Embed Size (px)
Citation preview

University of Central Florida University of Central Florida
STARS STARS
Electronic Theses and Dissertations
2019
Motor and Sensory Characterization of a Mouse Model of Motor and Sensory Characterization of a Mouse Model of
Charcot-Marie-Tooth Type 2O Disease Charcot-Marie-Tooth Type 2O Disease
Swaran Nandini University of Central Florida
Part of the Medical Sciences Commons
Find similar works at: https://stars.library.ucf.edu/etd
University of Central Florida Libraries http://library.ucf.edu
This Doctoral Dissertation (Open Access) is brought to you for free and open access by STARS. It has been accepted
for inclusion in Electronic Theses and Dissertations by an authorized administrator of STARS. For more information,
please contact [email protected].
STARS Citation STARS Citation Nandini, Swaran, "Motor and Sensory Characterization of a Mouse Model of Charcot-Marie-Tooth Type 2O Disease" (2019). Electronic Theses and Dissertations. 6386. https://stars.library.ucf.edu/etd/6386

MOTOR AND SENSORY CHARACTERIZATION OF A MOUSE MODEL OF CHARCOT-MARIE-TOOTH TYPE 2O DISEASE
by
SWARAN NANDINI M.S, University of Central Florida, 2013
B. Tech., Jaypee Institute of Technology (India), 2011
A dissertation submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy
in the Burnett School of Biomedical Sciences in the College of Medicine
at the University of Central Florida Orlando, Florida
Spring Term 2019
Major Professor: Stephen J. King

ii
©2019 SWARAN NANDINI

iii
ABSTRACT
Dynein is an essential motor protein required for the maintenance and survival of
cells. Dynein forms a motor complex to carry intracellular cargoes like organelles,
growth factors, peptides, and hormones along the microtubules inside the cells.
In neurons, the dynein is the retrograde motor protein that moves cargoes from
the neuronal tip to the neuronal soma along the length of an axon. Dynein has an
established role in neuronal nuclear migration, transport of neuronal survival
signals and growth factors, organelle positioning inside neurons etc. Hence, it is
not very surprising that numerous mutations in dynein have been reported in
association with neurodegenerative diseases in humans. The first human
mutation (H306R) in dynein heavy chain was reported to cause Charcot-Marie-
Tooth Type 2O disease (CMT2O) in humans. CMT2O patients display motor-
sensory neuropathy symptoms such as muscle weaknesses and wasting in legs,
skeletal deformities like pes cavus (high foot arching), difficulty in walking, and a
loss of sensation.
We developed a novel knock-in H304R mouse model with the corresponding
CMT2O linked dynein mutation to understand the disease’s molecular
mechanism. We investigated and characterized the motor-sensory phenotype of
the H304R mouse model (wildtype, heterozygous (H304R/+) and homozygous
(H304R/R) mice). First, we started with testing mice on motor skills behavior
tests such as tail suspension reflex, grip strength test, and rotarod test at 3, 6, 9

iv
and 12 months of age. Both male and female groups of heterozygous (H304R/+)
mice displayed mild defects in tail suspension reflex, grip strength, and rotarod
performance. In contrast, homozygous (H304R/R) mice exhibited severe defects
in the tail suspension reflex, grip strength, and rotarod performance right from an
early age. Next, I analyzed the sensory phenotype of the H304R mouse model.
Homozygous H304R/R mice appeared to have thinner sciatic nerves, reduced
total fascicular area of the sciatic nerve, and significantly quicker latency to tail
withdrawal from a pain stimulus than the wildtype and heterozygous H304R/+
mice.
Collectively, our motor and sensory characterization studies reveal that H304R
dynein mouse model recapitulates many of the phenotypes associated with CMT
symptoms. Hence, the H304R model is a useful tool in understanding the dynein
function in the onset and progression of CMT2O in humans.

v
I dedicate my PhD dissertation work to my grandparents, in honor of their
sacrifices, struggles, and hard work that afforded my family the privilege to
pursue our dreams.

vi
ACKNOWLEDGMENTS
First and foremost, I thank my PhD advisor, Dr. Stephen J. King for providing me
the opportunity to pursue my doctoral research in his lab. I specially thank him for
helping me develop my critical thinking and effective communication skills. His
mentorship has built me professionally both inside and outside the lab. Because
of his open-minded support, I was able to collaborate with other scientists and
learn many valuable scientific techniques and skills. In addition, I am also very
grateful to Dr. King for letting me be independent in pursuing my interests and
that has allowed me to build a strong leadership profile outside the lab. I also
thank him for creating a very welcoming and understanding environment in the
lab.
Next, I am very grateful to both the past and present members on my PhD
Dissertation Committee. Special thanks to Dr. Yoon-Seong Kim who has been a
long-term committee member for my M.S. thesis (2011-2013) and PhD
Dissertation (2013-2019) committees. Thank you, Dr. Kim, for showing me the
importance of thinking outside the box when looking for possible scientific
solutions. Special thanks to Dr. Annette Khaled for being a strong female role
model for me, both in science and in leadership roles. Thank you, Dr. Khaled, for
your direction and support for past so many years. Next, I want to thank
Dr. Stephen Lambert for being the most authentic person and well-rounded
scientist I know. Thank you, Dr. Lambert, for showing me the power of being

vii
passionately true to ourselves. Also, thank you for allowing me to learn the
specialized skills of mouse surgeries in your lab (thanks to Dr. Dale George and
Dr. Wesley Anderson). Last and of course not the least; I would like to thank Dr.
Alvaro Estevez who was a former M.S. and PhD committee member. Thank you,
Dr. Estevez for always having your office and lab door open for me walk in and
seek your guidance while you were at UCF. Thank you, Dr. Estevez, for always
believing in me. In addition, special thanks to Dr. Maria Clara Franco as well for
teaching me some novel techniques (mitochondrial isolation and oxygen
consumption assays) and the importance of designing in-depth experiments in
order to perform thorough science. Also, big thanks to Dr. Raheleh Ahangari for
being such a wonderful and kind advisor for my graduate teaching
assistantships. Thank you to all the above-mentioned scientists for challenging
me to be a well-trained PhD graduate student.
Thanks to the past and current members of King lab that made the environment
so collaborative and fun – Dr. Linda King, Rachal Love, Jami Conley Calderon,
Dr. Thywill Sabblah, Aaron Ledray, Julio Pasos, Alexis Ghersi, Rochelle Sadeghi,
Bryce Ordway, Keira Kraus, Emily Silva, Gabi Fiorino and Phil Hannoush. In
addition, special thanks to the former and current Burnett School of Biomedical
Sciences (BSBS) Program Leaders – Dr. Sampath Parthasarathy, Dr. Griffith
Parks, Dr. Saleh Naser, Dr. Jihe Zhao and Mr. Greg Norris for their support in
every step of the way of my graduate career. Many thanks to Lisa Vaughn,

viii
Allison Connally, Ka Yam, and Shannon Connally for being the strong backbone
of the BSBS PhD program.
Also special thanks to the former and current Lake Nona Staff members for their
efforts in making our lives easier so that we can solely focus on research – Lisa
Simcoe, David Frosch, Amy Postlewait, Jeanette Galloway, Abby Snipes, and
Susie Nisavic; the BSBS custodial staff (Ms. Lucy Bautista); the BSBS IT
(Robinson Pamplano and Alex Lazin); and the BSBS Engineering (Carol
Lanouette, Anthony Smith, and Joseph Myerson). And of course, special thanks
to the former and current BSBS Vivarium Staff members and Teri Krisch and
James Grant for making it so convenient for me to complete my experiments in
mice for past so many years. I also thank my peers in the BSBS program for
being so friendly and supportive of each other.
Serving multiple leadership roles in and outside of UCF has been a personal
career highlight for me. I am very thankful to all the leadership staff members I
had the opportunity to work with on the main campus. Moreover, I specially thank
them for always being so understanding and adaptable to my lab schedule and
letting me work on the leadership projects at my own schedule. Special thanks to
the UCF Recreation and Wellness Center (Mr. James Wilkening and the team),
UCF Internationalization Committee (Ms. Chantel Carter and the committee),
UCF Technology Fee Committee (Dr. Joel Hartman and the committee) for
wholeheartedly providing me with many valuable leadership opportunities for
past many years.

ix
Lastly, I want to thank my family and loved ones for being so selflessly patient
and understanding through the intense journey of the graduate school. Thank
you to all my friends who have shown me their unconditional support through
their actions, time and again. Last but not the least; deep heartfelt thanks to my
patrons (UCF students) that came to my group exercises classes every week for
past so many years. Their love, support, and energy are what kept me
encouraged and positive through this long journey of earning my doctoral degree.

x
TABLE OF CONTENTS
LIST OF FIGURES ..............................................................................................xv
LIST OF TABLES ............................................................................................. xviii
LIST OF ABBREVIATIONS .................................................................................xx
CHAPTER 1: INTRODUCTION ............................................................................ 1
1.1 Cytoplasmic Dynein Structure ................................................................. 3
1.1.1 Cytoplasmic Dynein Heavy Chain Structure ..................................... 4
1.2 Cytoplasmic Dynein Function .................................................................. 6
1.2.1 Dynein interactions with its binding partners ................................... 11
1.3 Charcot-Marie-Tooth Diseases in Humans ............................................ 20
1.3.1 Dynein mutation linked to CMT2O and SMA-LED .......................... 23
1.4 Mouse Models with Neurodegeneration Linked Dynein Mutations ........ 25
1.5 Peripheral Nervous System of Mouse ................................................... 30
1.5.1 Role of dynein mediated retrograde signaling in sensory neurons . 31
1.5.2 Sciatic Nerve anatomy in mice ........................................................ 35
1.5.3 Dorsal Root Ganglia anatomy in mice ............................................. 37
CHAPTER 2: MATERIALS AND METHODS ...................................................... 40
2.1 Generation of H304R mouse model ...................................................... 40
2.2 Mouse colony maintenance ................................................................... 42

xi
2.2.1 Mouse colony housing .................................................................... 42
2.2.2 Mouse colony breeding ................................................................... 42
2.2.3 Measurement of mouse body weight .............................................. 43
2.2.4 Mouse health supervision ............................................................... 43
2.3 Behavioral phenotype characterization test ........................................... 44
2.3.1 Tail suspension test ........................................................................ 44
2.3.2 Grip strength test ............................................................................ 46
2.3.3 Rotarod performance test ............................................................... 47
2.3.4 Tail flick test .................................................................................... 49
2.4 Mouse dissections (sciatic nerve and dorsal root ganglia) .................... 50
2.4.1 Tissue harvesting and cryopreservation ......................................... 50
2.4.2 Cryosectioning and immunohistochemistry of tissue samples ........ 50
2.4.3 Microscopy and data analysis ......................................................... 53
2.5 Motility analysis studies in primary H304R neurons .............................. 54
2.5.1 Tissue harvesting of embryonic primary neurons............................ 54
2.5.2 Live neuronal imaging with Rab7-GFP labeled vesicles ................. 55
2.5.3 Live neuronal imaging with Synaptophysin-RFP labeled vesicles .. 58
CHAPTER 3: BEHAVIORAL PHENOTYPE CHARACTERIZATIONS OF H304R
MICE .................................................................................................................. 59

xii
3.1 Introduction ............................................................................................ 59
3.2 Rationale ............................................................................................... 61
3.3 Results .................................................................................................. 64
3.3.1 H304R mice had normal life span ................................................... 64
3.3.2 H304R mice showed atypical tail suspension phenotype ............... 65
3.3.3 Heterozygous H304R/+ mice show mild defects in their motor
phenotype .................................................................................................... 69
3.3.4 Homozygous H304R/+ mice show severe defects in their motor
phenotype .................................................................................................... 75
3.4 Conclusions ........................................................................................... 81
CHAPTER 4: SENSORY PHENOTYPE CHARACTERIZATIONS OF H304R
MICE .................................................................................................................. 85
4.1 Introduction ............................................................................................ 85
4.1.1 Sensory deficits in previously published mutant dynein mice models
85
4.2 Rationale ............................................................................................... 88
4.3 Results .................................................................................................. 89
4.3.1 Altered latency to respond to pain stimulus in H304R mice ............ 89
4.3.2 Sciatic nerve appeared thinner in H304R mice ............................... 92

xiii
4.3.3 Reduced total sciatic nerve fascicular area in H304R mice ............ 93
4.3.4 Axonal counts in the sciatic nerve cross section ............................. 98
4.3.5 Identifying subpopulations of neurons in the adult DRG ............... 102
4.4 Conclusions ......................................................................................... 107
CHAPTER 5: MOTILITY STUDIES OF H304R MICE ...................................... 111
5.1 Introduction and rationale .................................................................... 111
5.2 Results ................................................................................................ 112
5.2.1 Synaptophysin-RFP positive vesicular motility in hippocampal
neurons ..................................................................................................... 112
5.2.2 Synaptophysin-RFP positive vesicular motility in cortical neurons 117
5.2.3 Rab7-GFP positive vesicular motility in cortical neurons .............. 122
5.3 Conclusions ......................................................................................... 125
CHAPTER 6: DISCUSSION ............................................................................. 127
6.1 All dynein mutations don’t act the same .............................................. 127
6.2 Relevance of H304R Mouse Model ..................................................... 129
6.3 Comparison of H304R Mouse Model with Other Dynein Mouse Models
133
6.4 Future Sensory Characterization Work of H304R Mouse Model ......... 136
6.5 Potential Dynein Disruptions in The H304R Mice and Future Studies . 139

xiv
APPENDIX A: DISSERTATION DEFENSE ANNOUCEMENT ......................... 142
REFERENCES ................................................................................................. 148

xv
LIST OF FIGURES
Figure 1-1 Intracellular motility inside a cell .......................................................... 2
Figure 1-2 Cytoplasmic Dynein Structure ............................................................. 3
Figure 1-3 Cytoplasmic Dynein Heavy Chain (DHC) peptide ............................... 4
Figure 1-4 Binding partners of dynein ................................................................. 13
Figure 1-5 Mutations in Dynein Heavy Chain (DHC) .......................................... 16
Figure 1-6 Clinical symptoms associated with Charcot Marie Tooth (CMT) ....... 21
Figure 1-7 CMT2O and SMA-LED linked mutation in dynein heavy chain ......... 23
Figure 1-8 Dynein heavy chain (DHC) mutations in mice ................................... 26
Figure 1-9 Pseudo-unipolar sensory neurons in DRGs ...................................... 30
Figure 1-10 Neurotrophic factors in sensory neurons development ................... 31
Figure 1-11 Neurotrophic factors activate Trk signaling in neurons .................... 32
Figure 1-12 Sciatic nerve anatomy in mice ......................................................... 36
Figure 1-13 Sciatic nerve is a mixed nerve ......................................................... 37
Figure 2-1 Generation of knock-in H304R mouse model .................................... 41
Figure 2-2 Tail suspension test ........................................................................... 45
Figure 2-3 Grip strength test meter ..................................................................... 46
Figure 2-4 Rotarod machine to measure motor coordination ............................. 47
Figure 2-5 Rotarod test profile ............................................................................ 48
Figure 2-6 Tail flick meter ................................................................................... 49
Figure 2-7 Schematic diagram of motility analysis using kymographs ................ 57
Figure 3-1 Comparison of dynein heavy chain peptide ....................................... 62

xvi
Figure 3-2 Gait phenotype of H304R mouse model ........................................... 65
Figure 3-3 Tail suspension phenotype of H304R mouse model ......................... 66
Figure 3-4 Atypical tail suspension response of H304R mouse model ............... 67
Figure 3-5 Grip strength phenotype of heterozygous H304R/+ mice .................. 70
Figure 3-6 Rotarod performance test of the H304R/+ mice ................................ 73
Figure 3-7 Grip strength phenotype of heterozygous H304R/R mice ................. 76
Figure 3-8 Rotarod performance test of the H304R/R mice ............................... 79
Figure 4-1 Sciatic nerve cross section ................................................................ 86
Figure 4-2 Tail flick pain sensitivity test of H304R male mice ............................. 89
Figure 4-3 Comparison of sciatic nerve thickness of H304R male mice ............. 93
Figure 4-4 Sciatic nerve cross sections of 3-month H304R male mice ............... 94
Figure 4-5 Sciatic nerve cross sections of 12-month H304R male mice ............. 95
Figure 4-6 Total fascicular area of sciatic nerve in H304R male mice ................ 96
Figure 4-7 The largest fascicle inside the sciatic nerve of each genotype .......... 99
Figure 4-8 Total count of axons inside a fascicle .............................................. 100
Figure 4-9 The percentage distribution of bundled axons vs single axons ....... 101
Figure 4-10 Serial sections of dorsal root ganglia of H304R mice .................... 103
Figure 4-11 Neuronal subpopulations staining of dorsal root ganglia ............... 106
Figure 5-1 Synaptophysin-RFP positive vesicles motility in hippocampal neurons
at DIV 10 .......................................................................................................... 114
Figure 5-2 Synaptophysin-RFP positive vesicles motility in hippocampal neurons
at DIV 14 .......................................................................................................... 116

xvii
Figure 5-3 Synaptophysin-RFP positive vesicles motility in cortical neurons at
DIV 10 .............................................................................................................. 119
Figure 5-4 Synaptophysin-RFP positive vesicles motility in cortical neurons at
DIV 14 .............................................................................................................. 121
Figure 5-5 Rab7-GFP positive vesicles motility in cortical neurons at DIV 10 .. 124
Figure 6-1 Severity of phenotypes associated with DHC mutations ................. 135

xviii
LIST OF TABLES
Table 1-1 Cellular functions of cytoplasmic dynein ............................................... 8
Table 1-2 Dynein heavy chain mutations in humans .......................................... 17
Table 1-2 Types of Charcot-Marie-Tooth disease in humans ............................. 22
Table 1-3 Dynein heavy chain mutations in mice ............................................... 27
Table 1-4 Major types of somatosensory neurons in the DRG ........................... 38
Table 2-1 Antibodies used for tissue immunohistochemistry .............................. 52
Table 2-2 Microscopic settings used for imaging ................................................ 53
Table 3-1 Tail suspension reflex of H304R mice ................................................ 68
Table 3-2 Grip strength in H304R/+ male mice .................................................. 71
Table 3-3 Grip strength in H304R/+ female mice ............................................... 72
Table 3-4 Rotarod performance in H304R/+ male mice ..................................... 74
Table 3-5 Rotarod performance in H304R/+ female mice .................................. 74
Table 3-6 Grip strength in H304R/R male mice .................................................. 77
Table 3-7 Grip strength in H304R/R female mice ............................................... 78
Table 3-8 Rotarod performance in H304R/R male mice ..................................... 80
Table 3-9 Rotarod performance in H304R/R female mice .................................. 80
Table 4-1 Tail flick data ...................................................................................... 91
Table 4-2 Total fascicular area in the sciatic nerve of H304R male mice ........... 97
Table 4-3 Axon counts inside the largest fascicle ............................................. 100
Table 4-4 Optimized conditions for identifying DRG neuronal subpopulations . 104
Table 4-5 Preliminary counts of wildtype DRG subpopulation .......................... 105

xix
Table 5-1 Synaptophysin-RFP positive vesicles motility in hippocampal neurons
at DIV 10 .......................................................................................................... 113
Table 5-2 Synaptophysin-RFP positive vesicles motility in hippocampal neurons
at DIV 14 .......................................................................................................... 115
Table 5-3 Synaptophysin-RFP positive vesicles motility in cortical neurons at DIV
10 ..................................................................................................................... 118
Table 5-4 Synaptophysin-RFP positive vesicles motility in cortical neurons at DIV
14 ..................................................................................................................... 120
Table 5-5 Rab7-GFP positive vesicles motility in cortical neurons at DIV 10 ... 123

xx
LIST OF ABBREVIATIONS
AAA+ - ATPase Associated Family of proteins
ATP – Adenosine Tri Phosphate
ALS – Amyotrophic Lateral Sclerosis
CMT – Charcot Marie Tooth
CMT2O – Charcot Marie Tooth Type 2O
Cra – Cramping
DHC – Dynein Heavy Chain
DIV – days in vitro
DRG – Dorsal Root Ganglia
GLUT1- Glucose transporter 1
H304R – Histidine 304 Arginine
H304R/+ - Heterozygous mutant of Histidine 304 Arginine
H304R/R – Homozygous mutant of Histidine 304 Arginine
H306R – Histidine 306 Arginine
ID – Intellectual Disability
Loa – Legs at odd angle

xxi
MCD – Malfunctions of Cortical Development
PBS – Phosphate Buffered Saline
SMA-LED – Spinal Muscular Atrophy with Lower Extremity Dominant
Swl – Sprawling

1
CHAPTER 1: INTRODUCTION
Dynein is a motor protein present ubiquitously in many living organisms ranging
from small microorganisms (yeast, fungi, parasite etc.) to humans. One of the
many functions of dynein is to carry intracellular cargo on microtubule tracks
(Paschal and Vallee, 1987). Microtubules are polarized cytoskeletal filaments
that are usually arranged with their growing plus ends at the cell periphery and
their minus ends at the microtubule-organizing center (MTOC) at the cell center.
Dynein utilizes adenosine triphosphate (ATP) as an energy source to power its
motor function and carries cellular cargoes from the cell periphery to the minus
end of the microtubule at the cell center (retrograde). In contrast, the motor
protein kinesin carries the cellular cargoes from the cell center to the plus end of
the microtubule that is usually located at the cell periphery (anterograde).
In eukaryotic cells, there are 3 classes of dynein – cytoplasmic dynein 1,
cytoplasmic dynein 2, and axonemal dynein (Wickstead and Gull, 2007) (Yagi,
2009). Cytoplasmic dynein 1 is involved in minus end directed transport within
the cell body and has extensive cellular functions ranging from intracellular
transport to cell division activities (listed in the Table 1-1). Cytoplasmic dynein 2
acts as a motor protein inside cilia/flagella and is involved in intraflagellar
transport of structural components for flagellar assembly (Pazour et al., 1999).

2
Figure 1-1 Intracellular motility inside a cell
Motor proteins dynein and kinesin carry cargoes like mitochondria and vesicles on the microtubule tracks. Dynein is minus end directed motor protein, while kinesin is plus end directed motor protein
Cytoplasmic dynein 2 transports cargoes from the tip to the base of cilia where
the minus end of microtubule is located (Porter et al., 1999). The axonemal
dynein is integrated with the microtubule to make the core structure of cilia called
axoneme (Gibbons and Rowe, 1965). Axonemal dynein is involved in powering
the ciliary beating to make the cilia and flagella motile. Cytoplasmic dynein 1 will
be referred just as ‘cytoplasmic dynein’ or ‘dynein’ from now on in this document.

3
1.1 Cytoplasmic Dynein Structure
Dynein is a large multimeric protein complex that is ~1.2 megaDalton in size. A
functional dynein motor unit (Figure 1.1) is comprised of many subunits – 1 pair
of dynein heavy chains (DHC), 1 pair of dynein intermediate chains (DIC), 2 pairs
of dynein light intermediate chains (DLIC), and 3 pairs of dynein light chains
(DLC). The dynein heavy chain (DHC) monomers dimerize at the N-terminal to
form a heavy chain dimer that interacts with other dynein subunits (DIC, DLIC,
DLC) to form a functional dynein motor complex.
Figure 1-2 Cytoplasmic Dynein Structure
Dynein is a multimeric protein comprising of subunits - dynein heavy chain (DHC), dynein light intermediate chain (DLIC), dynein intermediate chain (DIC) and dynein light chain (DLC). Dynein heavy chain has six ATPase heads and a microtubule (MT) binding domain.
Dynein heavy
chain (DHC)
DLIC
DIC
DLC

4
1.1.1 Cytoplasmic Dynein Heavy Chain Structure
The cytoplasmic dynein heavy chain (DHC) is encoded by the gene dync1h1 and
the peptide is ~532 kiloDalton in size. Cytoplasmic dynein heavy chain (DHC)
gene - dync1h1 is an essential gene. Animals with homozygous dynein null
mutations displayed embryonic lethality at embryonic day 8.5 where the embryo
development couldn’t proceed past the gastrulation stage due to disruptions in
Golgi trafficking and function (Harada et al., 1998). A single DHC monomer is
4646 amino acids long peptide in humans and 4644 amino acids long in mice.
The DHC can be divided into two main domains – N terminal tail domain and C
terminal globular head domain (Figure 1.2). Two monomers of DHC
homodimerize at the N-terminal tail region and interact with other subunits to
form a single functional unit of dynein. The C terminal globular head has 6
repeating AAA+ ATPase units forming a ring structure arrangement (Burgess et
al., 2004) (Sakato and King, 2004).
Figure 1-3 Cytoplasmic Dynein Heavy Chain (DHC) peptide
Dynein heavy chain has two domains – N terminal tail domain and C terminal head domain. There are also six AAA ATPase domains (AAA1-AAA6) that make up the motor dynein ring.
2

5
The six ATPase rings domains (AAA1-AAA6) are responsible for the motor
activity of DHC (Kon et al., 2012). Ring domain AAA1 has the p-loop motif that
can bind and hydrolyze ATP to generate energy for dynein motor activity
(Gibbons et al., 1991). Domains AAA2-AAA4 has p-loop motifs and ability to bind
ATP, whereas domains AAA5-AAA6 lack ATP binding sites. Overall, domains
AAA2-AAA6 lack ATP hydrolysis activity but do provide structural integrity and
regulate functions of the dynein motor ring (Mocz et al., 1991) (Neuwald et al.,
1999). A coiled-coil ‘stalk’ region is located between AAA4-AAA5 in the motor
ring and contains the microtubule binding domain (MTBD) of dynein heavy chain
at its tip (Koonce, 1997). ‘Linker’ domain lies across the six ATPase rings and it
changes its conformation based on the ATP binding state of dynein (Roberts et
al., 2012).
The cytoplasmic dynein undergoes a mechanochemical cycle where mechanical
conformational changes and chemical transitions regulate the motor activity. A
typical mechanochemical cycle of dynein consists of the following steps
(Burgess et al., 2003; Imamula et al., 2007; Kon et al., 2005) -
1. In the initial state, dynein is attached to the microtubule track via the
microtubule binding domain (MTBD) in the stalk region.
2. When an ATP molecule binds to the AAA+ ATPase domain of dynein
heavy chain, the MTBD of the stalk detaches from the microtubule.
3. Detachment of stalk from the microtubule causes a conformational change
in the linker domain and linker gets rotated.

6
4. Upon ATP hydrolysis, MTBD of stalk binds to a new position on the
microtubule and that leads to the release of ADP and Pi (inorganic
phosphate molecule).
5. Released Pi causes linker domain to straighten and generate a
“powerstroke” that extends dynein tail along with its cargo to move further
along towards the minus end of the microtubule track.
1.2 Cytoplasmic Dynein Function
Dynein has variety of roles and functions to play in a healthy functioning cell.
Dynein is involved in both cell division and interphase activities of the cell. Dynein
moves its cargoes on the microtubule in a “stop and go” manner. If the same
cargo has dynein and kinesin attached to it, then the transport of the cargo can
momentarily stop and the travel direction can get switched sometimes from the
retrograde to anterograde direction inside the cell (Steffen et al., 1997).
Sometimes the role of dynein is also distinct in different organisms – for example,
dynein transports vesicles in filamentous fungi (Sivagurunathan et al., 2012)
whereas dynein is responsible only for mitotic spindle positioning in yeast (Moore
et al., 2009). Even viruses utilize dynein driven transport of endocytotic vesicles
to reach the nucleus to replicate itself using the host machinery (Dodding and
Way, 2011). These functions of dynein are broadly categorized and summarized
in Table 1-1.

7
It is clear that cytoplasmic dynein is involved in many cellular functions that are
essential for the cell growth, maintenance, and survival. Total loss of dynein
function causes early embryonic lethality in Drosophila melanogaster (Dick et al.,
1996). Heterozygote dynein heavy chain knockout mice did not exhibit any
phenotype, however, the dynein-null homozygous mice were embryonic lethal
and did not survive past the gastrulation stage (Harada et al., 1998).

8
Table 1-1 Cellular functions of cytoplasmic dynein
Category Cargo Cellular functions References
Org
an
elle
tra
nsp
ort
Vesicles Transport of vesicles (Sivagurunathan et al., 2012),
(Koonce, 2000)
Early endosomes Receptor sorting and Rab5 positive early
endosome morphogenesis in HeLa cells
(Driskell et al., 2007)
Peroxisomes Coordinated movement of peroxisomes in
cultured Drosophila melanogaster cells
(Kural et al., 2005)
Melanosomes Regulation of melanosomes transport in
Xenopus melanophore cell line
(Gross et al., 2002)
Golgi vesicles Transport of vesicles from endoplasmic
reticulum to Golgi in mammalian cell lines
(Presley et al., 1997),
(Hoogenraad et al., 2001)
Mitochondria Transport of mitochondria in motor axons of
Drosophila melanogaster and in neurons
(Pilling et al., 2006),
(Schwarz, 2013)
Lysosomes and
late endosomes
Rab7-interacting lysosomal protein (RILP)
recruit dynein to Rab7 positive lysosomes and
late endosomes in mammalian cells
(Jordens et al., 2001),
(Cantalupo et al., 2001),

9
Category Cargo Cellular functions References
Recyclin
g a
nd
de
gra
datio
n
Phagosomes Dynein and dynactin regulate the maturation
and minus end directed motility of phagosomes
in mammalian cells
(Blocker et al., 1997)
Autophagosomes Maturation of autophagosomes (containing
organelles and soluble proteins for targeted
degradation) is regulated spatially by dynein in
primary neurons
(Maday et al., 2012)
Aggresomes Dynein drives the misfolded protein into
aggresomes for sequestration and eventual
degradation
(Johnston et al., 2002)
Cell
div
isio
n
Nuclear
positioning
Dynein positions nucleus in budding yeast
Saccharomyces cerevisiae during cell division
(Moore et al., 2009)
Spindle
positioning
Cortical dynein pulls and displaces microtubules
for mitosis and meiosis
(McNally, 2013)
Centrosomes Dynein transports and assembles gamma
tubulin and pericentrin at the centrosomes for
microtubule nucleation & organization
(Young et al., 2000)
Chromosome
separation
Dynein associated with the nuclear envelope
drives separation of chromosomes in prophase
(Raaijmakers et al., 2012)

10
Cyto
ske
leto
n &
pro
tein
tra
nspo
rt
Nuclear rotation Dynein rotates and directs the nucleus in the
leading edge of motile fibroblasts
(Levy and Holzbaur, 2008)
Microtubules Dynein maintains stable microtubule and
kinetochore orientation for aligning
chromosomes during metaphase
(Varma et al., 2008)
Neurofilaments Direct interaction and translocation of
neurofilaments by dynein in neuronal cells
(Shah et al., 2000)
Transcription
factors
Retrograde transport of transcription factor such
as glucocorticoid receptor from the cytoplasm to
the nucleus
(Harrell et al., 2004)
Ribonucleoprotein Correct mRNA localization by dynein mediated
apical transport in Drosophila melanogaster
(Wilkie and Davis, 2001)
Vira
l tr
an
spo
rt
HIV‐1 and HSV1 Dynein is hijacked by the viruses such as
Human immunodeficiency virus type 1 (HIV‐1)
and Herpes simplex virus (HSV1) to reach the
nucleus of the host cells for replication
(Dodding and Way, 2011)
Adenovirus Adenovirus capsid hexon directly interacts with
dynein for viral transport to the nucleus
(Bremner et al., 2009)
Category Cargo Cellular functions References

11
1.2.1 Dynein interactions with its binding partners
There are three families of cytoskeletal motor proteins inside a cell – microtubule-
based dynein, microtubule-based kinesin, and actin-based myosin. Multiple
subfamilies and isoforms of kinesins and myosin are available inside the cells to
perform distinct cellular functions (Vale, 2003). However, dynein has only three
classes inside the cell as mentioned previously – cytoplasmic dynein 1,
cytoplasmic dynein 2, and axonemal dynein. Cytoplasmic dynein 2 and axonemal
dynein are dedicated to ciliary and flagellar functions specifically. Therefore,
cytoplasmic dynein 1 (referred as dynein from now on) is solely responsible for
multiple cellular functions in all cell types besides cilia/flagella. It is no wonder
that dynein utilizes and interacts with various adaptor proteins to carry out
diverse cellular functions (Vallee et al., 2012).
Binding of dynein with its interacting partners controls different cellular functions
and is needed to load the cargo onto the motor complex. Dynein cargoes range
from membranous vesicles, organelles, proteins, cytoskeletal components and
even viruses. Furthermore, an active motor complex facilitates varied dynein-
based functions such are organelle transport and positioning (Burkhardt et al.,
1997), mitotic chromosome separation (Dujardin et al., 1998), and nuclear
migration (Levy and Holzbaur, 2008). Dynein interacts with many intracellular
binding partners via direct or indirect interactions (Figure 1-4). DHC participates
in the catalytic motor activity and other dynein subunits like LCs, ICs, LICs

12
interact with adaptor proteins to regulate various dynein functions in the cell
(Pfister, 2015).
The direct binding partners of dynein include nuclear distribution protein E
(NudE) (McKenney et al., 2011), lissencephaly (LIS 1) (Huang et al., 2012),
dynactin (dA) (Schroer, 2004), RAB-interacting lysosomal protein (RILP)
(Schroeder et al., 2014), Huntingtin protein (Htt) (Caviston and Holzbaur, 2009),
Snapin on late endosomes (Cai et al., 2010), adenoviral hexon protein (Bremner
et al., 2009), and pericentrin centrosomal protein (Tynan et al., 2000).
Dynactin regulates dynein driven motility by enhancing dynein’s binding with the
microtubule and making the transport more processive (Culver-Hanlon et al.,
2006). Additionally, dynactin also acts as an cargo adaptor for dynein and helps
dynein link with multiple cargo adaptors such as - Ankyrin 2 (AnkB) on
membranous cargoes (Lorenzo et al., 2014), RILP on Rab7 positive late
endosomes (Jordens et al., 2001), c-Jun N-terminal kinase-interacting proteins
(JIPs) on neuronal cargoes (like amyloid precursor protein) (Fu and Holzbaur,
2013), Huntington-associated protein 1 (HAP1) on neuronal vesicles (Engelender
et al., 1997), Spectrin on Golgi vesicles & late endosomes/lysosomes (Holleran
et al., 1996), bicaudal D/BICD2 on Golgi-ER vesicles (Liu et al., 2013; Matanis et
al., 2002), and TRAK/Milton adaptor proteins on mitochondria (van Spronsen et
al., 2013).

13
Figure 1-4 Binding partners of dynein
Dynein interacts with various cargo adaptors either directly (shown in blue arrows) or indirectly (shown in orange arrows) to achieve versatility in its cellular functions. Dynein heavy chain (DHC), dynein light intermediate chain (DLIC), dynein intermediate chain (DIC) and dynein light chain (DLC).
DHC
DLIC
DIC
DLC
Dynactin BicD, RILP, JIP, HAP1, AnkB, Spectrin, TRAK/Milton
Adenoviral hexon protein, pericentrin, RILP,
NudE/NudEL Lis1
Lis1
DIC Htt, Snapin
Direct interaction with dynein
Indirect interaction with dynein

14
1.2.2 Dynein mutations linked to neurodegeneration in humans
Neurons are highly polarized cells with axonal projections as long as up to a
meter in length. Consequently, it is understood that neurons need robust axonal
transport for survival. Dynein has an established role in retrograde transport of
cargoes from the neuronal tip to soma in neurons (Perlson et al., 2010).
Additionally, dynein interacts with dynactin to form a highly processive motor
complex (Culver-Hanlon et al., 2006) that has an important role in axonal
transport (Waterman-Storer et al., 1997). The dynein/dynactin motor protein
complex transports cargo from the tip of the axon towards the neuronal cell body
(retrograde direction). Other functions of dynein in neurons includes growth and
development of neurites (Barakat-Walter and Riederer, 1996), formation of
synapse in neurons (Cheng et al., 2006), axonal transport of microtubules &
neurofilaments (He et al., 2005) and intracellular organelles (Schnapp and
Reese, 1989), and neuronal migration inside the cortical layer of the brain
(Sasaki et al., 2000).
Seeing how crucial dynein is for neuronal growth and maintenance, it would not
seem very surprising to see a dynein heavy chain mutation leading to
neuropathies in humans (figure 1-5). The first dynein heavy chain mutation
H306R was reported in 2011, that lead to a hereditary motor-sensory
neurodegenerative disease called Charcot Marie Tooth Disease Type 2O in
humans (Weedon et.al. 2011). Since then at least 17 genetic mutations spanning
the motor and tail domains of the dynein heavy chain have been identified in

15
neurodegenerative diseases in humans (summarized in Table 1-2). The reported
dynein heavy chain mutation related diseases in humans can be broadly
classified as motor-sensory neuropathies, central nervous system deficits, and
congenital defects.

16
Figure 1-5 Mutations in Dynein Heavy Chain (DHC)
Mouse mutations (top, in blue text) and human mutations (bottom, in red text) in the DHC peptide. Mice mutations are reported in the tail domain of the DHC peptide, whereas the human mutations span the entire length of the DHC peptide.

17
Table 1-2 Dynein heavy chain mutations in humans
Disease DHC mutation Clinical symptoms Reference
Charcot Marie Tooth
Type 2O (CMT2O)
H306R muscle weakness and wasting, gait
abnormalities, skeletal deformities,
motor milestone delays, loss of
sensation
(Weedon et al.,
2011)
Spinal Muscular Atrophy
– Lower Extremity
Dominance (SMA-LED)
I584L, K671E, Y970C,
G1132E
Severe muscular atrophy in legs,
loss of motor neurons, delay in
motor milestones, waddling gait
(Harms et al.,
2012)
Malformation of Cortical
Development
deletion 659-662, K129I,
K3336N, R3384Q, R1567Q,
R3344Q, R1962C, K3241T,
R3344Q
Neuronal migration defects in
cerebellar cortex, microcephaly or
macrocephaly, polymicrogyria,
epilepsy/seizure in severe cases
(Poirier et al.,
2013)

18
Disease DHC mutation Clinical symptoms Reference
Intellectual disability E1518K, H3822P Defects in neuronal migration,
cortical development malfunctions,
impaired speech, reduced
intellectual and mental functions,
gait problems
(Willemsen et al.,
2012)
Congenital cataracts and
gut dysmotility
R2332C Cataract at birth, oral dysphagia,
dysmotility in gut, developmental
milestone delay, bifrontal
polymicrogyria
(Gelineau-Morel
et al., 2016)

19
It is interesting to note that even though these mutations lie in the same DHC
gene, they result in different clinical manifestations in humans. The DHC motor
domain mutations generally lead to cortical development type of defects,
whereas, the DHC tail domain mutations were observed to cause motor-sensory
type of defects in the reported human neuropathies. Moreover, it is important to
note that all dynein mutations in humans exhibit symptoms that are heavily
related to neuronal defects rather than any other type of defects in humans. This
suggests that dynein must have a specialized role in the neurons because even
the smallest single point mutation in dynein can disrupt the regular functions of
the central or peripheral nervous system. Hence, it is evident that mammalian
neurons must be particularly sensitive to the dynein mutations. It would be
valuable to understand how dynein heavy chain mutations cause diseases
specifically of the nervous systems in humans.

20
1.3 Charcot-Marie-Tooth Diseases in Humans
Charcot-Marie-Tooth (CMT) disease is the most common type of hereditary
motor-sensory neuropathy in humans with estimated prevalence rate is 1 in 2500
humans. CMT is characterized by symmetric distal limb weaknesses, muscular
atrophy in hands and/or feet, loss of sensation in lower limbs that may lead to
foot ulcerations, and skeletal defects like pes cavus (high arching of feet) (Figure
1-6).
More than 80 genes that are involved in neuronal structure maintenance signal
transduction, and axonal transport are reported to have mutations that are linked
with CMT (Bird, 1993). CMT patients are categorized into different types (table 1-
3) based on their nerve conduction velocity (NCV) and family history-based
inheritance of the disease. In each category, letters are assigned to represent the
gene involved. For example – CMT1A, CMT1B, CMT1C and so on, CMT2A,
CMT2B, CMT2C and so on. CMT1A indicates CMT Type 1A which is caused by
a mutation in PMP22 gene (myelination gene) (Patel et al., 1992) whereas
CMT1B indicates CMT Type 1B which is caused by a mutation in MPZ gene
(another myelination gene).
CMT is clinically diagnosed by molecular genetic testing and by identifying
peripheral neuropathy symptoms during their physical examinations and in their
previous medical history. CMT has a variable onset in patients, ranging from as
early as age at birth to as late as age 50. Currently there is no cure for CMT, and
the treatment is based on the clinical symptoms presented by the patients. CMT

21
patients often have to rely on surgical treatments, physical therapy, and use of
prosthetics like crutches/ankle braces to cope with the symptoms. About 5% of
all CMT patient are dependent on wheelchair for mobility.
Another challenge with CMT is the heterogeneity of the disease onset,
progression, and clinical symptoms even in related patients from the same family
pedigree. For example- in the Dominant Intermediate Charcot-Marie-Tooth
disease (DI-CMT), some patients from the same family may show demyelination
associated symptoms, whereas other patients from the same family may show
axonal degeneration, and some may even show both demyelination and axonal
demyelination form of the CMT symptoms.
Figure 1-6 Clinical symptoms associated with Charcot Marie Tooth (CMT)
CMT patients primarily show distal lower limb muscle wasting and atrophy, foot deformity (pes cavus), foot ulcerations, and gait abnormalities.

22
Table 1-3 Types of Charcot-Marie-Tooth disease in humans
Type of CMT Type of defect Inheritance Some examples of mutations (Bird, 1993)
CMT Type 1
(most common)
Demyelination with
reduced NCV of <35m/s
Autosomal
dominant
PMP22, MPZ, LITAF, ERG2, NEFL
CMT Type 2 Axonal degeneration with
normal NCV>45m/s
Autosomal
dominant
MFN2, RAB7, TRPV4, GARS, NEFL, MPZ,
DYNC1H1, GDAP1, HSPB8, LRSAM1, MORC2,
DI- CMT
(Dominant
Intermediate)
Demyelination and axonal
degeneration with NCV of
35-45m/s
Autosomal
dominant
GDAP1, MPZ, DNM2, DRP2, GNB4, INF2,
PLEKHG5, YARS,
CMT Type 4
(rare)
Demyelination and axonal
degeneration with NCV of
35-45m/s
Autosomal
recessive
GDAP1, MTMR2, SBF2, SH3TC2, NDRG1,
EGR2, PRX, FGD4, FIG4, HINT1
CMT Type X Demyelination and axonal
degeneration with NCV of
35-45m/s
X-linked DNM2, MPZ, YARS, GJB1

23
1.3.1 Dynein mutation linked to CMT2O and SMA-LED
A dynein heavy chain mutation linked to Charcot-Marie-Tooth type 2O (CMT2O)
in humans was first identified in 2011 (Weedon et al., 2011). Exome sequencing
of genes in a four generation European family identified a single point missense
substitution of Histidine to Arginine at position 306 (H306R) in cytoplasmic dynein
heavy chain 1 peptide (figure 1-7). The autosomal dominant H306R mutation
affected 23 members (male and female) in this family and they showed a varied
range of clinical symptoms.
Figure 1-7 CMT2O and SMA-LED linked mutation in dynein heavy chain
Missense substitution mutation (Histidine to Arginine) at position 306 in human cytoplasmic dynein heavy chain peptide has been linked to two distinct neuropathies – Charcot Marie Tooth type 2O (CMT2O) and spinal muscular atrophy with lower extremity dominant (SMA-LED).
CMT2O patients with autosomal dominant heterozygous H306R mutation have
varied disease onset ranging from early childhood to early adulthood. CMT2O
patients reportedly had delayed motor developmental milestones, problems in
walking or running, recurrent falls, slow progressing muscle weakness and
muscle wasting in lower legs, and skeletal deformities pes cavus (high arching of

24
foot). Physiological tests with CMT2O patients revealed normal nerve conduction
velocities, evidence for axonal degeneration in sural nerve biopsies, and
denervation defects in muscle biopsies. Several patients that were severely
affected with CMT2O also reported to have neuropathic pain and prickling
sensation (paresthesia) in their lower limbs. A few patients also reported loss of
reflexes and sensation, speech delays & behavioral problems, lumbar lordosis,
spinal and hip issues leading to waddling gait, and loss of proximal muscles in
the upper back (periscapular weakness).
In parallel, an independent study of a Japanese family with the same H306R
mutation presented clinical symptoms that were consistent with spinal muscular
atrophy with lower extremity dominant (SMA-LED) (Tsurusaki et al., 2012). The
patients in this family showed clinical features that were different from CMT2O
patients, such as proximal lower limb weakness instead of distal limb weakness
and non-progressive nature of muscular atrophy/wasting. SMA-LED patients with
H306R mutation showed motor neuron disease involvement but not sensory
neuron involvement. It is interesting to note that a family member in the
European family with CMT2O showed proximal limb atrophy like SMA-LED
patients in this Japanese family. In conclusion, the single point mis-sense
substitution of H306R in cytoplasmic dynein heavy chain has been reported to
cause to two separate neuropathies in humans – CMT2O and SMA-LED.

25
1.4 Mouse Models with Neurodegeneration Linked Dynein Mutations
There are three reported mouse models with DHC mutations – Legs at odd
angles (Loa), Cramping (Cra), Sprawling (Swl) (figure 1-8). These mice were
named after their abnormal hind limb clenching phenotype and their posture.
Heterozygotes of all three mouse models had normal lifespan, whereas the
homozygotes showed severe phenotype & did not survive embryonically or after
birth (Chen et al. (2007); (Hafezparast et al., 2003).
Loa and Cra mice were generated independently via N-ethyl-N-nitrosourea
(ENU) treated mice and were identified due to abnormal clenching of hind limbs
when lifted by tail (Hrabe de Angelis et al., 2000). Swl was created via radiation-
induced mutagenesis, also displayed abnormal clenching of hind limbs (Duchen,
1974). All three Loa, Cra, and Swl are autosomal dominant mutations that lead to
neurodegenerative phenotypes. Loa/+ and Cra/+ developed late-onset of motor
neuropathy, whereas Swl/+ showed early-onset of hereditary sensory
neuropathy. The key features of the three DHC mutant heterozygous mice are
listed below in table 1-4.

26
Figure 1-8 Dynein heavy chain (DHC) mutations in mice
Mouse mutations in the DHC peptide are reported in the tail domain and are associated with Loa, Cra, and Swl mice.

27
Table 1-4 Dynein heavy chain mutations in mice
DHC mutation Phenotype
Legs at odd angles (Loa) –
F580Y
(Hafezparast et al., 2003)
Loa/Loa homozygous lethal
late-onset motor neuron
degeneration in Loa/+
gait abnormalities
reduced front and hind limb strength,
thinner dorsal root and sciatic nerve
abnormal proprioception & absence of
H-reflex
lumbar DRG neurons loss
mild loss of spinal α- motor neurons
muscle spindle loss
neuronal migration defects
Cramping (Cra) – Y1055C
(Hafezparast et al., 2003)
Cra/Cra homozygous lethal
late-onset sensory neuron
degeneration in Cra/+
gait abnormalities
reduced hind limb strength
thinner dorsal root
abnormal muscle pathology,

28
mild loss of spinal α- motor neurons
behavioral hyperactivity
mitochondrial dysfunction in skeletal
muscles
Sprawling (Swl) –
[GIVT]1040[A]
(Chen et al., 2007)
Swl/Swl homozygous lethal
early-onset hereditary sensory
neuropathy in Swl/+
gait abnormalities
reduced hind limb strength,
thinner dorsal root and sciatic nerve
abnormal proprioception & absence of
H-reflex
lumbar DRG neurons loss
muscle spindle loss
To better understand the role of dynein in neurodegeneration, Loa/+, Cra/+, and
Swl/+ heterozygote mice were separately cross bred with the mutant SOD1G93A
mouse model of amyotrophic lateral sclerosis (ALS). ALS is a fast progressing
neurological disease marked by progressive loss of motor neurons, leading to a
total paralysis and lethality 3-5 years after onset. The gain of function mutation
G93A in superoxide dismutase 1 (SOD1) is autosomal dominant and kills motor

29
neurons rapidly (Valentine et al., 2005). It was interesting to note that when Loa/+
and Cra/+ were crossed separately with ALS SOD1G93A mice, their double
heterozygotes pups showed attenuation of the ALS SOD1G93A symptoms and
improved lifespan (Kieran et al., 2005) (Teuchert et al., 2006). This was a very
unsuspected finding and opposite of what was expected at the time. Loa/+ and
ALS SOD1G93A double heterozygotes also showed a remarkable increase in its
retrograde axonal transport. Swl/+ and SOD1G93A heterozygotes did not show any
attenuation or improvement of ALS SOD1G93A associated phenotype. It is
interesting to note that even though not all three mutations lie in the same tail
region of DHC, their attenuation response to ALS mutant SOD1G93A was the
same.
Undoubtedly, the three dynein heavy chain mutant mice (Loa, Cra, Swl) provided
many key insights into understanding the dynein function in neurodegenerative
disorders. However, Loa, Cra, and Swl mutations are not linked to any disease
alleles in humans and it is difficult to correlate their findings to human
neuropathies. Nevertheless, it would be very interesting to compare Loa, Cra,
and Swl mice studies with other DHC mice models that have dynein mutations
linked to an actual human disease.

30
1.5 Peripheral Nervous System of Mouse
The peripheral nervous system of mouse comprises of nerves and ganglia
arising out of the spinal cord. The dorsal root ganglia (DRG) is a cluster of
sensory neuron cell bodies located in pairs in the spinal column of mice that start
forming in the embryo at embryonic day 13 (E13) (Lawson and Biscoe, 1979).
The sensory neurons located in the DRGs are pseudo-unipolar in nature and do
not have dendrites. The DRG sensory neurons have a single axonal projection
that branches into two – peripheral and central branches (figure 1-9). The
peripheral branch brings in the nerve impulses from the innervated tissues (skin
or muscle) to the neuronal cell body located in the DRG. The central branch
carries the impulse away from the soma to the central nervous system i.e. the
spinal cord or brain. Both branches are myelinated and carry the nerve impulses
towards the central nervous system (shown by the arrows in figure 1-9).
Figure 1-9 Pseudo-unipolar sensory neurons in DRGs
Sensory neurons in the Dorsal Root Ganglia (DRG) are pseudo-unipolar in nature, i.e. it has one cell body and two functional axonal branches – peripheral and central branches.

31
1.5.1 Role of dynein mediated retrograde signaling in sensory neurons
Growth and maturation of sensory neurons inside the DRGs are dependent on
various neurotrophic factors such as nerve growth factor (NGF), glial derived
neurotrophic factors (GDNF), brain derived neurotrophic factors (BDNF), and
neurotrophins (NTs) (Kimpinski et al., 1997) (Markus et al., 2002) (Klusch et al.,
2018). The neurotrophic factors are secreted by the target tissue (muscle, skin,
or visceral organ) and stimulate the development and function of sensory
neurons (figure 1-10).
Figure 1-10 Neurotrophic factors in sensory neurons development
Sensory neurons and motor neurons growth, maturation and survival depends on the neurotrophic factors such as neurotrophins (NTs) and glial derived growth factors (GDNF) released by the target tissue they innervate.
The neurotrophic factors bind to the tyrosine kinases (Trk) receptors located on
the axonal terminals of the peripheral branch of the sensory neuron (table). NGF

32
specifically binds to TrkA receptors, NT4 and BDNF specifically bind to TrkB
receptors, and NT3 specifically binds to the TrkC receptors (figure 1-11).
Figure 1-11 Neurotrophic factors activate Trk signaling in neurons
Neurotrophic factors bind to a specific Trk receptor and activate downstream signaling of neuronal maintenance and survival factors. NGF= Nerve Growth Factor, BDNF= Brain Derived Neurotrophic Factors, NT3/4= Neurotrophins 3/4.
Binding of neurotrophic factors leads to dimerization of the receptors and their
autophosphorylation that recruits the effectors for downstream Trk signaling. The
activated Trk signals send survival factors such as mitogen-activated protein
kinase (MAPK); phosphatidylinositol 3-kinase (P13K); phospholipase (PLC-γ)

33
etc. (Chao, 2003) (Delcroix et al., 2003) to the neuronal cell body located in the
DRG via dynein mediated retrograde transport (Heerssen et al., 2004) (Ito and
Enomoto, 2016).
Neurotrophin mediated retrograde signaling also facilitates differentiation and
specification of neuronal subpopulations in the peripheral nervous system of
mice. Studies with BAX-/- and TrkA-/- double mutant mice lacking the NGF
mediated TrkA signaling showed an absence of calcitonin gene related peptide
(CGRP) expressing neuronal subpopulation in the mice DRGs (Suzuki et al.,
2010). Other studies have explored and established the role of target derived
neurotrophic factors (such as neurotrophins & BDNF) in determining the
cholinergic/peptidergic phenotype of the sensory neurons during their
development and differentiation in mice (Bergmann et al., 1997; Cellerino et al.,
1996).
Several in vitro studies have investigated and established the role and
importance of retrograde transport of neurotrophic factors. Neurotrophic factors
released by the target tissues bind to the specific receptors in the terminal ends
of the sensory axons innervating the tissue. Then the neurotrophic factors
mediated signaling cascade is activated at the axonal tip and it travels all the way
to the neuronal cell body in retrograde fashion.
Classical experiments with special compartmentalized cell chambers called
Campenot chambers were performed to investigate the spatial effect of

34
neurotrophin Nerve Growth Factor (NGF) on axonal growth (Campenot, 1977).
First, NGF was only supplied selectively to the compartment in the Campenot
chamber that had distal ends of axons growing in it. It was observed that axonal
growth was maintained even though the neuronal cell bodies did not receive
NGF. However, when the NGF was applied only to the Campenot compartment
containing neuronal cell bodies, the distal axons started retracting and
degenerating. These key experiments established that NGF mediated
neurotrophin signaling at the distal ends of neurons were required and adequate
for axonal growth and maintenance.
Subsequent studies then established that neurotrophins bound to the receptors
are internalized and packaged inside signaling endosomes for retrograde
transport to the neuronal cell bodies (Wang et al., 2016). Further investigation of
retrogradely transported signaling endosomes in rat sciatic nerves revealed that
these vesicles contained not only neurotrophin NGF and its receptor TrkA, but
also characteristic markers of early endosomes such as early endosome antigen
1 (EEA1) and Rab5 (small G-protein) (Deinhardt et al., 2006). Additionally,
components of mitogen-activated protein kinase (MAPK) and phosphatidylinositol
3-kinase (P13K) pathways were also found inside the signaling endosomes. The
same study also showed the evidence of dynein dependent retrograde transport
of these NGF-TrkA containing endosomes by colocalization, biochemical, and
nocodazole microtubule disruption assays. Several other independent studies
have also established the role of dynein in retrograde transport of neurotrophic

35
factors – colocalization of 14kDa Dynein Light Chain (DLC) with Trk receptors
(Yano et al., 2001), Rab7 positive vesicles in NGF-TrkA signaling (Saxena et al.,
2005), and p75 and TrkB signaling (Deinhardt et al., 2006).
In conclusion, dynein dependent retrograde signaling is necessary for the
development, specification and maintenance of sensory neurons and it is not
surprising that dynein disruption could cause sensory neuropathies. Numerous
studies have shown that disrupted neurotrophic factor mediated Trk signaling
causes severe sensory neuropathies in mice (Smeyne et al., 1994) (Patel et al.,
2000) (Sleigh et al., 2017).
1.5.2 Sciatic Nerve anatomy in mice
The sciatic nerve is the largest nerve innervating the hind limbs of mice. The
sciatic nerves arise from the sciatic notch near the spine, then continues all the
way from lumbosacral region to popliteal fossa (knee cavity) where it splits into
peroneal, sural and tibial nerves to innervate the lower hind limbs in mice (figure
1-12). The majority of sciatic nerves in C57BL/6J strain mice are made up of the
third lumbar (L3) and fourth lumbar (L4) spinal nerves and a lesser contribution is
received from the fifth lumbar (L5) spinal nerve (Rigaud et al., 2008). The
percentage contribution from each spinal nerve in a sciatic nerve is roughly about
L3 (~28%), L4 (~56%) and L5 (~16%) and no spinal nerve contribution is
reported from L6 at all.

36
Figure 1-12 Sciatic nerve anatomy in mice
Spinal nerves arising from lumbar vertebrae L3, L4, and L5 combine together to form a sciatic nerve in mice that splits up into peroneal, tibial, and sural nerves to innervate the lower hind limbs.
Because the sciatic nerve contains both motor and sensory neurons, it is also
regarded as a “mixed nerve” (Figure 1-13). Dorsal root supplies the sensory
neurons (afferent fibers) and ventral root (efferent fibers) supplies the motor
neurons at lumbar L3, L4, and L5 in mice to make up the “mixed” sciatic nerve in
mice. The sensory nerves bring in the afferent sensing signals from the periphery
target tissue to the spinal cord for processing. Sensing signals can be sensing
temperature, pain stimulus, touch, vibrations etc. In contrast, the motor neurons
send out the efferent action signals from the spinal to the target tissues. Action
signals can be voluntary or involuntary action of a muscle group to a stimulus.
Knee jerk test is a classic example of a reflex arc where the reflex hammer hits

37
the knee (stimulus) and the body reacts quickly by moving your leg away (action)
through rapid involuntary muscle contraction.
Figure 1-13 Sciatic nerve is a mixed nerve
Sciatic nerve has both sensory and motor neurons and is called a “mixed nerve”. Both dorsal and ventral roots supply spinal nerves to make up the sciatic nerve.
1.5.3 Dorsal Root Ganglia anatomy in mice
Neural crest migration gives rise to a cluster of sensory neurons called dorsal
root ganglia (DRG). There are 62 DRGs in total and they are located in pairs
along the spinal cord – 8 pairs in cervical, 13 pairs in thoracic, 6 pairs in lumbar
and 4 in sacral region in the C57BL/6J strain of mice (Malin et al., 2007). There
are many types of heterogeneous somatosensory neuronal subpopulations
present in the DRGs- nociceptors, mechanoreceptors, thermoreceptors, and
proprioceptors (table 1-4). Somatosensory neurons transmit sensory signals to

38
the spinal cord and brain from the target tissues that they innervate such as
muscle spindles, skin, and organs (Liu and Ma, 2011).
Table 1-5 Major types of somatosensory neurons in the DRG
Type of the neuron Size of the fiber Stimulus received
Nociceptors Small diameter (c-fibers) Pain stimulus
Mechanoreceptors Medium diameter (Aδ-
fibers)
Pressure, touch and
vibration
Thermoreceptors Small diameter (AΔ-fibers) Temperature (cold
and warm)
Proprioceptors Large diameter (Aβ-fibers) Muscle stretch and
orientation
Different somatosensory neurons express different types of receptor. Nociceptive
neurons have small sized diameter axons and express the TrkC receptors for
Nerve Growth Factor (NGF) (Suzuki et al., 2010). In addition, peptidergic
nociceptive neurons express calcitonin gene-related peptide (CGRP), whereas,
non-peptidergic nociceptive neurons express IsolecithinB4 (IB4) on its plasma
membrane (Bennett et al., 1996). Chemical stimuli such as capsaicin have been
reported to also stimulate responses in nociceptors expressing capsaicin
receptors called transient receptor potential cation channel subfamily V member
1 (TRPV1) channels (Amaya et al., 2004). Proprioceptive neurons have large

39
sized diameter axons and express TrkC receptors for neurotrophin 3 (NT-3)
uptake.
It is highly common for researchers to investigate the neuronal subpopulations in
the DRGs of mice exhibiting any sensory neurodegenerative symptoms. For
example - in sprawling (Swl) mice, it was reported that the heterozygous Swl/+
mice showed specific loss of its proprioceptive neurons and not in the nociceptive
neuron population in the lumbar dorsal root ganglia (Chen et al., 2007). As a
result, Swl/+ mice showed the corresponding phenotype of early-onset of
proprioceptive sensory neurological defects. In conclusion, investigation of sciatic
nerve anatomy and DRG neuronal subpopulation can provide key insights in
understanding molecular mechanisms behind sensory neuropathies.

40
CHAPTER 2: MATERIALS AND METHODS
2.1 Generation of H304R mouse model
The knock-in H304R mouse model was generated using gene targeting in mouse
embryonic cells (figure 2.1). Using 3-step PCR mutagenesis, the CMT2O disease
linked dynein mutation (CAC to CGC at position 304) was introduced in the
targeting vector with Neo cassette. Presence of the mutation was confirmed by
sequencing the PCR amplified targeted region. Then embryonic stem cells were
electroporated to take in the targeting vector. Embryonic stem cell clones with the
correct targeted single point mutation were confirmed using resistance selection
markers, PCR screening, and southern blot analysis. The embryonic stem cells
were then microinjected in C57BL/6 mouse blastocysts and implanted into
psuedopregnant foster host female mice. The offspring chimeric mice were
obtained, and cross bred to yield the heterozygous knock-in founder mice. Four
heterozygous founder mice with the desired single point mutation in dynein
heavy chain were outcrossed with B6129SF2/J mice (Jackson laboratories) to
produce a colony of different lineages. Heterozygous male and female mice from
different lineages were bred to obtain homozygous litters of mice.

41
Figure 2-1 Generation of knock-in H304R mouse model
Knock-in H304R mouse model was generated by introducing Neo cassette flanked targeting vector that was microinjected into embryonic stem cells. Chimeric mice were selected to obtain founder heterozygous knock-in founder mice.

42
2.2 Mouse colony maintenance
2.2.1 Mouse colony housing
All mice related procedures were carried out with the University of Central
Florida’s Institutional Animal Care and Use Committee (IACUC) guidance and
approval. The mice were checked daily by the trained vivarium staff and were
immediately treated in case of any health issues. Mice were housed in clear
plastic cages lined with sanitized bedding and were provided ad libitum access to
clean food and water. Mice cages were also provided with red colored housing
tube and a wooden block for enrichment of their surroundings. The cages were
kept in an environment-controlled room at 22 ± 2 °C and atmospheric humidity of
50 ± 10%. Mice were exposed to artificial light from 7:00am-7:00pm and to
darkness at 7:00pm-7:00am on a 12-hour diurnal light cycle.
2.2.2 Mouse colony breeding
Heterozygous H304R mice were repeatedly outcrossed to the B6129SF2/J mice
(Jackson Laboratories, stock#101045). Mice pups remained with their mother for
nursing until they were weaned on post-natal day 21 (3 weeks of age). Tail snips
were collected from the 3-week old pups upon weaning for PCR based
genotyping. Mice tails were also tattooed with unique identification numbers
using Labstamp Tattoo system (Somark Innovations).

43
2.2.3 Measurement of mouse body weight
Body weight of a mouse was recorded (in grams) every 2 weeks before the
mouse performed a behavior test. Bodyweight at 3 month of age were
statistically compared between the genotypes (wildtype, heterozygous,
homozygous) and gender (male and female) separately using One-way ANOVA
(GraphPad Prism 6.0 software).
2.2.4 Mouse health supervision
Mice colonies were kept under daily supervision of trained vivarium staff in case
any medical condition would arise. Mice were regularly monitored for their
posture, alertness, activeness, movement around their cages and consumption
of food and water. Physiological phenotypes of mice such as blood in urination,
and abscess on skins due to excessive scratching were immediately treated and
closely monitored. Mice teeth were routinely trimmed if they showed weight loss
due to inability to feed with longer teeth.
Other litter behaviors like fights with littermates, barbering each other, and
shredding of food were also noted. Phenotype like development of eye tumors,
recurrent seizures, and worsening health were also recorded. We immediately
euthanized any mice that showed any signs and symptoms of pain, suffering,
and unusual declining health.

44
2.3 Behavioral phenotype characterization test
All three groups of mice (wildtype, heterozygous H304R/+, homozygous
H304R/R) performed behavioral tests every two weeks starting from 1 month of
age to 24 months of age or until death (whichever came first). Later time points
such as 15, 18, 21 and 24 months of the longitudinal study yielded similar
phenotypic trend in mice at 12 months of age. Hence, the test readings until 12
months only were averaged in a 3-month bin to give time points of 3 month, 6
month, 9 month, and 12 month for data analysis.
The behavior assay data was collected by the researchers that were blinded to
the genotype of the H304R mice. Mice were genotyped by other lab members
and the mice genotypes were revealed after the completion of the study for data
grouping and statistical analyses. Both testers alternated their turn in testing the
same test mouse every two weeks. The behavior tests - tail suspension, grip
strength, rotarod, and tail flick tests were performed on all three groups of H304R
mice (wildtype, heterozygous, homozygous) as described below.
2.3.1 Tail suspension test
The tail suspension test was performed to assess the postural reflex phenotype
of mice. A mouse was gently lifted by its tail and its positioning of hind limbs were
categorized as 'typical' or 'atypical' tail suspension reflex. In a 'typical' tail
suspension reflex (figure 2-2), the mouse splayed its hind limbs away from the

45
body after being suspended by its tail. Whereas, in 'atypical' tail suspension
reflex, the mouse clenched its hind limbs tightly near its body when suspended
by its tail. The data was collected and recorded from two views (ventral and
dorsal) of the mice to get a better evaluation of the phenotype.
Figure 2-2 Tail suspension test
Mouse is gently suspended by its tail and the hind limb response is categorized as typical (shown here) or atypical (hind limbs clenching together).
The percentage of mice displaying ‘atypical’ reflexes was calculated for all three
groups of mice (wildtype, heterozygous, homozygous) at each time point - 3, 6,
9, and 12 months. Then the ‘atypical’ reflex data were statistically compared
between the wildtype and heterozygous H304R/+ and between wildtype and
homozygous H304R/R mice at each time point using the Fisher’s exact test (two-
tailed distribution, p<0.05) on the GraphPad Prism 6.0 software.

46
2.3.2 Grip strength test
A standard grip test was used to measure the muscular limb strength in mice
using a grip strength meter from Harvard Apparatus (BIOSEB GS3# 76-1068). All
limbs or front limbs of mouse were placed on the metal grid attached to the
readout equipment. Then the mouse was gently pulled across the metal grid
using its base of the tail. The equipment recorded the maximum force exerted (in
grams) by the all limbs or front limbs of the mouse being tested (figure 2-3).
Figure 2-3 Grip strength test meter
Grip strength in limbs is measured using a standard grip strength meter that records the maximum force exerted by the mouse.
A test mouse performed 4 consecutive trial each of all limbs and front limbs grip
test. The 4 trials were averaged for all limbs and front limbs respectively to yield
one single readout each on a test day. Next, the averaged all limbs grip strength
readings and the averaged front limbs grip strength readings of a mouse were

47
combined per each genotype at 3-month time intervals - 3, 6, 9, and 12 months.
The all limbs and front limbs grip strength data were statistically compared
between the wildtype and heterozygous H304R/+ and between wildtype and
homozygous H304R/R mice at each time point using Student’s t-test (two-tailed
distribution, p<0.05) on the GraphPad Prism 6.0 software.
2.3.3 Rotarod performance test
Rotarod test is a standard test used to measure motor coordination, muscle
endurance, and motor balance in mice. We performed rotarod test on our
wildtype, heterozygous, and homozygous mice cohorts using rotarod equipment
from IITC Life Sciences (model# 755). The optimized test parameters were input
into the equipment console and 5 mice were placed individually on a stationary
rod inside 5 different lanes before the start of the test (figure 2-4).
Figure 2-4 Rotarod machine to measure motor coordination
Motor coordination in mice is measured by the rotarod performance test. Five mice can simultaneously run on the rotating rod to perform the rotarod performance test.

48
The rotarod started rotating at 5 RPM (Revolutions Per Minute) and gradually
ramped up to 40 RPM in 60 seconds and stayed constant for next 120 seconds
(figure 2-5). The entire run duration of rotarod was 180 seconds including 60
seconds of ramp time followed by 120 seconds of constant RPM. The equipment
recorded a mouse's fall on the plate below the rod during the 180 seconds total
rotarod run window.
Figure 2-5 Rotarod test profile
The rotarod machine starts at 5 RPM and then ramps up to 40 RPM in 60 seconds. Then it stays at 40 RPM for rest of the rotarod test duration.
A mouse's fall on the plate triggered the magnetic switches at the bottom of the
plate and accurately detected and recorded the latency of fall (in seconds).
Software Series 8 was provided with the equipment and it was used to record the
latency of fall for the mice being tested. Each mouse performed three
consecutive trials of the rotarod test, and all three latency of fall readings were
averaged to yield a single test readout for a mouse at a time point. Next, the

49
averaged latency of fall readings per mouse in the three groups of mice
(wildtype, heterozygous, and homozygous) were combined respectively at 3-
month time intervals - 3, 6, 9, and 12 months. The rotarod data were statistically
compared between the wildtype and heterozygous H304R/+ and between
wildtype and homozygous H304R/R mice at each time point using Student’s t-
test (two-tailed distribution, p<0.05) on the GraphPad Prism 6.0 software.
2.3.4 Tail flick test
Tail flick test was used to quantify the pain sensitivity response of H304R mice to
thermal pain stimulus using Panlab tail flick meter (Harvard Apparatus #
LE7106). A mouse is placed in the restrainer provided with the equipment and
allowed to get habituated inside the restrainer for 2 minutes. Then the mouse tail
is placed under a beam of thermal light and the meter records the time (in
seconds) taken by the mouse to move its tail away from the source of thermal
pain stimulus. Tail flick test data was collected and compared between 1, 3 and
12 month wildtype, heterozygous H304R/+, and homozygous H304R/R male
mice.
Figure 2-6 Tail flick meter
Sensitivity and response to thermal pain stimulus in mice is measured by tail flick meter.

50
2.4 Mouse dissections (sciatic nerve and dorsal root ganglia)
Tissue from the peripheral nervous system of mice was dissected out by surgical
procedures. Sciatic nerves and the dorsal root ganglia were collected from mice
of 3 month and 12 months of age.
2.4.1 Tissue harvesting and cryopreservation
Mice were euthanized by administering CO2 and cervical dislocation. Sciatic
nerve and lumbar dorsal root ganglia (DRG) samples were collected from both
left and right side of the mice using Leica S8 AP0 Stereo Microscope. Collected
tissue was fixed in 4% paraformaldehyde solution (EMS catalog#15710) for 30
minutes at 4°C. Then the tissue was left in 30% Sucrose solution (weight/volume)
overnight at 4°C. Next day, the tissue samples were embedded in labelled
histology molds (Ted Pella, Inc. catalog # 27118) containing Tissue-Tek® O.C.T.
Compound (Sakura Finetek USA Inc. catalog #25608-930). The tissue samples
were rapidly frozen on dry ice and stored at -80°C in labelled freezer boxes.
2.4.2 Cryosectioning and immunohistochemistry of tissue samples
Frozen sciatic nerve or lumbar DRG was cryosectioned in 10µm size thin
sections on Leica Cryostat (model# CM 1850) and collected on glass slides. The
samples were then washed in 1X PBS (Gibco) thrice for 5 minutes each to clean
away any O.C.T compound around the tissue. Waterproof boundaries were
drawn around each tissue sample with ImmEdge Hydrophobic Barrier PAP Pen
(Vector laboratories, catalog #H-4000). The tissue was permeabilized for 10
minutes with permeabilization buffer (0.5% Triton-X100). Then block buffer (10%

51
normal goat serum and 0.1% Triton-X100 in PBS) was added for 60 minutes on
the samples. Tissue samples were incubated with appropriate dilutions of
primary antibodies (table 2-1) overnight at 4°C. Next day, the primary antibodies
were washed with block buffer twice and then incubated with respective
secondary antibodies for 2 hours at the room temperature. Unbound antibodies
were washed with block buffer twice, followed by 1X PBS wash twice. The tissue
samples were then mounted with ProLong™ Gold Antifade Mountant with DAPI
(ThermoFisher Scientific, catalog# P36931) and sealed with a glass coverslip.

52
Table 2-1 Antibodies used for tissue immunohistochemistry
Primary antibody Labels Catalog number Dilution
used
Mouse monoclonal
to Beta-III tubulin
(βIII tubulin)
Neuronal soma
and axons
Abcam # 78078 1:200
Rabbit polyclonal
to Myelin Basic
Protein (MBP)
Myelinated axons Abcam # 40390 1:200
Secondary
antibody
Labels Catalog number Dilution
used
Rabbit polyclonal
to Glucose
Transporter-1
(GLUT-1)
Perineurial glial
cells in the sciatic
nerve’s
perineurium
Abcam # 15309 1:200
Alexa Goat anti
mouse conjugated
with 488
Against mouse
antibodies
ThermoFisher
Scientific # A11001
1:200
Alexa Goat anti
rabbit conjugated
with 546
Against goat
antibodies
ThermoFisher
Scientific # A110035
1:200

53
2.4.3 Microscopy and data analysis
Images of sciatic nerve and dorsal root ganglia tissues were captured on the
Leica M205 FA Fluorescence Microscope with Leica DFC3000G CCD camera.
The microscope settings and configurations were kept uniform between all the
samples to avoid variability (table 2-2).
Table 2-2 Microscopic settings used for imaging
False color Wavelength
(nm)
Exposure
time
Gain Intensity Magnification
Green 515 nm 139 ms 2.5x 60% 80X
Red 660 nm 139 ms 2.5x 40% 80X
Blue 440 nm 2.6 sec 2.5x 100% 80X
Transmittance none 139 ms 2.5x 45% 80X
Collected sciatic nerve images were then analyzed on the ImageJ software
(National Institutes of Health and the Laboratory for Optical and Computational
Instrumentation) to calculate the cross-sectional area of the nerve. Threshold
function was used to trace out the boundary of the nerve that was stained by the
perineural marker GLUT-1 antibody. The area inside the traced-out threshold
was measured using the ‘measure area’ function. The measured area (in mm2)
was collected and compared between the wildtype, heterozygous H304R/+, and
homozygous H304R/R mice.

54
2.5 Motility analysis studies in primary H304R neurons
Primary neurons from the embryonic brains of H304R mouse model were
dissected out and grown in vitro. Intracellular motility parameters were recorded
in in vitro primary neurons to characterize the functions of dynein in H304R
mouse model.
2.5.1 Tissue harvesting of embryonic primary neurons
Timed pregnancy breeding was set-up between the heterozygous male and
female mice. The embryos were removed at embryonic day 16.5 from a
euthanized pregnant female mouse. Brains were removed from each embryo and
were dissected further to collect the cerebellar cortex and hippocampus from
each lobe of the brain. Leica S8 AP0 Stereo Microscope was used for embryonic
dissections. Cortices and hippocampi of wildtype, heterozygous (H304R/+), and
homozygous (H304R/R) were then pooled together respectively after the
genotype of the embryos was determined by the PCR based method. Cortical
and hippocampal neurons were isolated respectively by enzymatic digestion
technique with Papain (Worthington® Biochemical Corp# LS003118) and DNase
(Worthington® Biochemical Corp # LK003172) in Hibernate® E minus
Ca2+/Mg2+ media (BrainBits® # HECAMG500). Neurons were plated on
0.3µg/ml poly-D-Lysine (Millipore Sigma) and 10µg/ml Laminin (ThermoFisher
Scientific # 23017-015) coated 25mm Fisherbrand® glass coverslips (Catalog #
12-545-102) in 6-well plates. The neurons were incubated at 37°C in warm
Neurobasal™ media (ThermoFisher Scientific # 21103049) supplemented with

55
100X Antibiotic-Antimycotic (Gibco™ # 15240062), 50X B-27™ Supplement
(Gibco™ # 17504044), and 100x GlutaMAX™ (Gibco™ # 35050061). The
neurons were half fed by removing and replacing half of the media with fresh
neuronal media every other day.
2.5.2 Live neuronal imaging with Rab7-GFP labeled vesicles
Cortical neurons of all 3 genotypes (wildtype, heterozygous (H304R/+),
homozygous (H304R/R)) were nucleofected with Rab7-GFP plasmid using
standard protocol on the Nucleofector™ 2b Device (Lonza # AAB-1001). The
nucleofected neurons were then plated on the glass coverslips and incubated at
37°C in warm NeurobasalTM media (ThermoFisher Scientific # 21103049). The
nucleofected neurons were half fed with fresh media every other day.
Live neuronal movies were captured on the Nikon® Eclipse Ti Inverted
Microscope with Photometrics® Evolve EMCCD camera (#512) at D.I.V 10. The
live cell movies were recorded using the Metamorph® Automation and Image
Analysis Software (Molecular Devices). The movies were made at 100x
magnification with 500 frames captured at the frame rate of 10 fps (frames per
second) with the appropriate green fluorescence filter cube. The longest visible
projection of the neuron was selected visually and was used for making neuronal
movies to capture the motility events inside the neuronal projections.
Motility analysis on movies of neurons containing Rab7-GFP labeled vesicles
was done using the Kymograph function of the Metamorph® image analysis

56
software (Molecular Devices) and described in the next section. Motility
parameters of Rab7-GFP labeled vesicles such as total number of movements,
run length (µm), run velocity (µm/sec), and the direction of vesicular movement
were calculated (figure 2-7). Motility data from different genotypes (wildtype,
heterozygous (H304R/+), homozygous (H304R/R) were compared using one-
way ANOVA test on the GraphPad Prism®6 software.
Kymograph analysis description
The following Figure 2-7 is provided to illustrate the various features of
kymographs. For this purpose, a hypothetical set of two vesicles is shown.
Stationary vesicle is shown as a blue circle and a moving vesicle is shown as a
red circle inside a cell (figure 2-7). The red vesicle is shown moving on a
microtubule track shown with an orange line and the red vesicle is moving from
point B to A. A kymograph line is drawn from the cell center to the periphery
shown in the black line. The kymograph analysis of the black line is shown on the
right in the form of a time (sec) vs distance (µm) graph. The blue stationary
vesicle that does not move has a constant line as shown in blue indicating zero
displacement on the time vs distance graph. In contrast, the red vesicle that
moves shows the displacement as a slant line indicating a “run” on the time vs
distance graph. The slope of the slant line gives the run velocity (µm/sec) and the
slope angle gives the direction of the movement (towards or away from the cell
body).

57
Figure 2-7 Schematic diagram of motility analysis using kymographs
Kymograph is a time vs distance graph generated to analyze the motility parameters on vesicles (blue circle = stationary vesicle, red circle = moving vesicle in motion).

58
2.5.3 Live neuronal imaging with Synaptophysin-RFP labeled vesicles
Cortical and hippocampal primary neurons were each infected with Bacmam
Synaptophysin RFP (2µl/ml) (ThermoFisher Scientific # C10610) and incubated
for 48 hours. Live neuronal movies were captured as mentioned in the sections
above. Motility analysis of neurons containing Bacmam Synaptophysin RFP
labeled vesicles was done using the Kymograph function of the Metamorph®
Automation and Image Analysis Software (Molecular Devices). Motility
parameters of Synaptophysin RFP positive vesicles such as total number of
movements, run length (µm), run velocity (µm/sec), and the direction of vesicular
movement were calculated. Motility data from different genotypes of mice
(wildtype, heterozygous (H304R/+), homozygous (H304R/R)) were compared
using one-way ANOVA test on GraphPad Prism®6 software.

59
CHAPTER 3: BEHAVIORAL PHENOTYPE CHARACTERIZATIONS OF H304R MICE
3.1 Introduction
Prior to our study, three other dynein heavy chain mutant mice were reported in
the literature – legs at odd angle (Loa), Cramping (Cra), and Sprawling (Swl).
Loa and Cra mice were generated using ENU-mediated mutagenesis (Hrabe de
Angelis et al., 2000) and Swl was created using radiation induced mutagenesis
(Duchen, 1974). All three mice displayed abnormal posture and hind limb
clenching during the tail suspension. These transgenic mice were considered to
be the paradigm mouse models of neurological dysfunction and were extensively
studied. Some of the phenotypic studies on Loa, Cra, and Swl mice are outlined
below.
A multi-stage protocol called SHIRPA (Smith/Kline/Harwell/Imperial
College/Royal Hospital/Phenotype Assessment) was used to quantitatively asses
the phenotype of the Loa mouse in a longitudinal study (Rogers et al., 2001). The
Loa/+ test results were compared with age and gender matched wildtype control
mice in the behavioral and functional assessments done from 1 to 16 months of
age. During the SHIRPA screening, the heterozygote Loa/+ and wildtype mice
were assessed on tail suspension, grip strength, wire maneuver, and limb tone at
1, 3 and 14-16 months age.

60
For grip strength testing, mice were gently pulled across a steel mesh grip and
the grip strength exerted was recorded qualitatively with arbitrary scores: no
effort= 0, slight semi-effective effort= 1, moderate effective effort= 2, active
effort= 4, and unusual effective effort =5. For the wire maneuver test, mice were
suspended on a horizontal wire by their fore limbs and the efforts to bring their
hind limbs to the wire were scored as “falls immediately” to “active grip” with hind
limbs. Lastly, in the hind limb tone assessment, mice were restrained in the
supine position and their resistance to the pressure on the hind paws (plantar
side) was recorded as “slight” or “moderate” or “heavy” resistance. Overall, the
Loa/+ mice exhibited abnormal posture of hind limb clenching during tail
suspension, significant reductions in hind limb (median score of 1= slight grip),
reduced performance in the wire maneuver test (median score of 1= difficulty in
grasping with hind limbs), and deficits in hind limb tone (median score of 1= slight
resistance) as compared to the wildtype control mice.
For another stage of SHIRPA protocol, Loa/+ mice performed the rotarod test
and spontaneous locomotor activity test. For the rotarod test, the test mouse was
placed on a rotating rotarod and time when a mouse would fall off from a rotarod
was recorded. In locomotor activity test, a mouse was placed in a clear Perspex
activity cage fitted with infra-red signals, and the infra-red beam breaks induced
by the mouse locomotion was detected and recorded over a 30 minutes test
window. The Loa/+ mice had a significant reduction in its rotarod performance at
1, 3 and 16 months’ time points as they were unable to stay on the rotarod as

61
long as the control wildtype mice. During locomotion test, the Loa/+ mice
interestingly showed more locomotor activity than the wildtype control mice. In
conclusion, the SHIRPA protocol of behavioral assessments provides a
comprehensive methodology for documenting progressive neurological
dysfunction in mouse models (Rogers et al., 2001).
The other two DHC mutant mice – Cra and Swl were also characterized using
SHIRPA protocol for their neurological functions. Just like Loa/+ mice, both Cra/+
and Swl/+ mice displayed hind limb clenching during tail suspension, gait
abnormalities, no reductions in front grip strength, and significantly reduced grip
strength in hind limbs (Banks and Fisher, 2008; Chen et al., 2007). The
homozygous Loa/Loa, Cra/Cra, and Swl/Swl mice were either embryonically
lethal or did not survive 24 hours post birth due to their inability to feed (Chen et
al., 2007; Hafezparast et al., 2003). Overall, the DHC mutant Loa, Cra, and Swl
mice models provided some interesting evidences in the role of dynein in
neurological dysfunction.
3.2 Rationale
The mutagenesis generated Loa, Cra and Swl mice have provided many key
insights in understanding the dynein function, however, the mutations in Loa
(F8580Y), Cra (Y1055C), and Swl ([GIVT]1040[A]) are not linked to any
neurodegenerative disease alleles in humans. It is hard to correlate their key
findings in understanding the dynein function in the context of neurodegenerative

62
diseases in humans. Discovery of the first dynein mutation (H306R) in human
neuropathy (CMT2O) was an opportunity to study dynein function. Hence, we
generated the novel H304R mouse model as a tool to understand the dynein
function and mechanism in human neurodegenerative diseases.
The H304R mouse model has a mutation in its dynein heavy chain that is
reported to lead to human neuropathies such as Charcot-Marie-Tooth Type 2O
disease (CMT2O) and Spinal Muscular Atrophy with Lower Extremity Dominant
(SMA-LED). Both CMT2O and SMA-LED patients exhibit similar clinical
symptoms such as they display lower limb weaknesses, muscle wasting, skeletal
and joint deformities of foot, joint contractures, and problems in walking. CMT2O
patients also show a loss of sensation symptoms in addition to their motor
defects. The disease linked Histidine to Arginine substitution mutation is at
position 306 in the human dynein heavy chain, and the corresponding mutation is
at position 304 in mouse dynein heavy chain (Figure 3-1).
Figure 3-1 Comparison of dynein heavy chain peptide
CMT2O disease linked mutation is a missense substitution of Histidine (H) to Arginine (R) at position 306 in human dynein heavy chain peptide. The corresponding mutation is at position 304 in mouse dynein heavy chain peptide.

63
We generated the novel H304R mouse model using knock-in gene targeting
technique (Sabblah et al., 2018). It is critical to remember that because a mouse
has a disease linked mutation, it does not mean that it would be a relevant model
to study that disease. A mouse model has to recapitulate the clinical
manifestations of the disease to be considered as a relevant model for studying
that disease. Therefore, a key question for the H304R mouse model was if the
H304R mice would develop and display the symptoms associated with CMT2O.
To answer this question, we performed extensive studies to investigate and
characterize the behavioral phenotype of the H304R mouse model. Several
phenotypic assessments and motor-skills tests were performed at multiple time
points to investigate if the H304R mouse model recapitulated the corresponding
phenotype associated with CMT2O. First, we started with breeding and growing
the heterozygous H304R/+ colony for preliminary behavioral assessments. The
initial assessments such as tail suspension reflex, grip strength, and rotarod test
were performed with the heterozygous H304R/+ male and female mice and the
data was compared with the wildtype mice. The preliminary findings between the
heterozygous H304R/+ and wildtype mice were published first (Sabblah et al.,
2018).
After seeing a promising phenotype with the heterozygous H304R/+ mice, we
decided to start another behavioral skills study with the homozygous H304R/R
mice. The homozygous H304R/R mice were then bred and grown into a colony
of mice. We compared the tail suspension reflex, grip strength, and rotarod data

64
between the homozygous H304R/R mice and the wildtype control mice at
multiple ages (manuscript in submission).
3.3 Results
3.3.1 H304R mice had normal life span
The founder H304R knock-in mice were bred to yield both male and female
heterozygous H304R/+ mice. The heterozygous H304R/+ pups had a normal
birth and normal lifespan of 2 years (Sabblah et al., 2018). The heterozygous
H304R/+ mice did not appear to have any significant bodyweight differences and
were able to move around the cage just like their littermate wildtype control
animals.
The homozygous H304R/R pups were able to survive after birth unlike the other
dynein heavy chain homozygous mutant pups (Loa/Loa, Cra/Cra, Swl/Swl) and
had a normal lifespan (Chen et al., 2007; Hafezparast et al., 2003). The
homozygous H304R/R mice appeared to have gait defects such as a high
steppage gait in hind limbs (figure 3-2). Also, they were not as inclined to move in
the cage as compared to their littermate wildtype control and heterozygous
H304R/+ mice. They did not have any significant bodyweight differences
compared to their littermates.

65
Figure 3-2 Gait phenotype of H304R mouse model
Homozygous H304R/R mice showed gait defects such as high steppage gait in hind limbs (black arrow).
3.3.2 H304R mice showed atypical tail suspension phenotype
Tail suspension tests were performed to assess the phenotype of H304R mouse
model. Mice tend to splay their hind limbs away from their body when lifted by the
tails as their typical tail suspension reflex. However, in some cases, mice would
clench their hind limbs together and hold them close to their body as part of their
atypical tail suspension phenotype (Figure 3-3).

66
Figure 3-3 Tail suspension phenotype of H304R mouse model
Typical tails suspension reflex consists of splaying of hind limbs away from the body, whereas, atypical tail suspension involves clenching of hind limbs together closer to the body.
The tail suspension reflex test was performed on the H304R colony at multiple
ages and the mouse’s phenotype was scored as typical or atypical reflex. The tail
suspension reflex was then statistically compared between the 3 genotypes
using the Chi-square test (p<0.05). We observed that about 20-30% of male and
female heterozygous H304R/+ mice started to display the atypical tail suspension
phenotype at later ages (9 and 12 months) as compared to the wildtype control

67
mice (Figure 3-4). A majority (60-80%) of male and female homozygous
H304R/R mice significantly displayed the atypical tail suspension reflex
compared to the wildtype control and heterozygous H304R/+ mice right from an
early age. The tail suspension data is summarized in Table 3-1.
Figure 3-4 Atypical tail suspension response of H304R mouse model
Male and female heterozygous H304R/+ mice (gray bars) showed atypical tail suspension reflex at later time points as compared to the wildtype mice (black bars) (Chi-square test p<0.05). Whereas, both male and female homozygous H304R/R mice (white bars) showed atypical tail suspension reflex at all the time points (Chi-square test p<0.05). Note- * is the statistical significance as compared to the wildtype only and # is the statistical significance as compared to the wildtype and H304R/+.
* * * *
#
# # #
# # # #

68
Table 3-1 Tail suspension reflex of H304R mice
Tail
refl
ex
Time point
Percentage of mice showing atypical tail suspension
reflex
Number of animals (n) p-value (Chi-square
test)
Wild-type H304R/+ H304R/R Wild-type
H304R/+ H304R/R Wild-type vs
H304R/+
Wild-type vs
H304R/R
Mal
e m
ice
3 months
7.5% 2.0% 61.7% 34 25 26 0.0745 <0.0001
6 months
5.8% 11.2% 76.6% 46 34 20 0.0642 <0.0001
9 months
9.6% 21.6% 74.7% 29 27 20 0.0120 <0.0001
12 months
12.2% 27.3% 79.6% 40 42 18 0.0002 <0.0001
Fem
ale
mic
e
3 months
10.0% 15.4% 66.3% 9 8 27 0.5434 <0.0001
6 months
8.0% 12.5% 62.6% 35 21 20 0.2771 <0.0001
9 months
11.3% 18.6% 70.9% 54 37 19 0.0314 <0.0001
12 months
11.3% 18.5% 70.7% 56 44 19 0.0350 <0.0001

69
3.3.3 Heterozygous H304R/+ mice show mild defects in their motor phenotype
Initial behavioral assessments were done with the male and female heterozygous
H304R/+ mice and the data was statistically compared with age and gender
matched wildtype control mice. The behavioral test results are as follows:
Heterozygous H304R/+ mice show reductions in grip strength
Standard grip strength assay was performed to assess the muscular strength in
all limbs and front limbs of heterozygous H304R/+ mice. The male heterozygous
H304R/+ mice showed a statistically significant reduction of about 10% in all
limbs grip strength at all ages as compared to the wildtype control mice
(Student’s t-test, p<0.05) (Figure 3-5). Female heterozygous H304R/+ mice
showed a statistically significant reduction of about 5% in all limbs grip strength
at only 6 and 9 month ages as compared to the wildtype control animals
(Student’s t-test, p<0.05).
Male heterozygous H304R/+ mice showed a statistically significant reduction of
about 7% in its front grip strength at later ages of 9 and 12 months as compared
to the wildtype mice (Student’s t-test, p<0.05). The female heterozygous
H304R/+ mice did not have any reductions in their front grip strength as
compared to the wildtype mice at any age. The grip strength data for male
H304R/+ and female H304R/+ mice are summarized in table 3-2 and table 3-3
respectively.

70
Figure 3-5 Grip strength phenotype of heterozygous H304R/+ mice
Both male and female heterozygous H304R/+ mice showed significant reductions in all limbs grip strength as compared to the wildtype mice (A & B). Only male heterozygous H304R/+ mice showed significant reductions in front limbs grip strength at later time points (C). The female heterozygous H304R/+ mice did not show any reductions in front grip strength as compared to the wildtype control mice (D). Note- * Student’s t-test, p<0.05.

71
Table 3-2 Grip strength in H304R/+ male mice
Test (male mice)
Time point
Mean ± std. dev (grams) Number of animals (n)
p-value (Student’s t-test)
Wild-type H304R/+ Wild-type
H304R/+ Wild-type vs
H304R/+
All limbs
3 months
212.5 ± 23.2 190.4 ± 26.8 50 43 0.0001
6 months
225.1 ± 30.8 201.0 ± 29.9 62 58 0.0001
9 months
230.6 ± 37.9 206.8 ± 33.4 41 42 0.0033
12 months
244.5 ± 32.1 225.3 ± 33.5 40 42 0.0096
Front limbs
3 months
99.2 ± 12.9 95.6 ± 17.4 50 43 0.2654
6 months
88.8 ± 14.3 83.8 ± 15.0 62 58 0.0667
9 months
89.8 ± 12.9 82.1 ± 11.1 41 42 0.0045
12 months
92.0 ± 11.9 86.1 ± 13.4 40 42 0.0362

72
Table 3-3 Grip strength in H304R/+ female mice
Test (female mice)
Time point
Mean ± std. dev (grams) Number of animals (n)
p-value (Student’s t-test)
Wild-type H304R/+ Wild-type
H304R/+ Wild-type vs
H304R/+
All limbs
3 months
175.5 ± 24.0 171.9 ± 22.0 51 35 0.4820
6 months
192.9 ± 15.9 185.7 ± 16.9 56 41 0.0324
9 months
209.8 ± 18.8 199.8 ± 16.5 55 41 0.0068
12 months
222.2 ± 22.2 216.4 ± 20.0 54 41 0.1843
Front limbs
3 months
91.0 ± 12.5 89.1 ± 12.2 51 35 0.4801
6 months
81.5 ± 9.8 81.8 ± 11.0 56 41 0.8959
9 months
81.3 ± 11.8 79.8 ± 11.1 55 41 0.5148
12 months
85.9 ± 13.4 85.4 ± 13.1 54 41 0.8681

73
Heterozygous H304R/+ mice show deficits in the rotarod performance
Rotarod test was performed to assess the motor coordination phenotype of
H304R/+ mice. Mice were placed inside the rotarod equipment and their falling
off from the rotarod before the completion of the rotarod cycle was measured as
latency to fall (in seconds). The male heterozygous H304R/+ mice were falling
from the moving rotarod quicker than the wildtype mice (Table 3-5). They showed
significant reductions in their latency to fall (by about 25%) as compared to the
wildtype control animals at 3, 6 and 9 months of age (Student’s t-test, p value
<0.05) (Figure 3-6). Female heterozygous H304R/+ mice also showed a similar
trend (Table 3-5), but had a significant reduction of 20% in their latency to fall
only at an early age of 3 months as compared to the wildtype mice (Student’s t-
test, p value <0.05).
Figure 3-6 Rotarod performance test of the H304R/+ mice
Male and female heterozygous H304R/+ mice showed significant defects in their rotarod performance as compared to the wildtype at some time points (*Student’s t-test, p<0.05).

74
Table 3-4 Rotarod performance in H304R/+ male mice
Test (male mice)
Time point Mean ± std. dev (seconds)
Number of animals (n)
p-value (Student’s t-test)
Wild-type H304R/+ Wild-type
H304R/+ Wild-type vs
H304R/+
Rotarod
3 months 78.8 ± 40.9 55.2 ± 30.4 51 43 0.0019
6 months 95.5 ± 49.1 73.0 ± 42.4 48 41 0.0223
9 months 101.9 ± 52.9 74.6 ± 44.8 28 33 0.0359
12 months 100.7 ± 48.6 82.4 ± 45.7 20 25 0.2035
Table 3-5 Rotarod performance in H304R/+ female mice
Test (female mice)
Time point Mean ± std. dev (seconds)
Number of animals (n)
p-value (Student’s t-test)
Wild-type H304R/+ Wild-type
H304R/+ Wild-type vs
H304R/+
Rotarod
3 months 103.6 ± 47.6 82.7 ± 34.3 51 35 0.0205
6 months 113.7 ± 54.9 104.9 ± 45.2 51 35 0.4210
9 months 106.8 ± 61.4 105.9 ± 54.0 51 35 0.9386
12 months 98.8 ± 62.6 87.7 ± 51.3 50 35 0.3711

75
3.3.4 Homozygous H304R/+ mice show severe defects in their motor phenotype
After seeing the altered phenotype of motor skills in the heterozygous H304R/+
mice, we decided to perform behavioral assessments of the male and female
homozygous H304R/R mice. The data was statistically compared with age and
gender matched with the wildtype control mice. The behavioral test results are as
follows:
Homozygous H304R/R mice show reductions in grip strength
Standard grip strength assay was performed to assess the muscular strength in
all limbs and front limbs of heterozygous H304R/R mice. Male homozygous
H304R/R mice showed statistically significant reductions of about 30% in all
limbs grip strength as compared to the wildtype animals at all ages (Student’s t-
test, p value <0.05) (Figure 3-7). Female homozygous H304R/R mice also
showed statistically significant reductions of about 20% in all limbs grip strength
as compared to the wildtype animals at all ages tested (Student’s t-test, p value
<0.05). Both male and female homozygous H304R/R mice did not show any
reductions in their front grip strength as compared to the wildtype mice at all
ages. Grip strength data for male homozygous H304R/R (Table 3-6) and female
homozygous H304R/R (Table 3-7) mice are summarized in the table below.

76
Figure 3-7 Grip strength phenotype of heterozygous H304R/R mice
Both male and female homozygous H304R/R mice showed significant reductions in all limbs grip strength (A & B) as compared to the wildtype controls. No grip strength reductions were observed in the front limbs of both male and female homozygous H304R/R mice (C&D). Note- * Student’s t-test, p value <0.05.

77
Table 3-6 Grip strength in H304R/R male mice
Test (male mice)
Time point
Mean ± std. dev (grams) Number of animals (n)
p-value (Student’s t-test)
Wild-type H304R/R Wild-type
H304R/R Wild-type vs
H304R/R
All limbs
3 months
212.5 ± 23.2 162.0 ± 46.3 50 26 <0.0001
6 months
225.1 ± 30.8 162.9 ± 31.4 62 20 <0.0001
9 months
230.6 ± 37.9 162.2 ± 38.6 41 20 <0.0001
12 months
244.5 ± 32.1 167.0 ± 31.3 40 18 <0.0001
Front limbs
3 months
99.2 ± 12.9 96.0 ± 23.6 50 26 0.9995
6 months
88.8 ± 14.3 84.6 ± 21.7 62 20 0.9970
9 months
89.8 ± 12.9 85.8 ± 18.8 41 20 0.9987
12 months
92.0 ± 11.9 84.1 ± 20.2 40 18 0.7910

78
Table 3-7 Grip strength in H304R/R female mice
Test (female mice)
Time point
Mean ± std. dev (grams) Number of animals (n)
p-value (Student’s t-test)
Wild-type H304R/R Wild-type
H304R/R Wild-type vs
H304R/R
All limbs
3 months
175.5 ± 24.0 154.5 ± 23.0 51 27 0.0004
6 months
192.9 ± 15.9 164.9 ± 28.5 56 20 0.0004
9 months
209.8 ± 18.8 161.0 ± 29.5 55 19 <0.0001
12 months
222.2 ± 22.2 173.2 ± 39.2 54 19 <0.0001
Front limbs
3 months
91.0 ± 12.5 91.5 ± 15.5 51 27 >0.9999
6 months
81.5 ± 9.8 81.7 ± 17.0 56 20 >0.9999
9 months
81.3 ± 11.8 81.6 ± 17.9 55 19 >0.9999
12 months
85.9 ± 13.4 86.5 ± 21.1 54 19 >0.9999

79
Homozygous H304R/R mice show deficits in the rotarod performance
Homozygous H304R/R mice were unable to keep up with the rotating rotarod
and were falling off the rod a lot quicker than the wildtype mice. Male
homozygous H304R/R mice showed statistically significant reductions in their
latency to fall as compared to the wildtype at all ages (Student’s t-test, p value
<0.05) (Figure 3-8). Similarly, female homozygous H304R/R mice also showed
statistically significant reductions in their rotarod performance as compared to the
wildtype mice at all ages tested (Student’s t-test, p value<0.05). Both male (Table
3-8) and female (Table 3-9) homozygous H304R/R mice showed about 65%
reduction in their latency to fall as compared to the wildtype mice.
Figure 3-8 Rotarod performance test of the H304R/R mice
Both male and female homozygous H304R/R mice showed significant defects in their rotarod performance as compared to the wildtype at all ages (* - Student’s t-test, p value <0.05).

80
Table 3-8 Rotarod performance in H304R/R male mice
Test (male mice)
Time point Mean ± std. dev (seconds)
Number of animals (n)
p-value (Student’s t-test)
Wild-type H304R/R Wild-type
H304R/R Wild-type vs
H304R/R
Rotarod
3 months 78.8 ± 40.9 25.0 ± 9.6 51 26 <0.0001
6 months 95.5 ± 49.1 22.4 ± 9.7 48 20 <0.0001
9 months 101.9 ± 52.9 17.8 ± 9.6 28 20 <0.0001
12 months 100.7 ± 48.6 18.8 ± 9.7 20 18 <0.0001
Table 3-9 Rotarod performance in H304R/R female mice
Test (female mice)
Time point Mean ± std. dev (seconds) Number of animals (n)
p-value (Student’s t-test)
Wild-type H304R/R Wild-type
H304R/R
Wild-type vs
H304R/R
Rotarod
3 months 103.6 ± 47.6 28.1 ± 9.1 51 27 <0.0001
6 months 113.7 ± 54.9 30.8 ± 18.7 51 20 <0.0001
9 months 106.8 ± 61.4 23.9 ± 12.0 51 19 <0.0001
12 months 98.8 ± 62.6 23.2 ± 9.8 50 19 <0.0001

81
3.4 Conclusions
We generated a novel H304R mouse model for Charcot-Marie-Tooth type 2O
(CMT2O) with a disease mutation in its dynein heavy chain. We performed
several phenotypic and behavioral assessments on male and female
heterozygous H304R/+ and homozygous H304R/R mice. The heterozygous
H304R/+ had a normal lifespan and survived until 2 years (end point of the
study). Similarly, the homozygous H304R/R mice had a normal lifespan unlike
the other dynein heavy chain mutant mice (Loa/Loa, Cra/Cra, and Swl/Swl) that
were embryonic or post-natal lethal (Chen et al. (2007); (Hrabe de Angelis et al.,
2000).
Male and female heterozygous H304R/+ mice showed an atypical tail suspension
reflex of hind limb clenching at later ages (9 and 12 months). The atypical tail
suspension phenotype could be indicative of motor dysfunction in lower
extremities (or hind limbs) of the mice. Male and female heterozygous H304R/+
mice showed significant reductions of 5-10% in their hind limbs grip strength.
Only male heterozygous H304R/+ mice showed significant reductions in the front
limbs grip strength at later ages (9 and 12 months). Additionally, male
heterozygous H304R/+ mice also showed significant ~25% reduction in the
rotarod performance almost at all the time points. Female heterozygous H304R/+
mice showed a significant ~20% reduction in the rotarod performance test at an
early age only (3 months).

82
Male and female homozygous H304R/R mice appeared to have defects in their
gaits and were inclined to move less inside the cage as compared to their
littermates. The majority of homozygous H304R/R significantly showed atypical
tail suspension reflex right from an early age 3 months to all the way up to 12
months. Male and female homozygous H304R/R mice also showed significant
20-30% reductions in all limbs grip strength as compared to the wildtype mice,
but not in front limbs grip strength. Both male and female homozygous H304R/R
mice also performed poorly on the rotarod performance test with ~65% reduction
in latency to fall because they were unable to stay longer on the moving rod than
the wildtype control mice.
So far, our initial investigation and characterization of the novel H304R mice with
CMT2O mutation in DHC offered the following insights:
The H304R mouse model is a novel mouse model with a mutation in the
dynein heavy chain that is known to cause neuropathies in humans
(CMT2O and SMA-LED) (Tsurusaki et al. (2012); (Weedon et al., 2011).
The H304R mouse model was able to recapitulate and express the
symptoms of human neuropathies (CMT2O and SMA-LED) such as
muscle weaknesses, and motor coordination defects.
The heterozygous H304R/+ mutation expressed and recapitulated the
heterogeneity of motor phenotype in mice, similar to the heterogeneity of

83
clinical symptoms seen in humans with the autosomal dominant CMT2O
mutation.
Quantitatively, the heterozygous H304R/+ mice displayed a milder
reduction in its motor skills phenotype, whereas, the homozygous
H304R/R mice displayed a severe reduction in its motor skills phenotype.
The male heterozygous H304R/+ mice appeared to be affected more
significantly by the mutant dynein function than the female heterozygous
H304R/+ mice.
The male heterozygous H304R/+ mice had a variable onset of the motor
phenotype, sometimes at an early age (reductions in all limbs grip strength
and rotarod tests) and sometimes at a later age (atypical tail suspension
and reduced front limbs grip strength).
Both male and female homozygous H304R/R mice appeared to be
affected equally by the mutant dynein function.
The homozygous H304R/R mice consistently had an early onset of motor
phenotype that was severely deficient right from the start. In addition, the
homozygous H304R/R mutation phenotype was also static and non-
progressive in nature, i.e. it was not worsening drastically as the mice
aged.
A dynein molecule contains a dimer of two DHC monomeric peptides
linked together. A unique advantage of the homozygous H304R/R
mutation is that the homozygous mice will have a uniform composition of

84
dynein molecules with DHC dimers containing both mutant monomers. In
contrast, the heterozygous H304R/+ mutation leads to a mixed pool of
dynein in heterozygous mice, i.e. a quarter population of DHC dimers will
have both wildtype monomers, a quarter of DHC with both mutant
monomers, and half of the DHC dimers with one wildtype and one mutant
monomer.
In conclusion, the H304R mouse model appears to serve as a useful tool to
understand the role of dynein in neurodegenerative diseases. Future biochemical
studies with H304R mice can unveil the molecular mechanism of the mutant
dynein function in neurons.

85
CHAPTER 4: SENSORY PHENOTYPE CHARACTERIZATIONS OF H304R MICE
4.1 Introduction
Many CMT patients show the symptoms of sensory neuropathy along with motor
neuropathy. Common sensory deficits reported in CMT patients include loss of
sensation in distal limbs, paresthesia (tingling sensation), reduced
proprioception, and some patients also displayed reduction in their fine touch and
vibration sense (Weedon et al., 2011). Due to loss of sensation in their lower
limbs, some patients lose sensitivity to pain due to any foot wounding.
Consequently, that further leads to complications of foot ulcerations and usually a
follow up surgery is needed to remove the damaged foot/toes. It is worth noting
that previously published dynein heavy chain mutant mice (Loa, Cra and Swl)
reported sensory deficits in their phenotype (Chen et al., 2007; Dupuis et al.,
2009) as outlined below.
4.1.1 Sensory deficits in previously published mutant dynein mice models
The sciatic nerve is the largest nerve in the body, and it innervates the lower limb
muscles of mice. The outer layer of dense irregular connective tissue around a
sciatic nerve is called epineurium (Figure 4-1). The sciatic nerve is made up of
fascicles i.e. the several bundles of neuronal axons that are enclosed together.
The outer layer of a fascicle is lined with perineural glial cells and is called the
perineurium. The extracellular matrix space between neuronal axons inside a
fascicle is called the endoneurium.

86
Figure 4-1 Sciatic nerve cross section
The sciatic nerve is covered with a layer of connective tissue called epineurium and contains several nerve bundles called fascicles. The fascicles are lined with glial cells that make up a layer called perineurium. The extracellular space around the bundles of axons inside the fascicles is called endoneurium.
Deficits in the sciatic nerve anatomy have been linked to sensory dysfunction in a
previously reported dynein heavy chain mutant mouse (Chen et al., 2007). Both
heterozygote Loa/+ and heterozygote Swl/+ mice reportedly had thinner sciatic
nerves as compared to wildtype animals. Both ventral root and dorsal root
contribute their spinal nerves (neuronal axons) to make up the sciatic nerve. It
was observed and determined that the dorsal root size was smaller in both
heterozygote Loa/+ and Swl/+ mice as compared to wildtype controls, whereas
the ventral root size was unchanged. The Cra/+ mice also showed thinner dorsal
roots and loss of large caliber axons (Dupuis et al., 2009)

87
A dorsal root ganglion is a cluster of neuronal cell bodies or somas of pseudo-
unipolar sensory neurons. The dorsal root ganglia (DRG) receive input signals
from sensory neurons innervating target tissue such skin, muscles, and organs.
Input nerve signals travel along the sensory neuron axons in ‘afferent direction’
i.e. from target tissue to dorsal root ganglia and further passed onto the spinal
column for signal processing. Additionally, the dorsal root ganglion is composed
of different neuronal subpopulation involved in sensory stimulus reception. For
example, nociceptive neuronal subpopulation inside a dorsal root ganglion is
responsible for sensing pain stimulus from the target tissue such as skin. Another
example inside a dorsal root ganglion is the proprioceptive neuronal
subpopulation that is involved in sensing the muscle stretch and orientation.
In the study mentioned above, the DRG in heterozygote Loa/+ and heterozygote
Swl/+ mice were also examined, and they revealed interesting insights into the
DRG neuronal subpopulation composition (Chen et al., 2007). It was determined
that the total number of neurons were significantly reduced in dorsal root ganglia
from lumbar (L4) spinal region of heterozygote Loa/+ and heterozygote Swl/+
mice. Specifically, it was determined that proprioceptive neuronal subpopulations
were reduced by 69% in the dorsal root ganglia (L4) of newly born heterozygote
Swl/+ animals. Proprioceptive neuronal subpopulation count was not studied in
heterozygote Loa/+ mice. In conclusion, the sensory studies on heterozygote
Loa/+ and Swl/+ mice laid out the basis for investigating the sensory phenotype
of the H304R mouse model.

88
4.2 Rationale
We focused on the motor behavior skills characterization of the novel H304R
mouse in our initial studies and observed deficits in the tail suspension, grip
strength and the motor coordination phenotype of the mice. Here I start
investigating the sensory phenotypes of H304R mice to understand and compare
those to the sensory phenotypes of previously published dynein heavy chain
mutant mice (Loa/+ and Swl/+).
First, I started with assessing the sensory behavior phenotype by exposing
wildtype, heterozygous H304R/+, and homozygous H304R/R mice (both male
and female) to the tail flick sensitivity test and recorded their response to pain
stimulus at early (1 month and 3 months) and a later (12 months) time point.
Based on the tail flick data, I selected the male H304R mice cohort at 3 and 12
months’ time points for further sensory tissue investigation. I examined the gross
anatomy of sciatic nerve in the wildtype, heterozygous H304R/+, and
homozygous H304R/R male mice at 3 and 12 months of age and compared its
thickness in all three genotypes. I dived deeper into quantitative analysis of
sciatic nerve thickness by comparing the total fascicular area in the sciatic nerve
from the male H304R mice of all three genotypes. Because the dorsal root
ganglia (DRG) contribute its axons to the sciatic nerve, I refined and optimized
conditions to study the DRG composition to further understand the sciatic nerve
phenotype of the H304R mouse model.

89
4.3 Results
4.3.1 Altered latency to respond to pain stimulus in H304R mice
The tail flick test was used to measure the sensitivity of H304R mice to thermal
pain stimulus. A test mouse was restrained safely in a restrainer and its tail was
briefly exposed to a thermal light beam as a source of pain stimulus at a
moderate intensity setting (50 out of 99 on the setting scale). The tail flick meter
recorded the time (in seconds) that a mouse would take to remove its tail out of
the thermal light beam path (pain stimulus). Both male and female cohorts of
H304R mice were tested on the tail flick test at multiple time points and their data
was compared respectively between the three genotypes.
Figure 4-2 Tail flick pain sensitivity test of H304R male mice
Male homozygous H304R/R mice displayed a significantly quicker response to the pain stimulus at 3 months and 12 months of age as compared to their respective wildtype control male mice (One-way ANOVA, p<0.05). Only 3 month female homozygous H304R/R mice displayed a significant quicker response to the pain stimulus as compared to the wildtype control female mice (One-way ANOVA, p<0.05). 12 month old female homozygous H304R/R data is not presented due to the unavailability of mice.
Male mice Female mice
HH
HR
RR
0
5
1 0
1 5
1 m o n th m a le ta il f lic k
Tim
e t
o r
es
po
nd
(s
ec
)
HH
HR
RR
0
5
1 0
1 5
3 m o n th m a le ta il f l ic k w /o s in g le g e n o
Tim
e t
o r
es
po
nd
(s
ec
)
HH
HR
RR
0
5
1 0
1 5
3 m o n th fe m a le ta il f l ic k w /o s in g le g e n o
Tim
e t
o r
es
po
nd
(s
ec
)
HH
HR
RR
0
5
1 0
1 5
1 m o n th fe m a le ta il f lic k
Tim
e t
o r
es
po
nd
(s
ec
)
* *
HH
HR
RR
0
5
1 0
1 5
9 m o n th m a le ta il f lic k
Tim
e t
o r
es
po
nd
(s
ec
)
*
HH
HR
0
5
1 0
1 5
9 m o n th m a le ta il f lic k
Tim
e t
o r
es
po
nd
(s
ec
)
Res
po
nse
tim
e (s
ec)
Res
po
nse
tim
e (s
ec)

90
Interestingly, both heterozygous H304R/+ and homozygous H304R/R male mice
took less time to respond to the pain stimulus than the wildtype control mice
(Figure 4-2) at multiple time points. However, the quick response time was
statistically significant only in the homozygous H304R/R male mice at 3 months
and 12 months of age (One-way ANOVA, p<0.05) as compared to the wildtype
mice. There was no significant difference between the pain response time of
heterozygous H304R/+ and homozygous H304R/R male mice at any time points
tested (Table 4-1).
Female heterozygous H304R/+ and homozygous H304R/R mice did not show
any significant differences in their tail flick response times as compared to the
wildtype control female mice at 1 month of age. However, at 3 months of age,
both heterozygous H304R/+ and homozygous H304R/R female mice took lesser
time to respond to the pain stimulus. But only homozygous H304R/R female mice
had a statistically significantly quicker response time as compared to the wildtype
control mice (One-way ANOVA, p<0.05). The tail flick data for 12 month old
female homozygous H304R/R mice is not presented due to a lack of the mouse
availability. There was no significant difference between the pain response time
of heterozygous H304R/+ and homozygous H304R/R female mice at all time
points.

91
Table 4-1 Tail flick data
Average time of response (sec), number of animals (n), and the one-way ANOVA test p-values (with Tukey’s multiple
comparison test) for the wild-type, heterozygous (H304R/+) and homozygous (H304R/R) mice
Tail
flick
test
Time
point
(age)
Average time of response
to the pain stimulus
(seconds)
Number of animals (n) p-value (Tukey’s multiple
comparison)
Wild-
type
H304R/+ H304R/R Wild-
type
H304R/+ H304R/R Wild-
type vs
H304R/+
Wild-type
vs
H304R/R
H304R/+
vs
H304R/R
Ma
le m
ice 1 month 8.13 7.28 4.46 3 13 2 0.9051 0.4203 0.4743
3 months 8.64 6.40 5.21 7 18 6 0.0843 0.0284 0.5117
12 months 8.43 7.17 4.48 4 6 5 0.6272 0.0370 0.1227
fem
ale
mic
e 1 month 7.60 7.77 7.86 5 9 4 0.9876 0.9796 0.9968
3 months 8.43 7.04 4.59 9 12 4 0.3108 0.0163 0.1335
12 months 10.43 7.83 N/A 4 4 N/A N/A N/A N/A

92
4.3.2 Sciatic nerve appeared thinner in H304R mice
The sciatic nerve is the largest nerve in the body and it innervates the lower
limbs of mice. The sciatic nerve arises from spinal nerves L3, L4, and L5 from the
lumbar spinal column in mice. Dissections were performed on euthanized
wildtype, heterozygous H304R/+, and homozygous H304R/R male mice to
expose & collect both left and right sciatic nerves. Then, the tissue was frozen,
sectioned and stained with immunohistochemistry-based methods to compare
the sciatic nerve thickness between the three genotypes.
Sciatic nerve dissections were done on two litters each of 3 months and 12
month old wildtype, heterozygous H304R/+, and homozygous H304R/R male
mice. Visually, it appeared that both the left and right sciatic nerves were thinner
in the male homozygous H304R/R mice as compared to the wildtype and
heterozygous H304R/+ littermate control (Figure 4-3). Heterozygous H304R/+
mice appeared to have sciatic nerves of similar or slightly reduced thickness as
compared to the wildtype littermate control animals. These visual observations of
sciatic nerve were then further validated by quantitatively measuring the nerve
thickness and axonal counts using immunohistochemistry-based staining
methods as described next.

93
Figure 4-3 Comparison of sciatic nerve thickness of H304R male mice
The sciatic nerve appeared to be thinner in 3-month homozygous H304R/R male mice as compared to the wildtype and heterozygous H304R/+ littermates.
4.3.3 Reduced total sciatic nerve fascicular area in H304R mice
After seeing apparent differences in sciatic nerve thickness of H304R mice, I
wanted to quantify this observed difference by measuring the total fascicular
area. The sciatic nerve is made up of fascicles or bundle of neuronal axons
enclosed within the perineurium layer. Fascicular area was calculated by
measuring the area enclosed inside perineurium layer of fascicles that was
stained by a perineural marker - glucose transporter GLUT-1 antibody (details in
Chapter 2: Materials and methods). Total area of all the fascicles were added
together and compared between the 3-month (Figure 4-4) and 12-month (Figure
4-5) old wildtype, heterozygous H304R/+, and homozygous H304R/R male mice
respectively.
Wild-type
H304R/+
H304R/R

94
Figure 4-4 Sciatic nerve cross sections of 3-month H304R male mice
Sciatic nerve cross sections were stained with perineural glial maker – GLUT 1 (shown here in red) to label individual fascicles. Total fascicular area was calculated for all fascicles and compared between the genotypes. Homozygous H304R/R mice showed a significant reduction in the total fascicular area as compared to wildtype littermate control. No significance differences were observed between heterozygous H304R/+ and wildtype mice.
Wild-type H304R/+ H304R/R
Litter #1
Litter #2

95
Figure 4-5 Sciatic nerve cross sections of 12-month H304R male mice
Sciatic nerve cross sections were stained with perineural glial maker – GLUT 1 (shown here in red) to label individual fascicles. Total fascicular area was calculated for all fascicles and compared between the genotypes. Homozygous H304R/R mice showed a significant reduction in the total fascicular area as compared to wildtype littermate control. No significance differences were observed between heterozygous H304/R+ and wildtype mice.
Wild-type H304R/+ H304R/R
Litter #1
Litter #2

96
By visual observation, the sciatic nerve appeared thinner in homozygous
H304R/R male mice as compared to the wildtype littermate control animal at both
3 months and 12 months of age. This result was quantitatively confirmed by
observing a significant overall reduction of about 40% in total fascicular area of
homozygous H304R/R mice as compared to the wildtype control (One-way
ANOVA, Tukey’s multiple comparisons test, p<0.05) at 3 months and 12 months
age (Figure 4-6). Heterozygous H304R/+ mice also showed about 16% reduction
in the total fascicular area as compared to the wildtype littermate control animal
at 3 months and 12 months age, but this difference was not statistically
significant. Total fascicular area differences between homozygous H304R/R and
heterozygous H304R/+ male mice were not significant (Table 4-2).
Figure 4-6 Total fascicular area of sciatic nerve in H304R male mice
Homozygous H304R/R male mice had a statistically significant reduction of ~40% in its total fascicular area as compared to the wildtype control mice (1-way ANOVA, Tukey’s multiple comparisons test, p<0.05).
0
0.05
0.1
0.15
0.2
3 months 12 months
Tota
l fas
cicu
lar
area
(m
m2)
Sciatic nerve thickness
Wildtype
H304R/+
H304R/R
* *

97
Table 4-2 Total fascicular area in the sciatic nerve of H304R male mice
Total
fascicular area
Wildtype Heterozygous
H304R/+
Homozygous
H304R/R
3-month
littermate # 0.20 mm2 0.16 mm2 0.12 mm2
3-month
littermate #2 0.16 mm2 0.14 mm2 0.09 mm2
Average 3 -
month area 0.18 mm2 0.15 mm2 0.11 mm2
Total
fascicular area
Wildtype Heterozygous
H304R/+
Homozygous
H304R/R
12-month
littermate #1 0.19 mm2 0.20 mm2 0.12 mm2
12-month
littermate #2 0.17 mm2 0.10 mm2 0.10 mm2
Average 12 -
month area 0.18 mm2 0.15 mm2 0.11 mm2

98
4.3.4 Axonal counts in the sciatic nerve cross section
After seeing a reduction in the total fascicular area of the sciatic nerve, it was
apparent that the axon numbers inside the fascicles were needed to be counted
to reveal any alterations that may have led to this reduction. Alterations in total
fascicular area could be a result of reduced number of the axons or a reduction in
the myelination that could ultimately lead to the thinner nerves of heterozygous
H304R/+ and homozygous H304R/R mice.
The largest βIII tubulin stained fascicle from each genotype of 3-month littermate
male mice was selected (Figure 4-7). The axons were counted and recorded in
two categories based on their appearances – single axons and bundled axons. A
small and uniform green dot was counted as a single axon, while the larger
shaped green dot was counted as a bundle of multiple axons inside the fascicle.
Then the percentage of bundled axon over total axons and the percentage of
single axons over total axons were calculated and compared separately for all
three genotypes (Table 4-3). The preliminary results revealed a reduction in the
number of axons in both the heterozygous H304R/+ and homozygous H304R/R
mice (Figure 4-8). However, the percentage distribution of the total axons into
bundled axons or single axons categories were similar in all three genotypes
(Figure 4-9).

99
Figure 4-7 The largest fascicle inside the sciatic nerve of each genotype
The cross-sectional area of the largest fascicle inside each sciatic nerve was stained with the βIII tubulin antibody. Each green dot was counted as a single axon and a bunch of green dots were counted as a bundle of multiple axons.
Wildtype H304R/+ H304R/R

100
Table 4-3 Axon counts inside the largest fascicle
3 months Wildtype H304R/+ H304R/R
Bundled axons 629 461 362
Single axons 579 397 282
Total axons 1208 858 644
% of bundled axons 52.07% 53.73% 56.21%
% of single axons 47.93% 46.27% 43.79%
Figure 4-8 Total count of axons inside a fascicle
The total number of axons was counted in the cross-sectional area of the largest fascicle inside each sciatic nerve.

101
Figure 4-9 The percentage distribution of bundled axons vs single axons
The distribution of the total axons in the categories – bundled axons and single axons were similar in all three genotypes.

102
4.3.5 Identifying subpopulations of neurons in the adult DRG
After observing a significant reduction in sciatic nerve thickness and axonal count
in H304R mice, I wanted to determine if there was a corresponding alteration in
the dorsal root ganglia (DRG) composition. The DRG contribute its neuronal
axons to make up the sciatic nerves. The DRG contain a mixed subpopulation of
neurons such as nociceptive neurons (to sense pain) and proprioceptive neurons
(to sense muscle stretch and orientation). DRGs collected from the lumbar (L4)
region of wildtype, heterozygous H304R/+, and homozygous H304R/R male mice
were sectioned serially every 10µm and stained with anti-βIII tubulin antibody to
label all the neuronal somas (Figure 4-10).

103
Figure 4-10 Serial sections of dorsal root ganglia of H304R mice
The dorsal root ganglia from L4 lumbar region were serially sectioned. Best matched sections shown here in blue and orange boxes were picked for further analysis between the wildtype, heterozygous H304R/+ and H304R/R male mice.
Wil
dty
pe
H
30
4R
/+
H30
4R
/R

104
Best matched sections of L4 dorsal root ganglia were picked from wildtype,
heterozygous H304R/+, and homozygous H304R/R male mice for further staining
with neuronal subpopulation specific makers (Figure 4-11). I successfully
optimized the working conditions to stain dorsal root ganglia sections with anti-
CGRP antibody to label nociceptive neurons and with anti-Parvalbumin (PV)
antibody to label the proprioceptive neurons (Figure 4-11). This optimized
methodology (Table 4-4) can be further used to stain and count the anti-PV
positive neurons (proprioceptors) and the anti-CGRP positive neurons
(nociceptors).
Table 4-4 Optimized conditions for identifying DRG neuronal subpopulations
Primary antibody
Working concentration Notes
Mouse anti- βIII tubulin 1:200 Primary antibody
dilutions are to be
prepared in the Block
buffer prepared with
Bovine Serum Albumin
(BSA).
Overnight incubation at
4°C.
Goat anti-CGRP 1:100
Rabbit anti- parvalbumin 1:100
Secondary antibody
Working concentration Notes
Donkey anti- mouse 350 1:200 Goat secondary
antibody dilutions are to
be prepared in the Block
buffer prepared with
Bovine Serum Albumin
(BSA).
3 hours incubation at
room temperature.
Donkey anti- goat 488 1:100
Donkey anti- rabbit 568 1:200

105
Counting subpopulation numbers of anti-PV positive neurons and anti-CGRP
positive neurons can be used to calculate and compare the percentage of each
subpopulation over total neurons present in the DRG of H304R mice. Preliminary
data for the wildtype DRG is presented in table 4-5. Out of all the βIII tubulin
stained neurons, ~ 36% of the neurons were anti-PV positive and ~59% of the
neurons were anti-CGRP positive in the same section of the wildtype DRG.
Table 4-5 Preliminary counts of wildtype DRG subpopulation
Subpopulation Counts
Total cell bodies 39
anti-PV positive cell bodies 14
anti-CGRP positive cell bodies 23
% anti-PV positive cell bodies 35.90%
% anti-CGRP positive cell bodies 58.97%

106
Figure 4-11 Neuronal subpopulations staining of dorsal root ganglia
The dorsal root ganglia section from wildtype mice was stained with anti-βIII tubulin to identify all sensory neurons. The section was co-labelled with anti-parvalbumin and anti-CGRP primary antibodies to identify proprioceptive and nociceptive neuronal subpopulations respectively.

107
4.4 Conclusions
Charcot-Marie-Tooth patients exhibit sensory neuropathy symptoms in addition
to the motor neuropathy symptoms. In parallel, previously published dynein
heavy chain mutant Loa/+, Cra/+ and Swl/+ mice have also reported sensory
deficits (Chen et al., 2007; Dupuis et al., 2009). Based on the reports in human
CMT patients and previously published mutant dynein mice, I initiated the
sensory phenotype assessments of the H304R mouse model.
First, I started with characterizing the pain sensitivity response of H304R mice
with the tail flick test and found that both male and female homozygous H304R/R
mice had a statistically significant quicker response time to the pain stimulus than
the wildtype mice at 3 months of age. Male homozygous H304R/R mice also
showed a statistically significant quicker response time at 12 months of age as
compared to the wildtype mice. However, due to unavailability of female
homozygous H304R/R mice, no data was presented for comparison at 12
months. Heterozygous H304R/+ male mice also showed a quicker response to
tail flick test but didn’t have any significant differences as compared to the
wildtype control or homozygous H304R/R littermate mice at multiple time points.
It was previously reported that the heterozygote Loa/+ mice took a longer time to
respond to the tail flick test (unpublished data, (Banks and Fisher, 2008)). In
contrast, the H304R mutant mice took lesser time to respond on the tail flick test.
These contrasting observations indicate that the H304R mutation in the dynein

108
heavy chain might be affecting nociceptive neurons differently than the Loa
mutation (F580Y) in the dynein heavy chain.
Next, a closer examination of sciatic nerve anatomy revealed that homozygous
H304R/R mice had thinner sciatic nerves as compared to both the wildtype and
heterozygous H304R/+ littermate mice. Quantitatively homozygous H304R/R
mice had an overall reduction of ~40% in total fascicular area of its sciatic nerve
as compared to the wildtype littermates at 3 months and 12 months of age.
Heterozygous H304R/+ mice displayed a slight 16% reduction in the total
fascicular area at 3 months and 12 months of age, but this difference was not
statistically significant as compared to the wildtype littermate control animals.
Reductions in the sciatic nerve thickness of H304R mice were similar to the
observations of thinner sciatic nerves in the Loa/+ and Swl/+ mice (unpublished
data, (Banks and Fisher, 2008)). Repetition of sciatic nerve experiments needs to
be performed on more number of animals to confirm this preliminary observation.
Observations of the sciatic nerve appearing thinner and reduction in the total
fascicular area in the heterozygous H304R/+ and homozygous H304R/R mice
opened many interesting follow up questions. That is why counting the number of
axons inside the largest fascicle was the next key step to determine the potential
reason behind the observed reduction in the total fascicular area. The preliminary
data from a littermate set revealed that there was a reduction in the number of
the axons in both the heterozygous H304R/+ (~29%) and homozygous H304R/R
(47%) mice as compared to the wildtype control. However, the percentage

109
distribution of the bundled axons vs single axons was same in the fascicles from
all three genotypes. Reduced number of axons could be a potential reason
behind the thinner sciatic nerves inside the mutant H304R mice; however, further
investigations are necessary to establish this preliminary observation.
Because the sciatic nerve is a “mixed nerve”, it receives axons from both the
dorsal root (sensory neurons) and ventral root (motor neurons) of the spinal
column. The reduction in the diameter of sciatic nerve could be due to the loss of
axonal projections from the dorsal root and/or the ventral root. In order to answer
this question, I picked the sensory route or the dorsal root ganglia (DRG) to study
further.
As mentioned previously in this chapter, DRG has a mixed neuronal
subpopulation of sensory neurons that could be identified for quantitative
analysis. I successfully optimized the working conditions to label and identify the
neuronal subpopulation in the DRG of H304R mice. Future experiments using
these optimized conditions can be used to label all neurons with the βIII-tubulin
marker, proprioceptive neurons with the parvalbumin marker, and nociceptive
neurons with the calcitonin gene related protein (CGRP) marker. Next, these
labelled neuronal subpopulations can be quantitatively analyzed to calculate the
percentage of proprioceptive or nociceptive subpopulation over all the neurons
inside a DRG. Preliminary calculations with the wildtype DRG showed that ~36%
of all neuronal cell bodies were anti-parvalbumin positive cell bodies and nearly
~59% of all neurons were anti-CGRP positive cell bodies in the same stained

110
section of the DRG. Further quantitative subpopulation comparisons may reveal
some key differences in the composition of DRG in the three genotypes of the
H304R mice.
Overall, my preliminary work to characterize the sensory phenotype of H304R
mice revealed some interesting differences in the pain sensitivity response and
sciatic nerve anatomy of mutant heterozygous H304R/+ and homozygous
H304R/R mice. The sciatic nerve anatomy of mutant heterozygous H304R/+ and
homozygous H304R/R mice was also comparable with previously published
dynein mice (Loa/+ & Swl/+) as outlined above. Further in-depth studies with a
greater number of animals will be required to establish the preliminary
observation of reduced sciatic nerve thickness in the H304R mice. Subsequent
studies exploring and comparing the DRG neuronal subpopulations between the
three genotypes would be needed to get a deeper explanation of the preliminary
observations in the sensory phenotype of H304R mice.

111
CHAPTER 5: MOTILITY STUDIES OF H304R MICE
5.1 Introduction and rationale
Dynein is the key motor protein involved in the retrograde transport of cargoes
along the neuronal axon (Hirokawa et al., 1990). Direct role of dynein has been
established in the retrograde transport of neurotrophin mediated trk receptors
towards the neuronal soma for survival response (Heerssen et al., 2004; Yano et
al., 2001), fast axonal transport (Waterman-Storer et al., 1997), neurite growth
and organization (Ahmad et al., 1998), and neuronal migration (Tanaka et al.,
2004). Therefore, it is not surprising that the mutations in dynein have been
reported to show the impaired axonal transport in the DHC mutant Loa/+ mice
(Ori-McKenney et al., 2010) and in Swl/+ mice (Zhao et al., 2016).
Hence, we were also interested in investigating the intracellular motility
phenotype of H304R mouse model. Because previously studied DHC mutant
Loa/Loa mice showed defects in the development of its cortical and hippocampal
regions (Ori-McKenney and Vallee, 2011), we chose neurons from these areas
for our initial motility studies. We started with isolating cortical and hippocampal
neurons from H304R embryos (at E16.5) because embryonic neurons are easier
to grow in vitro than the adult mice neurons. Motility studies with established
dynein cargoes such as synaptophysin containing synaptic vesicles (Li et al.,
2000) and Rab7 positive vesicles (Johansson et al., 2007) were performed in
cortical and hippocampal embryonic neuronal cultures.

112
5.2 Results
5.2.1 Synaptophysin-RFP positive vesicular motility in hippocampal neurons
Hippocampal neurons from H304R mice were isolated and plated on coverslips
for intracellular motility experiments. The neurons from the same genotype were
pooled together independent of the gender of the embryo. Hippocampal neurons
were then infected with BacMam synaptophysin-RFP and were imaged at the
ages DIV 10 and 14 (more details in the methods chapter section 2.5). Various
parameters such as the number of movements in the inward (towards the cell
body) and outward (away from the cell body) directions, run length or distance
(µm), and velocity (µm/sec) were recorded of the synaptophysin-RFP positive
vesicles that were moving. The data was then analyzed and statistically
compared between the three genotypes. The wildtype mice are referred as HH,
the heterozygous mice are referred as HR, and the homozygous mice are
referred as RR in this chapter.
At DIV 10, synaptophysin-RFP positive vesicles in homozygous H304R/R mice
showed an unusual distribution of movement patterns i.e. about 83% of vesicles
were moving in the inward direction as compared to about 45% of inward
direction bound vesicles in both the wildtype and heterozygous H304R/+ mice
(Table 5-1). Subsequently, about 17% of vesicles were outward direction bound
in the homozygous H304R/R mice as compared to about 55% outward direction
vesicles in both the wildtype and heterozygous H304R/+ mice.

113
Table 5-1 Synaptophysin-RFP positive vesicles motility in hippocampal neurons at DIV 10
Furthermore, both inward and outward bound synaptophysin-RFP positive
vesicles in heterozygous H304R/+ mice showed a slight (but insignificant)
increase in the distance (µm) covered as compared to the wildtype and
homozygous H304R/R mice (Figure 5-1). Both heterozygous H304R/+ and
homozygous H304R/R mice showed a slight reduction in the velocity (µm/sec) of
the inward and outward bound synaptophysin-RFP positive vesicles as
compared to the wildtype mice, but this reduction was not found to be statistically
significant (Figure 5-1).

114
Figure 5-1 Synaptophysin-RFP positive vesicles motility in hippocampal neurons at DIV 10
The graphs show the distance (µm) covered by the vesicles in the inward (A) and outward (B) bound directions. The graphs show the velocity (µm/sec) of the vesicles in the inward (C) and outward (D) bound directions. Wildtype (black bars), heterozygous H304R/+ mice (gray bars), and homozygous H304R/R mice (blue bars).
At DIV 14, synaptophysin-RFP positive vesicles in homozygous H304R/R mice
showed an altered distribution of movement patterns. About 32% of
Synaptophysin-RFP positive vesicles were moving in inward direction as
compared to about 50-60% of inward bound vesicles in the wildtype and
heterozygous H304R/+ mice (Table 5-2).
A B
C D

115
Subsequently, 68% of synaptophysin-RFP positive vesicles were moving in
outward direction as compared to about 40-50% of outward-bound vesicles in the
wildtype and heterozygous H304R/+ mice.
Table 5-2 Synaptophysin-RFP positive vesicles motility in hippocampal neurons at DIV 14
Interestingly, synaptophysin-RFP positive vesicles in homozygous H304R/R mice
displayed a significant increase in the distance covered (µm) as compared to the
wildtype and heterozygous H304R/+ mice (One-way ANOVA, p<0.05) (Figure 5-
2). Heterozygous H304R/+ mice showed slight but insignificant increase in its
vesicular distance coverage as compared to the wildtype mice. However, the

116
synaptophysin-RFP positive vesicles in heterozygous H304R/+ mice showed a
significant decrease in its velocity (µm/sec) as compared to both wildtype and
homozygous H304R/R mice (One-way ANOVA, p<0.05) (Figure 5-2).
Figure 5-2 Synaptophysin-RFP positive vesicles motility in hippocampal neurons at DIV 14
The graphs show the distance (µm) covered by the vesicles in the inward (A) and outward (B) bound directions. The graphs show the velocity (µm/sec) of the vesicles in the inward (C) and outward (D) bound directions. Wildtype (black bars), heterozygous H304R/+ mice (gray bars), and homozygous H304R/R mice (blue bars). Note: * shows statistical significance (1-way ANOVA, p<0.05).
A B
C D

117
5.2.2 Synaptophysin-RFP positive vesicular motility in cortical neurons
Cortical neurons from H304R mice were isolated and plated on coverslips for
intracellular motility experiments. The neurons from the same genotype were
pooled together independent of the gender of the embryo. Cortical neurons were
then infected with BacMam synaptophysin-RFP and were imaged at the ages
DIV 10 and 14. Various parameters such as the number of movements in the
inward (towards the cell body) and outward (away from the cell body) directions,
run length or distance (µm), and velocity (µm/sec) were recorded of the
synaptophysin-RFP positive vesicles that were moving. The data was then
analyzed and statistically compared between the three genotypes.
At DIV 10, synaptophysin-RFP positive vesicles in both heterozygous H304R/+
and homozygous H304R/R mice showed an altered distribution of movement
patterns. About 63% of the vesicles in H304R mutant mice were moving in the
inward direction as compared to about 82% of inward direction bound vesicles in
the wildtype mice (Table 5-3). Subsequently, about 40% of vesicles were
outward direction bound in both heterozygous H304R/+ and homozygous
H304R/R mice as compared to only about 18% outward direction vesicles in the
wildtype mice.

118
Table 5-3 Synaptophysin-RFP positive vesicles motility in cortical neurons at DIV 10
Furthermore, both inward and outward bound synaptophysin-RFP positive
vesicles in heterozygous H304R/+ and homozygous H304R/R mice showed an
alteration in the distance (µm) covered as compared to the wildtype, but this
alteration was not significantly different statistically (Figure 5-3). Heterozygous
H304R/+ mice showed a slight (but insignificant) increase in the velocity (µm/sec)
of the inward and outward bound synaptophysin-RFP positive vesicles as
compared to the wildtype and homozygous H304R/R mice (Figure 5-3).

119
Figure 5-3 Synaptophysin-RFP positive vesicles motility in cortical neurons at DIV 10
The graphs show the distance (µm) covered by the vesicles in the inward (A) and outward (B) bound directions. The bottom graphs show the velocity (µm/sec) of the vesicles in the inward (C) and outward (D) bound directions. Wildtype (black bars), heterozygous H304R/+ mice (gray bars), and homozygous H304R/R mice (blue bars).
At DIV 14, synaptophysin-RFP positive vesicles showed a similar distribution of
movement patterns in all 3 genotypes. About 50-60% of the vesicles in H304R
mice was moving in the inward direction and about 40-50% of the vesicles were
outward direction bound in all three genotypes (Table 5-4).
A B
C D

120
Table 5-4 Synaptophysin-RFP positive vesicles motility in cortical neurons at DIV 14
Both inward and outward bound synaptophysin-RFP positive vesicles had similar
distance (µm) covered in all 3 genotypes as well (Figure 5-4). Heterozygous
H304R/+ mice showed a peculiar pattern in the velocity (µm/sec) characteristic of
the inward and outward bound synaptophysin-RFP positive vesicles as
compared to the wildtype and homozygous H304R/R mice (Figure 5-4). Velocity
of synaptophysin-RFP positive vesicles was decreased in the inward bound

121
direction and increased in the outward bound direction as compared to both the
wildtype and homozygous H304R/R mice, but this difference was not statistically
significant (Figure 5-4).
Figure 5-4 Synaptophysin-RFP positive vesicles motility in cortical neurons at DIV 14
The graphs show the distance (µm) covered by the vesicles in the inward (A) and outward (B) bound directions. The bottom graphs show the velocity (µm/sec) of the vesicles in the inward (C) and outward (D) bound directions. Wildtype (black bars), heterozygous H304R/+ mice (gray bars), and homozygous H304R/R mice (blue bars).
A B
C D

122
5.2.3 Rab7-GFP positive vesicular motility in cortical neurons
Cortical neurons from H304R mice were isolated and plated on coverslips for
intracellular motility experiments. The neurons from the same genotype were
pooled together independent of the gender of the embryo. Cortical neurons were
nucleofected with Rab7-GFP and were imaged at the age DIV 10. Various
parameters such as the number of movements in the inward (towards the cell
body) and outward (away from the cell body) directions, run length or distance
(µm), and velocity (µm/sec) were recorded of the synaptophysin-RFP positive
vesicles that were moving. The data was then analyzed and statistically
compared between the three genotypes.
At DIV 10, Rab7-GFP positive vesicles in both heterozygous H304R/+ and
homozygous H304R/R mice showed a similar distribution of movement patterns
in both inward and outward directions as compared to the wildtype mice (Table 5-
5). The Rab7-GFP positive vesicles in homozygous H304R/R mice showed an
increase in the distance (µm) covered by them in the inward direction, but this
difference was not significant compared to the wildtype and heterozygous
H304R/+ mice.

123
Table 5-5 Rab7-GFP positive vesicles motility in cortical neurons at DIV 10
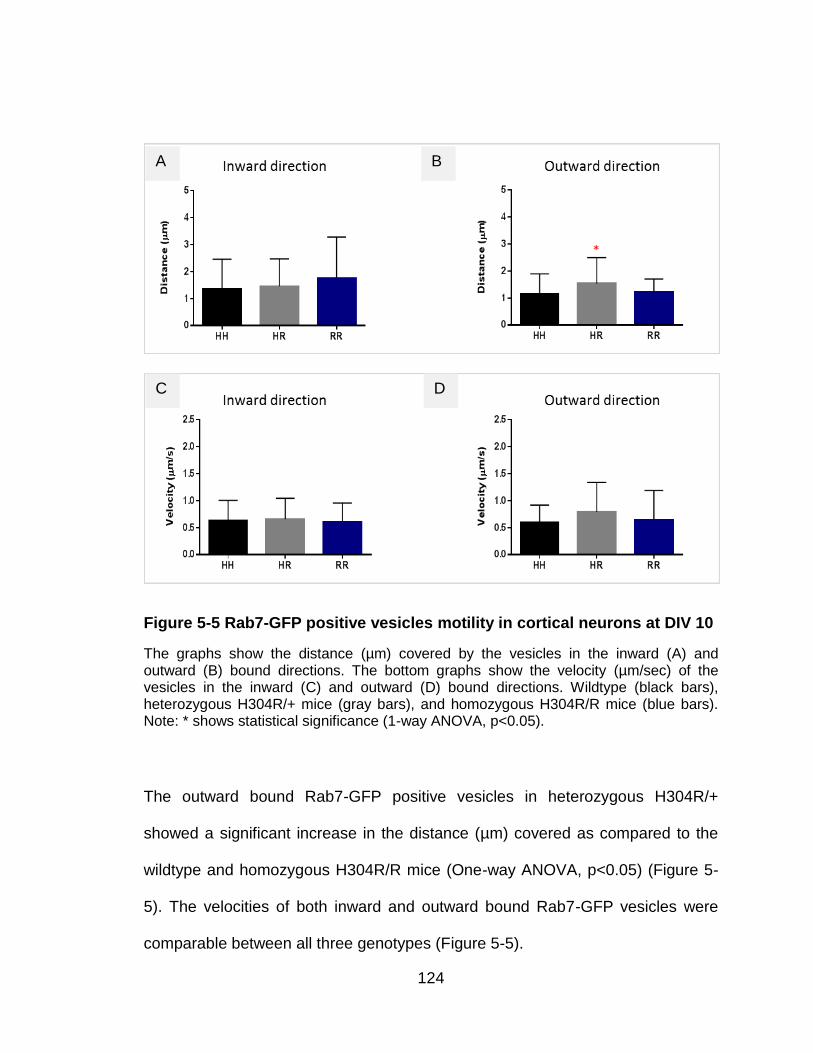
124
Figure 5-5 Rab7-GFP positive vesicles motility in cortical neurons at DIV 10
The graphs show the distance (µm) covered by the vesicles in the inward (A) and outward (B) bound directions. The bottom graphs show the velocity (µm/sec) of the vesicles in the inward (C) and outward (D) bound directions. Wildtype (black bars), heterozygous H304R/+ mice (gray bars), and homozygous H304R/R mice (blue bars). Note: * shows statistical significance (1-way ANOVA, p<0.05).
The outward bound Rab7-GFP positive vesicles in heterozygous H304R/+
showed a significant increase in the distance (µm) covered as compared to the
wildtype and homozygous H304R/R mice (One-way ANOVA, p<0.05) (Figure 5-
5). The velocities of both inward and outward bound Rab7-GFP vesicles were
comparable between all three genotypes (Figure 5-5).
A B
C D

125
5.3 Conclusions
The preliminary motility studies with synaptophysin-RFP and Rab7-GFP positive
vesicles in the hippocampal and cortical primary neurons from the H304R mice
presented highly variable and sometimes contradictory results. There could be
multiple reasons behind these variable results, such as:
Pooling of the primary neurons from the male and female embryos
together for each genotype might have masked any motility differences,
especially in the heterozygous H304R/+ mice neuronal cultures. We
previously showed that the male heterozygous H304R/+ mice are affected
more than the female heterozygous H304R/+ mice in their motor skills
phenotype. A better approach would be to perform the motility studies on
primary neuron cultures that are grown separately based on the genotype
and gender of the embryos.
The primary neurons were collected from the E16.5 developing embryos
that are hard to isolate and grow consistently in vitro.
The motility videos were collected from the longest visible axonal
projection in culture for the data analysis. The classification of “inward”
and “outward” direction bound moving vesicles may not be accurate if the
longest visible axonal projection was not an axon. Only the axons are
known to have an established microtubule polarity and the dendrites have
microtubules arranged in a random fashion with mixed polarities. Hence,

126
this could have introduced another source of variability in the direction
based motility data comparison.
The motility analysis was done on the primary neurons that were grown in
vitro for 10 (DIV 10) or 14 (DIV 14) days. Performing motility analysis at a
range of early and late time points might reveal some alterations in the
motility phenotype.
In conclusion, performing future motility experiments with the abovementioned
considerations might help resolve the variability issue observed in the preliminary
study results. Additionally, at the time of this preliminary study we chose neurons
from the central nervous system (CNS) due to the established experimental
protocols in CNS neurons. Future motility studies should also be performed on
the motor and sensory neurons from the peripheral nervous system in the H304R
mouse model.

127
CHAPTER 6: DISCUSSION
6.1 All dynein mutations don’t act the same
The motor protein dynein is responsible for retrograde transport of crucial
cargoes from the axonal tip to the soma in neurons. Various studies show the
specific role of dynein in neuronal functions such axonal transport (He et al.,
2005), retrograde signaling (Perlson et al., 2009), protein degradation and
recycling (Ligon et al., 2005), and neuronal migration (Ori-McKenney and Vallee,
2011). It is evident that neurons need a robust retrograde mediated transport for
its growth, maintenance, and survival. Hence, it is not surprising to see that
several mutations in cytoplasmic dynein have been reported in human
neurodegenerative diseases. These dynein mutations (Table 1-2) are associated
with motor and sensory neuropathies such as Charcot-Marie-Tooth type 2O
(CMT2O) (Weedon et al., 2011), Spinal Muscular Atrophy with Lower Extremity
Dominant (SMA-LED) (Harms et al., 2012) (Tsurusaki et al., 2012), in
malformations of cortical development (MCD) (Poirier et al., 2013), in intellectual
disability (Willemsen et al., 2012), and congenital cataract and gut dysmotility
(Gelineau-Morel et al., 2016).
The spectrum of neurodegenerative diseases in humans point out the
susceptibility of neurons to the dynein mutations in the peripheral and central
nervous systems. Perhaps, it is the distinct structure and function of neurons that
make them very sensitive to the altered functions of mutant dynein. For example,

128
the motor neurons have very long axons that can grow up to a meter in humans
and require robust axonal transport for its maintenance (Levy and Holzbaur,
2006). Not surprisingly, about 18 distinct mutations in dynein heavy chain are
reported thus far in SMA-LED patients with peripheral motor neuron loss (Scoto
et al., 2015). The same study also reported about 16 mutations in dynein heavy
chain associated with malformations of cortical development (MCD), signifying
the vulnerability of neurons in the central nervous system to the mutant dynein
functions.
The dynein heavy chain (DHC) mutations in motor domain and tail domain alter
the dynein function differentially, thus giving rise to a spectrum of human
neuropathies spanning both the peripheral and central nervous systems in
humans (Scoto et al., 2015). Interestingly, the same DHC mutation can cause
two different diseases as well. For examples, the H306R mutation in DHC is
associated with both CMT2O (Weedon et al., 2011) and SMA-LED (Tsurusaki et
al., 2012). In fact, both CMT2O and SMA-LED patients exhibit similar clinical
symptoms such as muscle weakness and wasting in legs, developmental
milestone delays, skeletal deformities, joint contractures, and gait problems. The
only difference is that the CMT2O patients also exhibit sensory loss symptoms, in
addition to the motor deficits.
Various mutations exist in the DHC peptide with overlapping phenotypes, but
they all do not cause just one exact same neuropathy in humans. Rather these
DHC mutations cause different type of neuropathies, thus signaling that the

129
mutations alter proteins in different ways. Our previous studies with filamentous
fungi N.crassa also recapitulated similar observations where various dynein
mutations resulted in distinct fungal morphology and vesicular motility
(Sivagurunathan et al., 2012). Because different dynein mutations do not behave
the same and cause the same phenotype in humans, there is a possibility that
there might be a distinct molecular mechanism of disease behind each dynein
mutation.
6.2 Relevance of H304R Mouse Model
Relevance of heterozygous H304R/+ mice
One key feature of Charcot-Marie-Tooth type 2O (CMT2O) disease is the
heterogeneity of its onset and the severity of the clinical symptoms in patients.
Family members with the same CMT2O dynein mutation reported variable ages
of onset and the severity of the clinical symptoms expressed. Similar trends were
observed in our study with the heterozygous H304R/+ mice behavioral and
phenotypic characteristics. The heterozygous H304R/+ mice showed variable
onset of phenotype, i.e. early onset of reduced all limbs grip strength and rotorod
test defects, while late onset of reduced front grip strength and tail suspension
reflex. Some heterozygous H304R/+ mice expressed a mild phenotype, whereas
some expressed a more severe phenotype just like as seen in the human
patients with the corresponding autosomal dominant CMT2O mutation.

130
A possible explanation to this heterogeneity might lie in the fact that dynein
heavy chains (DHC) dimerize with each other and interact with other subunits
(like intermediate chains, light intermediate chain, & light chains) to form a
functional dynein motor complex. In the heterozygous H304R/+ mice, there
would be a mix of DHC monomers that have been translated from the
heterozygous DHC alleles (H304R/+). From the mix of wildtype and H304R
mutant DHC monomers, three possible combinations of dynein population may
arise –
A quarter population of dynein containing wildtype DHC dimers only
A quarter population of dynein containing mutant DHC dimers only
And half of the population of dynein containing DHC dimers of 1 wildtype
monomer dimerized with 1 mutant DHC monomer
Thus, at any given time, a heterozygous H304R/+ mouse cells will have all three
types of DHC dimers combination – wildtype, homozygous, and heterozygous
DHC dimers. There could be a “dose-dependent” modulation of dynein motor
function depending on what percentage of wildtype, homozygous, and
heterozygous DHC dimers are present and interacting functionally with other
dynein subunits at a given time. The variable level of DHC dimers may also affect
the interaction of the dynein motor complex with its other binding partners and/or
cargoes. Future biochemical tests with purified dynein from H304R mouse model
can reveal key insights on the cellular dynamics that happen when different
“doses” of wildtype, homozygous, heterozygous DHC dimers are around in a cell.

131
One key experiment will be to test the binding efficiency of wildtype,
homozygous, and heterozygous DHC dimers with other dynein subunits and key
binding partners of dynein such as dynactin, BicD, Lis1, Nudel etc..
Another key observation in the heterozygous H304R/+ mice was the gender-
based differences in the exhibition of the motor skills phenotypic defects. Overall,
the male heterozygous H304R/+ mice appeared to be more affected by the
dynein mutation as evident in the various motor-skills assays. In contrast, the
female heterozygous H304R/+ mice showed mild to no defects in the exhibition
of the motor skills phenotypic defects. A potential reason behind this gender-
based observation could be the presence of the neuroprotective effects of
estrogen in the female mice (Green and Simpkins 2000). Thus, the female
heterozygous H304R/+ mice appear to be less sensitive to the effects of the
dynein mutation on their motor skills phenotype. Similar observations were made
in a small study with CMT1A patients where the male patients were shown to
have significantly reduced muscle strength in their lower limbs as compared to
the female patients (Wozniak and Gess, 2015).
Relevance of homozygous H304R/R mice
The homozygous H304R/R mice exhibited severe symptoms of atypical tail
suspension, reduced grip strength and rotorod deficits right from an early age.
The onset and the severity of homozygous H304R/R phenotype recapitulated the
severity of SMA-LED symptoms as seen in another and the only reported

132
homozygous DHC mutation (R399G) (Scoto et al., 2015). Offspring with this
DHC homozygous R399G mutation showed an early onset and severe motor
symptoms as compared to the parents that were heterozygous for the same DHC
mutation.
Furthermore, both male and female homozygous H304R/R mice showed similar
trends of deficiencies in their behavioral phenotype, unlike the heterozygous
H304R/+ mice that showed gender-based deficiencies in their phenotype
(Sabblah et al., 2018). As in the previous publication, only the male heterozygous
H304R/+ mice primarily exhibited an altered motor skills phenotype in most
behavioral tests. The female heterozygous H304R/+ mice exhibited little to no
difference in its behavioral phenotype as compared to the wildtype mice. This key
difference between the heterozygous H304R/+ and homozygous H304R/R mice
indicates that the homozygous H304R/R mutation has a distinct and more severe
effect on the dynein function that may not altered by the gender differences of the
mice.
Overall relevance of H304R mice in understanding the dynein function
In conclusion, both heterozygous H304R/+ and homozygous H304R/R mice with
DHC mutations recapitulate the onset and the phenotypes as seen with the
reported heterozygous (H306R) and homozygous (R399G) dynein mutations in
humans. Hence, our H304R mouse model would serve as a relevant and useful

133
mouse model to understand the molecular mechanism of mutant dynein in
human neuropathies.
6.3 Comparison of H304R Mouse Model with Other Dynein Mouse Models
Previously reported mutant DHC mice were generated using ENU or radiation
based mutagenesis method that exhibited dominant neurological phenotypes
(Hrabe de Angelis et al., 2000) (Duchen, 1974). The mutant DHC mice were
Legs at odd angle (Loa) mice with F580Y mutation, Cramping (Cra) mice with
Y1055C mutation, and Sprawling (Swl) mice with a nine base-pair deletion
[GIVT]1040[A] mutation in the cytoplasmic dynein heavy chain gene. Even
though all three mutations were located in the tail domain of the dynein heavy
chain, the Loa, Cra and Swl did not display the same exact phenotype.
A key difference is that the H304R mouse model has the corresponding dynein
mutation (H306R) that is reported to cause neuropathies in humans (CMT2O and
SMA-LED) (Tsurusaki et al., 2012; Weedon et al., 2011). In contrast, the
mutagenesis generated Loa, Cra, and Swl mice models do not have a
corresponding mutation that is associated with any human neuropathies. Hence,
it is hard to correlate their findings as a basis for human neuropathy. Another key
difference between H304R mice and other DHC mice is that homozygous
Loa/Loa, Cra/Cra, and Swl/Swl were embryonically lethal or did not survive 24
hours post birth due to inability to move and feed (Hafezparast et al., 2003)

134
(Zhao et al., 2016). In contrast, the homozygous H304R/R mice survived post
birth and had a normal lifespan.
The H304R mouse model shares many commonalities with other published
models of dynein heavy chain mutation Loa, Cra, and Swl. The first commonality
is that all the mutations are located in the tail domain of the dynein heavy chain.
Shared phenotypes between H304R model and Loa/+, Cra/+ and Swl/+ mice are
– atypical reflex of clenching hind limbs closer to the body upon tail suspension,
normal life span, gait abnormalities, reduced grip strength in hind limbs, and
thinner sciatic nerves. Additionally, H304R/+ and H304R/R mice showed deficits
in rotarod performance similar to Loa/+ mice.
In conclusion, the H304R model and previously published DHC mice model
display a varying range of phenotypes. The severity of the mouse DHC mutations
on the dynein function varies in the following order – the homozygous Loa/Loa,
Cra/Cra and Swl/Swl are the most severe mutations, followed by the
homozygous H304R/R and heterozygous Loa/+, Cra/+ and Swl/+ moderate
mutations, and finally the heterozygous H304R/+ mild mutation (Figure 6-1).

135
Figure 6-1 Severity of phenotypes associated with DHC mutations
Comparison of the severity of mouse DHC mutations with the human DHC mutations. The severity of the
phenotype associated with the DHC mutation increases from left to the right.
MOUSE
MUTATIONS
HUMAN MUTATIONS
Loa/+
Cra/+
Swl/+ H304R/+ H304R/R
H306R/+
(CMT2O)
Loa/Loa
Cra/Cra
Swl/Swl
LETHAL
R399G/G
No Mutation
NO SYMPTOM
No Mutation

136
6.4 Future Sensory Characterization Work of H304R Mouse Model
Collectively behavioral assessments like tail suspension, grip strength, and
rotarod performance tests have given us some robust insights into the motor
phenotype of H304R mouse model. Neuromuscular junction (NMJ) analysis gave
additional evidence supporting the defective motor phenotype of H304R mouse
model (Sabblah et al., 2018). NMJ analysis revealed the deficiencies in the
morphology and complexity in heterozygous H304R/+ and homozygous
H304R/R male mice. The synaptophysin occupancy in NMJs were also found to
be faulty, indicating a loss of motor neuron functionality in heterozygous H304R/+
and homozygous H304R/R male mice. The reason why the motor neurons are
not functional in mutant H304R mice is yet to be explored.
However, investigating the motor characterization is only one part of the equation
in understanding a motor-sensory neuropathy like CMT2O. Multiple studies have
indicated dynein’s role in sensory neuron maintenance and survival (Deinhardt et
al. (2006); (Saxena et al., 2005; Yano et al., 2001). In parallel, the previously
published dynein heavy chain mutant Loa/+, Cra/+ and Swl/+ mice have also
reported sensory deficits. Heterozygote Loa/+ and Swl/+ mice reportedly had
reduced size of dorsal root and fewer neurons in their lumbar dorsal root ganglia
(Chen et al., 2007). Furthermore, Loa/+ and Swl/+ also showed abnormal
proprioceptive phenotype as indicated by the absence of H-reflex and specific
loss of proprioceptive sensory neurons as compared to the wildtype mice.

137
Interestingly, Loa/+ and Swl/+ mice also had thinner sciatic nerves as compared
to the wildtype control mice. The Swl/+ mice showed no difference in the latency
to tail withdrawal as compared to the wildtype control mice. The Cra/+ mice
displayed subtler sensory phenotype than the Loa/+ and Swl/+ mice and had
reduced size of its dorsal root due to the loss of large caliber axon (potentially
proprioceptive) neurons (Dupuis et al. (2009). In conclusion, CMT2O patients
and all the other dynein mutant mice (Loa/+, Cra/+, and Swl/+) showed mild to
severe sensory phenotype and subsequently raised a question on the sensory
phenotype of the H304R mouse model. Hence, I started investigating the
sensory phenotype of H304R mice with tail flick test, sciatic nerve thickness
measurements, and optimizing the protocol to identify the dorsal root ganglia
neuronal subpopulation.
Preliminary results from the tail flick tests showed a quicker latency to tail
withdrawal from the pain stimulus in the homozygous H304R/R mice. One
possible explanation is that the homozygous H304R/R mice have hypersensitivity
to thermal pain stimulus due to reduced pain threshold as commonly seen in
mice models with sciatic nerve injury and persistent sensory neuropathic pain
(Wang et al., 2009; Yajima et al., 2002). Incidentally, severely affected CMT2O
patients also reported having neuropathic pain (Weedon et al., 2011).
Next, a closer examination of peripheral nervous system revealed thinner sciatic
nerves and reduced axonal counts in both the heterozygous H304R/+ and
homozygous H304R/R mice. It would be interesting to learn if the sciatic nerve

138
thinning in homozygous H304R/R is caused by the loss of motor and/or sensory
neurons. One way to test this would be to stain the sciatic nerve cross sections
with unique markers for both motor and sensory neurons. Then the number of
motor and sensory axons passing through the sciatic nerve can be counted and
normalized against the total number of neurons (labeled with a pan-neuronal
marker). This would help in identifying if any alteration in the number of motor
and/or sensory neurons led to thinner sciatic nerve in homozygous H304R/R
mutant mice.
To advance the sensory characterization, it would be of critical importance to
identify the neuronal subpopulation inside the dorsal root ganglia (DRG) as well
of the H304R mouse model. Previous work on Swl/+ mice revealed a significant
loss of proprioceptive neurons in the lumbar DRG (Chen et al., 2007). My
optimization work established the working protocol of how to stain and identify
nociceptive and proprioceptive subpopulations in the same section of the DRG of
H304R mice. The next step would be to stain DRG sections from the wildtype,
heterozygous H304R/+, and homozygous H304R/R mice with the unique
markers for each neuronal subpopulation. Then the number of neuronal cells
positive for nociceptive marker or proprioceptive marker can be counted and
normalized over the total count of neurons in the DRG. The results would yield if
there are any differences in the neuronal subpopulations count in the H304R
mouse model, and potentially reveal the neuropathology of mutant dynein in
sensory dysfunction.

139
6.5 Potential Dynein Disruptions in the H304R Mice and Future Studies
Our preliminary studies revealed both motor and sensory defects in our H304R
mouse model. Because large sized motor and sensory neurons have high
demands for energy and require robust axonal transport, it is not surprising that
motor and sensory neurons are vulnerable to the mutations that may alter the
dynein functions in retrograde axonal transports (LaMonte et al., 2002; Ori-
McKenney et al., 2010). Dynein has an established role in transporting nerve
growth factors via activated trk signaling from the neuronal tip to the soma for
neuronal survival (Heerssen et al., 2004; Mitchell et al., 2012; Yano et al., 2001).
Impaired retrograde signaling of growth factors or neurotropic factors in sensory
neurons has been shown to develop peripheral neuropathy phenotypes in mice
(Rishal et al., 2012). Based on this literature evidence, I speculate three different
potential mechanism of the H304R mutation in the disease progression.
The first disease mechanism is that the dynein mutation affects the retrograde
signaling of endosomes with the neurotrophic factors. This would lead to a loss of
sensory neurons in the H304R mice. As evident from my preliminary work with
the sensory phenotype characterization, I think the nociceptive (pain sensing)
subpopulation of the sensory neurons might be affected more as seen in the
quick tail withdrawal latency in the mutant H304R mice. Hence, impaired
retrograde signaling leading to the loss of nociceptive sensory neurons ultimately
causes the altered sensory phenotypes such as altered tail withdrawal, thinner
sciatic nerves, and a loss of axonal counts in the heterozygous H304R/+ and

140
homozygous H304R/R mice. Investigating the robustness of retrograde signaling
of trk receptors upon activation of growth factors such as brain derived
neurotrophic factors (BDNF), nerve growth factor (NGF), or neurotrophins (NTs)
would be of critical importance in understanding the role of mutant dynein in the
H304R mice.
The second disease mechanism is the impaired retrograde transport of length
sensing cellular factors leading to an altered growth of sensory neurons. This is
based on an interesting study that revealed a role of dynein in maintaining the
sensory neurons length (Ori-McKenney et al., 2010). In this study, adult sensory
neurons in the heterozygote Loa/+ mice had longer neurites than the wildtype
controls when grown in vitro. Additionally, heterozygote Loa/+ mice showed a
similar trend of significantly longer axis from cell periphery to the nucleus in
embryonic fibroblasts as compared to wildtype controls. These results indicated
that dynein might have a role in transporting the length sensing cellular factors
needed for sensory neuron axonal length maintenance. Based on the evidence
presented above I speculate that the H304R mice may have an impaired
retrograde transport of length sensing cellular factors. And that this may lead to
an alteration in the sensory neuron lengths. Hence, it would be an exciting idea
to measure and compare the lengths of the sensory neurons in vitro from the
H304R mice to reveal any alterations in the sensory neuron lengths.
Lastly, the third possible mechanism for the dynein dysfunction in the H304R
mice could be a biochemical interaction defect based on the location of the

141
H304R mutation on the dynein heavy chain (DHC) peptide. The H304R mutation
is located in the tail domain of the DHC peptide. The H304R mutation lies within
the homodimerization region of the tail domain in the DHC. The presence of the
H304R mutation in the homodimerization region may cause a hindrance to the
binding of two DHC monomers to form a functional DHC dimer. The H304R
mutation could also contribute to interference in the interactions of the DHC to
other dynein subunits like dynein intermediate chains (DICs) and dynein light
chains (DLCs). The interactions with DICs and DLCs allow for the indirect
interaction of the DHCs to its binding factors such as dynactin, Lis1, BicD and
these binding factors have been implicated in the proper functioning of the
neurons such as neurofilament transport, clearance of misfolded proteins, and
neuronal migration. Thus, it would be insightful to perform future studies to
investigate the functional binding of the mutant DHC with other dynein subunits
and binding factors using biochemical assays. These biochemical assays may
potentially uncover any altered dynein interaction that may contribute to the
pathogenesis of the disease as discussed above.

142
APPENDIX A: DISSERTATION DEFENSE ANNOUCEMENT

143
Announcing the Final Examination of Ms. Swaran Nandini for the degree of Doctor
of Philosophy in Biomedical Sciences
Dissertation Title: “MOTOR AND SENSORY CHARACTERIZATION OF A
MOUSE MODEL OF CHARCOT-MARIE-TOOTH TYPE 2O DISEASE”
Date: 04/03/2019 Time: 3:00pm
Location: Lake Nona BBS 103 (Live)/ BMS 136 (Simulcast)
ABSTRACT
Dynein is an essential motor protein required for the maintenance and survival of the
cells. Dynein forms a motor complex to carry intracellular cargoes like organelles,
growth factors, peptides, hormones etc. along the microtubules inside the cells. In
neurons, the dynein is the retrograde motor protein that moves cargoes from the neuronal
tip to the neuronal soma along the length of an axon. Dynein has an established role in
the neuronal nuclei migration, transport of neuronal survival signals and growth factors,
organelle positioning inside neurons etc. Hence, it is not very surprising that numerous
mutations in dynein have been reported in association with neurodegenerative diseases in
humans. The first mutation (H306R) in dynein heavy chain was reported to cause
Charcot-Marie-Tooth Type 2O disease (CMT2O) in humans. CMT2O patients display
motor-sensory neuropathy symptoms such as muscle weaknesses and wasting in legs,
skeletal deformities like pes cavus (high foot arching), difficulty in walking, and a loss of
sensation.
We developed a novel knock-in H304R mouse model with the corresponding CMT2O
linked dynein mutation to understand the disease’s molecular mechanism. We
investigated and characterized the motor-sensory phenotype of the H304R mouse model
(wildtype, heterozygous (H304R/+) and homozygous (H304R/R) mice). First, we started
with testing mice on motor skills behavior tests such as tail suspension reflex, grip
strength test, and rotarod test at 3, 6, 9 and 12 months of age. Both male and female
groups of heterozygous (H304R/+) mice displayed mild defects in tail suspension reflex,
grip strength, and rotarod performance at a later age time point. In contrast, homozygous
(H304R/R) mice exhibited severe defects in the tail suspension reflex, grip strength, and
rotarod performance right from an early age. Next, I analyzed the sensory phenotype of
the H304R mouse model. Homozygous H304R/R mice appeared to have thinner sciatic
nerve, reduced total fascicular area of the sciatic nerve, and significantly quicker latency
to tail withdrawal from a pain stimulus.
Collectively, our motor and sensory characterization studies reveal that H304R dynein
mouse model recapitulates the phenotype associated with CMT symptoms. Hence, the
H304R model is a useful tool in understanding the dynein function in the onset and
progression of CMT2O in humans.

144
Committee members:
Stephen J. King, Ph.D. (Committee Chair)
Annette Khaled, Ph.D.
Yoon-Seong Kim, M.D., Ph.D.
Stephen Lambert, Ph.D.
Publications:
Nandini, S.*, Conley Calderon, J.*, Sabblah, T. T., Love, R., King, L. E., King,
S. J. (2019). Mice with an autosomal dominant Charcot-Marie-Tooth type 2O
disease mutation in both dynein alleles display severe moto-sensory phenotypes.
(Submitted to Scientific Reports, * equal contribution)
Sabblah, T. T.*, Nandini, S.*, Ledray, A. P., Pasos, J., Conley Calderon, J., Love,
R., King, L. E., King, S. J. (2018). A novel mouse model carrying a human
cytoplasmic dynein mutation shows motor behavior deficits consistent with
Charcot-Marie-Tooth type 2O disease. Scientific reports, 8(1), 1739. (* equal
contribution)
Höing S, Yeh TY, Baumann M, Martinez NE, Habenberger P, Kremer L, Drexler
HCA, Küchler P, Reinhardt P, Choidas A, Zischinsky ML, Zischinsky G,
Nandini S., Ledray AP, Ketcham SA, Reinhardt L, Abo-Rady M, Glatza M, King
SJ, Nussbaumer P, Ziegler S, Klebl B, Schroer TA, Schöler HR, Waldmann H,
Sterneckert J. (2018) Dynarrestin, a Novel Inhibitor of Cytoplasmic Dynein. Cell
Chem Biol.,25(4), 357-369.
Sivagurunathan, S., Schnittker, R. R., Nandini, S., Plamann, M. D., & King, S. J.
(2012). A mouse neurodegenerative dynein heavy chain mutation alters dynein
motility and localization in Neurospora crassa. Cytoskeleton (Hoboken, N.J.),
69(9), 613-24.
Sivagurunathan, S., Schnittker, R. R., Razafsky, D. S., Nandini, S., Plamann, M.
D., & King, S. J. (2012). Analyses of dynein heavy chain mutations reveal
complex interactions between dynein motor domains and cellular dynein
functions. Genetics, 191(4), 1157-79.
Approved for distribution by Dr. Stephen J. King, Committee Chair
The public is welcome to attend.

145
APPENDIX B: RESUME

146
SWARAN NANDINI
EDUCATION & RESEARCH EXPERIENCES
Doctor of Philosophy - Biomedical Sciences, College of Medicine, University of Central Florida, FL o Received the UCF Graduate Dean's Dissertation Completion Fellowship o 7 research awards/scholarships, published 4 research papers & 5th paper
in submission, presented 18 research posters & 4 research talks
2013 - 2019
Master of Science - Biotechnology, College of Medicine, University of Central Florida (UCF), FL o Received the UCF COM Outstanding Master’s Thesis Award
2011 - 2013
Bachelor of Technology - Biotechnology, Jaypee Institute of Information Technology, Delhi, India o Completed Thesis research project and 2 summer research internships
2007 - 2011
LEADERSHIP EXPERIENCES (TOP 5 OUT OF 8)
American Society for Cell Biology (ASCB) – Ambassador & Regional Symposium Organizer o Involved in Annual conference promotion, moderated sessions, onsite
conference assistance o Partnered with the host school to organize the 2nd Annual FL Cell Biology
Symposium for 130+ attendees, recruited poster presenters & panel speakers, hosted career development workshop
2014 - 2019
University of Central Florida Recreation and Wellness Center (RWC) – Health & Fitness Intern o Achieved 100% client retention rate as a Group exercise instructor o Audited facility risks & developed creative solutions to mitigate risks (risk
management efforts) o Executed strategic projects to grow the diverse representation in the
RWC staff employment, and presented on LGBTQ+ clients customer service training (diversity & inclusion efforts)
2016 - 2019
University of Central Florida Technology Fee Committee – Graduate Student Representative o Reviewed 233 tech proposals to allocate committee funds of $22 million
2016 - 2019
National Association of Graduate Professional Students (NAGPS) – Vice Chair & Conference Chair o Organized the 2016 NAGPS Southeastern (SE) Regional Conference that
was hosted for the 1st time in FL, achieved the highest SE regional conference attendance in 4 years with participation from 7 out-of-state guest schools
2015 - 2017

147
Biomedical Sciences Graduate Student Association (BSGSA) at UCF – Student President o Achieved the highest student member enrollment ever o Recruited 6 guest speakers to talk on varied topics (bioentrepreneurship,
academia, industry) o Successfully led a cross functional team to organize the first ever 2-day
Career Symposium with limited budget, sold out registration, 1 attendee received an employment offer out of it
2014 - 2015
AWARDS
Received 16 awards and scholarships totaling $15,400 in amount. Key
highlights here-
2012 - 2019
o Graduate Dean's Dissertation Completion Fellowship University of Central Florida (UCF)
2019
o Risk Management Champion of the year UCF Recreation and Wellness Center
2018
o Diversity & Inclusion Champion of the year UCF Recreation and Wellness Center
2017
o Region II Top Educational Session Award National Intramural Recreational Sports Association
2016
o Volunteer UCF Service Award University of Central Florida (UCF)
2015
o Graduate Student Travel Award American Society for Cell Biology Education Committee
2014
COMMUNITY SERVICE
Biology Integrated Orlando Training and Enrichment Camp (BIOTEC) – Organizer & instructor Collaborated with multi-specialty teams to host 4 science outreach camps for high school students
2015 - 2018
Volunteer UCF – Event organizer & marketing specialist Led 2 different taskforce to organize two 24-hr long community service events for 200+ attendees
2015 - 2017
YMCA of Central Florida Teen Achievers Program – STEM Careers Outreach at a local high school
2015 - 2016

148
REFERENCES
Ahmad, F.J., Echeverri, C.J., Vallee, R.B., and Baas, P.W. (1998). Cytoplasmic dynein and
dynactin are required for the transport of microtubules into the axon. J Cell Biol 140,
391-401.
Amaya, F., Shimosato, G., Nagano, M., Ueda, M., Hashimoto, S., Tanaka, Y., Suzuki, H.,
and Tanaka, M. (2004). NGF and GDNF differentially regulate TRPV1 expression that
contributes to development of inflammatory thermal hyperalgesia. The European
journal of neuroscience 20, 2303-2310.
Banks, G.T., and Fisher, E.M. (2008). Cytoplasmic dynein could be key to understanding
neurodegeneration. Genome Biol 9, 214.
Barakat-Walter, I., and Riederer, B.M. (1996). Triiodothyronine and nerve growth factor
are required to induce cytoplasmic dynein expression in rat dorsal root ganglion
cultures. Brain Res Dev Brain Res 96, 109-119.
Bennett, D.L., Dmietrieva, N., Priestley, J.V., Clary, D., and McMahon, S.B. (1996). trkA,
CGRP and IB4 expression in retrogradely labelled cutaneous and visceral primary
sensory neurones in the rat. Neuroscience letters 206, 33-36.
Bergmann, I., Priestley, J.V., McMahon, S.B., Brocker, E.B., Toyka, K.V., and Koltzenburg,
M. (1997). Analysis of cutaneous sensory neurons in transgenic mice lacking the low
affinity neurotrophin receptor p75. The European journal of neuroscience 9, 18-28.
Bird, T.D. (1993). Charcot-Marie-Tooth (CMT) Hereditary Neuropathy Overview. In
GeneReviews((R)), M.P. Adam, H.H. Ardinger, R.A. Pagon, S.E. Wallace, L.J.H. Bean, K.
Stephens, and A. Amemiya, eds. (Seattle (WA)).
Blocker, A., Severin, F.F., Burkhardt, J.K., Bingham, J.B., Yu, H., Olivo, J.C., Schroer, T.A.,
Hyman, A.A., and Griffiths, G. (1997). Molecular requirements for bi-directional
movement of phagosomes along microtubules. J Cell Biol 137, 113-129.
Bremner, K.H., Scherer, J., Yi, J., Vershinin, M., Gross, S.P., and Vallee, R.B. (2009).
Adenovirus transport via direct interaction of cytoplasmic dynein with the viral capsid
hexon subunit. Cell Host Microbe 6, 523-535.

149
Burgess, S.A., Walker, M.L., Sakakibara, H., Knight, P.J., and Oiwa, K. (2003). Dynein
structure and power stroke. Nature 421, 715-718.
Burgess, S.A., Walker, M.L., Sakakibara, H., Oiwa, K., and Knight, P.J. (2004). The
structure of dynein-c by negative stain electron microscopy. J Struct Biol 146, 205-216.
Burkhardt, J.K., Echeverri, C.J., Nilsson, T., and Vallee, R.B. (1997). Overexpression of the
dynamitin (p50) subunit of the dynactin complex disrupts dynein-dependent
maintenance of membrane organelle distribution. J Cell Biol 139, 469-484.
Cai, Q., Lu, L., Tian, J.H., Zhu, Y.B., Qiao, H., and Sheng, Z.H. (2010). Snapin-regulated late
endosomal transport is critical for efficient autophagy-lysosomal function in neurons.
Neuron 68, 73-86.
Campenot, R.B. (1977). Local control of neurite development by nerve growth factor.
Proceedings of the National Academy of Sciences of the United States of America 74,
4516-4519.
Cantalupo, G., Alifano, P., Roberti, V., Bruni, C.B., and Bucci, C. (2001). Rab-interacting
lysosomal protein (RILP): the Rab7 effector required for transport to lysosomes. EMBO J
20, 683-693.
Caviston, J.P., and Holzbaur, E.L. (2009). Huntingtin as an essential integrator of
intracellular vesicular trafficking. Trends Cell Biol 19, 147-155.
Cellerino, A., Maffei, L., and Domenici, L. (1996). The distribution of brain-derived
neurotrophic factor and its receptor trkB in parvalbumin-containing neurons of the rat
visual cortex. The European journal of neuroscience 8, 1190-1197.
Chao, M.V. (2003). Neurotrophins and their receptors: a convergence point for many
signalling pathways. Nat Rev Neurosci 4, 299-309.
Chen, X.J., Levedakou, E.N., Millen, K.J., Wollmann, R.L., Soliven, B., and Popko, B.
(2007). Proprioceptive sensory neuropathy in mice with a mutation in the cytoplasmic
Dynein heavy chain 1 gene. The Journal of neuroscience : the official journal of the
Society for Neuroscience 27, 14515-14524.

150
Cheng, H.H., Liu, S.H., Lee, H.C., Lin, Y.S., Huang, Z.H., Hsu, C.I., Chen, Y.C., and Chang,
Y.C. (2006). Heavy chain of cytoplasmic dynein is a major component of the postsynaptic
density fraction. J Neurosci Res 84, 244-254.
Culver-Hanlon, T.L., Lex, S.A., Stephens, A.D., Quintyne, N.J., and King, S.J. (2006). A
microtubule-binding domain in dynactin increases dynein processivity by skating along
microtubules. Nat Cell Biol 8, 264-270.
Deinhardt, K., Salinas, S., Verastegui, C., Watson, R., Worth, D., Hanrahan, S., Bucci, C.,
and Schiavo, G. (2006). Rab5 and Rab7 control endocytic sorting along the axonal
retrograde transport pathway. Neuron 52, 293-305.
Delcroix, J.D., Valletta, J.S., Wu, C., Hunt, S.J., Kowal, A.S., and Mobley, W.C. (2003). NGF
signaling in sensory neurons: evidence that early endosomes carry NGF retrograde
signals. Neuron 39, 69-84.
Dick, T., Ray, K., Salz, H.K., and Chia, W. (1996). Cytoplasmic dynein (ddlc1) mutations
cause morphogenetic defects and apoptotic cell death in Drosophila melanogaster. Mol
Cell Biol 16, 1966-1977.
Dodding, M.P., and Way, M. (2011). Coupling viruses to dynein and kinesin-1. EMBO J
30, 3527-3539.
Driskell, O.J., Mironov, A., Allan, V.J., and Woodman, P.G. (2007). Dynein is required for
receptor sorting and the morphogenesis of early endosomes. Nat Cell Biol 9, 113-120.
Duchen, L.W. (1974). A dominant hereditary sensory disorder in the mouse with
deficiency of muscle spindles: the mutant Sprawling. J Physiol 237, 10P-11P.
Dujardin, D., Wacker, U.I., Moreau, A., Schroer, T.A., Rickard, J.E., and De Mey, J.R.
(1998). Evidence for a role of CLIP-170 in the establishment of metaphase chromosome
alignment. J Cell Biol 141, 849-862.
Dupuis, L., Fergani, A., Braunstein, K.E., Eschbach, J., Holl, N., Rene, F., Gonzalez De
Aguilar, J.L., Zoerner, B., Schwalenstocker, B., Ludolph, A.C., et al. (2009). Mice with a

151
mutation in the dynein heavy chain 1 gene display sensory neuropathy but lack motor
neuron disease. Experimental neurology 215, 146-152.
Engelender, S., Sharp, A.H., Colomer, V., Tokito, M.K., Lanahan, A., Worley, P., Holzbaur,
E.L., and Ross, C.A. (1997). Huntingtin-associated protein 1 (HAP1) interacts with the
p150Glued subunit of dynactin. Hum Mol Genet 6, 2205-2212.
Fu, M.M., and Holzbaur, E.L. (2013). JIP1 regulates the directionality of APP axonal
transport by coordinating kinesin and dynein motors. J Cell Biol 202, 495-508.
Gelineau-Morel, R., Lukacs, M., Weaver, K.N., Hufnagel, R.B., Gilbert, D.L., and
Stottmann, R.W. (2016). Congenital Cataracts and Gut Dysmotility in a DYNC1H1
Dyneinopathy Patient. Genes (Basel) 7.
Gibbons, I.R., Gibbons, B.H., Mocz, G., and Asai, D.J. (1991). Multiple nucleotide-binding
sites in the sequence of dynein beta heavy chain. Nature 352, 640-643.
Gibbons, I.R., and Rowe, A.J. (1965). Dynein: A Protein with Adenosine Triphosphatase
Activity from Cilia. Science 149, 424-426.
Gross, S.P., Tuma, M.C., Deacon, S.W., Serpinskaya, A.S., Reilein, A.R., and Gelfand, V.I.
(2002). Interactions and regulation of molecular motors in Xenopus melanophores. J Cell
Biol 156, 855-865.
Hafezparast, M., Klocke, R., Ruhrberg, C., Marquardt, A., Ahmad-Annuar, A., Bowen, S.,
Lalli, G., Witherden, A.S., Hummerich, H., Nicholson, S., et al. (2003). Mutations in
dynein link motor neuron degeneration to defects in retrograde transport. Science 300,
808-812.
Harada, A., Takei, Y., Kanai, Y., Tanaka, Y., Nonaka, S., and Hirokawa, N. (1998). Golgi
vesiculation and lysosome dispersion in cells lacking cytoplasmic dynein. J Cell Biol 141,
51-59.
Harms, M.B., Ori-McKenney, K.M., Scoto, M., Tuck, E.P., Bell, S., Ma, D., Masi, S., Allred,
P., Al-Lozi, M., Reilly, M.M., et al. (2012). Mutations in the tail domain of DYNC1H1
cause dominant spinal muscular atrophy. Neurology 78, 1714-1720.

152
Harrell, J.M., Murphy, P.J., Morishima, Y., Chen, H., Mansfield, J.F., Galigniana, M.D., and
Pratt, W.B. (2004). Evidence for glucocorticoid receptor transport on microtubules by
dynein. J Biol Chem 279, 54647-54654.
He, Y., Francis, F., Myers, K.A., Yu, W., Black, M.M., and Baas, P.W. (2005). Role of
cytoplasmic dynein in the axonal transport of microtubules and neurofilaments. J Cell
Biol 168, 697-703.
Heerssen, H.M., Pazyra, M.F., and Segal, R.A. (2004). Dynein motors transport activated
Trks to promote survival of target-dependent neurons. Nat Neurosci 7, 596-604.
Hirokawa, N., Sato-Yoshitake, R., Yoshida, T., and Kawashima, T. (1990). Brain dynein
(MAP1C) localizes on both anterogradely and retrogradely transported membranous
organelles in vivo. J Cell Biol 111, 1027-1037.
Holleran, E.A., Tokito, M.K., Karki, S., and Holzbaur, E.L. (1996). Centractin (ARP1)
associates with spectrin revealing a potential mechanism to link dynactin to intracellular
organelles. J Cell Biol 135, 1815-1829.
Hoogenraad, C.C., Akhmanova, A., Howell, S.A., Dortland, B.R., De Zeeuw, C.I.,
Willemsen, R., Visser, P., Grosveld, F., and Galjart, N. (2001). Mammalian Golgi-
associated Bicaudal-D2 functions in the dynein-dynactin pathway by interacting with
these complexes. EMBO J 20, 4041-4054.
Hrabe de Angelis, M.H., Flaswinkel, H., Fuchs, H., Rathkolb, B., Soewarto, D., Marschall,
S., Heffner, S., Pargent, W., Wuensch, K., Jung, M., et al. (2000). Genome-wide, large-
scale production of mutant mice by ENU mutagenesis. Nature genetics 25, 444-447.
Huang, J., Roberts, A.J., Leschziner, A.E., and Reck-Peterson, S.L. (2012). Lis1 acts as a
"clutch" between the ATPase and microtubule-binding domains of the dynein motor.
Cell 150, 975-986.
Imamula, K., Kon, T., Ohkura, R., and Sutoh, K. (2007). The coordination of cyclic
microtubule association/dissociation and tail swing of cytoplasmic dynein. Proceedings
of the National Academy of Sciences of the United States of America 104, 16134-16139.

153
Ito, K., and Enomoto, H. (2016). Retrograde transport of neurotrophic factor signaling:
implications in neuronal development and pathogenesis. J Biochem 160, 77-85.
Johansson, M., Rocha, N., Zwart, W., Jordens, I., Janssen, L., Kuijl, C., Olkkonen, V.M.,
and Neefjes, J. (2007). Activation of endosomal dynein motors by stepwise assembly of
Rab7-RILP-p150Glued, ORP1L, and the receptor betalll spectrin. J Cell Biol 176, 459-471.
Johnston, J.A., Illing, M.E., and Kopito, R.R. (2002). Cytoplasmic dynein/dynactin
mediates the assembly of aggresomes. Cell Motil Cytoskeleton 53, 26-38.
Jordens, I., Fernandez-Borja, M., Marsman, M., Dusseljee, S., Janssen, L., Calafat, J.,
Janssen, H., Wubbolts, R., and Neefjes, J. (2001). The Rab7 effector protein RILP controls
lysosomal transport by inducing the recruitment of dynein-dynactin motors. Curr Biol
11, 1680-1685.
Kieran, D., Hafezparast, M., Bohnert, S., Dick, J.R., Martin, J., Schiavo, G., Fisher, E.M.,
and Greensmith, L. (2005). A mutation in dynein rescues axonal transport defects and
extends the life span of ALS mice. J Cell Biol 169, 561-567.
Kimpinski, K., Campenot, R.B., and Mearow, K. (1997). Effects of the neurotrophins
nerve growth factor, neurotrophin-3, and brain-derived neurotrophic factor (BDNF) on
neurite growth from adult sensory neurons in compartmented cultures. J Neurobiol 33,
395-410.
Klusch, A., Gorzelanny, C., Reeh, P.W., Schmelz, M., Petersen, M., and Sauer, S.K. (2018).
Local NGF and GDNF levels modulate morphology and function of porcine DRG neurites,
In Vitro. PLoS One 13, e0203215.
Kon, T., Mogami, T., Ohkura, R., Nishiura, M., and Sutoh, K. (2005). ATP hydrolysis cycle-
dependent tail motions in cytoplasmic dynein. Nat Struct Mol Biol 12, 513-519.
Kon, T., Oyama, T., Shimo-Kon, R., Imamula, K., Shima, T., Sutoh, K., and Kurisu, G.
(2012). The 2.8 A crystal structure of the dynein motor domain. Nature 484, 345-350.
Koonce, M.P. (1997). Identification of a microtubule-binding domain in a cytoplasmic
dynein heavy chain. J Biol Chem 272, 19714-19718.

154
Koonce, M.P. (2000). Dictyostelium, a model organism for microtubule-based transport.
Protist 151, 17-25.
Kural, C., Kim, H., Syed, S., Goshima, G., Gelfand, V.I., and Selvin, P.R. (2005). Kinesin and
dynein move a peroxisome in vivo: a tug-of-war or coordinated movement? Science
308, 1469-1472.
LaMonte, B.H., Wallace, K.E., Holloway, B.A., Shelly, S.S., Ascano, J., Tokito, M., Van
Winkle, T., Howland, D.S., and Holzbaur, E.L. (2002). Disruption of dynein/dynactin
inhibits axonal transport in motor neurons causing late-onset progressive degeneration.
Neuron 34, 715-727.
Lawson, S.N., and Biscoe, T.J. (1979). Development of mouse dorsal root ganglia: an
autoradiographic and quantitative study. J Neurocytol 8, 265-274.
Levy, J.R., and Holzbaur, E.L. (2006). Cytoplasmic dynein/dynactin function and
dysfunction in motor neurons. Int J Dev Neurosci 24, 103-111.
Levy, J.R., and Holzbaur, E.L. (2008). Dynein drives nuclear rotation during forward
progression of motile fibroblasts. J Cell Sci 121, 3187-3195.
Li, J.Y., Pfister, K.K., Brady, S.T., and Dahlstrom, A. (2000). Cytoplasmic dynein
conversion at a crush injury in rat peripheral axons. J Neurosci Res 61, 151-161.
Ligon, L.A., LaMonte, B.H., Wallace, K.E., Weber, N., Kalb, R.G., and Holzbaur, E.L. (2005).
Mutant superoxide dismutase disrupts cytoplasmic dynein in motor neurons.
Neuroreport 16, 533-536.
Liu, Y., and Ma, Q. (2011). Generation of somatic sensory neuron diversity and
implications on sensory coding. Current opinion in neurobiology 21, 52-60.
Liu, Y., Salter, H.K., Holding, A.N., Johnson, C.M., Stephens, E., Lukavsky, P.J., Walshaw,
J., and Bullock, S.L. (2013). Bicaudal-D uses a parallel, homodimeric coiled coil with
heterotypic registry to coordinate recruitment of cargos to dynein. Genes Dev 27, 1233-
1246.

155
Lorenzo, D.N., Badea, A., Davis, J., Hostettler, J., He, J., Zhong, G., Zhuang, X., and
Bennett, V. (2014). A PIK3C3-ankyrin-B-dynactin pathway promotes axonal growth and
multiorganelle transport. J Cell Biol 207, 735-752.
Maday, S., Wallace, K.E., and Holzbaur, E.L. (2012). Autophagosomes initiate distally and
mature during transport toward the cell soma in primary neurons. J Cell Biol 196, 407-
417.
Malin, S.A., Davis, B.M., and Molliver, D.C. (2007). Production of dissociated sensory
neuron cultures and considerations for their use in studying neuronal function and
plasticity. Nat Protoc 2, 152-160.
Markus, A., Patel, T.D., and Snider, W.D. (2002). Neurotrophic factors and axonal
growth. Current opinion in neurobiology 12, 523-531.
Matanis, T., Akhmanova, A., Wulf, P., Del Nery, E., Weide, T., Stepanova, T., Galjart, N.,
Grosveld, F., Goud, B., De Zeeuw, C.I., et al. (2002). Bicaudal-D regulates COPI-
independent Golgi-ER transport by recruiting the dynein-dynactin motor complex. Nat
Cell Biol 4, 986-992.
McKenney, R.J., Weil, S.J., Scherer, J., and Vallee, R.B. (2011). Mutually exclusive
cytoplasmic dynein regulation by NudE-Lis1 and dynactin. J Biol Chem 286, 39615-
39622.
McNally, F.J. (2013). Mechanisms of spindle positioning. J Cell Biol 200, 131-140.
Mitchell, D.J., Blasier, K.R., Jeffery, E.D., Ross, M.W., Pullikuth, A.K., Suo, D., Park, J.,
Smiley, W.R., Lo, K.W., Shabanowitz, J., et al. (2012). Trk activation of the ERK1/2 kinase
pathway stimulates intermediate chain phosphorylation and recruits cytoplasmic dynein
to signaling endosomes for retrograde axonal transport. The Journal of neuroscience :
the official journal of the Society for Neuroscience 32, 15495-15510.
Mocz, G., Farias, J., and Gibbons, I.R. (1991). Proteolytic analysis of domain structure in
the beta heavy chain of dynein from sea urchin sperm flagella. Biochemistry 30, 7225-
7231.

156
Moore, J.K., Stuchell-Brereton, M.D., and Cooper, J.A. (2009). Function of dynein in
budding yeast: mitotic spindle positioning in a polarized cell. Cell Motil Cytoskeleton 66,
546-555.
Neuwald, A.F., Aravind, L., Spouge, J.L., and Koonin, E.V. (1999). AAA+: A class of
chaperone-like ATPases associated with the assembly, operation, and disassembly of
protein complexes. Genome Res 9, 27-43.
Ori-McKenney, K.M., and Vallee, R.B. (2011). Neuronal migration defects in the Loa
dynein mutant mouse. Neural Dev 6, 26.
Ori-McKenney, K.M., Xu, J., Gross, S.P., and Vallee, R.B. (2010). A cytoplasmic dynein tail
mutation impairs motor processivity. Nat Cell Biol 12, 1228-1234.
Paschal, B.M., and Vallee, R.B. (1987). Retrograde Transport by the Microtubule-
Associated Protein Map-1c. Nature 330, 181-183.
Patel, P.I., Roa, B.B., Welcher, A.A., Schoener-Scott, R., Trask, B.J., Pentao, L., Snipes,
G.J., Garcia, C.A., Francke, U., Shooter, E.M., et al. (1992). The gene for the peripheral
myelin protein PMP-22 is a candidate for Charcot-Marie-Tooth disease type 1A. Nature
genetics 1, 159-165.
Patel, T.D., Jackman, A., Rice, F.L., Kucera, J., and Snider, W.D. (2000). Development of
sensory neurons in the absence of NGF/TrkA signaling in vivo. Neuron 25, 345-357.
Pazour, G.J., Dickert, B.L., and Witman, G.B. (1999). The DHC1b (DHC2) isoform of
cytoplasmic dynein is required for flagellar assembly. J Cell Biol 144, 473-481.
Perlson, E., Jeong, G.B., Ross, J.L., Dixit, R., Wallace, K.E., Kalb, R.G., and Holzbaur, E.L.
(2009). A switch in retrograde signaling from survival to stress in rapid-onset
neurodegeneration. The Journal of neuroscience : the official journal of the Society for
Neuroscience 29, 9903-9917.
Perlson, E., Maday, S., Fu, M.M., Moughamian, A.J., and Holzbaur, E.L. (2010).
Retrograde axonal transport: pathways to cell death? Trends Neurosci 33, 335-344.

157
Pfister, K.K. (2015). Distinct functional roles of cytoplasmic dynein defined by the
intermediate chain isoforms. Exp Cell Res 334, 54-60.
Pilling, A.D., Horiuchi, D., Lively, C.M., and Saxton, W.M. (2006). Kinesin-1 and Dynein
are the primary motors for fast transport of mitochondria in Drosophila motor axons.
Mol Biol Cell 17, 2057-2068.
Poirier, K., Lebrun, N., Broix, L., Tian, G., Saillour, Y., Boscheron, C., Parrini, E., Valence,
S., Pierre, B.S., Oger, M., et al. (2013). Mutations in TUBG1, DYNC1H1, KIF5C and KIF2A
cause malformations of cortical development and microcephaly. Nature genetics 45,
639-647.
Porter, M.E., Bower, R., Knott, J.A., Byrd, P., and Dentler, W. (1999). Cytoplasmic dynein
heavy chain 1b is required for flagellar assembly in Chlamydomonas. Mol Biol Cell 10,
693-712.
Presley, J.F., Cole, N.B., Schroer, T.A., Hirschberg, K., Zaal, K.J., and Lippincott-Schwartz,
J. (1997). ER-to-Golgi transport visualized in living cells. Nature 389, 81-85.
Raaijmakers, J.A., van Heesbeen, R.G., Meaders, J.L., Geers, E.F., Fernandez-Garcia, B.,
Medema, R.H., and Tanenbaum, M.E. (2012). Nuclear envelope-associated dynein drives
prophase centrosome separation and enables Eg5-independent bipolar spindle
formation. EMBO J 31, 4179-4190.
Rigaud, M., Gemes, G., Barabas, M.E., Chernoff, D.I., Abram, S.E., Stucky, C.L., and
Hogan, Q.H. (2008). Species and strain differences in rodent sciatic nerve anatomy:
implications for studies of neuropathic pain. Pain 136, 188-201.
Rishal, I., Kam, N., Perry, R.B., Shinder, V., Fisher, E.M., Schiavo, G., and Fainzilber, M.
(2012). A motor-driven mechanism for cell-length sensing. Cell Rep 1, 608-616.
Roberts, A.J., Malkova, B., Walker, M.L., Sakakibara, H., Numata, N., Kon, T., Ohkura, R.,
Edwards, T.A., Knight, P.J., Sutoh, K., et al. (2012). ATP-driven remodeling of the linker
domain in the dynein motor. Structure 20, 1670-1680.

158
Rogers, D.C., Peters, J., Martin, J.E., Ball, S., Nicholson, S.J., Witherden, A.S., Hafezparast,
M., Latcham, J., Robinson, T.L., Quilter, C.A., et al. (2001). SHIRPA, a protocol for
behavioral assessment: validation for longitudinal study of neurological dysfunction in
mice. Neuroscience letters 306, 89-92.
Sabblah, T.T., Nandini, S., Ledray, A.P., Pasos, J., Calderon, J.L.C., Love, R., King, L.E., and
King, S.J. (2018). A novel mouse model carrying a human cytoplasmic dynein mutation
shows motor behavior deficits consistent with Charcot-Marie-Tooth type 2O disease.
Scientific Reports 8.
Sakato, M., and King, S.M. (2004). Design and regulation of the AAA+ microtubule motor
dynein. J Struct Biol 146, 58-71.
Sasaki, S., Shionoya, A., Ishida, M., Gambello, M.J., Yingling, J., Wynshaw-Boris, A., and
Hirotsune, S. (2000). A LIS1/NUDEL/cytoplasmic dynein heavy chain complex in the
developing and adult nervous system. Neuron 28, 681-696.
Saxena, S., Bucci, C., Weis, J., and Kruttgen, A. (2005). The small GTPase Rab7 controls
the endosomal trafficking and neuritogenic signaling of the nerve growth factor
receptor TrkA. The Journal of neuroscience : the official journal of the Society for
Neuroscience 25, 10930-10940.
Schnapp, B.J., and Reese, T.S. (1989). Dynein is the motor for retrograde axonal
transport of organelles. Proceedings of the National Academy of Sciences of the United
States of America 86, 1548-1552.
Schroeder, C.M., Ostrem, J.M., Hertz, N.T., and Vale, R.D. (2014). A Ras-like domain in
the light intermediate chain bridges the dynein motor to a cargo-binding region. Elife 3,
e03351.
Schroer, T.A. (2004). Dynactin. Annu Rev Cell Dev Biol 20, 759-779.
Schwarz, T.L. (2013). Mitochondrial trafficking in neurons. Cold Spring Harb Perspect
Biol 5.

159
Scoto, M., Rossor, A.M., Harms, M.B., Cirak, S., Calissano, M., Robb, S., Manzur, A.Y.,
Martinez Arroyo, A., Rodriguez Sanz, A., Mansour, S., et al. (2015). Novel mutations
expand the clinical spectrum of DYNC1H1-associated spinal muscular atrophy.
Neurology 84, 668-679.
Shah, J.V., Flanagan, L.A., Janmey, P.A., and Leterrier, J.F. (2000). Bidirectional
translocation of neurofilaments along microtubules mediated in part by
dynein/dynactin. Mol Biol Cell 11, 3495-3508.
Sivagurunathan, S., Schnittker, R.R., Razafsky, D.S., Nandini, S., Plamann, M.D., and King,
S.J. (2012). Analyses of dynein heavy chain mutations reveal complex interactions
between dynein motor domains and cellular dynein functions. Genetics 191, 1157-1179.
Sleigh, J.N., Dawes, J.M., West, S.J., Wei, N., Spaulding, E.L., Gomez-Martin, A., Zhang,
Q., Burgess, R.W., Cader, M.Z., Talbot, K., et al. (2017). Trk receptor signaling and
sensory neuron fate are perturbed in human neuropathy caused by Gars mutations.
Proc Natl Acad Sci U S A 114, E3324-E3333.
Smeyne, R.J., Klein, R., Schnapp, A., Long, L.K., Bryant, S., Lewin, A., Lira, S.A., and
Barbacid, M. (1994). Severe sensory and sympathetic neuropathies in mice carrying a
disrupted Trk/NGF receptor gene. Nature 368, 246.
Steffen, W., Karki, S., Vaughan, K.T., Vallee, R.B., Holzbaur, E.L., Weiss, D.G., and
Kuznetsov, S.A. (1997). The involvement of the intermediate chain of cytoplasmic dynein
in binding the motor complex to membranous organelles of Xenopus oocytes. Mol Biol
Cell 8, 2077-2088.
Suzuki, H., Aoyama, Y., Senzaki, K., Vincler, M., Wittenauer, S., Yoshikawa, M., Ozaki, S.,
Oppenheim, R.W., and Shiga, T. (2010). Characterization of sensory neurons in the
dorsal root ganglia of Bax-deficient mice. Brain research 1362, 23-31.
Tanaka, T., Serneo, F.F., Higgins, C., Gambello, M.J., Wynshaw-Boris, A., and Gleeson,
J.G. (2004). Lis1 and doublecortin function with dynein to mediate coupling of the
nucleus to the centrosome in neuronal migration. J Cell Biol 165, 709-721.

160
Teuchert, M., Fischer, D., Schwalenstoecker, B., Habisch, H.J., Bockers, T.M., and
Ludolph, A.C. (2006). A dynein mutation attenuates motor neuron degeneration in
SOD1(G93A) mice. Experimental neurology 198, 271-274.
Tsurusaki, Y., Saitoh, S., Tomizawa, K., Sudo, A., Asahina, N., Shiraishi, H., Ito, J., Tanaka,
H., Doi, H., Saitsu, H., et al. (2012). A DYNC1H1 mutation causes a dominant spinal
muscular atrophy with lower extremity predominance. Neurogenetics 13, 327-332.
Tynan, S.H., Purohit, A., Doxsey, S.J., and Vallee, R.B. (2000). Light intermediate chain 1
defines a functional subfraction of cytoplasmic dynein which binds to pericentrin. J Biol
Chem 275, 32763-32768.
Vale, R.D. (2003). The molecular motor toolbox for intracellular transport. Cell 112, 467-
480.
Valentine, J.S., Doucette, P.A., and Zittin Potter, S. (2005). Copper-zinc superoxide
dismutase and amyotrophic lateral sclerosis. Annu Rev Biochem 74, 563-593.
Vallee, R.B., McKenney, R.J., and Ori-McKenney, K.M. (2012). Multiple modes of
cytoplasmic dynein regulation. Nat Cell Biol 14, 224-230.
van Spronsen, M., Mikhaylova, M., Lipka, J., Schlager, M.A., van den Heuvel, D.J.,
Kuijpers, M., Wulf, P.S., Keijzer, N., Demmers, J., Kapitein, L.C., et al. (2013).
TRAK/Milton motor-adaptor proteins steer mitochondrial trafficking to axons and
dendrites. Neuron 77, 485-502.
Varma, D., Monzo, P., Stehman, S.A., and Vallee, R.B. (2008). Direct role of dynein motor
in stable kinetochore-microtubule attachment, orientation, and alignment. J Cell Biol
182, 1045-1054.
Wang, T., Martin, S., Nguyen, T.H., Harper, C.B., Gormal, R.S., Martinez-Marmol, R.,
Karunanithi, S., Coulson, E.J., Glass, N.R., Cooper-White, J.J., et al. (2016). Flux of
signalling endosomes undergoing axonal retrograde transport is encoded by presynaptic
activity and TrkB. Nature communications 7, 12976.

161
Wang, X., Ratnam, J., Zou, B., England, P.M., and Basbaum, A.I. (2009). TrkB signaling is
required for both the induction and maintenance of tissue and nerve injury-induced
persistent pain. The Journal of neuroscience : the official journal of the Society for
Neuroscience 29, 5508-5515.
Waterman-Storer, C.M., Karki, S.B., Kuznetsov, S.A., Tabb, J.S., Weiss, D.G., Langford,
G.M., and Holzbaur, E.L. (1997). The interaction between cytoplasmic dynein and
dynactin is required for fast axonal transport. Proceedings of the National Academy of
Sciences of the United States of America 94, 12180-12185.
Weedon, M.N., Hastings, R., Caswell, R., Xie, W., Paszkiewicz, K., Antoniadi, T., Williams,
M., King, C., Greenhalgh, L., Newbury-Ecob, R., et al. (2011). Exome sequencing
identifies a DYNC1H1 mutation in a large pedigree with dominant axonal Charcot-Marie-
Tooth disease. Am J Hum Genet 89, 308-312.
Wickstead, B., and Gull, K. (2007). Dyneins across eukaryotes: a comparative genomic
analysis. Traffic 8, 1708-1721.
Wilkie, G.S., and Davis, I. (2001). Drosophila wingless and pair-rule transcripts localize
apically by dynein-mediated transport of RNA particles. Cell 105, 209-219.
Willemsen, M.H., Vissers, L.E., Willemsen, M.A., van Bon, B.W., Kroes, T., de Ligt, J., de
Vries, B.B., Schoots, J., Lugtenberg, D., Hamel, B.C., et al. (2012). Mutations in DYNC1H1
cause severe intellectual disability with neuronal migration defects. J Med Genet 49,
179-183.
Yagi, T. (2009). Bioinformatic approaches to dynein heavy chain classification. Methods
Cell Biol 92, 1-9.
Yajima, Y., Narita, M., Narita, M., Matsumoto, N., and Suzuki, T. (2002). Involvement of a
spinal brain-derived neurotrophic factor/full-length TrkB pathway in the development of
nerve injury-induced thermal hyperalgesia in mice. Brain research 958, 338-346.
Yano, H., Lee, F.S., Kong, H., Chuang, J., Arevalo, J., Perez, P., Sung, C., and Chao, M.V.
(2001). Association of Trk neurotrophin receptors with components of the cytoplasmic

162
dynein motor. The Journal of neuroscience : the official journal of the Society for
Neuroscience 21, RC125.
Young, A., Dictenberg, J.B., Purohit, A., Tuft, R., and Doxsey, S.J. (2000). Cytoplasmic
dynein-mediated assembly of pericentrin and gamma tubulin onto centrosomes. Mol
Biol Cell 11, 2047-2056.
Zhao, J., Wang, Y., Xu, H., Fu, Y., Qian, T., Bo, D., Lu, Y.X., Xiong, Y., Wan, J., Zhang, X., et
al. (2016). Dync1h1 Mutation Causes Proprioceptive Sensory Neuron Loss and Impaired
Retrograde Axonal Transport of Dorsal Root Ganglion Neurons. CNS Neurosci Ther 22,
593-601.










![Identification and Characterization of Pleural Neurons ......dulin, sensory neuron, motor neuron, inhibition, neural cir- cuit, Aplysia] The sensory and motor neurons that mediate](https://img.pdfslide.us/doc/110x75/5fc497a9642d1777a877bb71/identification-and-characterization-of-pleural-neurons-dulin-sensory-neuron.jpg)