Embed Size (px)
Citation preview

Mutations inGAL2 or GAL4 alleviate catabolite repression produced bygalactose inSaccharomyces cerevisiae<
Cristina Rodrı´guez,a Carmen-Lisset Floresa,b,*aInstituto de Investigaciones Biome´dicas Alberto Sols C.S.I.C.-UAM. Unidad de Bioquı´mica y Gene´tica de Levaduras, Arturo Duperier 4, E-28029,
Madrid, Spain.bDpt. Bioquımica. Facultad de Biologı´a. Universidad de La Habana, Havana, Cuba
Received 24 March, 1999; received in revised form 14 October, 1999; accepted 1 November, 1999
Abstract
Galactose does not allow growth of pyruvate carboxylase mutants in media with ammonium as a nitrogen source, and inhibits growthof strains defective in phosphoglyceromutase in ethanol–glycerol mixtures. Starting withpyc1, pyc2, andgpm1strains, we isolated mutantsthat eliminated those galactose effects. The mutations were recessive and were nameddgr1-1anddgr2-1. Strains bearing those mutationsin an otherwise wild-type background grew slower than the wild type in rich galactose media, and their growth was dependent on respiration.Galactose repression of several enzymes was relieved in the mutants. Biochemical and genetic evidence showed thatdgr1-1was allelic withGAL2anddgr2-1with GAL4. The results indicate that the rate of galactose consumption is critical to cause catabolite repression. © 2000Elsevier Science Inc. All rights reserved.
Keywords:Catabolite repression; Galactose; Sugar transport; Yeast; Pyruvate carboxylase; Phosphoglyceromutase
1. Introduction
Utilization of non fermentable carbon sources by yeastsis not possible in the presence of sugars because of catab-olite repression. Catabolite repression is usually associatedwith glucose—it is even sometimes called glucose repres-sion [1]—but other sugars like galactose are also able torepress the expression of genes needed for utilization ofalternative carbon sources [2,3]. However, because thegenes that encode the enzymes for the galactose assimila-tory pathway are repressed by glucose [4], it is generally notrealized that galactose itself is a repressing sugar. Cataboliterepression caused by glucose has been extensively studiedand a coherent picture of its mechanisms begins to appear[1,5,6]. However, repression produced by galactose has notreceived, to our knowledge, much attention. It is not knownhow the yeast cells detect the presence of galactose and
transmit the corresponding signal(s) so that catabolite re-pression ensues. Because the initial steps of glucose andgalactose metabolism are different [7], one could imagine atleast two possibilities for these sugars to produce cataboliterepression: (1) the signals produced by glucose and galac-tose that trigger catabolite repression arise after the joiningof the two metabolic pathways; or (2) the two sugars pro-duce different signals that eventually converge at somefurther step. Mutants that affect catabolite repression byglucose have not been generally tested for their effects whengalactose is used as carbon source. To identify elementsimplicated in catabolite repression produced by galactose,we started the isolation of mutants with altered responses togalactose. To this end we used as starting material mutantsaffected in thePYC1, PYC2, or GPM1 genes. Mutants inbothPYC1andPYC2genes lack pyruvate carboxylase (E.C.6.4.1.1) and do not grow in a medium with galactose orother sugars as a carbon source and ammonium as the solenitrogen source because of their inability to replenish thetricarboxylic acid cycle [8,9]. Also, galactose or other sug-ars do not support growth ofgpm1mutants lacking phos-phoglyceromutase (E.C. 2.7.5.4) [10] and inhibit theirgrowth on alternative carbon sources likely due to cataboliterepression.
< This work was supported by Grants PB94-0091-CO2-01 and PB97-1213-CO2-01 from the Spanish DGICYT and B104-CT95-0132 from theEU project to C. Gancedo.
* Corresponding author. Tel.:134-91-585-46-20; fax:134-91-585-45-87.
E-mail address:[email protected] (C.-L. Flores).
www.elsevier.com/locate/enzmictecEnzyme and Microbial Technology 26 (2000) 748–755
0141-0229/00/$ – see front matter © 2000 Elsevier Science Inc. All rights reserved.PII: S0141-0229(00)00167-8

Our strategy was to search for mutants that with thosebackgrounds were able to grow in the presence of galactose.We reasoned that among these mutants some could bearmutations that affect the galactose sensing and/or transduc-ing machinery or catabolite repression. From about 200independent mutants with the desired phenotype, most wereaffected in theGAL2 gene and a single one in theGAL4gene.
2. Materials and methods
2.1. Yeast strains and growth conditions
The strains ofS. cerevisiaeused in this work are pre-sented in Table 1. Strains 22BHL and CLF002 were usedfor the search of the mutants; other strains, all in the W303genetic background [11], were used for crosses to segregatethe mutations obtained in either aGPM1 or a PYC1 PYC2background.
Yeasts were grown either in 2% peptone, 1% yeast ex-tract, or in 0.17% Yeast Nitrogen Base: 0.5% Ammoniumsulfate (Difco, Detriot, Michigan, USA) supplemented withthe adequate requirements. As the carbon source, 2% ga-lactose, 2% glucose, 3% glycerol, or mixtures of 3% glyc-erol 1 2.5% ethanol or 3% glycerol1 2.5% ethanol1 2%galactose were used. For solid media, 2% agar was added.Antimycin A, when used, was added at a final concentrationof 2 mg/l. When necessary, 40 mM aspartate was used as thenitrogen source instead of ammonium. Presporulation andsporulation plates were prepared as in [12]. Crosses andmicromanipulation of tetrads were done by standard proce-dures. Yeast weight is always given as wet weight (1 g ofdry weight equals approximately 5 g of wetweight).
2.2. Mutagenesis and isolation of mutants
Strains 22BHL,pyc1 pyc2, and CLF 002,gpm1, weremutagenized either with UV or with ethyl methane sulfo-nate essentially as in [13]. After mutagenesis cells fromstrain 22BHL were plated on YNB galactose ammonium
and cells from strain CLF002 were spread on YP ethanol1glycerol 1 galactose plates. Colonies appearing after 4–5days were selected and repurified on plates of the samecomposition.
2.3. Bacteria, plasmids, and library used
Escherichia coliDH5a was used for the propagation ofplasmids. LB ampicillin was used as selective medium fortheir propagation. Plasmid pBKGAL1 contains theGAL1gene with 845 bp upstream from the ATG up to 1257 bpafter it, cloned in theEcoRI site of pBluescript (Stratagene,La Jolla, CA, USA) (S. Cha´vez, personal communication).Plasmid pSEYC68-GAL2 contains theGAL2 gene underthe control of theGAL1 promoter and was constructed asfollows: the coding region ofGAL2 was amplified by PCRfrom genomic DNA of W303-1A with the following oligo-nucleotides as primers: upstream, 59-AAGCTTTCATAATGGCAGTTGAGGAG-39; downstream, 59-TGAAGCTTCTCACTTCAAACGC-39. HindIII sites (underlined) wereadded into the primers. The PCR products were clonedinto pGEM-T (Promega, Madison, Wisconsin, USA). Theresulting plasmid was disgested withHindIII, and afrag-ment of 1.7 Kb containing theGAL2gene was cloned in theHindIII site of plasmid pSEYC68 [14]. The genomic yeastlibrary GRF88 in YCp50 was obtained from the AmericanType Culture Collection, (Rockville, Maryland). To sub-clone fragments, plasmid YCp50 was used. Yeast was trans-formed as described by Ito et al. [15]. Plasmids were ex-tracted from yeast as described previously [16]. DNA wassequenced by the method of Sanger et al. [17].
2.4. RNA isolation and analysis
Total yeast RNA was extracted with TRIzol reagent byusing the protocol supplied by Gibco BRL, NY, USA. Afterelectrophoresis, RNA was transferred to nylon membranesas recommended by the supplier. The membranes werehybridized with the following probes radioactively labelledby using an oligolabelling kit (Pharmacia, Rainhan, UK):for GAL1, a 0.6-Kb NdeI-EcoRI fragment from plasmid
Table 1Yeast strains used in this work
Yeast Genotype
22BHLa Mat a ade2-1 his3-11,15 leu2-3,112 ura3-52 trp1-1 can1-100 pycl::LEU2 pyc2::HIS3CLF002 Mat a ade2-1 his3-11,15 leu2-3,112 ura3-52 trp1-1 can1-100 gpm1::LEU2 mth1::HIS3YCR001 Mat a ade2-1 his3-11,15 leu2-3,112 ura3-52 trp1-1 can1-100 pyc1::LEU2 pyc2::URA3YCR002 Mat a ade2-1 his3-11,15 leu2-3,112 ura 3-52 trp1-1 can1-100 pyc1:LEU2 pyc2::URA3 dgr1-1YCR004 Mat a ade2-1 his3-11,15 leu2-3,112 ura3-52 trp1-1 can1-100 dgr1-1CLF003 Mat a ade2-1 his3-11,15 leu2-3,112 ura3-52 trp1-1 can1-100 gpm1::LEU2 mthl::URA3CLF004 Mat a ade2-1 his3-11,15 leu2-3,112 ura3-52 trp1-1 can1-100 gpm1::LEU2 mth1::HIS3 dgr2-1CLF005 Mat a ade2-1 his3-11,15 leu2-3,112 ura3-52 trp1-1 can1-100 dgr2-1X3163-2Ab Mat a arg1 met1 trp3 ade5 lys7 gal2 mal CUPr
a Strain created by Bla´zquez et al. [9].b From Yeast Genetic Stock Center. (University of California, Berkeley, USA)
749C. Rodrıguez, C.-L. Flores / Enzyme and Microbial Technology 26 (2000) 748–755

pBKGAL1; for GAL2, a 0.7-KbPstI-PstI fragment from aPCR product fromGAL2 was used; forGAL5, a 0.4-Kbfragment from a PCR product from genomic DNA ampli-fied with the following primers: upstream, 59-ACGGTCCATTACTCGTTGAC-39; downstream, 59-AATTGCCACGGAGTCACCTGGAG-39; for GAL7, a 0.58-Kb fragmentamplified from genomic DNA with the following primers:upstream, 59-GTGCAATCTGTGAGAGGCAAT-39; down-stream, 59-CATTCAAAGGAGCCTGATGG-39; for GAL10,a 0.51-Kb fragment amplified from genomic DNA with thefollowing primers: upstream, 59-GCAATACCTAGAGGCCTACA39; downstream, 59-TAGCATACCTGCCGATCGTGG-39. As control for the RNA charge, the 0.8-kbNcoI-XbaI fragment internal to thePDA1 gene [18] was used asprobe.
2.5. Determination of enzyme activities
Cell extracts were obtained by shaking the yeasts withglass beads as described in Gamo et al [19]. Enzymes wereassayed in the crude extracts as follows: fructose-1,6-bisphosphatase as in [20], NAD-dependent glutamate dehy-drogenase as in [21], isocitrate lyase as in [22] and galac-tokinase as in [23]. Protein was determined with thebicinchoninic acid assay reagent (Pierce) with bovine serumalbumin as standard.
2.6. Acidification of the medium after sugar addition
The yeasts were grown in YPgalactose or YPglucose andcollected at the exponential phase of growth. Acidificationof the medium was measured at 20°C after addition of 10mM sugar as described by Serrano [24].
2.7. Transport of galactose
Transport of galactose was measured as described by DeJuan and Lagunas [25]. Essentially, the procedure was asfollows: yeasts were grown in YPgalactose, collected in theexponential phase of growth, and resuspended in 50 mMpotassium phosphate buffer, pH 6, at about 100 mg cells/ml.After equilibration at 20°C, 5ml of labelled 14C-galactose(0.25 mCi/mmol) was added to 50ml of this suspension(final galactose concentration, 5 mM). Next, 15 s aftergalactose addition, 10 ml of ice-cold water was added, andthe mixture was filtered under vacuum and washed on thefilter with 10 ml of ice-cold water. The filters then werecounted in scintillation vials in OptiPhase HiSafe liquid(Fisons Chemicals, Lougborough, United Kingdom). Con-trols for unspecific sugar adsorption were run in parallel byusing yeast cells previously heated for 15 minutes at 100°C.
2.8. Fermentation and respiration measurements
Fermentation and respiration of galactose were measuredat 30°C in a conventional Warburg respirometer with cell
suspensions of 20 mg cells/ml in 50 mM phosphate buffer,pH 6, with 2% galactose.
2.9. Determination of internal metabolites
Rapid sampling of yeast was done as in reference [26]and extraction of metabolites as in reference [27]. Determi-nation of glycolytic intermediates and ATP was done spec-trophotometrically as described by Bergmeyer [28]. To cal-culate the intracellular concentration of metabolites, weused the factor 1 g of yeast5 0.6 ml of cytoplasmic liquid[29].
3. Results
3.1. Isolation of mutations that suppress the toxic effectsof galactose on pyruvate carboxylase-less mutants
The reaction catalyzed by pyruvate carboxylase is theonly one available in yeasts to replenish the citric acid cycleduring growth in repressing sugars; therefore, a doublemutantpyc1 pyc2does not grow on a medium with galac-tose as the carbon source and ammonium as the nitrogensource [8,9]. The yeast strain 22BHL with mutations in bothPYCgenes was mutagenized as described in Section 2 andcolonies able to grow in the nonpermissive medium wereselected. Sixty-four independent colonies were studied. Theeffect of the mutation was specific for galactose; no growthwas observed in glucose–ammonium medium. Each se-lected mutant was crossed with thepyc1 pyc2 strainYCR001 and diploids isolated. They did not grow in thenonpermissive medium, indicating that the mutations wererecessive. Ten diploids were sporulated, and 12 to 15 com-plete tetrads of each of them were analyzed for growth ingalactose–ammonium medium. In all cases, a 21:22 segre-gation for the phenotype scored was observed, thus indicat-ing that the new phenotype was due to a nuclear, monogenicmutation. A positive scoring segregant of one of the crosses(YCR002) was chosen and crossed with all of the mutantsoriginally isolated. In no case was complementation ob-served indicating that all the mutants obtained belonged tothe same complementation group. We termed the affectedgeneDGR1 (decreased galactose repression) and the allelecarried by the arbitrarily chosen mutant straindgr1-1. Themutation was outcrossed to aPYC1 PYC2background andall the subsequent biochemical studies were done in such astrain (YCR004).
It was observed that thedgr1-1mutation was also able tosuppress the effect of galactose on agpm1 mutant (seebelow).
3.2. Isolation of mutations that relieve the toxic effects ofgalactose on phosphoglyceromutase deficient mutants
Phosphoglyceromutase encoded by the geneGPM1 cat-alyzes the interconversion between 3- and 2-phosphoglyc-
750 C. Rodrıguez, C.-L. Flores / Enzyme and Microbial Technology 26 (2000) 748–755
erates [10,30] and is therefore an enzyme essential forglycolysis as well as for gluconeogenesis. Mutations in theGPM1 gene cause inability to grow in galactose and thissugar inhibits growth of the mutants in mixtures of ethanoland glycerol.
Thegpm1strain CLF002 was mutagenized as describedin Section 2, plated on media with galactose1 glycerol 1ethanol as carbon source and colonies appearing after 4 dayswere selected. About 100 independent colonies were puri-fied in media from the same composition. Diploids from across of each mutant with agpm1strain (CLF003) did notgrow in galactose1 glycerol1 ethanol, indicating that themutations were recessive. The mutants did not grow onplates with a mixture of ethanol1 glycerol 1 glucose,indicating that the effect of the mutation was galactose-specific. The analysis of the mono- or polygenic nature ofthe mutation was not straightforward because homozygousgpm1diploids do not sporulate [19]. Therefore, we crossed10 mutants with aGPM1strain and 13-14 complete tetradsof each cross were analyzed. If the new phenotype werecaused by a nuclear monogenic mutation, and ifgpm1andthe new mutation were unlinked, a distribution 1:1:4 fortetrads of non-parental ditypes/parental ditypes/tetratypesought to be found [31]. In all cases, a pattern correspondingto a monogenic mutation not linked toGPM1 was found.
Because thedgr1-1mutation isolated by usingpyc1 pyc2mutants as starting material also allowed growth of thegpm1mutant on mixtures of galactose1 ethanol1 glyc-erol, all mutants isolated in this search were crossed with thestrain YCR004 bearing thedgr1-1mutation. Analysis of thediploids showed that all—except one—were allelic withdgr1-1. Therefore, the mutations isolated belonged to twocomplementation groups, which we termedDGR1 andDGR2. The mutant belonging to this last group was namedCFL004 and the mutation in itdgr2-1.
Both dgr1-1 and dgr2-1 mutation isolated in an other-wise wild-type background (strains YCR004 and CFL005)
caused inability to grow in galactose in the presence of therespiration inhibitor antimycin A.
3.3. Characterization of thedgr1-1mutation
Thedgr1-1mutant grew slower than the wild type in richgalactose medium (150 versus 105 min generation time).Although it was able to grow on minimal medium plateswith up to 10% galactose, we did not observe growth inmedium with 2% galactose when shake flasks were used.No problems were observed when glucose was used, show-ing that the mutation is specifically related to galactosemetabolism. A quantitative measurement of respirationshowed that the mutant in the presence of galactose con-sumed 3.5mmol O2/g yeast min, whereas the wild-typeconsumed 11mmol O2/g yeast min. Fermentation of galac-tose was not measurable indgr1-1strain, whereas the wildtype evolved 6mmol CO2/g yeast min from galactose.
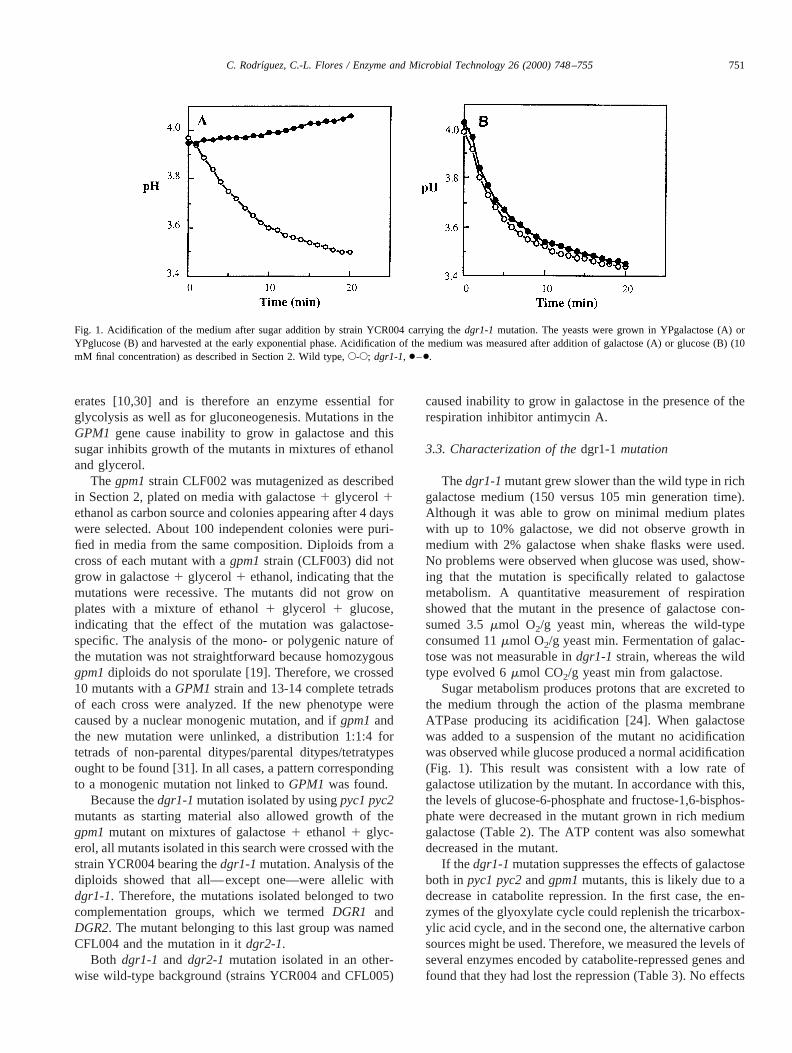
Sugar metabolism produces protons that are excreted tothe medium through the action of the plasma membraneATPase producing its acidification [24]. When galactosewas added to a suspension of the mutant no acidificationwas observed while glucose produced a normal acidification(Fig. 1). This result was consistent with a low rate ofgalactose utilization by the mutant. In accordance with this,the levels of glucose-6-phosphate and fructose-1,6-bisphos-phate were decreased in the mutant grown in rich mediumgalactose (Table 2). The ATP content was also somewhatdecreased in the mutant.
If the dgr1-1mutation suppresses the effects of galactoseboth inpyc1 pyc2andgpm1mutants, this is likely due to adecrease in catabolite repression. In the first case, the en-zymes of the glyoxylate cycle could replenish the tricarbox-ylic acid cycle, and in the second one, the alternative carbonsources might be used. Therefore, we measured the levels ofseveral enzymes encoded by catabolite-repressed genes andfound that they had lost the repression (Table 3). No effects
Fig. 1. Acidification of the medium after sugar addition by strain YCR004 carrying thedgr1-1 mutation. The yeasts were grown in YPgalactose (A) orYPglucose (B) and harvested at the early exponential phase. Acidification of the medium was measured after addition of galactose (A) or glucose (B) (10mM final concentration) as described in Section 2. Wild type,E-E; dgr1-1, ●–●.
751C. Rodrıguez, C.-L. Flores / Enzyme and Microbial Technology 26 (2000) 748–755
were observed during the growth in glucose (results notshown). In addition, we measured the activity of the firstenzyme of the galactose-utilizing pathway and found that itsactivity was about 50% of that measured in the wild type(Table 3).
3.4. Effect of thedgr1-1mutation on galactose transportand on the expression of genes of th galactose pathway
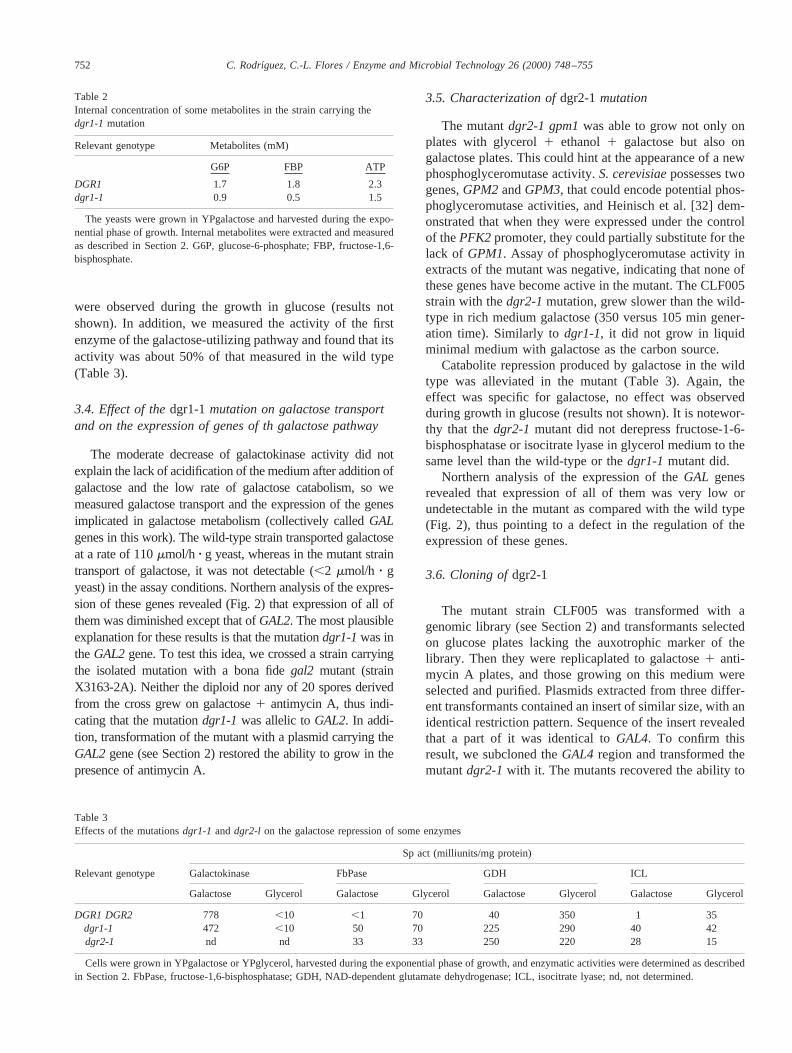
The moderate decrease of galactokinase activity did notexplain the lack of acidification of the medium after addition ofgalactose and the low rate of galactose catabolism, so wemeasured galactose transport and the expression of the genesimplicated in galactose metabolism (collectively calledGALgenes in this work). The wild-type strain transported galactoseat a rate of 110mmol/h z g yeast, whereas in the mutant straintransport of galactose, it was not detectable (,2 mmol/h z gyeast) in the assay conditions. Northern analysis of the expres-sion of these genes revealed (Fig. 2) that expression of all ofthem was diminished except that ofGAL2. The most plausibleexplanation for these results is that the mutationdgr1-1was intheGAL2gene. To test this idea, we crossed a strain carryingthe isolated mutation with a bona fidegal2 mutant (strainX3163-2A). Neither the diploid nor any of 20 spores derivedfrom the cross grew on galactose1 antimycin A, thus indi-cating that the mutationdgr1-1was allelic toGAL2. In addi-tion, transformation of the mutant with a plasmid carrying theGAL2gene (see Section 2) restored the ability to grow in thepresence of antimycin A.
3.5. Characterization ofdgr2-1mutation
The mutantdgr2-1 gpm1was able to grow not only onplates with glycerol1 ethanol 1 galactose but also ongalactose plates. This could hint at the appearance of a newphosphoglyceromutase activity.S. cerevisiaepossesses twogenes,GPM2andGPM3, that could encode potential phos-phoglyceromutase activities, and Heinisch et al. [32] dem-onstrated that when they were expressed under the controlof thePFK2promoter, they could partially substitute for thelack of GPM1. Assay of phosphoglyceromutase activity inextracts of the mutant was negative, indicating that none ofthese genes have become active in the mutant. The CLF005strain with thedgr2-1mutation, grew slower than the wild-type in rich medium galactose (350 versus 105 min gener-ation time). Similarly todgr1-1, it did not grow in liquidminimal medium with galactose as the carbon source.
Catabolite repression produced by galactose in the wildtype was alleviated in the mutant (Table 3). Again, theeffect was specific for galactose, no effect was observedduring growth in glucose (results not shown). It is notewor-thy that thedgr2-1 mutant did not derepress fructose-1-6-bisphosphatase or isocitrate lyase in glycerol medium to thesame level than the wild-type or thedgr1-1 mutant did.
Northern analysis of the expression of theGAL genesrevealed that expression of all of them was very low orundetectable in the mutant as compared with the wild type(Fig. 2), thus pointing to a defect in the regulation of theexpression of these genes.
3.6. Cloning ofdgr2-1
The mutant strain CLF005 was transformed with agenomic library (see Section 2) and transformants selectedon glucose plates lacking the auxotrophic marker of thelibrary. Then they were replicaplated to galactose1 anti-mycin A plates, and those growing on this medium wereselected and purified. Plasmids extracted from three differ-ent transformants contained an insert of similar size, with anidentical restriction pattern. Sequence of the insert revealedthat a part of it was identical toGAL4. To confirm thisresult, we subcloned theGAL4 region and transformed themutantdgr2-1with it. The mutants recovered the ability to
Table 2Internal concentration of some metabolites in the strain carrying thedgr1-1 mutation
Relevant genotype Metabolites (mM)
G6P FBP ATP
DGR1 1.7 1.8 2.3dgr1-1 0.9 0.5 1.5
The yeasts were grown in YPgalactose and harvested during the expo-nential phase of growth. Internal metabolites were extracted and measuredas described in Section 2. G6P, glucose-6-phosphate; FBP, fructose-1,6-bisphosphate.
Table 3Effects of the mutationsdgr1-1 anddgr2-l on the galactose repression of some enzymes
Sp act (milliunits/mg protein)
Relevant genotype Galactokinase FbPase GDH ICL
Galactose Glycerol Galactose Glycerol Galactose Glycerol Galactose Glycerol
DGR1 DGR2 778 ,10 ,1 70 40 350 1 35dgr1-1 472 ,10 50 70 225 290 40 42dgr2-1 nd nd 33 33 250 220 28 15
Cells were grown in YPgalactose or YPglycerol, harvested during the exponential phase of growth, and enzymatic activities were determined as describedin Section 2. FbPase, fructose-1,6-bisphosphatase; GDH, NAD-dependent glutamate dehydrogenase; ICL, isocitrate lyase; nd, not determined.
752 C. Rodrıguez, C.-L. Flores / Enzyme and Microbial Technology 26 (2000) 748–755

grow on galactose1 antimycin A plates. This result indi-cates that the mutationdgr2-1 is in theGAL4 gene.
4. Discussion
We have isolated suppressor mutants that allow growthof different glycolytic mutants in the presence of galactose.The two mutations isolated in this work interfere with ga-lactose utilization in two different ways: one of them,dgr1-1 that turned out to be allelic withGAL2, interfereswith an effective galactose uptake, and the other one,dgr2-1that was allelic withGAL4, hinders the induction of theGALgenes.
In contrast with the wide variety of glucose transporters,only Gal2 efficiently transports galactose [33–35]. How-ever, the mutantdgr1-1, was able to grow in galactose ifrespiration was not inhibited. A similar result was obtainedby Donnini et al [36] during the study of the mutationimp1that also turned out to be allelic withGAL2. Therefore, analternative transport system should be able to transportgalactose. An earlier report indicated that galactose may betransported by the glucose transport system [37], and morerecent results from Reifenberger et al. [35] indicated thatsome of the glucose transporter may transport galactosewith a Km higher than 250 mM. However, the particularglucose transporter responsible for this marginal but impor-tant activity has not been yet clearly identified.
The other mutation isolated,drg2-1, allelic with theGAL4gene, interferred with galactose metabolism—includ-ing uptake—because it prevented the expression of theGALgenes.GAL4 encodes an activator protein that binds tospecific regions in the promoters of theGAL genes [4].Curiously, the mutation isolated appears to affect in someway the derepression of fructose-1,6 bisphosphatase andisocitrate lyase. We have not yet a clear explanation for thiseffect.
The results with both mutants show a correlation be-tween the rate of galactose metabolism and galactose re-pression. Thus, the situation is reminiscent of that foundwith glucose. Different results indicate that in the case ofglucose, the strength of catabolite repression is correlatedwith the rate of its consumption and glycolytic flux [19,35].However, as pointed out by Gamo et al. [19], the flux byitself cannot be a regulatory factor, because regulatory sys-tems respond only to changes in the concentration of somemetabolite, not to changes in the rate of its formation. Howmay galactose exert catabolite repression? Two main pos-sibilities—not mutually exclusive—are either an interactionwith a membrane protein that will initiate some type ofregulatory cascade, or a change in the levels of metabolitesthat could activate or inhibit some regulatory proteins. In thecase of glucose, the two possibilities appear to be operative,although the second one has not been directly documented.Glucose interacts with either of two sensors, Snf3 or Rgt2[38] and controls the synthesis of some glucose transporters
Fig. 2. Northern blot analysis of the transcripts of theGAL genes in strains bearing the mutationsdgr1-1 and dgr2-1. The metabolic steps of galactosemetabolism form transport to funneling into the common glycolytic pathway are shown in the scheme. The name of the gene encoding the correspondingenzyme is indicated in each step. Total RNA was prepared from cells grown in YP galactose as indicated in Section 2. Each lane contained 15mg of totalRNA. Hybridization was carried out with the probes described in Section 2. As an internal control of mRNA charge, a probe of thePDA1 gene was used;see Section 2 [18].
753C. Rodrıguez, C.-L. Flores / Enzyme and Microbial Technology 26 (2000) 748–755

through the proteins Mth1 and Std1 [39,40]. In asnf3 rgt2mutant repression of theGAL1, SUC2[41] andFBP1 [40]is altered, likely as an indirect result of the lack on inductionof the relevant glucose transporters.
Concerning the second possibility, it is documented thatthe concentration of a series of metabolites is markedlydifferent in repression or derepression conditions [42]; how-ever, a direct effect of these changes on catabolite repres-sion has not been shown.
In the case of galactose, no equivalent to Snf3 or Rgt2have been found, and with the whole genome sequenced,there is no hint to expect so. Likely, Gal2 activity couldgenerate a signal triggering catabolite repression either di-rectly, with the protein acting as a sensor, or indirectlydetermining the changes in concentration of an intermediatethat could function as a signal. Our results do not allow todecide between the two possibilities.
Among 200 independent mutants, practically all wereallelic of GAL2, and hence the question arises why muta-tions in other genes have not been found. For example,mutations inGAL1 that would block phosphorylation ofgalactose or inGAL11the loss of function of which leads toa reduced expression of mostGAL genes [43]. There is nodefinite answer to this question, but we think that this resultindicates that transport has a high control coefficient in thegalactose pathway and that the screening favor the selectionof such mutants. It is also noteworthy that no mutations similarto HTR1-23, DGT1-1, or BPC1-1[9,19,44] were found. Thesemutations are allelic withMTH1 and control the synthesis ofsome glucose transporters in response to the concentration ofsugar in the medium [19,40]. Since the selection screenings inwhich these mutations were found were similar to those usedin this work—the only difference being the sugar used—itwould appear that the synthesis of Gal2 and that of the Hxttransporters are not controlled by similar mechanisms. Perhapsthe control exerted onGAL2expression by Gal4 and Mig1(4)is sufficient for efficient regulation.
Our results for galactose, together with those of otherauthors for glucose [19,44] highlight the importance of thetransport step for a coordinated function of the cell.
Acknowledgments
We thank Carlos Gancedo for continuous support alongthis work and help in the preparation of the manuscript,Juana M. Gancedo for critical reading of the manuscript, S.Chavez (Seville, Spain) for the gift of plasmid pBKGAL1,and Y. Steensma (Leiden, The Netherlands) for the plasmidwith thePDA1gene. This work is part of the project “Fromgene to products in yeasts: a quantitative approach” supportedby the European Community (DG XII Framework IV, Pro-gram of Cell factories). C.R. benefitted from a Fellowship fromthe Fundacio´n Ramon Areces, Madrid, Spain. C.-L. F. thanksthe academic authorities of the Facultad de Biologı´a, Univer-sidad de La Habana for a leave of absence.
References
[1] Ronne H. Glucose repression in fungi. Trends Genet 1995;11:12–7.[2] Scholler A, Schuller HJ. A carbon source-responsive promoter ele-
ment necessary for activation of the isocitrate lyase geneICL1 iscommon to genes of the gluconeogenic pathway in the yeastSaccha-romyces cerevisiae. Mol Cell Biol 1994;14:3613–22.
[3] Rodrıguez C, Gancedo JM. Glucose signalling in yeast is partiallymimicked by galactose and does not require the Tps1 protein. MolCell Biol Res Com 1999;1:55–8.
[4] Mechler K. Galactose metabolism inSaccharomyces cerevisiae: aparadigm for eukaryotic gene regulation. In: Zimmermann FK, EntianKD, editors. Yeast sugar metabolism, Basel, Switzerland: TechnomicPublishing, 1997. p 235–70.
[5] Johnston M, Carlson M. Regulation of carbon and phosphate utiliza-tion. In: Jones EW, Pringle JR, Broach JR. editors. The molecularbiology of the yeastSaccharomyces2nd ed., vol 2. Cold SpringHarbor, NY: Cold Spring Harbor Laboratory, 1992. p. 193–281.
[6] Gancedo JM. Yeast carbon catabolite repression. Microbiol Mol BiolRev 1998;62:334–61.
[7] Gancedo C, Serrano R. Energy Yielding Metabolism. In: Rose AH,Harrison JS, editors. The Yeasts, 2nd ed., vol 3. New York: AcademicPress, 1989. p. 205–60.
[8] Stucka R, Dequin S, Salmon JM, Gancedo C. DNA sequences inchromosomes II and VII code for pyruvate carboxylase isoenzymes inSaccharomyces cerevisiae. Mol Gen Genet 1991;229:307–15.
[9] Blazquez MA, Gamo FJ, Gancedo CA. A mutation affecting carboncatobolite repression suppresses growth deffects in pyruvate carbox-ylase mutants fromSaccharomyces cerevisiae. FEBS Lett 1995;337:197–200.
[10] Rodicio R, Heinisch JJ. Isolation of the yeast phosphoglycerate mu-tase gene and construction of deletion mutants. Mol Gen Genet1987;206:133–40.
[11] Thomas BJ, Rothstein R. Elevated recombination rates in transcrip-tionally active DNA. Cell 1989;56:619–30.
[12] Sherman F. Getting started with yeast. Meth Enzymol 1991;194:3–21.
[13] Lawrence CW. Clasical mutagenesis techniques. Meth Enzymol1991;194:273–81.
[14] Sutterlin C, Escribano MV, Gerold P, Maedas Y, Mazo´n MJ, Ki-noshita T, Schwarz RT, and Riezman H.Saccharomyces cerevisiaeGPI10, the functional homologue of human PIG-B, is required forglycosylphosphatidylinositol-anchor synthesis. Biochem J 1998;332:153–59.
[15] Ito H, Fukuda Y, Murata K, Kimura A. Transformation of intact yeastcells treated with alkali cations. J Bacteriol 1983;153:163–8.
[16] Robzyk K, Kassir Y. A simple and highly efficient procedure for rescu-ing autonomous plasmidss from yeast. Nuc Acid Res 1992;20:3790.
[17] Sanger F, Nicklen S, Coulson AR. DNA sequencing with chainterminating inhibitors. Proc Natl Acad Sci USA 1977;74:5463–67.
[18] Wenzel TJ, Teunissen AWRH, Steensma Y.PDA1 mRNA: a stan-dard for quantitation of mRNA inSaccharomyces cerevisiaesuperiorto ACT1mRNA. Nuc Acid Res 1995;23:883–4.
[19] Gamo FJ, Lafuente MJ, Gancedo C. The mutationDGT1–1decreasesglucose transport and alleviates carbon catabolite repression inSac-charomyces cerevisiae. J Bacteriol 1994;176:7423–29.
[20] Gancedo JM, Gancedo C. Fructose-1,6-diphosphatase, phosphofruc-tokinase and glucose-6-phosphate dehydrogenase in fermenting andnon-fermenting yeasts. Arch Microbiol 1971;76:132–8.
[21] DohertyDL-Glutamate dehydrogenases (yeast). Meth Enzymol 1970;17:850–56.
[22] Dixon GH, Kornberg HL. Assay methods for key enzymes of theglyoxylate cycle. Biochem J 1959;72:3.
[23] Maitra PKY, Lobo Z. A kinetic study of glycolytic enzyme synthesisin yeast. J Biol Chem 1971;246:475–88.
754 C. Rodrıguez, C.-L. Flores / Enzyme and Microbial Technology 26 (2000) 748–755

[24] Serrano R. Effect of ATPase inhibitors on the proton pump of respi-ratory deficient yeast. Eur J Biochem 1980;105:419–24.
[25] De Juan C, Lagunas R. Inactivation of the galactose transport systemin Saccharomyces cerevisiae.FEBS Lett 1986;207:258–61.
[26] Saez MJ, Lagunas R. Determination of intermediary metabolites in yeast.Critical examination of the effect of sampling conditions and recommen-dations to obtain true values. Mol Cell Biochem 1976;13:73–8.
[27] Gamo FJ, Portillo F, Gancedo C. Characterization of mutants that over-come the toxic effect of glucose on phosphoglucoseisomerase less strainsof Saccharomyces cerevisiae. FEMS Mic Lett 1993;106:233–8.
[28] Bergmeyer HU. Methods of enzymatic analysis. Weinheim, Ger-many: Verlag Chemie, 1983.
[29] Conway EJ, Downey M. An outer metabolic region of the yeast cell.Biochem J 1950;47:355.
[30] Fothergill–Gilmore LA, Watson HC. The phosphoglycerate mutases.Adv Enzymol Relat Areas Mol Biol 1989;62:227–313.
[31] Mortimer RK, Hawthorne DC. Yeast Genetics. In: Rose AH, Harri-son JS, editors. The Yeasts, 1st ed., vol. 1. New York: AcademicPress, 1969. p. 386–460.
[32] Heinisch JJ, Mu¨ller S, Schluter E, Jacoby J, Rodicio R. Investigationof two yeast genes encoding putative isoenzymes of phosphoglycer-ate mutase. Yeast 1998;14:203–13.
[33] Tschopp JF, Emr SD, Field C, Schekman R.GAL2 codes for amembrane-bound subunit of the galactose permease inSaccharomy-ces cerevisiae. J Bacteriol 1986;166:313–8.
[34] Boles E, Hollenberg CP. The molecular genetics of hexose transportin yeast. FEMS Microbiol Rev 1997;21:85–111.
[35] Reifenberger E, Boles E, Ciriacy M. Kinetic characterization ofindividual hexose transporters ofSaccharomyces cerevisiaeand their
relation to the triggering mechanisms of glucose repression. EurJ Biochem 1997;245:324–33.
[36] Donnini C, Lodi T, Ferrero I, Algeri A, Puglisi PP. Allelism ofIMP1 andGAL2genes ofSaccharomyces cerevisiae. J Bacteriol 1992;174:3411–15.
[37] Cirillo V. Galactose transport inSaccharomyces cerevisiae. 1. Non-metabolized sugars as substrates and inducers of the galactose trans-port system. J Bacteriol 1968;95:1727–31.
[38] Ozcan S, Dover J, Rosenwasl AG, Johnston MA. Two glucose trans-porters inSaccharomyces cerevisiaeare glucose sensors that generatea signal for induction of gene expression. Proc Natl Acad Sci 1996;93:12428–32.
[39] Schmidt MC, McCartney RR, Zhang X, Tillman TS, Solimeo H,Wolfl S, Almonte C, Watkins SC. Std1 and Mth1 proteins interactwith the glucose sensors to control glucose regulated expression inSaccharomyces cerevisiae. Mol Cell Biol 1999;19:4561–71.
[40] Lafuente MJ, Gancedo C, Jauniaux JC, Gancedo JM. Mth1 receivesthe signal given by the glucose sensors Snf3 and Rgt2 inSaccharo-myces cerevisiae. Mol Microbiol 2000;35:161–172.
[41] Ozcan S, Dover J, Johnston M. Glucose sensing and signaling by twoglucose receptors in the yeastSaccharomyces cerevisiae. EMBO J1998;17:2566–73.
[42] Gancedo JM, Gancedo C. Concentrations of intermediary metabolitesin yeast. Biochemie 1973;55:205–11.
[43] Amakasu H, Suzuki Y, Nishizawa M, Fukasawa T. Isolation andcharacterization ofSGE1: a yeast gene that partially suppresses thegal11 mutation in multiple copies. Genetics 1993;134:675–83.
[44] Ozcan S, Freidel K, Leuker A, Ciriacy M. Glucose uptake andcatabolite repression in dominantHTR1 mutants ofSaccharomycescerevisiae. J Bacteriol 1993;175:5520–8.
755C. Rodrıguez, C.-L. Flores / Enzyme and Microbial Technology 26 (2000) 748–755







![STUDIES OF THE DROSOPHILA BRAIN USING P[GAL4] ENHANCER …theses.gla.ac.uk/75487/1/13832077.pdf · 2019. 11. 19. · STUDIES OF THE DROSOPHILA BRAIN USING P[GAL4] ENHANCER TRAP LINES](https://img.pdfslide.us/doc/110x75/613774870ad5d2067648a163/studies-of-the-drosophila-brain-using-pgal4-enhancer-2019-11-19-studies-of.jpg)





















