Embed Size (px)
DESCRIPTION
nelson journal of pelontology
Citation preview

Paleontological Society
Cenozoic Strombidae, Aporrhaidae, and Struthiolariidae (Gastropoda: Stromboidea) from Chile:Their Significance to the Interpretation of Southeast Pacific Biogeography and ClimateAuthor(s): Sven N. NielsenSource: Journal of Paleontology, Vol. 79, No. 6 (Nov., 2005), pp. 1120-1130Published by: Paleontological SocietyStable URL: http://www.jstor.org/stable/4094998 .Accessed: 16/02/2011 21:03
Your use of the JSTOR archive indicates your acceptance of JSTOR's Terms and Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp. JSTOR's Terms and Conditions of Use provides, in part, that unlessyou have obtained prior permission, you may not download an entire issue of a journal or multiple copies of articles, and youmay use content in the JSTOR archive only for your personal, non-commercial use.
Please contact the publisher regarding any further use of this work. Publisher contact information may be obtained at .http://www.jstor.org/action/showPublisher?publisherCode=paleo. .
Each copy of any part of a JSTOR transmission must contain the same copyright notice that appears on the screen or printedpage of such transmission.
JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
Paleontological Society is collaborating with JSTOR to digitize, preserve and extend access to Journal ofPaleontology.
http://www.jstor.org

J. Paleont., 79(6), 2005, pp. 1120-1130
Copyright ? 2005, The Paleontological Society 0022-3360/05/0079-1120$03.00
CENOZOIC STROMBIDAE, APORRHAIDAE, AND STRUTHIOLARIIDAE (GASTROPODA: STROMBOIDEA) FROM CHILE: THEIR SIGNIFICANCE TO
THE INTERPRETATION OF SOUTHEAST PACIFIC BIOGEOGRAPHY AND CLIMATE
SVEN N. NIELSEN
Geologisch-Paliontologisches Institut und Museum, Universitit Hamburg, Bundesstrasse 55, D-20146 Hamburg, Germany, and Freie Universitdit Berlin, Institut ftir geologische Wissenschaften, Fachrichtung Paliontolgie, Malteserstrasse 74-100,
D-12249 Berlin, Germany, <[email protected]>
ABSTRACT-The stromboid fauna from Cenozoic deposits of Chile contains an unusual mixture of warm-water and cool-water taxa, showing relationships with either Argentinian or New Zealand taxa. Six species of Strombidae, Aporrhaidae, and Struthiolariidae are described, and their relationships and paleobiogeographic implications are discussed. Conus medinae Philippi, 1887 from the Miocene Navidad Formation is assigned to the genus Strombus and thereby constitutes the first record of Strombidae from southwestern South America. The new subgenus Austrombus is proposed for smooth species of Strombus without any spines or axial sculpture. Hemichen- opus araucanus (Philippi, 1887) from the ?Eocene of Peninsula Arauco shows strong affinities to Dicroloma zelandica Marshall, 1919 from the Eocene of New Zealand, which is here assigned to Hemichenopus. The New Zealand species Perissoptera (Hemichenopus) thomsoni Allan, 1926 does not belong in Hemichenopus but could not be reassigned to another aporrhaid genus. Struthiochenopus bandeli n. sp. and Struthiochenopus philippii Zinsmeister and Griffin, 1995 are reported from deepwater facies in Chile, rather than the nearshore facies favored by congeners in Argentina. Perissodonta ameghinoi (Ihering, 1897) and Perissodonta chilensis (Philippi, 1887) are the only species of this genus in the eastern Pacific, the former being an Atlantic species, reported for the first time from the Pacific. The term Neo-Weddellian is proposed for Miocene taxa having their origin in the Late Cretaceous-Paleogene Weddellian Province.
INTRODUCTION
ALL FOUR larger Recent families of Stromboidea are represented in Cenozoic strata from Chile. Species of Strombidae, Apor-
rhaidae, and Struthiolariidae are presented in this work; the Xeno- phoridae have been discussed recently by Nielsen and DeVries (2002). Strombidae have never been reported from southwestern South America, but Aporrhaidae and Struthiolariidae are known since Philippi (1887) described Chenopus araucanus and Struthio- laria chilensis from Chile. In this same work Philippi (1887) de- scribed Conus medinae, which is here transferred to the genus Strom- bus Linnaeus, 1758 and comprises the only record of Strombidae from Chile. Later, one aporrhaid, Struthiochenopus philippii Zins- meister and Griffin, 1995, and two struthiolariids, Perissodonta ameghinoi (Ihering, 1897) (Frassinetti and Covacevich, 1999) and P. hatcheri (Ortmann, 1902), were added to the Chilean fauna. Peris- sodonta ameghinoi was described originally from Argentina. The Chilean records of both struthiolariids are from Atlantic sediments to the east of the Andes near Chile Chico (Frassinetti and Covacev- ich, 1999) and Punta Arenas (Ortmann, 1902).
In Argentina, several Cenozoic aporrhaid (Zinsmeister and Griffin, 1995) and struthiolariid (Zinsmeister and Camacho, 1980) taxa are present. Strombidae and Xenophoridae have not been recorded from Cenozoic faunas. Strombus bravardi Borchert, 1901 was described from Miocene deposits of the Paranai For- mation. The holotype, however, is an internal mold (del Rio, 1998) and cannot be attributed reliably to any gastropod genus. In Peru, only Aporrhaidae are known from the Eocene to Oli- gocene of northern Peru (d'Orbigny, 1842; Woods, 1922; Olsson, 1928, 1931) and southern Peru (DeVries, 2004), and reported taxa belong to genera not known from Chile.
GEOLOGY OF FOSSIL-BEARING LOCALITIES
Navidad Formation, Central Chile.-The Navidad Formation (Darwin, 1846) is known for a rich molluscan fauna that was last reviewed extensively by Philippi (1887). Paleodepths, as indicated by environmental ranges of foraminifera and ostracodes, are from intertidal to outer shelf (Finger et al., 2003). Specimens of Stru- thiochenopus philippii have been recovered together with Xeno- phora paulinae Nielsen and DeVries, 2002 from grey deepwater
siltstone that today forms the intertidal platform at Punta Perro (PPP; Fig. 1.1) and is the type locality for both species. These deposits have been dated with foraminifers (Dremel in Herm, 1969, p. 71) as Early Miocene (Burdigalian), and correlation us- ing mollusks from well-dated sections in Peru indicates latest O1- igocene to early middle Miocene ages for typical Navidad mol- lusks in Chile (DeVries and Frassinetti, 2003). Tsuchi et al. (1990) and Ibaraki (1992), however, have pointed out that foraminiferal assemblages suggest a Late Miocene (Tortonian) age. These con- tradictory age data can be explained by the reworking and mixing of Early Miocene foraminifera and mollusks into the Late Mio- cene deposits of the Navidad Formation as shown by Finger et al. (2003), who also obtained a Late Miocene age. The possible reworking of mollusks encountered in the Navidad Formation has been discussed by Nielsen et al. (2003).
About I km north of the village of Matanzas a small fossiliferous sandlens (MAT; Fig. 1.1) contained a rich fauna, including Peris- sodonta chilensis (Philippi, 1887) and Xenophora paulinae. Addi- tional specimens of P. chilensis come from the intertidal platform near this locality (MAP; Frassinetti and Covacevich, 1993) where the neotype of Conus medinae also has been found. These localities have also been dated as Late Miocene (Finger et al., 2003).
Ranquil Formation, Peninsula Arauco, south-central Chile--Light- grey siltstones at several localities along the coast of Peninsula Arauco (FRM, RQT, MIB; Fig. 1.2) yield a molluscan fauna including genera like Dalium Dall, 1889, Exilia Conrad, 1860, and Borsonia Bel- lardi, 1839, indicating deeper water environments. These silt- stones are sometimes referred to the Eocene Millongue Formation (see Garcia, 1968), which, however, has a different lithology and faunal content (personal observations). A concordant boundary between the siltstone at Punta El Fraile (FRM) and an overlying brown sandstone typical of the Miocene Ranquil Formation, to- gether with the faunal composition of the siltstone, which includes a new Struthiochenopus Zinsmeister and Griffin, 1995 described in this paper, indicate a Late Miocene age for the siltstone. The
geological situation at Ranquil (RQT, MIB) is not as clear, but a Late Miocene age for the lithologic units at Punta El Fraile and Ranquil was confirmed using planktonic foraminifera (Finger et al., 2003).
1120
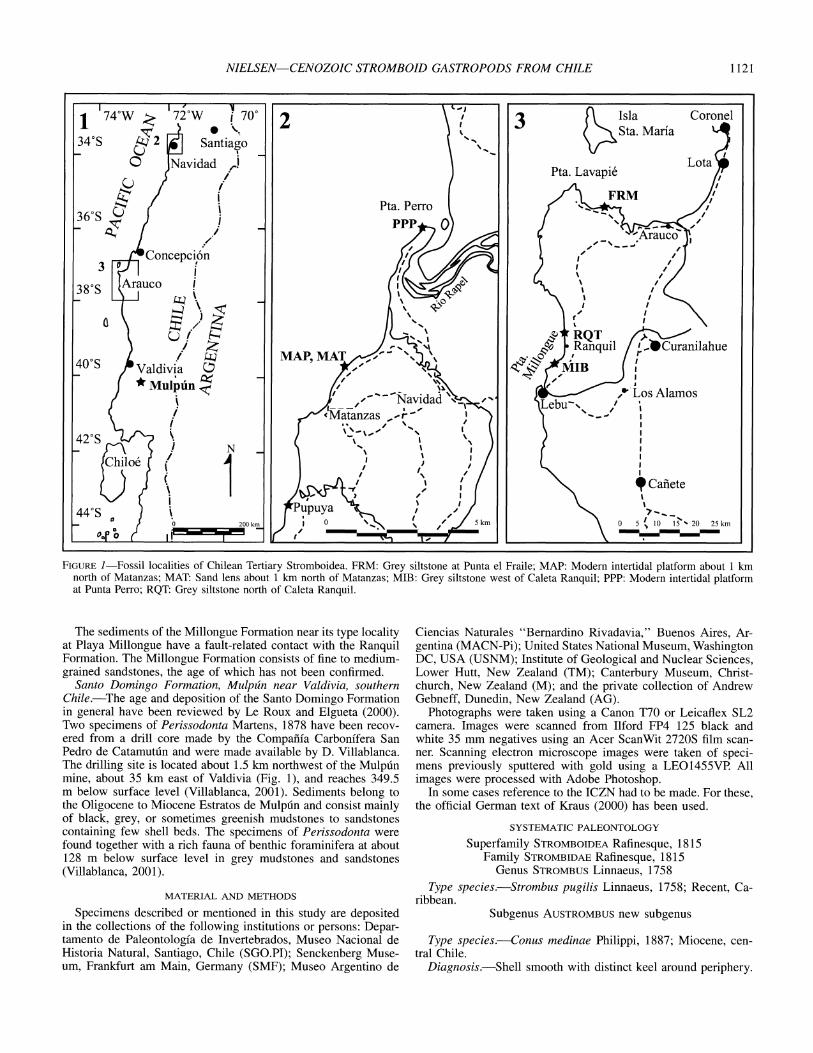
NIELSEN-CENOZOIC STROMBOID GASTROPODS FROM CHILE 1121
74W 72W 70 2 3 Isla Coronel
_ .
Sta. Maria 340S 2 Santiago S,
O Navidad , PtaLavaiLota Pta. Lavapi6 l
FRM
3s
Pta. Perro 360S PPP
/ *'Arauco
Concepci6n
384S Arauco I 380S
c f RQT 7 Ranquil *Curanilahue / k MAP, MAT
40S Valdivia MIB
a Mul*O'"- Lb Los Alamos a
I-or-"Lebu- / I Matanzas -r oro I
420S "
Chilo6
Ibi Cafiete
44<S Pupuya
1
I
2
0200 km 0. 5km 0 5I 10 15 20 25 km
o'er i\'
FIGURE 1-Fossil localities of Chilean Tertiary Stromboidea. FRM: Grey siltstone at Punta el Fraile; MAP: Modern intertidal platform about 1 km north of Matanzas; MAT: Sand lens about 1 km north of Matanzas; MIB: Grey siltstone west of Caleta Ranquil; PPP: Modern intertidal platform at Punta Perro; RQT: Grey siltstone north of Caleta Ranquil.
The sediments of the Millongue Formation near its type locality at Playa Millongue have a fault-related contact with the Ranquil Formation. The Millongue Formation consists of fine to medium- grained sandstones, the age of which has not been confirmed.
Santo Domingo Formation, Mulpuin near Valdivia, southern Chile.--The age and deposition of the Santo Domingo Formation in general have been reviewed by Le Roux and Elgueta (2000). Two specimens of Perissodonta Martens, 1878 have been recov- ered from a drill core made by the Compafiia Carbonffera San Pedro de Catamut in and were made available by D. Villablanca. The drilling site is located about 1.5 km northwest of the Mulpt6n mine, about 35 km east of Valdivia (Fig. 1), and reaches 349.5 m below surface level (Villablanca, 2001). Sediments belong to the Oligocene to Miocene Estratos de
Mulptin and consist mainly
of black, grey, or sometimes greenish mudstones to sandstones containing few shell beds. The specimens of Perissodonta were found together with a rich fauna of benthic foraminifera at about 128 m below surface level in grey mudstones and sandstones (Villablanca, 2001).
MATERIAL AND METHODS
Specimens described or mentioned in this study are deposited in the collections of the following institutions or persons: Depar- tamento de Paleontologia de Invertebrados, Museo Nacional de Historia Natural, Santiago, Chile (SGO.PI); Senckenberg Muse- um, Frankfurt am Main, Germany (SMF); Museo Argentino de
Ciencias Naturales "Bernardino Rivadavia," Buenos Aires, Ar- gentina (MACN-Pi); United States National Museum, Washington DC, USA (USNM); Institute of Geological and Nuclear Sciences, Lower Hutt, New Zealand (TM); Canterbury Museum, Christ- church, New Zealand (M); and the private collection of Andrew Gebneff, Dunedin, New Zealand (AG).
Photographs were taken using a Canon T70 or Leicaflex SL2 camera. Images were scanned from Ilford FP4 125 black and white 35 mm negatives using an Acer ScanWit 2720S film scan- ner. Scanning electron microscope images were taken of speci- mens previously sputtered with gold using a LE01455VP. All images were processed with Adobe Photoshop.
In some cases reference to the ICZN had to be made. For these, the official German text of Kraus (2000) has been used.
SYSTEMATIC PALEONTOLOGY
Superfamily STROMBOIDEA Rafinesque, 1815 Family STROMBIDAE Rafinesque, 1815
Genus STROMBUS Linnaeus, 1758
Type species.-Strombus pugilis Linnaeus, 1758; Recent, Ca- ribbean.
Subgenus AUSTROMBUS new subgenus
Type species.-Conus medinae Philippi, 1887; Miocene, cen- tral Chile.
Diagnosis.-Shell smooth with distinct keel around periphery.

1122 JOURNAL OF PALEONTOLOGY, V. 79, NO. 6, 2005
4
1 2 3 5
6 "9
r~|
'
b
P
6 7 8 9L
FIGURE 2-Strombus Linnaeus, 1758 and Hemichenopus Steinmann and Wilckens, 1908. 1-3, Strombus (Austrombus) medinae (Philippi, 1887), late Miocene, central Chile. 1, Lost holotype (from Philippi, 1887), height 90 mm. 2, 3, Neotype SGO.PI.5964, height 73.7 mm. 4, 5, 9, Hemichenopus araucanus (Philippi, 1887), ?Eocene, south-central Chile. 4, Syntype SGO.PI.4504, height 19 mm. 5, Syntype SGO.PI.4504 (on backside of sample). 9, Syntype SGO.PI.763, height 19 mm. 6-8, Hemichenopus zelandicus (Marshall, 1919), Middle Eocene, New Zealand. Hypotype TM 5131, height 25 mm.
Faint spiral threads on last whorl, coarsening anteriorly. Axial sculpture absent. Columella smooth.
Etymology.--Combination of australis, Latin for southern, and the genus Strombus.
Discussion.-Relations of Austrombus are unclear. According to Abbott (1960), species of Strombus are first known from the Miocene in the "southern Tethys realm" (southern Europe, Ca- ribbean, Indo-Pacific). Abbott also listed some Eocene species, which he supposed not to be closely related to Recent species of this genus. Comparison with several Neogene to Recent faunas (e.g., Woodring, 1957-1982; Abbott, 1960; Jung, 1965; Keen, 1971; Beu and Maxwell, 1990; Jung and Heitz, 2001) did not reveal any possible relationship for S. (A.) medinae. It does re- semble the extant S. (S.) gracilior Sowerby, 1825 and S. (Tricor- nis) tricornis Lightfoot, 1786 in general shell shape, although both Strombus and Tricornis Jousseaume, 1886 are characterized by a single row of shoulder spines or knobs (Abbott, 1960). The Ar- gentinian Miocene Strombus bravardi, despite being represented by a poorly preserved internal mold, clearly shows a row of.spines or nodes. If Strombus bravardi belongs in Strombus it is most probably related to species of Strombus sensu stricto from the Caribbean. The subgenus Laevistrombus Abbott, 1960 (Laevis- trombus Kira, 1955 is a nomen nudum, see Bieler and Petit, 1996)
has no spines but weak axial swellings are present. The type spe- cies, Strombus (Laevistrombus) canarium Linnaeus, 1758, is the only living species of the subgenus and occurs from southern India to Australia and Japan. Laevistrombus, however, has a dif- ferent shell form with a smooth rounded shoulder and varices on early whorls, characters not known for Austrombus. Abbott (1960) pointed out that the subgenera Strombus and Laevistrom- bus might be closely related. If that is true, Austrombus may be closely related to a common ancestor of those subgenera.
Species of the subgenus Canarium Schumacher, 1817 have ax- ial ribs or knobs, and may have a smooth last whorl. This sub- genus today reaches Easter Island and thus has the easternmost Pacific occurrence apart from Central American Strombus. Can- arium can be traced back to the Miocene of Indonesia (Abbott, 1960) and contains no representative similar to Strombus (Aus- trombus) medinae.
STROMBUS (AUSTROMBUS) MEDINAE (Philippi, 1887) new combination
Figure 2.1-2.3
Conus medinae PHILIPPI, 1887, p. 36, pl. 1, fig. 6.
Diagnosis.-As for subgenus.

NIELSEN-CENOZOIC STROMBOID GASTROPODS FROM CHILE 1123
Description.--Shell height at least up to 90 mm, with moder- ately high spire of about five whorls; protoconch of about three whorls, ornamentation unknown; protoconch and early teleoconch whorls convex; whorl profile of later whorls concave above and slightly concave below sharp, keel-like shoulder, constricted an- teriorly; surface of teleoconch with faint spiral threads coarsening anteriorly; aperture narrow, outer lip unknown; whorls smooth except for single keel-like angulation around shoulder; no strom- boid notch visible in growth increments; siphonal canal short, constricted, slightly bent backwards.
Material examined.-The holotype (Fig. 2.1) is not preserved in the Philippi collection at the SGO.PI and must be considered lost. The only available specimen, SGO.PI.5964 (Matanzas, height 73.7 mm), is well preserved and is here selected as the neotype to clarify taxonomic status and type locality. Philippi (1887, p. 36) gave the locality of the holotype as 'found in Na- vidad by D. J. Toribio Medina.' The type locality is here inter- preted as being the Navidad Formation around Navidad.
Occurrence.-Intertidal platform about 1 km north of Matan- zas, Navidad Formation of Late Miocene age, central Chile.
Discussion.--Strombus (Austrombus) medinae is easily sepa- rated from any other known species of Strombus by the total absence of spines and axial ornamentation. Philippi (1887) had seen only one specimen (the holotype) on which he based his new species, but this was not his own. The measurements of the ho- lotype as given by Philippi (1887, p. 36) are height 90 mm, width 47 mm. It is not present in his collection and must be considered as lost. The neotype designated herein was found by W. J. Zins- meister (Department of Earth and Atmospheric Sciences, Purdue University, Indiana, USA) about 1 km north of Matanzas. It comes from within the area of 'Navidad' (ICZN Article 75.3.6), the locality given by Philippi (1887) for the holotype of Conus medinae, and it is therefore the type locality according to the ICZN (Article 76.3).
Family APORRHAIDAE Philippi, 1836 Genus HEMICHENOPUS Steinmann and Wilckens, 1908
Type species.--Chenopus araucanus Philippi, 1887 by original designation;? Eocene, central Chile.
Other species.-Dicroloma zelandica Marshall, 1919 from the Middle to Late Eocene of New Zealand is here newly included. No other species are confirmed to belong to Hemichenopus.
HEMICHENOPUS ARAUCANUS (Philippi, 1887) Figure 2.4, 2.5, 2.9
Chenopus araucanus PHILLIPPI, 1887, p. 35, pl. 1, fig. 1.
Diagnosis.-Shell high-spired, with strongly keeled whorls; two prominent keels on whorl with two minor cords below; outer lip of aperture with two prominent digitations; posterior digitation directed away from and slightly behind aperture, anterior digita- tion directed forwards.
Description.-Shell about 20 mm high, with high turreted spire of at least six whorls; protoconch and initial whorls unknown; whorls with straight profile above and below prominent keel; sur- face sculpture mostly unknown; second subequal keel on last whorl; at least two further weaker cords on base of last whorl; aperture unknown, leading into siphonal canal; outer lip of ap- erture expanded into wing, with broad deep arcuate sinus poste- riorly, bearing two very prominent digitations about equal in length and width, corresponding in position to keels on exterior of teleoconch; posterior digitation directed away from and slightly behind aperture, anterior digitation directed forwards; digitations in one plane at 450 to spire axis; interior surface of wing with two deep narrow canals extending into digitations; outer margins of wing thickened; inner lip of aperture unknown.
Material examined.-Syntypes SGO.PI.763 (height 19 mm), SGO.PI.4500, 4501, 4502, 4503, 4504 (height 19 mm, additional wing on backside), 4505, 4506, 4507, 4508 (all probably from near Lebu). The specimen SGO.PI.763 is labelled as 'lectotype' but has never been published as such. According to the ICZN (Article 74.7.3), formal designation of a lectotype is not justified, because all syntypes are conspecific. There is no other material known.
Occurrence.-The age of this species remains unclear. Eocene, Miocene, and Pliocene sediments are found on Peninsula Arauco, the area of the type locality, and no new material has been found. The well-lithified sediment associated with the type specimens, however, suggests they came from Eocene beds.
Discussion.-A wing with two digitations distinguishes Hem- ichenopus araucanus from all other Chilean and Peruvian apor- rhaids. Steinmann and Wilckens (1908) thought Philippi's (1887) composite figure to be inaccurate and assigned their own material, which have only a blunt anterior digitation, to Hemichenopus ar- aucanus. This material is now known to belong to Struthiochen- opus magellanicus (Ihering, 1909) (Zinsmeister and Griffin, 1995). Zinsmeister and Griffin (1995, p. 698) further stated that "it should also be noted that Philippi's illustration (1887, pl. 1, fig. 1) is a composite reconstruction based on 17 fragments, and it is now well known that some of Philippi's illustrations are in- accurate representations of the actual material." While it is cer- tainly true that Philippi's figure is a composite drawing, the type material definitely allows such a reconstruction. Well-preserved wings present in Philippi's material (Fig. 2.5) clearly show the two prominent digitations.
HEMICHENOPUS ZELANDICUS (Marshall, 1919) new combination Figure 2.6-2.8
Dicroloma zelandica MARSHALL, 1919, p. 228-229, pl. 15, fig. 16; MAR- SHALL AND MURDOCH, 1920, p. 130, pl. 7, fig. 13; FLEMING, 1966, p. 51.
Dicroloma (sensu lato) zelandica BEU AND MAXWELL, 1990, p. 104, 407, pl. 6g.
Dicroloma (s.1.) zelandica MAXWELL, 1992, p. 91, pl. 9h, i.
Description.-Shell reaching 30 mm in height, with five to six spire-whorls; protoconch not definitely distinguishable from te- leoconch, consisting of four smooth, convex whorls. Spire-whorls strongly and evenly convex at first, developing a weak median angulation on fifth whorl. Penultimate whorl with sharply defined peribasal angulation, base gently convex. Spiral sculpture com- mencing as seven or eight fine, rounded, subequal cords, with interstitial threads. One cord becoming stronger than others on fifth whorl, forming median angulation, eventually becoming strong posterior keel on last whorl. Peribasal angulation marked by comparatively strong cord on last whorl of subadult shells, becoming anterior keel on adult shells. Base of last whorl with about eight low cords with some interstitial threads. Axial sculp- ture consisting of weak costellae, reaching from suture to suture on spire whorls, but stopping abruptly at peribasal angulation on penultimate whorl and becoming obsolete on last whorl. Aperture moderately large, columella vertical, almost straight, but pro- nounced anteriorly in adults to form a long, narrow, open siphonal canal curving slightly to left in apertural view. Inner lip with thin callus glaze in young shells, moderately callused in adults. Outer lip thickened, bearing two prominent digitations corresponding to keels. Posterior digitation directed away from, and slightly behind aperture, anterior digitation directed forwards. Both digitations similar in length to siphonal canal and bearing a shallow longi- tudinal groove along inner face. (For a more detailed description see Maxwell, 1992, p. 91).

1124 JOURNAL OF PALEONTOLOGY, V. 79, NO. 6, 2005
Types.--Holotype TM 5130 (not seen, height 28 mm), hypo- type TM 5131 (height 25 mm; Marshall and Murdoch, 1920, pl. 7, fig. 13; Maxwell, 1992, pl. 9i).
Discussion.-Dicroloma zelandica from the Bortonian (Middle Eocene) and Kaiatan (Late Eocene) of New Zealand resembles Hemichenopus araucanus and also has two similar digitations. It is therefore assigned to Hemichenopus. Beu and Maxwell (1990) and Maxwell (1992) already noted that its placement in Dicro- loma Gabb, 1868, a Mesozoic genus with a Jurassic type species, was not satisfactory, but left it in the genus because they could not find a more suitable alternative. Hemichenopus zelandicus dif- fers from H. araucanus chiefly in its less slender spire, in having more cords below the keels, and in the form of the outer lip digitations.
Hemichenopus has its outer lip expanded into a wing with two well-developed digitations; in this it resembles the Weddellian genus Austroaporrhais Zinsmeister and Griffin, 1995. Species of Austroaporrhais have "wavy nodose, axial ribs" while H. zelan- dicus has only relatively weak axial ornament. The spiral sculp- ture of H. araucanus, type species of Hemichenopus, is not known, so that distinguishing the two genera on the basis of the presence or absence of spiral sculpture with more or less strong nodes must remain provisional.
"HEMICHENOPUS" THOMSONI (Allan, 1926) Figure 3.1-3.4
Perissoptera (Hemichenopus) thomsoni ALLAN, 1926, p. 338, pl. 76, fig. 1. Hemichenopus thomsoni BEU AND MAXWELL, 1990, p. 407.
Description.--Shell up to 32 mm high, with high turreted spire of approximately eight whorls; spire angle about 550; protoconch worn; initial whorls convex, with a faint keel that becomes pro- gressively stronger on succeeding whorls, becoming obsolete on last two to three whorls; whorl profile slightly concave above and gently convex below keel or shoulder; surface of shell orna- mented with numerous spiral threads; very faint axial ribs visible only on initial whorls of teleoconch, but rapidly becoming ob- solete, appearing as faint nodes on keel, not present on last whorls; aperture elongated, subtriangular, with narrow, straight, siphonal anterior canal; outer lip simple in subadults, not known from adults. Inner lip covered by moderately thick callus pad, with conspicuous ridge along posterior basal edge.
Types.--Holotype Canterbury Museum, New Zealand, M243 (height 25.9 mm).
Other material examined.--One specimen, coll. Andrew Greb- neff AG1604 (height 31.53 mm).
Occurrence.-Waihao Downs, Waihao River, near Waimate (holotype, Allan, 1926) and silts of Evans Crossing section, Par- eora River, near Timaru (AG1604, Andrew Grebneff, personal commun.). Bortonian (Middle Eocene), southeast South Island, New Zealand.
Discussion.-Perissoptera (Hemichenopus) thomsoni has been included in Hemichenopus on the basis of the description of Stein- mann and Wilckens (1908) which, however, was based on a spe- cies of Struthiochenopus. The single spire described by Allan (1926) (Fig. 3.1, 3.2) indeed resembles species of Struthiochen- opus rather than Hemichenopus. However, it differs from species
of both genera in not having a second keel, the primary keel becoming obsolete on later whorls, whorl attachment slightly be- low primary keel, stronger spiral sculpture, and a subtriangular aperture. Hemichenopus thomsoni is similar to the middle Paleo- cene Struthioptera osiris Finlay and Marwick, 1937, from Wan- galoa, New Zealand, which is the least axially sculptured species of that genus. It differs from that species, and the whole genus, by the absence of a second keel. The most complete specimen available (Fig. 3.3, 3.4) is located in the private collection of A. Grebneff (AG1604) and lacks a protoconch and outer lip. It is very similar to some turrids, although Beu and Maxwell (1990) left it within Hemichenopus. Hemichenopus thomsoni does not belong in the genus Hemichenopus. However, at the moment it is not possible to reallocate H. thomsoni to any other aporrhaid ge- nus, nor can it be reliably attributed to that family. If it is an aporrhaid, the collection of adult specimens is necessary to clarify the generic position.
Genus STRUTHIOCHENOPUS Zinsmeister and Griffin, 1995
Type species.-Struthiochenopus magellanicus, designated herein (see discussion below); late Oligocene to early Miocene, Tierra del Fuego, Argentina.
Other species.-The previously known species have been de- scribed by Zinsmeister and Griffin (1995). Struthiochenopus ban- deli n. sp. from the Miocene of south-central Chile is added here.
Discussion.-Zinsmeister and Griffin (1995) named Perissop- tera nordenskjoldi Steinmann and Wilckens, 1908 as the type spe- cies of their new genus Struthiochenopus. However, Perissoptera nordenskjoldi was described by Wilckens only in 1910 (Wilckens, 1910). Also, in the definition of Struthiochenopus, Zinsmeister and Griffin (1995, p. 698) stated that they "propose the new genus Struthiochenopus for the species originally included in Hemi- chenopus araucanus by Steinmann and Wilckens from Tierra del Fuego;" which in fact is Struthiochenopus magellanicus (see Zinsmeister and Griffin, 1995, p. 699) and which therefore is recognized here as the type species. The name Dicroloma ma- gellanica Ihering, 1909 was introduced as a new name for Chen- opus araucanus Steinmann and Wilckens, 1908 (not of Philippi, 1887).
STRUTHIOCHENOPUS BANDELI new species Figure 3.5-3.10
Diagnosis.--Shell small, about 35 mm high, with about nine strongly keeled whorls; prosocline axial ribs along keel; outer lip of aperture with prominent posterior digitation; spire angle about
450. Description.---Shell up to 35 mm high, with high turreted spire
of approximately nine whorls; spire angle about 450; protoconch of 2.5 depressed whorls; initial teleoconch whorls convex, with a faint keel that becomes progressively stronger on succeeding whorls; second subequal keel present on last whorl; whorl profile flat above and gently concave below the keel; surface of teleo- conch ornamented with very fine spiral threads; very faint axial ribs visible only on initial teleoconch whorls, but rapidly becom- ing obsolete, appearing again as faint nodes on primary keel of last whorl; aperture elongated, sublenticular, with narrow, straight,
FIGURE 3-Aporrhaidae Philippi, 1836. 1-4, "Hemichenopus" thomsoni (Allan, 1926), Middle Eocene, New Zealand. 1, 2, Holotype Canterbury Museum M243, height 25.9 mm. 3, 4, Almost complete specimen, coll. Grebneff AG1604, height 31.53 mm, diameter 16.90 mm, Middle Eocene silts of Evans Crossing section, Pareora River, near Timaru, southeast South Island, New Zealand. 5-10, Struthiochenopus bandeli n. sp., late Miocene, south-central Chile. 5, 6, Paratype SMF 323628, height 22.5 mm. 7, SGO.PI.6076, height of sample 16 mm, width 23.5 mm. 8, Holotype SGO.PI.6074, height 30 mm. 9, 10, Paratype SGO.PI.6075. 11-14, Struthiochenopus philippii Zinsmeister and Griffin, 1995, late Miocene, central Chile. 11, 12, SMF 323630, height 20.5 mm. 13, 14, SGO.PI.6078.

NIELSEN-CENOZOIC STROMBOID GASTROPODS FROM CHILE 1125
400g l
1 2 3 4
5 6 7
9 10
8
11 12 13 400gm- 14

1126 JOURNAL OF PALEONTOLOGY, V. 79, NO. 6, 2005
anterior siphonal canal; outer lip expanded into subquadrate wing with prominent posterior digitation, anterior digitation reduced to blunt angulation; posterior margin of wing widely sinused; pos- terior keel extends along length of posterior digitation; inner sur- face of wing with deep narrow groove extending along digitation, inner lip covered by moderately thick callus pad, with conspicu- ous ridge along posterior basal edge.
Etymology.-Named after Klaus Bandel, who took part in sev- eral field trips where this material was collected and who guided me during revision of the Miocene gastropods from Chile.
Types.-Holotype SGO.PI.6074 (height 30 mm), paratypes SMF 323628 (height 22.5 mm), SGO.PI.6075, all from Punta El Fraile.
Other material examined.-SMF 323629 (11 fragments), SGO.PI.6076 (sample with four juvenile specimens, height 16 mm, width 23.5 mm), SGO.PI.6077, all from Punta El Fraile.
Occurrence.-Ranquil Formation of Late Miocene age, Punta El Fraile and Caleta Ranquil, Arauco, south-central Chile.
Discussion.-This species very much resembles Struthiochen- opus magellanicus, but its wider spire angle, which leads to a slightly broader shell form, as well as its whorl profile, which is more flattened above the keel and has a more prominent second keel, differentiate it from the Argentinian species. Since S. ma- gellanicus has a Late Oligocene to Early Miocene age, while S. bandeli is supposedly Late Miocene, it might be concluded that the latter arose from the former species. Struthiochenopus bandeli apparently comes from relatively deepwater facies, in contrast with the very similar Argentinian S. magellanicus, which is found in shallow-water facies. This ecological difference might turn out to be an additional character to separate these two species.
STRUTHIOCHENOPUS PHILIPPII Zinsmeister and Griffin, 1995 Figure 3.11-3.14
Hemychenopus araucanus (sic) TAVERA, 1979, p. 98, pl. 18, fig. 58 (not of PHILIPPI, 1887).
Struthiochenopus philippii ZINSMEISTER AND GRIFFIN, 1995, p. 698, fig. 3.2-3.5, 3.7.
Description.-Shell up to 32 mm high, with high turreted spire of up to 10 whorls; spire angle about 300; protoconch of 3.5 moderately depressed, smooth, convex whorls; keel development beginning on fourth whorl, second keel anterior to primary keel visible on last whorl; whorl profile nearly flat above and slightly convex below keel; surface of teleoconch ornamented with fine spiral threads; approximately 28 very faint axial ribs on early whorls, becoming restricted to keel region giving keel a nodose appearance on later whorls; aperture sublenticular, with narrow, straight anterior siphonal canal; outer lip expanded into broad wing with prominent posterior digitation, anterior digitation re- duced to blunt angulation; posterior margin of wing widely sin- used; inner lip covered by moderately thick callus pad, with con- spicuous ridge along posterior basal edge.
Types.-Holotype USNM 405855; paratypes USNM 405858 and USNM 405859 (all not seen).
Other material examined.~-SGO.PI.5307 (seven specimens, Pupuya), SGO.PI.5331 (23 specimens and one wing, Punta Alta); SMF 323630 (height 20.5 mm), SGO.PI.6078.
Occurrence.-Navidad Formation of Late Miocene age, Punta Perro and Punta Alta, Navidad area, central Chile.
Discussion.~-Zinsmeister and Griffin (1995) noted that this species is the only species of Struthiochenopus from a deepwater siltstone facies. However, the newly described S. bandeli n. sp. also comes from a deepwater siltstone, in contrast with all Ar- gentinian species, which are found in shallow-water facies (Zins- meister and Griffin, 1995). There are no aporrhaid species present
in the many nearshore deposits known along the modern Chilean coast.
Zinsmeister and Griffin (1995, p. 699) included their figure 3.7 under S. magellanicus, whereas the specimen clearly belongs to S. philippii as is correctly indicated under that species and in the figure captions.
Family STRUTHIOLARIIDAE Fischer, 1884 (in 1880-1887) Genus PERISSODONTA Martens, 1878
Type species.-Struthiolaria mirabilis Smith, 1875 by mono- typy; Recent, Kerguelen Islands.
Discussion.-Powell (1951) showed that Struthiolarella Stein- mann and Wilckens, 1908 is a synonym of Perissodonta because both 'genera' intergrade with each other in size and sculpture. He also commented on the fact, that earlier workers (Marwick, 1924; Finlay and Marwick, 1937) were not aware of Martens's name and consequently included several New Zealand species and also Struthiolaria mirabilis in Struthiolarella. However, Zinsmeister and Camacho (1980) later separated Struthiolarella again from Perissodonta on the basis of sculpture and protoconch morphol- ogy.
PERISSODONTA AMEGHINOI (Ihering, 1897) new combination Figure 4.1-4.3
Struthiolaria ameghinoi IHERING, 1897, p. 289, fig. 14; ORTMANN, 1902, p. 201, pl. 33, fig. ila (in part); PARODIZ, 1996, p. 201.
Struthiolarella ameghinoi STEINMANN AND WILCKENS, 1908, p. 53, pl. 6, fig. 7; ZINSMEISTER AND CAMACHO, 1980, text-fig. 3g-3i; FRASSI- NETTI AND COVACEVICH, 1999, p. 47, pl. 9, figs. 15, 16.
Struthiolarella cf. chilensis VILLABLANCA, 2001, p. 47, pl. 3, fig. 4.
Description.-Shell large with elevated spire consisting of six whorls; whorls shouldered; central portion of whorl slightly de- pressed anterior to shoulder; impressed suture; ornamentation consisting of elongated tubercles, approximately 12 per whorl; spiral ornamentation consisting of eight or nine narrow cords over axial ribs, six paired wider cords, and three or four narrow basal ones anterior to tubercles. Growth lines strongly sinusoidal, pos- terior sinus wide and squarish; prominent raised growth lines ir- regularly spaced. Columella sinusoidal, columellar callus thin.
Types.-There are 12 syntypes under the number #763 in the Ihering collection housed in the MACN-Pi (Parodiz, 1996, not seen).
Other material examined.-SMF 323631 (two specimens, height 21 mm and 17 mm).
Occurrence.-Mulp6n near Valdivia, Santo Domingo Forma- tion of Oligocene to Miocene age, southern Chile to Santa Cruz, Patagonia, Argentina.
Discussion.-Perissodonta ameghinoi is the type species of the genus Struthiolarella. Its assignment to Perissodonta confirms the synonymy of both genera as pointed out by Powell (1951).
Perissodonta ameghinoi has been reported from the Late Oli- gocene to Early Miocene of Ais6n (Frassinetti and Covacevich, 1999) and Punta Arenas (Ortmann, 1902) but revision of these Chilean faunas is beyond the scope of this work. These faunas are more similar to faunas from Argentina than to those from Pacific Chile because they have an Atlantic origin. Perissodonta ameghinoi was originally described from Santa Cruz, Argentina, and is among the very few gastropod species occurring on both sides of the Andes.
PERISSODONTA CHILENSIS (Philippi, 1887) new combination Figure 4.4-4.8
Struthiolaria chilensis PHILIPPI, 1887, p. 36, pl. 1, fig. 4. Struthiolarella chilensis COVACEVICH, FRASSINETTI, AND ALFARO, 1992,
p. 148, 150, pl. 1, figs. 3a-c, 4.
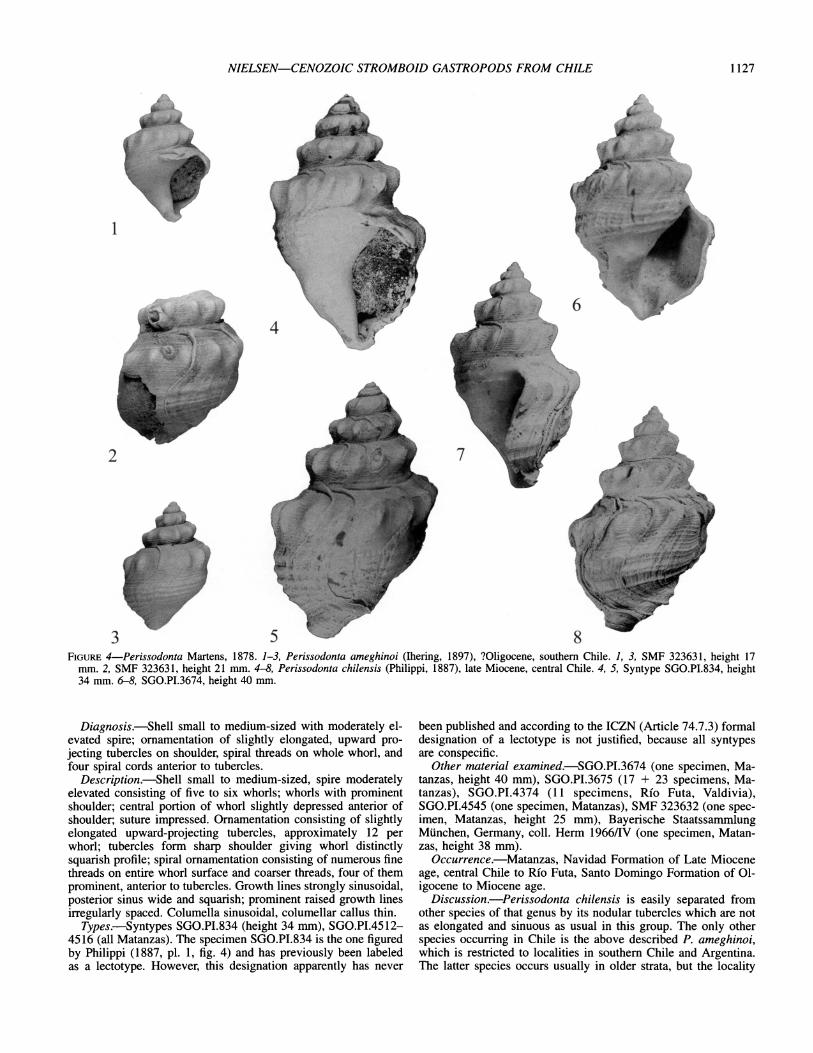
NIELSEN-CENOZOIC STROMBOID GASTROPODS FROM CHILE 1127
1
6 4
2 7
3 5 8 FIGURE 4-Perissodonta Martens, 1878. 1-3, Perissodonta ameghinoi (Ihering, 1897), ?Oligocene, southern Chile. 1, 3, SMF 323631, height 17
mm. 2, SMF 323631, height 21 mm. 4-8, Perissodonta chilensis (Philippi, 1887), late Miocene, central Chile. 4, 5, Syntype SGO.PI.834, height 34 mm. 6-8, SGO.PI.3674, height 40 mm.
Diagnosis.--Shell small to medium-sized with moderately el- evated spire; ornamentation of slightly elongated, upward pro- jecting tubercles on shoulder, spiral threads on whole whorl, and four spiral cords anterior to tubercles.
Description.-Shell small to medium-sized, spire moderately elevated consisting of five to six whorls; whorls with prominent shoulder; central portion of whorl slightly depressed anterior of shoulder; suture impressed. Ornamentation consisting of slightly elongated upward-projecting tubercles, approximately 12 per whorl; tubercles form sharp shoulder giving whorl distinctly squarish profile; spiral ornamentation consisting of numerous fine threads on entire whorl surface and coarser threads, four of them prominent, anterior to tubercles. Growth lines strongly sinusoidal, posterior sinus wide and squarish; prominent raised growth lines irregularly spaced. Columella sinusoidal, columellar callus thin.
Types.--Syntypes SGO.PI.834 (height 34 mm), SGO.PI.4512- 4516 (all Matanzas). The specimen SGO.PI.834 is the one figured by Philippi (1887, pl. 1, fig. 4) and has previously been labeled as a lectotype. However, this designation apparently has never
been published and according to the ICZN (Article 74.7.3) formal designation of a lectotype is not justified, because all syntypes are conspecific.
Other material examined.-SGO.PI.3674 (one specimen, Ma- tanzas, height 40 mm), SGO.PI.3675 (17 + 23 specimens, Ma- tanzas), SGO.PI.4374 (11 specimens, Rio Futa, Valdivia), SGO.PI.4545 (one specimen, Matanzas), SMF 323632 (one spec- imen, Matanzas, height 25 mm), Bayerische Staatssammlung Miinchen, Germany, coll. Herm 1966/IV (one specimen, Matan- zas, height 38 mm).
Occurrence.-Matanzas, Navidad Formation of Late Miocene age, central Chile to Rio Futa, Santo Domingo Formation of O1- igocene to Miocene age.
Discussion.-Perissodonta chilensis is easily separated from other species of that genus by its nodular tubercles which are not as elongated and sinuous as usual in this group. The only other species occurring in Chile is the above described P. ameghinoi, which is restricted to localities in southern Chile and Argentina. The latter species occurs usually in older strata, but the locality

1128 JOURNAL OF PALEONTOLOGY, V. 79, NO. 6, 2005
near Valdivia is not well enough dated to ensure that it did not extend into the middle or late Miocene. The material of Cova- cevich et al. (1992) is not well preserved and some specimens are unusually slim and high-spired. However, as stated correctly by Covacevich et al. (1992), this material fits better in P. chilensis than in P. ameghinoi. Perissodonta ameghinoi is the most likely ancestor to P. chilensis through reduction of elongated tubercles to blunt nodes, increase in shoulder angulation, reduction of spiral ornament, and narrowing of the anal sinus. The age discrepancy between the outcrops at Matanzas (Late Miocene) and Rio Futa (Oligocene to Miocene) can be explained by two different pos- sibilities. First, the latter outcrop is not well dated and this is true for the whole Santo Domingo Formation. Second, the specimens from Matanzas may be part of a reworked fauna of early Miocene age as discussed in Nielsen et al. (2003).
DISCUSSION
Climate.--Strombidae, like Xenophoridae, is an exclusively warm-water family (Abbott, 1960, 1974; Ponder, 1983). Apor- rhaidae today have an antitropical distribution (Kronenberg, 1991), but Kiel and Bandel (2002) and Kiel (2002) showed that this was not true during the Cretaceous. The aporrhaid taxa herein described lived in cool deepwater environments. Struthiolariidae is a Southern Hemisphere group, today including relatively warm- water (Struthiolaria, Pelicaria Gray, 1847, Tylospira Harris, 1897) and cool-water (Perissodonta) representatives (Powell, 1951; Zinsmeister and Camacho, 1980; Beu and Maxwell, 1990). The Miocene stromboid fauna is representative for the whole cen- tral Chilean fauna of that time in having this mixture of warm- water and cool-water taxa. Strombus and Xenophora Fischer von Waldheim, 1807, like the co-occurring genera Nerita Linnaeus, 1758 (Neritidae; Philippi, 1887; Nielsen et al., 2004), Zonaria Jousseaume, 1884 (Cypraeidae; Groves and Nielsen, 2003), and Olivancillaria d'Orbigny, 1840 (Olividae; Nielsen, 2004), are warm-water taxa while Struthiochenopus and Perissodonta are cool-water representatives. How these seemingly contradicting climate indicators should be interpreted is left for an evaluation of the complete fauna.
Relationships of stromboid fauna.-The affinities of Strombus medinae are still unclear. Morphologically closest are juveniles of the tropical eastern Pacific Strombus (Strombus) gracilior and the Indo-Pacific S. (Tricornis) tricornis that, however, as in all other species of this genus at least back to the Miocene (Abbott, 1960), have peripheral spines. No other forms without spines are known and more material of this rare species, especially adults with a preserved outer lip, is needed to clarify its relationships.
In the Aporrhaidae, a close relationship with New Zealand taxa, as shown for species of Xenophora (Nielsen and DeVries, 2002), applies only to Hemichenopus. The morphological similarities with the Eocene H. zelandicus from New Zealand provide further evidence for an Eocene age for H. araucanus. The Eocene genus Hemichenopus seems very close to the Maastrichtian to Paleocene genus Austroaporrhais from which it probably evolved by losing the nodes on the axial sculpture. While Austroaporrhais had a wide geographical distribution, being known from New Zealand, Antarctica, and southern South America, Hemichenopus is not known from Antarctica despite the rich, well-known mollusc fau- na from the Eocene of Seymour Island (Stilwell and Zinsmeister, 1992). Hemichenopus thomsoni from the middle Eocene of New Zealand definitely does not belong in this genus, but is retained there because a more appropriate genus was not found. The genus Struthiochenopus had its origin in the Weddellian Province (Zins- meister, 1979), being known from the Upper Cretaceous and Pa- leocene of the Antarctic Peninsula and, with a stratigraphic gap, from the Late Oligocene and Miocene of Argentina and Chile
(Zinsmeister and Griffin, 1995). Drepanocheilus (Tulochilus) ben- soni Finlay and Marwick, 1937, from the lower Paleocene of New Zealand (see Beu and Maxwell, 1990, p. 81, pl. 2e), is similar to some species of Struthiochenopus and might belong here. This, however, would place Struthiochenopus in synonymy with Tulo- chilus Finlay and Marwick, 1937, and thus reassignment should await careful examination of material of D. (T.) bensoni.
In the Struthiolariidae, Perissodonta is also a Weddelian taxon that had its highest diversity during the Eocene (Zinsmeister and Camacho, 1980). There is no fossil record after the Miocene and there are only two species living in the Southern Ocean. The age of P. chilensis remains unresolved (DeVries and Frassinetti, 2003; Finger et al., 2003; Nielsen et al., 2003), but it could be the last shallow-water inhabitant of this genus, becoming restricted to the cool deepwater environments of the Circumantarctic Region after the Miocene.
CONCLUSION
The composition of Tertiary stromboid species in western South America demonstrates a mixture of Pacific (Xenophora) and Weddellian (Hemichenopus, Struthiochenopus, Perissodonta) origins. Elements derived from the Weddellian fauna (Zinsmeister, 1979), such as Struthiochenopus and Perissodonta, which per- sisted at least until the Miocene, are referred to as Neo-Weddellian taxa here. Some of these Neo-Weddellian taxa apparently immi- grated from the southeastern Atlantic shores of Argentina through trans-Andean gateways that existed during the Miocene (Nielsen and Bandel, 2003). Meanwhile, faunal exchanges with the south- western Pacific that began during the Eocene persisted through the Miocene, not only between Argentina and New Zealand (Beu et al., 1997), but also between Chile and New Zealand. However, in the Stromboidea there is not a single taxon present which oc- curs in New Zealand, Chile, and Argentina at the same time. The Chilean taxa discussed here have either relationships with New Zealand taxa or with Argentinian ones, not both.
This mixed composition of faunas shows that the model of faunal separations during the Early Tertiary (e.g., Zinsmeister and Camacho, 1980) does not apply for the Miocene because Neo- Weddellian taxa occur in a fauna with elements strongly influ- enced by or influencing the New Zealand fauna. The presence of the genus Hemichenopus in Eocene sediments of New Zealand and Chile, and its absence in Argentina, suggest that central Chile could belong to a Chilean-New Zealand faunal province rather than the more southern Weddell Province of Zinsmeister (1979). However, Eocene faunas from Chile are too poorly known to evaluate faunal relationships on a broader scale.
ACKNOWLEDGMENTS
Thanks to K. Bandel (University of Hamburg, Germany) for company in the field, numerous stimulating discussions on gas- tropods in general and the Chilean fauna in particular, and for allowing access to the facilities of the Geologisch-Paldiontolo- gisches Institut und Museum (Universitdit Hamburg) after finish- ing my doctorate. D. Frassinetti (Museo Nacional de Historia Nat- ural, Santiago, Chile) gave access to the collections under his care and his own collections. D. Villablanca (Universidad Cat6lica del Norte, Antofagasta, Chile) and W.J. Zinsmeister (Purdue Univer- sity, West Lafayette, USA) made important specimens available for study. A.G. Beu and J. Simes (Institute of Geological and Nuclear Sciences, New Zealand) arranged the loan of the figured H. zelandicus. E. Vinx (University of Hamburg, Germany) made some of the photos. N. Hiller (Canterbury Museum, Christchurch, New Zealand) provided photos of the holotype of Hemichenopus thomsoni. A. Grebneff provided photos of H. thomsoni from his private collection. Discussions with PA. Maxwell (Waimate, New Zealand), T.J. DeVries (Burton, USA), and S. Kiel (FU Berlin,

NIELSEN-CENOZOIC STROMBOID GASTROPODS FROM CHILE 1129
Germany) helped me to understand better this group and its pa- leobiogeography. T.J. DeVries also helped to improved the lan- guage. For their comments on the manuscript I gratefully ac- knowledge L.C. Anderson (Louisiana State University, USA), PA. Maxwell, and A.G. Beu.
This work was financially supported during the years 2000 to 2003 by the University of Hamburg and the Deutsche For- schungsgemeinschaft (DFG), grant Ba 675/25.
REFERENCES
ABBOTT, T. A. 1960. The genus Strombus in the Indopacific. Indo-Pacific Mollusca, 1(2):33-146.
ABBOTT, T. A. 1974. American Seashells, the Marine Mollusca of the Atlantic and Pacific Coast of North America (second edition). Van Nostrand Reinhold, New York, 663 p.
ALLAN, R. S. 1926. Fossil Mollusca from the Waihao Greensands. Trans- actions of the New Zealand Institute, 56:338-346.
BELLARDI, L. 1839. Borsonia, nouveau genre de coquille fossile. Bulletin de la Soci6te gdologique de France, 10:30-31.
BEU, A. G., AND P A. MAXWELL. 1990. Cenozoic Mollusca of New Zealand. New Zealand Geological Survey Paleontological Bulletin, 58, 518 p.
BEU, A. G., M. GRIFFIN, AND P. A. MAXWELL. 1997. Opening of Drake Passage gateway and Late Miocene to Pleistocene cooling reflected in Southern Ocean molluscan dispersal: Evidence from New Zealand and Argentina. Tectonophysics, 281:83-97.
BIELER, R., AND R. E. PETIT. 1996. Additional notes on nomina first introduced by Tetsuaki Kira in "Coloured Illustrations of the Shells of Japan." Malacologia, 38:33-34.
BORCHERT, A. 1901. Die Molluskenfauna und das Alter der Parana-Stufe. Neues Jahrbuch fuir Mineralogie. Geologie und Paliontologie, 14:171- 245.
CONRAD, T. 1860. Descriptions of new species of Cretaceous and Eocene fossils of Mississippi and Alabama. Journal of the Philadelphia Acad- emy of Sciences, 2(4):279-296.
COVACEVICH, V., D. FRASSINETTI, AND G. ALFARO. 1992. Paleontologia y condiciones de depositaci6n del Mioceno marino en las nacientes del Rio Futa, Valdivia, Chile. Boletin del Museo Nacional de Historia Nat- ural de Chile, 43:143-154.
DALL, W. H. 1889. Reports on the results of dredging, under the super- vision of Alexander Agassiz, in the Gulf of Mexico (1877-78) and in the Caribbean Sea (1879-80), by the U.S. coast survey steamer "Blake"... XXIX. Report on the Mollusca. Part II. Gastropoda and Scaphopoda. Bulletin of the Museum of Comparative Zoology at Har- vard College, 18:1-492.
DARWIN, C. 1846. Geological Observations on South America. Smith, Elder, London, 279 p.
DEL Rio, C. J. 1998. Moluscos marinos Miocenos de la Argentina y del Uruguay. Monograffas de la Academia Nacional de Ciencias Exactas, Fisicas y Naturales, Buenos Aires, 15, 151 p.
DEVRIES, T. J. 2004. Eocene Mollusks from the Pisco Basin (southern Peru): Evidence for re-evaluating the age of the Otuma Formation. XII Congreso Peruano de Geologia, Lima, Peru, 26-29 October 2004.
DEVRIES, T. J., AND D. FRASSINETTI. 2003. Range extensions and bio-
geographic implications of Chilean Neogene mollusks. Boletifn del Mu- seo Nacional de Historia Natural de Chile, 52:119-135.
D'ORBIGNY, A. D. 1840. Voyage dans l'Am6rique Mdridionale, Volume 3, No. 3 (Mollusques). Levrault, Paris, p. 409-488.
D'ORBIGNY, A. D. 1842. Voyage dans l'Amerique M6ridionale. Vol. 3, No. 4 (Paldontologie). Pitois-Levrault, Paris, 187 p.
FINGER, K., A. ENCINAS, S. NIELSEN, AND D. PETERSON. 2003. Micro- faunal indications of late Miocene deep-water basins off the central coast of Chile. 10? Congreso Geol6gico Chileno. Concepci6n, Chile. Extended abstracts (CD-ROM), 8 p.
FINLAY, H. J., AND J. MARWICK. 1937. The Wangaloan and associated molluscan faunas of Kaitangata-Green Island Subdivision. New Zea- land Geological Survey Palaeontological Bulletin, 15, 140 p.
FISCHER, P. 1880-1887. Manuel de conchyliologie et de pal6ontologie conchyliologique, ou histoire naturelle des mollusques vivants et fos- sils. Librairie E Savy, Paris, 1369 p.
FISCHER VON WALDHEIM, G. 1807. Musdum-Demidoff. Mis en Ordre
Syst6matique et Descrit Par G. Fischer, Volume 3. V6g6taux et Ani- maux. Moscow, Universitd Imp6riau, 330 p.
FLEMING, C. A. 1966. Marwick's illustrations of New Zealands shells with a checklist of New Zealand Cenozoic Mollusca. New Zealand Department of Scientific and Industrial Research Bulletin, 173, 456 p.
FRASSINETTI, D., AND V. COVACEVICH. 1993. Bivalvos del Mioceno de Matanzas (Formaci6n Navidad, Chile Central). Boletin del Museo Na- cional de Historia Natural de Chile, 44:73-97.
FRASSINETTI, D., AND V. COVACEVICH. 1999. Invertebrados f6siles mar- inos de la Formaci6n Guadal (Oligoceno Superior-Mioceno Inferior) en Pampa Castillo, Regi6n de Aysdn, Chile. Servicio Nacional de Geo- logifa y Mineria Boletin, 51, 96 p.
GABB, W. M. 1868. An attempt at a revision of the two families, Strom- bidae and Aporrhaidae. American Journal of Conchology, 4:137-149.
GARCIA, E 1968. Estratigraffa del Terciario de Chile Central, p. 25-57. In G. Cecioni (ed.), Symposio sobre el Terciario de Chile, Zona Central. Andres Bello, Santiago.
GRAY, J. E. 1840. Mollusca, p. 86-89, 106-156. In Synopsis of the Con- tents of the British Museum (forty-second edition). G. Woodfall and Son, London.
GROVES, L. T., AND S. N. NIELSEN. 2003. A new late Miocene Zonaria (Gastropoda: Cypraeidae) from central Chile. The Veliger, 46:351-354.
HARRIS, G. E 1897. Catalogue of Tertiary Mollusca in the Department of Geology, British Museum (Natural History). Pt. 1. The Australasian Tertiary Mollusca. British Museum of Natural History, London, 407 p., 8 pls.
HERM, D. 1969. Marines Pliozin und Pleistozin in Nord- und Mittel- Chile unter besonderer Beriicksichtigung der Entwicklung der Mollus- ken-Faunen. Zitteliana, 2, 159 p.
IBARAKI, M. 1992. Planktonic Foraminifera from the Navidad Formation, Chile: Their geologic age and paleoceanographic implications, p. 91- 95. In K. Ishizaki and T. Saito (eds.), Centenary of Japanese Micro- paleontology. Terra Scientific, Tokyo.
IHERING, H. 1897. Os Molluscos dos terrenos terciarios da Patagonia. Revista do Museu Paulista, 2:217-382.
IHERING, H. 1909. Nouvelles recherches sur la Formation Magellanienne. Anales del Museo Nacional, Buenos Aires, serie 3,12:27-43.
JOUSSEAUME, E P 1884. Etude sur la famille des Cypraeidae. Bulletin de la Societ6 Zoologique de France, 9:81-100.
JOUSSEAUME, E P. 1886. Coquilles marines des cotes d'Abessinie et de Zanzibar, recueilis par M. Raffray en 1873 et en 1874. Le Naturaliste, 1886:220-222.
JUNG, P 1965. Miocene Mollusca from the Paraguana Peninsula, Vene- zuela. Bulletins of American Paleontology, 49:385-652.
JUNG, P, AND A. HEITZ. 2001. The subgenus Lentigo (Gastropoda: Strom- bidae) in tropical America, fossil and living. The Veliger, 44:20-53.
KEEN, A. M. 1971. Sea Shells of Tropical West America: Marine Mol- lusks from Baja California to Peru (second edition). Stanford Univer- sity Press, Stanford, California, 1,064 p.
KIEL, S. 2002. Notes on the biogeography of Campanian-Maastrichtian gastropods, p. 109-127. In M. Wagreich (ed.), Aspects of Cretaceous Stratigraphy and Palaeobiogeography. Osterreichische Akademie der Wissenschaften, Schriftenreihe der Erdwissenschaftlichen Kommissi- onen, 15.
KIEL, S., AND K. BANDEL. 2002. About some aporrhaid and strombid gastropods from the Late Cretaceous. Palaontologische Zeitschrift, 76: 83-97.
KIRA, T. 1955. Coloured Illustrations of the Shells of Japan. Hoikusha, Osaka, 204 p.
KRAUS, O. 2000. Internationale Regeln fuir die zoologische Nomenklatur. Abhandlungen des naturwissenschaftlichen Vereins in Hamburg, n.f., 34, 232 p.
KRONENBERG, G. C. 1991. The Recent species of the family Aporrhaidae. Vita Marina, 41:47-58.
LE Roux, J. P, AND S. ELGUETA. 2000. Sedimentologic development of a Late Oligocene-Miocene forearc embayment, Valdivia Basin Com- plex, southern Chile. Sedimentary Geology, 130:27-44.
LIGHTFOOT, J. R. 1786. A catalogue of the Portland Museum, lately the property of the Duchess Dowager of Portland, deceased, which will be sold by auction, by Mr. Skinner and co. on Monday the 24th of April, 1786. Skinner, London, 194 p.
LINNAEUS, C. 1758. Systema Naturae per Regna Triae Naturae (editio

1130 JOURNAL OF PALEONTOLOGY, V. 79, NO. 6, 2005
decima, reformata). Tomus 1. Holmiae, Laurentii Salvii, Stockholm, 823 p.
MARSHALL, P. 1919. Fauna of the Hampden beds and classification of the Oamaru System. Transactions of the New Zealand Institute, 51: 226-250.
MARSHALL, P., AND R. MURDOCH. 1920. Some Tertiary Mollusca, with descriptions of new species. Transactions of the New Zealand Institute, 52:128-136.
MARTENS, E. 1878. Einige Conchylien aus den kalteren Meeresgegenden der stidlichen Erdhilfte. Sitzungsbericht der Gesellschaft naturfor- schender Freunde zu Berlin, 1878:20-26.
MARWICK, J. 1924. The Struthiolariidae. Transactions and Proceedings of the New Zealand Institute, 55:161-190.
MAXWELL, P A. 1992. Eocene Mollusca from the vicinity of McCulloch's Bridge, Waihao River, South Canterbury, New Zealand: Paleoecology and systematics. Geological Survey Paleontological Bulletin, 65, 280 p.
NIELSEN, S. N. 2004. The genus Olivancillaria (Gastropoda, Olividae) in the Miocene of Chile: Rediscovery of an older species name and de- scription of a new species. The Nautilus, 118:88-92.
NIELSEN, S. N., AND K. BANDEL. 2003. Species migration in the Miocene Pacific-paleobiogeographic and paleoclimatic implications from Chi- lean gastropods. Abstracts 18. Geowissenschaftliches Lateinamerika Kolloquium, Freiberg. Terra Nostra, 2/2003:55.
NIELSEN, S. N., AND T. J. DEVRIES. 2002. Tertiary Xenophoridae of west- ern South America. The Nautilus, 116:71-78.
NIELSEN, S. N., D. FRASSINETTI, AND K. BANDEL. 2004. Miocene Veti- gastropoda and Neritimorpha (Mollusca, Gastropoda) of Central Chile. Journal of South American Earth Sciences, 17:73-88.
NIELSEN, S. N., T. J. DEVRIES, A. ENCINAS, K. L. FINGER, AND D. PE- TERSON. 2003. Towards an understanding of the age of the Navidad Formation. 100 Congreso Geol6gico Chileno. Concepci6n, Chile. Ex- tended abstracts (CD-ROM), 7 p.
OLSSON, A. A. 1928. Contributions to the Tertiary paleontology of north- ern Peru, Pt. 1, Eocene Mollusca and Brachiopoda. Bulletins of Amer- ican Paleontology, 14:47-154.
OLSSON, A. A. 1931. Contributions to the Tertiary paleontology of north- ern Peru, Pt. 4, The Peruvian Oligocene. Bulletins of American Pale- ontology, 17, number 63, 124 p.
ORTMANN, A. E. 1902. Tertiary invertebrates. Reports of the Princeton Expedition to Patagonia, 1896-1899, 4, Palaeontology, Pt. 2:45-332.
PARODIZ, J. J. 1996. The taxa of fossil Mollusca introduced by Hermann von Ihering. Annals of Carnegie Museum, 65:183-296.
PHILIPPI, R. A. 1836. Enumeratio Molluscorum Sicilixe cum viventium turn in Tellure Tertiaria Fossilium quae in itinere suo observavit. Be- rolini sumptibus Simonis Schroppii et Sociorum, 267 p.
PHILIPPI, R. A. 1887. Die tertiiren und quartairen Versteinerungen Chiles. EA. Brockhaus, Leipzig, 266 p.
PONDER, W. E 1983. Xenophoridae of the world. The Australian Museum Memoir, 17, 126 p.
POWELL, A. W. B. 1951. Antarctic and subantarctic Mollusca: Pelecypoda and Gastropoda. Discovery Reports, 26, 128 p.
RAFINESQUE, C. S. 1815. Analyses de la Nature, ou Tableau de l'univers et des Corps Organisdes. Barraveccia, Palermo, 224 p.
SCHUMACHER, H. C. E 1817. Essais d'un Nuveau Systeme des Habita- tions des Vers Testacds. Schultz, Copenhagen, 287 p.
SMITH, E. A. 1875. Descriptions of some new shells from Kerguelen's Island. The Annals and Magazine of Natural History, series 4, 14:67- 73.
SOWERBY, G. B. 1825. A Catalogue of the Shells Contained in the Col- lection of the Late Earl of Tankerville. London, 92 p. and 34 p., ap- pendix.
STEINMANN, G., AND O. WILCKENS. 1908. Kreide-und Tertidirfossilien aus den Magellanslindern gesammelt von der schwedischen Expedition 1895-1897. Arkiv for Zoologi, 4(6), 119 p.
STILWELL, J. D., AND W. J. ZINSMEISTER. 1992. Molluscan systematics and biostratigraphy, lower tertiary La Meseta Formation, Seymour Is- land, Antarctic Peninsula. American Geophysical Union, Antarctic Re- search Series, 55:1-192.
TAVERA, J. 1979. Estratigraffa y paleontologifa de la Formaci6n Navidad, Provincia de Colchagua, Chile (Lat. 30'50'-34?S). Boletin del Museo Nacional de Historia Natural de Chile, 36, 176 p.
TSUCHI, R., T. SHUTO, T. TAKAYAMA, I. KOIZUMI, A. FUJIYOSHI, M. IBARAKI, AND R. MARTINEZ-PARDO. 1990. Fundamental data on Ce- nozoic biostratigraphy of Chile-Supplement 3. Punta Perro section north of Navidad, central Chile. Reports of Andean Studies, Special Volume 3:70-71.
VILLABLANCA, D. V. 2001. Geologia y petrologia del carb6n: Yacimiento Carbonifero Mulptin, Provincia de Valdivia, X Regi6n de Los Lagos, Chile. Unpublished master's thesis, Universidad de Concepci6n, Chile, 156 p.
WILCKENS, 0. 1910. Die Anneliden, Bivalven und Gastropoden der An- tarktischen Kreideformation. Wissenschaftliche Ergebnisse der Schwedischen Stidpolar-Expedition 1901-1903, 3(12), 132 p.
WOODRING, W. P 1957-1982. Geology and paleontology of Canal Zone and adjoining parts of Panama. U.S. Geological Survey Professional Paper, 306 A-F, 759 p.
WOODS, H. 1922. Mollusca from Eocene and Miocene deposits of Peru, p. 51-113. In T. O. Bosworth (ed.), Geology of the Tertiary and Qua- ternary Periods in the Northern Part of Peru. Macmillan, London.
ZINSMEISTER, W. J. 1979. Biogeographic significance of the Late Meso- zoic and Early Tertiary molluskan faunas of Seymour Island (Antarctic Peninsula) to the final breakup of Gondwanaland, p. 344-347. In J. Gray and A. J. Boucot (eds.), Historical Biogeography, Plate Tectonics and Changing Environments. 37th Biological Colloquium. Oregon State University Press, Corvallis.
ZINSMEISTER, W. J., AND H. H. CAMACHO. 1980. Late Eocene Struthio- lariidae (Mollusca: Gastropoda) from Seymour Island, Antarctic Pen- insula and their significance to the biogeography of Early Tertiary shal- low-water faunas of the Southern Hemisphere. Journal of Paleontology, 54:1-14.
ZINSMEISTER, W. J., AND M. GRIFFIN. 1995. Late Cretaceous and Tertiary aporrhaid gastropods from the southern rim of the Pacific Ocean. Jour- nal of Paleontology, 69:692-702.
ACCEPTED 26 NOVEMBER 2004