Embed Size (px)
Citation preview

Toslron Vol . 29, No. 1, pp. 896, 1991 .
0041-0101/91 ß.00 + .00Printed in Great Britain.
~i 1990 Pcrgemon Pros plc
NEW TOXINS FROM THE VENOM OF THE COMMON TIGERSNAKE (NOTECHIS SCUTATUS SCUTATUS)
BRIAN FRANCIS, TED R. JOHN, CORRINE SEEBART and IVAN I . KAISER*
Department of Molecular Biology, University of Wyoming, Laramie, WY 82071, U.S.A .
(Accepted for publication 27 June 1990)
B. FRANCIS, T. R . JOHN, C. SEEBART and I . I . KAISER . New toxins from thevenom of the common tiger snake (Notechis scutatus scutatus) . Toxicon 29, 85-96, 1991 .-Scutoxin A and B represent two isoforms of a new toxic proteinfrom the venom of the Australian tiger snake, Notechis scutatus scutatus. Bothisoforms, of apparent mol. wt 13,000, are less basic than either notexin ornotechis II-5 . They both have similar i.v . LD Sp-values in mice of ca 0.006 ~g/g,and phospholipase activities of about 136 moles of fatty acid released/min/mg at 37°C when acting on phosphatidylcholine in the presence of TritonX-100. Toxicities of the scutoxins are the same as notexin and about seventimes more potent than notechis II-5 . ELISAs and western blot analysesindicate that the new toxins are immunologically similar to notexin andnotechis II-5, with phospholipase activities falling between these latter twoproteins . When crude venom is initially passed over a gel filtration column,each scutoxin isoform co-elutes in a different fraction with notexin . Gelfiltration experiments using purified samples of notexin and scutoxin havefailed to demonstrate any evidence for the formation of higher mol. wt proteincomplexes . Peptide mapping suggests the presence of five glutamate residues inone of the protein isoforms . These findings, together with the high toxicity andactive phospholipase levels, demonstrate that the new proteins are not thepreviously reported non-toxic and enzymatically inactive notechis II-1 . Thecombination of gel filtration on Sephacryl 5-200 and cation-exchangechromatography used to isolate the scutoxins also permits recovery of notexinand notechis II-5 in high purity .
INTRODUCTION
Toxic proteins isolated from the venom of the Australian tiger snake, Notechis scutatusscutatus, include the presynaptic neurotoxins, notexin and notechis II-5, and the post-synaptic neurotoxin, notechis III-4 (HALPERT and FAKER, 1975, 1976a; HALPERT et al .,1979). Notexin and notechis II-5 exhibit phospholipase A2 (PLAN activity (HAL1'ERT andFAKER, 1976b) . Another PLA2 homolog, notechis II-1, has both low toxicity and PLA2activity (HALPERT and FAKER, 1976b; LIND and FAKER, 1980). During our purification ofnotexin from crude venom ofN. s. scutatus, we encountered a protein which was less basicthan either notexin or notechis II-5 . Initially this protein was thought to be notechis II-1
'Author to whom correspondence should be addressed .
85

86
B. FRANCIS et al.
but it was found to be as toxic as notexin. Further study indicated that it also hadphospholipase activity and existed in two major isoforms which differed in charge .Experiments described elsewhere show that there can be substantial differences in theprotein components of venoms obtained from different Notechis subspecies (JoxN andKAISER, 1990), with the newly identified toxin found principally in N. s. scutatus. In thispaper we describe the purification and partial characterization of the isoforms of the newproteins which we shall call scutoxin A and B. We also provide an alternative procedurefor the isolation of highly purified notexin and notechis II-5 .
MATERIALS AND METHODSCrude, dried venom of Notechis scutatus scutatus, collected from the Victorian and South Australian region,
was obtained from Mr Pmt MIRTSCHIIV, Venom Supplies, Tanunda 5352, South Australia. Sephacryl5-200-HR, Mono S (cation exchanger) 0.5 x 5 cm, Mono Q (anion exchanger) 0.5 x 5 cm, the Superose 12analytical column, as well as the Fast Protein Liquid Chromatography (FPLC) system, were purchased fromPharmacia, Uppsala, Sweden. Sephacryl 5-200 gel filtration used 0.1 M ammonium acetate (pH 6 .9) as therunning buffer. Equilibrating buffer for cation-exchange chromatography on Mono S was 50 or 100 mM sodiumphosphate, with the eluting buffer consisting of 50 mM sodium phosphate plus 1 M sodium chloride . Phosphatebuffers were adjusted to either pH 6.5 or 5 .0, as indicated . Buffers for anion-exchange (Mono Q) were 50 mMTris-HCl and 50 mM Tris-HCl plus 1 M sodium chloride (pH 7 .0) . Absorbance of eluent from the S-200, MonoS, and Mono Q columns was monitored at 280 nm, although in some instances fractions were also measured at260 nm .Notexin and scutoxin A were reduced, carboxyamidomethylated and digested with glutamate specific
endoprotease, Glu-C (Boehringer-Mannheim) as described (Ktisee et al., 1990) . Reversed phase chromato-graphy of digested protein was carried out on the FPLC system using a Vydac 0.46 x 15 cm C, s column (5 pm) in0.1% trifluoroacetic acid (TFA) and eluted with 75% acetonitrile in 0.1% TFA . Peptides eluting from thecolumn were detected by monitoring at 214 nm .SDS-PAGE (15%)gels were Coomassie blue- or silver-stained as described (ne Sttvn et al., 1989) . Urea gels
were prepared and run as described elsewhere (True et al., 1971) . Phospholipase and lethality assays followedthe methods of A~rto and Iü~t (1985a, b) . Either male or female mice in the 20-30 g weight range were usedfrom our colony (Auto and K~~sex, 1985a) . The primary monoclonal antibody used in ELISAs and westernblots was kindly provided by Dr Joxx MIDDUuHROOK, (Pathology Division, USAMRIID, Frederick, MD,U.S.A .). It was raised against purified pseudexins (SCw~uUT and Mu»t.~axootc, 1989) isolated from PseudechCsporphyriacus, but was shown to cross-react with notexin (M~nor-eexoox, 1990) . ELISAs were carried out onchromatographic fractions by diluting the protein concentration of each fraction to about 1 pg/ml in Coat Buffer(35 mM NaHCO,, 15 mM Na2C0~ and 0.02% NaN3 at pH 9.6) and coating the miaotiter wells (Dynatech,Immulon II) by incubating at 4°C overnight . Plate wells were washed four times with a Wash Buffer of 0.5%Tween-20 (Bio-Rad EIA grade) in PBS (8 .1 mM NazHPO� 1 .5 mM KHZPO,, 2 .7 mM KCl and 140 mM NaCI atpH 7 .4) . Free sites on the plate surfaces were blocked by addition of 1% gelatin in PBS and incubated for I hr atroom temperature . Following a wash step as described above, monoclonal ascites fluid was serially diluted inmicrotiter wells from (0 to 10° times and incubated 1 hr at room temperature. Plate wells were washed again andSigma Goat Anü-mouse IgG Peroxidase Conjugate (diluted I : 500 in Wash Buffer) was added and incubated for2 hrs at room temperature. After another plate wash, color developer (1 mg/ml of 2,2'-azinobis(3-ethylbenz-thiazolinesulfonic acid) in 50 mM citric acid, pH 4.0 with 0.03% Hz02 added just before use) was added . T$iswas incubated at room temperature for 6 min and the reacÙon stopped by addition of 10% SDS . Absorbanoeswere read at 414 and 510 nm in a Titertech Multiskan MC obtained from Flow Labs (7655 Old Springhouse Rd,McLean, VA, U.S.A .).For western blots, proteins from SDS-PAGE were transferred to nitrocellulose filters using a Polyblot
electrotransfer apparatus from American Bionetics (Hayward, CA, U.S.A .) following the manufacturer'sinstructions . Visualization ofnotexin and notexin-like proteins was performed as described by Jottrr and K~tsex(1990) .
RESULTS
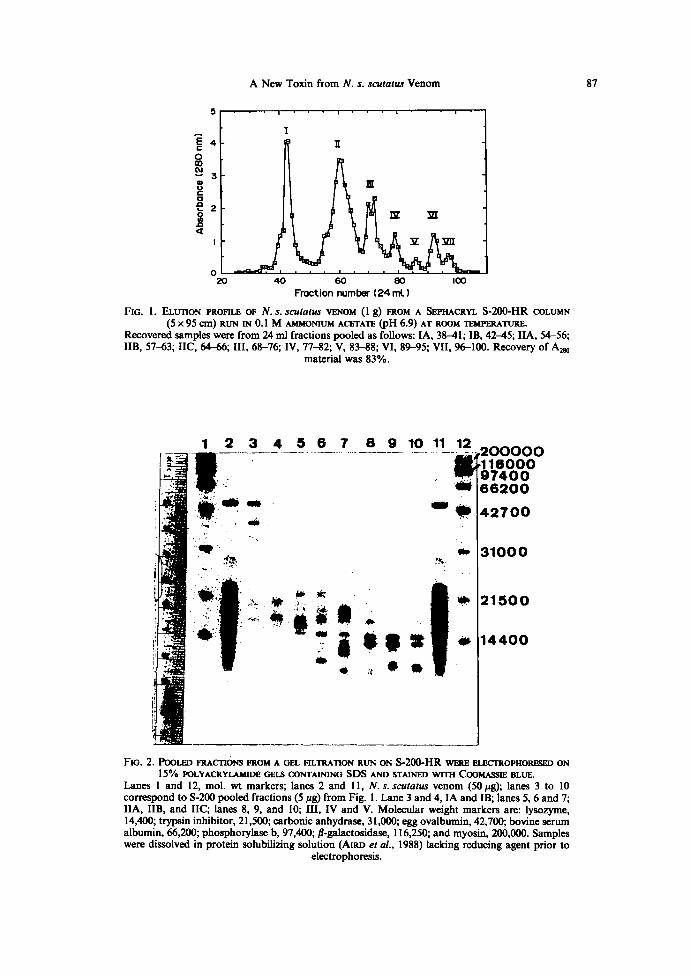
Figure 1 shows the 280nm absorbance profile of the column effluent from the5-200-HR cohunn with the peaks numbered from I to VII. On some runs peak II resolvedinto two components . Ultraviolet absorbance spectra of each peak (not shown) and260/280 nm absorbance ratios of peak tubes (peak I, 0.5 ; peak II, 0.6 ; peak III, 4.1 ; peak
c a
N 3muc0a 2
Q
A New Toxin from N. s . scutatus Venom
87
40 60 80 100Fraction number f24m1)
FIG . I . ELUTION PROFILE OF N. S. scutatus VENOM (I g) FROM A SEPHACRYL S-2OO-HR COLUMN(5 X9S Cm) RUN IN O .I M AMMONIUM ACETATE (pH 6.9) AT ROOM TEMPERATURE .
Recovered samples were from 24 ml fractions pooled as follows: IA, 381; IB, 425; IIA, 54-56;IIB, 57-63 ; IIC, 646; III, 68-76; IV, 77-82; V, 83--88; VI, 89J95 ; VII, 96-100 . Recovery of A~
material was 83% .
8 9 1011 12 200000. _ ._11600097400
~ 66200~42700
31000
21500
14400
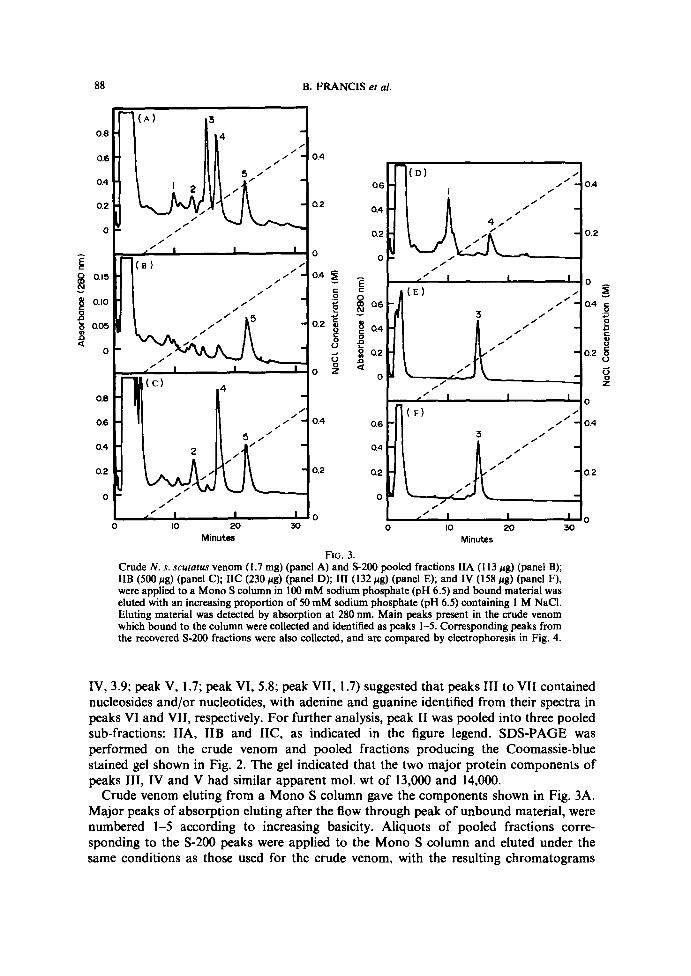
FIG . 2 . POOLED FRACTIONS FROM A GEL FILTRATION RUN ON S-2OO-HR WERE ELECTROPHORESED ONIS% POLYACRYLAMIDE GELS CONTAUiING SDS AND STAINED WITH COOMAS.StE BLUE.
Lanes 1 and 12, mol . wt markers; lanes 2 and 11, N. s . scatatus venom (50 pg); Lanes 3 to 10correspond to 5-200 pooled fractions (5 hg) from Fig . 1 . Lane 3 and 4, IA and IB; lanes 5, 6 and 7;IIA, IIB, and IIC ; lanes 8, 9, and 10; III, IV and V . Molecular weight markers are : lysozyme,14,400; trypsin inhibitor, 21,500; carbonic anhydrase, 31,000; egg ovalbumin, 42,700; bovine serumalbumin, 66,200 ; phosphorylase b, 97,400; ß-galactosidase, 116,250; and myosin, 200,000 . Sampleswere dissolved in protein solubilizing solution (AIRD et al., 1988) lacking reducing agent prior to
electrophoresis .
8 8
B. FRANCIS er al .
o.e
o.so.a0.2
O
Q15
0.10
Q05
0
O.B
0.6
0.4
0.2
0
Minutes
Minutes
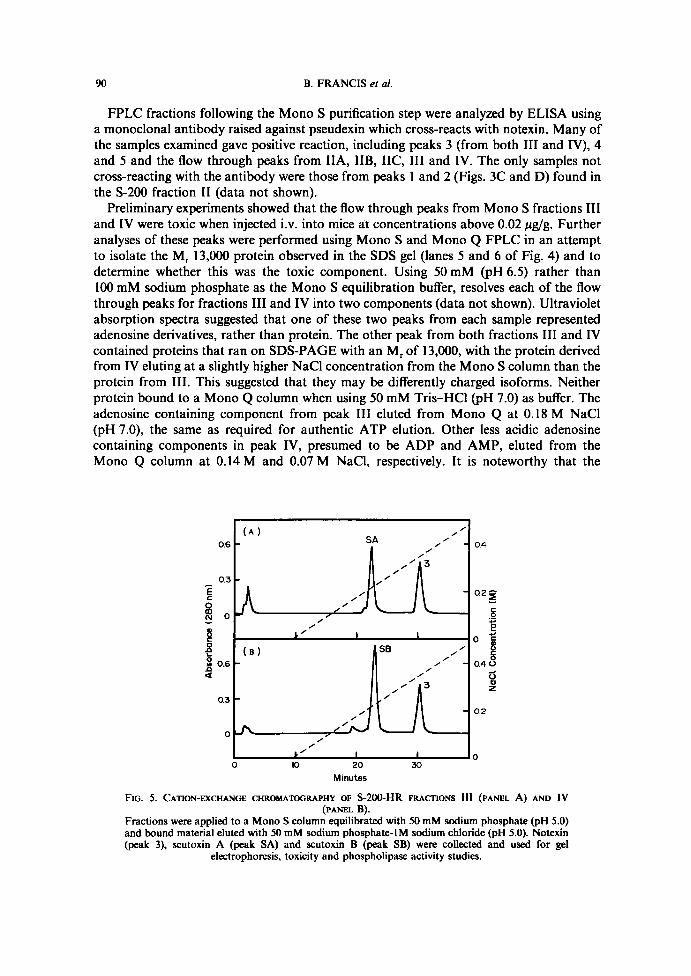
F~a . 3 .Crude N . .s. .scutatus venom (1 .7 mg) (panel A) and 5-200 pooled fractions IIA (113 ug) (panel B);IIB (500 Ng) (panel C) ; IIC (230 pg) (panel D) ; III (132 Ng) (panel E); and IV (158 ug) (panel F),were applied to a Mono S column in 100 mM sodium phosphate (pH 6.5) and bound material waseluted with an increasing proportion of 50 mM sodium phosphate (pH 6 .5) containing 1 M NaCI.Eluting material was detected by absorption at 280 nm. Main peaks present in the crude venomwhich bound to the column were collected and identified as peaks 1-5 . Corresponding peaks fromthe recovered 5-200 fractions were also collected, and are compared by electrophoresis in Fig. 4 .
0 .4
0.2
d
0.2U
ÛOZ
0
0.4
0.2
0
IV, 3 .9 ; peak V, 1 .7 ; peak VI, 5.8 ; peak VII, 1 .7) suggested that peaks III to VII containednucleosides and/or nucleotides, with adenine and guanine identified from their spectra inpeaks VI and VII, respectively . For further analysis, peak II was pooled into three pooledsub-fractions: IIA, IIB and IIC, as indicated in the figure legend . SDS-PAGE wasperformed on the crude venom and pooled fractions producing the Coomassie-bluestained gel shown in Fig. 2. The gel indicated that the two major protein components ofpeaks IIi, IV and V had similar apparent mol. wt of 13,000 and 14,000 .Crude venom eluting from a Mono S column gave the components shown in Fig. 3A .
Major peaks of absorption eluting after the flow through peak of unbound material, werenumbered 1-5 according to increasing basicity . Aliquots of pooled fractions corre-sponding to the S-200 peaks were applied to the Mono S column and eluted under thesame conditions as those used for the crude venom, with the resulting chromatograms

A New Toxin from N. s . scutatus Venom
89
1 2 3 4 5 6 7 8 9 1011 12 13141516 17181920-* . . . . .
FIG . 4 . SODIUM DOCECYL SULFATE-PAGE OF PROTEINS 3EPARATED HY MONO S FROM EITIiIIt CRUDEN. s . scatatus VENOM OR 5-200 POOLED FRACTIONS AND S'I'AINFD WITH SILVER.
Lanes 1 and 20, mol . wt standards as in Fig. 2 ; lanes 18 and 19, N. s . scutatus venom (1 and 2 Ng) ;lanes 2 to 6 correspond to flow through peaks from Mono S fractions (Fig . 3) IIA, IIB, IIC, IIIand IV, respectively. Lanes 7-12 correspond to Mono S peaks l-5 isolated from 5-200 fractions(Fig . 3B-F) . Lane 7, 1 ; 8, 2 ; 9, 3 (Fig . 3E) ; 10, 3 (Fig . 3F) ; ll, 4 ; and 12, 5 . Lanes 13-17,wrrespond to Mono S peaks 1-5 isolated from crude N. s . scutatus venom (Fig . 3A). Lane 13, 1 ;
14, 2 ; 15, 3 ; 16, 4 ; and 17, 5 . Lanes 2 to l7 contained 0 .5 Ng protein .
shown in Fig. 3. Peak IIA (Fig. 3B) contained mainly peak 5, IIB (Fig . 3C) peaks 2, 4 and5, IIC (Fig . 3D) peaks 1 and 4, III (Fig . 3E) peak 3, andN(Fig . 3F) peak 3. Peaks 3 fromS-200 fractions III andNeluted from the cation exchange column at the same concentra-tion of sodium chloride (Fig . 3E and 3F), suggesting similar basicity . An acid-ureapolyacrylamide gel of the five major peaks resolved in Fig. 3B to 3F, showed the expectedincrease in basicity from peaks 1-5, respectively (data not shown). Peak 3 from both S-200fractions III and Nco-migrated in this system . SDS-PAGE was carried out on materialrecovered from peaks 1 to 5 from the crude venom (Fig . 3A), and peaks 1-5 and the flowthrough from S-200 pooled fractions IIA, IIB, IIC, III and N (Figs 3B-F) giving thesilver-stained gel shown in Fig. 4. Protein present in peak 3 from S-200 fractions III andIV (Fig. 4, slots 9 and 10) had an M~ of 14,000 and therefore represented the upper of thetwo major bands found in III and IV before Mono S separation (slots 8 and 9, Fig. 2). Thelower, 13,000 M~ protein present in these two slots were found in the flow through peaksfrom III andN(slots 5 and 6, Fig. 4) . The flow through for III also included a 16,000 M,protein (slot 5, Fig. 4) . From published data the basic 14,000 M~ protein (peak 3, Fig. 3)was tentatively identified as notexin (KARLSSOIV et al., 1972 ; IIALPERT and EAxm, 1975,19766) and the more basic 14,000 M~ protein representing peak 4 (Fig . 3) as notechis 11-5(HALPERT and EAxER, 1976x) .
9 0
B.
FRANCIS et al
.
FPLC
fractions following the Mono S purification step were analyzed by ELISA using
a
monoclonal antibody raised against pseudexin which cross-reacts with notexin
.
Many of
the
samples examined gave positive reaction, including peaks 3 (from both III and IV), 4
and
5 and the flow through peaks from IIA, IIB, IIC, III and IV
.
The only samples not
cross-reacting
with the antibody were those from peaks 1 and 2 (Figs
.
3C and D) found in
the
S-200 fraction II (data not shown)
.Preliminary
experiments showed that the flow through peaks from Mono S fractions III
and
IV were toxic when injected i
.v .
into mice ât concentrations above 0
.02
~g/g
.
Further
analyses
of these peaks were performed using Mono S and Mono Q FPLC in an attempt
to
isolate the M~ 13,000 protein observed in the SDS gel (lanes 5 and 6 of Fig
.
4) and to
determine
whether this was the toxic component
.
Using 50mM (pH 6
.5)
rather than
100
mM sodium phosphate as the Mono S equilibration buffer, resolves each of the flow
through
peaks for fractions III and IV into two components (data not shown)
.
Ultraviolet
absorption
spectra suggested that one of these two peaks from each sample represented
adenosine
derivatives, rather than protein
.
The other peak from both fractions III and IV
contained
proteins that ran on SDS-PAGE with an M~ of 13,000, with the protein derived
from
IV eluting at a slightly higher NaCI concentration from the Mono S column than the
protein
from III
.
This suggested that they may be differently charged isoforms
.
Neither
protein
bound to a Mono Q column when using 50 mM Tris-HCl (pH 7
.0)
as buffer
.
The
adenosine
containing component from peak III eluted from Mono Q at 0
.18
M NaCI
(pH
7
.0),
the same as required for authentic ATP elution
.
Other less acidic adenosine
containing
components in peak IV, presumed to be ADP and AMP, eluted from the
Mono
Q column at 0
.14
M and 0
.07
M NaCI, respectively
.
It is noteworthy that the
0
b 20 30
MinutesFIG .
S
.
CATION-EXCHANGE CHROMATOGRAPHY OF S-2OO-HR FRACTIONS III (PANEL A) AND IV
(PANEL
B)
.Fractions
were applied to a Mono S column equilibrated with 50 mM sodium phosphate (pH S
.0)and
bound material eluted with 50 mM sodium phosphate-lM sodium chloride (pH S
.0).
Notexin
(peak
3), scutoxin A (peak SA) and scutoxin B (peak SB) were collected and used for gel
electrophoresis,
toxicity and phospholipase activity studies
.
A New Toxin from N. s. scutatus Venom
91
1 2 3 4 5 ß 7 8 91011 121314 15
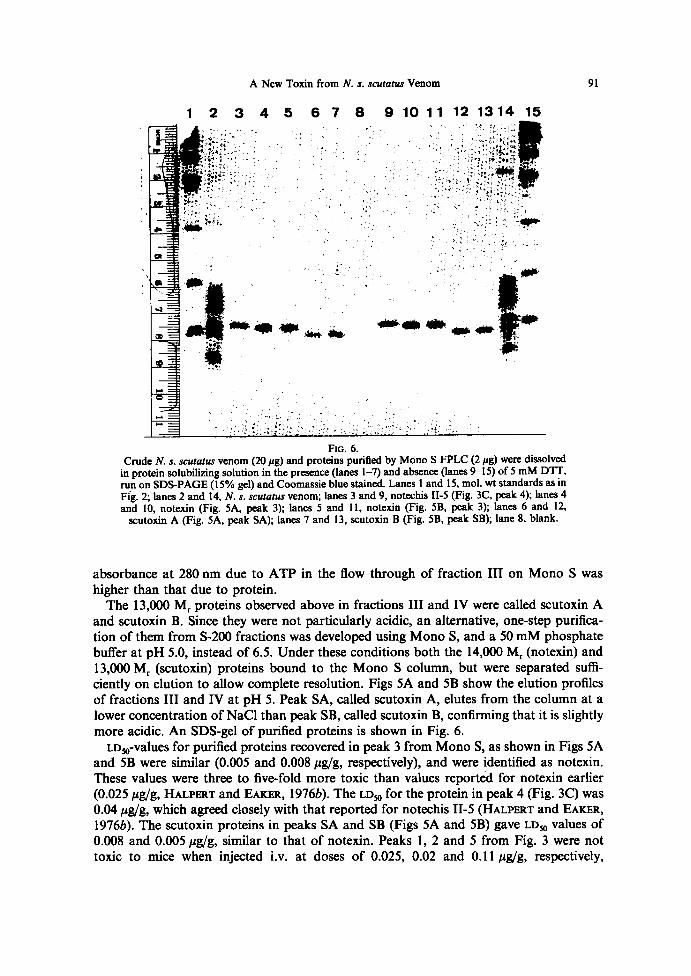
F~a. 6.CrudeN. s. scutatus venom (20 pg) and proteins purified by Mono SFPLC (2 pg) were dissolved
in protein solubilizing solution in the presence (lanes 1-~ and absence (lanes 9-15) of 5 mM DTT,run on SDS-PAGE (l5% gel) and Coomassie blue stained . Lanes 1 and 15, mol. wt standards as inFig. 2; lanes 2 and 14, N. s. scutatus venom; lanes 3 and 9, notechis II-5 (Fig . 3C, peak 4) ; lanes 4and 10, notexin (Fig . SA, peak 3) ; lanes 5 and 11, notexin (Fig . SB, peak 3) ; lanes 6 and 12,
scutoxin A (Fig. SA, peak SA); lanes 7 and 13, scutoxin B (Fig. SB, peak SB); lane 8, blank.
absorbance at 280 nm due to ATP in the flow through of fraction III on Mono S washigher than that due to protein.The 13,000 M~ proteins observed above in fractions III and IV were called scutoxin A
and scutoxin B . Since they were not particularly acidic, an alternative, one-step purifica-tion of them from 5-200 fractions was developed using Mono S, and a 50 mM phosphatebuffer at pH 5.0, instead of 6.5 . Under these conditions both the 14,000 M~ (notexin) and13,000 M~ (scutoxin) proteins bound to the Mono S column, but were separated suffi-ciently on elution to allow complete resolution . Figs SA and SB show the elution profilesof fractions III and IV at pH 5. Peak SA, called scutoxin A, elutes from the column at alower concentration of NaCI than peak SB, called scutoxin B, confirming that it is slightlymore acidic . An SDS-gel of purified proteins is shown in Fig. 6.LDspvalues for purified proteins recovered in peak 3 from Mono S, as shown in Figs SA
and SB were similar (0.005 and 0.008 f~gfg, respectively), and were identified as notexin.These values were three to five-fold more toxic than values reportéd for notexin earlier(0.025 ~g/g, HALPERT and EAxaIt, 19766) . The LDsp for the protein in peak 4 (Fig. 3C) was0.04 Ng/g, which agreed closely with that reported for notechis II-5 (HALPERT and FAKER,19766) . The scutoxin proteins in peaks SA and SB (Figs SA and SB) gave LDsp values of0.008 and 0.005 hg/g, similar to that of notexin. Peaks 1, 2 and 5 from Fig. 3 were nottoxic to mice when injected i.v . at doses of 0.025, 0.02 and 0.11 pg/g, respectively,
9 2
B. FRANCIS et a!.
although peak 5 was toxic at 0.5 Pg/g . All mice used in toxicity assays were larger than20 g.
Phospholipase activities of the purified proteins were determined and are reported inmoles of fatty acid released/min/mg, using 10 pg of each toxin per assay. Peak 2 from
Fig. 3 gave a value of 29 . Activities of notexin (peak 3 from Figs SA and SB) were nearlyidentical at 112 and 102, respectively . Scutoxin A and B also had similar phospholipaseactivites of 134 and 139, respectively . Notechis II-5 (Fig . 3C, peak 4), with an activity of182 had an activity ratio to notexin of ca 1 .7 : 1 . This is close to the notechis II-5 : notexinratio of 1 .65 : 1 reported by HALPERT and FAKER (1976b). Phospholipase activity ofscutoxin is between that of notexin and notechis II-5 . Peak 5 had an activity of 13t . Thephospholipase activity of both notechis II-5 and peak 5 showed sigmoidal time coursessimilar to that observed for intact crotoxin and related, dimeric rattlesnake presynapticneurotoxins. Notexin, scutoxin A and scutoxin B showed hyperbolic time courses like thatfound using the isolated basic subunit of crotoxin (AIRD and KAISER, 1985a) .
Experiments conducted to this point suggested that scutoxins A and B representedpreviously unreported toxins, with fractions 3 and 4 in Fig. 3, corresponding to notexinand notechis II-5, respectively. Reversed phase chromatography elution of notexin fromS-200 peak III and IV eluted at the same concéntration of acetonitrile, and a mixture ofthe two eluted as a single symmetrical peak . The same result was found for scutozin A andB, indicating that there was little hydrophobicity difference between these protein pairs.Earlier results with the Mono S and Mono Q columns showed that there were nodetectable differences in charge between the notexns recovered in S-200 fractions III andIV at either pH 5.0 or 6.5, suggesting that both fractions were identical . Althoughscutoxins A and B have the same hydrophobicity, they differ in charge, with SB beingmore basic at pH 5.0 (see Figs SA and B) .
o.so.a0.2
Ec
ON
D
0.8Q
D.a0.2
O
Minutes
so
15
O Ô
15
0
m
~ a
a°
F1G. i. PURIFIED SCUTOXIN A AND NOTEXIN FROM S-ZOO PEAK III WERE REDUCED, CARBOXYAMllI(YMECHYLATED (2 mg OF EACH) AND MIXED WITH GLUTAMA7E SPECIFIC PROTEASE (lO pg) FOR 24 hr AT
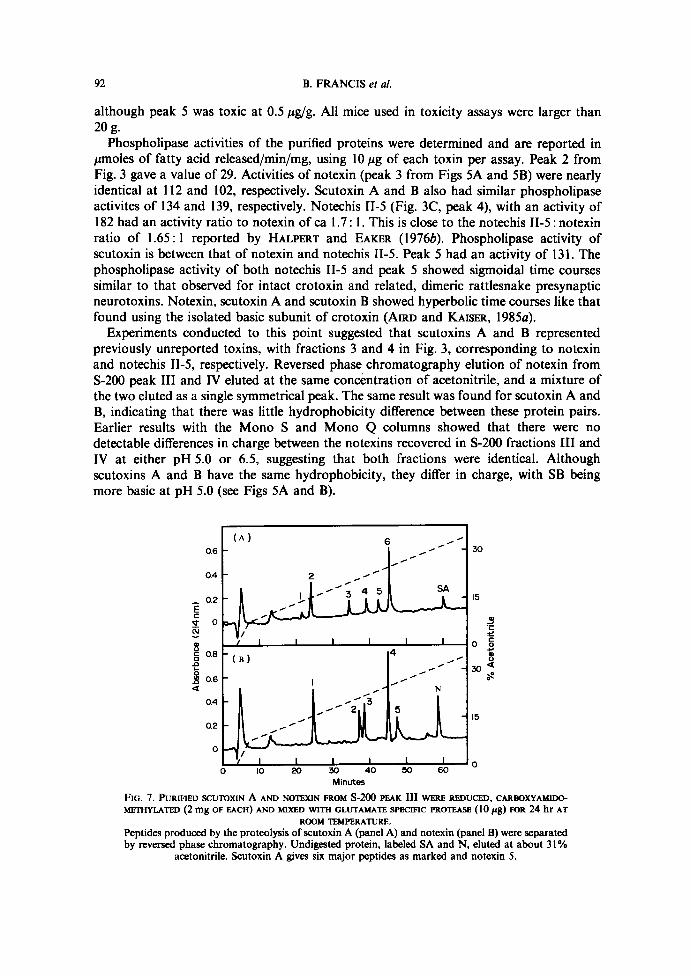
ROOM TEMPERATURE.Peptides produced by the proteolysis ofscutoxin A (panel A) and notexin (panel B) were separatedby reversed phase chromatography . Undigested protein, labeled SA and N, eluted at about 31%
acetonitrile . Scutoxin A gives six major peptides as marked and notexin 5 .
A New Toxin from N. s. scutatus Venom
93
1 2 3 4 5 6 7
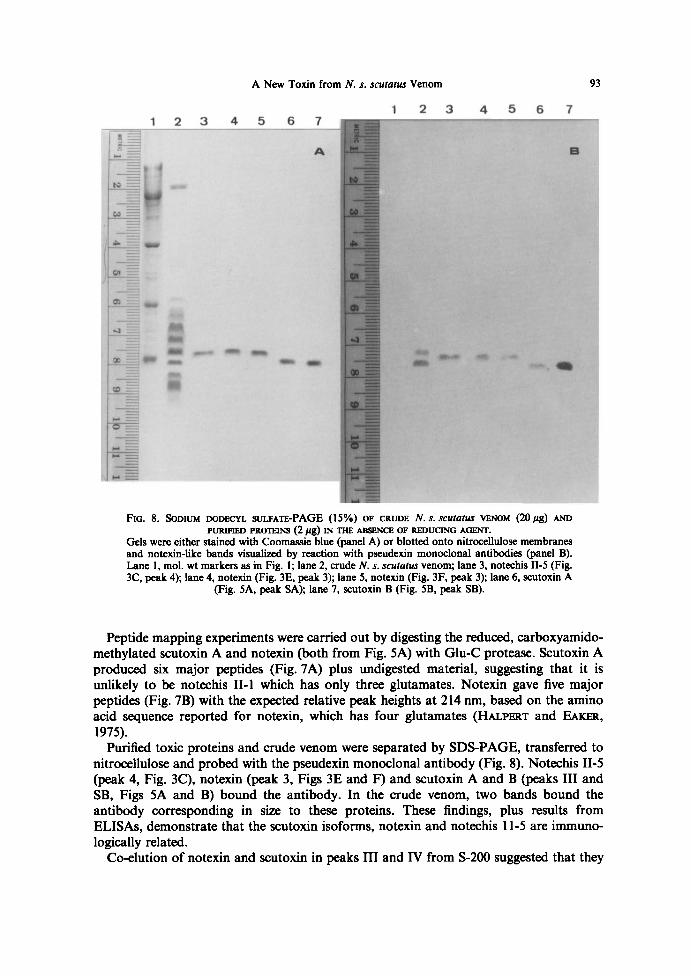
FiG. 8. SoDiue~ DoDECV~ suc.FATE-PAGE (15%) OF CRUDE N. s. scutntur VENOM (20 pg) ArroPURQ~fED PROTEINS (2 Pg) IN TF~ ABSENCE OF REDUCING AGENT.
Gels were either stained with Coomassie blue (panel A) or blotted onto nitrocellulose membranesand notexin-like bands visualized by reaction with pseudexin monoclonal antibodies (panel B).Lane 1, mol. wt markers as in Fig. l; lane 2, crude N. s. scutatus venom; lane 3, notechis II-5 (Fig.3C, peak 4); lane 4, notexin (Fig . 3E, peak 3); lane 5, notexin (Fig . 3F, peak 3); lane 6, scutoxin A
(Fig. SA, peak SA); lane 7, scutoxin B (Fig . SB, peak SB).
Peptide mapping experiments were carried out by digesting the reduced, carboxyamido-methylated scutoxin A and notexin (both from Fig. SA) with Glu-C protease . Scutoxin Aproduced six major peptides (Fig . 7A) plus undigested material, suggesting that it isunlikely to be notechis II-1 which has only three glutamates . Notexin gave five majorpeptides (Fig. 7B) with the expected relative peak heights at 214 nm, based on the aminoacid sequence reported for notexin, which has four glutamates (HALPERT and EAICER,
1975).Purified toxic proteins and crude venom were separated by SDS-PAGE, transferred to
nitrocellulose and probed with the pseudexin monoclonal antibody (Fig . 8) . Notechis II-5(peak 4, Fig. 3C), notexin (peak 3, Figs 3E and F) and scutoxin A and B (peaks III andSB, Figs SA and B) bound the antibody . In the crude venom, two bands bound theantibody corresponding in size to these proteins. These findings, plus results fromELISAs, demonstrate that the scutoxin isoforms, notexin and notechis 11-5 are immuno-logically related.
Co-elution of notexin and scutoxin in peaks III and IV from 5-200 suggested that they

94
B. FRANCIS et at.
TABLE I . PROTEIN AND NUCLEOTIDE ELUTION VOLUMES ON SUPER06E I2
Column was equilibrated with O .l M ammonium acetate (pH 6.9) atroom temperature. All runs were carried out at a flow rate of 0 .5ml/min and the efliuent monitored at 280 nm.
might differ or perhaps exist in different aggregation states to elute as two peaks. Wecould not detect any differences between the notexins present in peaks III and IV fromS-200. Scutoxin A (from III) was only slightly more acidic than scutoxin B (from IV). Anumber of gel filtration experiments using a Superose 12 column and FPLC were carriedout in an attempt to detect any molecular associations between the notexin, scutoxins andATP. Elution volumes for the major eluting peaks are shown in Table 1 . These resultsindicate that under the conditions used, notexin does not associate with scutoxin A toform a higher mol. wt complex and that ATP does not associate with notexin.
DISCUSSION
Two isoforms of a new toxin, which we called scutoxin A and B, were purified from thevenom of N. s. scutatus. The proteins have been shown to be different from notexin,notechis II-5, and notechis II-1 which were previously isolated from the same venom.Procedures for purifying notexin, notechis II-5, and scutoxin proteins using a combinationof gel filtration on Sephacryl S-200-HR and cation-exchange on Mono S have beendeveloped .When crude venom is applied to a S-200 gel filtration column, scutoxin A and scutoxin
B (M, 13,000) co-elute in peaks III and 1V with notexin (Mr 14,000). Peak V also appearsto contain notexin and scutoxin as shown by SDS-PAGE (compare Fig. 2 slot 10 for peakV with slots 8 and 9 for peaks III and IV) but this has not been proven . Notechis II-5 hasa mol. wt similar to both notexin and scutoxin but it elutes earlier (in peak II) in gelfiltration experiments with the crude venom. Based on its position of elution relative tonotexin and scutoxin, notechis II-5 appears to exist in crude venom as a higher mol. wtaggregate . Notexin and scutoxin may also exist as aggregates with decreasing molecularvolumes as one goes from peak III to IV; and perhaps exist as monomers in peak V.Presynaptic neurotoxins are known to exist in a variety of molecular forms, including thatof monomers (ammodytoxin A), heterologous dimers (crotoxin, ß-bungarotoxin), trimers(taipoxin), and pentamers (textilotoxin) . Thus, it is reasonable to suggest that there maybe molecular associations between scutoxin and notexin in peaks III to V. However,attempts to demonstrate an association between purified notexin and scutoxin using gelfiltration on Superose 12 FPLC have been unsuccessful . We are continuing to examinevarious combinations in an attempt to resolve this problem. Additional rations or anions,
Sample Elution volumes (ml)
Peak III (Fig . L) 17 .2 18 .5 19 .4Notexin (Fig . 3E, 3) 19 .4Scutoxin A (Fig. SA, SA) 18 .8Notexin + Scutoxin I7.0 18 .7 19 .7ATP 17 .4Notexin+ATP I7 .2 19 .4Notexin (Fig . 3F, 3) 19 .7Scutoxin B (Fig . SB, SB) I8 .4

A New Toxin from N. s. scutatus Venom
95
as well as minor protein components, may be essential for the reassociation of thesetoxins, and are perhaps removed during gel filtration . We were surprised to find that ATPelutes from the S-200-HR column at the same volume as notexin and scutoxin A (in peakIIn and that other less acidic adenosine nucleotides, presumably AMP and ADP, elutewith notexin and scutoxin B in peak IV . Again, preliminary studies suggest that there is noassociation between purified notexin and ATP. While the presence of nucleosides andnucleotides in snake venoms has been noted before (see review by BIEHBR, 1979), the ratherlarge amount of nucleotides found in N. s. sruratus venom was unexpected . Gel filtrationfractions from Bothrops riper venom all have 260/280 nm absorbance ratios of less than 1,suggesting the absence of nucleosides (unpublished data).The scutoxin isoforms have different charges but both are less basic than notechis II-5
and notexin. Their phospholipase activities in our Triton X-100-containing reactionmixture are higher than notexin but less than notechis II-5, while their toxicities are higherthan notechis II-5 and about the same as notexin. Like notechis II-5 and notexin, they arerecognized by a monoclonal antibody raised against pseudexin. These findings suggestthat scutoxin is a protein which is similar to the other phospholipase active proteins in theN. s. scutatus venom. A western blot of venoms from several other Notechis subspeciesusing the pseudexin monoclonal antibody has shown that scutoxin may be present insmall quantities in N. ater humphreysi, but is absent from N. ater serventyi and N. ater ater(JoxN and KA1sElt, 1990) . Thus scutoxin is a venom component which may be foundprincipally in N. s. scutatus . An unresolved question is why N. s. scutatus should secretemultiple proteins with phospholipase activity? Further experiments are being conducted toestablish the type of toxicity exhibited by scutoxin to determine if it is different fromnotexin and notechis II-5 which are presynaptic neurotoxins.
Scutoxin is clearly different from notechis II-1 as shown by the toxicity, phospholipaseactivity, and peptide mapping results. Notechis II-1 was not purified in this study. ELISAsshowed the presence of a component in the Mono S flow through of 5-200 peak II whichwas antigenic to the pseudexin antibody that cross-reacted with notechis II-5, notexin andscutoxin . Since notechis II-1 is highly homologous to notechis II-5, notechis II-1 may belocated in this fraction .
Note added in proofDrs Jas Sct~r and Joruv Mronrsaxoog (Pathology Division, USAMRIID, Frederick, MD, U.S.A .)
sequenced the first 15 N-terminal residues of our peak identified as notexin and found it identical to notexin(Hussar and F.~raue, 1975).
Acknowledgements-The authors thank Dr Jorua Mronr.eaxoorc (Pathology Division, USAMRIID, Frederick,MD, U.S.A .) for his generous gift of pseudezin monoclonal antibody. Supported in part by the U.S . ArmyMedical Acquisition Activity, Contract No. DAMD 17-89-C-9007 .
REFERENCES
Amn, S. D. and Kwtssrt, I. I . (1985x) Comparative atudies on three rattlesnake toxins . Toxicon 23, 361-374.Amn, S. D. and Kw>sEa, I . I . (1985b) Toxicity assays. Tozicon 23, 11-13 .Ausn, S. D., S~+xr, C. S. andKim, I. I. (1988) Preliminary fractionation and characterization of the venom
of the Great Basin rattlesnake (Crotales viridis tetoses) . Herpetotogica 44, 71,85.B~at=x, A. L. (1979) Metal and nonprotein constituents in snake venoms. In : Snake Yeno»u, pp. 295-306 (L~,C. Y., Ed .) . New York : Springer.
nA Su.ve, N. J., Aran, S. D., Sonar, C. and IG~t, I . I . (1989) A gyroxin analog from the venom of thebushmaster (Lachesis meta muta). Toxicon 27, 763-771.

96
B. FRANCIS et a!.
HALPERT, J. and E~x>:re, D., (1975) Amino acid sequence of a presynaptic neurotoxin from the venom ofNotechis scutatus scutatus (Australian tiger snake) . J. biol. Chem . 250, 6990997.
HALPBRr, J. and EAxEß, D., (1976x) Isolation and amino acid sequence of a neurotoxic phospholipase from thevenom of the Australian tiger snake Notechis scutatus scutatus . J. biol. Chem . 251, 7343-7347.
H~t.r>atr, J. and Eext :x, D. (1976b) Isolation of a non-neurotoxic, non~nrymatic phospholipase A homologfrom the venom of the Australian tiger snake Notechis scutatus scutatus. FEBS Lett. 71, 91-95.
HALPERT, J., FOI~aurr, J. and Entct:a, D., (1979) Amino acid sequence of a postsynaptic neurotoxin from thevenom of the Australian tiger snake Notechis scutatus scutatus . Biochimie 61, 719-723.
Jottx, T. R. and K~rsnrt, I. L, (L990) Comparison of venom constituents from four tiger snake (Notechis)subspecies. Toxicon 28, 1117-1122.
K~sne, I. L, Gur¢xxFZ, J. M., Pt,UUa`ffx, D., Autn, S. D. and On>a.L, G. V. (1990) The amino acid sequence ofa myotoxic phospholipase from the venom of Bothrops riper. Arch . Biochem. Biophys. 278, 319-325.
KnntssoN, E., E~tc©e, D. and Rm$rt, L. (1972) Purification of a presynaptic neurotoxin from the venom of theAustralian Tiger snake Notechis scutatus scutatus. Toxicon 10, 405-413.
Ltrm, P., and E~tcert, D., (1980) Complete amino-acid sequence of a non-neurotoxic, non~nzymatic phospholi-pase AZ homolog from the venom of the Australian tiger snake Notechis scutatus scutatus . Eur. J. Biochem.111, 403-~09.
Mtnnt.eeteoox, J. (1990) Monoclonal antibodies against pseudexin. III Symposium on Animal, Plant andMicrobial Toxins. IST Pan-American Section, Oaxtepec, Mexico, Jan. 9-12, p. 43 .
Sct~unr, J. J. and Mronteeaootc, J. L. (1989) Purification, sequencing and characterization of pseudexinphospholipases A2 from Pseudechis pxrphyriacus (Australian red-bellied black snake) . Toxicon 27, 805-818.
True, P., Mtzustanan, S., LOWRY, C. V. and No~ttrne, M. (1971) Reconstitution of ribosomes fromsubribosomal components. Methods in Enzymology 20, 39I~07 .





























