Embed Size (px)
Citation preview

(trieSpptfigst4ruhrtiponttnwSigraa
Sa
ip
Experimental Neurology 161, 571–584 (2000)doi:10.1006/exnr.1999.7315, available online at http://www.idealibrary.com on
Nerve Guides Seeded with Autologous Schwann Cells ImproveNerve Regeneration
Francisco J. Rodrıguez, Enrique Verdu, Dolores Ceballos, and Xavier NavarroDepartment of Cell Biology, Physiology and Immunology, Universitat Autonoma de Barcelona, E-08193 Bellaterra, Spain
Received June 4, 1999; accepted November 1, 1999
gi5egafioTjetdaTsHitBiritaalgtg
crtiusCasg
This study evaluates the ability of Schwann cellsSCs) transplantedintoanerveguidetoimproveregenera-ion and reinnervation after sciatic nerve resection andepair, leaving a 6-mm gap, in the mouse. SCs weresolated from predegenerated adult sciatic nerves andxpanded in culture using a chemically defined medium.yngeneic, isogeneic, and autologous SCs were sus-ended in Matrigel and seeded in resorbable, permeableoly(L-lactide-co-e-caprolactone) guides at 150,000 cells/ube. Guides containing SCs were compared to guideslled with Matrigel alone and with peroneal nerve auto-rafts. Functional reinnervation was assessed by noninva-ive methods to determine recovery of sweating, nocicep-ive, sensory, and motor functions in the hindpaw during
months postoperation. Morphological analysis of theegenerated nerves was performed at the end of follow-p. The group with an autograft achieved faster andigher levels of reinnervation and higher number ofegenerated myelinated fibers than groups repaired byubulization. The immunogenicity of transplanted SCsnfluenced the outcome of nerve regeneration. Trans-lants of autologous SCs resulted in slightly lower levelsf reinnervation than autografts, but higher recovery andumber of regenerated fibers reaching the distal nervehan transplants of isologous and syngeneic SCs, al-hough most of the differences were not statistically sig-ificant. Syngeneic SCs did not improve regenerationith respect to acellular guides. Prelabeled transplantedCs were found to survive into the guide 1–3 months after
mplantation, to a larger number when they were autolo-ous than syngeneic. Cellular prostheses composed of aesorbable guide seeded with autologous SCs appear asn alternative for repairing long gaps in injured nerves,pproaching the success of autografts. r 2000 Academic Press
Key Words: nerve regeneration; reinnervation;chwann cell; cellular prosthesis; tube repair; nerveutograft; sciatic nerve.
INTRODUCTION
After severe nerve injuries, loss of substance resultsn a significant gap between the nerve stumps that
revents repair by direct epineural suture. An autolo- 1571
ous graft from another nerve of lesser functionalmportance is usually employed to bridge the gap (36,4). The purpose of introducing a graft between the twonds of a transected nerve is to offer mechanicaluidance as well as an optimal environment for thedvancing axonal sprouts. The nerve graft may comerom the same individual (autograft), from anotherndividual of the same species (homograft or allograft),r from a different species (heterograft or xenograft).he best regeneration results are provided when in-
ured nerves are repaired with autografts (53). How-ver, autograft repair implies several problems such ashe need of a second surgical step, elimination of theonor nerve function, a limited supply of donor nerves,nd the mismatch between nerve and graft dimensions.ubulization is an alternative repair method that con-ists in suturing the nerve stumps into a tubular guide.owever, simple tubulization usually fails when bridg-
ng relatively long gaps of 6 mm in the mouse, 15 mm inhe rat, and longer in higher species (11, 22, 46, 48).esides providing directional guidance to the regenerat-
ng axons and preventing axonal escape into the sur-ounding tissue, nerve guides have the potential tontroduce into the local environment factors or cellshat may enhance regeneration, in attempts to developn artificial nerve graft. It has been reported thatddition of neurotrophic factors (28, 29 64) and extracel-ular matrix components (6, 37, 51) inside a tubularuide enhances regeneration of peripheral nerves, al-hough recovery was usually lower than with auto-rafts when applied to bridge long gaps.The Schwann cell (SC) and its basal lamina are
rucial components in the environment through whichegenerating axons grow to reach their peripheralargets. SCs of the injured nerve proliferate, helpnflammatory infiltrating cells to eliminate debris, andpregulate the synthesis of trophic and tropic factorsuch as NGF, CNTF, IGF, and laminin (10, 21, 44).onsidering the importance of the SCs in creating andequate environment for nerve regeneration, the con-truction of cellular prostheses consisting in a nerveuide seeded with isolated SCs has been attempted (5,
9, 23, 33, 35, 41). However, the number of axons that0014-4886/00 $35.00Copyright r 2000 by Academic Press
All rights of reproduction in any form reserved.

rtpIitwhSoorcgptahiaca
cpibtroo6wofz
S
aetesdcs0st0hD(
csiowaSaS
mtsmDL
mm(ammb(ntwD
E
an(mTSPat
lsiwnS3slfwB
572 RODRIGUEZ ET AL.
egenerate into the distal nerve and the degree ofarget reinnervation were in general lower with trans-lanted SCs than with nerve autografts (23, 33, 42).mmune compatibility between donor and host is anmportant factor that affects axonal regenerationhrough nerve grafts as well as with guides prefilledith transplanted SCs. Syngeneic (5, 19, 23, 33, 35),eterologous (23), xenogeneic (41), and autologous (42).Cs have been used in this experimental paradigm, butnly limited studies have directly compared the effectsf different types of SCs. Guenard and co-workers (23)eported that syngeneic adult SCs seeded in PAN/PVChannels enhanced nerve regeneration, while heterolo-ous SCs elicited a strong immune reaction that im-eded regeneration. They suggested that transplanta-ion of autologous SC would be desirable in order tovoid immunological rejection. Recently, Levi et al. (42)ave shown that autologous adult monkey SCs seeded
n acrylic guides enhance regeneration of peripheralxons with respect to guides filled with a laminin-ontaining gel alone but to a lesser degree than anutologous nerve graft.In the present study, we used a fast method for
ulture and expansion of adult SCs from small pieces ofredegenerated mouse nerves. We obtained syngeneic,sogeneic, and autologous SCs that were seeded inioresorbable guides in order to evaluate the effects ofransplanted SC immunogenicity on peripheral nerveegeneration. Nerve autograft and four different typesf prostheses were compared, by means of neurophysi-logical and histological techniques, for the repair of a-mm-long gap in the mouse sciatic nerve. The presentork was undertaken to try to increase the success ratef regeneration across a 6-mm gap, a limiting distanceor nerve regeneration in the mouse with simple tubuli-ation.
MATERIALS AND METHODS
chwann Cell Cultures
SC from adult mouse sciatic nerves were isolatedccording to a technique modified from that of Brockest al. (9). Adult outbred Swiss OF-1 mice were anesthe-ized (50 mg/kg pentobarbital, ip), the sciatic nerve wasxposed and cut at the midthigh, and the wound wasutured and disinfected with povidone–iodine. Five to 7ays later, the in vivo degenerating nerves were asepti-ally removed and stored in Hanks’ balanced saltolution (Sigma, St. Louis, MO) with Ca21 and Mg21 at°C. The epineurium and connective tissue weretripped off with fine forceps, and the nerves werereated with 0.25% trypsin, 0.1% collagenase A, and.1% DNase-I in 1 ml of Hanks’ solution at 37°C for 2–3. The enzymes were inactivated by addition of 10 ml ofulbecco’s minimum essential medium F-12 Ham
DMEM/F-12; Sigma) and the cell mixture recovered by a
entrifugation at 900 rpm for 7 min. The enzymaticolution was removed and the cells were resuspendedn culture medium. The cell suspension was seedednto 25-cm2 flasks or 24-well microplates precoatedith poly-L-lysine (10µg/ml) and incubated in 5% CO2
t 37°C. Culture medium was replaced every 2–3 days.ix nerves per flask were used for culturing syngeneicnd isogeneic adult SCs and one nerve for autologousCs at a density of 104 cells/cm2.The conditioned medium used is composed of a 1:1ixture of defined medium fresh and exposed for 48 h
o a culture of adult predegenerated nerve cells andupplemented with 1% fetal calf serum. The definededium was composed of a 3:1 mixture of MEM andMEM/F-12, supplemented with L-carnitine (2µg/ml),
-glutamine (2 mM), Hepes (15 mM), putrescine (8µg/l), taurine (4µg/ml), penicillin–streptomycin (50 UI/l–0.5µg/ml), gentamycin (40µg/ml), bovine albumin
1 mg/ml), ascorbic acid (1µg/ml), MEM nonessentialmino acid solution (1003) (1 ml/100 ml), MEM vita-in solution (1003) (1 ml/100 ml), GMS-S solution (1l/100 ml), lipoic acid (100 ng/ml), retinol (200 ng/ml),
iotin (20 ng/ml), linoleic acid (1µg/ml), linolenic acid1µg/ml), D,L-a-tocopherol (1µg/ml), progesterone (6,3g/ml), and vitamin B12 (1µg/ml). The effectiveness ofhe conditioned medium to promote SC proliferationas tested against the chemically defined medium andMEM/F-12 with 10% fetal calf serum (DF-10S).
valuation of Schwann Cell Cultures
The purity of SC cultures was evaluated at 3, 5, 7,nd 10 days in vitro. The total number of cells and theumber of SCs were counted in six random fields
3100, 1 mm2) on the basis of cell soma and nuclearorphology using phase contrast microscopy (9, 72).he SC purity was evaluated as the mean percentage ofCs with respect to the total number of cells counted.lating cell yield and proliferation were measured asn index of cell density per square millimeter and theotal number of cells.
SCs were also identified by immunocytochemicalabeling for S100 protein. Cells were washed with 0.1 Modium phosphate buffer (PBS, pH 7.4), fixed for 15 minn 4% paraformaldehyde, permeabilized for 30 minith 0.1 M PBS containing 0.3% Triton-X and 1%ormal goat serum, and incubated with rabbit anti-100 antibody (Incstar, Stillwater, MN) for 60 min at7°C. After washes in PBS, cells were incubated inecondary antisera, goat anti-rabbit cyanine 3.18-abeled IgG (1:200; Amersham, Arlington Heights, IL)or 30 min at room temperature. Following additionalashes, SC cultures were observed under an OlympusX-40 microscope equipped for epifluorescence using
ppropriate filters.
C
lo1s2cHstocgwatt2dCTaa3
ltfd2meer
S
eOwdsadlis6nwtaMM
fPiGsc
datTtlC
F
anoempeecnoatlai
bscsw(svswwdpppd(si
o
573SCHWANN CELL TRANSPLANTS AND NERVE REGENERATION
ellular Prostheses
Nerve guides were made of a bioresorbable poly(L-actide-e-caprolactone) copolymer (PLC) (60). The tubesffer a trabecular wall with high permeability, and have
mm i.d. and 150µm wall thickness (66). Prior toeeding, the guides were sterilized under UV light for4 h. Seven or 10 days after initial plating, cells fromonfluent adult SC cultures were rinsed twice with coldanks’ solution and detached by incubation with tryp-
in (0.05%; w/v)–EDTA (0.02%) for 2–3 min at roomemperature. Trypsin action was arrested by additionf DF-10S and the cell mixture was recovered byentrifugation. Cells were gently resuspended in Matri-el (Collaborative Research Inc., Bedford, MA) mixedith culture medium at a final concentration of 4 mg/mlnd the viability and number of cells were evaluated byrypan blue exclusion using a hemocytometer. Then,he volume was adjusted to obtain a cell density of5,000 cells/µl and 6 µl of the suspension were intro-uced with a micropipette into 8 mm long PLC tubes.ontrol tubes were filled with Matrigel (4 mg/ml) alone.he filled tubes were allowed to gelify at 37°C for 2–3 hnd then covered with a thin film of culture mediumnd left overnight in a humid ambiance with 5% CO2 at7°C.In some cases, SC were prelabeled with the cell
inker PKH26-GL (Sigma) in order to recognize theransplanted cells after implantation. SC were centri-uged and resuspended in 250µl of the commercialiluent C and then the suspension was mixed with50µl of diluent C containing 1µl of the dye and gentlyixed for 2 min. The reaction was stopped by adding an
qual volume of DF-10S and the excess of dye wasliminated by centrifugation. Finally, labeled SC wereesuspended and seeded into PLC guides as above.
urgical Procedures and Experimental Groups
Operations were performed under pentobarbital an-sthesia (60 mg/kg ip) on five groups of female SwissF1 mice, aged 2.5 months. First, the saphenous nerveas cut in the femoral space and a long segment of theistal stump removed to prevent regeneration. Theciatic nerve was exposed at the midthigh, transectedt a constant point, 45 mm from the tip of the thirdigit, and a segment of the distal stump resected. Theesion was repaired by fixing the nerve stumps 1 mmnside the ends of an 8 mm guide, by means of one 10-Outure stitch at each end, leaving an interstump gap ofmm, or with a 6-mm-long autograft of the peroneal
erve obtained from the resected segment, suturedith two 10-O stitches at each stump. The experimen-
al groups evaluated were Group PG (n 5 7), peronealutograft; Group M (n 5 6), PLC tube filled withatrigel; Group sSC (n 5 6), PLC tube filled with
atrigel seeded with 150,000 syngeneic SCs, obtained urom adult mice of the same strain; Group iSC (n 5 8),LC tube filled with Matrigel seeded with 150,000
sogeneic SCs, from adult mice of the same litter; androup aSC (n 5 8), PLC tube filled with Matrigel
eeded with 150,000 autologous SCs, derived from theontralateral nerve.Finally, the skin was closed with 5-O silk sutures and
isinfected with povidone–iodine solution. In order tovoid autotomy after denervation, animals were pre-reated with amitriptyline in the drinking water (57).he experimental procedures followed the recommenda-ions of the European Union for the care and use ofaboratory animals and were approved by the Ethicalommittee of our institution.
unctional Evaluation
Regeneration of large myelinated nerve fibers wasssessed by nerve conduction tests (59). The sciaticerve was stimulated by supramaximal square pulsesf 0.01 ms duration applied through a pair of needlelectrodes at the sciatic notch, and the compounduscle action potential (CMAP) was recorded from
lantar and gastrocnemius muscles with microneedlelectrodes, while for sensory nerve conduction, thelectrodes were inserted on the fourth toe to record theompound nerve action potential (CNAP) of the digitalerves. The evoked CMAP and CNAP were displayedn a storage oscilloscope (Tektronix 2221) at settingsppropriate to measure the amplitude from baseline tohe maximal negative peak and the latency from stimu-us to the onset of the first negative deflection. Thenimal temperature was controlled by a water circulat-ng heating pad.
Reinnervation by small nerve fibers was evaluatedy testing sympathetic sudomotor and nociceptive re-ponses. Sweating was stimulated by injection of pilo-arpine nitrate (5 mg/kg sc). Ten minutes later ailicone material (Elasticon, Kerr Co., Romulus, MI)as applied over the plantar surface of the hindpaw
59). As the material hardened, it retained the impres-ions made by the sweat droplets emerging from indi-idual sweat glands (SGs). The number of SG impres-ions was determined under a dissecting microscopeith transillumination. Recovery of pain sensitivityas tested by light pricking with a needle, under aissecting microscope, at five areas, from the mostroximal pawpad to the tip of the second digit on thelantar aspect of the denervated paw (59). A score toinprick (PP) was assigned from no response (0), re-uced or inconsistent responses (1), to normal reaction2) in each area tested, in comparison with the re-ponses to the same stimuli applied to the contralateralntact hindpaw.
Functional tests were performed before operation tobtain baseline control values and at several intervals
p to 105 days postoperation. For normalization, val-
uppfi(1rscmb
ratqtTa
I
lPniePmLssflacpodcs
M
p(tddEweqt2w
Mb5lsnfidcaawansm
S
cwbc
E
aacpmSv1cbwr
saatfagwp(c
574 RODRIGUEZ ET AL.
es obtained after operation were expressed as theercentage of preoperative values for each mouse andlotted against time. For all functions tested we identi-ed the day of the first response after denervationwhen there was no reinnervation an arbitrary value of20 days was assigned) and the maximal percentage ofeinnervation. A functional recovery index (FRI), repre-enting the area under the reinnervation curve, wasalculated by adding the rectangle areas obtained byultiplying the number of days of each testing interval
y the mean percentage of recovery of the interval (59).At the end of functional follow-up, animals were
eanesthetized, the operated hindlimb was dissected,nd the implanted tube or graft was inspected to verifyhe presence of a regenerated nerve and its grossuality. To assess regeneration across the repair site,he distal nerve was lightly pinched with fine forceps.hen a long segment including the implanted guidend nerve was removed.
mplant Evaluation
To study SC viability in the cellular prostheses, aive/dead–viability/cytotoxicity kit (L-3224; Molecularrobes, Eugene, OR) was used. Briefly, the regeneratederve cable was carefully pulled out from the tube and
mmersed in PBS containing calcein-AM (2µM) andthidium homodimer (4µM) for 90 min. After washes inBS, the cable was fixed in 4% paraformaldehyde for 30in and sectioned at 40µm with a sliding cryotome.ongitudinal sections were mounted on gelatin-coatedlides and observed under the epifluorescence micro-cope. The surviving transplanted SCs showed reduorescence due to the prelabeling with PKH26-GLnd green fluorescence due to retention of polyanionicalcein after intracellular esterase activity inside theirreserved membrane; dead transplanted SCs showednly red fluorescence since ethidium enters cells withamaged membranes (14). Finally, some regeneratedables were directly fixed in 4% paraformaldehyde andtained against S100 protein as described above.
orphometrical Evaluation
Regenerated nerves were fixed in glutaraldehyde–araformaldehyde (3–3%) in 0.1 M cacodylate bufferpH 7.4, 4 h, 4°C) and cut in two pieces to enableransverse sections at midtube or -graft and at theistal nerve. Samples were postfixed in OsO4 (2%, 2 h),ehydrated through ethanol series, and embedded inpon. Transverse semithin sections (0.5µm) of thehole nerve were stained with toluidine blue andxamined under light microscopy. Images were ac-uired with an Olympus DP10 camera to a PowerMacin-osh G3 computer and printed at final magnification of003 for measuring the cross-sectional area of the
hole nerve and 20003 for morphometrical analysis. sorphometry was performed from 7–10 fields choseny systematic sampling of squares containing at least00 myelinated fibers. If the whole nerve section hadess than 500 fibers, all regenerated fibers were mea-ured. The total number of myelinated fibers in theerve was estimated from the area occupied by thebers in the photographs. Images were redrawn on aigitizing tablet and processed through a Macintoshomputer using a morphometry software to obtain axonnd fiber perimeters (22). Areas and diameters of axonsnd fibers, myelin sheath thickness, and the g ratioere calculated. When there was no nerve regenerationt the cross-sectional level, zero values were entered forerve area and number of myelinated fibers, but theample was not used for morphometrical measure-ents.
tatistical Analysis
All data are expressed as means and SEM. Statisticalomparisons between groups were made by ANOVAith post hoc Scheffe test for multiple comparisonsetween groups. The differences were considered signifi-ant when P , 0.05.
RESULTS
valuation of Schwann Cell Cultures
Primary adult SCs have been successfully isolatednd expanded up to a number enough to seed anutologous cellular prosthesis from a small piece of theontralateral mouse sciatic nerve (10- to 15-mm-longiece). Primary cultures from adult prelesioned nervesaintained in conditioned medium showed a purity ofCs of about 80% at 5 days and 87% at 7 and 10 days initro. The SC density was about 50 at 5, 100 at 7, and88 SC/mm2 at 10 days in vitro. When adult SCs wereultured in defined medium, the SC purity and densityy 10 days was about 70% and 50 SC/mm2, respectively,hile in DF-10S medium these values were 25 and 30,
espectively (Table 1).In all primary cultures, SCs showed a spindle-like
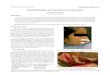
hape with birefringent soma and generally two largend thin processes (Fig. 1). Fibroblast-like cells showeddark soma with thick processes; in DF-10S medium
heir cytoplasm was spread and pleomorphic in shape,orming a monolayer on the substrate. Other cells withtypical macrophagic multivacuolated morphology be-
an to appear by 7 or 10 days. S100 immunostainingas seen only in spindle-like cells but not in theleomorphic and multivacuolated fried-egg-shaped cellsFig. 1). Counts of S100-positive cells were similar toounts of SCs based on morphological criteria in the
ame 10 days cultures.
F
pAcprmagpfriaaa
dmmaliwmiprd
t1awm
P
D
mm
575SCHWANN CELL TRANSPLANTS AND NERVE REGENERATION
unctional Reinnervation
Sudomotor nerve regeneration, judged by the reap-earance of secreting SGs over time, is shown in Fig. 2.fter denervation, the number of reactive SGs de-reased to 0 within 2 weeks. The first SGs reactive toilocarpine were observed from 30 days. The number ofeinnervated glands increased progressively to maxi-al values of about 85% in group PG, 80% in group
SC, 65% in group iSC, 48% in group M, and 43% inroup sSC (Table 2). The first withdrawal responses toinprick in proximal sites of the hindpaw were foundrom 30 days in group PG and by 60 days in theemaining groups. The total PP score progressivelyncreased as responses were elicited in more distalreas of the paw (Fig. 2), achieving a mean recovery ofbout 87% in groups PG and aSC, 82% in group iSC,nd 60% in groups sSC and M.The first low-amplitude CMAPs were recorded by 30
ays postoperation in plantar and gastrocnemiususcles of a few mice of group PG and 30 days later inice of the remaining groups (Fig. 3). The CMAP
mplitude increased progressively to reach mean finalevels of about 81% in group PG, 60% in group aSC, 42%n group iSC, 36% in group M, and 26% in group sSCith respect to control values for the gastrocnemiususcle and of 27% in group PG, 20% in group aSC, 13%
n groups M and iSC, and 6% in group sSC for thelantar muscle (Table 2). Similarly, digital CNAPs wereecorded by 30 days in a few mice of group PG; by 60ays in groups aSC, iSC, and M; and by 90 days
FIG. 1. Micrographs of Schwann cell cultures at 7 days in vitroaken under phase contrast from (A) a culture in medium containing0% serum (DF-10S), (B) a culture in chemically defined medium,nd (C) a culture in conditioned medium. D, Schwann cells stainedith S100 antiserum in conditioned medium culture. A–C, final
TABLE 1
Purity and Density of Schwann Cells on Primary Cultures ofAdult Sciatic Nerves Plated in Different Culture Media
Medium n 3 div 5 div 7 div 10 div
urity (% cells)DF-10S 3 43 6 4 43 6 1 31 6 1 25 6 0.3DM 9 81 6 8 82 6 1* 77 6 2* 73 6 2*CM 3 70 6 6* 80 6 2* 88 6 1*,** 87 6 3*,**
ensity (n · SC/mm2)DF-10S 3 15 6 5 24 6 8 30 6 3 31 6 7DM 9 27 6 5 34 6 4 39 6 3 49 6 6CM 3 13 6 3 47 6 4 97 6 4*,** 188 6 3*,**
Note. DF-10S, serum-containing medium; DM, chemically definededium; CM, Schwann-cell-conditioned medium; n, number of pri-ary cultures tested; div, days in vitro. Values are means 6 SEM.* P , 0.05 vs. DF-10S; ** P , 0.05 vs. DM.
agnification, 320; D, 340.

pi21pwd
cap
ts
M
i(itfirpngt
Staggcldawflaa
tcmatatantsstiataasnstmtg
PrMa
576 RODRIGUEZ ET AL.
ostoperation in group sSC. The CNAP amplitudencreased throughout the study to reach values above0% in groups PG, aSC, and iSC; 18% in group M; and1% in group sSC. The latency of the evoked actionotentials decreased over time to reach values thatere slightly longer than controls, but not significantlyifferent between experimental groups (Fig. 3).The FRI, indicative of the overall reinnervation
ourse, was highest in group PG followed by groupsSC and iSC and finally groups sSC and M. The
FIG. 2. Percentage of (A) the number of reactive SGs and (B) theP score over time in groups of mice with sciatic nerve resection andepair with a peroneal autograft (PG) or PLC guides containingatrigel alone (M) or seeded with syngeneic (sSC), isologous (iSC),
nd autologous (aSC) Schwann cells.
ercentage of the FRI with respect to that achieved by (
he nerve graft group was about 42% in groups M andSC, 58% in group iSC, and 68% in group aSC (Table 2).
orphological Regeneration and SC Viability
During microscopic dissection we found a regenerat-ng nerve cable inside the PLC guide in four of six mice66%) of groups M and sSC, six of eight (75%) of groupSC, and seven of eight (87%) of group aSC (Table 3). Inhese cases the tube was covered by a vascularized thinbrous tissue. Pinching the distal nerves evoked aeflex response in the anesthetized animals, whileinching and cutting the fibrous cover of the tube didot, indicating that axonal regeneration had pro-ressed inside the guide. In group PG regenerationhrough the graft occurred in all the mice.
After 30 and 90 days transplanted PKH26-labeledCs were visualized in the regenerated nerve cables byhe red fluorescence of the label in their cytoplasm. Inddition, by 30 days postimplantation in nerves ofroup aSC all red fluorescent cells also showed intensereen calcein fluorescence, while fewer double-stainedells were observed in groups iSC and sSC (Fig. 4). Theower density of labeled cells found in the latter groups,espite that the same amount was implanted initially,lso suggests a certain degree of cell death. As thereere only few evident green-labeled cells without reduorescence, migration of host cells into the graftppeared to be limited during the short time (30 days)ssayed.Under light microscopy, the nerve cables regenerated
hrough guides were round in shape and located at theenter of the tube lumen. Sections taken at the tubeidpoint demonstrated typical regenerates, includingthin newly formed perineurium surrounding a cen-
ral cable composed of small nerve fascicles of myelin-ted and unmyelinated axons with neovascularizationhroughout. The myelinated nerve fibers were of vari-ble size, with thinner myelin sheaths than in controlerves (Fig. 5). In some of the samples of all groupshere was a mild infiltrate of monocytic cells within andurrounding the regenerated cable. At the distal nervetump, there was a lower density of regenerated fibershan at midtube, particularly in groups M, sSC, andSC, while in group aSC both values were similar. Withn autologous peroneal graft there was abundant scarissue at the suture lines. At the midgraft the regener-ted nerve showed numerous myelinated and unmyelin-ted axons within the endoneurium, but also somemall size fascicles in the epineurium. In the distalerve, a lower number (57%) of regenerated axons oflightly smaller caliber were observed with respect tohe midgraft sections. The number of regeneratedyelinated fibers was significantly higher in group PG
han in groups M and sSC at both levels and than inroup iSC at midtube, but not from values of group aSC
Table 3).
w(cendwahwi
etPalrist
ianleM
toaaebiiraeRtptrr
Et
nR
O
R
F
(b
577SCHWANN CELL TRANSPLANTS AND NERVE REGENERATION
Morphometrical analysis was performed only in miceith a regenerated nerve through the guide or graft
Table 3). The areas of the intratubular regeneratedable ranged between 0.07 and 0.22 mm2 in the differ-nt animals, being in most cases smaller than theormal mouse sciatic nerve (0.22 mm2). The averageiameter of myelinated axons and myelin thicknessere slightly larger in the PG than in the other groups,lthough differences did not attain significance. Theistogram distribution of fiber and axon diametersere skewed to the left with respect to control nerves, more
n groups M and sSC than in groups iSC, aSC, and PG.
DISCUSSION
The results of this study show that SCs, isolated andxpanded from small pieces of predegenerated nerves,ransplanted within a bioresorbable and permeableLC guide enhance peripheral nerve regenerationcross an interstump gap of 6 mm in the mouse, aength that normally limits regeneration after tubeepair (11, 22). In addition, we demonstrate that SCmmunogenicity affects peripheral regeneration withinuch cellular prostheses. The levels of distal reinnerva-
TAB
Reinnervation Success Rate, Onset Day, and Maximal Degach Function Tested and the Average during 105 Days of Fol
he Five Groups of Mice Studied
Group: PG M
: 7 6einnervation: 7 (100%) 4 (66%)
nset daySG 30 6 0 65 6 18PP 43 6 13 85 6 12a
G-CMAP 38 6 8 80 6 15a,b
P-CMAP 39 6 8 85 6 12a
CNAP 60 6 11 100 6 10a
Mean 43 6 8 84 6 13a
ecovery (%)SG 84 6 10 48 6 17PP 86 6 14 58 6 20G-CMAP 81 6 8 36 6 14a
P-CMAP 27 6 6 13 6 7CNAP 21 6 5 18 6 9Mean 60 6 8 35 6 13
RISG 5557 6 877 2322 6 948a,b
PP 6667 6 1120 2887 6 1162a
G-CMAP 3954 6 675 1539 6 654a
P-CMAP 1173 6 265 453 6 257a
CNAP 1083 6 318 655 6 428Mean 3698 6 567 1571 6 666a
Note. Values are means 6 SEM. Groups were repaired with an autoM) or containing syngeneic Schwann cells (sSC), isologous Schwannvs group aSC; c vs group iSC.
ion and the number of myelinated fibers regenerated A
nto the distal stump were similar with transplantedutologous SCs to those attained with an autologouserve graft, while transplanting isologous SCs yielded
ower results and syngeneic SCs did not improve regen-ration with respect to the control guides filled withatrigel alone.Successful regeneration in tube repair depends on
he formation of a new extracellular matrix scaffold,ver which blood vessels, fibroblasts, and SCs migratend form a new nerve structure (44, 70). Survivingxons in the proximal stump develop growth cones andxtend into the tube along the connective strandsridging the gap, and eventually axons and accompany-ng SCs progress into the distal nerve stump. Thismplies a delay in axonal elongation and failure ofegeneration if the nerve stumps are not able to provide
proper cable with enough regenerative promotinglements inside the tube, as often occurs in long gaps.egeneration across synthetic guides is dependent on
he presence as a distal insert of a nerve stump thatrovides both cellular and humoral factors for regenera-ion (38, 45, 71). If the distal nerve stump was excluded,egeneration through tubular guides was significantlyeduced or even absent (46, 47, 71). Madison and
2
e of Reinnervation and Functional Recovery Index (FRI) for-Up after Resection and Tube Repair Leaving a 6-mm Gap in
sSC iSC aSC
6 8 84 (66%) 6 (75%) 7 (87%)
80 6 18a,c 30 6 1 41 6 1175 6 15 67 6 7a 67 6 7a
65 6 18 60 6 0a,b 41 6 1180 6 13a 86 6 9a 67 6 7a
109 6 7a 92 6 8a 75 6 1082 6 14a 69 6 5a 63 6 8a
43 6 19a 65 6 9 80 6 1163 6 20 82 6 13 87 6 1226 6 13a 42 6 13 60 6 14
6 6 3a 13 6 6 19 6 411 6 9 23 6 7 23 6 634 6 15 45 6 8 54 6 8
2130 6 982a,b 3695 6 536a 4814 6 6993166 6 1022a,b 3937 6 697a 4987 6 758a
996 6 555a 1502 6 551a 2421 6 662203 6 106a 449 6 224a 539 6 144a
267 6 246 678 6 278 1019 6 2841585 6 687a 2140 6 424a 2530 6 421a
us peroneal graft (PG) or with a PLC guide filled with Matrigel alones (iSC), and autologous Schwann cells (aSC). P , 0.05 a vs group PG;
LE
relow
logocell
rchibald (50) have shown that rat SCs are not able to

tc
G
r
578 RODRIGUEZ ET AL.
FIG. 3. Percentage of the amplitude of (A) the plantar CMAP, (B)o the plantar CMAP over time in groups of mice with sciatic nervontaining Matrigel alone (M) or seeded with syngeneic (sSC), isologo
TAB
Morphometrical Parameters of the Regenerated Nin the Five Grou
roup n
No. myelinated fibers
nMidgraft Distal
PG 7 3884 6 767 2233 6 230 4M 6 1370 6 491* 515 6 217* 4sSC 6 981 6 431* 622 6 299* 4iSC 8 1456 6 384* 1039 6 252 4aSC 8 1768 6 333 1901 6 319 4
Note. Values are means 6 SEM. All animals were included for num
the gastrocnemius CMAP, and (D) the digital CNAP and (C) the latencye resection and repair with a peroneal autograft (PG) or PLC guidesus (iSC), and autologous (aSC) Schwann cells.
LE 3
erve at Mid- and Distal Levels of the Graft or Tubeps of Mice Studied
Axonal diameter (µm) Myelin thickness (µm)
Midgraft Distal Midgraft Distal
2.75 6 0.15 2.57 6 0.14 1.04 6 0.08 1.03 6 0.062.32 6 0.16 2.18 6 0.11 0.84 6 0.03 0.86 6 0.032.45 6 0.10 2.29 6 0.09 0.85 6 0.06 0.86 6 0.052.33 6 0.19 2.35 6 0.12 0.85 6 0.05 0.90 6 0.042.62 6 0.06 2.35 6 0.10 0.86 6 0.04 0.89 6 0.05
ber of myelinated fibers, while only a sample of animals with successfulegeneration in the graft or tube was included for axonal diameter and myelin thickness calculations. Group definition as in Table 2.
* P , 0.05 vs group PG.
bagicataoowr
cpeeadtapSssriia
repanaastirbnirti
o(dPiw
579SCHWANN CELL TRANSPLANTS AND NERVE REGENERATION
S
ridge a 10-mm gap if the distal nerve insert iscellular. Therefore, regeneration fails through longaps, such as 6 mm in the mouse (11, 22, 58) or 15 mmn the rat (46, 47), most likely because the regenerativeapabilities of the nerve stumps have been exceedednd fibroblasts and SCs are not able to migrate throughhe gap and provide a permissive environment forxonal elongation. In these situations, the introductionf a neuritotropic matrix and a transplanted populationf activated SCs into the guide prior to implantationould mimic the initial cable formation and provide
egenerating axons with a proper support.A number of developments have made it possible to
onsider using an artificial nerve graft or cellularrosthesis consisting in a nerve guide seeded with SCsxpanded in culture to repair gaps within the periph-ral nervous system as an alternative to the classicalutologous nerve grafts, while avoiding the secondaryeficit resulting from obtaining graft donor nerves andhe limitations in the amount of donor material avail-ble for grafting. Advances toward applying cellularrostheses should, at least, include the ability to isolateCs from small segments of peripheral nerve within ahort time period, the proof that transplanted SCsurvive in the host and effectively promote axonalegeneration and functional recovery, and the availabil-ty of tubulization materials with adequate propertiesn that they are biocompatible, bioresorbable, perme-ble, and flexible.The practical use of SC transplants for nerve repair
equires conclusive information on the success of regen-ration and effective distal reinnervation. The neuro-hysiological tests used in this study are designed tossess reinnervation by different types of peripheralerve fibers, allowing serial evaluation in the samenimal at desired intervals (11, 58, 59). The reappear-nce of CMAP and CNAP evoked by nerve electricaltimulation, SG secretion under cholinergic stimula-ion, and response to pricking the skin with time arendicative of the rate of axonal regeneration and targeteinnervation. The degree of reinnervation, expressedy the amplitude of compound action potentials, theumber of reactive SGs, and the PP score, is an
ndication of the number of axons that are able toegenerate and reconnect with adequate targets andherefore of the functional recovery. Recovery of sweat-ng and nociception achieved in all animals higher
FIG. 4. Micrographs showing longitudinal sections (40µm thick)f the regenerated nerve cable inside guides seeded with autologousA, B, E) and syngeneic (C, D) Schwann cells at 30 days (A–D) and 90ays (E) postimplantation. A and C, red fluorescence produced byKH26-GL-prelabeled Schwann cells. B and D, green fluorescence
nduced by calcein indicating viable cells. Arrowheads point to cellsith PKH26-GL but not calcein label. E, Schwann cells stained with
100 antiserum. Bar: A–D, 100µm; E, 200µm.
pp6da
ftpt
aam
580 RODRIGUEZ ET AL.
ercentages than recovery of muscle and sensory actionotentials, as also shown in previous studies (11, 58, 59,6). The nerve conduction velocity is an index of theegree of myelination and maturation of the regener-
FIG. 5. Representative transverse sections (0.5µm) of regeneratefter repair with a peroneal autograft (A, B) and a PLC guide seedednd distal sections are from the same mouse in each group. Note thyelin thickness. Bar, 6 µm in all the micrographs.
ted fibers, and it usually remains lower than normal a
or a long time after severe nerve injuries. The quantita-ive measurements of distal target reinnervation at theaw are more discriminative between experimentalreatments than measures of proximal targets or than
rves at the midgraft (A, C, E) and the distal nerve (B, D, F) 110 daysh autologous (C, D) and with syngeneic (E, F) Schwann cells. Medialer grouping in small fascicles and the changes in fiber density and
d newite fib
nalysis of the walking track (56, 65, 67). On the other

haitogafeartfsnl
N
totgt3atogormtgfemTmdAbetsgbn
Sibcfiie
pfsecAiirp
S
bagSsnfcsrmtSctnmfifcdpctc
mmiawasweattpgs
581SCHWANN CELL TRANSPLANTS AND NERVE REGENERATION
and, morphometrical evaluation of the regeneratedxons at the site of repair, although providing valuablenformation on the quality and quantity of regenera-ion, may give an unreliable prediction of functionalutcome, as the number of axons counted in grafts oruides may be higher than the number of neuronsctually regenerating due to sprouting and neuromaormation (30, 66), and the limitations of axons tolongate in the distal nerve for long distances toppropriate targets (49). The number of axons thategenerate into the distal nerve and reach denervatedissues is more relevant as it can be related to theunctional reinnervation of targets (33). Our resultshow that the degree of target reinnervation and theumber of regenerated fibers in the distal nerve fol-
owed a similar gradation between the studied groups.
erve Guide and Medium for Cellular Prostheses
Although apparently of secondary importance, theype of nerve guide plays a key role in the constructionf cellular prostheses. Several physical parameters ofhe guides used, such as inner diameter, surface micro-eometry, thickness, permeability and composition, andhe gap length, affect nerve regeneration (1, 2, 11, 22,1, 46, 58). The PLC guides allow regeneration throughlonger gap and of a higher number of axons than other
ubes made of synthetic (silicone, Teflon, polysulfone)r biological materials (collagen) (15, 58, 66). The PLCuides implanted are well tolerated by the host, elicitnly a mild foreign body reaction, and have a lowesorption rate for over 6 months (60), maintainingechanical stability for longer time than required for
he axons to cross the gap. The high permeability of theuides further enhances nerve regeneration and allowsor an exchange of molecules and nutrients between thextraneural ambiance and the intratubular space thatay contribute to the survival of transplanted cells.
ransplanted SCs survived inside the guide even 3onths after implantation. Semipermeable guides of
ifferent materials, such as PAN/PVC (23, 41, 42),quavene biopolymer (33), or collagen (5, 35), haveeen used previously in such a model, but recently Levit al. (42) pointed out the need to improve flexibility ofhe guides to overcome failures due to dislodgment andecondary lesions to the repaired nerve. The PLCuides soften as biodegradation takes place in vivo andecome completely resorbed from the body, avoiding theeed to retrieve them at a later time.The ideal medium in which to suspend transplanted
Cs within the guide has not yet been defined. Directnjection of a cell suspension in culture medium haseen used (5, 19), but in order to avoid the transplantedells to escape from the lumen, the guide should betted in dimension to the repaired nerve or the open-
ngs occluded; both alternatives may have deleterious
ffects on the regenerating axons because of late com- gression or neuroma formation at the tube–guide inter-ace (17, 38). Laminin-containing gels seem the mostuitable matrix to seed SCs (23, 41, 42), based onvidence that laminin is a good substrate for cultureell adhesion and exerts neuritotropic effects (39).lthough a concentrated Matrigel matrix was shown to
mpair axonal regeneration probably due to an obstruct-ng effect (69), diluted Matrigel (4 mg/ml) improvedegeneration through prefilled tubes in long gaps anderformed better than collagen gels (37).
chwann Cell Immunogenicity and NerveRegeneration
Acellular nerve grafts allow for axonal regeneration,ut to a lesser degree and extension than cellularutografts or isografts (13, 24, 26). Regenerating axonsrow through the acellular graft only in the presence ofCs that proliferate and migrate from the proximaltump (27). The limited axonal growth in acellularerve grafts is due to the absence of SCs that provide a
avorable environment for sustained regeneration inellular autografts and through the degenerated distaltump. Similarly, within the central nervous systemegeneration of retinal ganglion cell axons, that nor-ally fails in the damaged optic nerve, is possible
hrough a peripheral nerve graft, but only when viableCs are present in the graft (7, 16). Even a lowerompliment of SCs than the normal amount present inhe injured nerve, provided by introducing a smallerve segment in silicone guides, is sufficient to pro-ote regeneration across longer gaps than in saline-
lled guides (20, 32, 65). This beneficial effect wasound only with a cellular autologous transplant, espe-ially if predegenerated, while acellular transplantsid not improve regeneration and heterologous trans-lants impaired regeneration (65), indicating that theellular component and the immunological fate of nerveransplants are relevant factors to be taken into ac-ount.The targets of rejection in peripheral nerves areainly the cells and the myelin sheaths, because theirembranes carry the antigens of the major histocompat-
bility complex (18). After transplantation of a nervellograft in the mouse, donor SCs were rejected in a feweeks and host-derived SCs invaded the graft associ-ted with regenerating axons. Under immunosuppres-ion donor SCs persisted in the allograft, associatedith and myelinating regenerated axons (3, 52). How-ver, despite the use of immunosuppressive treatment,chronic rejection process ensued in some cases, and
he number of myelinated fibers regenerated throughhe allograft was even lower than without immunosup-ression (52). By using acellular basal lamina allo-rafts that exhibit reduced immunogenicity with re-pect to cellular allografts, Gulati (24) showed that
rafts repopulated by coculture with isogeneic SCs
sradfi
bartda(thwncrwcSapAn1MtcKawwvW(thipmr(i
P
bnfirad
tp(ads(aerleSbicLnpdGwapas
ttdaiqaTcrgoapt
aPGDf
582 RODRIGUEZ ET AL.
urvived and supported axonal regeneration compa-able to cellular isografts in the rat. In contrast,cellular allografts cocultured with allogeneic SCs un-erwent immunological rejection and were unsuccess-ul, indicating that cultured SCs continue to exhibitmmunogenicity.
Our results demonstrate that immune compatibilityetween donor and host is an important factor thatffects the success of cellular prostheses to promoteegeneration and reinnervation. All the SCs used inhis report were allogeneic (from the same species), buterived from adult mice of the same (isogeneic) ornother litter (syngeneic) or from the same individualautologous). Previous studies (5, 23, 35, 41) showedhat syngeneic SCs transplanted in nerve guides en-ance axonal regeneration with respect to control guidesith saline solution or extracellular matrix compo-ents. However, our results show that a better out-ome, in terms of functional recovery, morphologicalegeneration, and proportion of success, was obtainedith autologous than with isogeneic and syngeneic
ells. It is worthy of comment that, although syngeneicCs have been shown to survive after implantation atn unknown proportion (35), a number of the trans-lanted cells die because of mild host rejection (33, 42).concentration-dependent effect was reported by Gue-
ard et al. (23), who found that guides seeded with 4 305 syngeneic SCs did not improve regeneration overatrigel-filled guides bridging a short, 8-mm gap, in
he rat while myelinated axon counts increased signifi-antly when the seeding density was doubled or tripled.im et al. (35) found similar values of CNAP amplitudend number of regenerated fibers between guides seededith 4 3 105 syngeneic SCs and sural autografts,hereas Keeley et al. (33) reported significantly loweralues in guides with 1 3 104 SCs than in autografts.hen repairing a long gap, above the limiting length
15 mm in the rat), regeneration was only improved byransplanting a large number of SCs (5). Thus, a veryigh number of syngeneic SCs should be transplanted
nitially to ensure that a number sufficient enough toromote regeneration remain viable during the firstonth after implantation. On the other hand, our
esults show that a lower amount of autologous SCs1.5 3 105) provide better results than syngeneic orsogeneic cells.
redegenerated Nerve as a Source of TransplantedSchwann Cells
This report also shows that adult autologous SCs cane expanded from a small piece of predegeneratederve (one mouse sciatic nerve) without mitogens. Thisnding opens the possibility of using nerve fragmentsesected from injured human nerves for designingutologous cellular prostheses to be implanted at a
elayed intervention. Several studies have reportedhat predegenerated nerve grafts (13, 25, 34) andredegenerated nerve transplants in synthetic tubes65) shorten the delay of axonal elongation and increasexonal regeneration. Contrarily, regeneration fails or iselayed when growing axons face a normal nerveegment or if distal Wallerian degeneration is slowed61, 68). Within the degenerated nerve, SCs deprived ofxonal contact proliferate, help inflammatory cells toliminate debris, and upregulate the synthesis andelease of a variety of neurotrophic factors and basalamina components (10, 21, 44). Thus, in vivo predegen-rated nerves are an excellent source of adult mitoticC, easily expandable in vitro (4, 8). Adult SCs can alsoe obtained from intact nerves if allowed to degeneraten vitro and incubated 2–3 days prior to dissociation inulture medium with mitogens (12, 43, 55). Recently,evi et al. (42) observed that after 14 days in vitro theumber of autologous SCs increased by 10-fold with aurity of 99% when SCs were plated in a chemicallyefined medium containing heregulin, forskolin, andas6. In the present study, using a conditioned mediumithout SC mitogens, we observed a similar patternfter 7 to 10 days. Conditioned media increase theroliferation of SCs (62, 63) that are able to synthesizend secrete several factors that promote its own adhe-ion, migration, and proliferation.The demonstration that human SCs isolated in cul-
ure retain functional capabilities after transplanta-ion in the peripheral nervous system of immune-eficient rodents (40, 41), as they survive, enhancexonal regrowth, and myelinate regenerated axons, ismportant evidence for clinical expectations. Fre-uently, during clinical nerve repair, small nerve piecesre resected from the nerve stumps prior to repair.hese nerve fragments might be a source of SCs toonstruct autologous cellular prostheses. The toolsequired for developing an autotransplantation pro-ram for the treatment of severe nerve injuries—btaining a small segment of injured nerves, isolatingnd expanding its SCs, and constructing a cellularrosthesis to repair the gap in the injured nerve—arehus already available (42).
ACKNOWLEDGMENTS
This work was supported by grants from the CYCIT (SAF97–0247)nd the Societat Catalana de Neurologia-Fundacio Uriach, Spain.oly(L-lactide-e-caprolactone) guides were kindly provided by Dr.abriele Perego, Novamont SpA, Novara, Italy. We are grateful tors. Manuel Nieto-Sampedro and Graciela Gudino-Cabrera for help-
ul comments and advice.
REFERENCES
1. Aebischer, P., V. Guenard, and S. Brace. 1989. Peripheral nerveregeneration through blind-ended semipermeable guidancechannels: Effect of the molecular weight cutoff. J. Neurosci. 9:
3590–3595.
1
1
1
1
1
1
1
1
1
1
2
2
2
2
2
2
2
2
2
2
3
3
3
3
3
3
3
3
3
583SCHWANN CELL TRANSPLANTS AND NERVE REGENERATION
2. Aebischer, P., V. Guenard, and R. F. Valentini. 1990. Themorphology of regenerating peripheral nerves is modulated bythe surface microgeometry of polymeric guidance channels.Brain Res. 531: 211–218.
3. Aguayo, A. J., M. Attiwell, J. Trecarten, S. Perkins, and G. M.Bray. 1977. Abnormal myelination in transplanted Tremblermouse Schwann cells. Nature 265: 73–75.
4. Ansselin, A. D., S. D. Corbeil, and D. F. Davey. 1995. Culture ofSchwann cells from adult animals. In Vitro Cell Dev. Biol. 31:253–254.
5. Ansselin, A. D., T. Fink, and D. F. Davey. 1997. Peripheral nerveregeneration through nerve guides seeded with adult Schwanncells. Neuropathol. Appl. Neurobiol. 23: 387–398.
6. Bailey, S. B., M. E. Eichler, A. Villadiego, and K. M. Rich. 1993.The influence of fibronectin and laminin during Schwann cellmigration and peripheral nerve regeneration through siliconchambers. J. Neurocytol. 22: 176–184.
7. Berry, M., L. Rees, S. M. Hall, P. Yiu, and J. Sievers. 1988. Opticaxons regenerate into sciatic nerve isografts only in the pres-ence of Schwann cells. Brain Res. Bull. 20: 223–231.
8. Bolin, L. M., T. P. Iismaa, and E. M. Shooter. 1992. Isolation ofactivated adult Schwann cells and a spontaneusly immortalSchwann cell clone. J. Neurosci. Res. 33: 231–238.
9. Brockes, J. P., K. L. Fields, and M. C. Raff. 1979. Studies oncultured rat Schwann cells. I. Establishment of purified popula-tions from cultures of peripheral nerve. Brain Res. 165: 105–118.
0. Bunge, R P. 1993. Expanding roles for the Schwann cell:Ensheathment, myelination, trophism and regeneration. Curr.Opin. Neurobiol. 3: 805–809.
1. Butı, M., E. Verdu, R. O. Labrador, J. J. Vilches, J. Fores, and X.Navarro. 1996. Influence of physical parameters of nerve cham-bers on peripheral nerve regeneration and reinnervation. Exp.Neurol. 137: 26–33.
2. Casella, G. T. B., R. P. Bunge, and P. M. Wood. 1996. Improvedmethod for harvesting human Schwann cells from matureperipheral nerve and expansion in vitro. Glia 17: 327–338.
3. Danielsen, N., J. M. Kerns, B. Holmquist, Q. Zhao, G. Lundborg,and M. Kanje. 1995. Predegeneration enhances regenerationinto acellular nerve grafts. Brain Res. 681: 105–108.
4. Decherchi, P., P. Cochard, and P. Gauthier. 1997. Dual stainingassessment of Schwann cell viability within whole peripheralnerves using calcein-AM and ethidium homodimer. J. Neurosci.Methods 71: 205–213.
5. Den Dunnen, W. F. A., B. van der Lei, J. M. Schakenraad, E. H.Blaauw, I. Stokroos, A. J. Pennings, and P. H. Robinson. 1993.Long term evaluation of nerve regeneration in a biodegradablenerve guide. J. Microsurg. 14: 508–515.
6. Dezawa, M., K. Kawana, and E. Adachi-Usami. 1997. The role ofSchwann cells during retinal ganglion cell regeneration inducedby peripheral nerve transplantation. Invest. Ophtalmol. Vis.Sci. 38: 1401–1410.
7. Ducker, T. B., and G. J. Hayes. 1968. Experimental improve-ments in the use of Silastic cuff for peripheral nerve repair. J.Neurosurg. 28: 582–587.
8. Evans, P. J., R. Midha, and S. E. Mackinnon. 1994. Theperipheral nerve allograft: A comprehensive review of regenera-tion and neuroimmunology. Progr. Neurobiol. 43: 187–233.
9. Feltri, M. L., S. S. Herer, L. Wrabertz, J. Kamholz, and M. E.Shy. 1992. Mitogen-expanded Schwann cells retain the capacityto myelinate regenerating axons after transplantation into ratsciatic nerve. Proc. Natl. Acad. Sci. USA 89: 8827–8831.
0. Francel, P. C., T. J. Francel, S. E. Mackinnon, and C. Hertl.
1997. Enhancing nerve regeneration across a silicone tubeconduit by using interposed short-segment nerve grafts. J.Neurosurg. 87: 887–892.
1. Fu, S. Y., and T. Gordon. 1997. The cellular and molecular basisof peripheral nerve regeneration. Mol. Neurobiol. 14: 67–116.
2. Gomez, N., J. Cuadras, M. Butı, and X. Navarro. 1996. Histo-logic assessment of sciatic nerve regeneration following resec-tion and graft or tube repair in the mouse. Restor. Neurol.Neurosci. 10: 187–196.
3. Guenard, V., N. Kleitman, T. K. Morrissey, R. P. Bunge, and P.Aebisher. 1992. Syngeneic Schwann cells derived from adultnerves seeded in semipermeable guidance channels enhanceperipheral nerve regeneration. J. Neurosci. 12: 3310–3320.
4. Gulati, A. K. 1995. Immunological fate of Schwann cell-populated acellular basal lamina nerve allografts. Transplanta-tion 59: 1618–1622.
5. Gulati, A. K. 1996. Peripheral nerve regeneration throughshort- and long-term degenerated nerve transplants. Brain Res.742: 265–270.
6. Hall, S. M. 1986. Regeneration in cellular and acellular auto-grafts in the peripheral nervous system. Neuropathol. Appl.Neurobiol. 12: 27–46.
7. Hall, S. M. 1986. The effect of inhibiting Schwann cell mitosis onthe re-innervation of acellular autografts in the peripheralnervous system of the mouse. Neuropathol. Appl. Neurobiol. 12:401–414.
8. Harman, K., J. Katnick, R. Lim, A. Zaheer, and J. C. de la Torre.1991. Glia maturation factor b stimulates axon regeneration intransected sciatic nerve. Brain Res. 564: 332–335.
9. Hollowell, J. P., A. Villadiego, and K. M. Rich. 1990. Sciaticnerve regeneration across gaps within silicone chambers: Long-term effects of NGF and consideration of axonal branching. Exp.Neurol. 110: 45–51.
0. Jenq, C. B., and R. E. Coggeshall. 1985. Long-term patterns ofaxon regeneration in the sciatic nerve and its tributaries. BrainRes. 345: 34–44.
1. Jenq, C. B., and R. E. Coggeshall. 1987. Permeable tubesincrease the length of the gap that regenerating axons can span.Brain Res. 408: 239–242.
2. Jenq, C. B., and R. E. Coggeshall. 1987. Sciatic nerve regenera-tion after autologous sural nerve transplantation in the rat.Brain Res. 406: 52–61.
3. Keeley, R., T. Atagi, E. Sabelman, J. Padilla, P. Kadlcik, J. Agras,L. Eng, T. W. Wiedman, K. Nguyen, A. Sudekum, and J. Rosen.1993. Synthetic nerve graft containing collagen and syntheticSchwann cells improves functional, electrophysiological, andhistological parameters of peripheral nerve regeneration. Re-stor. Neurol. Neurosci. 5: 353–366.
4. Kerns, J. M., N. Danielsen, B. Holmquist, M. Kanje, and G.Lundborg. 1993. The influence of predegeneration on regenera-tion through peripheral nerve grafts in the rat. Exp. Neurol.122: 28–36.
5. Kim, D. H., S. E. Connolly, D. G. Kline, R. M. Voories, A. Smith,M. Powell, T. Yoes, and J. K. Daniloff. 1994. Labeled Schwanncell transplants versus sural nerve grafts in nerve repair. J.Neurosurg. 80: 254–260.
6. Kline, D. G. 1990. Surgical repair of peripheral nerve injury.Muscle Nerve 13: 843–852.
7. Labrador, R. O., M. Butı, and X. Navarro. 1998. Influence ofcollagen and laminin gels concentration on nerve regenerationafter resection and tube repair. Exp. Neurol. 149: 243–252.
8. Lebeau, J. M., M. H. Ellisman, and H. C. Powell. 1988.Ultrastructural and morphometric analysis of long-term periph-eral nerve regeneration through silicone tubes. J. Neurocytol.
17: 161–172.
3
4
4
4
4
4
4
4
4
4
4
5
5
5
5
5
5
5
5
5
5
6
6
6
6
6
6
584 RODRIGUEZ ET AL.
9. Letourneau, P., S. Rogers, I. Pech, S. Palm, J. Mccarthy, and L.Furcht. 1988. Cellular biology of neuronal interactions withfibronectin and laminin. In Current Issues in Neural Regenera-tion Research (P. J. Reier, R. P. Bunge, and F. J. Seil, Eds.), pp.137–146. A. R. Liss, New York.
0. Levi, A. D. O., and R. P. Bunge. 1994. Studies of myelinformation after transplantation of human Schwann cells intothe severe combined immunodeficient mouse. Exp. Neurol. 130:41–52.
1. Levi, A. D. O., V. Guenard, P. Aebischer, and R. P. Bunge. 1994.The functional characteristics of Schwann cells cultured fromhuman peripheral nerve after transplantation into a gap withinthe rat sciatic nerve. J. Neurosci. 14: 1309–1319.
2. Levi, A. D. O., V. K. H. Sonntag, C. Dickman, J. Mather, R. H. Li,S. C. Cordoba, B. Bichard, and M. Berens. 1997. The role ofcultured Schwann cell grafts in the repair of gaps within theperipheral nervous system of primates. Exp. Neurol. 143:25–36.
3. Levi, A. D. O. 1996. Characterization of the technique involvedin isolating Schwann cells from adult human peripheral nerve.J. Neurosci. Methods 68: 21–26.
4. Liu, M. H. 1996. Growth factors and extracellular matrix inperipheral nerve regeneration, studied with a nerve chamber. J.Periph. Nerv. Syst. 2: 97–110.
5. Longo, F. M, E. G. Hayman, G. E. Davis, E. Rouslahti, E.Engvall, M. Manthorpe, and S. Varon. 1984. Neurite-promotingfactors and extracellular matrix components accumulating invivo within nerve regeneration chambers. Brain Res. 309:105–117.
6. Lundborg, G., L. B. Dahlin, N. Danielsen, R. H. Gelberman,F. M. Longo, H. C. Powell, and S. Varon. 1982. Nerve regenera-tion in silicone chambers: Influence of gap length and of distalstump components. Exp. Neurol. 76: 361–375.
7. Lundborg, G., R. H. Gelberman, F. M. Longo, H. C. Powell, andS. Varon. 1982. In vivo regeneration of cut nerves encased insilicone tubes. J. Neuropathol. Exp. Neurol. 41: 412–422.
8. Mackinnon, S. E., and A. L. Dellon. 1990. A study of nerveregeneration across synthetic (Maxon) and biologic (collagen)nerve conduits for nerve gaps up to 5 cm in the primate. J.Reconstr. Microsurg. 6: 117–121.
9. Madison, R. D., S. J. Archibald, and T. M. Brushart. 1996.Reinnervation accuracy of the rat femoral nerve by motor andsensory neurons. J. Neurosci. 16: 5698–5703.
0. Madison, R. D., and S. J. Archibald. 1994. Point sources ofSchwann cells result in growth into a nerve entubulation repairsite in the absence of axons: Effects of freeze–thawing. Exp.Neurol. 128: 266–275.
1. Madison, R. D., C. F. Dasilva, P. Dikkes, R. L. Sidman, and T. H.Chiu. 1987. Peripheral nerve regeneration with entubulationrepair: Comparison of biodegradable nerve guides versus poly-ethylene tubes and the effects of a laminin-containing gel. Exp.Neurol. 95: 378–390.
2. Midha, R., P. J. Evans, S. E. Mackinnon, T. J. Best, G. M. T.Hare, D. A. Hunter, and J. A. Wade. 1993. Comparison ofregeneration across nerve allografts with temporary or continu-ous cyclosporin A immunosuppression. J. Neurosurg. 78: 90–100.
3. Midha, R., S. E. Mackinnon, and L. E. Becker. 1994. The fate ofSchwann cells in peripheral nerve allografts. J. Neuropathol.Exp. Neurol. 53: 316–322.
4. Millesi, H. 1981. Reappraisal of nerve repair. Surg. Clin. N. Am.61: 321–341.
5. Morrisey, T. K., N. Kleitman, and R. P. Bunge. 1991. Isolationand functional characterization of Schwann cells derived fromadult peripheral nerve. J. Neurosci. 11: 2433–2442.
6. Munro, C. A., J. Jp. Szalai, S. E. Mackinnon, and R. Midha.1998. Lack of association between outcome measures of nerveregeneration. Muscle Nerve 21: 1095–1097.
7. Navarro, X., M. Butı, and E. Verdu. 1994. Autotomy preventionby amitriptyline after peripheral nerve section in differentstrains of mice. Restor. Neurol. Neurosci. 6: 151–157.
8. Navarro, X., F. J. Rodrıguez, R. O. Labrador, M. Butı, D.Ceballos, N. Gomez, J. Cuadras, and G. Perego. 1996. Periph-eral nerve regeneration through bioresorbable and durablenerve guides. J. Periph. Nerv. Syst. 1: 53–64.
9. Navarro, X., E. Verdu, and M. Butı. 1994. Comparison ofregenerative capabilities of different functional types of nervefibers. Exp. Neurol. 129: 217–224.
0. Perego, G., G. D. Cella, N. N. Aldini, M. Fini, and R. Giardino.1994. Preparation of a new nerve guide from a poly(L-lactide-co-6-caprolactone). Biomaterials 15: 189–193.
1. Perry, V. H., M. C. Brown, E. R. Lunn, P. Tree, and S. Gordon.1990. Evidence that very slow Wallerian degeneration in C57BL/Ola mice is an intrinsic property of the peripheral nerve. Eur. J.Neurosci. 2: 802–808.
2. Peulve, P., A. Laquerrierre, M. Paresy, J. Hemmet, and M.Tadie. 1994. Establishment of adult rat Schwann cell cultures:effect of b-FGF, a-MSH, NGF, PDGF, and TGF-b on cell cycle.Exp. Cell Res. 214: 543–550.
3. Raabe, T. D., D. R. Clive, T. J. Neuberger, D. Wen, and G. H.Devries. 1996. Culture neonatal Schwann cells contain andsecrete neuregulins. J. Neurosci. Res. 46: 263–270.
4. Rich, K. M., T. D. Alexander, J. C. Pryor, and J. P. Hollowell.1989. Nerve growth factor enhances regeneration though sili-cone chambers. Exp. Neurol. 105: 162–170.
5. Rodrıguez, F. J., N. Gomez, R. O. Labrador, M. Butı, D. Ceballos,J. Cuadras, E. Verdu, and X. Navarro. 1999. Improvement ofregeneration with predegenerated nerve transplants in siliconechambers. Restor. Neurol. Neurosci. 14: 65–79.
66. Rodrıguez, F. J., N. Gomez, G. Perego, and X. Navarro. 1999.Highly permeable polylactide-caprolactone nerve guides en-hance peripheral nerve regeneration through long gaps. Bioma-terials 20: 1489–1500.
67. Shenaq, J. M., S. M. Shenaq, and M. Spira. 1989. Reliability ofsciatic function index in assessing nerve regeneration across a 1cm gap. Microsurgery 10: 214–219.
68. Tapia, M., N. C. Inestrosa, and J. Alvarez. 1995. Early axonalregeneration: Repression by Schwann cells and a protease? Exp.Neurol. 131: 124–132.
69. Valentini, R. F., P. Aebischer, S. R. Winn, and P. M. Galletti.1987. Collagen- and laminin-containing gels impede peripheralnerve regeneration through semipermeable nerve guidancechannels. Exp. Neurol. 98: 350–356.
70. Williams, L. R., F. M. Longo, H. C. Powell, G. Lundborg, and S.Varon. 1983. Spatial–temporal progress of peripheral nerveregeneration within a silicone chamber: Parameters for a bioas-say. J. Comp. Neurol. 218: 160–170.
71. Williams, L. R., H. C. Powell, G. Lundborg, and S. Varon. 1984.Competence of nerve tissue as distal insert promoting nerveregeneration in a silicone chamber. Brain Res. 293: 201–211.
72. Wood, P. M. 1976. Separation of functional Schwann cells andneurons from normal peripheral nerve tissue. Brain Res. 115:361–375.