Embed Size (px)
Citation preview

294 Journal of Lipid Research Volume 56, 2015
Copyright © 2015 by the American Society for Biochemistry and Molecular Biology, Inc.
This article is available online at http://www.jlr.org
effi cient therapeutic strategies for preventing and treating metabolic disorders associated with obesity.
Adipogenesis is controlled by a balance of internal and external factors that either stimulate or repress adipogenic differentiation. In the early phase of adipogenic differ-entiation, CCAAT/enhancer binding protein (C/EBP) � and C/EBP � induce expression of C/EBP � and PPAR � , which are the principal adipogenic transcription factors that control the early differentiation of preadipocytes into lipid-accumulating fat cells ( 5, 6 ). In addition, PPAR � appears to suppress canonical Wingless/int-1 class (Wnt) signaling by accelerating proteasome-dependent degrada-tion of � -catenin. Conversely, � -catenin, a transcriptional co-activator in the Wnt signaling pathway, blocks adipogenesis by repressing PPAR � and C/EBP � ( 7, 8 ). A number of re-ports have suggested a relationship between Wnt signaling and diabetes and adipogenesis ( 9, 10 ). Adipose tissue-specifi c expression of Wnt10b reduces adiposity and improves insulin sensitivity in the ob/ob obesity model ( 11 ). Although the extensive downregulation of � -catenin expression and its antadipogenic effects during adipogenic differentiation have been characterized, less is known about the factors that modulate � -catenin activity during adipogenesis.
Wnt signaling is initiated upon binding of Wnt ligands to transmembrane Frizzled receptors. In the canonical Wnt signaling pathway, Frizzled receptors transduce sig-nals through Dishevelled (Dvl) to inhibit glycogen synthase kinase 3 (GSK3 � ), resulting in hypophosphorylation and subsequent stabilization of � -catenin. Following nuclear translocation, active � -catenin binds to and coactivates members of the T-cell factor/lymphoid-enhancer factor
Abstract Nucleoredoxin (NRX) is a member of the thiore-doxin family of proteins that controls redox homeostasis in cell. Redox homeostasis is a well-known regulator of cell differentiation into various tissue types. We found that NRX expression levels were higher in white adipose tissue of obese ob/ob mice and increased in the early adipogenic stage of 3T3-L1 preadipocyte differentiation. Knockdown of NRX decreased differentiation of 3T3-L1 cells, whereas overexpression increased differentiation. Adipose tissue-specifi c NRX transgenic mice showed increases in adipocyte size as well as number compared with WT mice. We further confi rmed that the Wingless/int-1 class (Wnt) / � -catenin pathway was also involved in NRX-promoted adipogenesis, consistent with a previous report showing NRX regulation of this pathway. Genes involved in lipid metabolism were downregulated, whereas inflammatory genes, including those encoding macrophage markers, were signifi cantly up-regulated, likely contributing to the obesity in Adipo-NRX mice. Our results therefore suggest that NRX acts as a novel proadipogenic factor and controls obesity in vivo. —Bahn, Y. J., K-P. Lee, S-M. Lee, J. Y. Choi, Y-S. Seo, and K-S. Kwon. Nucleoredoxin promotes adipogenic differentiation through regulation of Wnt/ � -catenin signaling. J. Lipid Res. 2015. 56: 294–303.
Supplementary key words Wingless/int-1 class (Wnt) signaling • adi-pogenesis • obesity • infl ammation
Adipose tissues play important roles in the regulation of whole-body energy homeostasis ( 1 ). Excess energy intake in-creases the number and size of fat cells ( 2 ). These obese adi-pose tissues facilitate chronic low-grade infl ammation and insulin resistance, which result in severe obesity and diabetes ( 3, 4 ). Thus, understanding adipocyte development and adi-pogenesis could lay the groundwork for the development of
This work was supported by Bio & Medical Technology Development Program Grants 20110030133 and 2013M3A9B6076413 (K-S.K.) of the National Research Foundation funded by the Korea government (MSIP), and the KRIBB Research Initiative Program.
Manuscript received 18 August 2014 and in revised form 26 December 2014.
Published, JLR Papers in Press, December 29, 2014 DOI 10.1194/jlr.M054056
Nucleoredoxin promotes adipogenic differentiation through regulation of Wnt/ � -catenin signaling
Young Jae Bahn , * ,† Kwang-Pyo Lee , † Seung-Min Lee , † Jeong Yi Choi , † Yeon-Soo Seo , * and Ki-Sun Kwon 1,†
Department of Biological Science,* Korea Advanced Institute Science and Technology (KAIST) , Daejeon 305-701, Republic of Korea; and Aging Research Institute, † Korea Research Institute of Bioscience and Biotechnology (KRIBB) , Daejeon 305-806, Republic of Korea
Abbreviations: Adipo-NRX mice, adipose tissue-specific NRX transgenic mice; C/EBP, CCAAT/enhancer binding protein; Dvl, Di-shevelled; Fabp4, fatty acid binding protein 4; GTT, glucose tolerance test; ITT, insulin tolerance test; LSL-NRX mice, loxP-stop-loxP-NRX transgenic mice; NRX, nucleoredoxin; TRX, thioredoxin; WAT, white adipose tissue; Wnt, Wingless/int-1 class.
1 To whom correspondence should be addressed. e-mail: [email protected]
The online version of this article (available at http://www.jlr.org) contains supplementary data in the form of three fi gures, one table, and Methods.
by guest, on August 29, 2018
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2014/12/29/jlr.M054056.DC1Supplemental Material can be found at:

Nucleoredoxin regulates adipogenesis 295
BSA and 1.5 mg/ml type I collagenase (Worthington, Lakewood, NJ). The digested tissue was fi ltered through a 300 � m nylon mesh to remove undigested tissue and centrifuged at 500 g for 5 min. The fl oating adipocyte fraction was removed, washed with buffer, and recentrifuged to isolate free adipocytes. The stromal-vascular pellet was resuspended in erythrocyte lysis buffer (154 mM NH 4 Cl, 10 mM KHCO 3 , and 1 mM EDTA), filtered through a 45 � m nylon mesh to remove endothelial cells, and pelleted at 500 g for 5 min. Enriched cells were cultured in a growth me-dium composed of DMEM, 20% fetal bovine serum, 100 U/ml penicillin, and 100 mg/ml streptomycin (Gibco-Invitrogen, Carlsbad, CA) at 37°C in a humidifi ed 5% CO 2 atmosphere.
Cell culture and adipogenic differentiation The preadipocytes cell line 3T3-L1, derived from mouse em-
bryo fi broblasts, was grown at 37°C in DMEM containing 10% heat-inactivated bovine calf serum (Gibco-Invitrogen), 100 U/ml penicillin, and 100 mg/ml streptomycin in a humidifi ed 5% CO 2 atmosphere. 3T3-L1 cells were induced to differentiate into ma-ture adipocytes according to the procedure of Student et al. ( 22 ), with minor modifi cations. Briefl y, 2 days after reaching confl u-ence (day 0), cells were placed in differentiation medium composed of DMEM, 10% fetal bovine serum, and MDI, a differentiation cocktail consisting of 0.5 mM 3-isobutyl-1-methyl-xanthine (IBMX), 1 � M dexamethasone, and 10 � g/ml insulin (Sigma, St. Louis, MO). The medium was replenished every other day.
Generation of stable 3T3-L1 cell lines 3T3-L1 cells stably expressing FLAG -tagged NRX were gener-
ated using a lentivirus-mediated infection system. FLAG is a short, hydrophilic fusion tag consisting of eight amino acids (Asp-Tyr-Lys-Asp-Asp-Asp-Asp-Lys). For expression of NRX, cDNA encod-ing FLAG-tagged NRX was cloned into the multicloning site of the green fl uorescent protein (GFP)-tagged pLentiM1.4 vector. Lentiviruses were subsequently produced by transiently cotrans-fecting HEK293T cells with pLP1, pLP2, and pVSV-G plasmid (Invitrogen, Carlsbad, CA) using Lipofectamine (Invitrogen). Forty-eight hours after transfection, supernatants containing len-tiviral particles were collected and used to infect 3T3-L1 cells in the presence of 4 µg/ml polybrene. Infected cells were selected by incubation with 2 µg/ml puromycin for 2–3 weeks and used in experiments as indicated. NRX cDNA was kindly provided by Dr. Tasuku Honjo (Kyoto University, Japan) ( 15 ). An NRX mutant in which catalytic Cys205 and Cys208 residues were replaced with Ser (CS-NRX), provided by Dr. Sung-Kyu Ju, was constructed us-ing site-directed mutagenesis.
RNA interference NRX knockdown in 3T3-L1 cells was accomplished using
shRNA against mouse NRX in pLKO.1-puro lentiviral vectors obtained from Sigma (clone ID NM_022463.3-1358s1c1 and NM_022463.3-452s1c1), according to the manufacturer’s proto-col. Briefly, shRNA lentiviral particles were generated in HEK293T cells by transient transfection with pLP1, pLP2, pVSV-G, and shRNA lentiviral vector or pLKO.1-scrambled (control) vector (SHC002V; Sigma) using Lipofectamine. Forty-eight hours after transfection, supernatants containing lentiviral parti-cles were collected and used to infect 3T3-L1 cells in the pres-ence of 4 µg/ml polybrene. Infected cells were selected by incubation with 2 µg/ml puromycin for 2–3 weeks and used in experiments as indicated. An shRNA-resistant NRX incapable of shNRX binding and subsequent NRX degradation was con-structed by site-directed mutagenesis of the target region of shNRX. The construct was obtained by standard methods using
family of transcription factors, leading to activation of tar-get genes ( 12, 13 ).
Nucleoredoxin (NRX) is member of the thioredoxin (TRX) family of proteins. TRX family proteins commonly possess a pair of redox-active, oxidation-sensitive cysteine residues in the catalytic center that are directly involved in the reduction of disulfi de bonds in target proteins ( 14 ). Although NRX activity has been demonstrated in in vitro assays, whether the TRX-related oxidoreductase activity of NRX plays a role in vivo is unknown ( 15 ). Previous studies have shown that NRX is a multifunction protein that regu-lates target proteins through its direct binding activity rather than its oxidoreductase activity ( 16, 17 ). Endoge-nous NRX protein is predominantly localized in the cyto-sol of cells, and its transcripts are widely expressed in all adult tissues ( 18 ). Knockout of the NRX gene in mouse embryos is perinatally lethal; NRX -knockout embryos (day 18.5) are smaller than their WT littermates and exhibit craniofacial defects with short frontal regions ( 19 ). Inter-estingly, a genomic region around the mouse NRX gene is involved in type 1 and type 2 diabetes ( 20 ). However, a role for NRX in adipogenesis and obesity has not been reported.
Here, we investigated the role of NRX in preadipocyte differentiation and the obesity phenotype using a 3T3-L1 preadipocyte differentiation system and adipose tissue-specifi c transgenic mice. We show that NRX mediates adi-pogenesis by modulating � -catenin activity in vitro and in vivo. Our fi ndings suggest that NRX might act as a proad-ipogenic factor that is involved in adipocyte differentia-tion and aspects of the obesity phenotype.
MATERIALS AND METHODS
Generation of adipose tissue-specifi c NRX transgenic mice
The loxP-stop-loxP-NRX transgenic mice (LSL-NRX mice) were obtained by microinjection and germ-line transmission of the transgenic construct (supplementary Fig. 1A). The LSL-NRX mouse strain was backcrossed with the C57BL/6 strain for more than eight generations to create a uniform genetic background. Adipose tissue-specifi c NRX transgenic mice (Adipo-NRX mice) were produced by crossing LSL-NRX mice with adiponectin-Cre transgenic mice, generating mice with adipose tissue-specific overexpression of hNRX. All animal experiments were per-formed according to protocols approved by the Animal Care and Use Committee of the Korea Research Institute of Bioscience and Biotechnology (KRIBB).
Potential founder mice were genotyped by PCR analysis using genomic DNA isolated from mouse tail clips. Primers used for the detection of the transgenic product were 5 ′ -TGC GAG ATT ACA CCA ACC TG-3 ′ and 5 ′ -CCC ATA TGT CCT TCC GAG TG-3 ′ .
Isolation and culturing of primary adipocytes Primary adipocytes were isolated from epididymal fat pads us-
ing the collagenase method, as described previously ( 21 ). Briefl y, freshly excised fat pads were minced and digested for 45 min to 1 h at 37°C in Krebs-Ringer bicarbonate (pH 7.4) containing 4%
by guest, on August 29, 2018
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2014/12/29/jlr.M054056.DC1Supplemental Material can be found at:

296 Journal of Lipid Research Volume 56, 2015
and photomicrographed. Oil Red O staining was then quantifi ed as described previously ( 23 ). Incorporated Oil Red O dye was extracted by adding absolute isopropanol to the stained cell-culture dish and shaking the dish for 30 min. Triplicate samples were read at 510 nm using an Ultrospec2000 Spectrophotometer (Pharmacia Biotech, Piscataway, NJ).
Hematoxylin and eosin staining Adipose tissue, muscle, and liver samples were fi xed for 12–16 h
at room temperature in 10% formalin (Sigma) and then em-bedded in paraffi n. Five-micron sections cut at 50 � m intervals were mounted on charged glass slides, deparaffi nized in xylene, and stained with hematoxylin and eosin (H and E). Adipocyte cross-sectional area was quantifi ed for each adipocyte in each fi eld using National Institutes of Health Image J software (http://rsb.info.nih.gov/ij/). To quantify cell number, adipocytes were isolated from 100 mg of adipose tissue by using 2 mg/ml collage-nase type II-S (Sigma) digestion buffer and were counted using a cell counter (Logos Biosystems, Annandale, VA) ( 24 ).
Glucose and insulin tolerance test Glucose tolerance test (GTT) was performed with 8 h fasted
animals. After determination of fasted blood glucose levels, each animal was injected intraperitoneally with 20% glucose (1 g/kg). The insulin tolerance test (ITT) was performed, with an initial fasting for 4 h, and subsequent intraperitoneal injection of insu-lin (1 U/kg). In all tests, tail blood glucose levels were measured with a glucometer (Roche Diagnostics, Mannheim, Germany) at the indicated times after injection.
Immunohistochemistry Formalin-fi xed, paraffi n-embedded sections of 5 µm were
mounted on charged glass slides, deparaffi nized in xylene, stained with hematoxylin, and processed for immunohistochemi-cal detection of F4/80 according to standard immunoperoxidase procedure using VECTASTAIN Elite ABC kit (Vector Labs, Burl-ingame, CA) and anti-F4/80 antibody (Abcam, Cambridge, UK).
Statistical analysis Quantitative data are presented as means ± SD unless indi-
cated otherwise. Differences between means were evaluated us-ing Student’s unpaired t -test. A P -value < 0.05 was considered statistically signifi cant.
RESULTS
NRX levels increase in the early stage of 3T3-L1 preadipocyte differentiation
We analyzed the expression of NRX during 3T3-L1 cells differentiation induced by a differentiation cocktail composed of IBMX, dexamethasone, and insulin. NRX transiently increased in the early stages of adipocyte differentiation at both mRNA and protein levels. Quanti-tative RT-PCR revealed that NRX mRNA reached a maxi-mum 1 day after differentiation induction ( Fig. 1A ). Protein levels of NRX were proportional to mRNA levels ( Fig. 1B ). We next determined whether hyperglycemic conditions affected NRX levels in ob/ob mice. Interestingly, NRX protein was upregulated in WAT of ob/ob mice com-pared with that in WT mice ( Fig. 1C ). These results sug-gest that increased expression of NRX may be associated with adipogenesis and, ultimately, obesity.
the primers 5 ′ -CTT TTG TGA ATG ACT TC T T G G C T G AAA AAC TC-3 ′ and 5 ′ -TCA GGC TTG AGT TTT TC A GC C A A G AAG TCA TT-3 ′ . Underlined regions indicate the sites that were mu-tated without changing amino acid residues.
RNA extraction and real-time PCR analysis Total RNA was extracted using the Easy-Blue reagent (iN-
tRON Biotechnology, Korea), according to the manufactur-er’s instructions, and treated with RNase-free DNase I (Takara, Shiga, Japan) to remove contaminating genomic DNA. cDNA was then synthesized from total RNA by RT using a DiaStar RT kit (Solgent, Korea). Quantitative RT-PCR analysis was per-formed using the Step One Plus real-time PCR system (Ap-plied Biosystems, Waltham, MA) with the corresponding primers.
Immunoblotting and immunoprecipitation Cells were lysed in a lysis buffer containing 20 mM HEPES (pH
7.2), 150 mM NaCl, 0.5% Triton X-100, 0.1 mM Na 3 VO 4 , 1 mM NaF, 1 mM 4-(2-aminoethyl)-benzenesulfonyl fl uoride hydro-chloride, and 5 mg/ml aprotinin (Sigma). Soluble proteins in cell lysates were separated by SDS-PAGE and analyzed by immu-noblotting using anti-NRX (R&D Biosystems, Minneapolis, MN, AF5719), anti-PPAR � (Cell Signaling Technology, Danvers, MA, #2430), anti- � -actin (Santa Cruz Biotechnology, Santa Cruz, CA, SC-47778), anti-Dvl-1 (Santa Cruz Biotechnology, SC-8025), anti-lamin A/C (Santa Cruz Biotechnology, SC-6215), anti-cyclin D1 (Cell Signaling Technology, 2922), anti- � -catenin (Cell Signaling Technology, 9562), anti- � -tubulin (Millipore, 05-829), anti-v-akt murine thymoma viral oncogene homology (AKT; Santa Cruz Biotechnology, SC-1618), anti-phospho-AKT (Cell Signaling Technology, 9271L), and anti-fatty acid binding protein 4 (FABP4; Cell Signaling Technology, 2120) antibodies. White adi-pose tissue (WAT) from ob/ob mice was kindly provided by Dr. Chul-Ho Lee (KRIBB). For immunoprecipitation, lysates were incubated with anti-FLAG agarose (Sigma) or anti-NRX antibody at 4°C for several hours, after which the beads were washed three times with cell lysis buffer. The beads were resuspended in 1× SDS-PAGE sample buffer, and the eluted proteins were resolved by SDS-PAGE.
In vitro gene transfer For Amaxa nucleofections, pellets containing 0.5–1.5 × 10 6
3T3-L1 cells were carefully resuspended in 100 µl of Nucleofec-tor solution (Lonza, Allendale, NJ), mixed with 1–2 µg of plas-mids, and subjected to nucleofection using T-30 Amaxa protocols. The cells were then gently transferred into a 6-well plate and cul-tured until analysis.
Luciferase assays and reagents Luciferase activity was assessed using the Luciferase Assay sys-
tem (Promega, Madison, WI), according to the manufacturer’s instructions. 3T3-L1 cells were transduced with TOPfl ash and FOPfl ash exogenous reporter constructs together with pCMV- � -galactosidase. Luciferase activity was normalized to � -galactosidase activity to adjust for transfection effi ciency.
Oil Red O staining Cultured cells were washed twice with PBS and fi xed by incu-
bating with 3.7% paraformaldehyde for 15 min at room tempera-ture. The cells were then washed with distilled water and stained for 30 min with 0.3% fi ltered Oil Red O solution in 60% isopro-panol. The stained cells were washed twice with distilled water
by guest, on August 29, 2018
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2014/12/29/jlr.M054056.DC1Supplemental Material can be found at:

Nucleoredoxin regulates adipogenesis 297
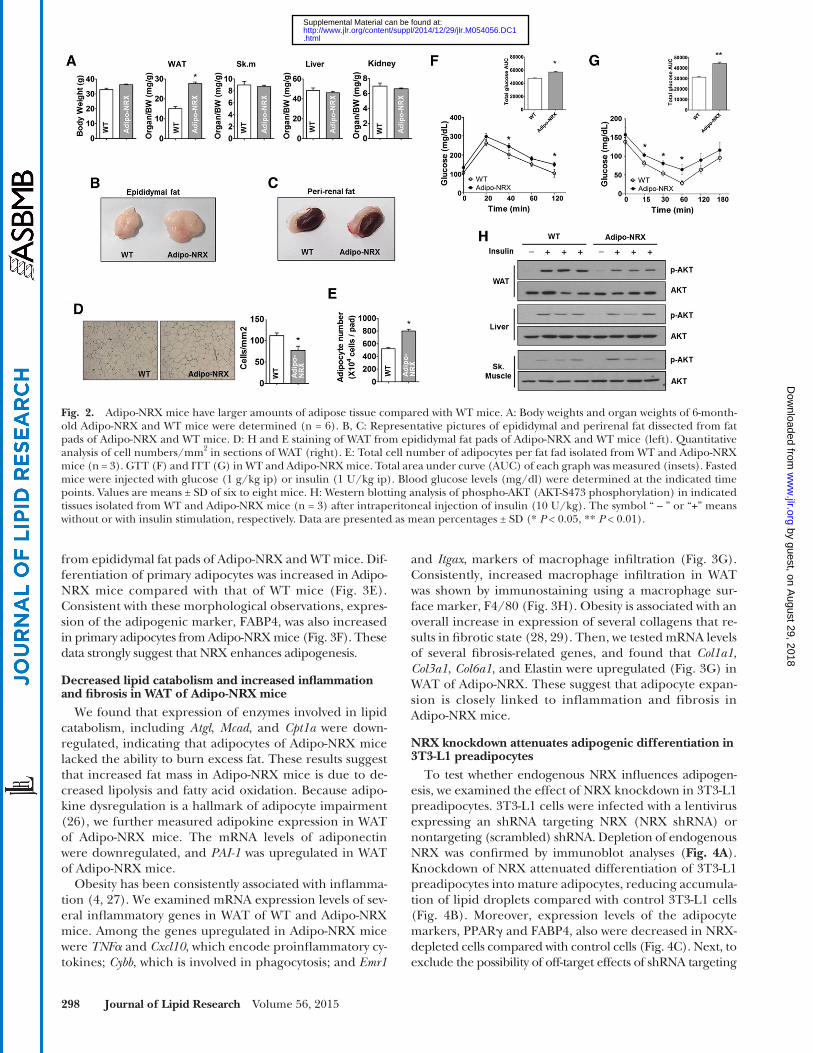
adipocyte size, suggesting that hypertrophy was accompa-nied by hyperplasia in adipose tissue of Adipo-NRX mice.
The plasma analysis revealed that blood glucose and in-sulin levels were higher in Adipo-NRX mice than in WT mice (supplementary Table 1). We further assessed glu-cose homeostasis in WT and Adipo-NRX mice via a GTT and ITT. Adipo-NRX mice showed the tendency to glu-cose intolerance compared with WT mice ( Fig. 2F ) and exhibited impaired insulin tolerance that was associated with reduced insulin sensitivity in WAT but not in liver and skeletal muscle ( Fig. 2G, H ). Although the adipose tissue appeared lipid accumulation, there was no signifi cant change in lipid deposition in liver and skeletal muscle (supplementary Fig. 2). These data demonstrate that adipocyte-specific NRX overexpression increases lipid accumulation in WAT, resulting in insulin resistance.
NRX overexpression leads to increased adipogenic differentiation
To evaluate whether overexpression of NRX facilitates adipogenic differentiation in cell culture system, we in-fected 3T3-L1 preadipocytes with a lentivirus expressing GFP-tagged mouse NRX or a control lentivirus. Stable overexpression of NRX was monitored by immunoblotting ( Fig. 3A ) and fl uorescence imaging. 3T3-L1 preadipocytes stably overexpressing NRX showed increased accumula-tion of lipid droplets following induction of adipocyte differentiation compared with control cells ( Fig. 3B, C ). We confi rmed that the adipogenic markers, PPAR � and FABP4, were also upregulated in NRX-overexpressing cells compared with control cells ( Fig. 3D ). To confi rm that NRX overexpression positively regulates adipocyte dif-ferentiation, we next isolated and cultured primary adipocytes
NRX overexpression leads to adipocyte hypertrophy combined with hyperplasia in mouse adipose tissue
To further investigate the function of NRX in adipogene-sis in vivo, we generated LSL-NRX mice, then crossed these mice with adiponectin-Cre mice to generate Adipo-NRX mice. To validate the specifi c overexpression of NRX in adi-pose tissues, we analyzed NRX protein levels in several tis-sues isolated from Adipo-NRX and WT mice. As expected, overexpression of NRX was observed only in adipose tissues, including white and brown adipose tissue (supplementary Fig. 1B). To clarify activity of the adiponectin promoter, we checked expression levels of adiponectin during adipogen-esis. We confi rmed that the expression level of adiponectin was very low before induction of differentiation but gradu-ally increased after induction of differentiation (supple-mentary Fig. 1C), which was consistent with a previous report ( 25 ). We also observed that expression of adiponec-tin promoter-driven NRX was induced at early differentia-tion stage in primary adipocytes isolated from Adipo-NRX mice (supplementary Fig. 1D).
Although there was a slight increase in body weight in Adipo-NRX mice compared with WT littermate controls, a comparison of organ weights revealed a notable increase in epididymal WAT in Adipo-NRX mice ( Fig. 2A ). Epidid-ymal and perirenal fat masses were also signifi cantly larger in Adipo-NRX mice than in WT mice ( Fig. 2B, C ). There was no difference in food intake between Adipo-NRX mice and WT littermate controls. An analysis of adipocyte cross-sectional areas showed that epididymal fat of Adipo-NRX mice contained hypertrophied adipocytes ( Fig. 2D ). A quantitative analysis revealed that adipose tissue expan-sion of Adipo-NRX mice was caused by an increase of adi-pocyte numbers ( Fig. 2E ) as well as an enlargement of
Fig. 1. NRX expression during adipogenic differentiation of 3T3-L1 preadipocytes and in WAT of obese mice. A, B: 3T3-L1 cells were induced to differentiate into adipocytes by treatment with MDI medium. A: Expression of NRX was analyzed using real-time quantitative RT-PCR. Total RNA was extracted at the indicated days of differentiation. � -Actin was used as an internal control. B: NRX expression was analyzed by Western blotting at the indicated days of differentiation. PPAR � and FABP4 were used as adipocyte differentiation markers. � -Actin was used as a loading control. C: Western blot analysis of NRX expression in visceral WAT of WT and ob/ob mice. Data are presented as mean fold-changes ± SD (* P < 0.05, ** P < 0.01).
by guest, on August 29, 2018
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2014/12/29/jlr.M054056.DC1Supplemental Material can be found at:
298 Journal of Lipid Research Volume 56, 2015
and Itgax , markers of macrophage infi ltration ( Fig. 3G ). Consistently, increased macrophage infi ltration in WAT was shown by immunostaining using a macrophage sur-face marker, F4/80 ( Fig. 3H ). Obesity is associated with an overall increase in expression of several collagens that re-sults in fi brotic state ( 28, 29 ). Then, we tested mRNA levels of several fi brosis-related genes, and found that Col1a1 , Col3a1 , Col6a1 , and Elastin were upregulated ( Fig. 3G ) in WAT of Adipo-NRX. These suggest that adipocyte expan-sion is closely linked to inflammation and fibrosis in Adipo-NRX mice.
NRX knockdown attenuates adipogenic differentiation in 3T3-L1 preadipocytes
To test whether endogenous NRX infl uences adipogen-esis, we examined the effect of NRX knockdown in 3T3-L1 preadipocytes. 3T3-L1 cells were infected with a lentivirus expressing an shRNA targeting NRX (NRX shRNA) or nontargeting (scrambled) shRNA. Depletion of endogenous NRX was confi rmed by immunoblot analyses ( Fig. 4A ). Knockdown of NRX attenuated differentiation of 3T3-L1 preadipocytes into mature adipocytes, reducing accumula-tion of lipid droplets compared with control 3T3-L1 cells ( Fig. 4B ). Moreover, expression levels of the adipocyte markers, PPAR � and FABP4, also were decreased in NRX-depleted cells compared with control cells ( Fig. 4C ). Next, to exclude the possibility of off-target effects of shRNA targeting
from epididymal fat pads of Adipo-NRX and WT mice. Dif-ferentiation of primary adipocytes was increased in Adipo-NRX mice compared with that of WT mice ( Fig. 3E ). Consistent with these morphological observations, expres-sion of the adipogenic marker, FABP4, was also increased in primary adipocytes from Adipo-NRX mice ( Fig. 3F ). These data strongly suggest that NRX enhances adipogenesis.
Decreased lipid catabolism and increased infl ammation and fi brosis in WAT of Adipo-NRX mice
We found that expression of enzymes involved in lipid catabolism, including Atgl , Mcad , and Cpt1a were down-regulated, indicating that adipocytes of Adipo-NRX mice lacked the ability to burn excess fat. These results suggest that increased fat mass in Adipo-NRX mice is due to de-creased lipolysis and fatty acid oxidation. Because adipo-kine dysregulation is a hallmark of adipocyte impairment ( 26 ), we further measured adipokine expression in WAT of Adipo-NRX mice. The mRNA levels of adiponectin were downregulated, and PAI-1 was upregulated in WAT of Adipo-NRX mice.
Obesity has been consistently associated with infl amma-tion ( 4, 27 ). We examined mRNA expression levels of sev-eral infl ammatory genes in WAT of WT and Adipo-NRX mice. Among the genes upregulated in Adipo-NRX mice were TNF � and Cxcl10 , which encode proinfl ammatory cy-tokines; Cybb , which is involved in phagocytosis; and Emr1
Fig. 2. Adipo-NRX mice have larger amounts of adipose tissue compared with WT mice. A: Body weights and organ weights of 6-month-old Adipo-NRX and WT mice were determined (n = 6). B, C: Representative pictures of epididymal and perirenal fat dissected from fat pads of Adipo-NRX and WT mice. D: H and E staining of WAT from epididymal fat pads of Adipo-NRX and WT mice (left). Quantitative analysis of cell numbers/mm 2 in sections of WAT (right). E: Total cell number of adipocytes per fat fad isolated from WT and Adipo-NRX mice (n = 3). GTT (F) and ITT (G) in WT and Adipo-NRX mice. Total area under curve (AUC) of each graph was measured (insets). Fasted mice were injected with glucose (1 g/kg ip) or insulin (1 U/kg ip). Blood glucose levels (mg/dl) were determined at the indicated time points. Values are means ± SD of six to eight mice. H: Western blotting analysis of phospho-AKT (AKT-S473 phosphorylation) in indicated tissues isolated from WT and Adipo-NRX mice (n = 3) after intraperitoneal injection of insulin (10 U/kg). The symbol “ – ” or “ + ” means without or with insulin stimulation, respectively. Data are presented as mean percentages ± SD (* P < 0.05, ** P < 0.01). by guest, on A
ugust 29, 2018w
ww
.jlr.orgD
ownloaded from
.html http://www.jlr.org/content/suppl/2014/12/29/jlr.M054056.DC1Supplemental Material can be found at:

Nucleoredoxin regulates adipogenesis 299
through inhibition of GSK3 � , leading to the stabili-zation and nuclear translocation of � -catenin ( 12 ). It has been reported that NRX regulates Wnt/ � -catenin signal-ing by directly binding and inhibiting Dvl and thereby controls cell proliferation ( 16 ). Wnt/ � -catenin signaling is also a central negative regulator of adipogenesis ( 7 ). Here, we found that NRX is involved in adipogenic differentia-tion, prompting us to examine whether NRX regulates adipogenesis through Wnt/ � -catenin regulation. First, we tested the interaction between NRX and Dvl in adipocyte. Endogenous Dvl interacted with ectopically expressed WT NRX, but not with a cysteine-mutant NRX defective in thiol reducing activity, in 3T3-L1 preadipocytes ( Fig. 5A ). Moreover, unlike WT NRX, ectopically expressed cysteine-mutant NRX did not increase 3T3-L1 differentiation ( Fig. 5B ). These results suggest that NRX controls adipogenesis
the NRX coding sequence, we transfected NRX-knock-down 3T3-L1 cells with an expression plasmid for an shRNA-resistant NRX (resNRX) with a codon usage differ-ent from that of the original plasmid. resNRX expression restored PPAR � and FABP4 expression levels ( Fig. 4E ) as well as accumulation of lipid droplets ( Fig. 4D ). These re-sults clearly demonstrate that NRX is involved in adipo-genic differentiation.
NRX regulates adipogenesis through modulation of � -catenin signaling in 3T3-L1 preadipocytes
The conserved Wnt/ � -catenin signaling pathway medi-ates a variety of cellular responses, from cell proliferation to cell-fate determination ( 30, 31 ). Dvl transduces signals from the Wnt receptor, Frizzled, to downstream components
Fig. 3. Overexpression of NRX promotes adipogenic differentiation of 3T3-L1 preadipocytes and primary adipocytes. 3T3-L1 cells were infected with lentiviruses expressing NRX (pLenti-NRX-GFP-fl ag) or control (pLenti-GFP-fl ag), selected with puromycin, and induced to differentiate using MDI medium. A: Ectopic expression of NRX was monitored by Western blot analysis with anti-NRX antibody. B: After 8 days of differentiation, the degree of lipid accumulation was visualized by staining cells with Oil Red O. C: Staining in cells was quantifi ed using a dye-extraction solution (n = 4). D: Whole-cell lysates were extracted on day 0 or day 8 for Western blot analysis of NRX, PPAR � , FABP4, and � -actin. E: Primary white adipocytes isolated from Adipo-NRX and WT mice were cultured in MDI medium containing rosigli-tazone to induce differentiation into adipocytes. After 10 days of differentiation, cells were stained with Oil Red O dye. F: Whole-cell lysates were extracted from differentiated white adipocytes on day 10 for Western blot analysis of NRX, FABP4, and � -actin (n = 4). G: Relative mRNA abundance was examined in WAT from WT and Adipo-NRX mice by real-time quantitative RT-PCR (n = 3). Data are presented as mean fold-changes ± SD (* P < 0.05, ** P < 0.01, *** P < 0.001). H: Frozen sections of WAT isolated from WT and Adipo-NRX were immu-nostained with antibody against F4/80, a macrophage surface marker. Images were taken at magnifi cation ×200 (upper panels) and ×400 (lower panels). Atgl, adipose triglyceride lipase; Ccl5, chemokine (C-C motif) ligand 5; Cpt1a, carnitine palmitoyltransferase 1a; Cxcl10, chemokine (C-X-C motif) ligand 10; Cybb, cytochrome b-245, beta polypeptide; Emr1, Epidermal growth factor (EGF) -like module con-taining, mucin-like, hormone receptor-like 1; HSL, hormone-sensitive lipase; Itgax, integrin alpha X; Mcad, acyl-CoA dehydrogenase, me-dium chain; PAI-1, plasminogen activator inhibitor type 1; Col1a1, Collagen I; Col3a1, Collagen III; Col6a1, Collagen VI.
by guest, on August 29, 2018
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2014/12/29/jlr.M054056.DC1Supplemental Material can be found at:

300 Journal of Lipid Research Volume 56, 2015
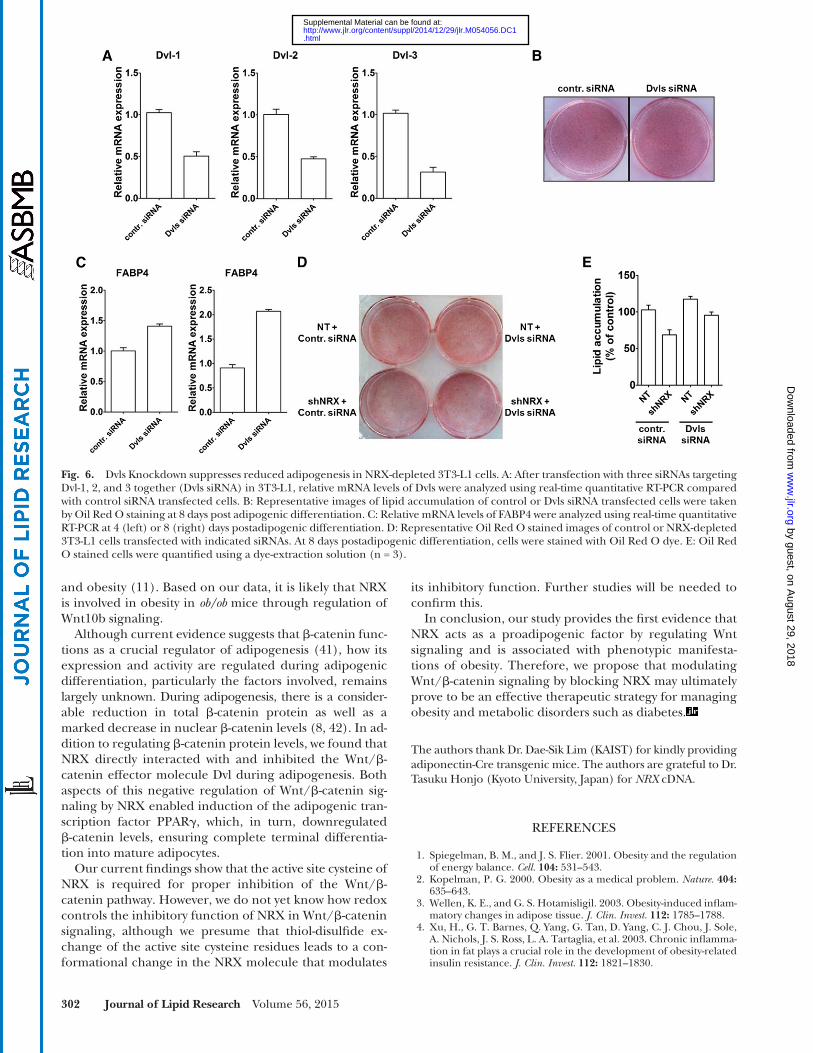
lipid accumulation by 31.2%, while knockdown of Dvls together reduced lipid accumulation only by 10.5%. These results suggest that NRX regulates adipogenesis through inhibition of Dvl- � -catenin axis in adipocyte differentiation.
DISCUSSION
Our study demonstrated that NRX mRNA and protein were increased in the early stages of adipocyte differentia-tion and were also increased in WAT of ob/ob mice, a leptin-defi ciency model of obesity. Using NRX-depleted and NRX-overexpressing 3T3-L1 preadipocytes, we showed that NRX is associated with adipogenic differentiation of 3T3-L1 cells. We found that differentiation of primary adi-pocytes from Adipo-NRX mice was increased in vitro, and epididymal and perirenal fat mass were increased in Adipo-NRX mice in vivo. These observations, taken to-gether with our in vitro and in vivo adipogenesis data, thus uncover a novel role for NRX as a proadipogenic factor.
Adipose tissue expansion occurs through an enlarge-ment in adipocyte size (hypertrophy) and/or an increase in adipocyte number (hyperplasia). We showed that Adipo-NRX mice exhibited hyperplasia in adipocytes, which is linked to enhanced adipogenic differentiation ( 32, 33 ), as well as hypertrophy ( Fig. 2D, E ). Adipo-NRX mice showed decreased expression of enzymes involved in lipolysis and fatty acid oxidation in WAT, while unchanged in lipogenic enzymes compared with WT mice. Decreased triacylglycerol catabolism is associated with the occurrence of prevalent metabolic diseases, such as obesity and type 2 diabetes ( 34–36 ). Hypertrophic adipocyte in our model is
by interacting with Dvl through a thiol-based mechanism. We further observed that endogenous NRX-Dvl binding was dissociated by Wnt3a treatment ( Fig. 5C ), suggesting that NRX controls adipogenesis via Wnt/ � -catenin signal-ing. To test this hypothesis, we determined whether NRX controls the nuclear localization of � -catenin in 3T3-L1 cells and primary adipocytes isolated from Adipo-NRX mice. Nuclear � -catenin levels and transcriptional activity were increased in NRX-knockdown cells but were de-creased in these cells following restoration of NRX expres-sion ( Fig. 5D ). Enhancing NRX expression by increasing the amount of transfected plasmid induced a dose-dependent decrease in the levels of nuclear � -catenin and its down-stream effector cyclin D1 in Wnt3a-treated 3T3-L1 cells ( Fig. 5E ), resulting in recovery from Wnt3a-mediated inhibition of adipogenesis ( Fig. 5F , third panel). Consis-tent with the results obtained in 3T3-L1 preadipocytes ( Fig. 5D, E ), the expression of � -catenin target genes was decreased in WAT of Adipo-NRX mice ( Fig. 5G ).
Next, we examined whether the interaction of NRX with Dvl plays a role in adipogenesis. While single Dvl gene ( Dvl-1 , Dvl-2 , or Dvl-3 ) knockdown showed no sig-nifi cant changes in adipocyte differentiation compared with control group (supplementary Fig. 3), triple knock-down of all Dvl genes enhanced differentiation of 3T3-L1 into mature adipocytes ( Fig. 6A , B ) accompanying with an elevated expression of FABP4, an adipocyte differentiation marker, compared with control ( Fig. 6C ). To test whether NRX role is dependent on Dvl, we checked the effect of NRX on adipogenesis in the presence or absence of three Dvls. We found that the Dvls knock-down suppressed reduced 3T3-L1 differentiation by NRX knockdown ( Fig. 6D, E ). NRX knockdown reduced
Fig. 4. NRX shRNA attenuates adipogenic differentiation in 3T3-L1 preadipocytes. A: NRX protein levels in NRX-knockdown and con-trol, nontarget (NT) 3T3-L1 cells were monitored by Western blot analysis with anti-NRX antibody. B: NRX-knockdown and control 3T3-L1 cells were differentiated for 8 days in MDI medium and then stained with Oil Red O dye. C: After induction of differentiation, whole-cell lysates were prepared from nontarget and NRX-knockdown cells on the indicate days for Western blot analysis of NRX, PPAR � , FABP4, and � -actin. � -Actin was used as a loading control. D: Rescue experiments were performed by transfecting NRX-knockdown 3T3-L1 cells with plasmids encoding resNRX. Eight days after differentiation, cells were stained with Oil Red O dye. E: Analysis of protein levels during ad-ipogenic differentiation of NRX-knockdown and NRX-rescued cells. Whole-cell lysates were extracted on day 8 for Western blot analysis of NRX, PPAR � , FABP4, and � -actin.
by guest, on August 29, 2018
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2014/12/29/jlr.M054056.DC1Supplemental Material can be found at:

Nucleoredoxin regulates adipogenesis 301
the molecular links between obesity and infl ammation have not been completely elucidated. Adipo-NRX mice might be a potential model system for detailed mechanistic studies in the relationship between obesity and infl ammation.
Wnt signaling is an important regulator of fate decisions in mesenchymal cells ( 31 ). In multipotent mesenchymal precursors in vitro, activation of Wnt/ � -catenin signaling promotes osteoblastogenesis but inhibits adipogenesis ( 39 ). Given that NRX inhibits Wnt/ � -catenin signaling ( 16 ), it is possible that NRX reciprocally modulates adipo-genic and osteogenic differentiation. Activation of � -catenin in primary osteoblasts isolated from calvariae of NRX � / � embryos results in enhanced osteoblastic differen-tiation ( 19 ). However, there have been no reports of a role for NRX in adipogenesis. Here, we provide the fi rst dem-onstration that NRX positively regulates adipogenesis by inhibiting the Wnt/ � -catenin pathway. Wnt10b is a likely candidate for the endogenous Wnt that participates in adi-pogenesis. The expression level of Wnt10b, which is highly expressed in preadipocytes, declined upon initiation of differentiation ( 40 ), exhibiting a negative correlation with NRX expression ( Fig. 1A ).Wnt10b has been reported to attenuate the development of obesity in ob/ob mice by stabi-lizing � -catenin and subsequently inhibiting adipogenesis. Conversely, blocking Wnt signaling promotes adipogenesis
likely caused by the decrease of triacylglycerol catabolism rather than an increase in triacylglycerol synthesis. How-ever, the mechanism by which NRX regulates fatty acid catabolism remains to be determined.
Lipid accumulation increased exclusively in WAT of Adipo-NRX mice, with no change in liver and skeletal muscle compared with WT mice (supplementary Fig. 2). In plasma analysis, Adipo-NRX mice exhibited increased fasting insulin level accompanying with increased fasting glucose level, suggesting that pancreas function was nor-mal (supplementary Table 1). Adipo-NRX mice showed the tendency to glucose intolerance in GTT analysis ( Fig. 2F ) and also showed mild insulin resistance as shown in impaired insulin tolerance and reduced insulin sensitivity in WAT but not in liver and skeletal muscle ( Fig. 2G, H ). Therefore, insulin resistance in our model is likely caused by excessive fat deposition in WAT.
Infi ltration of adipose tissue by infl ammatory cells has been described as a common feature of obesity ( 27 ). We also found increased expression of infl ammatory gene and mac-rophage markers as well as dysregulation of adipokines in Adipo-NRX mice ( Fig. 3G ). Many rodent models of obesity are associated with chronic infl ammation, and it is known that obesity-induced insulin resistance is strongly correlated with expression of infl ammatory markers ( 37, 38 ). However,
Fig. 5. NRX regulates adipogenesis via modulation of � -catenin in 3T3-L1 cells. A, B: WT NRX interacts with Dvl and increases 3T3-L1 differentiation. A: 3T3-L1 cells were transfected with vectors expressing FLAG-tagged WT or mutant (Mut) NRX, lysed and immunoprecipi-tated with anti-FLAG antibody, and analyzed by immunoblotting with anti-Dvl antibody. B: After 8 days of incubation in differentiation media, cells were stained with Oil Red O dye. C: 3T3-L1 cell lysates were immunoprecipitated using anti-NRX antibodies after treatment with 200 ng/ml Wnt3a followed by immunoblotting, as indicated. D–F: NRX inhibits � -catenin activity and promotes recovery from Wnt3a-mediated inhibition of adipogenesis. D: Reporter gene (TOPfl ash) expression assays in NRX-knockdown 3T3-L1 cells after cotransfection of increasing amounts of resNRX. Nuclear (N) and cytosolic (C) fractions were analyzed by immunoblotting. Lamin A/C was used as a control for the nuclear fraction. E: 3T3-L1 cells transfected with vectors expressing FLAG-tagged NRX were treated with Wnt3a. Cytosolic and nuclear proteins were isolated for Western blot analysis of FLAG-NRX, � -catenin, lamin A/C, and cyclin D1. F: 3T3-L1 cells overex-pressing NRX were induced to differentiate into adipocytes with Wnt3a for the fi rst 2 days. After 8 days of differentiation, cells were stained with Oil Red O dye. Whole-cell lysates were prepared on day 8 for Western blot analysis of NRX, FABP4, and � -actin. G: � -Catenin target genes were examined in WAT from WT and Adipo-NRX mice by real-time quantitative RT-PCR (n = 3). Data are presented as mean fold-changes ± SD (* P < 0.05, ** P < 0.01). MMP-7, matrix metalloproteinase-7.
by guest, on August 29, 2018
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2014/12/29/jlr.M054056.DC1Supplemental Material can be found at:
302 Journal of Lipid Research Volume 56, 2015
its inhibitory function. Further studies will be needed to confi rm this.
In conclusion, our study provides the fi rst evidence that NRX acts as a proadipogenic factor by regulating Wnt signaling and is associated with phenotypic manifesta-tions of obesity. Therefore, we propose that modulating Wnt/ � -catenin signaling by blocking NRX may ultimately prove to be an effective therapeutic strategy for managing obesity and metabolic disorders such as diabetes.
The authors thank Dr. Dae-Sik Lim (KAIST) for kindly providing adiponectin-Cre transgenic mice. The authors are grateful to Dr. Tasuku Honjo (Kyoto University, Japan) for NRX cDNA.
REFERENCES
1 . Spiegelman , B. M. , and J. S. Flier . 2001 . Obesity and the regulation of energy balance. Cell . 104 : 531 – 543 .
2 . Kopelman , P. G. 2000 . Obesity as a medical problem. Nature . 404 : 635 – 643 .
3 . Wellen , K. E. , and G. S. Hotamisligil . 2003 . Obesity-induced infl am-matory changes in adipose tissue. J. Clin. Invest. 112 : 1785 – 1788 .
4 . Xu , H. , G. T. Barnes , Q. Yang , G. Tan , D. Yang , C. J. Chou , J. Sole , A. Nichols , J. S. Ross , L. A. Tartaglia , et al . 2003 . Chronic infl amma-tion in fat plays a crucial role in the development of obesity-related insulin resistance. J. Clin. Invest. 112 : 1821 – 1830 .
and obesity ( 11 ). Based on our data, it is likely that NRX is involved in obesity in ob/ob mice through regulation of Wnt10b signaling.
Although current evidence suggests that � -catenin func-tions as a crucial regulator of adipogenesis ( 41 ), how its expression and activity are regulated during adipogenic differentiation, particularly the factors involved, remains largely unknown. During adipogenesis, there is a consider-able reduction in total � -catenin protein as well as a marked decrease in nuclear � -catenin levels ( 8, 42 ). In ad-dition to regulating � -catenin protein levels, we found that NRX directly interacted with and inhibited the Wnt/ � -catenin effector molecule Dvl during adipogenesis. Both aspects of this negative regulation of Wnt/ � -catenin sig-naling by NRX enabled induction of the adipogenic tran-scription factor PPAR � , which, in turn, downregulated � -catenin levels, ensuring complete terminal differentia-tion into mature adipocytes.
Our current fi ndings show that the active site cysteine of NRX is required for proper inhibition of the Wnt/ � -catenin pathway. However, we do not yet know how redox controls the inhibitory function of NRX in Wnt/ � -catenin signaling, although we presume that thiol-disulfi de ex-change of the active site cysteine residues leads to a con-formational change in the NRX molecule that modulates
Fig. 6. Dvls Knockdown suppresses reduced adipogenesis in NRX-depleted 3T3-L1 cells. A: After transfection with three siRNAs targeting Dvl-1, 2, and 3 together (Dvls siRNA ) in 3T3-L1, relative mRNA levels of Dvls were analyzed using real-time quantitative RT-PCR compared with control siRNA transfected cells. B: Representative images of lipid accumulation of control or Dvls siRNA transfected cells were taken by Oil Red O staining at 8 days post adipogenic differentiation. C: Relative mRNA levels of FABP4 were analyzed using real-time quantitative RT-PCR at 4 (left) or 8 (right) days postadipogenic differentiation. D: Representative Oil Red O stained images of control or NRX-depleted 3T3-L1 cells transfected with indicated siRNAs. At 8 days postadipogenic differentiation, cells were stained with Oil Red O dye. E: Oil Red O stained cells were quantifi ed using a dye-extraction solution (n = 3).
by guest, on August 29, 2018
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2014/12/29/jlr.M054056.DC1Supplemental Material can be found at:

Nucleoredoxin regulates adipogenesis 303
5 . Farmer , S. R. 2006 . Transcriptional control of adipocyte formation. Cell Metab. 4 : 263 – 273 .
6 . Rosen , E. D. , and B. M. Spiegelman . 2000 . Molecular regulation of adipogenesis. Annu. Rev. Cell Dev. Biol. 16 : 145 – 171 .
7 . Ross , S. E. , N. Hemati , K. A. Longo , C. N. Bennett , P. C. Lucas , R. L. Erickson , and O. A. MacDougald . 2000 . Inhibition of adipogenesis by Wnt signaling. Science . 289 : 950 – 953 .
8 . Moldes , M. , Y. Zuo , R. F. Morrison , D. Silva , B. H. Park , J. Liu , and S. R. Farmer . 2003 . Peroxisome-proliferator-activated receptor gamma suppresses Wnt/beta-catenin signalling during adipogen-esis. Biochem. J. 376 : 607 – 613 .
9 . Grant , S. F. , G. Thorleifsson , I. Reynisdottir , R. Benediktsson , A. Manolescu , J. Sainz , A. Helgason , H. Stefansson , V. Emilsson , A. Helgadottir , et al . 2006 . Variant of transcription factor 7-like 2 (TCF7L2) gene confers risk of type 2 diabetes. Nat. Genet. 38 : 320 – 323 .
10 . Longo , K. A. , W. S. Wright , S. Kang , I. Gerin , S. H. Chiang , P. C. Lucas , M. R. Opp , and O. A. MacDougald . 2004 . Wnt10b inhibits development of white and brown adipose tissues. J. Biol. Chem. 279 : 35503 – 35509 .
11 . Wright , W. S. , K. A. Longo , V. W. Dolinsky , I. Gerin , S. Kang , C. N. Bennett , S. H. Chiang , T. C. Prestwich , C. Gress , C. F. Burant , et al . 2007 . Wnt10b inhibits obesity in ob/ob and agouti mice. Diabetes . 56 : 295 – 303 .
12 . Behrens , J. , B. A. Jerchow , M. Wurtele , J. Grimm , C. Asbrand , R. Wirtz , M. Kuhl , D. Wedlich , and W. Birchmeier . 1998 . Functional interaction of an axin homolog, conductin, with beta-catenin, APC, and GSK3beta. Science . 280 : 596 – 599 .
13 . Nelson , W. J. , and R. Nusse . 2004 . Convergence of Wnt, beta-catenin, and cadherin pathways. Science . 303 : 1483 – 1487 .
14 . Lillig , C. H. , and A. Holmgren . 2007 . Thioredoxin and related mol-ecules—from biology to health and disease. Antioxid. Redox Signal. 9 : 25 – 47 .
15 . Kurooka , H. , K. Kato , S. Minoguchi , Y. Takahashi , J. Ikeda , S. Habu , N. Osawa , A. M. Buchberg , K. Moriwaki , H. Shisa , et al . 1997 . Cloning and characterization of the nucleoredoxin gene that en-codes a novel nuclear protein related to thioredoxin. Genomics . 39 : 331 – 339 .
16 . Funato , Y. , T. Michiue , M. Asashima , and H. Miki . 2006 . The thio-redoxin-related redox-regulating protein nucleoredoxin inhibits Wnt-beta-catenin signalling through dishevelled. Nat. Cell Biol. 8 : 501 – 508 .
17 . Hayashi , T. , Y. Funato , T. Terabayashi , A. Morinaka , R. Sakamoto , H. Ichise , H. Fukuda , N. Yoshida , and H. Miki . 2010 . Nucleoredoxin negatively regulates Toll-like receptor 4 signaling via recruitment of fl ightless-I to myeloid differentiation primary response gene (88). J. Biol. Chem. 285 : 18586 – 18593 .
18 . Funato , Y. , and H. Miki . 2007 . Nucleoredoxin, a novel thiore-doxin family member involved in cell growth and differentiation. Antioxid. Redox Signal. 9 : 1035 – 1057 .
19 . Funato , Y. , T. Terabayashi , R. Sakamoto , D. Okuzaki , H. Ichise , H. Nojima , N. Yoshida , and H. Miki . 2010 . Nucleoredoxin sustains Wnt/ � -catenin signaling by retaining a pool of inactive dishevelled protein . Curr. Biol. 20 : 1945 – 1952 .
20 . Babaya , N. , H. Ikegami , T. Fujisawa , K. Nojima , M. Itoi-Babaya , K. Inoue , T. Ohno , M. Shibata , and T. Ogihara . 2005 . Susceptibility to streptozotocin-induced diabetes is mapped to mouse chromosome 11. Biochem. Biophys. Res. Commun. 328 : 158 – 164 .
21 . Soukas , A. , N. D. Socci , B. D. Saatkamp , S. Novelli , and J. M. Friedman . 2001 . Distinct transcriptional profi les of adipogenesis in vivo and in vitro. J. Biol. Chem. 276 : 34167 – 34174 .
22 . Student , A. K. , R. Y. Hsu , and M. D. Lane . 1980 . Induction of fatty acid synthetase synthesis in differentiating 3T3-L1 preadipocytes. J. Biol. Chem. 255 : 4745 – 4750 .
23 . Ramirez-Zacarías , J. L. , F. Castro-Munozledo , and W. Kuri-Harcuch . 1992 . Quantitation of adipose conversion and triglycerides by staining intracytoplasmic lipids with Oil red O. Histochemistry . 97 : 493 – 497 .
24 . Carnevalli , L. S. , K. Masuda , F. Frigerio , O. Le Bacquer , S. H. Um , V. Gandin , I. Topisirovic , N. Sonenberg , G. Thomas , and S. C. Kozma . 2010 . S6K1 plays a critical role in early adipocyte differen-tiation. Dev. Cell . 18 : 763 – 774 .
25 . Díez , J. J. , and P. Iglesias . 2003 . The role of the novel adipocyte-derived hormone adiponectin in human disease . Eur. J. Endocrinol. 148 : 293 – 300 .
26 . Huh , J. Y. , Y. Kim , J. Jeong , J. Park , I. Kim , K. H. Huh , Y. S. Kim , H. A. Woo , S. G. Rhee , K. J. Lee , et al . 2012 . Peroxiredoxin 3 is a key mole-cule regulating adipocyte oxidative stress, mitochondrial biogenesis, and adipokine expression. Antioxid. Redox Signal. 16 : 229 – 243 .
27 . Weisberg , S. P. , D. McCann , M. Desai , M. Rosenbaum , R. L. Leibel , and A. W. Ferrante , Jr . 2003 . Obesity is associated with macrophage accumulation in adipose tissue. J. Clin. Invest. 112 : 1796 – 1808 .
28 . Halberg , N. , T. Khan , M. E. Trujillo , I. Wernstedt-Asterholm , A. D. Attie , S. Sherwani , Z. V. Wang , S. Landskroner-Eiger , S. Dineen , U. J. Magalang , et al . 2009 . Hypoxia-inducible factor 1alpha induces fi brosis and insulin resistance in white adipose tissue. Mol. Cell. Biol. 29 : 4467 – 4483 .
29 . Wang , Q. A. , P. E. Scherer , and R. K. Gupta . 2014 . Improved meth-odologies for the study of adipose biology: insights gained and op-portunities ahead. J. Lipid Res. 55 : 605 – 624 .
30 . Clevers , H. 2006 . Wnt/beta-catenin signaling in development and disease. Cell . 127 : 469 – 480 .
31 . Teo , R. , F. Mohrlen , G. Plickert , W. A. Muller , and U. Frank . 2006 . An evolutionary conserved role of Wnt signaling in stem cell fate decision. Dev. Biol. 289 : 91 – 99 .
32 . Wang , Q. A. , C. Tao , R. K. Gupta , and P. E. Scherer . 2013 . Tracking adipogenesis during white adipose tissue development, expansion and regeneration. Nat. Med. 19 : 1338 – 1344 .
33 . Sun , K. , C. M. Kusminski , and P. E. Scherer . 2011 . Adipose tissue remodeling and obesity. J. Clin. Invest. 121 : 2094 – 2101 .
34 . Langin , D. , A. Dicker , G. Tavernier , J. Hoffstedt , A. Mairal , M. Ryden , E. Arner , A. Sicard , C. M. Jenkins , N. Viguerie , et al . 2005 . Adipocyte lipases and defect of lipolysis in human obesity. Diabetes . 54 : 3190 – 3197 .
35 . Zechner , R. , P. C. Kienesberger , G. Haemmerle , R. Zimmermann , and A. Lass . 2009 . Adipose triglyceride lipase and the lipolytic ca-tabolism of cellular fat stores. J. Lipid Res. 50 : 3 – 21 .
36 . Jocken , J. W. , D. Langin , E. Smit , W. H. Saris , C. Valle , G. B. Hul , C. Holm , P. Arner , and E. E. Blaak . 2007 . Adipose triglyceride li-pase and hormone-sensitive lipase protein expression is decreased in the obese insulin-resistant state. J. Clin. Endocrinol. Metab. 92 : 2292 – 2299 .
37 . Blackburn , P. , J. P. Despres , B. Lamarche , A. Tremblay , J. Bergeron , I. Lemieux , and C. Couillard . 2006 . Postprandial variations of plasma infl ammatory markers in abdominally obese men. Obesity (Silver Spring) . 14 : 1747 – 1754 .
38 . Aron-Wisnewsky , J. , J. Tordjman , C. Poitou , F. Darakhshan , D. Hugol , A. Basdevant , A. Aissat , M. Guerre-Millo , and K. Clement . 2009 . Human adipose tissue macrophages: m1 and m2 cell surface markers in subcutaneous and omental depots and after weight loss. J. Clin. Endocrinol. Metab. 94 : 4619 – 4623 .
39 . Cawthorn , W. P. , A. J. Bree , Y. Yao , B. Du , N. Hemati , G. Martinez-Santibanez , and O. A. MacDougald . 2012 . Wnt6, Wnt10a and Wnt10b inhibit adipogenesis and stimulate osteoblastogenesis through a beta-catenin-dependent mechanism. Bone . 50 : 477 – 489 .
40 . Bennett , C. N. , S. E. Ross , K. A. Longo , L. Bajnok , N. Hemati , K. W. Johnson , S. D. Harrison , and O. A. MacDougald . 2002 . Regulation of Wnt signaling during adipogenesis. J. Biol. Chem. 277 : 30998 – 31004 .
41 . Kennell , J. A. , and O. A. MacDougald . 2005 . Wnt signaling inhibits adipogenesis through beta-catenin-dependent and -independent mechanisms. J. Biol. Chem. 280 : 24004 – 24010 .
42 . Hou , J. C. , S. Shigematsu , H. C. Crawford , P. Z. Anastasiadis , and J. E. Pessin . 2006 . Dual regulation of Rho and Rac by p120 catenin controls adipocyte plasma membrane traffi cking. J. Biol. Chem. 281 : 23307 – 23312 .
by guest, on August 29, 2018
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2014/12/29/jlr.M054056.DC1Supplemental Material can be found at:





























