-
7/24/2019 Neutrophil Function From Mechanism to Disease
1/33
Neutrophil Function:From Mechanisms to Diseas
Borko Amulic, Christel Cazalet,Garret L. Hayes, Kathleen D.
Metzler,and Arturo Zychlinsky
Department of Cellular Microbiology, Max Planck Institute for
Infection Biology,Charit eplatz 1, 10117 Berlin, Germany; email:
[email protected],[email protected],
[email protected],
[email protected]@mpiib-berlin.mpg.de
Annu. Rev. Immunol. 2012. 30:45989
First published online as a Review in Advance onJanuary 3,
2012
TheAnnual Review of Immunologyis online
atimmunol.annualreviews.org
This articles doi:10.1146/annurev-immunol-020711-074942
Copyright c2012 by Annual Reviews.All rights reserved
0732-0582/12/0423-0459$20.00
All authors contributed equally to the work andare listed
alphabetically.
Keywords
inflammation, antimicrobial, granule, phagocytosis, NET
Abstract
Neutrophils are the most abundant white blood cells in
circula
and patients with congenital neutrophil deficiencies suffer from
seinfections that are often fatal, underscoring the importance of
t
cells in immune defense. In spite of neutrophils relevance in
immuresearch on these cells has been hampered by their
experimentall
tractable nature. Here, we present a survey of basic neutrophil
bio
with an emphasis on examples that highlight the function of
neutronot only as professional killers, but also as instructors of
the immsystem in the context of infection and inflammatory disease.
We f
on emerging issues in the field of neutrophil biology, address
ques
in this area that remain unanswered, and critically examine the
exmental basis for common assumptions found in neutrophil
literatu
459
Click here for quick links to
Annual Reviews content online,
including:
Other articles in this volume
Top cited articles
Top downloaded articles
Our comprehensive search
FurtherANNUAL
REVIEWS
-
7/24/2019 Neutrophil Function From Mechanism to Disease
2/33
INTRODUCTION
In the late nineteenth century, Paul Ehrlich,
dissatisfied with what he considered an in-excusable disinterest
in the white blood cell,
began to utilize newly developed cell-staining
techniques to examine subpopulations of
leuko-cytes.Hisexperimentationledtoanewappreci-
ation for the heterogeneity of white blood cellsand to the
discovery of several novel leukocyte
subpopulations. Ehrlich named one of thesenewly discovered cell
types, characterized by a
polymorphous nucleus and a tendency to re-tain neutral dyes, the
neutrophil (1) (see also
the sidebar, A Natural History of Neutrophils).The function of
neutrophils was initially
shrouded in considerable mystery; their con-
spicuous presence during infections led severalresearchers to
arrive hastily at a rather ironic
conclusion: They surmised that neutrophilspromote infection,
serving as cellular shuttles
for bacteria (2). Their actual function, that ofantimicrobial
actors in the immune response,
was eventually demonstrated conclusively by acontemporary of
Ehrlich, Elie Metchnikoff, an
A NATURAL HISTORY OF NEUTROPHILS
Phagocytes are ancient cells that evolved to allow
multicellularorganisms to thrive in the face of constant
competition with mi-
crobes for resources. Metchnikoffs seminal theory of
cellularimmunity was based on comparative embryology and
observa-
tions of phagocytes in various simpleorganisms, includingthe
mi-croscopic crustacean Daphnia. Remarkably, even the slime
mold
Dictyostelium discoideumhas phagocytic cells that protect it
from
infection (200). The short-lived neutrophil with a lobulated
nu-cleus andgranule-packed cytoplasm is a more
recentevolutionary
adaptation. In insects, phagocytes are long lived and have
roundnuclei. They do, however, produce hydrogen peroxide and
carry
distinct classes of granules (201). Bony fish and frogs have
bonafide neutrophils that are functionally similar to mammalian
ones
(202, 203). In both zebrafish and rodents, neutrophils are
lessabundant than in humans, comprising only 1520% of immune
cells. In chimpanzees, neutrophils account for more than 50%
ofthe differential blood count (204).
early and enthusiastic evolutionary biologist i
terested in the phagocytic capacity of cells.Metchnikoff
demonstrated that injury
starfish embryos resulted in recruitment phagocytic cells to the
site of injury (3). H
theorized (correctly) that these cells migrate
injured sites and participate in microbe dige
tion. Remarkably, this prescient view of netrophil action still
aptly summarizes, more tha century later, the basic role of
neutroph
in immunity. The uniquely lobulated nucleof the neutrophil also
inspired Metchnikoff
rename these cells: He called them polymophonuclear leukocytes
(or PMNs), a title th
still enjoys frequent use and that is used intechangeably with
neutrophil throughout this r
view. Together with two other developmentarelated cell types,
the eosinophils and basoph
(also discovered by Ehrlich), PMNs form t
granulocyte family of white blood cells, a
familywhosehallmarkisthepresenceofgranule
unique storage structures important in antimcrobial functions
(see section on Granules an
Degranulation, below).Neutrophils were discovered at the daw
of the immunological sciences; consequentelucidation of their
role in the immune r
sponse has been an ongoing process stretchinover more than a
century. We now know th
they are key components of the innate immuresponse and vital in
immune function; unfo
tunately, their importance has often been ove
shadowed by breakthroughs in the study of tadaptive immune
response (4). Admittedly, th
situation is exacerbated by neutrophils notorous experimental
intractability: They exhibi
short life span and are terminally differentiatepreventing
growth in tissue culture. The sta
dard tools of molecular biology, such as tranfection and RNA
interference, are of little u
when applied to these cells, and immortalizneutrophil-like cell
lines rarely reflect t
functional diversification of neutrophils. Fu
thermore, neutrophil-like cells studied in tisolation of a
culture dish most certainly do n
mimic the complex biological reality in tissuor circulation.
Conclusions from in vitro stu
ies should, therefore, be carefully interprete
460 Amulic et al.
-
7/24/2019 Neutrophil Function From Mechanism to Disease
3/33
Unfortunately, in vivo studies of neutrophil
function also raise concerns. Mouse neu-trophils, the preferred
model for in vivo
studies, differ in important aspects from theirhuman
equivalents. This is perhaps best
exemplified by the differences in the respectiveantimicrobial
repertoires and the numbers of
PMNs in circulation (30% versus 70% in miceand humans,
respectively).Despite these difficulties, no picture of the
immune response can be complete withouta comprehensive
understanding of the neu-
trophil and its functions. The extensive natureof neutrophil
research, however, precludes a
comprehensive review of the subject matter.In this review, we
intend to provide a survey
of basic neutrophil biology and function, whileemphasizing
recent advances in neutrophil re-
search and providing a critical assessment ofsome current
reports on PMN action.
Our survey of the neutrophil begins in
adult bone marrow where, under the in-struction of growth
factors and cytokines,
pluripotent hematopoietic cells differentiateinto myeloblasts, a
developmental cell type
committed to becoming granulocytes. As theseprecursor cells
mature to neutrophils, they syn-
thesize proteins that are sorted into differentgranules (5).
Traditionally, granules have been
subdivided into three different classes basedon their resident
cargo molecules: azurophilic,
specific, and gelatinase granules. Although this
subdivision is practical, these designations arelargely
artificial. Granules areformedthrough a
continuous process; vesicles bud from the Golgiapparatus and
fuse, producing granular struc-
tures. The content of these structures is dic-tated by the
transcriptional program active at
the time of their formation. As the maturingneutrophil
sequentially alters its transcriptional
profile, granule content changes, resulting in acontinuum of
granule species with overlapping
cargoes (6).
The release of neutrophils from the bonemarrow is tightly
regulated in healthy in-
dividuals: Chemokines control the passageof PMNs into
circulation and maintain a
pool of cells ready for release in case of
infection. Indeed, the number of neutrophils
drastically increases during infection and somediseases.
Interestingly, neutrophils circulate
for only approximately 68 h and are amongthe shortest-lived
cells in the human body.
Although the reason for this short life is unclear,
it may ensure neutrophil integrity; this hypoth-
esis is bolstered by observations that apoptosisprevents the
release of noxious molecules.Still, the question of why evolution
opted for
eliminating neutrophils quickly as opposedto reducing leakage of
their dangerous cargo
remains an unanswered and intriguing mystery.Mature neutrophils
emerge from the bone
marrow intent on pursuing one simple, yetessential, question:
Has host integrity been
compromised by potentially harmful invaders?
Should the answer prove to be yes, theneutrophil must swiftly
enact a carefully
choreographed process to locate, attack, anddestroy the
potential threat. At its disposal is
an impressive arsenal of antimicrobial weaponsthat are deadly,
indiscriminate, and brutish in
their application. Although effective in theirdestructive
capacity, these weapons can prove
to be just as dangerous to the host cells as totheir intended
targets, the microbial invaders.
Therefore, their deployment must be executedwith exquisite
precision and timing, at locations
where they are both contained and effective.
How then does the neutrophil locate andidentify infections? How
does it transition
at the correct time and place from an in-active cellular
bystander to a fully activated
microbial killing machine? This transitionprocess, during which
the neutrophil inte-
grates a complex barrage of environmentalcues and translates
them into specific actions,
is known as neutrophil activation. As itpursues microbes, the
neutrophil will enact an
impressive multitude of cellular mechanisms:It will mobilize
secretory vesicles and granules,
identify chemotactic gradients and traverse
them through destruction and reorganizationof the actin
skeleton, penetrate the endothelial
barrier and navigate a course through thebasement membrane, and
begin transcription
of cytokines for recruitment of new immune
www.annualreviews.org Neutrophil Functions 461
-
7/24/2019 Neutrophil Function From Mechanism to Disease
4/33
Selectins:transmembraneglycoproteins thatmediate cell
adhesionvia binding to sugarmoieties
Integrins:transmembranereceptors that mediateattachment to
theextracellular matrix, aswell as direct cell-cellinteraction
andsignaling
Oxidative/respiratoryburst: a rapid increasein oxygenconsumption
upon
neutrophil activationdue to production ofROS by the
NADPHoxidase
cells. Ultimately, upon arriving at the infection
site, it will seek the insulting pathogens andunleash its
extensive arsenal of antimicrobial
weapons. The initiation of these processes oc-curs in the
bloodstream, where the neutrophil
acts as a monitor for host distress, patrollingvessels and
vigilantly seeking out indications of
an incipient inflammatory response.
NEUTROPHIL ACTIVATION
At inflammatory sites, bacterial-derived and
host-produced inflammatory signals areabundant; these compounds
stimulate the
endothelial cells near the inflammatory site.These stimulants,
such as the bacterial-derived
lipopolysaccharide (LPS) and fMLP, as wellas the classical
chemoattractants and cytokines
tumor necrosis factor (TNF)-, interleukin(IL)-1, and IL-17,
prompt endothelial cells to
produce adhesion molecules on their luminal
side: the P-selectins, E-selectins, and severalmembers of the
integrin superfamily, the
ICAMs (5). As neutrophils traverse the circu-latory system, they
continuously and randomly
probe the vessel wall; the postcapillary venules,where flow
dynamics and the constricted space
are particularly amenable to increased randomprobing, are often
the best-suited location
for neutrophils to encounter the stimulatedendothelial cells (7,
8).
On the surface of neutrophils, two constitu-
tively expressed proteins are critical for recog-nition of the
endothelial inflammatory signals:
the glycoprotein P-selectin glycoproteinligand-1 (PSGL-1)
andL-selectin(9, 10). Upon
random contact with the endothelium, thesemolecules engage the
P- and E-selectins of
endothelial cells, resulting in selectin-mediatedtethering of
neutrophils to the vessel wall.
This is followed by a characteristic rolling ofneutrophils along
the endothelium. It is here
that the complex activation cascade begins
and the neutrophil commitment to microbialkilling commences.
What changes occur in the
neutrophil at this early time point?The engage-ment of PSGL-1
and L-selectin on neutrophils
activates a variety of kinases, including Src
family kinases, Syk, phosphoinositide 3-kina
(PI3K), and p38 mitogen-activated protekinase (1113). This
cascade initiates a numb
of changes in neutrophil biology and sets tstage for integrin
activation and firm adhesio
After selectin-mediated rolling, neutroph
enter a firm adhesion state mediated by t
2 integrin family of proteins (LFA-1 anMac-1 proteins on the
neutrophil); firm adh
sion is characterized by the arrest of neutroph
rolling in preparation for transendothelmigration (13, 14). As
the neutrophil ro
along the endothelium, interaction wiselectins,
chemoattractants, cytokines, a
bacterial products results in activation aclustering of the2
integrins on the surface
the neutrophil (15, 16). The 2 integrins th
engage their endothelial ligands, members the ICAM-1
immunoglobulin superfami
resulting in arrest of neutrophil rolling anfirm adhesion. This
integrin engagement,
well as continuing input from inflammatochemoattractants and
cytokines, prepares t
neutrophil for its final chemotactic pursuit: Tcell spreads,
producing a leading-edge lame
lipodium where chemokine and phagocyreceptors are concentrated,
the cytoskeleton
rebuilt and targeted toward movement alonchemotactic gradients,
and initiation of t
neutrophil oxidative burst begins (17, 18).
Now firmly adhered, the neutrophil munegotiate a path through
the endothelium in
the underlying tissue. In a process dependeon 2 integrins and
ICAMs, neutroph
crawl along the vessel wall until a preferrsite of
transmigration is reached (192
Upon arrival at an endothelial cell junctioncomplex interaction
between (a) the neutroph
integrins and their endothelial partners a(b) neutrophil surface
proteins and vario
endothelial junction molecules results in tranmigration through
the endothelial juncti
(13). Once through the endothelial linin
the neutrophil must navigate the basememembrane, a protein mesh
consisting large
of laminins and collagen type IV. Speculatioabounds that granule
proteases assist in th
migration by digesting the protein me
462 Amulic et al.
-
7/24/2019 Neutrophil Function From Mechanism to Disease
5/33
-
7/24/2019 Neutrophil Function From Mechanism to Disease
6/33
extracellular traps) (see the section on Neu-
trophils and the Elimination of Microbes,below).
The initiation of these microbicidal actionsindicates the final
stage of the neutrophils
journey through the activation process. How-ever, a prominent
question remains largely
unanswered by the preceding exposition: Whatexactly is meant by
the (admittedly ambiguous)phrase neutrophil activation? A quick
scan
of the literature presents the inexperiencedreader with a
sometimes rather conflicting (and
overwhelming) view of neutrophil activation.In fact, one could
be (erroneously) led to
believe that neutrophil activation refers only todirect
stimulation of the oxidative burst, as this
has been the canonical in vitro activation assayfor decades.
This is, however, an oversimpli-
fied view of a complex process. The myriadinteractions that
occur during a neutrophils
journey toward an inflammatory site must be
parsed by the complex neutrophilic signalin
mechanisms, a process that gradually leato complete activation
and culminates in th
premiere killing functions of phagocytosdegranulation, and
NETosis. It is, therefor
more insightful to view neutrophil activatio
as a continuum of processes, priming step
and signal cascades with varying effects anoutcomes, all focused
on the realization one goal: the transition of naive,
circulatin
neutrophils to their microbe-eliminatintissue-resident
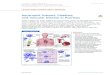
counterparts (Figure 1).
NEUTROPHILS AND THEELIMINATION OF MICROBES
The basic instruction set of the activatneutrophil is both
effective and ruthless
its simplicity: (1) kill microbes, (2) do nharm to the host, and
(3) when in doubt, s
rule 1. To fulfill this antimicrobial agend
Neutrophil
Endothelial cell
PSGL-1,L-selectin
P-selectin andE-selectin
IntegrinICAM
Phagocytosis
Degranulation
Cytokine secretion
NETs
a Capture b Rolling c Firm adhesion
Figure 1
Neutrophil recruitment to sites of inflammation. The circulating
neutrophil must recognize signs of
inflammation and migrate to areas where its antimicrobial
arsenal is needed for the elimination of infection(a) Close to the
inflammatory sites, stimulated endothelial cells expose a class of
molecules, the selectins,which serve to capture circulating
neutrophils and tether them to the endothelium. (b)
Selectin-mediatedrolling along chemoattractant gradients then
ensues, followed by (c) integrin-mediated firm
adhesion.Subsequently, the neutrophil traverses through the
endothelium and arrives at the site of inflammation.Here, the
neutrophil releases cytokines that recruit other immune cells, and
it begins to implement itsantimicrobial agenda. Among the processes
employed are engulfment of microbes via
receptor-mediatedphagocytosis, release of granular antimicrobial
molecules through degranulation, and formation ofneutrophil
extracellular traps (NETs).
464 Amulic et al.
-
7/24/2019 Neutrophil Function From Mechanism to Disease
7/33
Inflammation:recruitment andactivation of imcells upon
infecinjury; whenuncontrolled it
tissue damage
neutrophils possess an array of toxic weapons
that are carefully regulated through controlledmechanisms. These
antimicrobial weapons
vary considerably in their methods of actionand thus reflect the
neutrophils attempt to
exploit any and all weaknesses that microbesmight present during
the course of infection.
An understanding of these weapons, their
action, and their method of release is criticalto understanding
neutrophil function.
Granules and Degranulation
The neutrophil must safely transport a plethora
of dangerous substances through the blood-stream and then
correctly deploy them at the
appropriate time. Therefore, it comes as nosurprise that a
specialty storage organelle has
evolved in neutrophils: the granule. Expect-edly, these
structures are replete with specifi-
cally tuned mechanics that address the unique
needs of neutrophils. Granules are, however,
far more than just latent repository organellesfordangerous
substances; they areactiveand in-
dispensable participants in almostall neutrophilactivities
during inflammation.
As mentioned above, there are threefundamental types of granules
in neutrophils
(Figure 2). Azurophilic granules (also known
as peroxidase-positive or primary granules) arethe largest,
measuring approximately 0.3 M
in diameter, and are the first formed duringneutrophil
maturation. They are named for
their ability to take up the basic dye azure A andcontain
myeloperoxidase (MPO), an enzyme
critical in the oxidative burst (32, 33). Othercargo of this
granule class include the defensins,
lysozyme, bactericidal/permeability-increasingprotein(BPI), anda
number of serine proteases:
neutrophil elastase (NE), proteinase 3 (PR3),and cathepsin G
(CG) (34). As such, these
granules are brimming with antimicrobial
Granule type Primary
(azurophilic)
Myeloblast Promyelocyte Myelocyte Metamyelocyte Band cellStage
offormation
Myeloperoxidase
Defensin
Degranulationpropensity
Lysozyme
Elastase
Lactoferrin
Gelatinase
Complement receptor Characteristicproteins
Otherproteins
Cathepsin G, PR3,BPI, azurocidin,sialidase,-glucuronidase
Gp91phox/p22phox,CD11b, collagenase,hCAP18, NGAL, B12BP,SLPI,
haptoglobin,pentraxin 3,oroscomucoid,2-microglobulin,heparanase,
CRISP3
Gp91phox/p22phox,CD11b,
MMP25,arginase-1,2-microglobulin,CRISP3
Gp91phox/p22pCD11b, MMP25,FPR, alkalinephosphatase, CDCD13,
CD14,plasma proteins
FcRIII
Secretovesicle
Tertiary(gelatinase)
Secondary(specic)
PMN
Figure 2
Neutrophil granules. Neutrophil granules carry a rich variety of
antimicrobials and signaling molecules. They are typically
dividthree types (primary or azurophilic, secondary or specific,
and tertiary or gelatinase). Additionally, structures called
secretory vesare also considered to be a granule subset.
Considerable overlap exists in the cargo of the different granules,
and their contents sdetermined by the timepoint during
hematopoiesis at which they are produced (5). Granules also differ
in their ability to mobilizsecretory vesicles being the first to
fuse with the plasma membrane and the azurophilic granules
demonstrating the least degranupropensity.
www.annualreviews.org Neutrophil Functions 465
-
7/24/2019 Neutrophil Function From Mechanism to Disease
8/33
compounds and function as a primary reposi-
tory for the molecular weaponry of neutrophils.The second class
of granules, the specific (or
secondary) granules, are smaller (0.1 Mdiameter), do not contain
MPO, and are char-
acterized by the presence of the glycoproteinlactoferrin. These
granules are formed after
azurophilic granules; they also contain a widerange of
antimicrobial compounds includingNGAL, hCAP-18, and lysozyme (33,
35). The
third class, the gelatinase (tertiary) granules, arealso
MPO-negative, are smaller than specific
granules, and contain few antimicrobials,but they serve as a
storage location for a
number of metalloproteases, such as gelatinaseand leukolysin.
These granules are also the
last population of granules formed duringneutrophil maturation
(5). Finally, a fourth set
of structures, the secretory vesicles, are alsocommonly
considered part of the neutrophil
granule family. In contrast to the classical
granules, these do not bud from the Golgi,but instead are formed
through endocytosis
in the end stages of neutrophil maturation(36). Consequently,
their cargo consists pre-
dominantly of plasma-derived proteins such asalbumin. The
membrane of secretory vesicles
serves as a reservoir for a number of importantmembrane-bound
molecules employed during
neutrophil migration.As a neutrophil proceeds through
activation,
granules are mobilized and fuse with either the
plasma membrane or the phagosome, releasingtheir contents into
the respective environment.
In both cases, the membrane of the granulebecomes a permanent
part of the target mem-
brane, thus altering its molecular composition(6). The different
classes of granules demon-
strate varying propensities for mobilization inresponse to
inflammatory signals: Azurophilic
granules are the most difficult to mobilize, fol-lowed by
specific granules, gelatinase granules,
and finally, secretory vesicles (3741). The
underlying mechanisms for this differentialmobilization are not
entirely understood, al-
thoughregulationofintracellularcalciumlevelsappears to play a
salient role (32, 39). Because
of this varying mobilization propensity, each
granule subset has been traditionally associat
with a particular stage of neutrophil activatioAfter neutrophils
contact the endothelium
stimulation through selectins and chemoattratants induces
mobilization of secretory ve
cles, whose membranes are rich in key facto
necessary for continued activation of the ne
trophil, including, among others, the
2 intgrins, complement and fMLP receptors, as was the FcRIII
receptor CD16 (5, 38, 39, 42
Fusion of the secretory vesicles with the plasmmembrane exposes
these components to the e
ternal environment. This results in the trantion to firm
adhesion, mediated by2 integr
interaction with the endothelium. As they prceed through the
endothelium, neutrophils a
exposed to further activationsignals that initia
mobilization of gelatinase granules, thereby rleasing
metalloproteases. The activity of the
proteases may help neutrophils traverse tbasement membrane,
although this has n
been conclusively demonstrated (43, 44).At the inflammatory
site, complete ac
vation of the neutrophil ensues, promptiinitiation of the
oxidative burst and mobiliz
tion of the azurophilic and specific granuleThese granules
either fuse with the phagosom
(see section on Phagocytosis, below), cotributing to the
antimicrobial activities of th
compartment, or fuse with the plasma mem
brane, releasing their potent antimicrobiinto the tissue. The
fusion of specific granul
with the plasma or phagosomal membrane isparticular importance
for the oxidative bur
as flavocytochrome b558, a component of thNADPH oxidase
machinery, resides in t
specific granule membrane (45). This fusiopermits assembly of
the NADPH oxidase com
plex and allows reactive oxygen species (ROproduction both
inside the phagolysosome an
outside of the cell. Degranulation of primaand secondary
granules contributes to t
creation of an antimicrobial milieu at the i
flammatory site and produces an environmeinhospitable to
invading pathogens.
The release of granular proteins during dgranulation presents
the astute observer wi
a tempting proposition: Could these granul
466 Amulic et al.
-
7/24/2019 Neutrophil Function From Mechanism to Disease
9/33
-
7/24/2019 Neutrophil Function From Mechanism to Disease
10/33
Table 1 Mechanism of action of neutrophil antimicrobial
proteins
Antimicrobial peptide Antimicrobial mechanisma
Cationic antimicrobial peptides
-defensins (HNP-1, HNP-2,
HNP-3, HNP-4)
Permeabilize membrane bilayers containing negatively charg
phospholipids Inhibit DNA, RNA as well as protein biosynthesis
Inhibition of bacterial cell wall synthesis
LL-37 Transmembrane pore-formingBPI Increase bacterial
permeability and hydrolysis of bacterial
phospholipids by binding to LPS
Histones Unknown mechanism
Proteolytic enzymes
Lysozyme Degrades bacterial cell wall
Proteinase 3 (PR3) Mechanism independent of a proteolytic
activity by binding to the
bacterial membrane
Neutrophil elastase (NE),
cathepsin G (CG)
Cleaves bacterial virulence factors and outer membrane
proteins Mechanism independent of a proteolytic activity by
binding t
the bacterial membrane
Azurocidin Mechanism independent of a proteolytic activity by
binding to the
bacterial membrane
Metal chelator proteins
Lactoferrin Alters bacterial growth by binding to iron, an
essential bacteri
nutrient Binds to the lipid A part of LPS, causing a release of
LPS fro
the cell wall and an increase in membrane permeability
Calprotectin Alters bacterial growth by sequestering manganese
and zinc
aOnly direct actions of neutrophil antimicrobial proteins on
microbes are listed in the table.
antimicrobials, is essential for designing appro-
priate in vitro conditions to probe mechanismsof action.
The neutrophil cationic antimicrobialpeptides include defensins
and cathelicidins.
Neutrophils mostly produce -defensins, aprotein family whose
members possess multi-
ple disulfide bonds and whose structures maychange under
physiological conditions and
increase their activity (48). A surprising num-
ber of functions are assigned to defensins, butnone have been
validated in vivo. Interestingly,
inhibition of bacterial cell wall synthesis (49)was recently
shown at low concentrations that
may be more similar to those present at inflam-matory sites.
Cathelicidins, including the well-
studied LL-37, are proteolytically processed
from larger proteins, and in addition to the
antimicrobial activity, they may potentiaDNA activation of
dendritic cells (DCs) (50)
Neutrophils also contain a number full-length cationic
antimicrobial protein
including BPI and histones. BPI is cationand binds LPS avidly,
much like its structur
cousin the LPS binding protein. BPI bindingLPS results in
increased bacterial permeabili
and hydrolysis of bacterial phospholipids; c
death then follows (51). Interestingly, histonare extremely
effective antimicrobials a
were one of the first antimicrobials describ(52). The
significance of histones (and of th
peptides derived from them) as microbiremains to be demonstrated
in vivo (5
Given their dual role as an architectu
468 Amulic et al.
-
7/24/2019 Neutrophil Function From Mechanism to Disease
11/33
Chronicgranulomatousdisease (CGD)caused by mutatrendering
theNADPH oxidas
nonfunctional,characterized bysusceptibility toinfection
andautoinflammatio
scaffold for DNA and as antimicrobials, their
in vivo significance is particularly difficult
todemonstrate.
The second class of neutrophil antimi-crobials encompasses a
broad assortment of
proteolytic enzymes that participate in microbedestruction.
Lysozyme destroys the bacterial
wall, making it an obvious antimicrobial, asshown in mice
deficient in this enzyme (54).Surprisingly, this occurred
independently of its
enzymatic activity (55). Neutrophils also con-tain several
serine proteases (including PR3,
CG, and NE, collectively known as the serpro-cidins) that
exhibit differing specificities. They
are tightly regulated intra- and extracellularlyby serpins,
indicating that their activity is
deployed under specific conditions. NE cleavesenterobacterial
virulence factors with high
specificity (56), indicating the possibility of thecoevolution
of microbial virulence factors and
antimicrobial effectors. Of further interest, NE
mutations in humans, but not genetic ablationof this enzyme in
mice, result in neutropenia.
This can be rescued by the administration ofrecombinant
granulocyte macrophage colony-
stimulating factor (GM-CSF); however, thesepatients still
exhibit significant susceptibility
to infections. Mice deficient in NE or CGare highly susceptible
to bacterial and fungal
infections (57, 58). Another protein, azuro-cidin, is a member
of the same family but lacks
protease activity. Unexpectedly, it still kills
microbes, suggesting that these proteins mayall have
antimicrobial activity independent
of proteolysis, perhaps as a result of theircationicity. These
serine proteases also play a
salient role in autoimmunity (see discussion insection on
Autoimmunity, below) (59).
The final class of neutrophil antimicrobialsconsists of a number
of proteins that chelate
essential metals from microbes and possiblyimpact bacterial
growth. Two of these chela-
tors are lactoferrin, first identified in milk,
which binds preferentially to iron, and cal-protectin (also
called S100A and many other
names),whichsequesterszinc(60)andresultsinnutritional immunity
(61).
Reactive Oxygen Species
Upon activation, neutrophils produce ROS ina process called the
respiratory burst. It is mis-
leading to think of ROS as a single entity be-cause they differ
in their stability, reactivity, and
permeability to membranes (62). However, allROS can modify and
damage other molecules,
properties exploited by the host cell for signal-ing and
antimicrobial action.
The NADPH oxidase complex assembleson the phagosomal and plasma
membranes
and begins the reactive oxygen cascade by
reducing molecular oxygen to superoxide.Downstream of
superoxide, many potential
reactions can occur (for details, see References6264).
Superoxide, though not a strong
oxidant, rapidly dismutates, forming hydrogenperoxide.
Superoxide can also react with nitric
oxide, which is produced at high levels atinflammatory sites, to
form peroxynitrite, a
strong oxidant. Upon degranulation into thephagosome, MPO can
react with hydrogen
peroxide to produce various reactive species,including
hypohalous acids. Hypochlorous
acid, thought to be the major product of MPO
in the phagosome, is more reactive than su-peroxide and is
antimicrobial in vitro. Thus, it
is assumed to have direct antimicrobial effectsin the phagosome.
However, a theoretical
model of the phagosome suggests that most ofthe hypochlorous
acid produced would react
with host proteins before reaching the bac-terium. This model
predicts that chloramines,
produced when hypochlorous acid reactswith amine groups, may be
the most relevant
antimicrobial actors in the phagosome (65).ROS are clearly
important for neutrophil
antimicrobial activity: Neutrophils from
chronic granulomatous disease (CGD) patientskill microbes
poorly, making these patients
susceptible to many infections. Interestingly,CGD patients can
control catalase-negative
bacteria, which produce, but do not degrade,their own hydrogen
peroxide, thus providing
a substrate for reactions downstream in thereactive oxygen
cascade (66). NADPH ox-
idase is also implicated in the regulation of
www.annualreviews.org Neutrophil Functions 469
-
7/24/2019 Neutrophil Function From Mechanism to Disease
12/33
inflammation, which explains why CGD
patients often suffer from autoinflammatorydiseases (67).
Paradoxically, although MPO is requiredfor neutrophil
microbicidal activity in vitro,
MPO-deficient individuals do not have strikingclinical
manifestations (68, 69). Some MPO-
deficient individuals suffer from frequent or se-vere
infections, especially
withCandidaspecies,andafewhavebeenmistakenforCGDpatients.
However, most MPO-deficient individuals inthe developed world
have apparently normal
immunity. The mild effects of MPO deficiencysuggest that MPOs
products are not essential
for antimicrobial action. Indeed, in the absenceof MPO, other
reactive species (e.g., superox-
ide, hydrogen peroxide, hydroperoxyl radical,peroxynitrite) can
still be produced in the
neutrophil phagosome; hydroperoxyl radical
ispredictedtobepresentatantimicrobialconcen-
trations (65). However, there may be a broader
reason for this discrepancy. Modern technolo-gies can
distinguish between individuals who
are partially and completely MPO deficient,and partial MPO
deficiency does not correlate
with pathology (70). Residual activity of MPOmay be sufficient
for antimicrobial activity: In
the case of CGD, even 1% of normal NADPHoxidase activity leads
to an improved prognosis
(71). Epidemiological studies distinguishingthe degrees of MPO
deficiency and their
correlation with clinical manifestations may be
necessary to understand the function of MPO.In addition to
direct antimicrobial action,
ROS can modify host molecules. Becausethese species are highly
reactive, they are often
thought to be too nonspecific to be involved insignaling.
However, specificity can be achieved
on the submolecular level, by cellular redoxbuffering systems
and by limited diffusion of
ROS owing to their short half-lives (72). Awell-studied example
of ROS in signaling is
the reversible regulation of various targets
(including phosphatases, metalloproteinases,and caspases) by
direct oxidation of cysteine
residues. In addition, neutrophil granuleproteases can be
regulated by oxidative inacti-
vation of their inhibitors or by direct oxidation
(73, 74). Furthermore, superoxide generati
leads to an ionic influx into the phagosome compensate for
charge; this may activate gra
ule proteases by releasing them from their ptative matrix (75).
There is controversy aroun
which ions and which channel are responsib
for charge compensation, but this theory
protease activation is certainly intriguing (69Studies of ROS
are hampered by variotechnical issues. Ideally, a probe for RO
should be specific, targetable to particuintracellular
compartments, and capable
being used in vivo. Traditional probes fROS do not meet these
specifications;
addition, the probes often become radicspecies (76). One
promising new approa
for ROS detection that meets these criteria
the use of redox-sensitive fluorescent proteibased probes, such
as roGFP and HyP
(76). Other methods that can be used in viinclude transcription
profiling of superoxi
or hydrogen peroxidesensitive genes as was the detection of
relatively stable products
reactive oxygen using mass spectrometry (76
Phagocytosis
Phagocytosis is the major mechanism to rmove pathogens and cell
debris. It is an activ
receptor-mediated process during which a paticle is internalized
by the cell membrane in
a vacuole called the phagosome. As with oth
phagocytes, the mechanistic details of internaization depend on
the type of interaction b
tween the neutrophil and the microorganismInteraction can be
direct, through recognitio
of PAMPs by pattern-recognition receptors, opsonin mediated. The
latter mechanism is be
ter characterized and includes two prototypicexamples:
FcR-mediated phagocytosis, whi
relieson theformationof pseudopodextensiofor engulfment of
IgG-opsonized particles, a
complement receptor-mediated phagocytoswhich does not require
membrane extensio
or pseudopods (77).
After engulfment, the nascent phagosomis relatively benign to
microorganisms, acqu
ing its lethal properties only after a drast
470 Amulic et al.
-
7/24/2019 Neutrophil Function From Mechanism to Disease
13/33
Autophagy: a pin which cellulacontents are degin
lysosomes,especially inconditions of nu
scarcity and infe
maturation process. Our understanding of
this process is largely based on studies inmacrophages, and
although these are certainly
instructive, essential differences exist in neu-trophils.
Macrophage phagocytosis follows an
endocytic maturation pathway: In neutrophils,phagosome
maturation happens upon fusion of
granules to the phagosome, whereby deliveryof antimicrobial
molecules into the phagoso-mal lumen occurs. Simultaneously,
assembly
of the NADPH oxidase on the phagosomalmembrane allows ROS
production, and jointly,
these two mechanisms create an environmenttoxic to most
pathogens. Neutrophil phago-
somal pH regulation also differs significantlyfrom that observed
in macrophages. While the
macrophage phagosome gradually acidifies,neutrophil phagosomal
pH is initially alkaline
(78) and remains neutral for prolonged periodsof time (79). The
maintenance of this alkaline
pH is essential for the activation of the major
serine proteases NE and CG, and it is sustainedvia NADPH oxidase
activity, despite contin-
uing fusion of acidic granules. Key events ofthe maturation
process are described in more
detail in Reference 80.Not all pathogens succumb to the
hostile
environment of the phagosome. In fact, somehave evolved
strategies to survive inside neu-
trophils. These strategies include interferingwith engulfment,
modulating phagosome
maturation, and creating a more hospitable
intraphagosomal environment. The polysac-charide capsule
expressed by Staphylococcus
aureusconfers antiphagocytic properties (81).Helicobacter pylori
can disrupt targeting of
NADPH oxidase to the phagosome so thatsuperoxide anions
accumulate extracellularly
rather than in the phagosome (82). Francisella
tularensisprevents triggering of the oxidative
burst and also inhibits ROS production inresponse to other
stimuli (83). Finally, other
pathogens, such asSalmonella typhimuriumand
Streptococcus pyogenes, can efficiently block gran-ule fusion
with the phagosome (84, 85). The
variety of mechanisms evolved by intracellularpathogens to
resist killing and enable survival
within the phagosome further emphasizes the
importance of phagocytosis in the innate
immune defense.
Neutrophil Extracellular Traps
Upon stimulation, neutrophils can undergoNETosis, an active form
of cell death that
leads to release of decondensed chromatin intothe extracellular
space (86, 87). The fibrousstructures termed NETs contain histones
as
well as antimicrobial granular and cytoplasmicproteins (88).
NETs trap many types of mi-
crobes ex vivo and have been found in variousdisease models in
vivo; they are thought to
kill microbes by exposing them to high localconcentrations of
antimicrobials (89).
The mechanism of NET formation is notcompletely understood. The
reactive oxygen
pathway is involved, as NADPH oxidase andMPO are required for
NET formation in re-
sponse to chemical and biological stimuli (87,
90, 91). Nitric oxide donors can induce NETsvia a mechanism that
also requires ROS (90), a
finding that awaits genetic confirmation. All ac-tivators of NET
formation tested so far require
ROS production. S. aureusmaybe an exception,although those
experiments were done using
pharmacological inhibitors, not cells deficientin ROS production
(92). Upstream of NADPH
oxidase, the Raf-MEK-ERK pathway is impli-cated in NET formation
(93), but further along
in the process, NE translocates from the gran-
ules to the nucleus and degrades histones, lead-ing to chromatin
decondensation (94). Histone
citrullination may also play a role in NET for-mation, although
this has not been confirmed
in primary human neutrophils (9597). Au-tophagy is also thought
to be required for NET
formation, but this has so far been shown onlyusing a
nonspecific inhibitor of autophagy (98).
The majority of research on NETs has beenconducted ex vivo.
Ideally, to test the relevance
of NETs, a NETs knockout organism should
be generated to investigate its response topathogens.
Unfortunately, it is not possible to
eliminate the main components of NETsDNA and histonesfrom an
infection model.
Moreover, the factors that are important for
www.annualreviews.org Neutrophil Functions 471
-
7/24/2019 Neutrophil Function From Mechanism to Disease
14/33
Cystic fibrosis:caused by defects inthe CFTR
iontransporter,characterized by thick,sticky mucus and
decreases in lung anddigestive function
NET formation, such as NADPH oxidase,
MPO, and NE, are also critical for other an-timicrobial
neutrophil functions. For now, the
evidence for the relevance of NETs is indirect.On the one hand,
bacteria that express DNases
as virulence factors disseminate more efficientlyin the host,
which may point to evolutionary
pressure to avoid entrapment by NETs (99,100). In addition, a
persistent Aspergillusinfection in a CGD patient was cleared
after
gene therapy, which restored NADPH oxidaseactivity, NET
formation, and NET-mediated
but not phagocytosis-mediated killing by thepatients neutrophils
ex vivo(101). On the other
hand, the immune system has redundant mech-anisms to fight
infection, and it may be that
NETs are especially important under certainconditions, such as
during infections with large
pathogens that are not readily phagocytosed.NETs can also have
detrimental effects on
the host. Because NETs expose self molecules
extracellularly, they lead to autoimmunity:NETs have been
implicated in systemic
lupus erythematosus (SLE), an autoimmunedisease characterized by
the formation of
autoantibodies, often against chromatin andneutrophil components
(102106) (see section
on Autoimmunity, below). Platelet-inducedNETs, formed during
sepsis, are associated
with hepatotoxicity due to tissue damage(107). Platelets also
bind to NETs, raising the
possibility that NETs nucleate blood clots in
the context of deep vein thrombosis (108).NETs have also been
observed in the airway
fluids of cystic fibrosis patients, where theymay increase the
viscosity of the sputum and
decrease lung function (109).
NEUTROPHILS IN IMMUNECELL CROSS TALK
Neutrophils participate in the communica-
tion networks that form the foundations ofimmunity, issuing
instructions to practically
all other immune cells. As one of the first celltypes to arrive
at sites of infection, neutrophils
secrete cytokines and chemokines critical in the
unfolding of the inflammatory response and in
establishing the correct environmental cond
tions to launch the adaptive immune responThe cytokines released
by PMNs are oft
synthesized de novo. Although neutrophtranscribe little after
leaving the bone marro
once activated, these cells undergo a trascriptional burst that
results in the synthe
of signaling molecules (110, 111). Comparwith other immune cells
(e.g., macrophageneutrophils typically produce lower amoun
of cytokines per cell, but they are so abundaat inflammatory
sites that their contributi
to total cytokine levels is significant (4). Futhermore,
neutrophil-secreted proteases c
modulate signaling networks in vivo throucytokine processing
(112).
The initial neutrophil cytokine responsean appeal for
immunological reinforcemen
The most abundantly produced cytokine, IL-primarily serves to
recruit other neutroph
(113). Similarly, neutrophil-derived proinflam
matory IL-1 and TNF-induce other ceto produce neutrophil
chemoattractants (11
115) (for a comprehensive list of cytokinproduced by
neutrophils, please see Referenc
115, 116). In addition to cytokines, neutrophrelease other
signaling mediators, includi
granule contents (117), lipids (118), and ROsuch as hydrogen
peroxide (119). They al
communicate via cell-cell contact (120). Hewe provide examples
of how neutroph
interact with other cells to shape the immun
response (seeFigure 3).
Monocytes and Macrophages
As they respond to infection or injurneutrophils and their
relatives in the mon
cyte/macrophage lineage coordinate thactivities, leading to
alternating waves of r
cruitment of these two cell types. Macrophagand patrolling
monocytes are among the init
detectors of PAMPsand endogenousactivatothe danger-associated
molecular patterns (12
and these cells work to summon large numbe
of neutrophils to the inflammatory locus. Tinflux of neutrophils
is followed closely by t
arrival of monocytes, suggesting a causal lin
472 Amulic et al.
-
7/24/2019 Neutrophil Function From Mechanism to Disease
15/33
Neutrophil
Neutrophil
Neutrophil
Neutrophil Monocyte
T cell
T cell
Macrophage
Lymph
Blood
Tissue
Activation anddifferentiation
ROS?
Arginase?
IFN-
IFN-
IL-12
NK cell
DC
DC
DC
Activation
Activation
Crosspriming
Bacteria
Th1
Antigenpresentation
CD4+
T cell
CD8+
T cell
DC
DC
Figure 3
Neutrophil communication with other immune cells. Neutrophils
interact with a variety of cell types. They are important both
recruitment of monocytes and dendritic cells (DCs) to infected
tissues and for enhancement of macrophage and DC activity.
Incontrast, in the lymph nodes, neutrophils impede DC function by
inhibiting antigen presentation to CD4+ cells. Neutrophils
alinteract with the adaptive arm of the immune system: They can act
as antigen-presenting cells by cross-presenting antigen to CDcells;
they also secrete IL-12, which activates T cells. T cells, in turn,
activate neutrophils by secreting IFN-. Finally, neutrophDCs and
natural killer (NK) cells colocalize and enhance each others
activity via receptor-receptor interactions and soluble med
behind these temporal dynamics. Indeed, neu-
trophils recruit monocytes via several differentmechanisms. They
express classical monocyte
chemoattractants such as CCL2 (MCP-1)(122), CCL3 (MIP-1) (123),
CCL20 (MIP-
3), and CCL19 (MIP-3) (124). Additionally,
and perhaps more unexpectedly, neutrophilsuse granule proteins
to induce extravasation
of monocytes in vivo, as shown for LL-37,azurocidin (HBP/CAP37),
and CG (125127).
Monocyte recruitment is also affectedindirectlyby neutrophils:
via upregulation of endothelial
adhesion factors, increase of transendothelialpermeability,
enhancement of production of
chemoattractants by other cell types, and mod-ulation of the
activities of these chemokines
via proteolytic processing (reviewed in 128).In addition to
recruitment, neutrophils mod-
ulate monocyte and macrophage cytokine
production (128), directly enhancing their
microbicidal activity (129). The circuitous
nature of the cross talk of these two cell typesbecomes obvious
during inflammation abate-
ment: Monocytes, recruited by neutrophils
and differentiated into macrophages, repressfurther neutrophil
chemotaxis and ensure
the appropriate removal of their postmortemremains (see section
on Neutrophils and
Resolution of Inflammation, below).
Dendritic Cells
Neutrophils can also recruit and activateDCs in vivo. This was
recently illustrated
in a mouse model of Leishmaniasis, wheresubcutaneous inoculation
of Leishmania majortriggered a massive and rapid infiltration
ofneutrophils (130). These cells secrete the
chemokine CCL3, recruiting DCs to the
site of inoculation and initiating a protective
www.annualreviews.org Neutrophil Functions 473
-
7/24/2019 Neutrophil Function From Mechanism to Disease
16/33
DC-SIGN:dendritic cellspecificintercellular
adhesionmolecule-3-grabbingnonintegrin
Granulocytereceptor 1 (Gr1):the anti-Gr1 antibodyRB6-8C5 reacts
withboth Ly6G (specificfor neutrophils) andLy6C (present onmany
immune celltypes)
Th17 cells: subset ofT helper cells thatproduce IL-17,important
in
inflammation andimplicated inautoimmunity
Th1 response (131). Interestingly, activated
neutrophils can induce the maturation of DCsin vitro through
specific receptor-receptor
interactions between Mac-1 and DC-SIGN,leading to local
secretion of TNF- (120).
In this case, the reduced levels of cytokineproduction foster
specificity, as only proximal
DCs receive the maturation signal. A similaractivation model was
earlier proposed forTox-oplasma gondii(132). Neutrophil-activated
DCs
produce the proinflammatory cytokine IL-12and induce
proliferation of T cells (120, 132).
However, some of these experiments shouldbe interpreted
cautiously because they are
based on the injection of the anti-Gr1 antibody(RB6), which
depletes neutrophils but may also
result in depletion of many other cell types inmice. The
anti-Ly6G monoclonal antibody is
more specific and hence a betterreagent for thistype of
experiment (133). The crucial role of
neutrophils in DC activation was recently con-
firmed using anti-Ly6G antibody depletion: InMycobacterium
tuberculosisinfection, timely traf-
ficking of DCs to lymph nodes and activation ofCD4+ T cells were
both dependent on PMNs.
Furthermore, this study demonstrated thatDCs presented bacterial
antigens when they
ingested infected neutrophils just as efficientlyas they did via
direct uptake ofMycobacterium(134). In sharp contrast to the above
findings,a separate study using an immunization model
showed that neutrophils recruited to lymph
nodes compete for antigen with DCs andmacrophages and that these
neutrophils inhibit
their interactions with T cells (135). It is possi-ble that
neutrophils have site-specific effects on
DCs and can be stimulatory at peripheral sitesand inhibitory in
the lymph nodes. Neutrophils
exhibit fascinating and somewhat enigmatic be-havior in the
lymph nodes, where they engage
in swarming activity in response to parasiticinfection (136).
The functions and mechanistic
details of these swarms are unknown andrepresent questions of
immense interest.
Natural Killer Cells
Studies of interactions between neutrophil and
natural killer (NK) cells have historically been
performed in vitro, and their interpretation
frustratingly difficult owing to the questioable purity of cell
preparations. Recently,
was shown that neutrophils, NK cells, and DCinteract in a menage
a trois involving bo
cytokine signaling and direct cell-cell conta(137, 138). In one
report, infection of mi
with Legionella pneumophila triggered prodution of IFN-by NK
cells; this was dependeon both PMN-derived IL-18 and DC-deriv
IL-12 (137). Similarly, human neutrophils, Ncells, and DCs
colocalize at inflammatory sit
and a positive feedback loop has been proposon the basis of in
vitro data. In this scheme, ne
trophils interact with a specific subset of DC(via CD18-ICAM-1
interactions), promptin
the DCs to produce IL-12p70, which in tustimulates
IFN-production by NK cells an
further activates neutrophils. Simultaneousneutrophils
alsoactivate NK cells by direct co
tact (139). Additional in vitro interactions b
tween neutrophils and NK cells are extensivereviewed in
Reference 138.
Lymphocytes
A surprising finding in recent years is the exte
sive cross talk between cells located at opposiends of the
immune spectrum. Previous
thought to belong to isolated compartmenneutrophils and T cells
shape and impa
each others functions, both qualitatively an
quantitatively (140). Neutrophils affect T cfunction indirectly
via DCs, as outlined abov
but can also influence T cell function directlPMNs secrete
IL-12, which may be crucial f
Th1 cell differentiation (141, 142). They alexpress several T
cell chemoattractants (11
as well as B cell development and maturatifactors (143, 144).
Cytokine communicati
occurs in both directions: For instance, IFN-which is secreted
by T cells, prolongs ne
trophil life span, induces gene expression, anincreases
phagocytic capacity (145). The
helper 17 (Th17) cell subset secretes IL-1
a key cytokine in the control of neutrophdynamics, which acts by
upregulating expre
sion of CXCL8 (IL-8), G-CSF, and TNF
474 Amulic et al.
-
7/24/2019 Neutrophil Function From Mechanism to Disease
17/33
Ulcerative colitype of inflammbowel diseasecharacterized byand
tissue erosiothe colon and re
by epithelial, endothelial, and stromal cells
(146). Collectively, these Th17-associatedcytokines increase
granulopoeisis as well as the
recruitment and life span of neutrophils.Neutrophils potentially
have suppressive ef-
fects on T cells via two proposed mechanisms:(a) L-arginine
depletion by release of arginase,
which inhibits T cell responses in vitro (147),and (b) hydrogen
peroxidemediated suppres-sion, as proposed in a cancer model (119)
(see
section on Cancer, below). Direct evidence ofsuch interactions
in vivo is still missing.
Interestingly, neutrophils influence CD8+
T cell responses by cross-presenting exogenous
antigens in vivo. Using mice in which profes-sional
antigen-presenting cells do not express
functional MHC class I, Beauvillain et al. (148)showed that
antigen-pulsed neutrophils can
induce differentiation of cytotoxic T cells.These striking
findings imply that neutrophils
have characteristics of antigen-presenting cells.
Neutrophils also appear capable of expressingMHC class II and
costimulatory molecules
under inflammatory conditions (149151),and they can present
antigen to CD4+ T cells
in vitro (152154). However, the functionalsignificance for
protective immunity remains
unclear, especially in light of the finding thatmouse
neutrophils that migrate to the lymph
node have a negative effect on CD4 responsesin an immunization
system (135). In humans,
there are large variations in the ability of
donors to express MHC class II (149, 151),suggesting concomitant
variations in the ability
to activate T cells, a finding that could haveimplications for
susceptibility to autoimmune
diseases. Therefore, neutrophil modulation ofadaptive immunity
seems to be highly complex
and is only now starting to be unraveled.
NEUTROPHILS ANDRESOLUTION OFINFLAMMATION
The lethal cargo of neutrophils is not onlydestructive toward
invading microbes, but
also harmful to host cells. Thus, neutrophil
deployment must be tightly controlled.
Although some collateral damage to host
tissues is inevitable during infection, neu-trophils must be
removed before they have
serious, detrimental effects on inflamed tissues.Resolution of
inflammation is an active process
that limits further leukocyte infiltration andremoves apoptotic
cells from inflamed sites.
This process is essential for maintenance oftissue homeostasis
and, if impeded, leads tononresolving inflammation, a
problematic
condition that contributes to many diseases.
Apoptosis and Clearance
Apoptosis is a central aspect of inflammation
resolution. Once neutrophils have executedtheir antimicrobial
agenda, they die via a built-
in cell-death program. However, not only doesapoptosis reduce
the number of neutrophils
present, it also produces signals that abro-
gate further neutrophil recruitment. Phagocy-tosis of apoptotic
neutrophils also reprograms
macrophages to adopt an anti-inflammatoryphenotype.
Neutrophil death is influencedby inflamma-tory mediators such as
GM-CSF and LPS and
by environmental conditions such as hypoxia,all of which prolong
neutrophil survival. The
signaling networks that regulate survival havealso been well
characterized. These networks
also control the expression of known antiapo-
ptotic (Mcl-1 and A1) or proapoptotic proteins(Bad, Bax, Bak,
and Bid), and they also activate
caspases (for an extensive review, see Reference155). Given that
neutrophils are terminally
differentiated, it is unexpected that moleculescontrolling cell
proliferation regulate survival.
Proposed to have prosurvival effects, one suchprotein is
survivin. It is expressed more highly
in immature neutrophils than in mature ones,but its expression
can be restored in mature
cells by inflammatory signals such as G-CSF or
GM-CSF. In line with these findings, survivinis also highly
expressed in neutrophils at sites
of inflammation, such as cystic fibrosis sputum,appendix
infiltrates, and intestines of patients
with ulcerative colitis (156).
www.annualreviews.org Neutrophil Functions 475
-
7/24/2019 Neutrophil Function From Mechanism to Disease
18/33
Wegenersgranulomatosis:vasculitis affecting thelungs, nose,
andkidneys; inflammationleads to reduced blood
flow, tissuedestruction, anddamage of vital organs
Prostaglandins andleukotrienes: lipidssynthesized
bycyclooxygenases and5-lipoxygenase,respectively, in thearachidonic
acidpathway; haveproinflammatoryfunctions including
leukocyte recruitment
Similarly, cyclin-dependent kinases func-
tion as prosurvival factors in neutrophils.Pharmacological
inhibition of these cell cycle
regulators induce caspase-dependent apoptosisand block life-span
extension by survival factors
(157). More recently, prosurvival effects werealso attributed to
proliferating cell nuclear
antigen (PCNA). This factor usually residesin the nucleus, where
it is involved in DNAreplication, but in neutrophils, it
associates
with procaspases in the cytosol and is thoughtto prevent their
activation. During apoptosis,
PCNA is targeted for proteosomal degradation,which correlates
with an increase in caspase-3
and caspase-8 activities. This mechanism is rel-evant in
Wegeners granulomatosis and sepsis,
where stabilization of PCNA is associated withresistance of
neutrophils to apoptosis (158).
Equally important for the resolution of in-flammation is the
proper removal of apoptotic
cells. This relies on the release of find-me
signals at early stages of cell death, which at-tract
phagocytes. Likewise, distinct eat me
signals are required for specific recognition ofapoptotic cells.
Ingestion of apoptotic cells by
macrophages drives the production of the anti-inflammatory
cytokines tumor growth factor
(TGF)- and IL-10 (155). Failure to clear theseapoptotic cells,
by contrast, results in secondary
necrosis and release of products that generateproinflammatory
signals (Figure 4).
Lipid Mediator Class Switch
Soluble mediators play a crucial role in theresolution of
inflammation. In neutrophils,
a particularly prominent role is assumed bylipid mediators. The
successful progression
of inflammation appears to hinge on a shiftin the composition of
secreted lipids. At early
stages of inflammation, neutrophils synthesizeproinflammatory
lipid mediators, such as
prostaglandins and leukotrienes. These arederived from
arachidonate precursor molecules
and are synthesized through the cyclooxy-
genase and lipoxygenase pathways. Duringthe later stages of the
inflammatory response,
neutrophils interact with various cell types in
their vicinity (epithelial cells, endothelial cel
fibroblasts, platelets, and leukocytes) and paticipate in the
transcellular biosynthesis of lip
mediators with anti-inflammatory and prorsolving activities,
such as lipoxins,resolvins, a
protectins. A major lipid mediator class swit
thus exists, governed by temporally regulat
expression of different lipoxygenases and tmobilization of
different fatty acid substrateThe different biosynthesis pathways
of pror
solving lipid mediators have been reviewed detail elsewhere
(118). Interestingly, microo
ganisms are also a source of lipid precursothat can be used by
neutrophils for resolv
synthesis. Thus, microbes also likely participain synthesis of
mediators with proresolvi
functions at the site of infection (159, 160).How do lipid
mediators contribute
the termination of inflammation? Lipoxin
resolvins, and protectins exert cell-type specieffects,
promoting monocyte/macropha
recruitment and activation while inhibitineutrophil functions.
The inhibitory effe
extends to all essential steps of neutropresponses: migration,
adhesion, and activatio
All three lipid mediators reduce neutrophrecruitment, a process
that involves the lipoxi
A4 receptor and the leukotriene B4 recept(BLT1) (161167). Ariel
et al. (168) also pr
posed an interesting mechanism of action flipoxins,resolvins,
andprotectinsin clearing i
flammatory sites. They showed that neutroph
exposure to these lipids increases expressiof CCR5 on the
surface of late apoptotic ne
trophils, leading to efficientsequestration of tchemoattractants
CCL3 and CCL5. The s
questration of these chemokines means they aunavailable to
recruit neutrophils to inflam
sites (168) (Figure 4). This mechanism complements other
anti-inflammatory process
in which chemokines are inactivated by netrophil proteases. Of
these lipids, lipoxins a
the most completely understood. In addition
neutrophil recruitment, lipoxins can inhibit tshedding of
L-selectin and the upregulation
2 integrins in response to proinflammatostimuli, thereby
reducing adhesion of ne
trophils to endothelial cells (169, 170). Final
476 Amulic et al.
-
7/24/2019 Neutrophil Function From Mechanism to Disease
19/33
TNF-IL-6
IL-10TGF-PGE-2
Neutrophil
Monocyte
Platelets
Lipoxins
Macrophage
Macrophage
ChemokinesApoptotic
neutrophil
NEToticneutrophil
LeukotrienesProstaglandins
?Microorganisms
LipoxinResolvinsProtectins
Chemokines
CCR5
Initiationof inammation
Resolutionof inammationLeukotrienes Prostaglandins TNF-
TGF-Lipoxins Resolvins Protectins IL-10
Chemokine clear
Figure 4
From inflammation to homeostasis: neutrophil apoptosis and lipid
mediator class switching in the resolution of inflammation. Asite
of infection, resident macrophages initiate an inflammatory
response, secreting proinflammatory cytokines and chemokines alert
the immune system and promote neutrophil recruitment. In the early
stages of inflammation, microbes trigger the productproinflammatory
lipid mediators, such as leukotrienes and prostaglandins, which
also recruit neutrophils. As inflammation progrswitch occurs, and
anti-inflammatory lipid mediators such as lipoxins, resolvins, and
protectins are produced. Notably, interactioneutrophils with
platelets induces the production of lipoxins. Anti-inflammatory
lipid mediators initiate the resolution of inflammby blocking
neutrophil and promoting monocyte recruitment. Monocytes
differentiated into macrophages ingest apoptotic neutdriving the
production of the anti-inflammatory cytokines tumor growth factor
(TGF)- and IL-10 and prostaglandin-E2 (PGEwhich drive the lipid
mediator class switch. Proresolving lipid mediators also promote
the expression of CCR5 on the surface of
apoptotic neutrophils, providing a means of scavenging
chemokines. Chemokine clearance upon phagocytosis of apoptotic
neutrby macrophages further contributes to the reduction of
neutrophil infiltration and the return to tissue homeostasis. The
contribumacrophages to the clearance of NETotic neutrophils, and
how this could impact inflammation resolution, is currently
unknowtimeline of the inflammation process from initiation to
resolution is summarized in the upper part of the figure.
Chronic obstrupulmonary dise(COPD): lung caused by
noxioparticles or gas,tobacco smokininflammation lelung
obstruction
lipoxins also impact neutrophil activation byinhibiting ROS and
peroxynitrite production,
NF-B activation, and IL-8 expression (170).In addition to
directly impacting neu-
trophil functions, lipid mediators promotenonphlogistic
(noninflammatory) phagocyto-
sis of apoptotic neutrophils by monocytes
and macrophages. In the presence of anti-inflammatory lipids,
engulfment of apoptoticneutrophils is notaccompanied by
thereleaseof
proinflammatory mediators, as typically occurs
during macrophage activation.Instead, produc-tionof
theanti-inflammatory cytokines TGF-and IL-10 is increased (163,
171).
Disorders Associated withNonresolved Inflammation
The failure of neutrophils to apoptose or mal-
functions in the removal of their apoptotic re-mains result in
chronic inflammation. These
conditions lead to the accumulation of cyto-toxic substances and
are associated with severe
pathologies, including cystic fibrosis, chronic
obstructive pulmonary disease (COPD), andrheumatoid arthritis
(RA). The severity of in-
flammation often directly correlates with poorclinical
outcome.
COPD is a major cause of death in indus-trialized nations, where
smoking is a prime
www.annualreviews.org Neutrophil Functions 477
-
7/24/2019 Neutrophil Function From Mechanism to Disease
20/33
Rheumatoid arthritis(RA): chronicinflammatory diseasethat
affects manytissues and organs butprimarily synovial
joints; severeinflammation causesdeformity
instigator of this disease. A chronic neutrophil
infiltration in the lungs of COPD patientspromotes tissue damage
and organ dysfunc-
tion. One of the key molecules controllingthe inflammatory
response in the lung is
leukotriene A4 hydrolase (LTA4H). Thisenzyme has two opposing
activities. First, its
hydrolase activity converts leukotriene A4 intoleukotriene B4, a
potent neutrophil chemoat-tractant and proinflammatory agent.
Second,
LTA4H is an aminopeptidase that inactivatesa specific neutrophil
chemoattractant, the
proline-glycine-proline tripeptide (PGP), thusconferring the
enzyme with anti-inflammatory
properties. Interestingly, tobacco smoke selec-tively inhibits
only the aminopeptidase activity
of LTA4H, promoting the accumulation ofboth leukotriene B4 and
PGP. This in turn
promotes neutrophil recruitment and fuelschronic lung
inflammation (172).
Another prime example of a disease linked to
nonresolving inflammation is RA. Neutrophilsare the most
abundant leukocytes present in the
synovial fluid of RA patients, and their role inpathogenesis has
been demonstrated in several
animal models. These models primarily usedneutrophil depletion
or adoptive transfer of
wild-type neutrophils in leukotriene-deficientmice (173175). In
one model, synthesis
of leukotriene B4 by neutrophils in jointsis essential for
disease development (174).
Leukotriene B4 can act in an autocrine manner
via the neutrophil receptor BLT1 to promotethe recruitment of a
first wave of neutrophils
into the joint. Later, the recruitment of asecond wave of
neutrophils is independent of
this leukotriene B4BLT1 pathway. At thisstage, immune complexes
are essential for
stimulating infiltrating neutrophils to deliverIL-1into the
joint. This in turn induces the
production of chemokines by synovial tissuecells and sustains
neutrophil recruitment (175,
176). These studies exemplify the complex
regulation cascades involving lipids, cytokines,and chemokines
that orchestrate neutrophil
recruitment in chronic inflammation. Theyalso demonstrate the
cross talk between neu-
trophils and other immune cells discussed in
the previous section. It is, however, unknow
whether all neutrophils are capable of adaptinto the changing
chemoattractant environme
or if different subsets of neutrophils are sucessively involved.
The relevance of this mod
in human disease remains to be establishe
although the clinical similarities between th
mouse model and human RA are encouragin
NEUTROPHILS IN DISEASE
Neutrophils areprominent players in theinna
immune response and the clearance of infetion, a subject
addressed in several promine
reviews. However, neutrophil action can al
support disease progression in other illnesseA host of
autoimmune disorders belong to th
category. In addition, certain malignant canceare also prime
examples of illnesses in whi
neutrophils play a salient role.
Cancer
The link between cancer and inflammati
was noted as early as 1863 by Rudolf Vircho(177). Since then, it
has been proposed th
neutrophil-derived ROS have the potential initiate tumor
formation by genotoxic stre
and induction of genomic instability. Althouthis has been
demonstrated in vitro (178, 179
convincing evidence for PMN-mediated DNmutagenesis in vivo is
still lacking. Neutroph
do, however, impact cancer progressio
They are abundant in tumors and influentumor development through
several secret
mediators, including cytokines, ROS, amatrix-degrading proteases
(reviewed in Re
erence 180). The majority of findings suppoa protumor and
antihost effect of the
cells; clinical studies indicate that neutrophinfiltration of
tumors is associated with poor
prognosis (181, 182). Indeed, some canceseem to actively recruit
neutrophils throug
production of IL-8 (183). In agreement withis, antibody
depletion of neutrophils reduc
tumor growth (184). The protumor functio
of neutrophils operates at multiple leveincluding production of
angiogenic facto
(185), enhancement of metastasis (186), an
478 Amulic et al.
-
7/24/2019 Neutrophil Function From Mechanism to Disease
21/33
Acute-phaseproteins: secreliver, concentraplasma changes 25% or
more duinflammation
suppression of the antitumor immune response
(119, 187). Using the anti-Ly6G antibody,Fridlender and
colleagues (187) depleted neu-
trophils and confirmed their tumorigenic role.Moreover, the
study showed that neutrophils in
the tumor microenvironment could, under cer-tain circumstances,
be induced to target their
cytotoxic arsenal at tumor cells, whose growththey usually help
to fuel. Pharmacologicalinhibition of TGF- signaling led tumor-
associated neutrophils to assume a heightenedproinflammatory
state, causing a reduction in
tumor growth. These alternatively activatedneutrophils underwent
a complete reversal in
their effect on CD8+T cells, serving to activaterather than
suppress these cells. Differential
neutrophil responses were also demonstrated ina melanoma study.
In this instance, increased
systemic levels of the acute-phase proteinserum amyloid A
(SAA-1) induced neutrophils
to secrete the anti-inflammatory cytokine IL-
10, which also inhibited T cell responses. Crosstalk with
invariant NKT cells could counter
this response, restoring a proinflammatoryactivation status
(188). Thus, investigation of
neutrophils in cancer has revealed considerableplasticity in
their responses. Although little
evidence currently supports the existence ofdifferent
populations, it is likely that neutrophil
responses are more flexible and less stereotypedthan previously
thought.
Another major mechanism of tumor escape
from immune control has recently beenattributed to a
heterogeneous category of im-
mature myeloid cells, called myeloid-derivedsuppressor cells
(MDSCs) (189). In healthy
individuals, MDSCs are found in the bonemarrow, where they
differentiate into mature
neutrophils and monocytes. In cancer andsome autoimmune and
infectious diseases,
differentiation is partially blocked, leading toaccumulation of
these precursors, which act as
powerful suppressors of T cell functions. MD-
SCs have characteristics of neutrophils, and inmice, they are
typically detected using the neu-
trophil surface markers CD11b+ and Gr-1+,
although they consist of variable proportions
of monocytic and granulocytic cells (189). In
human renal cell carcinoma, MDSCs have
identical morphology and express the same sur-face markers as do
activated neutrophils (190,
191). MDSCs inhibit T cell proliferation bylimiting L-arginine
availability via arginase and
NOS activities, both of which use this aminoacid as a
substrate(189, 191, 192). Furthermore,
MDSCs are strong producers of ROS, whichsuppresses T cell
responses (119, 192). Inter-fering with the release of MDSCs or
using drug
interventions to polarize neutrophil responsesin the tumor
microenvironment could repre-
sent novel therapeutic strategies against cancer.
Autoimmunity
Deregulated neutrophil cell death and/orclearance often
accompanies autoimmune syn-
dromes (193195) and may play a major rolein disease
pathogenesis, given that release of
proteolytic and cytotoxic molecules from neu-
trophils can trigger organ damage. Neutrophilproducts act as
both targets and mediators of
autoimmunity. MPO and PR3 are the main tar-gets of
antineutrophil cytoplasmic antibodies
(ANCA), autoantibodies directed against anti-gens present in the
cytoplasm of neutrophils.
Wegeners granulomatosis is consistently as-sociated with the
presence of ANCA. Further-
more, the extent of organ damage in patientswith Wegeners
granulomatosis correlates with
the PMN infiltrate rather than with traditional
autoimmunity parameters such as T cell acti-vation or
autoantibody titers (196). Likewise,
ANCA bind MPO and PR3 expressed on thesurface of activated
neutrophils, promoting
degranulation and release of chemoattractantsand ROS, which
together lead to a vicious
cycle of tissue damage and inflammation.
Earlyreportsalsosuggestthat,inaninflammatoryen-
vironment, ANCA accelerate
ROS-dependentneutrophilapoptosis,suggestingafeed-forward
cycle culminating in organ damage (194, 195).
SLE is another chronic autoimmune diseaseaffecting multiple
tissues and organs. Autoan-
tibodies produced in SLE are predominantlyeither ANCA or
directed against chromatin.
Although neutrophils had long been suspected
www.annualreviews.org Neutrophil Functions 479
-
7/24/2019 Neutrophil Function From Mechanism to Disease
22/33
Vasculitis:inflammation of bloodvessels
to be causative agents, their role in SLE patho-
genesis remained elusive. The recent discoveryof a link between
SLE and NET formation
has helped to shed light on this quandary.It was proposed that
TNF- and IFN-prime cells for NET formation in response toanti-PR3,
antiribonucleoprotein, anti-HNP,
or anti-LL-37 autoantibodies (103, 104, 106).Thus, high levels
of inflammatory cytokines inautoimmune patients are believed to
sensitize
neutrophils to NETosis, whereas autoanti-bodies may trigger a
switch from apoptosis to
NETosis. Additional evidence suggesting arole for NETs in
autoimmune pathology was
obtained when NETs were identified in renaland/or skin biopsies
from patients with SLE
and small vessel vasculitis (103106). Severalstudies have
reported the presence of a particu-
lar subset of neutrophils in PBMC preparationsfrom pediatric and
adult SLE patients. These
low-density granulocytes display phenotypic
characteristics of immature neutrophils withnonsegmented nuclei
and higher expression
of MPO, NE, and defensin-3, and they maybe related to the MDSCs
discussed previously
(see section on Cancer, above) (197, 198).An increased capacity
to form NETs and a
heightened cytotoxicity toward endothelialcells could bestow
them with pathogenic
properties in lupus (105).Because NETs appear to be formed
during
autoimmune disease, their timely removal may
be an essential mechanism for maintainingtissue homeostasis.
Human serum contains the
nuclease DNase I, which degrades NETs invitro. Notably, a
familial form of SLE is linked
to a mutation in DNase I (199). Furthermore,in a cohort of SLE
patients, 36% exhibited
either elevated titers of autoantibodies directedagainst NET
components or inhibitors of
DNase I, both of which may protect NETsfrom degradation. Most
notably, impaired
NET degradation correlates with development
of lupus nephritis, one of the most severemanifestations of SLE
(102).
Can it be that NETs play a general rolein modulation of
autoimmune responses? We
know that NETs induce plasmacytoid DCs
to produce IFN-, a central cytokine in SL
pathogenesis (103, 104). However, it remainsbe determined if DCs
can present NET com
ponents or if they contribute to autoreactivecell activation. It
is also possible that NETs a
involved in other autoimmune diseases. Shou
this prove to be the case, understanding t
role of NETs may provide critical insights inthe role of
microbial infections as a trigger autoimmunity.
CONCLUDING REMARKS
Neutrophils are specialized phagocytes th
arose as an evolutionary adaptation in
vertbratestocoordinateandexecuteoneofthemo
fundamental physiological responses: inflammation. They are
endowed with antimicrob
mechanisms that make them the preeminemicrobe exterminators of
the immune system
In addition to this important role, PMNs al
network with many other immune cells anhelp regulate the
initiation of specific T an
B cell immunity. However, neutrophils do nalwaysactin ways
beneficialto thehost: Unco
trolled neutrophil responses can exacerbate aeven cause
autoimmune and inflammatory d
eases. Many challenges remain in understaning neutrophil
function: Is there specializatio
among PMNs? Are they more plastic than wsuspect? How do they
make decisions befo
deploying their armamentaria? How do th
kill microbes? How specific are their instrutions to other
cells? Answering these questio
will better define neutrophils role in defenand disease and will
provide a rational path f
pursuing new therapies. Moreover, neutrophcan potentially
provide insights into sever
unique aspects of basic cell biology. Their striingly short life
spans make them excellent mo
els for investigating cell death, whereas thereliance on ROS as
biochemical effectors m
reveal novel ways for relaying intracellul
signals. The uniquely lobulated neutropnucleus is a feat of
higher-order nucle
architecture that is just beginning to yieits secrets. In short,
exciting times await t
humble neutrophil.
480 Amulic et al.
-
7/24/2019 Neutrophil Function From Mechanism to Disease
23/33
DISCLOSURE STATEMENT
The authors are not aware of any affiliations, memberships,
funding, or financial holdings that
might be perceived as affecting the objectivity of this
review.
ACKNOWLEDGMENTS
We thank Diane Schad for assistance with graphic design and
Cornelia Heinz for administrative
help. G.H. is an Alexander von Humboldt Foundation Scholar,
andB.A. is supported by an EMBOLong-Term Fellowship.
LITERATURE CITED
1. Ehrlich P. 1880. Methodologische Beitrage zur Physiologie und
Pathologie der verschiedenen Formen
der Leukocyten.Z. Klin. Med.1:553602. Waller A. 1846.
Microscopic observation on the perforation of capillaries by the
corpuscles of blood and
the origin of mucus and pus globules. Lond. Edinburgh Philos.
Mag. J. Sci. 2:271803. Metchnikoff E. 1893. Lecon sur la pathologie
comparee de inflammation.Ann. Inst. Pasteur7:348574. Nathan C.
2006. Neutrophils and immunity: challenges and opportunities.Nat.
Rev. Immunol.6:173825. Borregaard N. 2010. Neutrophils, from marrow
to microbes.Immunity33:65770
6. Borregaard N, Cowland JB. 1997. Granules of the human
neutrophilic polymorphonuclear leukocyte.Blood89:350321
7. Finger EB, Puri KD, Alon R, Lawrence MB, von Andrian UH,
Springer TA. 1996. Adhesion through
L-selectin requires a threshold hydrodynamic
shear.Nature379:266698. Lawrence MB, Kansas GS, Kunkel EJ, Ley K.
1997. Threshold levels of fluid shear promote leukocyte
adhesion through selectins (CD62L,P,E).J. Cell Biol.136:717279.
Kansas GS. 1996. Selectins and their ligands: current concepts and
controversies.Blood88:325987
10. McEver RP, Cummings RD. 1997. Role of PSGL-1 binding to
selectins in leukocyte recruitment.
J. Clin. Investig.100:S9710311. Yago T, Shao B, Miner JJ, Yao L,
Klopocki AG, et al. 2010. E-selectin engages PSGL-1 and CD44
through a common signaling pathway to induce integrin
L2-mediated slow leukocyte rolling. Blood
116:4859412. Mueller H, Stadtmann A, Van Aken H, Hirsch E, Wang
D, et al. 2010. Tyrosine kinase Btk regulates
E-selectin-mediated integrin activation and neutrophil
recruitment by controlling phospholipase C
(PLC) 2 and PI3K pathways.Blood115:31182713. LeyK, Laudanna C,
Cybulsky MI,NoursharghS. 2007. Getting tothe site of
inflammation:the leukocyte
adhesion cascade updated.Nat. Rev. Immunol.7:6788914. Campbell
JJ, Hedrick J, Zlotnik A, Siani MA, Thompson DA, Butcher EC. 1998.
Chemokines and the
arrest of lymphocytes rolling under flow conditions.
Science279:3818415. Constantin G, Majeed M, Giagulli C, Piccio L,
Kim JY, et al. 2000. Chemokines trigger immediate2
integrin affinity and mobility changes: differential regulation
and roles in lymphocyte arrest under flow.
Immunity13:7596916. Liu S, Kiosses WB, Rose DM, Slepak M, Salgia
R, et al. 2002. A fragment of paxillin binds the 4
integrin cytoplasmic domain (tail) and selectively inhibits
4-mediated cell migration. J. Biol. Chem.
277:2088794
17. Goldfinger LE, Han J, Kiosses WB, Howe AK, Ginsberg MH.
2003. Spatial restriction of4 inte-grin phosphorylation regulates
lamellipodial stability and 41-dependent cell migration. J. Cell
Biol.
162:7314118. Xu J, Wang F, Van Keymeulen A, Herzmark P, Straight
A, et al. 2003. Divergent signals and cytoskeletal
assemblies regulate self-organizing polarity in
neutrophils.Cell114:2011419. Schenkel AR, Chew TW, Muller WA. 2004.
Platelet endothelial cell adhesion molecule deficiency or
blockade significantly reduces leukocyte emigrationin a majority
of mouse strains.J. Immunol. 173:6403
8
www.annualreviews.org Neutrophil Functions 481
-
7/24/2019 Neutrophil Function From Mechanism to Disease
24/33
20. Phillipson M, Heit B, Colarusso P, Liu L, Ballantyne CM,
Kubes P. 2006. Intraluminal crawling
neutrophils to emigration sites: a molecularly distinct process
from adhesion in the recruitment cascad
J. Exp. Med.203:256975
21. Wong CH, Heit B, Kubes P. 2010. Molecular regulators of
leucocyte chemotaxis during inflammatio
Cardiovasc. Res.86:18391
22. Zarbock A, Ley K. 2008. Mechanisms and consequences of
neutrophil interaction with the endotheliu
Am. J. Pathol.172:17
23. Selvatici R, Falzarano S, Mollica A, Spisani S. 2006. Signal
transduction pathways triggered by select
formylpeptide analogues in human neutrophils.Eur. J.
Pharmacol.534:11124. Chen Y, Yao Y, Sumi Y, Li A, To UK, et al.
2010. Purinergic signaling: a fundamental mechanism
neutrophil activation.Sci. Signal.3:ra45
25. Sabroe I, Dower SK, Whyte MK. 2005. The role of Toll-like
receptors in the regulation of neutrop
migration, activation, and apoptosis.Clin. Infect. Dis.41(Suppl.
7):S42126
26. Parker LC, Whyte MK, Dower SK, Sabroe I. 2005. The
expression and roles of Toll-like receptors
the biology of the human neutrophil.J. Leukoc. Biol.77:88692
27. Ley K. 2002. Integration of inflammatory signals by rolling
neutrophils.Immunol. Rev.186:818
28. Guthrie LA, McPhail LC, Henson PM, Johnston RB Jr. 1984.
Priming of neutrophils for enhanc
release of oxygen metabolites by bacterial lipopolysaccharide.
Evidence for increased activity of t
superoxide-producing enzyme.J. Exp. Med.160:165671
29. El-Benna J, Dang PM, Gougerot-Pocidalo MA. 2008. Priming of
the neutrophil NADPH oxida
activation: role of p47phox phosphorylation and NOX2
mobilization to the plasma membrane. SemImmunopathol.30:27989
30. Didsbury JR, Uhing RJ, Tomhave E, Gerard C, Gerard N,
Snyderman R. 1991. Receptor class dese