Embed Size (px)
Citation preview

MOSS LANDING MARINE LABORATORIES
VERTICAL TRANSPORT AND EXCHANGE OF MATERIALS IN THE UPPER WATERS OF THE OCEANS (VERTEX):
INTRODUCTION TO THE PROGRAM, HYDROGRAPHIC CONDITIONS AND MAJOR COMPONENT FLUXES DURING VERTEX I
Moss Landing Marine Laboratories Technical Publication 83-2
by
John H. MartinI George A. Knauer l
William W. Broenkow l
Kenneth W. Bruland 2
David M. Kar1 3 Lawrence F. Smal1 4
r'la ry W. Sil ver2
Marcia M. Gowing2
December 1983 Moss Landing, CA 95039
IMoss Landing Marine Laboratories, Moss Landing, CA 95039 2University of California, Santa Cruz, CA 95064
3University of Hawaii, Honolulu, HI 96822 40regon State University, Corvallis, OR 97331

VERTICAL TRANSPORT AND EXCHANGE OF ~iATERIALS IN THE UPPER WATERS OF THE OCEANS (VERTEX):
INTRODUCTION TO THE PROGRAM, HYDROGRAPHIC CONDITIONS AND MAJOR COMPONENT FLUXES DURING VERTEX I
by
John H. MartinI George A. Knauer l
William W. Broenkow l Kenneth W. Bruland 2
Dav id t·t Kar1 3 Lawrence F. Smal1 4
Mary W. Silver2 Marcia M. Gowing 2
IMoss Landing Marine Laboratories, Moss Landing, CA 95039 2University of California, Santa Cruz, CA 95064
3University of Hawaii, Honolulu, HI 96822 40regon State University, Corvallis, OR 97331

ABSTRACT
This paper introduces VERTEX, a multi-disciplinary research program
dealing with various aspects of particle transport in the upper,
high-energy layers (0-2000 m) of the ocean. Background information is
presented on hydrography, biological composition of trapped
particulates, and major component fluxes observed on a cruise off
central California (VERTEX I). Organic C fluxes measured with two trap
systems are compared with several other estimates taken from the
literature. The intent of this overview paper is to provide a common
setting in an economical manner, and avoid undue repetition of
background and ancillary information in subsequent publications.

INTRODUCTION
The mechanisms and rates of biogeochemical cycling of elements and
organic compounds in the sea and the incorporation and flux of energy
through biological compartments have interested oceanographers for
years. Indeed, cycling and flux studies of one sort or another have
involved almost every discipline of oceanography. However, the factors
responsible for controlling the distributions of elements and compounds
in oceanic water columns are poorly understood. This is especially true
when considering the role of organisms and their remains in these
cycling processes, in spite of the fact that everyone recognizes their
importance.
Within the past few years, the development of particle traps has
enabled the measurement of fluxes of materials, not only to the sea
floor, but also within various portions of the water column. Recent
advances in methodology and instrumentation have also made possible "the
accurate determination of a reasonably large suite of elements and
compounds at the very low levels at which they exist in sea water. This
combination of developments now enables oceanographers to measure rates
of change and residence times in the water column by comparing fluxes at
various depth intervals.
A group sharing mutual interests in studying these processes in the
upper "high-energy" layers (0-2000 m) of the ocean was formed in 1979.
The participants named the resea-rch program VERTEX, an acronym which
stands for vertical !ransport and exchange of materials in the upper
ocean. The overall objectives of the program are to: (1) accurately

2
determine the amounts of elements and/or compounds in the water column
in both particulate and dissolved form; (2) measure the fluxes of these
elements and compounds at selected depths using particle traps; (3)
using the information gained in objectives land 2, estimate residence
times and rates of change; and (4)' understand how the transport and
exchange system works in general and in specific ocean regions.
The purpose of this paper is to introduce the VERTEX program and to
provide background information on the first cruise (VERTEX I) which took
place off central California in August-September 1980. This basic
information will provide a common setting in an economical manner for
future publications by the participants that will deal with various
aspects of the program, such as primary production and C, H, N fluxes
(Knauer and Martin, Moss Landing Marine Laboratories [MLML]),
zooplankton (Small, Oregon State University), micro-organisms (Karl,
University of Hawaii), marine snow (Silver and Gowing, University of
California at Santa Cruz [UCSCJ), hydrography and currents (Broenkow,
MLML), trace elements (Bruland, UCSC: Martin and Knauer, MLML) , natural
series radionuclides (Bruland, UCSC); transuranics (Fowler,
International Laboratory of Marine Radioactivity, Monaco), 1ipids
(Wakeham and Farrington, Woods Hole Oceanographic Institution [WHOI]),
organo-nitrogen compounds (Lee, WHOI) and higher molecular weight
hydrocarbons (Risebrough, Bodega Marine Laboratory, University of
California at Berkeley).

3
METHODS
Particle Traps: Two types of particle traps are used in the VERTEX
program: large (0.25 m2 ) cone-shaped traps designed by Andrew Soutar of
Scripps Institution of Oceanography (Fig. 1), and small (0.0039 m2 )
cylindrical traps designed by George Knauer of MLML (Fig. 2). The
Soutar design, funnel-shaped particle interceptor traps (PITs), consist
of a pair of teflon-coated fiberglass funnels with acrylic cod ends.
The PIT collecting surface is a cellular grid array (square openings 1
cm on a side) where particle trap interaction (rejection vs entrapment)
is believed to occur. PITs for organic material collections are of the
same design, except that the cones are teflon-coated stainless steel and
the cod ends are electro-polished stainless steel. Preservation in the
trace element/radionuclide PITs is accomplished using buffered (pH = 8)
formalin. The formalin-sodium borate solution in a 60-ml polyethylene
bottle weighted with a teflon-coated magnet is placed in the PIT cod
ends, where it slowly diffuses out through small holes in the bottle
cap. Mercuric chloride is used for preservation in the stainless steel
PITs. All interior PIT surfaces are thoroughly cleaned with either acid
or organic solvents prior to launch.
The MLML traps (Fig. 2) consist of a frame constructed of
high-impact PVC, which is fitted with 12 identical cylinders (10 = 7.0
cm; 1ength = 60 cm). The mouth of each cyl i nder has a baffl e system
consisting of 16 smaller tubes (10 = 1.3 cm; length = 5 cm). The tops
of the tubes are milled to a wall thickness of 0.06 mm to maximize open
surface area. The cylinders are made of lucite with polyethylene cod

,j'
Soutar Trap centra I post
I em gridI I0.6 m flow control
1111111111111111111111111""" II /I, /I 11111111111" 1111111111" 111111 11111111111111111111/111111111111111111.11111111111111111111 lip
~rudder
~fiberglass
_____I support member
1.3 m funnel wall
~ Delrin swivel -, on clevice post
T sample collector
0.2m ~
FIG. 1. Schematic drawing of the Soutar design particle interceptor traps. Traps were deployed byBruland of UCSC and are referred to as UCSC PITs throughout the text.
1

POLYPROPYLENE REMOVABLE LINE BAFFLE SYSTEM
7.0 em. TO
E u to r<>
TOPVIEW BAFFLE RETAINING
GRID SYSTEM / COLLER
E o
STABI LIZI NG RETAINING lO C\J
LANYARD CORD COLLECTION
CUP
U1o. M LML Frame b.MLML Trap Cylinder FIG. 2. Schematic drawing of the MLML traps used during VERTEX I.

6
ends, except for those designed to collect materials for organa-nitrogen
compounds, which consist of glass cylinders lined with teflon bags.
The ~1LML trap cylinders are filled vJith either a NaCl solution
(Knauer, Martin) or a sucrose solution (Silver, Gowing) having a density
-3)( p = 1.05 g cm greater than that of sea water. The solution
maintains its integrity for periods of weeks, preventing significant
exchange. The density solution also retains added preservatives and
substances that dissolve from the trapped particles (e.g., Cd and P04;
Knauer and Martin, 1981). Various preservatives or tracers are added to
the gradient solutions, depending on the intended analyses or
experiments (see Table 1).
A similar flotation system is used for the two trap systems (Fig.
3). The major portion of the array·s ~Jeight is carried by submerged
glass, hard-hat floats set at depth below turbulent wave action. Both
arrays are equipped with spar buoys fitted with strobe lights and OAR
rad; 0 t ransmi tters. The ~lL~1L buoys are a1,so equ i pped v.Ji th an Argos
satellite transponder.
Both the Soutar and MLML type traps were tested during the Sediment
Trap 1ntercomparison Experinlent (ST1E; Spencer, et al., 1981), which
took place in the Panama Basin from 28 July to 1 December 1979.
Generally good agreement was obtained among the VERTEX traps and two
other trap systems, those of Honjo (Woods Hole Oceanographic
Institution) and Gardner (Lamont-Doherty Geological Observatory) (Fig.
4). This agreement is remarkable in view of the fact that the traps
were of different design and size. For example, Honjo·s traps have a
1.5 m2 collecting surface in comparison to the 0.0039 m2 for the MLML
traps. The obvious loss of CaC03 in the MLML traps resulted from the

7
TABLE 1. Intended analyses, preservatives used, and investigators for
the 12 MLML trap cylinders during VERTEX I.
NUTnbe r 0 f Analysis Samples Pre serva ti ve Investigator
Particle
Identification 1 Glutaraldehyde Silver
C, N, H 2 Forma1in Knauer-Martin
Trace metals 3 Formalin Knauer-Martin
ATP 2 Phosphoric acid Karl
Uptake kinetics 1 H3 adenine Karl (no preservative)
Organic nitrogen 2 Chloroform Lee
Radionuclides 1 Formalin Bruland

8
spar bUoy w/strobe radio transmitter and surface floats
wave damper
su b- surface floats
PIT - 100m
PIT- 250m
PIT - 750m
PIT -1500 m
weight FIG. 3. Flotation system used for both UCSC and MLML traps during VERTEX I.

Org Matter Si Iicate Carbonate
a 15 30 0 50 100 0 25 500' iii iii • • • iii iii
o e e- 0 •i 1000, •• •
~ 0 +0 0
I~
E r • 1'. t 0
1- 2000 0.. Ir • A + •OA toAI •W 0
o
3000 l 0 i· MLML 0 I 0 o WHOI e LOGO ~ SIO o lot 0
I , • ,4 000' , , , , • I , I I , •
FIG. 4. Major component fluxes (mg m- 2 day-I) measured during Sediment Trap Intercomparison \.0
Experiment by Knauer and Martin (MLML), Honjo (WHOI), Gardner (LOGO), and Soutar (SID). Figure drawn from data in Spencer, et al., 1981.

10
dissolution of CaC03 in the NaCl trap solution. CaC03 is now added to
selected MLML trap cylinders to prevent this problem.
Hydrographic methods are presented in Broenkow and Greene (1981).
Nutrients were analyzed immediately after collection using an
autoanalyzer via the methods of Atlas, Gordon, Hager and Park (1971).
Biological composition methods are described in Silver and Alldredge
(1981 ) .

11
RESULTS AND DISCUSSION
Hydrographic Data
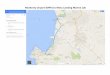
The study area was surveyed prior to deployment of the traps (17 to
21 August 1980) using a 20-station CTO grid centered on the intended
trap launch site. Near-surface temperature and chlorophyll fluorescence
were measured via continuous surface profiles among the CTO stations.
The survey data showed a pattern typical of the late upwelling season
(Lynn 1967) with cool « 13°C) temperatures and relatively high (> 20
mg m-2) chlorophyll levels in the nearshore waters (Fig. 5). The local
center of strong upwelling is near Point Sur, and tongues of
nutrient-rich, cool, high-salinity waters have been often observed
penetrating into Monterey Bay from the south (Broenkow and Smethie,
1978 ) . The surface temperature distribution was consistent with
geostrophic flow at the surface (Fig. 6). The pre-deployment survey
showed two eddies: cyclonic flow was centered at 37°N, l23°30'W,
somewhat west of a surface temperature minimum; and an anticyclonic eddy
was centered at 35°40 ' N, 124°W in an area of warmer surface waters.
Flow between these two features was strong and onshore. To avoid the
possibility of shoreward drift, the traps were launched south of the
survey area rather than in the center of the shoreward flow, as
originally planned.
Vertical temperature, salinity and dissolved oxygen distributions
near the traps (Fig. 7) are representative of the California Current
along the central California coast. Mixed-layer depths vary seasonally
in the study area, from near zero during periods of intense upwelling

12
•
Q)
-0 :J 37°
-+-'. -+-' 0
--.J
-C -+-'
L
0 z
36°
FIG.
.= .:~:.: · : : .: :.: · :.: N l '-r·a •••~ ••• - •• -. • MI ~
San ·e: e_e: . 0 , 0 20 30 40 50
Francisco -;~. -.. : I I '. I 'I 1'1 I' I I' .. - 0 30 60 90
Kilometers
Cruz\ \ ...•. +._0 .- +--
_.~
0: 14 15 ~
.tt. ·15
.. .. ..
o •••• .~....... - _. . ....0.·., .. . · . . .: ..- . -... .. .. .. . . .~..~..'{::~~:~~: .
.....:e; ·~
Morro 8ay}~• : :
West Longitude Sa. COT Stations (+) and surface temperatures (oC).
Trajectory of MLML PIT mooring shown by dashed line.

13
b .::/.: ::: ~.:.:: ::: N Mn~ :~:.. :.. 0 10 2·0 30 40 50
.· .: I I 'I I ' ii' I "I .' .... 0 30 60 90 ::.: ;.. Kilometers
..-...
Q)
-0 + Cruz37° + +\::J 4-J
5.4-J 0 . .
.--J +
...c ++4-J L <5 a . . . . .Z • + . . . .:. .. . .. .36°
++
.: :. I •• • .. ... . . .+ .: ... . .... . + .c start . . .. ..
.. ' \. : ., }., , .. ;-.:..:.:.=.., ..- .• ".,4 • . ':.:
•
West Longitude
3FIG. 5b. Surface pigments (mg m- ) from in vivo fluorescence. _
-------~~---------

14
. -....-... .=::.< e: : :: : ·:. : ::..
·-... -. 0 10 ... : I I I. -.. a
N. Mn~s 20 30 40 50 . '. I I I I" "
30 60 90
(1)
-0 :J
-+-J.-+-J o
--.J 164
-C -+-J L o
Z
174=::J,,•
Kilometers
+ + +
++
.. . ... ..:e· · _. _
: :. I -. •..... . . . .: ... . .... .. .. .. . . .~. :..':\:..:.::
• :. e: •••• ....
West Longitude
FIG. 6. Dynamic height anomalies (dyn mm) surface/I500 db, and trajectory of MLML PIT mooring.

.-------- Salinity (ppt) 15
32.5 33.0 33.5 34.0 34.5 35.0
-- Temperature (deg C) o 4 8 12 16 20 -------...-..........-----..------.......--.....-~---.--.........--......-----.O i
.....,0.' -"'::~.:......
00••...0•....•0...... --._"-•• 0· ..., ..
-It
.0 .., .... " ~. ,
...... e,.
':,400 ,"'••.., ...., '\ I
" ",, \,,,
800 "'\
I,,,, I \ ,
..t: \ ,"~ ,J
~ ,,I I
,,~ 1200 , , "'\ C
, "'\,,I\
( J, \ \,,•1600 ,
"J,, ,\
2000 ~~ ~ ~--",,----,- -.a.-~
o 100 200 300 400 500 _ Oxygen (J-Lmoles/kg)
FIG 6 7. Vertical temperature, salinity, and dissolved oxygen, Station 28; 350 37.1 I N, 1230 40.8 I W, 31 AuguSt 1980.

16
(generally in the spring and summer months) to nearly 100 m during the
winter (Eber, 1977). The 20 to 30 m mixed layer depths observed during
VERTEX I are typical of mid- to late-summer conditions. Minimum
dissolved oxygen concentrations (about 12 umo1e kg-lor 4% saturation)
were found at 750 m, and oxygen concentrations of < 10% saturation were
present between 450 and 1200 m (Fig. 7). The intrusive features shown
in the oxygen profile between 200 and 400 m are probably real (because
they were observed at several nearby stations) and result from
small-scale advective mixing processes.
Salinity distributions (Fig. 8a) show low salinity (33.2 < S <
33.7) surface waters formed in the transition zone between Pacific
Subarctic and Pacific Equatorial Waters (Sverdrup, Johnson and Fleming,
1942). An interesting feature of the salinity distribution along the
California coast is the presence of relatively high salinity "southern"
water (Wickham, 1975) which flows northward along the continental slope
in the California Countercurrent. The salinity and geostrophic flow
sections across the study area (Fig. 8) show a small subsurface salinity
maximum (S ~ 34.2) in the area of northerly flow nearshore. From spring
to fall, the California Countercurrent is generally subsurface, with its
core between 200 and 300 m (Wickham, 1975; Hickey, 1979), though
occurrences of northerly flow at the surface may be observed whenever a
favorable interplay of forces permits. McLain and Thomas (in prep.)
suggest that the surface countercurrent is caused primarily by local
onshore Ekman transport during winter, and by remote forcing due to
poleward propagation of coastal trapped waves from the tropics.

17
Distance (km)
200 150 100 50 a Or------,.--------y"----=-=---:----~..---~--
0------------------33.4 ~33.6 1 :
--_.- , ..-------=-33.8 34 0 :.. ~.:.::a . ~ ..,-r•..., ~ ::.J: : ---- -- 34.1 --.........----C .:.::=:' ·
----..... 3~:2 ~ ..'.-34.2 ~ ...500 :.- ... ...E ~
..c -+-' --------34.4---a. (1)
0 1000
34.5-
1500 ~__""--__.....Io....-__--L.-_----':"---I.-..:.._.-.----1
'b
~ 500E ~
..c -+-' a. Q) ----- -1
0 1000
1500 a.....-__-......- ......Io--__--L__----:....:.....L.... • ----I.
FIG. 8. Transect west of Point Sur, 17-19 August 1980. (a) Sa1i ni ty.(b) Geostrophie flow (em sec-I). Northerly flow is hatched.

18
The ~1LML and UCSC trap moori ngs acted as 2000 and 1500 m-long
(respectively) drift poles, and their trajectories were determined by
the water column-mean velocity. (The ratio of the submerged and exposed
surface area was 36:1 for the MLML trap line.) Both the MLML and UCSC
moorings drifted in a clockwise sense around the anticyclonic
geostrophic eddy (Figs. 6, 9). Because the MLML mooring was accurately
tracked by 70 LORAN-C fixes during the 13-day deployment, and the UCSC
array was located by only 16 such fixes, detailed current results are
available only for the MLML trap array, on which two Endeco Type 741,
neutrally-buoyant current meters were set at 150 and 1500 m. Broenkow
(1982) found good agreement between daily-mean relative currents
measured by the freely drifting meters when adjustments were made for
trap array drift and the geostrophic currents near the traps on day 8 of
the experiment. He showed that the daily mean mooring line drift rate
varied from 4.2 to 11.4 em see- l (mean 7.5 em see- l ) during the 13-day
deployment. The relative velocities as measured by the current meters
showed both tidal and inertial (20.5 hr) periodicity at 150 and 1500 m,
and relative velocities at these depths were essentially out of phase by
180°. Gardner (1980a, b) has shown that currents can affect the
performance and accuracy of certain particle interceptor traps to
accurately catch sinking particles. Relative current speed measurements
at 150 and 1500 m (PITs closest to the meters were at 100, 200 and 1400
m) showed that the instantaneous (based on a 2-minute recording
interval) maximum current speed was 21 em see-1 at 150 m and 17 em see-l r
at 1500 m. _Thirty-minute sustained maximum velocities of 18.3 and 15.2 lem see- were observed at these depths (Fig. 10). The bimodal (8 and 13

19
36°00'
LORAN
UCSC -_ ... ---_ ...... start \_1
\
Q) 35°45' . 13 ~ \
\* \J \ \:J \
\
*3,.+J \ I.-+-' t\ , --1 \
\ , 0 *\
t5 ...c
\ \ ,, .......,
L-I \11 * a \ , I
,z 35°30' , , ,
,I , '~ , ,it 7
, , ~
.....' ~
,~ ~ --. ...
N. Miles ~ ........---a 5 10 15 '9
I I II i i i i
0 10 20 30 Kilometers
35°15' 124°15' 124°00' 123
0
45' 123°30'
West Longitude FIG. 9. Observed PIT positions 26 August to 8 September 1980 .
. Dashed line is UCSC mooring; solid line is MLML mooring. Numbers show interpolated daily noon positions. Adapted from Broenkow (1982).

20
100
80 a ~ () c 60
150 m
Q.)
:J 0Q) 40 L
l.L
20
100
80 b ~ 1500 m ()
c 60 Q) :J CT Q) 40 l-
l.L
20
5 10 15 20
Mean Speed (em/sec) FIG. 10. Frequency fur 3D-min. average relative current
speeds on MLML PIT moorings, 26 August to 8 Seftember 1980. (a (b)
150 nl. 1500 m.

21
cm sec- l ) frequency distribution for the 150-m depth was caused by the
water column acceleration following day 8 of the experiment (Fig. 10).
lAt 1500 m, the modal relative speed was 5 cm sec- . Mean relative
lcurrent speeds during the experiment were 9.7 and 6.0 cm sec- . In view
of these velocities and the trap designs (MLML cylinders and UCSC cones,
"both with baffles), we believe it unlikely that horizontal water
movements were significantly affecting trap accuracy, especially in
light of the fact that they were free drifting (see Staresinic, Von
Brockel, Snodlake and Clifford, 1982).
Nutrients
Nutrient profiles typical of the California Current were observed
during VERTEX I (Fig. 11). Nitrate plus nitrite and silicate
concentrations were near zero at the surface (0. 1; 3. 5 umo1 kg-1 ,
respectively), while phosphate levels were relatively high (0.5 umol
kg- l ). Typical increases with depth were observed for these nutrients;
P0 and N03 + N02 maxima coincided with the oxygen minimum (Figs. 7,4
11), while 5i02 amounts increased continuously in the 2000-m water
column. Anmonia concentrations (data not shown) were variable (0-0.9
umole kg- l ) throughout the water column, and no definite trend with
depth was observed.
Biological Composition of Trapped Particulates
Amorphous organic detritus dominated the trapped particulate
contents at all depths. Intact or readily recognizable fecal pellets
were the next most common particles. Skeletal debris as well as
apparently living organisms were also present at all depths. We now

.,
P04
° N03+N02
° Si02
o
° 2 4 20 40 100 200
500
-E-IIOOO b: w o
1500
N2000 N
FIG. 11. Vertical nutrient distributions (umoles kg-I) at VERTEX I PIT site.

23
describe these major classes of materials based on examination of the
samples using light and scanning electron microscopy.
Individual particles of amorphous detritus corresponded generally
to one of the two major categories described previously as IIflakes ll and
IIflocs" (Wieve and Pomeroy, 1972). The flocs,. or mucus-like aggregates,
were similar in appearance to fragments from hand-collected specimens of
marine snow, and were especially prominent in the upper 100 m, and again
between the depths of 500 and 1100 m. At the other depths, the
amorphous organic detritus was dominated by flake-like particles
resembling flattened plates or membranes.
The sources of most of the flocculent detritus were not obvious,
except for two types with distinctive mucus. One recognizable type was
produced by the diatom Thalassiosira subtilis; T. subtilis colonies were
abundant in the upper 100 m, and the mucus was found in small quantities
to the bottom trap at 2000 m (Fig. 12a). (The taxonomic designation of
T. subtilis for these specimens was confirmed by G. Fryxell.) Thick
mats of threads (Fig. 12b) surround T. subtilis cells, and the
persistence of the mucilage to depths is explained partially by the
resistance of the threads to digestion, as evidenced by their presence
within fecal pellets in the traps. A chitin-like composition may
explain the chemical hardiness of these fibrils (Hasle, 1972). A second
recognizable source of flocculent mucus was larvaceans. Larvacean
houses (together with occasional specimens of intact larvaceans)
occurred in modest numbers from the surface traps to those at 2000 m.
In contrast to the flocculent detritus, the origins of the flake-like
materials were never evident.

24
.. - ,
FIG. 12. (a) Fibrillar mat produced by Thalassiosira subtilis and T. subtilis cells from the 50-m trap;(b) Thalassiosira subtilis entwined in fibrous mucilage; (c) Skeletonema costatum, a common neritic diatom, from the 2000-m trap;(d) Strombidium sp., an oligotrich ciliate from the 2000-m trap.

25
Phytop1ankton as soci ated wi th the detri tus ; nd i cated three
distinctive sources of materials in the traps. Traps from the upper 100
m contained an abundance of coccolithophores, dominated by Emiliania
huxley;, the primary coccolithophore of subarctic Pacific and north
equatorial Pacific waters, and a common species of the central North
Pacific (Okada and Honjo, 1973; Reid, 1980). The specimens of E.
huxleyi in the traps within the euphotic zone, as well as those from
greater depths, possessed coccoliths characteristic of cells from both
warm and cold water environments, mixtures we have found previously from
surface waters fo the outer California Current at this time of year.
Thalassiosira subtilis, the diatom forming mats of mucliage fibers
discussed above, was another abundant species whose presence indicated
warm oceanic water masses (Hasle, 1972). Moderate numbers of the very
large diatoms Rhizosolenia castracanei and R. imbricata var. shrubsolei
were also present in the upper traps. These diatom species occured in
i ntertwi ni ng "mats II of frus tul es in the water, associ ati ons especi ally
characteristic of oceanic, nutrient depleted regimes and water masses of
oceanic gyres (Alldredge and Silver, 1982). The combination of E.
huxleyi, and T. subtilis, and Rhizosolenia species indicates inputs of
materials to the traps from oceanic and outer California Current water
masses.
A second source of materials in the traps was indicated by the
presence of the heavily armored dinoflagellate, Ceratium dens. This
species also occurred in considerable numbers in traps from the upper
100 nl, and had been present in the Monterey Bay area for at least a
month prior to the cruise, causing a "red tide". This dinoflagellate

26
species indicated a source of materials from inshore, near-surface
waters of the central California coast during conditions of nutrient
depletion that often occur after the relaxation of upwelling (Garrison,
pers. comm.). Recognizable Ceratium debris disappeared almost
immediately below lOa m, but fragments of Rhizosolenia occurred sparsely
below the epipelagic depths, and coccolith debris occurred consistently
and abundantly to the bottom trap.
At depths of 1100 m and below, a third source of material was
indicated by the presence of new phytoplankton species in the traps.
Populations that clearly were derived from communities like those in
overlying waters were also present, and thus the mixture of the
populations suggested inputs from different sources at depth. These new
populations were dominated by Skeletonema costatum (Fig. 12c), a neritic
diatom commonly characteristic of the early stages of upwelling in the
nearshore area (Garrison, 1979). Alumino-silicate fluxes increased at
these same depths '(see below), indicating lateral input of terrigenous
materials. The change in phytoplankton composition at these depths
further suggests that these materials were originally from the coastal
zone.
In addition to the skeletal and organic detritus discussed above,
many heterotrophi c protozoans, algae, and metazoans -- judged intact
with light microscopy -- occurred in the traps. Ciliates were the most
conspicuous fonms (Fig. 12d) among the protozoans, but flagellates
(including non-pigmented, naked dinoflagellates) were also moderately
abundant. Most of the algae within the detritus were either in advanced
stages of decomposi ti on or cons; sted of empty-wa11 ed spec;mens, but

27
occasional specimens showed autofluorescence, indicating the presence of
intact protoplasts with pigment.
Metazoans were present in all the traps, and were most abundant in
the 50 to 200 m traps. In sediment traps, such organisms are usually
interpreted as being II swimners" (Knauer, et al., 1979), forms that swam
into the traps on their own. However, it is probable that some of
these, particularly the smaller forms, were associated with the detrital
debris that sank into the traps, and were part of detrital food chains . . Copepods were the most common swinmers in our samples, followed by
ostracods and copepod naup1i i • La rvaceans, gymnosomatous pteropods,
polychaetes, hydrozoan jellies (including siphonophores), amphipods, and
chaetognaths occurred in lower numbers.
The larvaceans are a particularly interesting group of "swirrmers",
because they bring with them their mucous houses', which are one of the
most readily recognized forms of floc or marine snow. Because
larvaceans are almost constantly in their houses as protection from
predators (Alldredge, 1976), the larvaceans may actually inject this
form of "detritus" when they swim into the traps. Larvacean houses were
not numerous relative to other forms of detritus, and thus would not
have introduced a significant amount of material into the traps during
VERTEX I.
Major Component Fluxes
Major component fluxes estimated using the two trap systems are
shown in Fig. 13 and Table 20 Total mass fluxes were similar at 80-100

~
!/
I' Total Part Org Matter CaC03 AI-Si
o 400 8000· 200 400 0 40 80 0 20 40 o
500
~ LV\ E
-......,
I 1000 fa. w 0
1500
.LV\ .l- I" .I 0
2000
FIG. 13. Major component fluxes (mg m 2 day-I) measured with MLML squares) trap systems during VERTEX I.
(filled squares) and UCSC (open ~

29
TABLE 2. VERTEX I major component fluxes measured with MLML and UCSC
traps. Organic matter, CaC03 and alumino-silicate fluxes
were estimated by multiplying organic C values by 2, Ca
values by 2.5, and Al quantities by 12.
Depth Total Org. Mat. CaC03 A1-Si
(m) ------------------mg -2
m -1
day ---------------
50 720 440 89 3.3
80* 300 190 57 0.84
100 210 130 53 1.9
200 160 88 53 8.5
250* 260 150 63 5.3
300 120 63 39 14
500 110 48 41 21
600 120 45 43 29
700 120 33 24 33
750* 86 18 33 15
900 150 61 26 43
1100 150 45 30 46
1700 100 30 29 39
2000 97 31 18 37
*ucsc traps.

30
m and 700-750 m, while the UCSC trap flux was about 2X higher than the
MLML trap flux in the 200-300 m depth interval.
Organic matter fluxes were estimated by doubling the organic C
flux. Good agreement was obtained between the two trap systems, with
the exception of the 200-300 m depth interval, where the UCSC trap flux
was about twice as high as the trend observed with the MLML traps. This
may have resulted from incomplete removal of zooplankton which actively
swam into the UCSC traps, or because of poor replication in the MLML C
values at 100 and 200 m. Similar calcium carbonate fluxes were obtained
wi th the two trap systems!t The 1argest di screpancy was observed for
alumino-silicate fluxes, estimated by multiplying the Al values by 12.1,
based on Taylor's (1964) crustal abundance estimate for this element
(8.23%). The MLML traps yielded fluxes about twice as high as those
obtained by UCSC. At the present time, it is unknown whether
overtrapping by MLML or undertrapping by UCSC is involved. Certainly
the trapping of rarer larger particles is not involved because the
probability of catching these particles in the MLML traps would be 64X
less than in the UCSC traps based on collecting area (0.0039 vs 0.25
m2 ). The difference in alumino-silicate fluxes might be due to the fact
that the MLML trap samples were concentrated via filtration, while the
UCSC PIT materials were concentrated via centrifugation. If significant
amounts of Al were associated with fine clay particles, it ;s possible
that they were not concentrated via the latter technique. Nevertheless,
the overall agreement between the two trap systems ;s quite good ;n view
of their different sizes and geometries, the fact that this was the
first time they were compared in the free-floating mode in the

31
California Current, and that different trap sample processing methods
were emp 1oyed.
Our VERTEX I organic C fluxes are compared with other estimates
made in the Atlantic and Pacific (Fig. 14). To simplify this task, we
did not include fluxes measured in the upper 300 m of the water column
(e.g., Staresinic, Rowe, Shaughnessey and Williams, 1978; Zeitzschel and
Zenk, 1978; Tsunogai, Uenatsu, Taneka and Harade, 1980; Sasaki and
Nishizawa, 1981; Staresinic, et al, 1982), nor those measured in bays
and enclosed basins (e.g., Bishop, Ketten and Edmond, 1978; Knauer et
al., 1979; Crisp, Brenner, Venatesan, Ruth and Kaplan, 1979; Dunbar and
Berger, 1981).
The data shown in Fig. 14 can be summarized as follows: Fluxes in
the 300-1500 m portion of the water column range from 0.1 to 2.6 mmol C
2- day-:. 1 l' · d·
near-shore waters and lowest in oligotrophic open-ocean areas. The same
applies to the 1500-4000 m depth interval, except that maximum fluxes
are only half as high (1.3 mmol C m-2 day-I). Available flux estimates
for the 4000-6000 m portion of the water column are very low, on the
order of 0.05 to 0.2 mmol C m-2 day-I. This summary is similar to that
compiled by Suess (1980).
It appears that there is order of magnitude agreement in near-shore
and open ocean C flux estimates. Hopefully, agreement will improve as
more is learned about the physics governing the entrapment of particles,
and the chemistry and biology occurring within the traps themselves.
m ... As expected, f uxes are genera .IY hi ghest 1n pro uctlve

32
FIG. 14. A comparison of VERTEX I organic C fluxes with previously pUblished Pacific and Atlantic Oceans estimates. Data for upper 300 m of water column plus those for bays, basins, etc. are not included.
Legend Key:(1) This paper, central California
A = MLML trapsB = UCSC traps
(2) Knauer, et ale (1979), northeast Pacific open ocean (3) Honjo (1980)
A = central Sargasso Sea B = tropical Atlantic C = north central Pacific
(4) Spencer, et ale (1981), Honjo STIE data, Panama (5) Honjo (1978), Sargasso Sea (6) Deuser and Ross (1980), Deuser, et ale (1981),
Sargasso Sea, bar = seasonal range (7) Rowe and Gardner (1979), northwest Atlantic Ocean (8) Hinga, et ale (1979), north Atlantic deep-sea floor (9) Martin and Knauer (1982) equatorial north Pacific
(10) Bishop, et ale (1977), equatorial Atlantic (11) Bishop, et ale (1980), Panama basin (12) Cobler and Dymond (1980), Galapagos (13) Wiebe, et ale (1976), Bahamas

33
. mmol m-2 day-I oO,......__r-I __~2r=--_~3p:..-._~4 ,
~ <]v () .~ . ••
o~ etL}I <)<1 0
~
~IA2 'I ~
-IS &2E
~ + o 3A ~
t;i ~ 383 • • 3C ~40
(;i C>5 ~ HD6
~7
08 09 EBIO () II +12 <V13
6..-.--....I.---......L-----L----'

34
ACKNOWLEDGEMENTS
We thank Andrew Soutar of Scripps Institution of Oceanography for
the schematic shown in Fig. 1. This research was supported by grants
from the National Science Foundation Marine Chemistry and Biological
Oceanography Programs (aCE #s 79-23321, 79-23322, 79-26797, 80-03200,
and 80-05180).

35
REFERENCES
ALLDREDGE, A.L. (1976) Field behavior and adaptive strategies of
Appendicularians (Chordata: Tunicata). Marine Biology 38, 29-39.
ALLDREDGE, A.L. and M.W. SILVER. (1982) Abundance and production rates
of floating diatom mats (Rhizosolenia castracanei and~. imbricata
yare shrubsolei) in the Eastern Pacific Ocean. Marine Biology 66,
83-88.
ATLAS, E.l., l.I. GORDON, S.W. HAGER and P.K. PARK. (1971) A practical
manual for use of the Technicon autoanalyzer in seawater nutrient
analyses (revised). Tech. Rept. 215, Ref. 71-22, Oregon State
University School of Oceanography. 48 p.
BISHOP, J.K.B., J.M. EDMOND, D.R. KETTEN, M.P. BACON and W.B. SIlKER.
(1977) The chemistry, biology and vertical flux of particulate
matter from the upper 400 m of the equatorial Atlantic Ocean.
Deep-Sea Research 24,511-548.
BISHOP, J.K.B., D.R. KETTEN and J.M. EDMOND. 1978. The chemistry, bio
logy and ve~tical flux of particulate matter from the upper 400 m
of the Cape Basin in the southeast Atlantic Ocean. Deep-Sea
Research 25,1121-1162.
BISHOP, J.K.B., R.W. COLLIER, D.R. KETTENS and J.M. EDMOND. (1980) The
chemistry, biology and vertical flux of particulate matter from the
upper 1500 m of the Panama Basin. Deep-Sea Research 27,615-640.
BROENKOW, W.W. (1982) A comparison between geostrophic and current
meter observations in a California Current eddy. Deep-Sea
Research 29,1303-1311.

36
BROENKOW, W.W. and W.M. SMETHIE, JR. (1978) Surface circulation and
replacement of water in Monterey Bay. Estuarine and Coastal
Marine Science 6, 583-603.
BROENKOW, W.W. and N. GREENE. (1981) Oceanographic results during
VERTEX particle interceptor trap experiment, 17 August to 8 Septem
ber 1980. Moss Landing Marine Laboratories Tech. Pub1. 81-1. Moss
Landing, CA. 109 p.
COBLER, R. and J. DYMOND. (1980) Sediment trap experiment on the Gala
pagos spreading center, equatorial Pacific. Science 209, 801-803.
CRISP, P.T., S. BRENNER, M.I. VENTAKESAN, E. RUTH and I.R. KAPLAN.
(1979) Organic chemical characterization of sediment-trap particu~
lates from San Nicholas, Santa Barbara, Santa Monica, and San
Pedro Basins, California. Geochimica et Cosmochimica Acta 43,
1791-1801.
DEUSER, W.G. and E.H. ROSS. (1980) Seasonal change in the flux of or
ganic carbon to the deep Sargasso Sea. Nature 283, 364-365.
DEUSER, W.G., E.H. ROSS and R.F. ANDERSON. (1981) Seasonality in the
supply ~f sediment to the deep Sargasso Sea and implications for
the rapid transfer of matter to the deep ocean. Deep-Sea Research
28A, 495-505.
DUNBAR, R.B. and W.H. BERGER. (1981) Fecal pellet flux to modern
bottom sediment of Santa Barbara Basin (California) based on sedi~
ment trapping. Geological Society of America Bulletin 92, 212-218.

37
I:BER, L.E. (1977) Contoured depth-time charts (0 to 200 m, 1950 to
1966) of temperature, salinity, oxygen and sigma-t at 23 CalCOFI
stations in the California Current. California Cooperative Oceanic
Fisheries Investigations Atlas No. 25. 231 p.
GARDNER, W.O. (1980a) Sediment trap dynamics and calibration: a labo
ratory evaluation. Journal of Marine Research 38, 17-40.
GARDNER, W.O. (1980b) Field assessment of sediment traps. Journal of
Marine Research 38, 41-52.
GARRISON, D.l. (1979) Monterey Bay phytoplankton. I. Seasonal cycles
of phytoplankton assemblages. Journal of Plankton Research 1,
241-265.
HASlE, G.R. (1972) Thalassiosira subtilis (Bacillariophyceae) and two
allied species. Norwegian Journal of Botany 19, 111-137.
HICKEY, S.M. (1979) The California Current system: hypothesis and
facts. Progress in Oceanography 8, 191-279.
HINGA, K.R., J. MC N°. SIEBURTH and G.R. HEATH. (1979) The supply and
use of organic material at the deep-sea floor. Journal of Marine
Research 37,557-579.
HONJO, S. (1978) Sedimentation of materials in the Sargasso Sea at a
5367 m deep station. Journal of Marine Research 36, 469-492.
HONJO, S. (1980) Material fluxes and modes of sedimentation in the
mesopelagic and bathypelagic zones. Journal of Marine Research
38, 53-97.
HONJO, S., D.W. SPENCER and J.W. FARRINGTON. (1982) Deep advection
transport of lithogenic particles in Panama Basin. Science 216,
516-518.

38

•

40
•
•
WICKHAM, J.B. (1975) Observations of the California Countercurrent.
Journal of Marine Research 33, 325-340.
WIEBE, W.J. and L.R. POMEROY. (1972) Microorganisms and their associa
tion with aggregates and detritus in the sea: A microscopic study.
Memorie dell· Instituto Italiano ~ Idrobiologia 29 (supplement),
325-352.
WIEBE, P.H., S.H. BOYD and C. WINGET. (1976) Particulate matter sink
ing to the deep-sea floor at 2000 m in the Tongue-of-the-Ocean,
Bahamas, with a description of a new sedimentation trap. Journal
of Marine Research 34, 341-354.