Embed Size (px)
Citation preview

A
ertctm(Ahfrt©
K
1
w2ptitft[
0d
Brain Research Bulletin 71 (2007) 601–609
Nucleus reuniens of the midline thalamus: Link between the medialprefrontal cortex and the hippocampus
Robert P. Vertes a,∗, Walter B. Hoover a, Klara Szigeti-Buck b, Csaba Leranth b
a Center for Complex Systems and Brain Sciences, Florida Atlantic University, Boca Raton, FL 33431, United Statesb Department of Obstetrics, Gynecology and Reproductive Sciences and Neurobiology, Yale University,
School of Medicine, New Haven, CT 05620, United States
Received 22 September 2006; received in revised form 6 November 2006; accepted 5 December 2006Available online 3 January 2007
bstract
The medial prefrontal cortex and the hippocampus serve well recognized roles in memory processing. The hippocampus projects densely to, andxerts strong excitatory actions on, the medial prefrontal cortex. Interestingly, the medial prefrontal cortex, in rats and other species, has no directeturn projections to the hippocampus, and few projections to parahippocampal structures including the entorhinal cortex. It is well establishedhat the nucleus reuniens of the midline thalamus is the major source of thalamic afferents to the hippocampus. Since the medial prefrontalortex also distributes to nucleus reuniens, we examined medial prefrontal connections with populations of nucleus reuniens neurons projectingo hippocampus. We used a combined anterograde and retrograde tracing procedure at the light and electron microscopic levels. Specifically, we
ade Phaseolus vulgaris-leuccoagglutinin (PHA-L) injections into the medial prefrontal cortex and Fluorogold injections into the hippocampusCA1/subiculum) and examined termination patterns of anterogradely PHA-L labeled fibers on retrogradely FG labeled cells of nucleus reuniens.t the light microscopic level, we showed that fibers from the medial prefrontal cortex form multiple putative synaptic contacts with dendrites ofippocampally projecting neurons throughout the extent of nucleus reuniens. At ultrastructural level, we showed that medial prefrontal cortical fibers
orm asymmetric contacts predominantly with dendritic shafts of hippocampally projecting reuniens cells. These findings indicate that nucleuseuniens represents a critical link between the medial prefrontal cortex and the hippocampus. We discuss the possibility that nucleus reuniens gateshe flow of information between the medial prefrontal cortex and hippocampus dependent upon attentive/arousal states of the organism.2007 Elsevier Inc. All rights reserved.
eywords: Prelimbic cortex; Working memory; Mediodorsal nucleus; Entorhinal cortex; Rat
thnBwutc
. Introduction
The hippocampus and medial prefrontal cortex (mPFC) serveell recognized roles in memory processing [1,2,15–17,20–2,50,54]. The hippocampus distributes heavily to the medialrefrontal cortex [12,18,28,29,31,43,47] and exerts strong exci-atory actions on the mPFC [18,30,33,34]. Despite the directnnervation/influence of the hippocampal formation (HF) onhe mPFC, interestingly, there are no direct return projections
rom the mPFC to HF, and rather moderate mPFC projectionso parahippocampal structures including the entorhinal cortex6,9,27,33,37,39,40,45,49].∗ Corresponding author. Tel.: +1 5612972362; fax: +1 5612972363.E-mail address: [email protected] (R.P. Vertes).
cdhpd
l
361-9230/$ – see front matter © 2007 Elsevier Inc. All rights reserved.oi:10.1016/j.brainresbull.2006.12.002
The nucleus reuniens (RE) of the ventral midline thalamus ishe principal (or virtually sole) source of thalamic input to theippocampus [8,24,42,53,55–57]. RE stimulation produces pro-ounced excitatory actions at CA1 of the hippocampus [7,14].ertram and Zhang [7] compared the effects of stimulation of REith stimulation of the CA3 region of the hippocampus on pop-lation responses (field EPSPs and spikes) at CA1, and reportedhat RE actions on CA1 were equivalent to, and in some casesonsiderably greater than, those of CA3 on CA1. They con-luded that the RE projection to the hippocampus “allows for theirect and powerful excitation of the CA1 region. This thalamo-ippocampal connection bypasses the trisynaptic/commissural
athway that has been thought to be the exclusive excitatoryrive to CA1.”It has recently been shown [48,49] that the mPFC, particu-arly the infralimbic (IL) and prelimbic (PL) cortices, distribute

6 arch
pmmpdp(olfitdRH(
2
2
iUfa
aflrsms4GH
siamswaT+
pcifibv
2
fiiitaas
Fsg
1dcdtfararP1ssiba0
f(iEv3wfD2rPiErdrAa
bolaiSAc
2
ti0tmmbw
02 R.P. Vertes et al. / Brain Rese
rominently to RE. The combined demonstration, then, thatPFC projects to RE and RE in turn to HF, suggests that REay represent an important relay between the mPFC and hip-
ocampus. It remains to be determined whether mPFC fibersistributing to RE contact RE neurons projecting to the hip-ocampus. To assess this, we made anterograde tracer injectionsPHA-L) in the ventral mPFC and retrograde injections (Flu-rogold) in the CA1/subiculum of HF and examined, at theight and ultrastructural level, synaptic connections of mPFCbers on RE neurons projecting to the hippocampus. We showed
hat mPFC fibers form asymmetric (excitatory) contacts pre-ominantly on dendritic shafts of RE cells projecting to HF.E thus appears to be a critical link between the mPFC andF, completing an important loop between these structures
HF → mPFC → RE → HF).
. Materials and methods
.1. Animals and surgical protocols
Twelve male Sprague–Dawley rats (Charles River, Wilmington, MA) weigh-ng 275–350 g were used. Experiments were approved by the Florida Atlanticniversity Institutional Animal Care and Use Committee and conform to all
ederal regulations and the National Institute of Health guidelines for the carend use of laboratory animals.
Under sodium pentobarbital anesthesia (50 mg/kg, ip), each rat receivedn injection of Phaseolus vulgaris-leucoagglutinin (PHA-L) in the medial pre-rontal cortex and an injection of Fluorogold (FG) in the hippocampus. Powderedectin from P. vulgaris-leucoagglutinin (Vector Labs, Burlingame, CA) waseconstituted to 5% in 0.05 M sodium phosphate buffer, pH 7.4. The PHA-Lolution was iontophoretically deposited in the brains of anesthetized rats byeans of a glass micropipette with an outside tip diameter of 40–60 �m. The
tereotaxic coordinates were: AP, +2.6 to 2.8 mm to bregma, L, 0.5 mm and V,.5–5.2 mm. Positive direct current pulses (5–10 �A) were applied through arass stimulator (Model 88) coupled with a high voltage stimulator (Frederickaer Co., Brunswick, ME) at 2 s “on”/2 s “off” intervals for 40–50 min.
Fluorogold (Fluorochrome, LLC, Denver, CO) was dissolved in a 0.1 Modium acetate buffer (pH 5.0) to yield a 5% concentration. FG was eitherontophoretically deposited in the hippocampus or pressure injected using
1 �l Hamilton syringe. FG was iontophoretically injected through a glassicropipette with an outside tip diameter of 40–50 �m for 5–10 min using the
ame protocol described above for PHA-L. The pressure injections (0.03 �l)ere made slowly over 10–15 min and the syringe was left in place for an
dditional 10 min to prevent the tracer from spreading up the injection path.he stereotaxic coordinates were: AP, −5.6 to −5.9 mm to bregma, L, 5.4 to5.8 mm, and V, 4.6–6.2 mm.
After a survival period of 7–10 days, rats were deeply anesthetized anderfused transcardially with 50 ml of heparinized saline, followed by a fixativeontaining 4% paraformaldehyde, 0.1% glutaraldehyde, and 15% picric acidn 0.1 M phosphate buffer (PB) (pH 7.4). The brains were removed and post-xed overnight in 4% paraformaldehyde and 15% picric acid in 0.1 M PB. Therains were then washed with 0.1 M PB subsequently sectioned (50 �m) with aibratome and collected in 0.1 M PB for light and electron microscopic analysis.
.2. Light microscopic procedures
Two procedures were used for the dual visualization of PHA-L labeledbers and FG retrogradely labeled neurons. The first procedure involved an
ntensification of FG retrogradely labeled neurons and the second procedure an
ntensification of PHA-L anterogradely labeled fibers. All sections were initiallyreated with 1% sodium borohydride in 0.1 M PB for 30 min to remove excessiveldehydes. They were then washed three times for 5 min each (3 × 5 min) in PBnd incubated for 60 min in 0.5% bovine serum albumin in 0.1 M Tris-bufferedaline (TBS, pH 7.6) at room temperature (RT) to minimize nonspecific labeling.oodvi
Bulletin 71 (2007) 601–609
ollowing this, sections were incubated for 20 h at RT in diluent [0.1% bovineerum albumin (BSA) in TBS containing 0.25% Triton X-100] and biotinylatedoat anti-PHA-L (Vector Labs, Burlingame, CA) at a dilution of 1:500.
For the first procedure sections were washed (0.1 M PB) and placed for 1 h in:400 concentration of biotinylated rabbit anti-goat immunoglobulin (IgG) andiluent. Following another PB wash, sections were incubated for 1 h in 1:100oncentration of peroxidase–avidin complex (ABC) from the Vector Elite kit andiluent. The peroxidase reaction product was visualized by incubation in a solu-ion containing 0.022% 3,3′-diaminobenzidine (DAB) and 0.003% H2O2 in TBSor 6 min. Sections were then re-incubated in 0.5% BSA in 0.1 M TBS for 15 min,nd incubated for 48 h at RT in 1:200 concentration of rabbit anti-FG (Fluo-ochrome, LLC, Denver, CO) and diluent. Sections were then washed (0.1 M PB)nd placed for 2 h in a 1:100 concentration of a secondary antiserum, donkey anti-abbit immunoglobulin (Jackson ImmunoResearch Laboratories, West Grove,A) and diluent. Following another PB wash, sections were incubated for 2 h in a:200 concentration of rabbit peroxidase-anti-peroxidase (Jackson ImmunoRe-earch Laboratories, West Grove, PA) and diluent. Following 0.1 M PB washes,ections were re-incubated in the same concentrations in donkey anti-rabbitmmunoglobulin and rabbit peroxidase-anti-peroxidase for 2 h each—doubleridge procedure. After final washes (0.1 M PB), the reaction product was visu-lized by an incubation in a solution containing 0.015% 3,3′-diaminobenzidine,.015% cobalt acetate and 0.003% H2O2 in TBS for 6 min.
For the second procedure, sections were washed (0.1 M PB) and placedor 1 h in 1:400 concentration of biotinylated horse anti-goat immunoglobulinIgG) and diluent. Following another PB wash, sections were incubated for 1 hn 1:100 concentration of peroxidase–avidin complex (ABC) from the Vectorlite kit and diluent. After a final PB wash, the peroxidase reaction product wasisualized by incubation in a solution containing 0.015% cobalt acetate, 0.022%,3′-diaminobenzidine (DAB) and 0.003% H2O2 in TBS for 5 min. Sectionsere then re-incubated in 0.5% BSA in 0.1 M TBS for 15 min, and then incubated
or 48 h at RT in 1:200 concentration of rabbit anti-FG (Fluorochrome, LLC,enver, CO) and diluent. Sections were then washed (0.1 M PB) and placed forh in a 1:400 concentration of a secondary antiserum, biotinylated donkey anti-
abbit immunoglobulin (Jackson ImmunoResearch Laboratories, West Grove,A) and diluent. Following another PB wash, sections were incubated for 1 hn 1:100 concentration of peroxidase–avidin complex (ABC) from the Vectorlite kit and diluent. Following a final PB wash, the peroxidase reaction product
eaction was visualized by an incubation in a solution containing 0.015% 3,3′-iaminobenzidine and 0.003% H2O2 in TBS for 5 min. All sections were re-insed in PBS (3 × 1 min) and mounted onto chrome-alum gelatin-coated slides.n adjacent series of sections from each rat was stained with cresyl violet for
natomical reference. Sections were examined using light and darkfield optics.The first procedure gave rise to brown appearing PHA-L labeled fibers on
lack FG labeled cells and the second procedure to black PHA-L labeled fibersn brown FG-labeled cells. The lightfield photomicrographs of injection sites,abeled fibers and cells were taken with a Nikon DXM1200 camera mounted onNikon Eclipse E600 microscope. The captured images were then assembled
nto photomontages using Image-Pro Plus software (Media Cybernetics, Inc.,ilver Spring, MD). Single photomicrographs (non-montages) were taken withCT-1 software (Nikon, USA). Digital images were enhanced (brightness andontrast) using Adobe PhotoShop 9.0 (Mountain View, CA).
.3. Electron microscopic procedures
To allow for antibody penetration, the sections were incubated in a cryopro-ective solution (20% sucrose in 0.1 M PB) until sections sank and freeze-thawedn liquid nitrogen. The tissue was then blocked with a diluent solution (1% BSA,.1% glycine, 0.1% l-lysine, and 4% normal goat serum in 0.1 M PB) for 30 mino reduce non-specific labeling and then incubated for 48 h in a cocktail of pri-ary antibodies against both the PHA-L and FG as described above for lighticroscopic procedures. Following copious washes in PB, sections were incu-
ated in ABC at a concentration of 1:50 in 0.1 M PB for 2 h. Following furtherashes, sections were immersed for 4 min in a nickel-intensified DAB/glucose
xidase solution (15 mg DAB, 12 mg ammonium chloride, 0.12 mg glucosexidase, 600 �l of 0.05 M nickel ammonium sulphate, and 600 �l of 10% B--glucose in 40 ml 0.1 M PB) to visualize the PHA-L reaction product. FG wasisualized using silver intensification of a gold secondary antibody. Follow-ng a DAB reaction, the tissue was washed and blocked again in preparation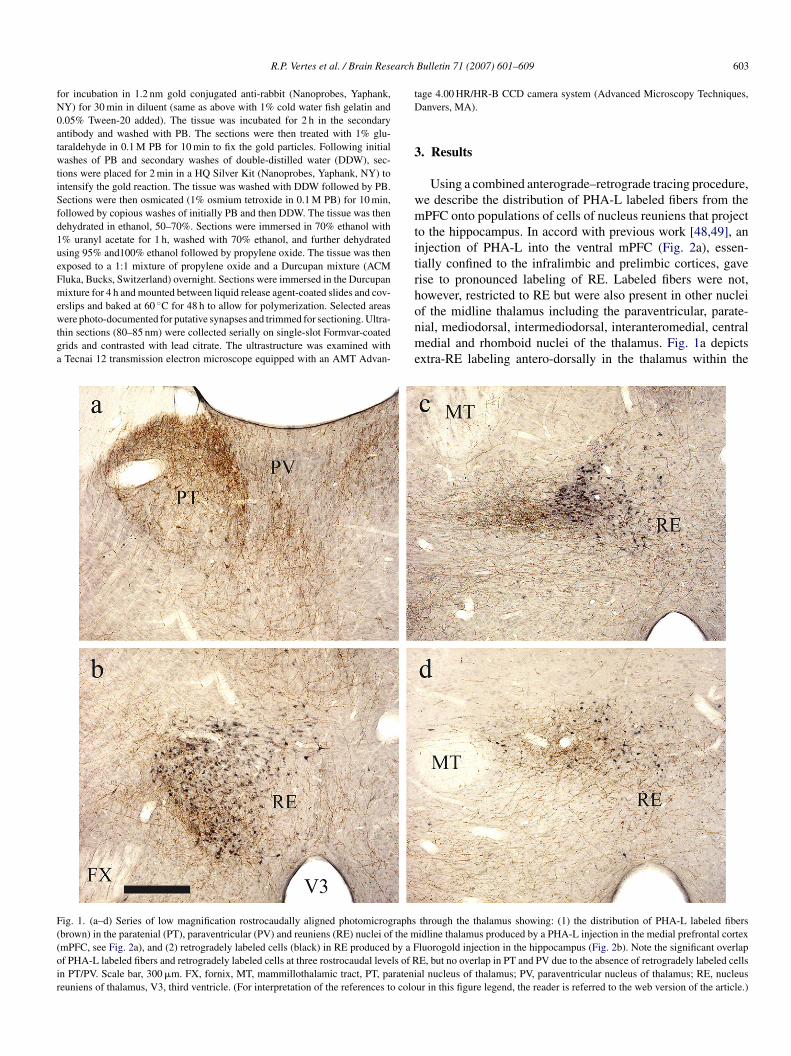
arch
fN0atwtiSfd1ueFmewtga
tD
3
wmtitrh
F((oir
R.P. Vertes et al. / Brain Rese
or incubation in 1.2 nm gold conjugated anti-rabbit (Nanoprobes, Yaphank,Y) for 30 min in diluent (same as above with 1% cold water fish gelatin and.05% Tween-20 added). The tissue was incubated for 2 h in the secondaryntibody and washed with PB. The sections were then treated with 1% glu-araldehyde in 0.1 M PB for 10 min to fix the gold particles. Following initialashes of PB and secondary washes of double-distilled water (DDW), sec-
ions were placed for 2 min in a HQ Silver Kit (Nanoprobes, Yaphank, NY) tontensify the gold reaction. The tissue was washed with DDW followed by PB.ections were then osmicated (1% osmium tetroxide in 0.1 M PB) for 10 min,ollowed by copious washes of initially PB and then DDW. The tissue was thenehydrated in ethanol, 50–70%. Sections were immersed in 70% ethanol with% uranyl acetate for 1 h, washed with 70% ethanol, and further dehydratedsing 95% and100% ethanol followed by propylene oxide. The tissue was thenxposed to a 1:1 mixture of propylene oxide and a Durcupan mixture (ACMluka, Bucks, Switzerland) overnight. Sections were immersed in the Durcupanixture for 4 h and mounted between liquid release agent-coated slides and cov-
rslips and baked at 60 ◦C for 48 h to allow for polymerization. Selected areasere photo-documented for putative synapses and trimmed for sectioning. Ultra-
hin sections (80–85 nm) were collected serially on single-slot Formvar-coatedrids and contrasted with lead citrate. The ultrastructure was examined withTecnai 12 transmission electron microscope equipped with an AMT Advan-
onme
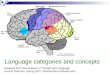
ig. 1. (a–d) Series of low magnification rostrocaudally aligned photomicrographsbrown) in the paratenial (PT), paraventricular (PV) and reuniens (RE) nuclei of the mmPFC, see Fig. 2a), and (2) retrogradely labeled cells (black) in RE produced by a Ff PHA-L labeled fibers and retrogradely labeled cells at three rostrocaudal levels of Rn PT/PV. Scale bar, 300 �m. FX, fornix, MT, mammillothalamic tract, PT, paratenieuniens of thalamus, V3, third ventricle. (For interpretation of the references to colo
Bulletin 71 (2007) 601–609 603
age 4.00 HR/HR-B CCD camera system (Advanced Microscopy Techniques,anvers, MA).
. Results
Using a combined anterograde–retrograde tracing procedure,e describe the distribution of PHA-L labeled fibers from thePFC onto populations of cells of nucleus reuniens that project
o the hippocampus. In accord with previous work [48,49], annjection of PHA-L into the ventral mPFC (Fig. 2a), essen-ially confined to the infralimbic and prelimbic cortices, gaveise to pronounced labeling of RE. Labeled fibers were not,owever, restricted to RE but were also present in other nuclei
f the midline thalamus including the paraventricular, parate-ial, mediodorsal, intermediodorsal, interanteromedial, centraledial and rhomboid nuclei of the thalamus. Fig. 1a depictsxtra-RE labeling antero-dorsally in the thalamus within the
through the thalamus showing: (1) the distribution of PHA-L labeled fibersidline thalamus produced by a PHA-L injection in the medial prefrontal cortexluorogold injection in the hippocampus (Fig. 2b). Note the significant overlapE, but no overlap in PT and PV due to the absence of retrogradely labeled cellsal nucleus of thalamus; PV, paraventricular nucleus of thalamus; RE, nucleusur in this figure legend, the reader is referred to the web version of the article.)
604 R.P. Vertes et al. / Brain Research Bulletin 71 (2007) 601–609
Fig. 2. (a) Low magnification photomicrograph of a PHA-L injection in the medial prefrontal cortex, virtually confined to the prelimbic cortex. (b) Low magnificationphotomicrograph of a FG injection in the ventral hippocampus, with the core of the injection (arrow) centered in stratum lacunosum-moleculare of ventral CA1.(c) Low magnification photomicrograph showing pronounced overlap in RE (dorsolateral to the third ventricle) of PHA-L labeled fibers (black) and FG labeledcells (brown). (d) High magnification photomicrograph showing putative synaptic contacts of a PHA-L labeled fiber (arrow) on a dendrite of a FG labeled cell. (eand f) High magnification photomicrographs showing putative synaptic contacts (arrows) of PHA-L labeled fibers on proximal dendrites FG labeled cells (f, highermagnification of d). Scale bars, a and b, 1200 �m; for c, 60 �m; for d, 25 �m; for e and f, 15 �m. AC, anterior cingulate cortex; ACC, nucleus accumbens; AGl,lateral agranular (frontal) cortex; CA1, CA1 field of Ammon’s horn of hippocampal formation; EC, entorhinal cortex; IL, infralimbic cortex; OC, occipital cortex;PL, prelimbic cortex; SUBv, ventral subiculum of hippocampal formation. (For interpretation of the references to colour in this figure legend, the reader is referredto the web version of the article.)

R.P. Vertes et al. / Brain Research Bulletin 71 (2007) 601–609 605
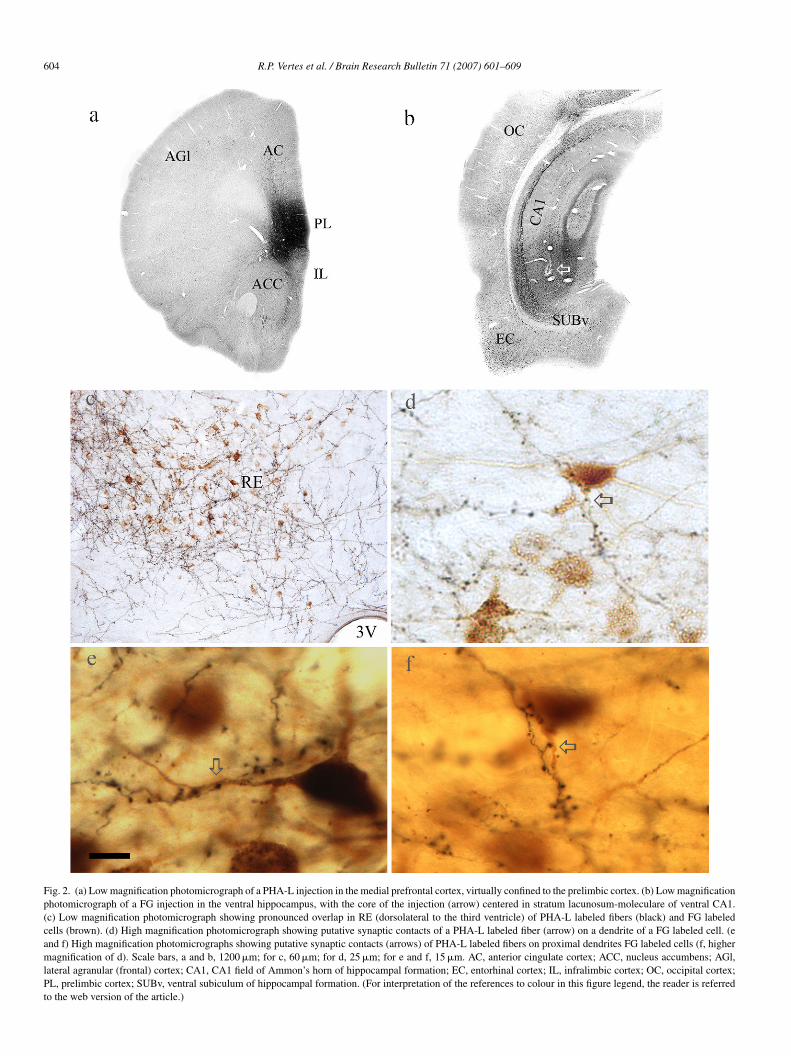
Fig. 3. (a–d) Series of consecutive electron micrographs within nucleus reuniens showing asymmetric contacts of a single PHA-L labeled (F) fiber (from the mPFC)on a labeled dendritic shaft, identified by the presence of numerous silver intensified gold deposits (arrows in D), of a RE cell retrogradely from the hippocampus.Note also the presence of asymmetric contacts of an unlabeled fiber (A) on the same labeled dendrite (D). Scale bar, 1 �m.
6 arch
pt
fifltmtlRsmsfiipr
a
dtin
ausmdtAmufo
FRpb
06 R.P. Vertes et al. / Brain Rese
aratenial nucleus and less so in the medially adjacent paraven-ricular nucleus.
Although RE was a main, but not the sole, target of labeledbers from mPFC, labeled cells were virtually restricted to REollowing retrograde FG injections in HF. As depicted for threeevels of RE (Fig. 1b–d), labeled neurons were present ros-rocaudally throughout RE following a HF injection (Fig. 2b),ost densely concentrated ipsilaterally (left side) within ros-
ral, ventral and lateral subdivisions of RE [38,44]. Very fewabeled neurons were observed within the thalamus outside ofE. Accordingly, RE was the only nucleus of the thalamus
howing a convergence of anterogradely labeled fibers (fromPFC) and retrogradely labeled neurons (from HF). As further
hown in Fig. 1, there was a significant overlap of PHA-L labeledbers (brown) and FG labeled cells (black), at rostral (Fig. 1b),
ntermediate (Fig. 1c) and caudal (Fig. 1d) levels of RE, most
ronounced rostrally—the site of the heaviest concentration ofetrogradely labeled neurons in RE.Fig. 2c shows a dense plexus of labeled fibers interspersedmong, and in close proximity to, labeled cells in RE. As
aac
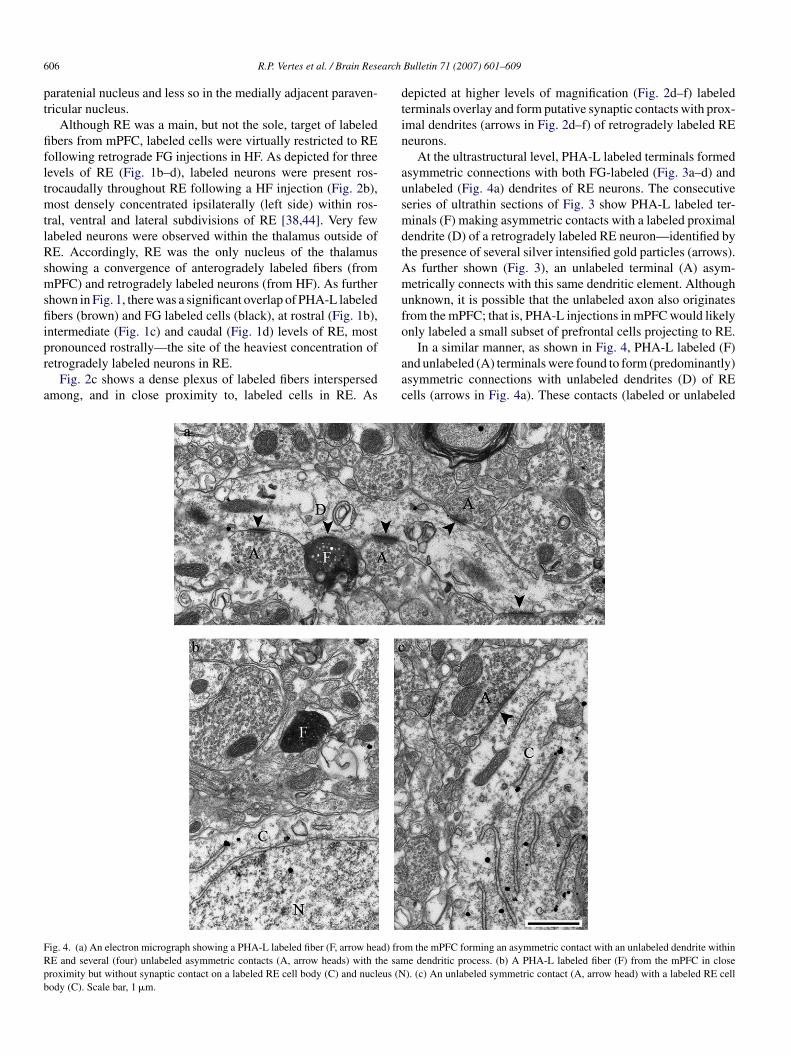
ig. 4. (a) An electron micrograph showing a PHA-L labeled fiber (F, arrow head) froE and several (four) unlabeled asymmetric contacts (A, arrow heads) with the samroximity but without synaptic contact on a labeled RE cell body (C) and nucleus (Nody (C). Scale bar, 1 �m.
Bulletin 71 (2007) 601–609
epicted at higher levels of magnification (Fig. 2d–f) labelederminals overlay and form putative synaptic contacts with prox-mal dendrites (arrows in Fig. 2d–f) of retrogradely labeled REeurons.
At the ultrastructural level, PHA-L labeled terminals formedsymmetric connections with both FG-labeled (Fig. 3a–d) andnlabeled (Fig. 4a) dendrites of RE neurons. The consecutiveeries of ultrathin sections of Fig. 3 show PHA-L labeled ter-inals (F) making asymmetric contacts with a labeled proximal
endrite (D) of a retrogradely labeled RE neuron—identified byhe presence of several silver intensified gold particles (arrows).s further shown (Fig. 3), an unlabeled terminal (A) asym-etrically connects with this same dendritic element. Although
nknown, it is possible that the unlabeled axon also originatesrom the mPFC; that is, PHA-L injections in mPFC would likelynly labeled a small subset of prefrontal cells projecting to RE.
In a similar manner, as shown in Fig. 4, PHA-L labeled (F)nd unlabeled (A) terminals were found to form (predominantly)symmetric connections with unlabeled dendrites (D) of REells (arrows in Fig. 4a). These contacts (labeled or unlabeled
m the mPFC forming an asymmetric contact with an unlabeled dendrite withine dendritic process. (b) A PHA-L labeled fiber (F) from the mPFC in close). (c) An unlabeled symmetric contact (A, arrow head) with a labeled RE cell

arch
fittrotiRtcRn
4
omhtpfissRsh
itdfF
nHld(uvdmdnr
uggHtttpwl
pgirR
4
mrmcio
stoiiFd
irttepmsamiItfl
tjR[rIiahobfl
R.P. Vertes et al. / Brain Rese
bers with unlabeled dendrites) does not negate the possibilityhat the unlabeled dendrites belong to cells that also project tohe hippocampus. Specifically, FG injections in HF (or thoseestricted to the ventral CA1/subiculum) would undoubtedlynly retrogradely fill a small percentage of RE cells projectingo HF. Fig. 4b shows a labeled terminal (F) in close proxim-ty, but without synaptic contact, on a labeled cell body (C) inE. No labeled terminals were found to make synaptic connec-
ions with cell bodies. Fig. 4c depicts a unlabeled (A) symmetricontact on a labeled cell body (C), indicating local or extra-E inhibitory connections with hippocampally projecting REeurons.
. Discussion
At the light microscopic level, we demonstrated a pronouncedverlap in nucleus reuniens of PHA-L labeled fibers from theedial prefrontal cortex and retrogradely labeled cells from the
ippocampus, as well as multiple putative contacts of labelederminals onto dendrites of RE cells projecting to the hip-ocampus. At the ultrastructural level, we showed that mPFCbers form asymmetric contacts predominantly with dendritichafts of hippocampally projecting RE cells. These findingsuggest that mPFC exerts excitatory actions on populations ofE cells that project to the hippocampus, and as such RE may
erve as an important relay between the ventral mPFC and theippocampus.
Although the inputs and outputs of RE have been describedn separate studies (see Ref. [51]) the present report is the firsto examine RE afferents/efferents in a single study, and by sooing demonstrated a strong convergence within RE of fibersrom the mPFC onto RE hippocampally projecting cells (seeig. 1).
Medial prefrontal fibers were shown to form asymmetric con-ections with proximal dendrites of RE neurons projecting toF. Unlabeled fibers also made asymmetric connections with
abeled dendrites, and in some instances the same (labeled) den-rites were contacted by labeled and unlabeled mPFC fibersFig. 3). Since even large PHA-L injections in mPFC wouldndoubtedly only label a small percentage of mPFC cells, it isery possible that many of the unlabeled profiles on labeled den-rites also represent fibers originating from mPFC. In a similaranner, labeled fibers forming connections with unlabeled den-
rites could represent contacts on hippocampally projecting REeurons since retrograde injections were restricted to a smallegion of the terminal RE field in HF [8,13,38,53,57].
RE fibers innervating the hippocampus appear to mainlytilize glutamate. Specifically, RE is densely populated bylutamatergic cells [26] and RE neurons were shown to be retro-radely labeled following [3H]d-aspartate injections in HF [8].alasy et al. [23] recently described a tight band of VGLUT2
erminals in stratum lacunosum-moleculare of CA1 that werehought to originate from RE. Taken together, this suggests
hat mPFC can indirectly exert excitatory actions on the hip-ocampus, mediated through an excitatory, glutamatergic, relayithin RE. Finally, RE selectively targets CA1 and the subicu-um but avoids CA3 and the dentate gyrus ([8,13,38,53,57],
C
Bulletin 71 (2007) 601–609 607
resent results), and the CA1/subiculum is the source of ori-in of efferents to the mPFC [12,18,28,29,31,43,47]. Thisndicates that projections between structures are directlyouted through CA1 (and subiculum): CA1 → mPFC →E → CA1.
.1. Functional considerations
As discussed, in the absence of direct projections from thePFC to the hippocampus, the nucleus reuniens appears to
epresent an important link from the mPFC to HF involved inemory processing. The mnemonic function commonly asso-
iated with the prefrontal cortex, and one extensively examined,s working memory—or the temporary storage and utilizationf information over short intervals [1,2,20,22].
As well documented for monkeys [22], the mPFC of rats, orpecifically the prelimbic cortex of mPFC [51], has been showno participate in tasks requiring the maintenance of informationver time—or working memory. The reversible or irreversiblenactivation of the ventral PFC (or PL) produces marked deficitsn delayed response tasks involving short or long delays [19,51].urther, cells of the ventral mPFC in rats show sustained activityuring the delay period of delay tasks [3,4,36].
Whereas information is retained for the short term in the PFC,t is widely acknowledged the long term storage of informationequires the hippocampus and associated structures of the medialemporal lobe [15–17,41,58]. Regarding the dual (and interac-ive) roles of the PFC and HF in memory processing, Bucknert al. [10,11] recently put forth the intriguing hypothesis that therefrontal cortex uses information in the short term (workingemory) to deal with impending task demands and its long term
torage may be simply a by product of its use in meeting immedi-te behavioral requirements. The conversion of short (workingemory) to long term stores might then involve a transfer of
nformation from mPFC to the hippocampus/parahippocampus.n the absence of direct mPFC inputs to the HF, indirect projec-ions from PFC to HF via RE may represent an important routeor the transfer of information from PFC to the medial temporalobe involved in memory processing.
Although speculative, it is possible that information relayedhrough RE (in contrast to direct cortical-hippocampal pro-ections) may be subject to modulating influences acting onE that affect that transfer. For instance, Van der Werf et al.
46] have proposed that the midline thalamus, including RE,egulates processes such as arousal, attention and awareness.n support of this, the midline thalamus receive pronouncednput from ‘arousal related’ sites of the brainstem [32,35,52]s well as from ‘wake-promoting’ orexin neurons of the lateralypothalamus [5,25]. It is possible that by modulating levelsf arousal/attention, RE serves to gate the flow of informationetween the mPFC and HF thereby controlling the type of pre-rontal information that gains access to the hippocampus for itsong term storage.
onflict of interest
No conflict of interest.

6 arch
A
MM
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[[
08 R.P. Vertes et al. / Brain Rese
cknowledgments
This research was supported by NIMH grants MH35883 andH63519 to RPV, and NIH and NIMH grants NS 042644 andH 060858 to CL.
eferences
[1] A. Baddeley, Working Memory, Clarendon Press, Oxford, 1986.[2] A. Baddeley, Recent developments in working memory, Curr. Opin. Neu-
robiol. 8 (1998) 234–238.[3] E.H. Baeg, Y.B. Kim, K. Huh, I. Mook-Jung, H.T. Kim, M.W. Jung, Dynam-
ics of population code for working memory in the prefrontal cortex, Neuron40 (2003) 177–188.
[4] A.S. Batuev, N.P. Kursina, A.P. Shutov, Unit activity of the medial wall ofthe frontal cortex during delayed performance in rats, Behav. Brain Res.41 (1990) 95–102.
[5] L. Bayer, E. Eggermann, B. Saint-Mleux, D. Machard, B.E. Jones, M.Muhlethaler, M. Serafin, Selective action of orexin (hypocretin) on non-specific thalamocortical projection neurons, J. Neurosci. 22 (2002) 7835–7839.
[6] R.M. Beckstead, Autoradiographic examination of corticocortical and sub-cortical projections of the mediodorsal-projection (prefrontal) cortex in therat, J. Comp. Neurol. 184 (1979) 43–62.
[7] E.H. Bertram, D.X. Zhang, Thalamic excitation of hippocampal CA1 neu-rons: a comparison with the effects of CA3 stimulation, Neuroscience 92(1999) 15–26.
[8] H. Bokor, A. Csaki, K. Kocsis, J. Kiss, Cellular architecture of the nucleusreuniens thalami and its putative aspartatergic/glutamatergic projection tothe hippocampus and medial septum in the rat, Eur. J. Neurosci. 16 (2002)1227–1239.
[9] S.L. Buchanan, R.H. Thompson, B.L. Maxwell, D.A. Powell, Efferent con-nections of the medial prefrontal cortex in the rabbit, Exp. Brain Res. 100(1994) 469–483.
10] R.L. Buckner, W.M. Kelley, S.E. Petersen, Frontal cortex contributes tohuman memory formation, Nat. Neurosci. 2 (1999) 311–314.
11] R.L. Buckner, J. Logan, D.I. Donaldson, M.E. Wheeler, Cognitive neuro-science of episodic memory encoding, Acta. Psychol. 105 (2000) 127–139.
12] D.B. Carr, S.R. Sesack, Hippocampal afferents to the rat prefrontal cortex:synaptic targets and relation to dopamine terminals, J. Comp. Neurol. 369(1996) 1–15.
13] M.J. Dolleman-Van der Weel, M.P. Witter, Projections from the nucleusreuniens thalami to the entorhinal cortex, hippocampal field CA1, and thesubiculum in the rat arise from different populations of neurons, J. Comp.Neurol. 364 (1996) 637–650.
14] M.J. Dolleman-Van der Weel, F.H. Lopes da Silva, M.P. Witter, Nucleusreuniens thalami modulates activity in hippocampal field CA1 throughexcitatory and inhibitory mechanisms, J. Neurosci. 17 (1997) 5640–5650.
15] Y. Dudai, The Neurobiology of Memory. Concepts, Findings, Trends,Oxford University Press, Oxford, 1989.
16] H. Eichenbaum, N.J. Cohen, From Conditioning to Conscious Recollec-tion: Memory Systems of the Brain, Oxford University Press, New York,2001.
17] H. Eichenbaum, G. Schoenbaum, B. Young, M. Bunsey, Functional orga-nization of the hippocampal memory system, Proc. Natl. Acad. Sci. 93(1996) 13500–13507.
18] F. Ferino, A.M. Thierry, J. Glowinski, Anatomical and electrophysiologicalevidence for a direct projection from Ammon’s horn to the medial prefrontalcortex in the rat, Exp. Brain Res. 65 (1987) 421–426.
19] S.B. Floresco, J.K. Seamans, A.G. Phillips, Selective roles for hippocam-pal, prefrontal cortical, and ventral stiatal circuits in radial arm maze tasks
with or without delay, J. Neurosci. 17 (1997) 1880–1890.20] J.M. Fuster, The prefrontal cortex—an update: time is of the essence,Neuron 30 (2001) 319–333.
21] P.S. Goldman-Rakic, The issue of memory in the study of prefrontal func-tion, in: A.M. Thierry, P.S. Goldman-Rakic, Y. Christen (Eds.), Motor
[
[
Bulletin 71 (2007) 601–609
and Cognitive Functions of the Prefrontal Cortex, Springer-Verlag, Berlin,1994, pp. 112–123.
22] P.S. Goldman-Rakic, Cellular basis of working memory, Neuron 14 (1995)477–485.
23] K. Halasy, T. Hajszan, E.G. Kovacs, T.T. Lam, C. Leranth, Distribution andorigin of vesicular glutamate transporter 2-immunoreactive fibers in the rathippocampus, Hippocampus 14 (2004) 908–918.
24] M. Herkenham, Connections of nucleus reuniens thalami: evidence fora direct thalamo-hippocampal pathway in the rat, J. Comp. Neurol. 177(1978) 589–610.
25] H. Huang, P. Ghosh, A.N. van den Pol, Prefrontal cortex-projectingglutamatergic thalamic paraventricular nucleus-excited by hypocretin: afeedforward circuit that may enhance cognitive arousal, J. Neurophysiol.95 (2006) 1656–1668.
26] E.E. Hur, L. Zaborszky, Vglut2 afferents to the medial prefrontal andprimary somatosensory cortices: a combined retrograde tracing in situhybridization study, J. Comp. Neurol. 483 (2005) 351–373.
27] K.M. Hurley, H. Herbert, M.M. Moga, C.B. Saper, Efferent projectionsof the infralimbic cortex of the rat, J. Comp. Neurol. 308 (1991) 249–276.
28] A. Ishikawa, S. Nakamura, Convergence and interaction of hippocam-pal and amygdalar projections within the prefrontal cortex in the rat, J.Neurosci. 23 (2003) 9987–9995.
29] T.M. Jay, M.P. Witter, Distribution of hippocampal CA1 and subicularefferents in the prefrontal cortex of the rat studied by means of antero-grade transport of Phaseolus vulgaris-leucoagglutinin, J. Comp. Neurol.313 (1991) 574–586.
30] T.M. Jay, F. Burette, S. Laroche, NMDA receptor dependent long termpotentiation in the hippocampal afferent fibre system to the prefrontal cortexin the rat, Eur. J. Neurosci. 7 (1995) 247–250.
31] T.M. Jay, J. Glowinski, A.M. Thierry, Selectivity of the hippocampal pro-jection to the prelimbic area of the prefrontal cortex in the rat, Brain Res.505 (1989) 337–340.
32] K.E. Krout, R.E. Belzer, A.D. Loewy, Brainstem projections to midlineand intralaminar thalamic nuclei of the rat, J. Comp. Neurol. 448 (2002)53–101.
33] S. Laroche, S. Davis, T.M. Jay, Plasticity at hippocampal to prefrontalcortex synapses: dual roles in working memory and consolidation, Hip-pocampus 10 (2000) 438–446.
34] S. Laroche, T.M. Jay, A.M. Thierry, Long-term potentiation in the prefrontalcortex following stimulation of the hippocampal CA1/subicular region,Neurosci. Lett. 114 (1990) 184–190.
35] J.T. McKenna, R.P. Vertes, Afferent projections to nucleus reuniens of thethalamus, J. Comp. Neurol. 480 (2004) 115–142.
36] A.A. Orlov, N.P. Kurzina, A.P. Shutov, Activity of medial wall neuronsin frontal cortex of rat brain during delayed response reactions, Neurosci.Behav. Physiol. 18 (1988) 31–37.
37] R.L. Reep, J.V. Corwin, A. Hashimoto, R.T. Watson, Efferent connectionsof the rostral portion of medial agranular cortex in rats, Brain Res. Bull. 19(1987) 203–221.
38] P.Y. Risold, R.H. Thompson, L.W. Swanson, The structural organizationof connections between hypothalamus and cerebral cortex, Brain Res. Rev.24 (1997) 197–254.
39] P. Room, F.T. Russchen, H.J. Groenewegen, A.H.M. Lohman, Efferentconnections of the prelimbic (area 32) and the infralimbic (area 25) cortices:an anterograde tracing study in the cat, J. Comp. Neurol. 242 (1985) 40–55.
40] S.R. Sesack, A.Y. Deutch, R.H. Roth, B.S. Bunney, Topographical organi-zation of the efferent projections of the medial prefrontal cortex in the rat:an anterograde tract-tracing study with Phaseolus vulgaris leucoagglutinin,J. Comp. Neurol. 290 (1989) 213–242.
41] L.R. Squire, Memory and Brain, Oxford University Press, New York, 1987.42] H.S. Su, M. Bentivoglio, Thalamic midline cell populations projecting to
the nucleus accumbens, amygdala, and hippocampus in the rat, J. Comp.
Neurol. 297 (1990) 582–593.43] L.W. Swanson, A direct projection from Ammon’s horn to prefrontal cortexin the rat, Brain Res. 217 (1981) 150–154.
44] L.W. Swanson, Brain Maps: Structure of the Rat Brain, Elsevier, New York,1998.

arch
[
[
[
[
[
[
[
[
[
[
[
[
[
R.P. Vertes et al. / Brain Rese
45] M. Takagishi, T. Chiba, Efferent projections of the infralimbic (area 25)region of the medial prefrontal cortex in the rat: an anterograde tracerPHA-L study, Brain Res. 566 (1991) 26–39.
46] Y.D. Van der Werf, M.P. Witter, H.J. Groenewegen, The intralaminar andmidline nuclei of the thalamus. Anatomical and functional evidence forparticipation in processes of arousal and awareness, Brain Res. Rev. 39(2002) 107–140.
47] T. van Groen, J.M. Wyss, The connections of presubiculum and parasubicu-lum in the rat, Brain Res. 518 (1990) 227–243.
48] R.P. Vertes, Analysis of projections from the medial prefrontal cortex to thethalamus in the rat, with emphasis on nucleus reuniens, J. Comp. Neurol.442 (2002) 163–187.
49] R.P. Vertes, Differential projections of the infralimbic and prelimbic cortexin the rat, Synapse 51 (2004) 32–58.
50] R.P. Vertes, Hippocampal theta rhythm: a tag for short term memory, Hip-pocampus 15 (2005) 923–935.
51] R.P. Vertes, Interactions among the medial prefrontal cortex, hippocampusand midline thalamus in emotional and cognitive processing in the rat,Neuroscience 142 (2006) 1–20.
52] R.P. Vertes, G.F. Martin, Autoradiographic analysis of ascending projec-tions from the pontine and mesencephalic reticular formation and the
[
Bulletin 71 (2007) 601–609 609
median raphe nucleus in the rat, J. Comp. Neurol. 275 (1988) 511–541.
53] R.P. Vertes, W.B. Hoover, A.C. do Valle, A. Sherman, J.J. Rodriguez, Effer-ent projections of reuniens and rhomboid nuclei of the thalamus in the rat,J. Comp. Neurol. 499 (2006) 768–796.
54] R.P. Vertes, W.B. Hoover, G. Viana Di Prisco, Theta rhythm of the hip-pocampus: subcortical control and functional significance, Behav. Cogn.Neurosci. Rev. 3 (2004) 173–2000.
55] M.P. Witter, H.J. Groenewegen, F.H. Lopes da Silva, A.H. Lohman,Functional organization of the extrinsic and intrinsic circuitry of theparahippocampal region, Prog. Neurobiol. 33 (1989) 161–253.
56] F.G. Wouterlood, Innervation of entorhinal principal cells by neurons ofthe nucleus reuniens thalami. Anterograde PHA-L tracing combined withretrograde fluorescent tracing and intracellular injection with lucifer yellowin the rat, Eur. J. Neurosci. 3 (1991) 641–647.
57] F.G. Wouterlood, E. Saldana, M.P. Witter, Projection from the nucleus
reuniens thalami to the hippocampal region: light and electron microscopictracing study in the rat with the anterograde tracer Phaseolus vulgaris-leucoagglutinin, J. Comp. Neurol. 296 (1990) 179–203.58] S. Zola-Morgan, L.R. Squire, Neuroanatomy of memory, Annu. Rev. Neu-rosci. 16 (1993) 547–563.

![(12) United States Patent US 8,487,246 B2 Vertes et a ...vertes.columbian.gwu.edu/publicat_html/Vertes 2013 US 8,487,246 B… · (12) United States Patent Vertes et a]. US008487246B2](https://img.pdfslide.us/doc/110x75/5f0788457e708231d41d7319/12-united-states-patent-us-8487246-b2-vertes-et-a-2013-us-8487246-b.jpg)