Embed Size (px)
Citation preview

ARTHRITIS & RHEUMATISMVol. 58, No. 11, November 2008, pp 3419–3424DOI 10.1002/art.23990© 2008, American College of Rheumatology
Novel Detection of In Vivo HLA–B27 ConformationsCorrelates With Ankylosing Spondylitis Association
Helen Fussell,1 Darren Nesbeth,2 Izabela Lenart,2 Elaine C. Campbell,3 Sarah Lynch,3
Susana Santos,4 Keith Gould,5 Simon J. Powis,3 and Antony N. Antoniou2
Objective. The class I major histocompatibilitycomplex (MHC) molecule HLA–B27 exhibits a strongassociation with the autoimmune inflammatory arthri-tis disorder ankylosing spondylitis (AS) and with otherrelated spondylarthropathies. In the absence of both adefined autoimmune response and a target autoanti-gen(s), the propensity of HLA–B27 to misfold has beenhypothesized to be a major parameter in disease patho-genesis. We undertook this study to test the hypothesisthat HLA–B27 misfolding is due to exposure of cysteineresidues within the heavy chain to the oxidizing envi-ronment of the endoplasmic reticulum.
Methods. A rapid acidification and alkylationmodification method was used to examine cysteineresidue exposure and accessibility within AS-associatedand non-AS–associated HLA–B27 subtypes.
Results. This novel approach to probing in vivoclass I MHC structure revealed that the HLA–B27 heavy
chain adopts conformations not previously described.Furthermore, amino acid residues specific to subtypesHLA–B*2706, B*2709, and B*2704 can have an impacton these novel conformations and on cysteine residueexposure.
Conclusion. HLA–B27 can adopt novel conforma-tions, resulting in differential accessibility of cysteineresidues, which can explain the propensity to misfold.Cysteine exposure in the HLA–B27 heavy chain is alsoaffected by residues within the 114 and 116 regions,thereby providing a potential biochemical basis for theassociation of HLA–B27 subtypes with AS.
HLA–B27 is strongly associated with develop-ment of the autoimmune inflammatory arthritis diseaseankylosing spondylitis (AS) and other spondylarthropa-thies (SpA) (1,2). The majority of HLA–B27 subtypesare associated with the SpA, but HLA–B*2706 andB*2709 generally exhibit little or no correlation with AS(3–5). Compared with disease-associated subtypes,B*2706 and B*2709 exhibit subtle amino acid sequencechanges, predominantly at the structurally importantp114 and p116, which constitute the F pocket of thepeptide-binding groove and can determine the require-ment for the class I major histocompatibility complex(MHC)–specific accessory molecule tapasin (6). Com-pared with the AS-associated B*2705 subtype, B*2709differs by a single aspartic acid–to-histidine substitutionat p116 (D1163H116), while B*2706 exhibits an aspar-tic acid and a tyrosine at p114 and p116, respectively(H1143D114, D1163Y116), in addition to a serine and aglutamic acid at p77 (D773S77) and p152 (V1523E152),respectively, with the latter 2 residues also present in theAS-associated B*2704 subtype.
Class I MHC molecules possess 4 conservedstructural cysteine (Cys) residues in the endoplasmicreticulum (ER)–luminal domain, C101, C164, C203, andC259. However, HLA–B27 possesses an additional ER-
Dr. Fussell’s work was supported by an Arthritis ResearchCampaign (ARC) UK project grant (17222). Drs. Nesbeth and Gould’swork was supported by the Medical Research Council, UK (ref.G-0400418). Ms Lenart’s work was supported by an ARC studentship(17868). Ms Lynch’s work was supported by the University of St.Andrews Maitland-Ramsay PhD studentship. Dr. Santos’ work wassupported by a fellowship from the Portuguese Foundation for Scienceand Technology (grant SFRH/BPD/20964/2004). Dr. Antoniou’s workwas supported by an ARC UK Career Development Fellowship(15293).
1Helen Fussell, PhD: National Health Service Blood andTransplant, Colindale Blood Centre, London, UK; 2Darren Nesbeth,PhD, Izabela Lenart, BSc, MSc, Antony N. Antoniou, PhD: UniversityCollege London, London, UK; 3Elaine C. Campbell, PhD, SarahLynch, BSc, Simon J. Powis, PhD: University of St. Andrews, Fife, UK;4Susana Santos, PhD: University of Dundee, Dundee, UK; 5KeithGould, PhD: Imperial College London, London, UK.
Address correspondence and reprint requests to Antony N.Antoniou, PhD, Department of Immunology and Molecular Pathol-ogy, Division of Infection and Immunity/Centre of Rheumatology,University College London, Windeyer Institute of Medical Science, 46Cleveland Street, London W1T 4JF, UK. E-mail: [email protected].
Submitted for publication July 7, 2007; accepted in revisedform July 21, 2008.
3419

luminal Cys residue at p67, as well as Cys residues in thetransmembrane and cytosolic domains at p308 and p325,respectively. Apart from forming monomeric heavychain, which associates with �2-microglobulin (�2m) andpeptide, HLA–B27 has an enhanced propensity to mis-fold and form heavy-chain dimers, which can involveC67 and C164 (7).
It has not been determined why HLA–B27 exhib-its such structural anomalies. Disulfide bond formationrequires appropriate exposure and reactivity of Cysresidues (8); therefore, inappropriate disulfide bondformation may be essential to understanding HLA–B27misfolding. In this study, we adopted a unique approachto determine Cys residue accessibility within HLA–B27heavy chains in vivo, using a rapid acidification andalkylation method to demonstrate novel heavy-chainconformations, and we found that residues 114 and 116had the greatest influence on Cys accessibility.
MATERIALS AND METHODS
Cell lines and antibodies. C58, CEM, and CEM.NKRcells were maintained in R10 medium (RPMI 1640 plus 10%fetal bovine serum; Globepharm, Guildford, UK), and HeLaand HEK 293 cells were maintained in Dulbecco’s modifiedEagle’s medium. C58.HLA–B*2705, C58.HLA–B*2704,C58.HLA–B*2709, C58.HLA–B*2706, C58.H114D, C58.D116Y,C58.H114D.D116Y, C58.D77S, C58.V152E, C58.HLA–B*0702, C58.C67S, and C58.HLA–B27.human tapasin (hutpn)transfectants were generated by electroporation and selectedin medium plus G418 (1 mg/ml). Monoclonal antibody HC10recognizes unfolded HLA–B and C molecules. Anti-V5 anti-body was obtained from Serotec (Oxford, UK), and horserad-ish peroxidase–conjugated goat anti-mouse secondaryantibody was obtained from Dako (Glostrup, Denmark).
Rapid acidification and alkylation. Briefly, �5 � 105
cells were incubated at 37°C with or without 5 mM dithiothre-itol (DTT) with or without 0.5 mM diamide for 15 minutes,precipitated with 200 �l of 10% trichloroacetic acid (TCA)/phosphate buffered saline (PBS) at 4°C for 10 minutes, thencentrifuged at 14,000 revolutions per minute at 4°C for 10minutes. Pellets were washed in acetone, air-dried, and resus-pended in nonreducing sample buffer with or without AMS(Invitrogen, San Diego, CA) at 10 mg/ml.
Site-directed mutagenesis. Mutagenesis was per-formed based on the QuickChange (Stratagene, La Jolla, CA)protocol (9).
Single-chain constructs. The single-chain dimer con-sists of human �2m, a (GGGGS)3 linker, and HLA–B*2705class I heavy-chain sequence. The single-chain trimer com-prises human �2m amino-terminal hydrophobic signal sequence,the peptide sequence SRYWAIRTR (residues 383–391 of influ-enza virus nucleoprotein peptide [NP]), a GGGGGG(SGG)3
linker, the human �2m sequence, a second (GGGGS)3 linker,and B*2705 class I heavy-chain sequence.
Immunoblotting. Samples were resolved by sodiumdodecyl sulfate–polyacrylamide gel electrophoresis, trans-ferred and blocked with 5% milk powder in PBS/0.1% Tween,then incubated with appropriate antibodies. Images wererevealed by chemiluminescence using SuperSignal Femto(Pierce, Rockford, IL) and developed on a Fluor-S Multi-Imager Max imager (Bio-Rad, Richmond, CA) using QuantityOne 1D analysis software (Bio-Rad, Hercules, CA).
RESULTS
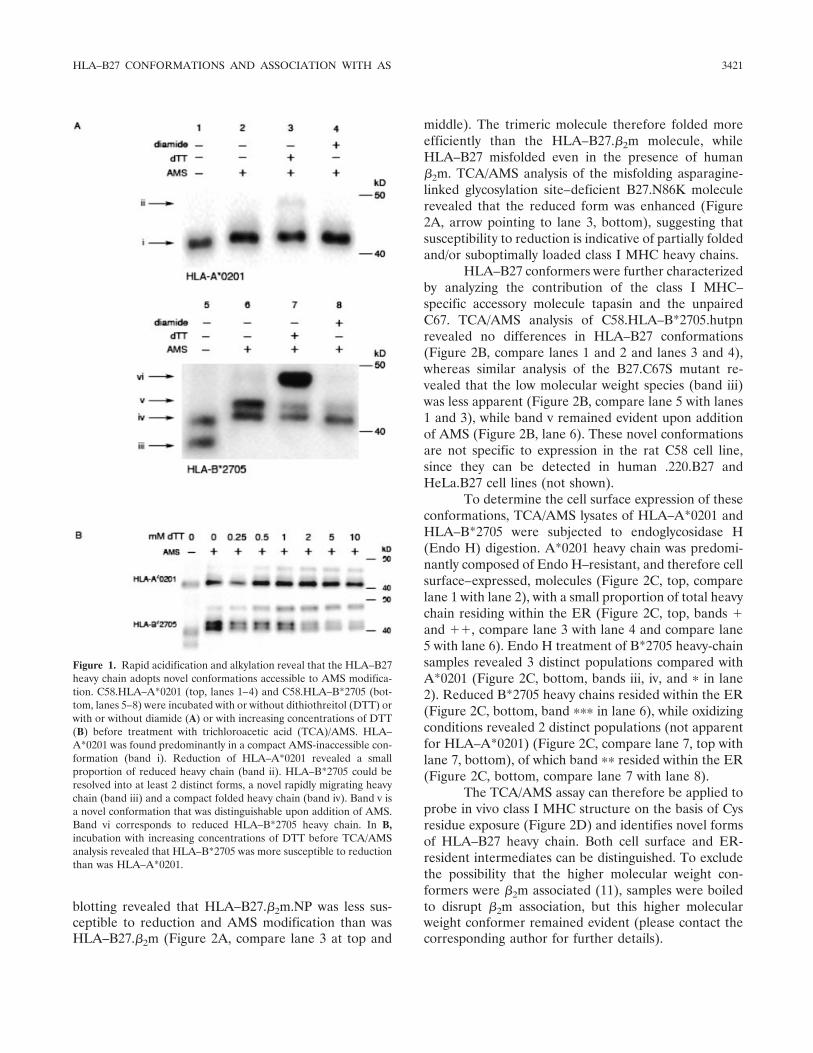
Detection of novel in vivo conformations adoptedby HLA–B27. As a first step toward determining Cysresidue exposure within HLA–B27, a rapid cellularacidification and alkylation approach was used to com-pare and determine the redox status of HLA–B27 withthat of HLA–A2. Rat thymoma C58 cell lines expressingeither C-terminal V5-tagged HLA–B*2705 or HLA–A*0201 heavy chains were incubated with either the cell-permeable reducing agent DTT or the oxidizing agentdiamide to establish the conformations of reduced andoxidized class I MHC heavy chains, respectively. Thiswas followed by cellular acidification with TCA to “trap”the heavy-chain redox status and prevent postlysis oxi-dation events and alkylation in nonreducing samplebuffer with or without the alkylating agent AMS (10).Lysates were immunoblotted for class I MHC heavychains with the anti-V5 antibody.
HLA–A*0201 exhibited no significant change inmolecular weight following the addition of AMS (Figure1A, band i, compare lane 1 with lane 2), with a smallproportion of heavy chain accessible to AMS followingreduction (Figure 1A, band ii, lane 3). Strikingly, HLA–B*2705 (Figure 1A, lanes 5–8) revealed several differ-ences compared with HLA–A*0201. In the absence ofAMS, 2 discernible HLA–B*2705 conformers (Figure 1A,bands iii and iv, lane 5) were detected. AMS addition led tothe resolution of 2 distinct species (Figure 1A, bands ivand v, lane 6) and an apparent absence of band iii. Asignificant proportion of heavy chain at steady state wassusceptible to reduction (Figure 1A, band vi, lane 7). Byexposing HLA–A*0201 and HLA–B*2705 to increasingconcentrations of DTT, TCA/AMS analysis revealedthat HLA–B*2705 was more sensitive to reduction thanHLA–A*0201 (Figure 1B).
Association of AMS-modifiable HLA–B27 heavychains with misfolding. Sensitivity to reduction is indic-ative of suboptimal peptide loading and/or partial fold-ing of the heavy chain (9). To determine the influenceof peptide on HLA–B27 thiol accessibility, we useda single-chain HLA–B27 molecule that was either di-rectly fused to human �2m or was fused to human �2mand a B27-specific peptide (influenza virus NP). Immuno-
3420 FUSSELL ET AL
blotting revealed that HLA–B27.�2m.NP was less sus-ceptible to reduction and AMS modification than wasHLA–B27.�2m (Figure 2A, compare lane 3 at top and
middle). The trimeric molecule therefore folded moreefficiently than the HLA–B27.�2m molecule, whileHLA–B27 misfolded even in the presence of human�2m. TCA/AMS analysis of the misfolding asparagine-linked glycosylation site–deficient B27.N86K moleculerevealed that the reduced form was enhanced (Figure2A, arrow pointing to lane 3, bottom), suggesting thatsusceptibility to reduction is indicative of partially foldedand/or suboptimally loaded class I MHC heavy chains.
HLA–B27 conformers were further characterizedby analyzing the contribution of the class I MHC–specific accessory molecule tapasin and the unpairedC67. TCA/AMS analysis of C58.HLA–B*2705.hutpnrevealed no differences in HLA–B27 conformations(Figure 2B, compare lanes 1 and 2 and lanes 3 and 4),whereas similar analysis of the B27.C67S mutant re-vealed that the low molecular weight species (band iii)was less apparent (Figure 2B, compare lane 5 with lanes1 and 3), while band v remained evident upon additionof AMS (Figure 2B, lane 6). These novel conformationsare not specific to expression in the rat C58 cell line,since they can be detected in human .220.B27 andHeLa.B27 cell lines (not shown).
To determine the cell surface expression of theseconformations, TCA/AMS lysates of HLA–A*0201 andHLA–B*2705 were subjected to endoglycosidase H(Endo H) digestion. A*0201 heavy chain was predomi-nantly composed of Endo H–resistant, and therefore cellsurface–expressed, molecules (Figure 2C, top, comparelane 1 with lane 2), with a small proportion of total heavychain residing within the ER (Figure 2C, top, bands �and ��, compare lane 3 with lane 4 and compare lane5 with lane 6). Endo H treatment of B*2705 heavy-chainsamples revealed 3 distinct populations compared withA*0201 (Figure 2C, bottom, bands iii, iv, and � in lane2). Reduced B*2705 heavy chains resided within the ER(Figure 2C, bottom, band ��� in lane 6), while oxidizingconditions revealed 2 distinct populations (not apparentfor HLA–A*0201) (Figure 2C, compare lane 7, top withlane 7, bottom), of which band �� resided within the ER(Figure 2C, bottom, compare lane 7 with lane 8).
The TCA/AMS assay can therefore be applied toprobe in vivo class I MHC structure on the basis of Cysresidue exposure (Figure 2D) and identifies novel formsof HLA–B27 heavy chain. Both cell surface and ER-resident intermediates can be distinguished. To excludethe possibility that the higher molecular weight con-formers were �2m associated (11), samples were boiledto disrupt �2m association, but this higher molecularweight conformer remained evident (please contact thecorresponding author for further details).
Figure 1. Rapid acidification and alkylation reveal that the HLA–B27heavy chain adopts novel conformations accessible to AMS modifica-tion. C58.HLA–A*0201 (top, lanes 1–4) and C58.HLA–B*2705 (bot-tom, lanes 5–8) were incubated with or without dithiothreitol (DTT) orwith or without diamide (A) or with increasing concentrations of DTT(B) before treatment with trichloroacetic acid (TCA)/AMS. HLA–A*0201 was found predominantly in a compact AMS-inaccessible con-formation (band i). Reduction of HLA–A*0201 revealed a smallproportion of reduced heavy chain (band ii). HLA–B*2705 could beresolved into at least 2 distinct forms, a novel rapidly migrating heavychain (band iii) and a compact folded heavy chain (band iv). Band v isa novel conformation that was distinguishable upon addition of AMS.Band vi corresponds to reduced HLA–B*2705 heavy chain. In B,incubation with increasing concentrations of DTT before TCA/AMSanalysis revealed that HLA–B*2705 was more susceptible to reductionthan was HLA–A*0201.
HLA–B27 CONFORMATIONS AND ASSOCIATION WITH AS 3421

Residues at p114 and p116 affect accessibility ofHLA–B27 cysteine residues to thiol modification. TCA/AMS was applied to B27 mutants mimicking the non-AS–associated HLA–B*2706 and B*2709 subtypes andthe AS-associated B*2704 subtype. B*2705 was mutatedas follows: H114D(06), D116Y(06), D116H(09),H114D.D116Y(06), D77S(04), and V152E(04) and ex-pressed in C58. Residue changes at p116 demonstrateda dramatic impact on HLA–B27 Cys residue exposure(Figure 3A). B27.H114D was more susceptible to reduc-tion (Figure 3A, band vi, lane 3), but the introduction ofD116Y or H114D.D116Y resulted in a conformationless accessible to AMS modification, especially underreducing conditions (Figure 3A, compare band vi withband iv, lane 3). Interestingly, the low molecular weightstructure (Figure 3A, band iii, lane 1) was still evident inall HLA–B27 molecules. However, there was an appar-
ent change in resolution of the conformation depicted byband v, following changes at p116 (Figure 3A, lanes 2and 4). Quantitation of complementary DNA and pro-tein levels of the 114 and 116 mutants revealed expres-sion within 2–3-fold of each other, except for B*2709(please contact the corresponding author for furtherdetails). Fluorescence-activated cell sorting analysisshowed that most mutants (except for H114D) wereexpressed at levels similar to those of B*2705 (Fussell H,Antoniou AN: unpublished observations), suggestingthat these conformations were not due to differentialexpression levels.
We then analyzed the effects of residues ex-pressed by both the AS-associated subtype (B*2704) andthe non-AS–associated subtype (B*2706) (i.e., S77 andE152). Interestingly, V152E appeared to be more sus-ceptible to reduction than D77S. Furthermore, B*2704
Figure 2. Modification of misfolding HLA–B27 heavy chains by AMS. A, The �2-microglobulin (�2m) and the influenza virus nucleoprotein peptide(NP) transfectants C58.HLA–B*2705.�2m (HLA–�27.�2m) and C58.HLA–B*2705.�2m.NP (HLA–�27.�2m.NP) were subjected to TCA/AMS.C58.HLA–B*2705.�2m was more susceptible to DTT-mediated reduction than was C58.HLA–B*2705.�2m.NP (arrow) (lanes 1–4). The transfectantC58.HLA–B*2705.N86K (HLA–�27.N86K) was much more susceptible to reduction (lanes 1–4). B, C58.HLA–B*2705 with or without humantapasin (tpn) (lanes 1–4) and HLA–B27.C67S (lanes 5 and 6) was subjected to TCA/AMS analysis. Human tapasin did not influence the presenceof HLA–B27 conformers (bands iii and v), while C67S had a modest influence on band iii. C, HLA–A*0201 (lanes 1–8, top) and HLA–B*2705 (lanes1–8, bottom) TCA/AMS-treated samples were incubated with or without endoglycosidase H (Endo H) at 37°C for 1 hour. Bands modified by AMSwere susceptible to Endo H digestion and therefore resided within the endoplasmic reticulum (ER). Endo H digestion of HLA–A*0201 revealedunfolded molecules that resided within the ER (bands � and ��). Following Endo H digestion, HLA–B*2705 could be resolved into 3 bands (iii,iv, and �). Reduced HLA–B*2705 heavy chain (band ���) and a novel AMS-bound form (band ��) resided within the ER. D, HLA–B27 is thoughtto exist in multiple forms, and TCA/AMS analysis can enhance the detection of misfolded/alternatively oxidized forms within the ER andcompact/folded cell surface conformations. S � oxidized thiol; SH � reduced thiol (see Figure 1 for other definitions).
3422 FUSSELL ET AL

was more susceptible to reduction than B*2706 (Figure3A) (please contact the corresponding author for furtherdetails).
To determine whether band v was dependent onthe oxidizing environment, we exposed B27.H114D.D116Y to increasing diamide concentrations. However,band v was not enhanced but exhibited an apparentchange in molecular weight (Figure 3B, band v, B*2705,lanes 1–4 and B27.H114D, lanes 5–8), indicative of amore compact conformation, while this conformationremained unaffected by an increasingly reducing envi-ronment (Figure 3C, band v, lanes 2–4 and 6–8). In-creasing reducing conditions predominantly affectedband iv in the H114D mutant (Figure 3C, band iv,compare lanes 6, 7, and 8), while it appeared more resistantto reduction in the H114D.D116Y mutant (Figure 3C,lanes 11 and 12). Thus, the distinct conformer depicted byband v suggests an alternatively oxidized state that isrelatively resistant to reduction but prone to oxidation.
To determine the specificity of these conforma-tions, we tested other HLA products expressed endog-enously in human HeLa, CEM, CEM.NKR, and HEK293 cell lines or transfected into the C58 cell line (C58.B*0702). TCA/AMS analysis did not reveal conforma-tions similar to those detected for HLA–B27 (Figure3D), suggesting that these conformations could beunique to HLA–B27.
DISCUSSION
The role of HLA–B27 in AS remains elusive, butHLA–B27 misfolding and dimerization may be involved(12). Since heavy-chain dimers were originally described,it remains unclear how and why these structures form aswell as the extent to which they participate in ASdevelopment and pathogenesis.
We adopted a unique approach to probe the invivo structure of HLA–B27 to understand this tendencyto misfold. The current study used TCA/AMS for prob-ing in vivo conformations of class I MHC heavy chain.We have demonstrated that within the ER, HLA–B27heavy chain exists 1) as a fast migrating structure, 2) ina reduced and/or novel oxidized state, and 3) as apeptide-bound/compact conformation (Figure 2D). Pre-vious attempts to probe Cys accessibility revealed thatHLA–B27 can exist in at least 2 distinct conformationsfollowing cell surface expression (13), which can beaccounted for by partially folded/misfolded forms ofHLA–B27 (14). Our study identifies additional ER-resident HLA–B27 heavy-chain conformations with ex-posed Cys residues.
The accessibility of HLA–B27 to DTT-mediatedreduction may be the result of slow folding kinetics orbinding of low-affinity peptides (6,7). Susceptibility toreduction reflects exposed Cys residues and is analogous
Figure 3. Residues at p114 and p116 affect accessibility of HLA–B27cysteine residues to thiol modification. A, HLA–B*2705, B*2704,B*2706, B*2709, the 114 and 116 series of mutants, and D77S andV152E were subjected to TCA/AMS analysis (lanes 1–4). D116Y ledto the adoption of a more compact, less reducible form of heavy chain(band iv versus band vi). B*2709 (D116H) and D116Y substitutions ledto reduced detection and change in resolution of the conformationdepicted by band v. HLA–B*2704 was more susceptible to reductionthan HLA–B*2706. All mutants expressed band iii (although theHLA–B*2704 image does not show band iii; please contact thecorresponding author for further details). Note that the differences inband intensities between lanes are due to differential solubility. B andC, HLA–B*2705 (lanes 1–4), B27.H114D (lanes 5–8), and B27.H114D.D116Y (lanes 9–12) were incubated with increasing concentrations ofdiamide (B) and DTT (C). In B, increasing oxidative conditions did notinduce the alternative form (band v) but changed its conformation. InC, HLA–B*2705 and B27.H114D were more susceptible to reductionthan was B27.H114D.D116Y. band iii � novel rapidly migrating heavychain; band iv � compact folded heavy chain; band v � novel AMSdistinguishable form; band vi � reduced heavy chain. D, TCA/AMSanalysis of C58.B*0702 and HLA alleles expressed by HeLa, HEK 293,CEM, and CEM.NKR (NKR) cell lines was performed (lanes 1–4).Novel conformations detected for HLA–B27 appeared not to be sharedby other alleles. ut � untreated (see Figure 1 for other definitions).
HLA–B27 CONFORMATIONS AND ASSOCIATION WITH AS 3423

to our previous finding that poorly peptide-loaded classI MHC molecules were prone to ERp57-mediated re-duction (9). Resistance to reduction results from thecompact nature that class I MHC molecules adopt oncethey are fully folded, leaving Cys residues inaccessibleto further chemical modification. Such reduced accessi-bility to DTT by HLA–B27 heavy chain was observedfollowing residue changes at p116.
Misfolding models for HLA–B27 have been pro-posed (15); however, this is the first direct visualizationof misfolding HLA–B27 monomeric heavy chains in anin vivo setting. The TCA/AMS assay has revealed novelconformations associated with HLA–B27 biosynthesisand, significantly, has demonstrated that Cys residues ofHLA–B27 heavy chains are exposed to the oxidizingenvironment of the ER lumen. There appears to be acorrelation between novel HLA–B27 heavy-chain con-formations, folding rates, and heavy-chain dimer forma-tion (data not shown). Ultimately, Cys residue exposureto the ER environment and disulfide bond formationcould affect the extent to which misfolding moleculesparticipate in AS disease pathogenesis.
ACKNOWLEDGMENTS
We would like to thank Dr. Anthony Williams (CancerSciences Division, University of Southampton) and Dr. AdamBenham (Durham University) for helpful suggestions.
AUTHOR CONTRIBUTIONS
Dr. Antoniou had full access to all of the data in the study andtakes responsibility for the integrity of the data and the accuracy of thedata analysis.Study design. Powis, Antoniou.Acquisition of data. Fussell, Nesbeth, Lenart, Campbell, Lynch,Santos, Antoniou.Analysis and interpretation of data. Antoniou.Manuscript preparation. Gould, Powis, Antoniou.Statistical analysis. Fussell.Supply of essential materials. Gould.Generation of reagents. Antoniou.
REFERENCES
1. Brewerton DA, Hart FD, Nicholls A, Caffrey M, James DC,Sturrock, RD. Ankylosing spondylitis and HL-A 27. Lancet 1973;1:904–7.
2. Schlosstein L, Terasaki PI, Bluestone R, Pearson CM. Highassociation of an HL-A antigen, W27, with ankylosing spondylitis.N Engl J Med 1973;288:704–6.
3. Ramos M, Lopez de Castro JA. HLA-B27 and the pathogenesis ofspondyloarthritis. Tissue Antigens 2002;60:191–205.
4. Olivieri I, D’Angelo S, Scarano E, Santospirito V, Padula A. TheHLA–B*2709 subtype in a woman with early ankylosing spondyl-itis. Arthritis Rheum 2007;56:2805–7.
5. Cauli A, Vacca A, Mameli A, Passiu G, Fiorillo MT, Sorrentino R,et al. A Sardinian patient with ankylosing spondylitis andHLA–B*2709 co-occurring with HLA–B*1403. Arthritis Rheum2007;56:2807–9.
6. Williams AP, Peh CA, Purcell AW, McCluskey J, Elliott T.Optimization of the MHC class I peptide cargo is dependent ontapasin. Immunity 2002;16:509–20.
7. Antoniou AN, Ford S, Taurog JD, Butcher GW, Powis SJ.Formation of HLA-B27 homodimers and their relationship toassembly kinetics. J Biol Chem 2004;279:8895–902.
8. Huppa JB, Ploegh HL. The eS-Sence of -SH in the ER. Cell1998;92:145–8.
9. Antoniou AN, Ford S, Alphey M, Osborne A, Elliott T, Powis SJ.The oxidoreductase ERp57 efficiently reduces partially folded inpreference to fully folded MHC class I molecules. EMBO J 2002;21:2655–63.
10. Mezghrani A, Fassio A, Benham A, Simmen T, Braakman I, SitiaR. Manipulation of oxidative protein folding and PDI redox statein mammalian cells. EMBO J 2001;20:6288–96.
11. Tran TM, Horejsi V, Weinreich S, Pla M, Breur BS, Capkova J,et al. Strong association of HLA-B27 heavy chain with � 2-micro-globulin. Hum Immunol 2000;61:1197–201.
12. Kollnberger S, Bird LA, Roddis M, Hacquard-Bouder C,Kubagawa H, Bodmer HC, et al. HLA-B27 heavy chain ho-modimers are expressed in HLA-B27 transgenic rodent models ofspondyloarthritis and are ligands for paired Ig-like receptors.J Immunol 2004;173:1699–710.
13. Whelan MA, Archer JR. Chemical reactivity of an HLA-B27 thiolgroup. Eur J Immunol 1993;23:3278–85.
14. Santos SG, Antoniou AN, Sampaio P, Powis SJ, Arosa FA.Lack of tyrosine 320 impairs spontaneous endocytosis and en-hances release of HLA-B27 molecules. J Immunol 2006;176:2942–9.
15. Luthra-Guptasarma M, Singh B. HLA-B27 lacking associated�2-microglobulin rearranges to auto-display or cross-display resi-dues 169-181: a novel molecular mechanism for spondyloarthropa-thies. FEBS Lett 2004;575:1–8.
3424 FUSSELL ET AL





























