horizontally within the central ±5° on a frontal,unstructured screen at 1.5 m distance. A detailed de-scription of the training procedure is given else-where.22 Background luminance was in the uppermesopic range (1 cd/m2). Standard ophthalmologicinspection of the eyes and the fundus showed no ob-vious abnormalities. One animal (FA20) had normalrefraction, and the other (FA21) was myopic (-4
diopters) in both eyes. The monkeys were seated inan upright position with the head (stereotaxic hori-zontal plane perpendicular to the gravity vector)firmly attached to the upper part of the primate chairby means of a headgear, which was stereotacticallyimplanted into the skull. The primary position ofgaze (defined on the screen at 1.5 m distance) wasassumed to be in the horizontal plane.
Eye movements of the pursuing eye were measuredwith an infrared oculometer23 which produced analogvoltage signals linearily (at least within the measuredrange between 5° left and 5° right) related to horizon-tal and vertical eye position with a resolution of<0.1° (0-100 Hz bandwidth). The oculometer wascalibrated by means of the visual target, which themonkeys were trained to fixate at different positions.Possible refraction errors were compensated for by acorrecting lens in front of the pursuing eye. The hori-zontal and vertical eye and the visual target positionsignals were digitized (8 bit, 100 Hz) by an Applemicrocomputer and displayed on the monitor as sep-arate cursors moving in real time along a calibratedscale in the upper half of the video frame (Fig. 1,bottom).
A schematic illustration of the TV-ophthalmo-scope is shown in the top of Figure 1. The fundus ofthe nonpursuing eye was illuminated from the side byophthalmoscopic lenses (13 diopters) and by a nor-mal and a half-reflective mirror (HM) placed in frontof this eye. To minimize interfering reflections fromthe cornea, the entering light cone was confined to adiameter of a little less than that of the pharmacologi-cally dilated pupil and thus illuminated a circularfundus area of about 20° in diameter. The reflectedlight from the fundus was deflected by the HM andcollimated by a second ophthalmoscopic lens to anaerial image of the fundus, which in turn was re-corded by a light-sensitive CCD-video camera (Sie-mens K211). The camera video signal (image repeti-tion rate, 50 Hz) was electronically mixed into thecomputer video signal to yield a composite video sig-nal comprising the ocular fundus image and the posi-tion signals of the visual target and of the contralat-eral eye (Fig. 1, bottom).
In each monkey the fundus motion of both eyeswas successively recorded during horizontal pursuit(or fixation) of the contralateral eye. Pursuit frequen-cies ranged from 0.3 to 0.9 Hz. (Beyond 0.9 Hz thefundus images were blurred and later could not beanalyzed properly.) The fundus luminance (approxi-mately 20,000 cd/m2, lower than in a conventionalfundus camera) was fairly well tolerated by bothmonkeys and did not cause obvious impairment totheir pursuit performance. Our investigation adhered
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933145/ on 07/13/2018

2514 INVESTIGATIVE OPHTHALMOLOGY & VISUAL SCIENCE / December 1989 Vol. 30
Screen
TV - Ophthalmoscope
Lamp
Video
CameraOculometer
OS
VideoCamera
Computer
OD
Target HorizontalPosition Eye Vertical
Position EyePosition
Video-Tape
PictureSplitting
Unit
Fig. 1. Schematic repre-sentation of the experimen-tal setup used in the mon-key experiments. Top: Op-tical arrangement of theTV-ophthalmoscope, thepresentation of the visualtarget, and the measure-ment of eye movements bymeans of infrared oculome-try. L,, L2, L3 = ophthal-moscopic lenses; M = mir-ror; HM = half-reflectivemirror; OS = left eye; OD= right eye. Bottom: Video-electronic arrangement forgeneration of a compositevideo signal comprising theocular fundus image of oneeye and the position signals,represented by computergraphics, of the visual targetand the contralateral eye.MUX = multiplexer; A/D= analog to digital con-verter. See text for details.
to the ARVO Resolution on the Use of Animals inResearch.
To evaluate possible errors induced by misalign-ment of the optical axes in the TV-ophthalmoscope(and to test the image analysis procedure), ocular tor-sion in one monkey was additionally measured bydirect, frontal recording of the rotation of two smallink cross-marks applied to the corneal surface.
In additional measurements, both monkeys pur-
Monitor
sued the visual target moving sinusoidally along ahorizontal trajectory 5° above and 5° below the hori-zontal plane.
Recording Technique in Humans
In five healthy human subjects with normal uncor-rected vision, the fundus motion of the pursuing eyewas measured by means of a prototype SLO (Roden-stock Instruments, Miinchen, FRG). A detailed de-
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933145/ on 07/13/2018

No. 12 OCULAR TORSION DURING HORIZONTAL PURSUIT / Orr ond Eckmiller 2515
scription of the SLO is given elsewhere.24'25 Briefly, acollimated He-Ne laser beam focused by the eye to aspot of about 2-4' arc in diameter was rapidlyscanned across the fundus by two orthogonal deflect-ing scanning mirrors. The reflected light from anygiven illuminated fundus point was collected by aphotodetector, thus generating a time-resolved(video-compatible, image repetition rate 50 Hz) se-quential imaging of the ocular fundus. As a result ofthe scanning process, the subject perceived a moder-ately bright, homogeneous, rectangular red scan ras-ter covering a visual field of 16° horizontally and 12°vertically with an average luminance of about 5000cd/m2. By varying the intensity of the scanning laserbeam (with an acousto-optic modulator placed be-tween the laser source and the scanners), any dy-namic stimulus pattern could be produced on thesubject's retina; the pattern was simultaneouslyviewed by the subject and, on the video monitor,superimposed on the generated fundus image. Theretinal position of each point of the stimulus patternwas thus seen directly on the retina, as were retinalshifts (due to eye movements) that the subject per-formed in observing the pattern. In the current study,the intensity modulation of the scanning laser beamwas achieved by means of a video camera (externallysynchronized by the SLO) which recorded the sinu-soidal motion of a small light dot. The pursuit stimu-lus (the moving dot) was perceived as a black dotagainst the red, homogeneous laser raster.
Subjects looked with the pursuing eye (accommo-dation at infinity) into the SLO with the head hori-zontally fixed by a bite-board in an upright position(forehead and mouth in a common frontal plane) andpursued the visual target (a dot of 8' arc in diameter)moving horizontally (0.3-0.9 Hz, ±5°) against thehomogeneous, red scan raster. The conjugate (non-pursuing) eye was not covered. In one subject (SC)both eyes were measured successively; in the otherfour subjects, measurements were made only with theright eye. Subjects were informed about the nature ofthe experiments and participated voluntarily.
As a control, pure horizontal eye movements (withno torsional component) were simulated by horizon-tal rotations of a Zeiss artificial eye (with the sameoptical properties and retinal dimensions as a naturalhuman eye) in both the TV-ophthalmoscope andSLO setups.
Data Analysis
The image sequences generated by the TV-oph-thalmoscope or the SLO were stored on VHS video-tape (JVC HRD-755) and subsequently subjected toan offline, frame-by-frame rotation analysis. A digitalvideo pointer26 in combination with an Apple micro-
computer and a graphics tablet was used to determinethe Cartesian coordinates of the foveal pit (sharplyimaged as a small bright reflection point) and thebifurcation of a clearly visible retinal vessel within thestanding fundus image. The slope of a line passingthrough these two retinal landmarks yielded the rawangle of ocular torsion.* This procedure was repeatedfor a sequence of about 100 successive video framescorresponding to a recording time of 4 sec. (Due tothe limited capability of the VHS videotape recorder,only the first field of a given frame could be captured,thus yielding a temporal resolution of 40 ms). We areaware that this temporal resolution was inadequatefor the detection of short-duration saccades, but itproved sufficient for normal smooth pursuit eyemovements as measured in this study.f The spatial
* The fundus image on the monitor represented a planar projec-tion of the spherically curved fundus, causing nonlinear distortionof the retinal coordinates at eccentric, horizontal eye positions.This projection-caused distortion could be eliminated by trans-forming the Cartesian coordinates of the foveal pit and the blood-vessel bifurcation (measured at a given horizontal eye position) tothe corresponding values at primary position. Cartesian coordi-nates (x, anterior; y, left; z, superior) are related to spherical coordi-nates by:
x = sin (0) cos (p)
y = sin (i?) sin (p)
z = cos (i?)
with t? = the vertical angle, and p = the horizontal angle of a givenpoint on the fundus. The calibration of the two coordinate systemswas done by means of the known angular distance (16°) betweenthe foveal pit and the optic disc. The transformation matrix M forhorizontal rotation about the z-axis is:
cos (-0) sin (-0) 0
M = -sin (-0) cos (-0) 0
0 0 1
with 0 = the angle of horizontal eye rotation. The primary positioncoordinates were obtained by multiplying the Cartesian xyz-vectorwith the rotation matrix M.
f Since submission of the current paper our fundus recordingequipment was improved in the following ways: 1) An U-maticVTR (Sony VO-9600P) with Frame Code Generator (SonyFCG-700) was used, which allowed computer-controlled search ofindividual video fields, thus yielding a temporal resolution of 20 msfor data analysis (which allows the monitoring even of short-dura-tion saccadic eye movements). 2) In the production model of theSLO (which became available from Rodenstock Instruments in thespring of 1989, the maximal size of the scanned retinal surface was40°, thus allowing the measurement of much greater eye move-ment amplitudes. 3) The video pointer has been replaced by adigital video frame memory (Leutron Vision, Gernlinden, FRG),which, by setting a computer controlled cursor to the overlay plane,increases the number of resolvable fundus points to 512* 512. Thevideo frame memory is also used to control the acousto-optic mod-ulator that varies the intensity of the scanning laser beam. By thismeans, dynamic stimulus patterns can be programmed easily on apersonal computer.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933145/ on 07/13/2018
2516 INVESTIGATIVE OPHTHALMOLOGY 6 VISUAL SCIENCE / December 1989 Vol. 30
resolution of the video pointer was 410*285 pixelsallowing an angular resolution of ocular torsion ofabout 0.2°.
In the monkey recordings, the position of the visualtarget and the pursuing eye were displayed within theocular fundus image and could therefore be deter-mined directly by decoding the cursor positions onthe calibrated scale. In the human recordings, thehorizontal and vertical eye position was determinedby means of the translatory excursions of the fovealpit on the monitor. These excursions were calibratedwith respect to the angular distance (16°) between theoptic disc and the fovea.
Results
Ocular Torsion during Fixation
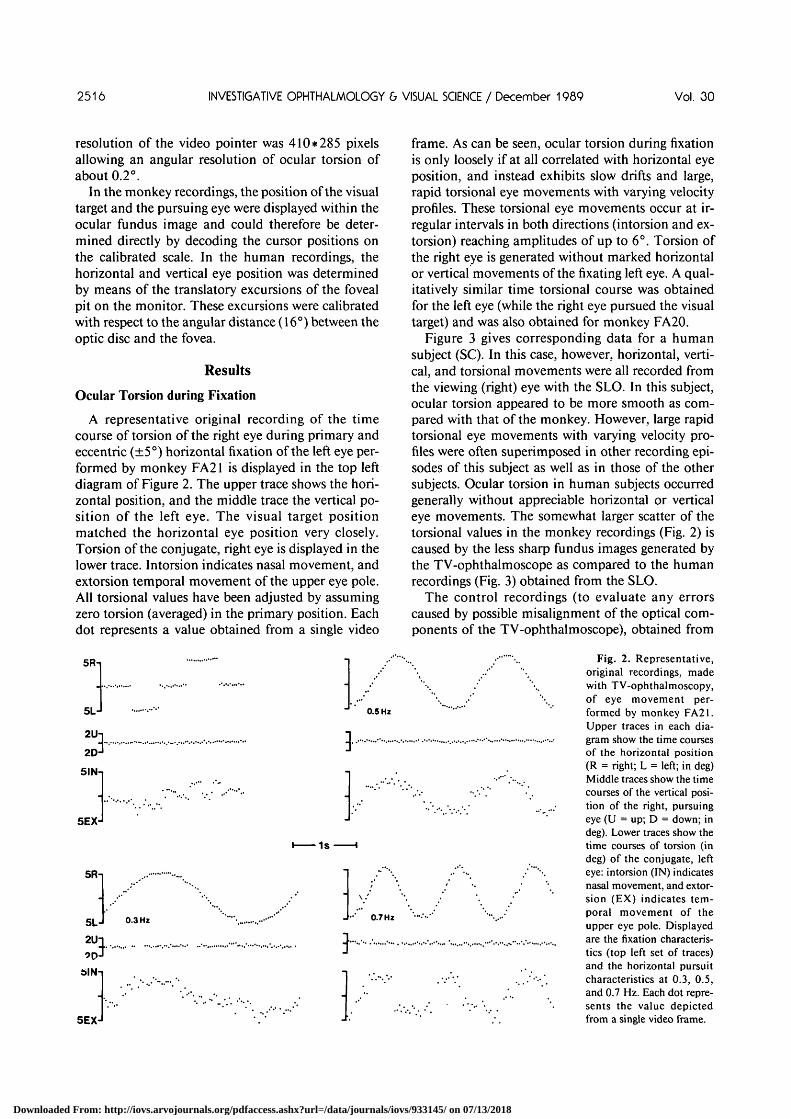
A representative original recording of the timecourse of torsion of the right eye during primary andeccentric (±5°) horizontal fixation of the left eye per-formed by monkey FA21 is displayed in the top leftdiagram of Figure 2. The upper trace shows the hori-zontal position, and the middle trace the vertical po-sition of the left eye. The visual target positionmatched the horizontal eye position very closely.Torsion of the conjugate, right eye is displayed in thelower trace. Intorsion indicates nasal movement, andextorsion temporal movement of the upper eye pole.All torsional values have been adjusted by assumingzero torsion (averaged) in the primary position. Eachdot represents a value obtained from a single video
5Rn
5LJ
2 U ]2DJ
5INn
5EXJ
5Rn
2U
5IN-
5EX-
0.5 Hz
1s
0.3 Hz 0.7 Hz
}.,„.
frame. As can be seen, ocular torsion during fixationis only loosely if at all correlated with horizontal eyeposition, and instead exhibits slow drifts and large,rapid torsional eye movements with varying velocityprofiles. These torsional eye movements occur at ir-regular intervals in both directions (intorsion and ex-torsion) reaching amplitudes of up to 6°. Torsion ofthe right eye is generated without marked horizontalor vertical movements of the fixating left eye. A qual-itatively similar time torsional course was obtainedfor the left eye (while the right eye pursued the visualtarget) and was also obtained for monkey FA20.
Figure 3 gives corresponding data for a humansubject (SC). In this case, however, horizontal, verti-cal, and torsional movements were all recorded fromthe viewing (right) eye with the SLO. In this subject,ocular torsion appeared to be more smooth as com-pared with that of the monkey. However, large rapidtorsional eye movements with varying velocity pro-files were often superimposed in other recording epi-sodes of this subject as well as in those of the othersubjects. Ocular torsion in human subjects occurredgenerally without appreciable horizontal or verticaleye movements. The somewhat larger scatter of thetorsional values in the monkey recordings (Fig. 2) iscaused by the less sharp fundus images generated bythe TV-ophthalmoscope as compared to the humanrecordings (Fig. 3) obtained from the SLO.
The control recordings (to evaluate any errorscaused by possible misalignment of the optical com-ponents of the TV-ophthalmoscope), obtained from
Fig. 2. Representative,original recordings, madewith TV-ophthalmoscopy,of eye movement per-formed by monkey FA21.Upper traces in each dia-gram show the time coursesof the horizontal position(R = right; L = left; in deg)Middle traces show the timecourses of the vertical posi-tion of the right, pursuingeye (U = up; D = down; indeg). Lower traces show thetime courses of torsion (indeg) of the conjugate, lefteye: intorsion (IN) indicatesnasal movement, and extor-sion (EX) indicates tem-poral movement of theupper eye pole. Displayedare the fixation characteris-tics (top left set of traces)and the horizontal pursuitcharacteristics at 0.3, 0.5,and 0.7 Hz. Each dot repre-sents the value depictedfrom a single video frame.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933145/ on 07/13/2018

No. 12 OCULAR TORSION DURING HORIZONTAL PURSUIT / Orr ond Eckmiller 2517
Fig. 3. Representative,original recordings, madewith SLO, of eye movementperformed by subject SC.This diagram is analogousto that of Figure 2, with theexception that horizontal(upper trace), vertical (mid-dle trace), and torsional(lower trace) eye move-ments here were all re-corded from the pursuing(right) eye. R = right; L= left; U = up; D = down;IN = intorsion; EX = ex-torsion. Eye movements indeg.
5LJ
2U
2D5IN-i
5EXJ
5R-i
5L-I2 U :2D-
5IN-i
5EX-
1.
0.3 Hz
-*-' 0.5 Hz
•1S
0.7 Hz
direct frontal recording of the rotation of the inkcrosses on the corneal surface yielded torsional timecourses qualitatively and quantitatively similar tothose obtained from the rotation of the retinal land-marks.
Ocular Torsion during Pursuit
Figures 2 and 3 show representative original re-cordings of ocular torsion during horizontal fovealpursuit eye movements in monkey FA21 (Fig. 2) andsubject SC (Fig. 3) at different pursuit frequencies(0.3, 0.5, and 0.7 Hz). Ocular torsion during pursuitbetween 5° left and 5° right sinusoidally modulatedup to 8° peak-to-peak, approximately in phase withhorizontal eye position. Intorsion occurred alwaysduring pursuit in the temporal direction and extor-sion during pursuit in the nasal direction, yielding eyemovements comparable to the motion of windshield-wipers. Smooth position-dependent ocular torsionwas often superimposed by irregularly occurringrapid torsional eye movements similar to those oc-curing during fixation. These rapid movements oc-curred as single events (at 0.5 Hz, as in Fig. 2) or inquick repetition (at 0.5 Hz, as in Fig. 3). Also, there isno qualitatively noticeable difference between thetorsional modulation of the pursuing eye (humans,Fig. 3) and the conjugate, nonpursuing eye (monkeys,Fig. 2). Ocular torsion at 0.9 Hz pursuit frequency(not shown) was qualitatively similar to that at thelower frequencies. Also, the torsional characteristicsof pursuit episodes 5° above and below the eye hori-
zontal plane showed no measurable increase or de-crease in ocular torsion as compared to the normalstimulation paradigm (stimulus moving in the hori-zontal plane).
The torsional peak-to-peak modulation and phase(re. horizontal eye position) of all eyes tested in bothmonkeys and human subjects are listed in Table 1.The values show the mean (+ standard deviation,SD) pooled for n = 30 cycles of pursuit. Positive (ornegative) phase values indicate that torsion leads (orlags) horizontal eye position. The data of this tablecan be summarized as follows:
1. Ocular torsion reached modulation values of upto 7.6° peak-to-peak (mean + 1 SD) for horizontalpursuit modulation of 10° peak-to-peak, thus yield-ing a dynamic torsional gain (torsional modulation/horizontal pursuit modulation) GT of up to 0.76.
2. Ocular torsion of the pursuing eye (humans)was very similar to ocular torsion of the conjugate,nonpursuing eye (monkeys).
3. Torsional phase and modulation depth showedconsiderable intra-individual fluctuation (SD) andlarge interindividual variability. In two subjects (PHand DO), the peak-to-peak modulation was below 1 °and could therefore not be evaluated with sufficientaccuracy. Nevertheless, these subjects too generatedrapid torsional eye movements similar to those of theother subjects (Fig. 3).
4. Ocular torsion exhibited no significant depen-dence on the pursuit frequency, at least not in therange from 0.3 to 0.9 Hz.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933145/ on 07/13/2018

2518 INVESTIGATIVE OPHTHALMOLOGY & VISUAL SCIENCE / December 1989 Vol. 30
Table 1. Average (±SD) modulation depth and phase of ocular torsion of all eyes tested in both monkeysand human subjects at various pursuit frequencies
Subject
Monkey 1(FA20)
Monkey 2(FA21)
Subject 1(SC)
Subject 2(PS)
Subject 3(PH)
Subject 4(DO)
Subject 5(RE)
Eye
OD*OS*OD*OS*ODfOSfODf
ODf
ODf
ODf
0.3 Hz
Modulation(deg)
5.8 ± 1.04.3 ± 1.45.4 ± 1.26.0 ± 1.43.2 ± 1.04.2 ± 0.93.7 ± 0.6
3.2 ± 0.4
Phase (deg)
3.8 ±6.1 ±2.8 ±2.0 ±
-3.1 ±5.7 ±
11.4 ±
5.0 ±
3.86.78.39.66.47.2
19.3
7.1
0.5
Modulation(deg)
4.0 ± 0.64.8 ± 1.06.0 ± 0.84.8 ± 0.85.0 ± 0.94.5 ± 1.15.2 ± 0.5
Pursuit frequency
Hz
Phase (deg)
-3.3 ± 7.50.8 ± 8.8
14.4 ± 14.69.7 ± 9.4
14.2 ± 12.39.6 ± 11.88.7 ± 7.3
0.7 Hz
Modulation(deg)
4.0 ± 0.64.2 ± 0.66.2 ± 1.06.2 ± 0.64.4 ± 0.84.4 ± 0.84.0 ± 1.4
— only weak modulation (below 0.8)
— only weak modulation (below 1.0)
4.0 ± 0.2 0.2 ± 1.8 2.9 ± 0.4
Phase (deg)
1.35.1
18.35.5
10.57.5
11.1
25.2
± 7.1± 8.0± 10.5± 9.9± 8.6± 9.0± 12.8
± 2.1
0.9 Hz
Modulation(deg)
—6.4 ± 1.26.0 ± 0.45.4 ± 1.25.0 ± 1.14.7 ± 0.7
3.2 ±0.5
Phase (deg)
—-15.3 ±-7.0 ±20.4 ±13.8 ±12.3 ±
14.3 ±
13.411.97.48.2
13.7
6.9
* Torsion from conjugate, nonpursuing eye.t Torsion from pursuing eye.OD = right eye; OS =left eye.
Fig. 4 shows a comparison between ocular torsionmeasured during fixation (5° left, primary position,5° right) and horizontal pursuit (±5°, 0.5 Hz) forsubject SC (top diagram) and monkey FA21 (bottomdiagram). Data were averaged over all recording epi-sodes (n > 80). The average amount of ocular torsionduring fixation is close to zero; the occurrence of bidi-rectional, rapid torsional eye movements explains therelatively large standard deviation. In contrast, dur-ing horizontal pursuit, ocular torsion is significantlycorrelated with horizontal eye position and exhibits asmaller standard deviation.
The control measurements made with the artificialeye yielded no measurable difference in ocular tor-sion between horizontal position 5° left and 5° right.
Discussion
In the current study, the time courses of oculartorsion (about the visual axis) during fixation andfoveal pursuit (±5°, 0.3-0.9 Hz) in the horizontalplane were measured as the ocular fundus rotationusing TV-ophthalmoscopy in monkeys and SLO inhuman subjects. The major finding was that oculartorsion was clearly correlated with horizontal eye po-sition only during horizontal pursuit and not duringfixation.
Our findings are in part corroborated by those of astudy by Ferman et al.12 These investigators mea-sured ocular torsion during circular pursuit by meansof a scleral search coil. The data in Figures 6 and 7 oftheir report led us to deduce that ocular torsionwould reach values of 7° peak-to-peak when the eye
passed through the 20°-left and 20°-right positionsduring circular pursuit (corresponding to a torsionalgain GT of up to 0.18). However, the directions ofocular torsion that they measured (extorsion withtemporal movements) are opposite to the directionsmeasured in our study (intorsion with temporalmovements). The reason for this discrepancy in tor-sional direction is unclear. Also, their recordings areinterspersed with rapid torsional eye movements ofvarying amplitudes of up to several degrees and withdifferent velocity profiles similar to those demon-strated in the current study during pure horizontalpursuit.
Possible Sources of ErrorOur method of measuring ocular torsion (in addi-
tion to horizontal and vertical eye movements) bymeans of analyzing fundus movements appears tohave a number of advantages over alternativemethods:
1. There is no possibility of slip of a sensor relativeto the eye (as, for example, in techniques using ascleral search coil or an optical lever).
2. Extensive goniometric corrections are not re-quired (as they are in search coil and external video-based techniques).
3. The SLO allows for the first time the exactmonitoring of the retinal projection of a visual stimu-lus (by intensity modulation of the scanning laserbeam) directly on the fundus pattern of the seeingeye. All external measurement techniques (evaluatingmovements of the eyeball rather than the fundus, on
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933145/ on 07/13/2018

No. 12 OCULAR TORSION DURING HORIZONTAL PURSUIT / Orr ond Eckmiller 2519
5 I N -
,£, 5EXJ
HUMAN
OF• P
Z
oenccO
5IN-1
5EXJ
MONKEY
5L 0 5REYE POSITION (DEG)
Fig. 4. The difference in ocular torsion during fixation (F, opencircles) and foveal pursuit at 0.5 Hz (P, filled circles) for subject SC(top) and monkey FA21 (bottom). Pooled data from all recordingepisodes (n > 80). IN = intorsion; EX = extorsion; L = left; R= right. Bars = standard deviations.
which the target projection occurs) presuppose thatthe retina is rigidly attached to the bulbus (rotatingabout a stationary center) and consequently that itfollows exactly the same movement trajectory as theanterior eye pole. This assumption, however, wouldbe justified only if the eye were an ideal, rigid body.Rather, the orbit plant has considerable elastic prop-erties and consequently, friction (from the orbital tis-sues), deformation (due to unbalanced extraocularmuscle innervation), and inertial forces (during eyemovements) may cause relative retinal motion. Fur-thermore, the center of rotation actually is not sta-tionary;27 the assumption that it is stationary causesmisreadings for torsional as well as horizontal or ver-tical rotations of the bulbus.
4. Our measurement of ocular torsion was basedon the evaluation of the shift of retinal coordinatesprojected onto the monitor plane. Whereas this possi-ble source of error is uncritical at the primary posi-tion, ocular torsion measured at eccentric eye posi-tions needs to be corrected for nonlinear projection-caused errors. These errors can easily be calculated bysimple arithmetic operations (see Methods) and havebeen considered in the presented data.
5. The control measurements using the artificialeye (with the same optical properties and retinal di-
mensions as the natural human eye) revealed no mea-surable amount of ocular torsion as a function ofhorizontal rotations in the range of ±5°.
Neurally Controlled Real Ocular Torsion vsKinematically Controlled Torsionaccording to Listing's law
One might argue that the observed torsional modu-lation during pursuit was caused kinematically byorbit plant mechanics ("false" torsion) rather thanactively generated, since we may not have measuredthe primary position as the true center of the pursuitmovements. In this case pursuit might have passedthrough tertiary instead of secondary positions. Withmonkeys, we assumed that the primary positioncould be reached when the line of gaze was in thehorizontal plane of the stereotaxic coordinate system.In humans, we assumed that the line of gaze couldreach the primary position by having the subject lookstraight ahead in a horizontal plane perpendicular tothe vertical forehead-mouth plane. These assump-tions may have caused positive or negative verticaldeviations from the primary position by several de-grees. However, several points exclude this possi-bility:
1. "False" torsion according to Listing's law hasbeen calculated14 and is far below 1° for (theoretic)vertical head deviations of up to 10° and horizontaleye rotations of +5° and can thus be excluded inaccounting for the large torsional modulation foundin the present study.
2. Our control measurements of ocular torsionduring horizontal pursuit at a level 5° above andbelow the normal horizontal plane yielded no notice-able changes in torsional modulation (although"false" torsion according to Listing's law does includedifferent directions for horizontal eye rotations aboveand below the horizontal plane).
3. Ocular torsion was measured by means ofsteady fixation at various positions in the horizontalplane. We found no significant eye-position-depen-dent ocular torsion during fixation, in agreementwith Listing's law.14
Our current findings support the hypothesis20'21
that smooth ocular torsion approximately in phasewith eye position during horizontal pursuit is activelygenerated, presumably by a separate neural oculo-motor subsystem, which generates pursuit-specificretinal events. While the pursued moving target ismaintained close to the foveal center, the stationaryvisual background (eg, a large window frame) yieldsan extrafoveal retinal projection with an uneven dis-tribution of retinal slip velocities. For example, theslip velocity vector field of the window frame projec-
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933145/ on 07/13/2018

2520 INVESTIGATIVE OPHTHALMOLOGY 6 VISUAL SCIENCE / December 1989 Vol. 30
tion results from a vectorial addition of horizontalpursuit velocity and torsional velocity. Thus, the re-sulting slip velocity of a window component pro-jected onto a retinal location above the fovea (as tor-sional center) may be considerably different from thevelocity corresponding to a retinal location below thefovea. It is hypothetized here that this ocular torsion-induced uneven distribution of extrafoveal slip veloc-ities significantly reduces the ability of the visual sys-tem to reassemble the "Gestalt"28 of the backgroundpatterns. In other words, the uneven and constantlychanging slip velocity vector field of the stationarybackground projection can be assumed to assist thenecessary figure-background discrimination29 duringpursuit. The available literature addresses the depen-dence of visual acuity on the location of retinal stim-ulation30-31 as well as the dependence of the visualpattern recognition performance on the orientationof the test pattern relative to the retinal merid-ians.32'33 A test of our hypothesis will require the fol-lowing future experiments: 1) figure-background dis-crimination in the presence of moving extrafovealpatterns during fixation, and 2) figure-backgrounddiscrimination during pursuit in the presence of sta-tionary extrafoveal patterns.
Key words: ocular torsion, monkey, human, smooth pur-suit eye movements, scanning laser ophthalmoscopy
Acknowledgments
The authors are indebted to Drs. Andreas Plesch andUlrich Klingbeil (Rodenstock Instruments) for the kinddisposition of a prototype version of the scanning laser oph-thalmoscope and for valuable technical support. Thanksare also due to Petra Sarfert, Sabine Canditt, and PeterHoltermann for serving as patient and interested subjects.
References
1. Duke-Elder WS: Textbook of Ophthalmology, Vol. 4. St.Louis, CV Mosby, 1949, p. 3945.
2. Diamond SG and Markham CH: Ocular counterrolling as anindicator of vestibular otolith function. Neurology 33:1460,1983.
3. Collewijn H, Van der Steen J, Ferman L, and Jansen TC:Human ocular counterroll: Assessment of static and dynamicproper ties from electromagnetic scleral coil recordings. ExpBrain Res 59:185, 1985.
4. Brecher GA: Die optokinetische auslosung von augenrollungund rotatorischem nystagmus. Pfliigers Archiv f d ges Physiol234:13, 1934.
5. Howard IP and Templeton WB: Visually induced eye torsionand tilt adaptation. Vision Res 4:433, 1964.
6. Balliet R and Nakayama K: Training of voluntary torsion.Invest Ophthalmol Vis Sci 17:303, 1978.
7. Merton PA: Compensatory rolling movements of the eye. JPhysiol (Lond) 132:25, 1956.
8. Petrov AP and Zenkin GM: Torsional eye movements andconstancy of the visual field. Vision Res 13:2465, 1973.
9. Fender PH: Torsional motions of the eyeball. Br J Ophthalmol39:65, 1955.
10. Boeder P: An analysis of the general type of uniocular rota-tions. Arch Ophthalmol 57:200, 1957.
11. Ferman L, Collewijn H, and van den Berg AV: A direct test ofListing's law: I. Human ocular torsion measured in static ter-tiary positions. Vision Res 27:929, 1987.
12. Ferman L, Collewijn H, and van den Berg AV: A direct test ofListings law: II. Human ocular torsion measured under dy-namic conditions. Vision Res 27:939, 1987.
13. Ferman L, Collewijn H, Jansen TC, and van den Berg AV:Human gaze stability in the horizontal, vertical and torsionaldirection during voluntary head movements, evaluated with athree-dimensional scleral induction coil technique. Vision Res27:811, 1987.
14. Helmholtz H: Handbuch der Physiologischen Optik, Gull-strand A, 3rd ed, von Kries J, and Nagel W, editors. Hamburg,Leipzig, Voss Verlag, 1910, pp. 34-104.
15. Moses RA: Torsion of the eye on oblique gaze. Arch Ophthal-mol 44:136, 1950.
16. Quereau JVD: Rolling of the eye around its visual axis duringnormal ocular movements. Arch Ophthalmol 53:807, 1955.
17. Ditchburn RW and Ginsborg BL: Involuntary eye movementsduring fixation. J Physiol (Lond) 119:1, 1953.
18. Enright JT: Saccadic anomalies: Vergence induces large de-partures from ball-on-socket behaviour. Vision Res 24:301,1984.
19. Eckmiller R: Neural control of pursuit eye movements. Phys-iol Rev 67:797, 1987.
20. Eckmiller R and Ott D: Torsional eye movements in the mon-key during horizontal foveal pursuit: Behavioural and neuro-physiological evidence. Soc Neurosci Abstr 13:171, 1987.
21. Eckmiller R and Ott D: Function of ocular torsion duringhorizontal pursuit in man and monkey. ARVO Abstracts. In-vest Ophthalmol Vis Sci 29(Suppl):166, 1988.
22. Eckmiller R and Mackeben M: Pursuit eye movements andtheir neural control in the monkey. Pflugers Arch 377:15,1978.
23. Bach M, Bouis D and Fischer B: An accurate and linear infra-red oculometer. J Neurosci Methods 9:9, 1983.
24. Webb RH, Hughes GW, and Delori FC: Confocal scanninglaser ophthalmoscope. Applied Optics 26:1492, 1987.
25. Plesch A, Klingbeil U, and Bille J: Digital laser scanningfundus camera. Applied Optics 26:1480, 1987.
26. Ott D: A digital video pointer for the analysis of videotapedocular fundus motion. Neurosci Lett Suppl 173, 1988.
27. Enright JT: The aftermath of horizontal saccades: Saccadicretraction and cyclotorsion. Vision Res 26:1807, 1986.
28. Zusne L: Visual Perception of Form. New York, AcademicPress, 1970, p. 113.
29. Rock I: The description and analysis of objects and event per-ception. In Handbook of Perception and Human Perfor-mance, Vol 2: Cognitive Processes and Performance, BoffKR,Kaufman L, and Thomas JP, editors. John Wiley & Sons,1986, p. 33.
30. Kerr JL: Visual resolution in the periphery. Perception andPsychophysics 9:375, 1971.
31. Westheimer G and McKee S: Visual acuity in the presence ofretinal image motion. J Opt Soc Am [A] 65:847, 1975.
32. Mitchell DE, Freeman RD, and Westheimer G: Effect of ori-entation on the modulation sensitivity for interference fringeson the retina. J Opt Soc Am [A] 57:246, 1967.
33. Berkley MA, Kitterle F, and Watkins DW: Gratings visibilityas a function of orientation and retinal eccentricity. Vision Res15:239, 1975.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933145/ on 07/13/2018





























