Embed Size (px)
Citation preview

1
New lager brewery strains obtained by crossing techniques using cachaça 1
(Brazilian spirit) yeasts 2
3
Bruna Inez Carvalho Figueiredo1, &
, Margarete Alice Fontes Saraiva1, &
, Paloma Patrick de 4
Souza Pimenta1, Miriam Conceição de Souza Testasicca
2, Geraldo Magela Santos 5
Sampaio1, Aureliano Claret da Cunha
1, Luis Carlos Crocco Afonso
2, Marisa Vieira de 6
Queiroz3, Ieso de Miranda Castro
1, Rogelio Lopes Brandão
1# 7
8
1Laboratório de Biologia Celular e Molecular,
2Laboratório de Imunoparasitologia, Núcleo 9
de Pesquisas em Ciências Biológicas, Universidade Federal de Ouro Preto, Ouro Preto, 10
Minas Gerais, Brazil, 3Laboratório de Genética de Microrganismos, Departamento de 11
Microbiologia, Universidade Federal de Viçosa, Viçosa, Minas Gerais, Brazil. 12
13
&These authors contributed equally to this work. 14
15
#Correspondence should be addressed to Rogelio Lopes Brandão at Laboratório de 16
Biologia Celular e Molecular, Núcleo de Pesquisas em Ciências Biológicas, Universidade 17
Federal de Ouro Preto, Campus do Morro do Cruzeiro - 35.400-000. Ouro Preto, MG, 18
Brazil. Email: [email protected]. Tel: +55 31 3559 1680. 19
20
Running title: Lager brewery yeasts obtained by crossing techniques 21
22
AEM Accepted Manuscript Posted Online 4 August 2017Appl. Environ. Microbiol. doi:10.1128/AEM.01582-17Copyright © 2017 American Society for Microbiology. All Rights Reserved.
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

2
ABSTRACT 23
The development of hybrids has been an effective approach to generate novel yeast strains 24
with optimal technological profile for use in beer production. This study describes the 25
generation of a new yeast strain for lager beer production by direct mating between two S. 26
cerevisiae strains isolated from cachaça distilleries: one strongly flocculent and other with 27
higher production of acetate esters. The first step toward this procedure was to analyze the 28
sporulation ability and reproductive cycle of the strains belonging to a specific collection of 29
yeasts isolated from cachaça fermentation vats. Most strains showed high rate of 30
sporulation, spore viability, and homothallic behavior. In order to get new yeast strains with 31
desirable properties useful for lager beer production, we compare haploid-to-haploid and 32
diploid-to-diploid mating procedures. Moreover, assessment of parental phenotype traits 33
showed that the segregant diploid C2-1d generated from diploid-to-diploid mating 34
experiment showed good fermentation performance at low temperature, high flocculation 35
capacity and desirable production of acetate esters that was significantly better than one 36
type lager strain. Thereby, the strain C2-1d might be an important candidate for the 37
production of lager beer with distinct fruit trace, originated from a non-GMO approach. 38
39
Keywords: Aroma volatile compounds; Beer; Flocculation; Hybrid; Mating type. 40
41
42
43
44
45
46
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

3
IMPORTANCE 47
48
Recent work has suggested the utilization of hybridization techniques for generation of 49
novel non-genetically modified brewing yeast strains with combined properties, not 50
commonly found in a unique yeast strain. We have observed remarkable traits, especially 51
low temperature tolerance, maltotriose utilization, flocculation ability and production of 52
volatile aroma compounds, among a collection of Saccharomyces cerevisiae strains isolated 53
from cachaça distilleries, which allow their utilization in the production of beer. The 54
significance of our research is in the use of breeding/hybridization techniques to generate 55
yeast strains that would be appropriate to produce new lager beers by exploring the capacity 56
of cachaça yeast strains to flocculate and to ferment maltose at low temperature with the 57
concomitant production of flavoring compounds. 58
59
60
61
62
63
64
65
66
67
68
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

4
INTRODUCTION 69
Alcoholic fermentation is a crucial step in the production of different beverages, 70
including beer, wine and spirits; in all these examples, it is carried out mainly by yeast. 71
There is quite a large variety of different yeast species that are able to ferment sugar into 72
ethanol; among them, those belonging to the genera Saccharomyces are used for bioethanol 73
production and dominate also wine, spirits and beer industry (46). 74
In general, to produce a high-quality beer, yeast must be effective in fermenting the 75
sugars from the wort (maltose, maltotriose and glucose); be able to tolerate high ethanol 76
levels and to produce desired aroma compounds (5). Moreover, two main beer styles can be 77
distinguished according to the species of yeast used: ale and lager beers. Ale beers are 78
produced by strains of S. cerevisiae, but lager beers are traditionally obtained by yeast 79
species that result from hybridization events between S. cerevisiae and non-cerevisiae that 80
were selected in conditions of low-temperature fermentations for beer production (23). 81
Beyond that, lager yeast also presents another interesting feature: the ability to flocculate 82
that leads cells to the setting at the bottom of the fermentation tank, enabling easy removal 83
and re-inoculation; in contrast, ale yeast rises to the surface (46). 84
An additional and important difference between lager and ale strains is related to the 85
capacity to produce different types and levels of flavoring compounds. Ale yeasts produce 86
higher concentrations of esters and higher alcohols, while lager yeasts are acknowledged by 87
their “clean flavor profile”, i.e. lack of ester-derived fruity or floral aroma (21). However, 88
there are two distinct genotypes of lager yeasts, designed Saaz and Frohberg (24). Saaz-89
type strains produce lower concentrations of aroma compounds like ethyl acetate, 3-90
methylbutanol and 3-methylbutyl acetate than the more aroma-rich Frohberg yeasts (14). 91
This phenotypic difference resulted in the diversified and differentiated aroma of lager 92
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

5
beers in the market and also would explain the preference for Frohberg type lager yeasts 93
over Saaz-type lager yeasts in beer industry (26). 94
Volatile acetate esters, such as 3-methylbutyl acetate and ethyl acetate, are 95
considered one of the most important groups of aroma-active yeast metabolites because 96
they are responsible for the highly desired fruity, candy and floral character of beverages 97
(41). They are produced by an enzyme-catalyzed condensation reaction between acyl-98
coenzyme A (CoA) and a higher alcohol (38), and small changes in the concentration of 99
these secondary metabolites can have large effects on the sensorial quality of beer (34). 100
Moreover, higher alcohols, like 3-methylbutanol, also contribute to the final quality of 101
different beverages, including beers. Higher alcohols are formed by anabolism or 102
catabolism (Ehrlich pathway) of amino acids (31). As esters are synthesized through higher 103
alcohols and acyl-CoA, the ratio of levels of higher alcohols/esters may affect the beer 104
quality (22). 105
Interestingly, in our previous studies, we have been able to demonstrate a 106
remarkable large phenotypic diversity among a collection of S. cerevisiae strains isolated 107
from fermentation vats during the production of cachaça (the Brazilian spirit that is usually 108
obtained from the distillation of fermented sugarcane juice). Our culture collection was 109
phenotyped on different properties for bioethanol production, including the ability to 110
tolerate ethanol, methanol, aluminum, zinc, different pH values, 5-HMF stress as well as 111
foam production (8). Additionally, different flocculation traits have been observed in yeast 112
strains isolated from fermentation vats during the production of cachaça. Among these 113
strains, we have found some that present a higher flocculation rate and cellular growth at 114
low temperature (1) and other strains that present distinct properties that allow their 115
utilization in the production of beer (M.T. de Souza and T.M. Araújo, unpublished data). 116
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

6
Taking into account all these data, we have decided to use our yeast collection to try 117
to generate yeast strains that would be appropriate to produce lager beers by exploring the 118
capacity of some yeast cachaça strains to flocculate and to be able to ferment maltose at 119
low temperatures with the concomitant production of flavoring compounds. To accomplish 120
this goal, there are different strategies and the choice among them is based on the genetic 121
nature of traits (monogenic or polygenic), the knowledge of the genes involved and 122
phenotypic trait requirement (4, 16). Moreover, because of the complex legislation and 123
consequently the lack of acceptance for the use of recombinant yeast (genetically modified 124
organism - GMO) for food production, only classical techniques such as selection of 125
variants, random mutation, and mating/hybridization are used to produce food-grade starter 126
cultures (33, 37). 127
In our case, the generation of new lager yeasts would require the combination in one 128
single yeast strain the capacity to produce volatile aroma compounds and the flocculation 129
ability. In the present study, we described the use of mating techniques to generate yeast 130
hybrid from a strongly flocculent S. cerevisiae strain and other S. cerevisiae with high 131
acetate esters production. The hybrids were tested for their flocculation ability, maltose 132
fermentation, and production of flavoring compounds in conditions of wort fermentation 133
performed at lower temperatures that are typical for lager yeasts. Therefore, the aims of this 134
work were: (a) to classify the strains of our culture collection regarding their reproductive 135
cycle (hetero or homothallism), as well as their sporulation rates; (b) to use this data as tool 136
to select strains/segregants for further phenotypic improvement using hybridization 137
strategies (direct mating); (c) to compare two mating hybridization techniques using a 138
flocculent strain unable to ferment maltose with another able to ferment maltose and to 139
produce higher concentrations of flavoring compounds strains by using haploid-to-haploid 140
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

7
mating and diploid-to-diploid mating; (d) to test the resulting hybrids for production of beer 141
by comparing the results obtained with classical reference brewery strains. 142
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

8
MATERIALS AND METHODS 143
Strains and growth conditions 144
The yeast strains used in this study were previously isolated from cachaça 145
distilleries according the methodology described before (2, 9, 30, 43). They are stored in 146
YP broth (1% (wt/vol) yeast extract, 2% (wt/vol) meat peptone) added of 30% (vol/vol) 147
glycerol at -80 ºC. All strains are available in the culture collection of the Federal 148
University of Ouro Preto ([email protected]). The commercial strain Saccharomyces 149
pastorianus var. Weihenstephan 34/70 (tetraploid) (Fermentis, Germany) and the laboratory 150
strains Saccharomyces cerevisiae BY4743 (MATa/α, diploid) and BY4742 (MATα, 151
haploid) (both purchased from EUROSCARF, Germany) were used as controls. 152
Yeast cells were grown in YPD broth containing 1% (wt/vol) yeast extract, 2% 153
(wt/vol) meat peptone, 2% (wt/vol) glucose and incubated at 30 ºC with 200 rpm in orbital 154
incubator New Brunswick Model G200 (New Brunswick Scientific, New Jersey, USA). 155
156
General molecular biology techniques 157
Yeast genomic DNA was extracted as previously described (25). PCRs were 158
performed using thermal cycle equipment T100TM
(Bio-Rad Laboratories, California, USA) 159
and for diagnostic purposes was used GoTaq® DNA polymerase (Promega, Wisconsin, 160
USA) according to manufacturer’s protocols. PCR products were visualized using an Alpha 161
Imager Mini System (Alpha Innotech Corporation, San Leandro, USA) after electrophoretic 162
separation at 6 V.cm-1
in a 1.2% (wt/vol) agarose gel stained with GelRedTM
Nucleic Acid 163
(Biotium Inc, Fremont, USA). The primers were synthesized by Integrated DNA 164
Technologies (IDT Corporation, Newark, USA). Yeast strains were transformed using the 165
lithium acetate/polyethylene glycol method (15). 166
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

9
To perform the molecular identification of the species used in this work, internal 167
transcribed spacer (ITS) regions (ITS1/5, 8S/ITS2) were amplified, and the amplicon (880 168
pb) was analyzed by restriction fragment length polymorphism (RFLP) using HhaI, HaeIII, 169
and HinfI and then compared with a control correspondent sequence originating from S. 170
cerevisiae BY4741 (EUROSCARF, Germany). ITS PCR products were purified and 171
sequenced by capillary electrophoresis (Sanger method) using the ABI3130 platform Life 172
Technologies (Myleus Biotechnology, Belo Horizonte, Minas Gerais, Brazil). Sequences 173
were analyzed using the National Centre for Biotechnology Information (NCBI) database. 174
175
Sporulation, tetrad dissection and spore viability 176
Yeast cells (128 isolates) pre-grown in YPD broth for 24 h at 30 ºC, 200 rpm, were 177
centrifuged at 1000 x g, 5 min, 4 ºC (AllegraTM
X-12R centrifuge, SX4750/SX4750A 178
rotors, Beckman Coulter Life Sciences, California, USA); the pellets were washed twice 179
with cold sterile water. Cells were then spot-inoculated on sporulation medium containing 180
1% (wt/vol) potassium acetate, 0.05% (wt/vol) potassium bicarbonate, 1.5% (wt/vol) agar, 181
pH 6.0 and incubated at 23 ºC for 5-10 days until asci were visible microscopically. The 182
ascus wall was digested with 5 µl of the lyticase solution (5000 U mL-1
lyticase, 1 M 183
sorbitol, 0.1 M Na3-citrate, 0.06 M EDTA and 0.14 M β-mercaptoethanol) and incubated 184
for 5 min at room temperature. Approximately, forty single-spore cultures from ten tetrads 185
were dissected using a micromanipulator MSM System 400 (Singer Instruments, 186
Roadwater, United Kingdom), on YPD-agar (1% (wt/vol) yeast extract, 2% (wt/vol) meat 187
peptone, 2% (wt/vol) glucose, 1.5% agar) and incubated at 30 ºC for 3-5 days. The spores 188
that formed visible colonies were designed viable. Spore viability was determined as the 189
number of viable spores divided by the total number of spores seeded. 190
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

10
The sporulation frequency and efficiency were evaluated by microscopic 191
observation and quantified according to the methodology already described (11). The 192
sporulation frequency was calculated after counting the number of asci (dyads, triads, and 193
tetrads) in a total cell population of at least 300 (total asci divided by sum of cells and asci 194
counted). The sporulation efficiency was calculated as the sum of tetrads divided by the 195
total number of asci. 196
197
Mating type characterization 198
The heterothallic and homothallic nature of the yeast strains was determined by 199
mating type assaying of all clones of cells produced by viable spores originated from four 200
different tetrads. Mating type was analyzed by PCR as previously demonstrated (17). PCR 201
was carried out directly from colonies of yeast using combinations of three specific primers 202
to MATlocus, MATa and MATα listed in Table 1, under the following conditions: 4 min at 203
94 °C; followed by 30 cycles of 1 min at 94 °C, 2 min at 58 °C, and 1 min at 72 °C; with a 204
final 10 min at 72 °C. 205
206
Determination of ploidy by flow cytometry 207
The ploidy of each parental and hybrid strain was determined fluorescently labeling the cell 208
nucleic acid and analyzing the fluorescence by flow cytometry as described (21) with some 209
modifications. Cells were grown for eight hours in YPD broth centrifuged (3269 x g, 10 210
min, 4 ºC) and washed with PBS buffer (137 mM NaCl, 2.7 mM KCl, 10 mM Na2HPO4, 2 211
mM KH2PO4, pH 7.4) and resuspended in 250 mM EDTA diluted in the PBS buffer. A 212
sample of 2 x 107 cells were fixed with 70% (v/v) cold ethanol and maintained overnight at 213
4 °C. Cells were centrifuged, resuspended in Tris-HCl buffer (50 mM Tris-HCl, 15 mM 214
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

11
MgCl2, pH 7.7), RNase (0.5 mg mL-1
, Sigma-Aldrich, Missouri, USA) and incubated 215
overnight at 37 °C. Cells were treated with papain and proteinase K (1 mg mL-1
, Sigma-216
Aldrich, Missouri, USA) for 1 h at 37 °C and 2 h at 50 °C, centrifuged and ressupended in 217
PBS buffer. One hour before analysis, cells samples were stained with 2 µM SYTOX®
218
Green (Thermo Fisher Scientific, Massachusetts, USA). Cells were analyzed using a BD 219
LSR Fortessa™ cell analyzer (Becton Dickinson Bioscience, New York, USA) and 220
detected at 488 nm (FITC-A). DNA content relative was estimated based on the 221
fluorescence intensity compared with the yeast reference strains (S. pastorianus var. 222
Weihenstephan 34/70 – tetraploid; S. cerevisiae BY4743 - MATa/α, diploid; and BY4742 - 223
MATα, haploid) and plotted to exponential scale. 224
225
Electrophoretic karyotype analysis 226
Chromosomal DNA was prepared as described before (42), with minor 227
modifications. Yeast strains were grown on YPD broth at 30 °C to stationary phase. 228
Approximately 4 x 108 cells mL
-1 were washed with 50 mM EDTA pH 8.0, resuspended in 229
lyticase buffer (5000 U mL-1
lyticase, 1 M sorbitol, 0.1 M Na3-citrate, 0.06 M EDTA and 230
0.14 M β-mercaptoethanol) and incubated at 37 °C for 3 hours. An aliquot of this 231
suspension was mixed with an equal volume of 1% (wt/vol) Certified™ low melting 232
agarose (Bio-Rad Laboratories, California, USA) in the buffer (0.01 M EDTA, 0.05 M 233
Tris-HCl, pH 8.0) and distributed into wells of plugs to solidification. The plugs were 234
transferred to 1 mL of a solution containing 1 mg mL-1
proteinase K, 0.45 M EDTA, pH 8.0 235
and 1% (wt/vol) SDS, incubated overnight at 50 °C, washed with 0.5 M EDTA, pH 8.0 and 236
stored at 4 °C in this last solution. Before use, agarose plugs prepared were washed two 237
times for 30 min at 50 °C in TE buffer (0.01 M Tris-HCl, 0.001 M EDTA, pH 8.0), two 238
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

12
times for 30 min at room temperature in the same buffer and inserted in the wells of a 1% 239
(wt/vol) agarose gel. Electrophoresis was performed using CHEF DR-III pulsed field 240
system (Bio-Rad instruments, California, USA) under 0.5 X TBE (45 mM Tris, 45 mM 241
boric acid, 1.0 mM EDTA, pH 8.3) recirculated, and cooled at 10 °C in the following 242
conditions: 18 h with a switching time of 60 s, 6 h with a switching time of 35 s and a 243
constant current at 150 mA. 244
245
ho deletion mutants of Saccharomyces cerevisiae LBCM92 246
The expression of the HO gene results in a MAT locus change (MATa to MATα or 247
the other way around) due to the activity of a specific endonuclease Ho. The HO gene was 248
deleted in LBCM92 strain using separately two cassettes conferring resistance to geneticin 249
and to hygromycin in order to delete two copies of the same gene. The deletion cassettes 250
were PCR-amplified from pJET1,2-attB-KanMX-attP and pJET1,2-attB-hph-attP (provided 251
by Laboratory of Molecular Cell Biology, University of Leuven, Belgium) with the 252
corresponding pdel fw and pdel rv primers (Table 1) using a Phusion® High-Fidelity DNA 253
Polymerase (New England BioLabs Inc., Massachusetts, USA). The primers contain 254
sequences of 20 bp that flaking the cassettes and sequences with homology to HO gene 255
(Table 1). 256
PCR were performed in 50 µL and contained 10 ng of template plasmids, 2.5 mM of 257
each dNTP, 50 pmol of each primer, and 2.0 units of Phusion DNA Polymerase and buffer 258
according to the manufacturer’s instructions for use. The PCR thermal cycle program 259
included an initial denaturation at 94 °C for 3 min, followed by 5 cycles, with a 260
denaturation step at 94 °C for 1 min, an annealing step of 1 min at 62 °C, an elongation step 261
at 72 °C for 2 min, and 30 cycles, with a denaturation step at 94 °C for 1 min, an annealing 262
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

13
step of 30 sec at 71,5 °C and extension step during 2 min at 72 °C. Final extension was 263
performed at 72 °C for 5 min. The desired band after PCR amplification was cut from the 264
gel and purified with NucleoSpin® PCR clean up and Gel extraction kit (Macherey-Nagel, 265
Duren, Germany). Subsequently, 50 µL of PCR product was used for transformation of the 266
strain LBCM92. After heat shock (42 °C), the cells were centrifuged 1500 x g, 2 min 267
(5415D centrifuge, Eppendorf, Hamburg, Germany), recovered in YPD medium for 4 hours 268
(30 °C, 200 rpm) and plated on YPD-agar containing either 400 µg mL-1
of geneticin 269
(Sigma-Aldrich, Missouri, USA) or 600 µg mL-1
of hygromycin (Invitrogen, Thermo 270
Fisher Scientific, Massachusetts, USA). The first transformation was performed using 271
geneticin cassette, and a selected transformant was used in the second transformation with 272
hygromycin cassette. 273
274
Mating type switching in the Saccharomyces cerevisiae LBCM78 and LBCM92 strains 275
To obtain yeast diploid strains (MATa/a or MATα/α) the mating type was switched 276
using a CEN plasmid (pFL39GAL1HOKanMX) containing a functional HO open reading 277
frame (ORF) controlled by GAL1 promoter (7). Yeast strains were transformed with 1 µg of 278
plasmid and after heat shock (42 °C), the cells were centrifuged (2 min, 1500 x g), 279
recovered in YPD medium for 4 hours (30 °C, 200 rpm) and plated on YPD-agar with 400 280
µg mL-1
of geneticin. Yeast cells transformed were grown in YPD broth with geneticin 281
(400 µg mL-1
) overnight at 30 °C. The cells were centrifuged and resuspended in YP broth 282
supplemented with 2% (wt/vol) galactose, geneticin (400 µg mL-1
) and incubated at 30 °C. 283
After 1 hour of growth was added 2% (wt/vol) glucose. Cell samples were collected each 284
hour during a period of 12 hours of incubation and plated in YPD-agar to obtain single 285
colonies. Mating type of each single colony was verified by PCR according described 286
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

14
above. Yeast cells presenting mating type a/a (originated from LBCM92 strain) and α/α 287
(obtained from LBCM78 strain) were selected and cultivated in YPD broth without 288
geneticin for 48 h to induce plasmid curing. 289
290
Direct mating 291
The diploid strains resulting from the mating type switching of the parental strains 292
(LBCM92 MATa/a and LBCM78 MATα/α) as well as the haploid segregants of both 293
original parental strains (LBCM92 and LBCM78) presenting opposite mating types were 294
streaked on YPD-agar. One colony of each strain of opposite mating type was mixed over 295
the surface of YPD-agar plate with ten microliter of sterile water. The plate was dried and 296
incubated for 48 h at 37 °C. A small sample of the spots was serially diluted and plated in 297
YPD-agar. After 48 h of incubation, a few colonies resulting from the crossing were 298
analyzed by mating type PCR and flow cytometry to confirm the poidy. Furthermore, in 299
order to obtain segregant diploids, the tetraploids strains were inoculated on solid 300
sporulation medium as described before. 301
302
Evaluation of cachaça yeast strains for beer production 303
To evaluate the selected cachaça yeast strains for their use in beer production, we 304
tested them for maltose/maltotriose fermentation, cold tolerance, production of phenolic 305
compounds and flocculation capacity. 306
Maltose fermentation was carried out in tubes with YP broth added of 2% (wt/vol) 307
maltose containing inverted Durham vials in order to detect gas production. Maltotriose 308
fermentation by the yeast strains was assayed by growing them in YP both-2% maltotriose 309
in the absence or presence of antimycin A (2 mg L-1
, Sigma-Aldrich, Missouri, USA) at 30 310
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

15
°C for 72 h (47). Cold tolerance was evaluated by yeast grown on YPD broth at 12 ºC. 311
Yeast growth was monitored following A600nm in a SpectraMax 340PC384 microplate 312
Reader using a Soft Max® Pro software (Molecular Devices, California, USA). The growth 313
rate for each strain at the exponential growth phase was determined by linear regression of 314
the plot ln A600nm versus time (h). 315
Phenolic compound (4VP) was analyzed using the protocol described before (40) 316
with modifications. The yeast strains (initial O.D600 of approx. 0.1) were grown in Yeast 317
Nitrogen Base Broth (Sigma-Aldrich, Missouri, USA) 2% (wt/vol) glucose supplemented 318
with 1 mM ferulic acid at 25 ºC for 5 days. One milliliter of supernatant sample was 319
collected and diluted with an equal volume of methanol. Sample was filtered through 0.25 320
µm pore membrane (Millipore, Massachusetts, USA) and 30 µL was injected into HPLC 321
N02104 (Shimadzu, Kyoto, Japan) equipped with a degasser (DGU-20A5), a binary pump 322
(LC-20AT) and Shimp-pack CLC-ODS column (particle diam. 5 µm, 25 cm x 4.6 mm). 323
The fingerprint was recorded at a wavelength of 260 nm by using a diode array detector 324
(SPD-M20A). A linear gradient of the solvent A, 0.5% (vol/vol) acetic acid in water, and 325
solvent B (100% acetonitrile) was used in the following manner at a flow rate of 1 mL min-
326
1 at 40 ºC: 15 min, 5-20%; 15-40 min, 20-40%; 40-50 min, 40-5%. 327
Flocculation was evaluated by formation of settling cell flocs after growth of yeast 328
in YPD broth at 30 °C for 48 h. Flocculation was quantified using a modified Helm’s assay 329
as described before (1). Yeast strains were grown in YPD broth for 48 h (30 °C, 200 rpm) 330
and washed twice with sterile water. The flocculation test was carried out with 331
approximately 108
cells (1 mL of cell samples at D.O600 of approx. 0.4) washed with CaSO4 332
solution (0.51 g L-1
). Cells in test tubes were harvested by centrifugation and stirred 333
vigorously in vortex, for 30 s, in the solution containing CaSO4 (0.51 g L-1
) sodium acetate 334
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

16
(6.8 g L-1
), acetic acid (4.05 g L-1
) and 4% (vol/vol) ethanol, pH 4.5. Yeast cells in control 335
tubes were resuspended in 0.5 M EDTA (pH 7). After a sedimentation period of 15 min, 336
samples were taken from just below the meniscus and diluted 10X, then the absorption at 337
600 nm was measured. Six replicates were performed for each sample of test tubes, and the 338
extent of flocculation was expressed as the mean percentage using the following formula: 339
Flocculation (%) = 100 × [A600 (control) - A600 (sample)] /A600 (control) 340
341
Screening of haploid segregants of Saccharomyces cerevisiae LBCM78 strain for 342
volatile aroma compounds 343
Yeast pre-grown in 3 mL of YPD broth at 30 ºC overnight was used to inoculate 50 344
mL of YP 250 broth (0.27% (wt/vol) yeast extract, 0.54% (wt/vol) meat peptone, pH 4.5) 345
with 5% (wt/vol) glucose. This pre culture was used for inoculation on YP 250 broth with 346
10% (wt/vol) glucose on fermentation tubes. Batch fermentations were started at 347
approximately 5 x 106 cells mL
-1 and incubated statically at 30 °C for 4 days. The 348
headspace of fermentation samples was taken immediately after the end of batch to prevent 349
evaporation of volatiles. 350
The quantification of volatile aroma compounds was analyzed by gas 351
chromatography coupled with flame ionization detection (CP-3380, Varian) standardized 352
for volatile acetate esters and higher alcohols; n-pentanol was used as internal standard. The 353
GC was equipped with a DB - WAX polietilenoglicol column (60m x 0,25mm x 0,50µm) 354
(J&W, Albany, New York) and the gas used were N2 (gas make up): 29 mL min-1
, H2: 30 355
mL min-1
e Ar: 300 mL min-1
. Samples of 5 mL were collected in glass tubes with 2% (v/v) 356
n-pentanol each. After heating for 30 min at 60 °C, 5 mL of headspace fraction was 357
collected with syringe gastight e injected in the splitless mode (2 min) at 225 ºC. The 358
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

17
temperature program was 50 ºC/min, then 50 °C to 100 ºC, 5 ºC/min to 100 ºC for 3 min, 359
100 °C to 250 ºC at 30 ºC/min and 250 ºC for 3 min and detected at 280 ºC. Compound was 360
quantified by the ratio between the peak area of each sample and the peak area of internal 361
standard using a standard curve. 362
363
Laboratory-scale lager-like fermentations 364
Hybrid and parental strains were tested in mimicked lager fermentations, performed 365
in wort-like medium consisted of dry malt extract (Brewferm® 8 EBC, Belgium) 16% 366
(wt/vol) - 13.2º Brix. The yeasts were pre-cultivated in 3 mL of YPD medium at 30 °C for 367
16 h. Next, a second pre-cultivation was executed consisting of the inoculation of 1 mL of 368
the first pre-culture also in 50 mL of YPD, but now presenting 5% (wt/vol) glucose and 369
incubation at 30 °C for 48 h. These cultures were used for inoculation of the laboratory-370
scale fermentation at an OD600 of 5.0, approximately 5 x 107 cells mL
-1 on tubes (5.5 x 14 371
cm, handmade at Katholieke Universiteit Leuven, Belgium) without shaking (to simulate an 372
anaerobic condition) at 12 °C for 11 days. All fermentations were performed in triplicates. 373
Weight loss was measured to verify fermentation progress and the quantification of 374
volatiles compounds produced was analyzed as described above. 375
376
Statistical analysis 377
Data from fermentation trials and production of volatile aroma compounds were 378
analyzed using the D’Agostino and Pearson omnibus normality test followed by one-way 379
ANOVA with post hoc Tukey’s test. The statistical level of significance was set at P < 380
0.05. 381
382
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

18
RESULTS 383
Sporulation, spore viability and identification of the reproductive cycle 384
In order to develop a strategy for yeast breeding, we initially studied the sporulation 385
rate, spore viability and sexual cycle of yeast strains belonging to a collection obtained 386
from cachaça distilleries. Figure 1 shows the percentage of frequency and efficiency of 387
sporulation as well as the viability of the spores obtained from yeast strains isolated from 388
cachaça distilleries. In general, the major part of the isolates (ninety-six strains) displayed 389
percentage of sporulation above 50% (Fig. 1A). However, fourteen of the analyzed strains 390
were unable to sporulate in the media tested. All strains that presented sporulation were 391
able to form asci with four spores, although in some cases, the number of asci with two 392
spores (dyads) was relatively high resulting in low efficiency of the sporulation (Fig. 1B). 393
Twenty-four yeast strains showed between 90 and 100% of viability, while seven strains 394
presented non-viable spores (Fig. 1C). For these yeast strains and the strains that presented 395
low spores viability, it was necessary to dissect a large number of tetrads and keep spores 396
growing during 5-8 days in order to obtain viable samples. 397
All strains that sporulate were examined through tetrad analysis in order to verify 398
the sexual life cycle. The four spores of one tetrad were selected from each strain and the 399
mating type was determined by PCR. A single band showed that the spores were haploid 400
(MATa, generating a 544 bp product, or MATα, generating a 404 bp product) and two 401
bands illustrated that the spores were diploid (MATa/α) and homothallic yeast (Fig. 2A). 402
Only three strains (LBCM73, LBCM78 and LBCM115) demonstrated a clear heterothallic 403
pattern (two segregants MATa and two segregants MATα) (Fig. 2A, lines 2-16). All other 404
yeast strains examined are homothallic; as one example we have strain LBCM92 (Fig. 2A, 405
lines 17-21). 406
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

19
Selection of yeast strains to mating procedures 407
As stated before, our intention was to generate new yeast strains with characteristics 408
of lager yeasts that require the combination in one single strain the capacity to ferment at 409
low temperatures, to present a constitutive flocculation and with production of flavoring 410
compounds. 411
The first strain chosen for crossing was LBCM78, because this strain has already 412
been identified as a good producer of flavoring compounds as demonstrated in a recent 413
work (2). Besides, it is able to ferment maltose, with a high frequency of sporulation 414
capacity, reasonable spore viability and with a typical heterothallic life cycle (Fig. 1 and 415
Fig. 2A, lines 7-11). After sporulation and tetrad analysis, 23 segregants were screened to 416
identify those that showed higher production of flavoring compounds. From these 417
segregants, eight were randomly phenotyped in small-scale fermentation, with subsequently 418
volatile compounds analysis, and one segregant (78-2b) produced a concentration of 3-419
methylbutyl acetate and the ratio between 3-methylbutyl acetate and 3-methylbutanol 420
similar to its parental strain (LBCM78) (Fig. 3). The mating type of the 78-2b strain was 421
identified as “α” (Fig. 2A, line 9). 422
Among all flocculent strains in the yeast cachaça that we have selected, LBCM92 423
was the strain that presented the highest level of flocculation, confirming data obtained 424
before (1). Nevertheless, this strain presents a sporulation frequency of 10% and it is unable 425
to ferment maltose. In addition, strain LBCM92 is homothallic, generating haploid 426
segregants with unstable mating types, with four spores of all analyzed tetrads by PCR 427
showing two bands, MATa/α (Fig. 2A, lines 17-21). Therefore, as our aim was to compare 428
two mating hybridization techniques, we constructed the LBCM92 strain with deletion of 429
two copies of HO gene. By this way, six transformants were obtained (data not shown). 430
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

20
One of these transformants, identified as 92hoΔ-IIaD, was selected and after sporulation, 431
tetrads were dissected and the stability of all haploid segregants was confirmed. Figure 2A, 432
lines 23-26 shows mating type analysis of four spores of one of these tetrads dissected. In 433
total, 49 segregants were obtained. From these, 17 segregants showing mating type “a” 434
were subsequently evaluated for their flocculation ability. Among these segregants eight 435
(92hoΔ-2b, 92hoΔ-2f, 92hoΔ-9f, 92hoΔ-11c, 92hoΔ-11e, 92hoΔ-16f, 92hoΔ-17d and 436
92hoΔ18c) showed intermediate flocculation ability, or cells fully flocculent compared to 437
the parental LBCM92 (Table S1). The segregant 92hoΔ-16f was selected for crossing 438
because it showed the highest flocculation rate (99%) (Table 2). 439
The taxonomical identity of LBCM78 and LBCM92 strains were previously 440
confirmed by amplification and sequencing of both ITS region. The obtained sequences 441
presented in both cases up to 95% identity to the corresponding region from S. cerevisiae 442
KDLYS9-5 (GenBank: JN599148.1) (data not shown). These results indicate that both 443
strains are S. cerevisiae. 444
445
Hybrid strains by haploid-to-haploid mating 446
Based on these results, and after confirming the ploidy (Fig. 4), we have crossed the 447
haploids 78-2b (MATα) and 92hoΔ-16f (MATa) to obtain the diploid hybrids. All 448
seventeen hybrids obtained were able to ferment maltose, but only four of them (B16, B22, 449
C28 and C33) presented flocculation ability similar to 92hoΔ-16f (Table S1). The 450
confirmation of the mating type of these hybrids is shown in Figure 2B, lines 6-9. Hybrid 451
B22 was chosen for further studies because it presented a flocculation rate of 70%, although 452
with an inferior value observed in the original 92hoΔ-16f haploid strain (approximately of 453
98%) (Table 2). 454
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

21
Hybrid strains by diploid-to-diploid mating 455
In order to carry out the cross between the diploid S. cerevisiae LBCM78 and 456
LBCM92 (both MATa/α), a procedure that force mating type change in diploid cells was 457
used by which LBCM78 and LBCM92 yeast strains were transformed with 458
pFL39GAL1HOKanMX and grown in medium containing galactose to induce the HO gene 459
expression. Ninety-seven colonies (55 from LBCM78 strain and 42 from LBCM92 strain) 460
were analyzed; and mating type homozygotes was observed in two colonies from LBCM92 461
strain (identified as 92 IX MATa/a and 92 C MATα/α) and four colonies from LBCM78 462
strain (identified as 78 a32 MATa/a, 78 a69 MATa/a, 78 a21 MATα/α, 78 a34 MATα/α) 463
(data not shown). After plasmid curing procedure, it was observed that 78 a34 MATα/α 464
(15) and 92 IX MATa/a (7) strains lost the ability to grow in YPD with 400 µg mL-1
of 465
geneticin. 466
Thus, yeast strains 92 IX (7) MATa/a and 78 a34 (15) MATα/α (Fig. 2C, lines 3 and 467
4) were subsequently chosen for hybridization using the cell-to-cell mating procedure. A 468
total of 33 hybrids from this crossing were isolated and thirty-one were able to ferment 469
maltose (Table S2). Interestingly, none of these hybrids shown to possess the ability of 470
flocculation; even so, eight hybrids (B15, C1, C2, C3, C4, C13, C14 and C16 - Fig. 2C, 471
lines 6-13) were randomly chosen to be sporulated and to generate new diploid segregants. 472
Among the 495 segregants obtained and analyzed, twenty-five recovered flocculation 473
ability, where twelve presented cells fully flocculent (Table S2). However, three segregants 474
(C2-1d, C3-7b and C4-1g) presented a flocculation ratio (around 91-95%) compared to the 475
parental strain – LBCM92 (Table 2). Figure 2C, lines 14-16 shows mating type profile of 476
these three segregants (C2-1d and C4-1g: MATα; C3-7b: MATa). 477
478
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

22
Confirmation of ploidy and hybrid conditions 479
Considering their promising characteristics, yeast strains C2-1d, C3-7b, C4-1g, B22 480
and parental were submitted to ploidy analysis by using Sytox Green staining and flow 481
cytometry to measure the DNA content. All yeast strains generated dual peaks of 482
fluorescence, with the second peak attributed to cells undergoing DNA synthesis (G2/M). 483
Haploid, diploid and tetraploid yeast strains are used as controls being easily 484
distinguishable each other, with cells displaying second peaks respectively of 485
approximately double and quadruple fluorescent levels of the haploid control strain (Fig. 486
4A to 4D). Parental yeast strains (LBCM78 and LBCM92) were confirmed as diploid (Fig. 487
4G and 4L), and their segregants 78-2b and 92hoΔ-16f as haploids as well (Fig. 4F and 488
4K). Hybrid strain B22 originated from the crossing between these two haploid strains gave 489
fluorescent peaks equivalent to diploid genome content (Fig. 4N). Hybrid strains (C2, C3 490
and C4) generated from diploid-to-diploid mating (LBCM78 X LBCM92) displayed a 491
similar profile to that exhibited by the tetraploid control (Fig. 4H, 4I and 4M). Moreover, 492
their segregants (C2-1d, C3-7b and C4-1g) were confirmed as diploids (Fig. 4E, 4J and 493
4O). Furthermore, hybrid status was confirmed by karyotyping and the gel banding patterns 494
are shown in the supplementary material, Fig. S2. The tetraploid hybrid strains C2, C3 and 495
C4 (derivative of diploid-to-diploid mating) presented band of chromosomes from both 496
parental strains and their segregants have maintained several bands of LBCM78 and 497
LBCM92. Although less pronounced, the hybrid character was also shown by strain B22 498
(derivate of haploid-to-haploid mating) that presented bands from parental strains, 78-2b 499
and 92hoΔ-16f as indicated by arrows (Fig. S2). 500
501
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

23
Production of flavoring compounds during fermentation 502
Next, we assessed the potential of the new yeast strains generate in this work to 503
produce aromatic lager beer. The diploid hybrid strain (B22), its ancestral segregant 504
haploids (78-2b and 92hoΔ-16f) and the segregants obtained from tetraploids (C4-1g and 505
C2-1d), as well as the original wild type strains (LBCM78 and LBCM92) were tested in 506
laboratory-scale fermentation mimicking lager style (see Materials and Methods for 507
details). One Frohberg-type lager strain S. pastorianus W34/70 was included as reference 508
strain. 509
Since the fermentation of lager beers takes place at low temperatures, we verified 510
the growth of the new strains in such conditions. Both parental, the hybrid B22, the 511
segregants C2-1d and C4-1g were able to grow on YPD broth at the temperature of 12 °C 512
with a rate of growth of approximately 0.05 h-1
as lager strain W34/70 (Table S3). On the 513
other hand, the segregant strain C3-7b grew poorly on these conditions and its fermentation 514
performance was not evaluated. In addition, the parental (LBCM78, LBCM92, 78-2b, and 515
92hoΔ-16f), hybrid (B22) and segregant (C2-1d and C4-1g) strains did not fermented 516
maltotriose (Table S3). All yeast strains have not shown growth in presence of antimycin A 517
(inhibitor of cell respiration), except the lager strain S. pastorianus W34/70 (Table S3). All 518
yeast strains converted ferulic acid into the phenolic compound (2-methoxy-4-vinilphenol, 519
4VP), and only the strains, C3-7b, B22, C4-1g that produced below of limit detection in the 520
tested conditions (Table S4). 521
Fermentation efficiency was monitored by weight loss generated by CO2 production 522
and the results are shown in Figure 5. The results show that the C2-1d displayed the best 523
performance since it reached the same ethanol production level after 6 days as well as the 524
reference lager strain W34/70 and higher levels than the parental strain LBCM78 (Fig. 5). 525
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

24
On the other hand, hybrid strains C4-1g and B22 showed an inferior performance when 526
compared to their parental strains, LBCM78 and 78-2b, respectively. However, at the end 527
of fermentation, all hybrid strains showed similar ethanol levels but below to the lager 528
strain (Fig. 5). As expected, the strains LBCM92 and its segregant 92hoΔ-16f displayed 529
poor ethanol production due to deficiency in maltose fermentation (Fig. 5). 530
Analysis of the aroma production of the hybrid strains revealed that breeding 531
generated changes in aroma production (even in the segregants originated by tetraploid 532
hybrid strains). Concentration of aroma compounds in the fermented malt extract is shown 533
in Figure 6. Some hybrids, such as B22 and C2-1d, showed 3-methylbutyl acetate 534
concentrations exceeding the levels of the parental strain LBCM78, but similar at segregant 535
78-2b (Fig. 6B). Production of 3-methylbutanol was also increased although to a lesser 536
extent (Fig. 6A). All hybrid strains showed ethyl acetate concentration higher than the 537
parental strain LBCM78, but similar to the lager strain W34/70 (Fig. 6C). Among the three 538
tested hybrid strains, C2-1d produced higher ratio 3-methylbutyl acetate/3-methylbutanol 539
when compared to all other strains including the lager strain W34/70 (Fig. 6D). Given that 540
the strain C2-1d showed good fermentation performance, it might be an important 541
candidate for the production of lager beer with distinct sensorial characteristics. 542
543
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

25
DISCUSSION 544
The demand for yeast strains with optimal and heterogeneous technological profiles 545
for use in beer production has increased. In this study, we demonstrated that it is possible to 546
breed yeast strains isolated from cachaça distilleries to develop novel lager brewery strains 547
with desired traits. 548
First, we tried to understand the characteristics of 128 cachaça yeast strains 549
regarding their sporulation ability; spore viability and life cycle. Tetrad analysis revealed 550
levels of sporulation varying from high to very low and the frequency of sporulation 551
(proportion of four spores for asci among total asci) was also variable. We used optimal 552
conditions for sporulation (medium containing acetate and incubation temperature 553
approximately 6 ºC lower than the optimal temperature for growth as suggest before (10); 554
however, our results showed fourteen yeast strains unable to sporulate, even when the 555
incubation time was increased to 30 days. These results are consistent with those obtained 556
for S. cerevisiae strains isolated from wine fermentation (11, 18, 27). Moreover, as low 557
frequency of sporulation is usually associate to aneuploidy and other types of unbalance in 558
the genome, the fact that the majority of cachaça yeast strains are diploids (data not 559
shown), suggest that still unknown mechanisms must be involved in the control of the 560
sporulation. 561
The yeast spore viability assay is generally performed to check the percentage of 562
viable spores obtained among spores resulting from the sporulation process after 2-5 days 563
of dissection and incubation at temperature of 30 ºC (11, 18). In our study, spore viability 564
was also quite variable among the isolates; while only 37 strains presented spore viability 565
above 80%, seven strains presented non-viable spores. Another interesting characteristic 566
was that some strains presented spore colonies visible only after long time of incubation. In 567
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

26
general, there was no relationship among the parameters frequency and efficiency of 568
sporulation as well as the viability of the spores, because several strains presented a low 569
percentage of sporulation, but had high spore efficiency (i.e. LBCM50) or high percentage 570
of sporulation and low or no viability of spores (i.e. LBCM94). 571
Regarding the sexual life cycle, and considering the sporulating strains, the majority 572
of the cachaça yeast strains analyzed seem to be homothallic (98%) and only three strains 573
heterothallic. Interestingly, different data have shown that several heterothallic S. cerevisiae 574
strains have been isolated from wine and natural sites (11, 19). However, the majority of 575
natural isolates of S. cerevisiae are considered able to undergo mating-type switching and 576
therefore be homothallic (6, 28). If this is indeed the case, then the HO endonuclease should 577
be predominantly functional in S. cerevisiae strain in nature, and homothallism represents 578
the most common life cycle similar to results shown here. Among the homothallic yeast 579
strains of our study, 12 strains presented at least one inactive copy of HO gene (date not 580
shown). The mutations found in the HO sequences of S. cerevisiae strains are strong 581
indicators of full or partial loss of endonuclease Ho function and yeast cells are unable to 582
switch mating type and to occur crossing between neighboring “sister” cells (self-583
diploidization) (19). 584
On the other hand, due to its complex sexual life cycle, homothallic yeast can 585
eliminate cells with deleterious recessive mutations from the population, while generating 586
homozygous diploid cells in which these mutations are not present (28, 39, 45). This would 587
be important in the cachaça fermenting conditions due to existence of different stressing 588
conditions, that together with lack of nutrients lead to sporulation of yeast cells for survival 589
(32); nevertheless, and because the presence of an active HO gene, some of the descendants 590
of these haploid spores will do self-diploidization. Therefore, homothallism can be 591
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

27
important to select cachaça yeast strains resistant to different types of stress (28, 32). 592
Nevertheless, and from a breeding perspective, the use of spores that display homothallic 593
behavior is not appropriate forcing the deletion of the HO gene and at same time generating 594
a genetic modified yeast cells (35, 36). 595
Based on these results, we identified yeast strains suitable for use in sexual 596
hybridization techniques, with good sporulation rate and viable spores. The development of 597
hybrids has proven to be a powerful approach to generate novel yeast variants with 598
enhanced characteristics for lager beer (21, 26, 36). This is very important for the brewing 599
industry because, as recently demonstrated (13), the available commercial strains have been 600
cultivated over the years suffering an intense domestication process that lead to a strong 601
industry-specific selection for many important traits (stress tolerance; sugar utilization; 602
flavor production; etc). Nevertheless, it was also associated with loss of the sexual cycle 603
and decay of survival in nature. Consequently, the genome of brewing yeast strains has 604
become aneuploidy or polyploidy, with reduction of the capacity of sporulation and with 605
the generation of spores with low viability. This situation is still more serious for lager 606
strains because they are originated from a natural hybridization between S. cerevisiae and S. 607
eubayanus (29) and the resulting hybrid S. pastorianus is a tetraploid strain that cannot be 608
used for breeding practices. Therefore, we believe that our strategy can contribute to 609
overcome all these natural difficulties creating possibilities of generation of new interesting 610
lager yeast strains for brewery industry. 611
Thus, we used two cachaça S. cerevisiae strains LBCM92 and LBCM78 that 612
present a higher flocculation rate and higher levels of flavoring compounds, respectively. In 613
order to get new yeast strains with desirable properties useful for lager beer production, we 614
compare two hybridization methods: haploid-to-haploid and diploid-to-diploid mating 615
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

28
procedures. In spite of the fact that the haploid-to-haploid mating increases the possibility 616
of yielding a superior hybrid (21, 26, 38), in our case the selected superior segregant 617
haploid from LBCM92 MATa/α HO::KanMX/HO::hph (LBCM92hoΔ) should be 618
considered a genetically modified organism, and the use of GM yeasts in food fermentation 619
processes is still controversial (39). 620
Putative hybrids from haploid-to-haploid mating events were confirmed using PCR 621
mating type analysis. However, an assessment of parental phenotype traits showed that only 622
one hybrid called B22 inherited high flocculation capacity, although with a rate below the 623
value of haploid parent strain. Several authors consider that the biggest advantage of the 624
haploid-to-haploid mating is the phenotyping of the haploid segregants before to the mating 625
(in our case the capacity to ferment maltose and higher production of flavoring compound – 626
78-2b, and high capacity to flocculate – 92hoΔ-16f); since parental traits can be transferred 627
to diploid hybrid increasing the chances to obtain strains with the desired characteristics 628
(38). However, the results shown that this technique was not a promising approach because 629
we did not have any hybrids with a flocculation rate similar to or higher than that of the 630
parent strain. This also suggests that high flocculation ability may be a polygenic and 631
multifactorial trait (1), and that most probably the ancestral 78-2b (originated from the 632
parental strain LBCM78) must carry out negative regulators of the flocculation. 633
In addition, none of the yeast strains originated from diploid-to-diploid mating 634
showed flocculation ability. In this case, as flow cytometry analysis further revealed, DNA 635
content of these hybrids was equivalent to tetraploid genome (Fig. 4), we initially wondered 636
that this loss of phenotype should be regulated by ploidy (12). Interestingly, our results 637
have shown that the flocculation phenotype was recovered in several segregants obtained 638
from dissection of tetraploid hybrids (Table S1), with three segregants (C2-1d, C3-7b and 639
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

29
C4-1g) displaying stronger flocculation ability. However, the situation seems to be more 640
complex since the diploids resulting from the crossing between the haploids 78-2b and 641
92hoΔ-16f presented a variable flocculation rate reinforcing the idea that flocculation 642
would be a polygenic and multifactorial trait (1), and that in the genetic background of the 643
parental wild strain LBCM78 there would be strong negative regulators of the flocculation. 644
Interestingly, the segregation of such diploids generated from the approach haploid-to-645
haploid breeding also lead to a recovery of a high rate of flocculation (compared to the 646
parental wild strain LBCM92 – data not shown), but it was observed a high level of loss of 647
spore viability, a trait inherited from both parental strains that present spore viability 648
ranging between 51 and 60% (strain LBCM78) and 21 and 30% (strain LBCM92). Beyond 649
that, the presence of the genetic markers originated from the deletion of HO gene in the 650
parental wild strain LBCM92 present an obstacle to the use of such strains in the brewery 651
industry. 652
In contrast, triploid and tetraploid hybrids obtained by crossing between S. 653
cerevisiae and S. eubayanus showed a much higher flocculation ability when compared to 654
the correspondent segregant diploid hybrids (20). Some phenotypic traits were also affected 655
by the ploidy level in other interspecific Saccharomyces hybrids, where triploid and 656
tetraploid hybrids were able to produce more ethanol and 3-methylbutyl acetate than their 657
parental strains and diploid hybrids (26). Therefore, it seems that the influence of ploidy is 658
rather circumstantial and that most probably the presence of dominant negative regulators 659
of flocculation is the main reason for the decrease of the flocculation rate in the different 660
hybrids obtained in this work. 661
Additionally, lager beers are traditionally fermented at lower temperatures using 662
bottom-fermenting yeast strains. The cold adaptation of lager yeast seems have been 663
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

30
inherited from S. eubayanus part of the genome (3, 14, 44, 46). However, we found 664
intraspecific hybrid strains (S. cerevisiae) able to ferment malt extract at 12 °C and which 665
produced more ethanol than their corresponding parental strains. Indeed, the hybrid C2-1d 666
also showed interesting fermentation performance in laboratory-scale lager beer 667
fermentation comparable to lager strain S. pastorianus W34/70 (Figure 5). In fact this is not 668
a surprising finding since it has been demonstrated that ale brewing S. cerevisiae strains can 669
tolerate low temperatures (13). 670
However, not all of the hybrids developed were able to outperform their respective 671
parental strains in laboratory-scale lager beer fermentation. This was particularly true in the 672
case of the haploid-to-haploid mating technique, yielding the so-called crippled strains, 673
which show improvement for some selected traits, but a worse performance for other 674
industrially important phenotypes (in our case, reduced flocculation rate) (38). 675
Three of the most important aroma compounds in beer are 3-methylbutyl acetate (a 676
banana-like aroma), ethyl acetate (an alcoholic, fruity, but also solvent-like aroma) (38) and 677
3-methylbutanol (fruity, sweet) (35). In general, the production of 3-methylbutyl acetate 678
and ethyl acetate was found to be significantly higher in the hybrids than parental strains 679
(Figure 6). In addition, production of 3-methylbutyl acetate was accompanied by increase 680
in ethyl acetate, since these compounds share part of their metabolic pathway (31). Despite 681
the fact that ethyl acetate often contributes positively to the aroma, it can sometimes be 682
perceived negatively because it can emanate a solvent-like aroma when it is present in high 683
concentrations (34). However, in our results, ethyl acetate production of the hybrids did not 684
reach high levels; the maximal concentration detected was 18 mg L-1
in the strains C4-1g, 685
while the odor threshold in lager beer should be approximately 30 mg L-1
(34, 41). The 3-686
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

31
methylbutyl acetate concentration obtained with hybrid strain C2-1d was markedly high, 687
reached concentrations around its report threshold level in beer (1.2 mg L-1
) (34). 688
In conclusion, our study showed the improvement of yeast strains S. cerevisiae 689
isolated from cachaça distilleries for production of lager style beer by using haploid-to-690
haploid and diploid-to-diploid mating experiments. The best hybrid generated showed good 691
fermentation performance at low temperature, high flocculation capacity and desirable 692
production of acetate esters that was significantly better than one commercial lager strain. 693
Nevertheless, and considering that other non-typical characteristics were found in the new 694
hybrid strain, i.e. incapacity to ferment maltotriose and production of phenolic off-flavors 695
compounds, additional work still using classical breeding and/or mutagenesis procedures 696
must be done to improve the characteristics of such new hybrid. In any case, our work 697
demonstrates that alternative techniques can be used to found new and interesting strains to 698
be used in beer production. 699
700
Acknowledgments 701
We thank Rodrigo Dian O. A. Soares for technical assistance in the cytometer 702
analysis. This work was supported by grants from Fundação de Capacitação de Pessoal de 703
Nível Superior from the Ministry of Education – CAPES/Brazil (PCF-PVE 021/2012; 704
Edital 76/2014) and from Universidade Federal de Ouro Preto, Fundação de Amparo à 705
Pesquisa do Estado de Minas Gerais - FAPEMIG (Process APQ-00263-10) and a research 706
fellowship from Conselho Nacional de Desenvolvimento Científico e Tecnológico – CNPq 707
(Brazil) Process 304815/2012. 708
709
710
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

32
Conflicts of interest 711
Authors wish to confirm that there are no known conflicts of interest associated with 712
this publication and there has not been significant financial support for this work that could 713
have influenced its outcome. 714
715
References 716
717
1. Alvarez, F., L. F. da Mata Correa, T. Macedo Araújo, B. E. Fernandes Mota, 718
L. E. F. Ribeiro da Conceição, I. de Miranda Castro, and R. Lopes Brandão. 719
2014. Variable flocculation profiles of yeast strains isolated from cachaça 720
distilleries. Int J Food Microbiol 190:97-104. 721
2. Barbosa, E. A., M. T. Souza, R. H. S. Diniz, F. Godoy-Santos, F. Faria-Oliveira, 722
L. F. M. Correa, F. Alvarez, M. X. Coutrim, R. J. d. C. F. Afonso, and I. M. 723
Castro. 2016. Quality improvement and geographical indication of cachaça 724
(Brazilian spirit) by using locally selected yeast strains. J App Microbiol 121:1038-725
1051. 726
3. Bing, J., P.-J. Han, W.-Q. Liu, Q.-M. Wang, and F.-Y. Bai. 2014. Evidence for a 727
Far East Asian origin of lager beer yeast. Curr Biology 24:R380-R381. 728
4. Bizaj, E., A. G. Cordente, J. R. Bellon, P. Raspor, C. D. Curtin, and I. S. 729
Pretorius. 2012. A breeding strategy to harness flavor diversity of Saccharomyces 730
interspecific hybrids and minimize hydrogen sulfide production. FEMS Yeast Res 731
12:456-465. 732
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

33
5. Bokulich, N. A., and C. W. Bamforth. 2013. The microbiology of malting and 733
brewing. Microbiol Mol Biol R 77:157-172. 734
6. Butler, G., C. Kenny, A. Fagan, C. Kurischko, C. Gaillardin, and K. H. Wolfe. 735
2004. Evolution of the MAT locus and its Ho endonuclease in yeast species. Proc 736
Natl Acad Sci U S A 101:1632-7. 737
7. Connolly, B., C. I. White, and J. E. Haber. 1988. Physical monitoring of mating 738
type switching in Saccharomyces cerevisiae. Mol Cell Biol 8:2342-2349. 739
8. da Conceição, L., M. Saraiva, R. Diniz, J. Oliveira, G. Barbosa, F. Alvarez, L. 740
da Mata Correa, H. Mezadri, M. Coutrim, R. d. Afonso, C. Lucas, I. Castro, 741
and R. Brandão. 2015. Biotechnological potential of yeast isolates from cachaça: 742
the Brazilian spirit. J Ind Microbiology Biot 42:237-246. 743
9. de Souza, A. P., M. e. A. Vicente, R. C. Klein, L. G. Fietto, M. X. Coutrim, R. J. 744
de Cássia Franco Afonso, L. D. Araújo, P. H. da Silva, L. E. Bouillet, I. M. 745
Castro, and R. L. Brandão. 2012. Strategies to select yeast starters cultures for 746
production of flavor compounds in cachaça fermentations. Antonie Van 747
Leeuwenhoek 101:379-92. 748
10. Elrod, S. L., S. M. Chen, K. Schwartz, and E. O. Shuster. 2009. Optimizing 749
Sporulation Conditions for Different Saccharomyces cerevisiae Strain Backgrounds, 750
p. 21-26. In S. Keeney (ed.), Mol Gen Method, vol. 1. Humana Press, Totowa, NJ. 751
11. Fernández-González, M., J. Úbeda, and A. Briones. 2015. Study of 752
Saccharomyces cerevisiae Wine Strains for Breeding Through Fermentation 753
Efficiency and Tetrad Analysis. Curr Microbiol 70:441-449. 754
12. Galitski, T., A. J. Saldanha, C. A. Styles, E. S. Lander, and G. R. Fink. 1999. 755
Ploidy regulation of gene expression. Science 285:251-254. 756
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

34
13. Gallone, B., J. Steensels, T. Prahl, L. Soriaga, V. Saels, B. Herrera-Malaver, A. 757
Merlevede, M. Roncoroni, K. Voordeckers, and L. Miraglia. 2016. 758
Domestication and divergence of Saccharomyces cerevisiae beer yeasts. Cell 759
166:1397-1410. e16. 760
14. Gibson, B. R., E. Storgårds, K. Krogerus, and V. Vidgren. 2013. Comparative 761
physiology and fermentation performance of Saaz and Frohberg lager yeast strains 762
and the parental species Saccharomyces eubayanus. Yeast 30:255-266. 763
15. Gietz, R. D., R. H. Schiestl, A. R. Willems, and R. A. Woods. 1995. Studies on 764
the transformation of intact yeast cells by the LiAc/SS-DNA/PEG procedure. Yeast 765
11:355-60. 766
16. Giudici, P., L. Solieri, A. M. Pulvirenti, and S. Cassanelli. 2005. Strategies and 767
perspectives for genetic improvement of wine yeasts. Appl Microbiol Biotechnol 768
66:622-8. 769
17. Huxley, C., E. D. Green, and I. Dunbam. 1999. Rapid assessment of 770
Saccharomyces cerevisiae mating type by PCR. Trends Genet 6:236. 771
18. Johnston, J. R., C. Baccari, and R. K. Mortimer. 2000. Genotypic 772
characterization of strains of commercial wine yeasts by tetrad analysis. Res 773
Microbiol 151:583-590. 774
19. Katz Ezov, T., S.-L. Chang, Z. E. Frenkel, A. V. Segrè, M. Bahalul, A. W. 775
Murray, J.-Y. Leu, A. Korol, and Y. Kashi. 2010. Heterothallism in 776
Saccharomyces cerevisiae isolates from nature: effect of HO locus on the mode of 777
reproduction. Mol Ecol 19:121-131. 778
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

35
20. Krogerus, K., M. Arvas, M. De Chiara, F. Magalhães, L. Mattinen, M. Oja, V. 779
Vidgren, J.-X. Yue, G. Liti, and B. Gibson. 2016. Ploidy influences the functional 780
attributes of de novo lager yeast hybrids. App Microbiol Biot 100:7203-7222. 781
21. Krogerus, K., F. Magalhães, V. Vidgren, and B. Gibson. 2015. New lager yeast 782
strains generated by interspecific hybridization. J Ind Microbiol Biot 42:769-778. 783
22. Landaud, S., E. Latrille, and G. Corrieu. 2001. Top Pressure and Temperature 784
Control the Fusel Alcohol/Ester Ratio through Yeast Growth in Beer Fermentation. 785
J I Brewing 107:107-117. 786
23. Libkind, D., C. Hittinger, E. Valerio, C. Goncalves, and J. Dover. 2011. 787
Microbe domestication and the identification of the wild genetic stock of lager-788
brewing yeast. Proc. Natl. Acad. Sci. USA 108:14539. 789
24. Liti, G., D. M. Carter, A. M. Moses, J. Warringer, L. Parts, S. A. James, R. P. 790
Davey, I. N. Roberts, A. Burt, V. Koufopanou, I. J. Tsai, C. M. Bergman, D. 791
Bensasson, M. J. T. O'Kelly, A. van Oudenaarden, D. B. H. Barton, E. Bailes, 792
A. N. Nguyen Ba, M. Jones, M. A. Quail, I. Goodhead, S. Sims, F. Smith, A. 793
Blomberg, R. Durbin, and E. J. Louis. 2009. Population genomics of domestic 794
and wild yeasts. Nature 458:337-341. 795
25. López, V., A. Querol, D. Ramón, and M. T. Fernández-Espinar. 2001. A 796
simplified procedure to analyse mitochondrial DNA from industrial yeasts. Int J 797
Food Microbiol 68:75-81. 798
26. Mertens, S., J. Steensels, V. Saels, G. De Rouck, G. Aerts, and K. J. Verstrepen. 799
2015. A Large Set of Newly Created Interspecific Saccharomyces Hybrids Increases 800
Aromatic Diversity in Lager Beers. App Environ Microbiol 81:8202-8214. 801
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

36
27. Mortimer, R., and M. Polsinelli. 1999. On the origins of wine yeast. Res. 802
Microbiol. 150:199. 803
28. Mortimer, R. K. 2000. Evolution and variation of the yeast (Saccharomyces) 804
genome. Genome Res 10:403-409. 805
29. Nguyen, H. V., J. L. Legras, C. Neuveglise, and C. Gaillardin. 2011. 806
Deciphering the hybridisation history leading to the Lager lineage based on the 807
mosaic genomes of Saccharomyces bayanus strains NBRC1948 and CBS380. PLoS 808
One 6:e25821:1-19. 809
30. Oliveira, V. A., M. A. Vicente, L. G. Fietto, I. M. Castro, M. X. Coutrim, D. 810
Schüller, H. Alves, M. Casal, J. O. Santos, L. D. Araújo, P. H. da Silva, and R. 811
L. Brandão. 2008. Biochemical and molecular characterization of Saccharomyces 812
cerevisiae strains obtained from sugar-cane juice fermentations and their impact in 813
cachaça production. Appl Environ Microbiol 74:693-701. 814
31. Pires, E. J., J. A. Teixeira, T. Brányik, and A. A. Vicente. 2014. Yeast: the soul 815
of beer’s aroma—a review of flavour-active esters and higher alcohols produced by 816
the brewing yeast. App Microbiol Biot 98:1937-1949. 817
32. Plech, M., J. A. G. M. de Visser, and R. Korona. 2013. Heterosis Is Prevalent 818
Among Domesticated but not Wild Strains of Saccharomyces cerevisiae. G3: 819
Genes|Genomes|Genetics 4:315-323. 820
33. Pretorius, I. 2000. Tailoring wine yeast for the new millennium: novel approaches 821
to the ancient art of winemaking. Yeast 16:675. 822
34. Saerens, S. M. G., F. R. Delvaux, K. J. Verstrepen, and J. M. Thevelein. 2010. 823
Production and biological function of volatile esters in Saccharomyces cerevisiae. 824
Microbial Biot 3:165-177. 825
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

37
35. Saerens, S. M. G., P. J. Verbelen, N. Vanbeneden, J. M. Thevelein, and F. R. 826
Delvaux. 2008. Monitoring the influence of high-gravity brewing and fermentation 827
temperature on flavour formation by analysis of gene expression levels in brewing 828
yeast. Appl Microbiol Biot 80:1039-1051. 829
36. Sanchez, R. G., N. Solodovnikova, and J. Wendland. 2012. Breeding of lager 830
yeast with Saccharomyces cerevisiae improves stress resistance and fermentation 831
performance. Yeast 29:343-355. 832
37. Schilter, B., and A. Constable. 2002. Regulatory control of genetically modified 833
(GM) foods: likely developments. Toxicol Lett 127:341-349. 834
38. Steensels, J., E. Meersman, T. Snoek, V. Saels, and K. J. Verstrepen. 2014. 835
Large-scale selection and breeding to generate industrial yeasts with superior aroma 836
production. App Environ Microbiol 80:6965-6975. 837
39. Steensels, J., T. Snoek, E. Meersman, M. P. Nicolino, K. Voordeckers, and K. J. 838
Verstrepen. 2014. Improving industrial yeast strains: exploiting natural and 839
artificial diversity. Fems Microbiol Rev 38:947-995. 840
40. Tanruean, K., and N. Rakariyatham. 2016. Efficient Synthesis of 4-Vinyl 841
Guaiacol via Bioconversion of Ferulic Acid by Volvariella volvacea. Chiang Mai J. 842
Sci 43:158 - 168. 843
41. Verstrepen, K. J., G. Derdelinckx, J.-P. Dufour, J. Winderickx, J. M. 844
Thevelein, I. S. Pretorius, and F. R. Delvaux. 2003. Flavor-active esters: Adding 845
fruitiness to beer. J Biosci Bioeng 96:110-118. 846
42. Vezinhet, F., B. Blondin, and J.-N. Hallet. 1990. Chromosomal DNA patterns and 847
mitochondrial DNA polymorphism as tools for identification of enological strains of 848
Saccharomyces cerevisiae. App Microbiol Biot 32:568-571. 849
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

38
43. Vicente, M. A., L. G. Fietto, I. M. Castro, A. N. dos Santos, M. X. Coutrim, and 850
R. L. Brandão. 2006. Isolation of Saccharomyces cerevisiae strains producing 851
higher levels of flavoring compounds for production of "cachaça" the Brazilian 852
sugarcane spirit. Int J Food Microbiol 108:51-9. 853
44. Walther, A., A. Hesselbart, and J. Wendland. 2014. Genome sequence of 854
Saccharomyces carlsbergensis, the world's first pure culture lager yeast. G3: Genes| 855
Genomes| Genetics 4:783-793. 856
45. Wang, C., and Y. Liu. 2013. Dynamic study of yeast species and Saccharomyces 857
cerevisiae strains during the spontaneous fermentations of Muscat blanc in 858
Jingyang, China. Food Microbiol 33:172. 859
46. Wendland, J. 2014. Lager yeast comes of age. Eukaryot Cell 13:1256-1265. 860
47. Zastrow, C. R., C. Hollatz, P. S. de Araujo, and B. U. Stambuk. 2001. 861
Maltotriose fermentation by Saccharomyces cerevisiae. J Ind Microbiol Biotechnol 862
27:34-8. 863
864
865
866
867
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

39
Figures legends 868
869
FIG 1 Sporulation analysis of yeast strains isolated from cachaça distilleries. A, 870
sporulation frequency; B, Sporulation efficiency; C, Spore viability. Numbers 871
corresponding to each yeast strain of the collection LBCM is placed inside bars. 872
873
FIG 2 Determination of yeast strain mating type by PCR. Colony-PCR using three primers 874
to amplify either a MATa (544bp) or MATα (404bp) specific band. M, Promega™ 100bp 875
DNA Ladder. (A) S. cerevisiae LBCM73, LBCM78, LBCM115, LBCM92 and their 876
segregants denoted with a number and one small letter. (B) Yeast strains B16, B22, C28, 877
C33 are hybrids originated from the crossing between segregants 78-2b and 92hoΔ-16f, and 878
control strains S. cerevisiae BY4742 MATα and BY4743 MATa/α. (C) Yeast strains 92 IX 879
(7) diploid MATa/a, 78 a34 (15) diploid MATα/α; Strains C1, C2, C3, C4, B15, C13, C14 880
and C16 are tetraploid hybrids; Strains C2-1d, C4-1g and C3-7b are diploid segregants. 881
882
FIG 3 Concentration of aroma compounds produced by parental strain S. cerevisiae 883
LBCM78 and its meiotic segregants at the end of fermentation in YP 250 broth with 10% 884
(wt/vol) glucose. Results are the mean and standard deviation of two independent 885
experiments. Means with the same letter are not significantly different from each other 886
(Tukey’s test, P < 0.05). 887
888
FIG 4 DNA content analysis of yeast strains by fluorescence flow cytometry. Cells were 889
processed through the staining protocol with SYTOX® Green (see Material and Methods). 890
SYTOX® Green fluorescence was analyzed using Flowjo® software. Fluorescence 891
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

40
histograms present cell peaks at the G0/G1 and G2/M border. A, B, C control strains; F, K 892
haploid segregants; N, strain originated from haploid-to-haploid mating; G, L, parental 893
strains; H, I, M tetraploid strains originated from diploid-to-diploid mating; E, J, O 894
segregants originated from dissection of tetraploid strains. 895
896
FIG 5 Fermentation kinetics (depicted as cumulative weight loss) of parental strains S. 897
cerevisiae LBCM78, LBCM92, 78-2b and 92hoΔ-16f, hybrid strains B22, C4-1g, C2-1d 898
and S. pastorianus var. Weihenstephan 34/70. Results are the mean and standard deviation 899
of two independent experiments. 900
901
FIG 6 The concentrations of aroma compounds in the malt extract fermented by parental 902
strains S. cerevisiae LBCM78, LBCM92, 78-2b and 92hoΔ-16f, hybrid strains B22, C4-1g, 903
C2-1d and S. pastorianus var. Weihenstephan 34/70. Results are the mean and standard 904
deviation of two independent experiments. Means with the same letters are not significantly 905
different from each other (Tukey’s test, P < 0.05). 906
907
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

41
Table 1 PCR primers employed in this study
Primer Sequence (5’- 3’)
MATLocus AGTCACATCAAGATCGTTTATGG
MATa ACTCCACTTCAAGTAAGAGTTTG
MATα GCACGGAATATGGGACTACTTCG
pdelfw TAGCAGATGCGCGCACCTGCGTTGTTACCACAACTCTTATGAGGCCC
GCGGAGTGGTCGGCTGGAGATCGG
pdelrv ATGCTTTCTGAAAACACGACTATTCTGATGGCTAACGGTGAAATTAA
AGACATCGAGCCGTTATGGCGGGCATC
Primer set: MATLocus/MATa/MATα (the DNA sequence present at the MATLocus); pdelfw/pdelrv
(deletion cassettes amplification).
908
909
910
911
912
913
914
915
916
917
918
919
920
921
922
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from

42
Table 2 Flocculation ability of different yeast strains 923
Strain Description Flocculation percentage (SD)*
LBCM92 Parental diploid strain MATa/α 92.9 (1.0) j,i
LBCM78 Parental diploid strain MATa/α 43.7 (3.9) b
78-2B Segregant haploid from LBCM78 MATα 55.4 (1.0) c,d
92-11c Segregant haploid from LBCM92 MATa 90.4 (0.7) j
92-17d Segregant haploid from LBCM92 MATa 91.4 (1.4) j,i
92-16f Segregant haploid from LBCM92 MATa 99.0 (0.4) i
B16 Hybrid diploid MATa/α a 63.8 (0.2) e,f,g
B22 Hybrid diploid MATa/α a 70.1 (0.4) g,h,i
C28 Hybrid diploid MATa/α a 55.5 (0.3) c,d,e
C33 Hybrid diploid MATa/α a 44.6 (1.9) b
92 IX (7) Parental diploid strain MATa/a 93.8 (1.1) j,i
78 a34 (15) Parental diploid strain MATα/α 41.9 (2.4) b
C2 Hybrid tetraploid MATa/α b 28.3 (2.1) a
C3 Hybrid tetraploid MATa/α b 28.9 (2.1) a
C4 Hybrid tetraploid MATa/α b 45.2 (3.7) b
C1-2a Segregant diploid from C1 MATα b 65.3 (0.7) f,g,h
C1-8c Segregant diploid from C1 MATa/α b 68.7 (6.7) f,g,h,i
C2-1d Segregant diploid from C2 MATα b 91.4 (0.4) j,i
C2-8h Segregant diploid from C2 MATa/α b 73.3 (0.0) h,i
C3-7b Segregant diploid from C3 MATa b 90.8 (5.0) j,i
C4-1g Segregant diploid from C4 MATα b 95.4 (2.9) j,i
B15-1d Segregant diploid from B15 MATa/α b 75.2 (0.3) i
B15-7c Segregant diploid from B15 MATa/α b 56.5 (0.5) c,d,e
C13-4h Segregant diploid from C13 MATa/α b 61.5 (0.3) d,e,f
C13-4f Segregant diploid from C13 MATα b 67.6 (0.1) f,g,h,i
C14-2c Segregant diploid from C14 MATa/α b 62.6 (0.2) d,e,f,g
C16-6a Segregant diploid from C16 MATα b 50.3 (0.4) b,c
*Values are the mean and standard deviation of two independent experiments. Means followed by different 924 letters are significantly different by the Tukey’s test, P < 0.05; aHybrid originated from haploid-to-haploid 925 mating; bHybrid originated from diploid-to-diploid mating. 926
on February 1, 2019 by guest
http://aem.asm
.org/D
ownloaded from









![LOUDSPEAKER Web: Telephone: 01582 405694 …pmc-speakers.com/sites/default/files/attachments/twenty21 review... · Telephone: 01582 405694 Web: . K. ... Report, p63]. The John Howes](https://img.pdfslide.us/doc/110x75/5acc4bd37f8b9a27628c5869/loudspeaker-web-telephone-01582-405694-pmc-reviewtelephone-01582-405694.jpg)

![WELCOME [d2jzxcrnybzkkt.cloudfront.net] · Rum/Cachaça Bacardi Carta Blanca 15/180 Maracatu Cachaça 17/210 Havana 3YO 15/180 Mount Gay Eclipse 15/180 Ron Zacapa 23 19/230 Prices](https://img.pdfslide.us/doc/110x75/5c0e385c09d3f258548cb0b4/welcome-rumcachaca-bacardi-carta-blanca-15180-maracatu-cachaca-17210.jpg)








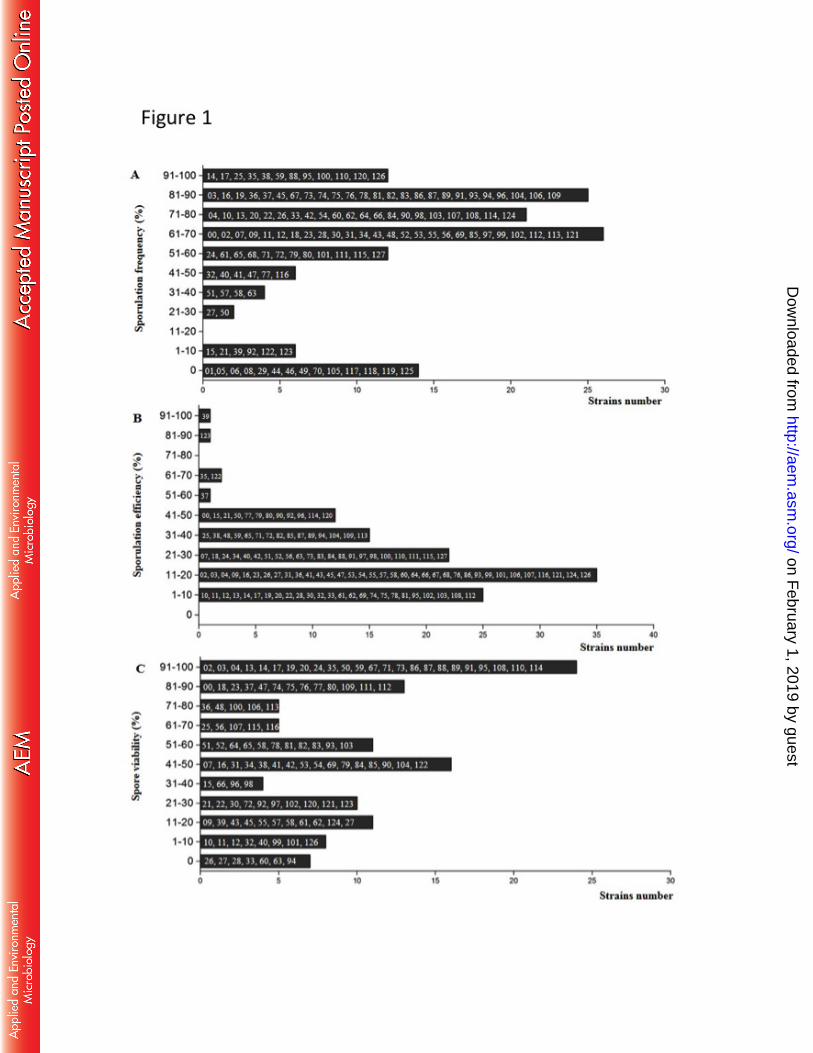
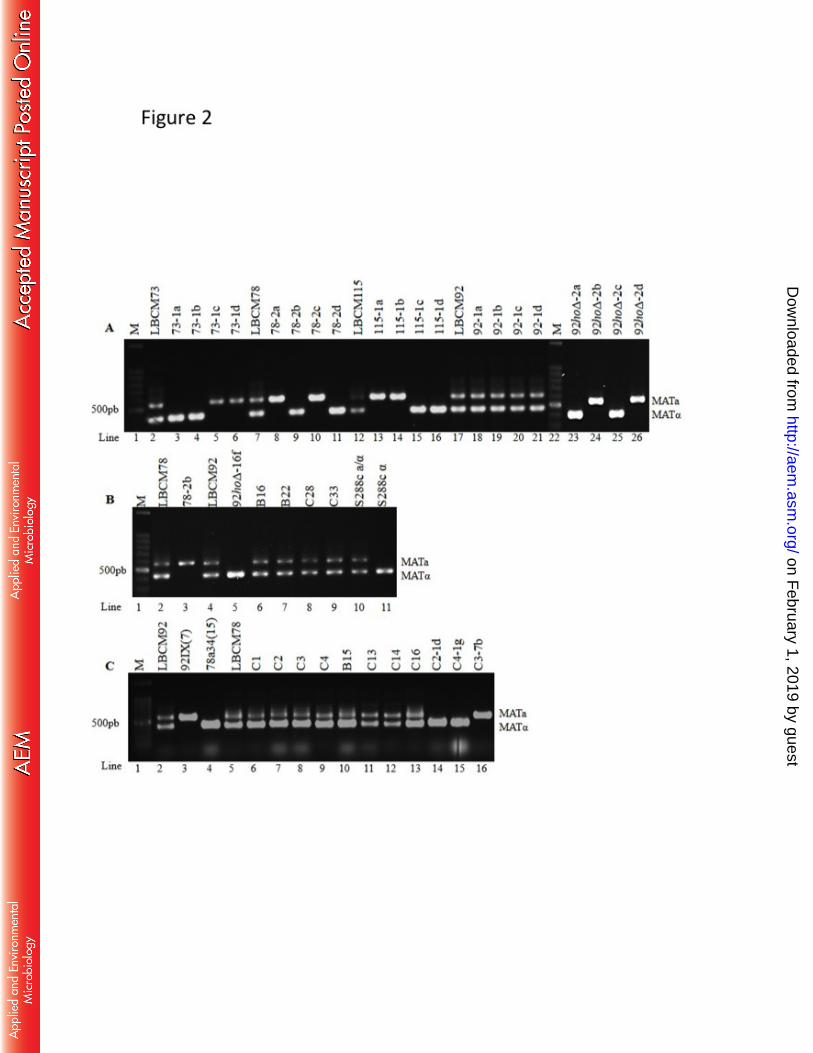
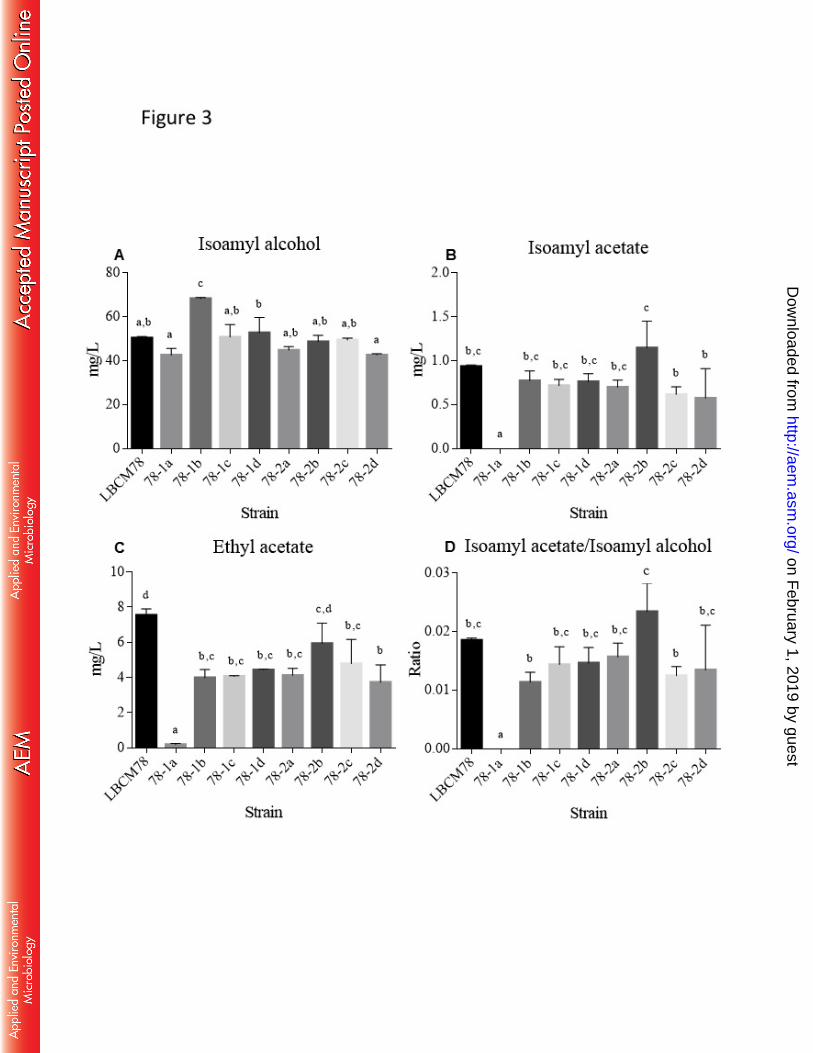
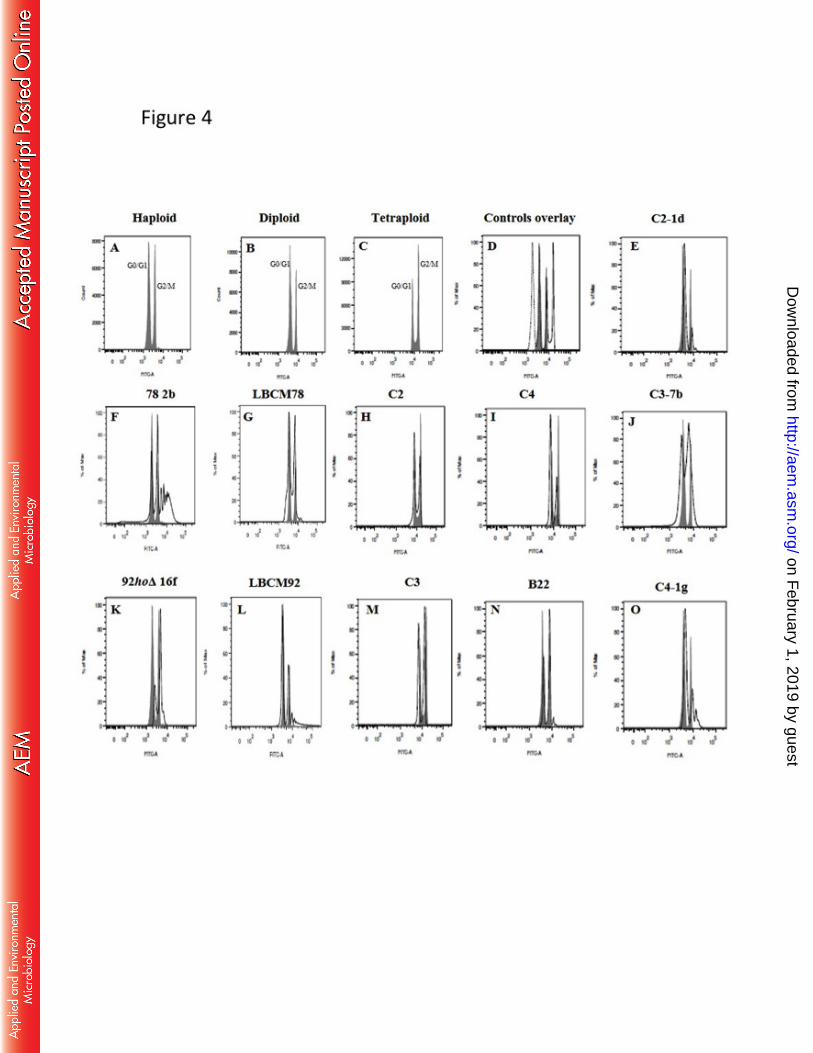
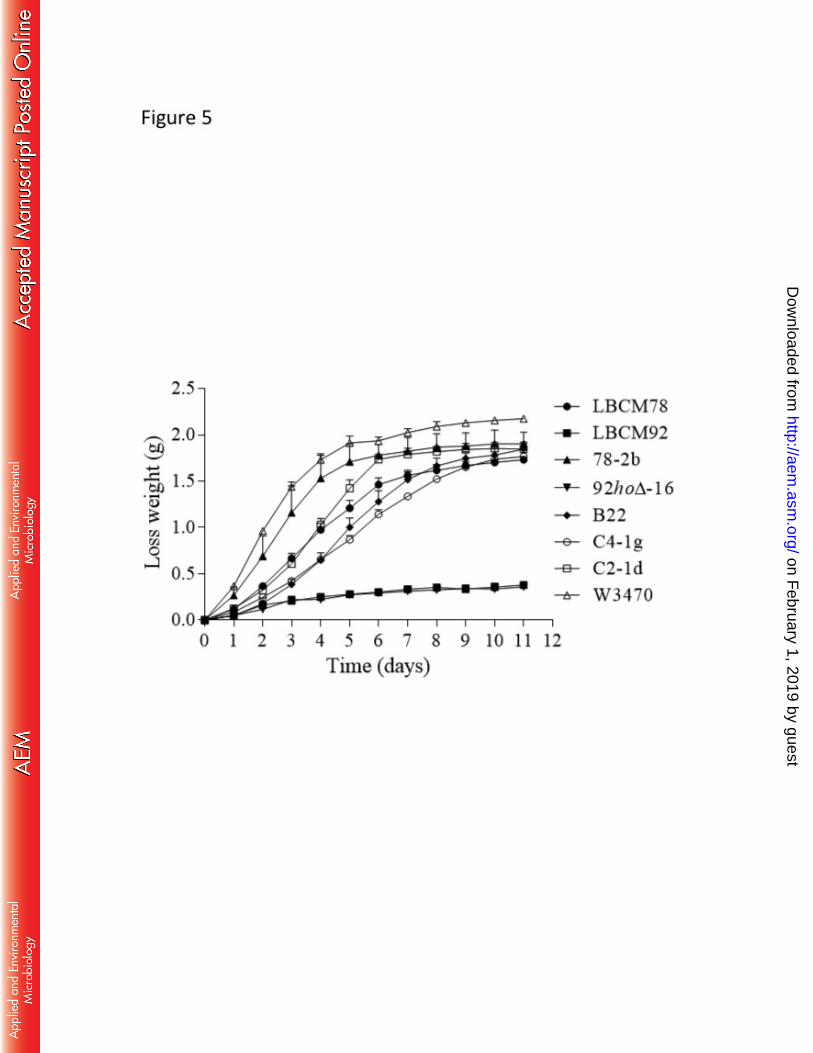
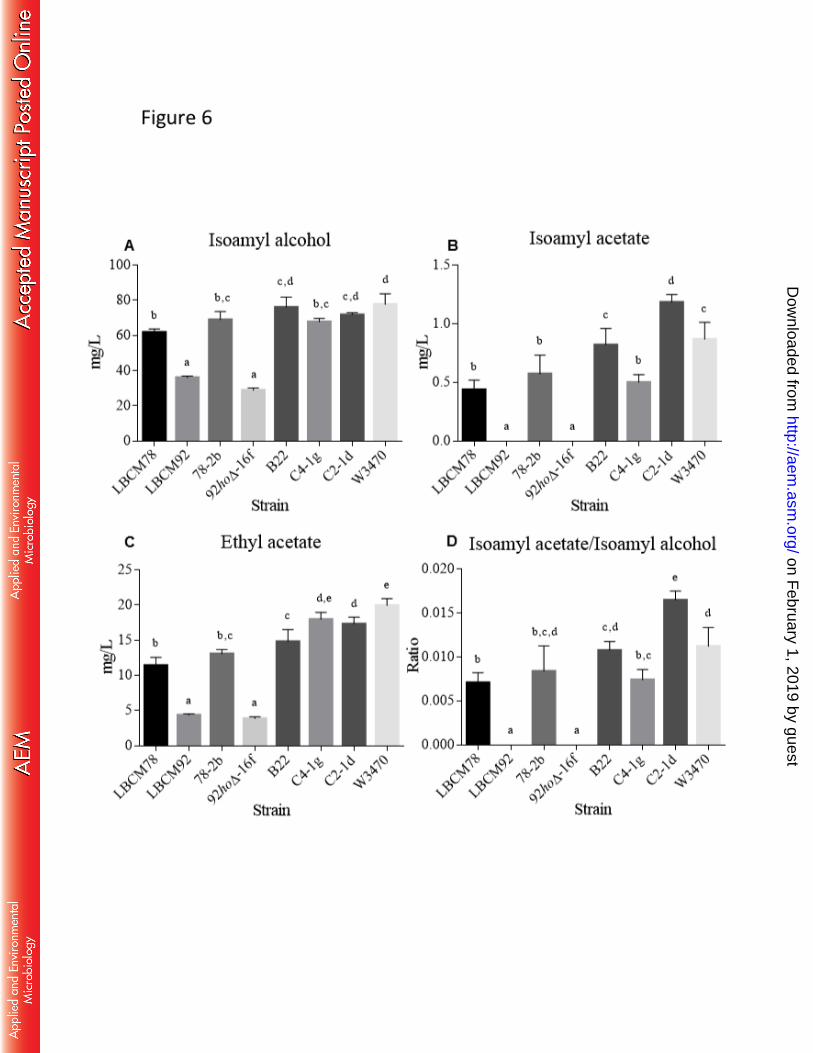

![Flat Fee Internet Recruitment 01582 691414 [email protected]](https://img.pdfslide.us/doc/110x75/620392e8da24ad121e4afd89/flat-fee-internet-recruitment-01582-691414-emailprotected.jpg)







