Embed Size (px)
Citation preview

___________________________Corresponding author: Sohail Ahmad Jan, Department of Biotechnology, Quaid-i-AzamUniversity, Islamabad, Pakistan ,Phone: +92-345-9118133, E-mail: [email protected];[email protected]
UDC 575.630DOI: 10.2298/GENSR1601323J
Original scientific paper
MORPHO- BIOCHEMICAL EVALUATION OF Brassica rapa SUB-SPECIES FOR SALTTOLERANCE
Sohail Ahmad JAN1*, Zabta Khan SHINWARI1, Malik Ashiq RABBANI2
1Department of Biotechnology, Quaid-i- Azam University, Islamabad, Pakistan2Plant Genetic Resources Institute, National Agricultural Research Centre, Islamabad, Pakistan
Jan A. S., Z. K.Shinwari, M. A.Rabbani (2016): Morpho- biochemical evaluation ofBrassica rapa sub-species for salt tolerance- Genetika, Vol 48, No.1, 323 -338.Salt stress is one of the key abiotic stresses that affect both the qualitative and quantitativecharacters of many Brassica rapa sub-species by disturbing its normal morpho-biochemical processes. Therefore, the present research work was designed to study theeffect of different NaCl events (0, 50,100 and 150 mmol) on morphological andbiochemical characters and to screen salt tolerant genotypes among brown, yellow andtoria types of B. rapa sub-species. The plants were grown in test tubes with addition offour level of NaCl (0, 50,100 and 150 mmol). The effect of salinity on shoot and rootlength, shoot/ root fresh and dry weight, relative water content (RWC), proline andchlorophyll a, b, a+b contents was recorded after 4 weeks of sowing. The genotype 22861(brown type) showed excellent morphological and biochemical performance at all stresslevels followed by Toria-Sathi and Toria-A respectively as compared to Check varietyTS-1. The genotype 26158 (yellow type) gave very poor performance and retard growth.The %RWC values and chlorophyll a, b and a+b contents were decreased several foldswith the increase of salt concentration. While, the proline contents was increased withraising of salt stress. The brown and toria types showed maximum tolerance to salt stressat early germination stages as compare to yellows one. The present study will serve asmodel to develop quick salt tolerant genotypes among different plant sub-species againstsalt stress.
Key words: Brassica rapa, early germination, salt stress, salt tolerance.
INTRODUCTIONSalt is one of major environmental extreme that badly affect both soil texture and
composition (MARTINEZ-BELTRAN and MANZUR, 2005). The high concentrations of salts in soil

324 GENETIKA, Vol. 48, No.1, 323-338, 2016
reduce the normal morpho-physiological and other biological processes by nutrient and ionunbalances (ASHRAF and HARRIS, 2004; GARTHWAITE et al., 2005). The low to high salt stressleads cell injury by negatively inhibiting reactive oxygen species (ROS). The salt stress alsodecrease the amount of essential nutrients that retard normal plant growth and lead plant death(CUIN and SHABALA, 2007, VITAL et al., 2008, SHINWARI et al., 1998, MASOOD et al., 2005).Drought and salinity are the two major abiotic stresses that affect both the qualitative andquantitative characters of many brassica species such as rapeseed (Brassica napus L.), mustard(Brassica juncea L.), cabbage (Brassica oleracea L.) and turnip rape (Brassica rapa L.). Thediploid species of brassica (Brassica rapa) is more sensitive to salt stress then other polyploidspecies (KUMAR, 1995). The high salt concentration of salts in soil and irrigation water inhibitsseed germination of many brassica species. In some brassica species, germination occurs but plantsshow retard growth and development (ZAMANI et al., 2010). So, it is important to screen improvedgenotypes against these environmental extremes by using efficient identification and selectionmethod (ASHRAF and MCNEILLY, 2004, ALMODARES et al., 2007, ISLAM and KARIM, 2010).
The effect of salinity on both morpho-physiological and biochemical processes of manyimportant plant species have been reported by many researchers. QUESADA et al. (2002) describedthat low to high salt stress inhibit normal germination rate and plant fresh and dry weight up toseveral folds of many important Arabidopsis genotypes. MUNIR et al. (2013) calculated thephotosynthetic rate, proline content and intracellular or intercellular ion content of many canolacultivars. Their findings showed the low photosynthetic rate and increase proline and ion contentsat high NaCl levels (ULFAT et al., 2007). Different salt levels retards normal growth anddevelopment of many brassica species (KUMAR et al., 2009). The total chlorophyll content ofwinter type radish (Raphanus sativus L.) was decreased up to several folds at low to moderate andhigh salt stress levels (MUNIR et al., 2013). The salt stress also leads abnormal Na+ ion level inshoot, which negatively inhibit normal plant biochemical processes (KHAYAT et al., 2010). Salttolerant ability varies among B. rapa sub-species. Many B. rapa sub-species are so sensitive to saltstress and have no optimized protocol to screen salt tolerant genotypes at early germination stage.Therefore, the present study was conducted to develop a quick, novel screening system for theidentification and further evaluation of brown, yellow and Toria sub-species of B. rapa at earlygermination stage.
MATERIALS AND METHODSThe present trial was conducted in Plant Genetic Resources Institute (PGRI), National
Agriculture Research Centre (NARC), Islamabad, Pakistan. The fully mature seeds of threeBrassica rapa sub-species were provided by the gene bank of PGRI, NARC, Islamabad, Pakistan.
Sterilization of SeedsThe seeds were first treated with doubled distilled water for 5-10 minutes with continuous
shaking. The seeds were then treated with 10% Clorox bleach for 10-15 minutes with gentleshaking followed by a single washed with double distilled water for minutes. The sterilize seedswere transferred into sterilize petri dishes having autoclaved filter paper.
Seeds Germination with Different Salt LevelsThe sterilize seeds were kept on the top of filter papers inside test tubes. About 3-4 ml of NaClwas added at four different concentrations (0, 50, 100 and 150 mmol). The tubes were stored in

S .A..JAN et al: SALT TOLERANCE OF Brassica rapa SUB SPECIES 325
incubator at 25±2 ºC under 16 hours light and 8 hours dark condition. The controlled plants weretreated with distilled water only.
Morpho-Biochemical ScreeningThe effect of salt stress on various morphological characters was tested. Many
morphological variations such as shoot length, root length, shoot fresh and dry weight and rootfresh and dry weight were noted at different salt concentrations after 4 weeks of germination.
Measurement of Chlorophyll a, b, a+b and Proline ContentsAfter 4 weeks of salt stress, young leaves samples were taken from both controlled and
salt treated plants and were subjected to biochemical screening tests. In first step leaves weretreated with 80% acetone, followed by centrifugation at 8000 rpm for 10 minutes. The clearsupernatant was separated from rest of mixture and its absorbance was tested at UV-visiblespectrophotometer at 645 and 663 nm. The chlorophyll a, b and a+b contents were also recordedfrom both controlled and salt tested plants by using protocol of ARNON (1949). The proline testwas carried out by using standard protocol of BATES et al. (1973) with little modification.
Measurement of Relative Water Content (RWC)The percent of Relative Water Content (% RWC) was calculated by using method of
BARRS and WEATHERLY (1962). The percent Relative Water Content (RWC) was measured byusing following formula;
RWC (%) = [(FW–DW)/(TW–DW)]×100
The Fresh Weight (FW) was measured by separating all of leaf disks from each plant andplaced in distilled water for 8 hours at dark condition and weighted. Turgid Weight (TW) wasmeasured by dehydrating leaves, to removes excess moistures. The Dry Weight (DW) wasmeasured by desiccating leaves at 80 °C for 1-2 days.
Data AnalysisThe data was analysed by using statistical software’s, Statistic v. 8.1 (ANALYTICAL and
SOFTWARE, 2005) and Origin 8.1.
RESULTSSalt is one the key environmental extreme that directly affects the normal morphological
and other biological processes of many B. rapa sub-species. The low to high salt concentrations insoil disturb its normal plant growth and development. The present protocol was designed to studythe effect of different salt stress on some of the important morpho-biochemical process of B. rapaecotypes. In present experiment the effect of low to moderate and high salt stress were studied theeffect of salt stress on three important Brassica rapa sub-species i.e. 22861 (brown type), 26158(yellow type), Toria types (Toria-A and Toria-Sathi) were tested along with a check variety TS-1.The morpho-biochemical responses varied with genotypes and different NaCl concentrations (0,50, 100 and 150 mmol).
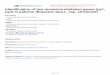
326 GENETIKA, Vol. 48, No.1, 323-338, 2016
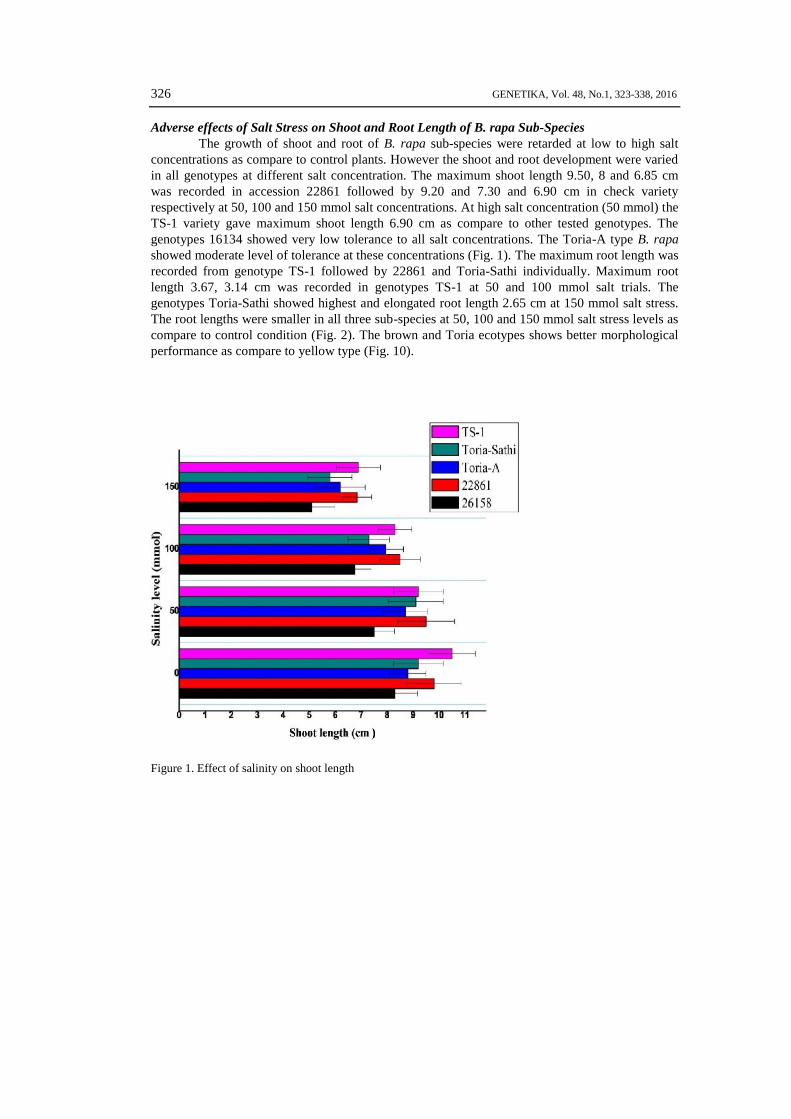
Adverse effects of Salt Stress on Shoot and Root Length of B. rapa Sub-SpeciesThe growth of shoot and root of B. rapa sub-species were retarded at low to high salt
concentrations as compare to control plants. However the shoot and root development were variedin all genotypes at different salt concentration. The maximum shoot length 9.50, 8 and 6.85 cmwas recorded in accession 22861 followed by 9.20 and 7.30 and 6.90 cm in check varietyrespectively at 50, 100 and 150 mmol salt concentrations. At high salt concentration (50 mmol) theTS-1 variety gave maximum shoot length 6.90 cm as compare to other tested genotypes. Thegenotypes 16134 showed very low tolerance to all salt concentrations. The Toria-A type B. rapashowed moderate level of tolerance at these concentrations (Fig. 1). The maximum root length wasrecorded from genotype TS-1 followed by 22861 and Toria-Sathi individually. Maximum rootlength 3.67, 3.14 cm was recorded in genotypes TS-1 at 50 and 100 mmol salt trials. Thegenotypes Toria-Sathi showed highest and elongated root length 2.65 cm at 150 mmol salt stress.The root lengths were smaller in all three sub-species at 50, 100 and 150 mmol salt stress levels ascompare to control condition (Fig. 2). The brown and Toria ecotypes shows better morphologicalperformance as compare to yellow type (Fig. 10).
Figure 1. Effect of salinity on shoot length
S .A..JAN et al: SALT TOLERANCE OF Brassica rapa SUB SPECIES 327
Figure 2. Effect of salinity on root length
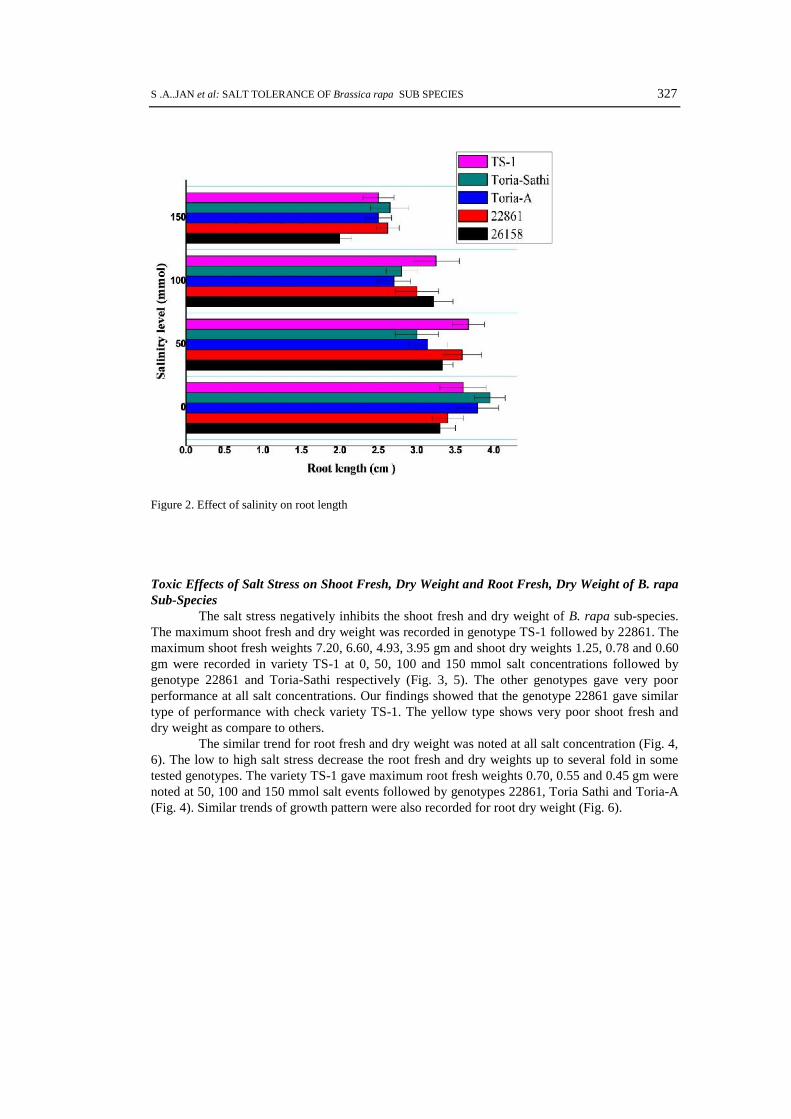
Toxic Effects of Salt Stress on Shoot Fresh, Dry Weight and Root Fresh, Dry Weight of B. rapaSub-Species
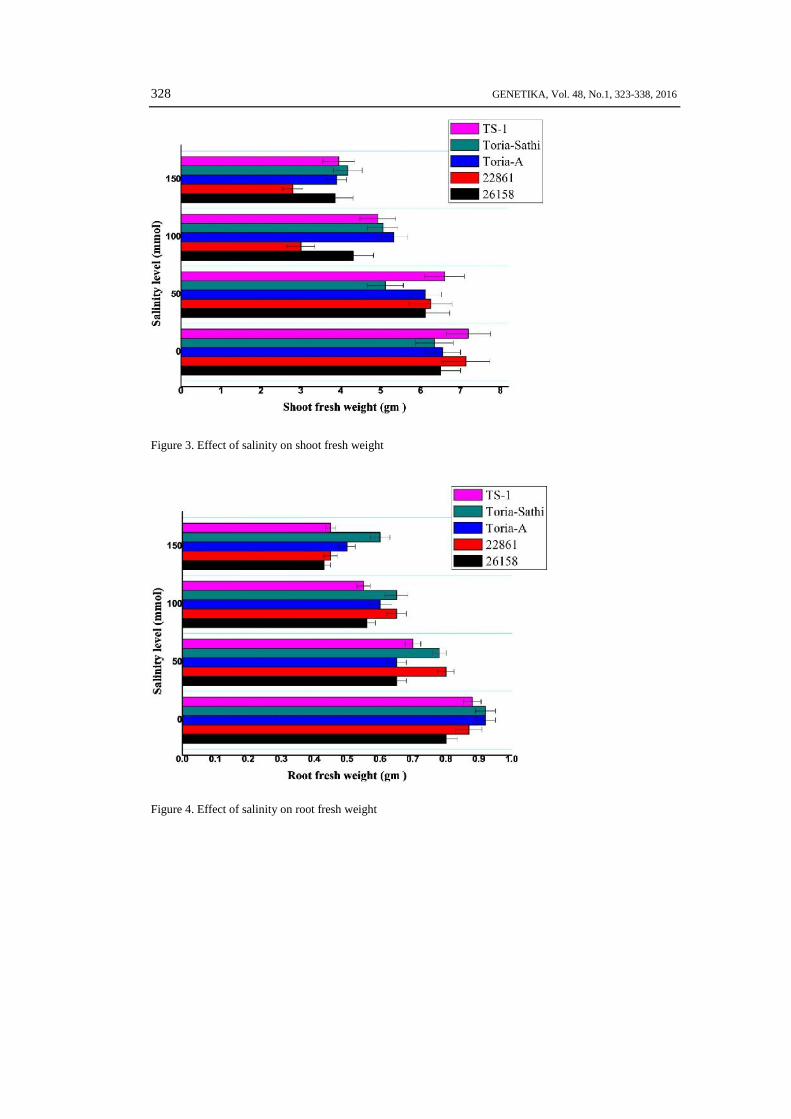
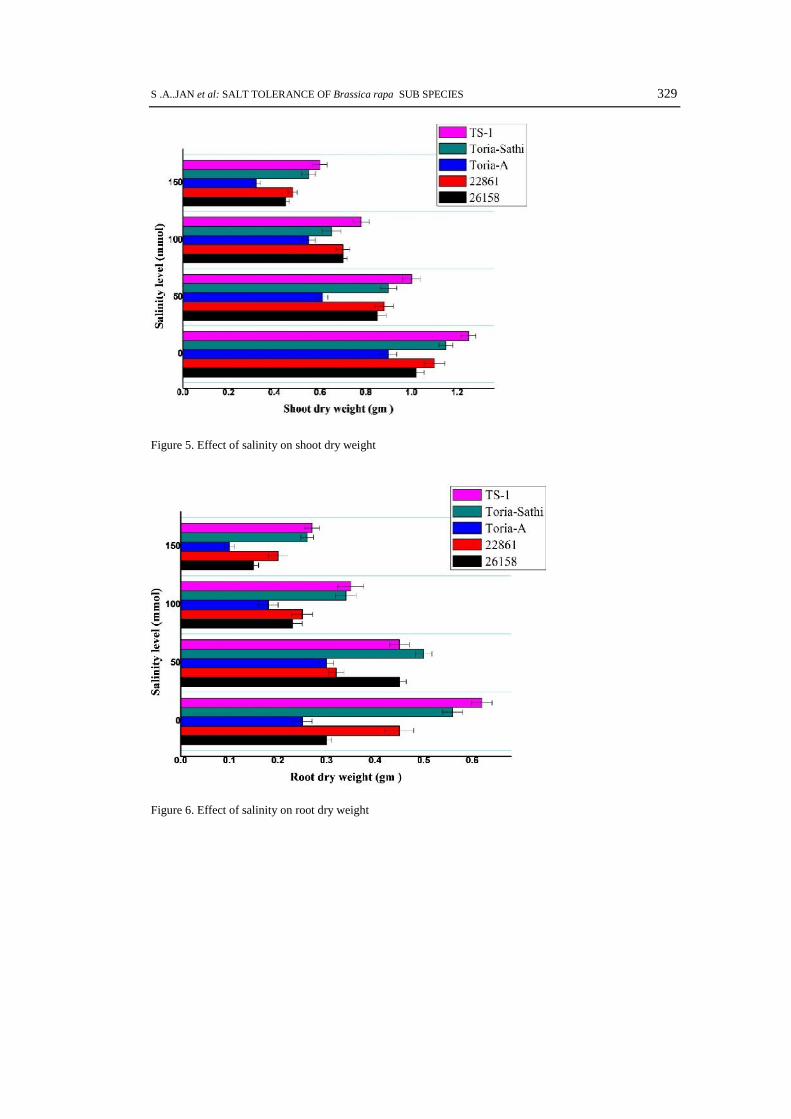
The salt stress negatively inhibits the shoot fresh and dry weight of B. rapa sub-species.The maximum shoot fresh and dry weight was recorded in genotype TS-1 followed by 22861. Themaximum shoot fresh weights 7.20, 6.60, 4.93, 3.95 gm and shoot dry weights 1.25, 0.78 and 0.60gm were recorded in variety TS-1 at 0, 50, 100 and 150 mmol salt concentrations followed bygenotype 22861 and Toria-Sathi respectively (Fig. 3, 5). The other genotypes gave very poorperformance at all salt concentrations. Our findings showed that the genotype 22861 gave similartype of performance with check variety TS-1. The yellow type shows very poor shoot fresh anddry weight as compare to others.
The similar trend for root fresh and dry weight was noted at all salt concentration (Fig. 4,6). The low to high salt stress decrease the root fresh and dry weights up to several fold in sometested genotypes. The variety TS-1 gave maximum root fresh weights 0.70, 0.55 and 0.45 gm werenoted at 50, 100 and 150 mmol salt events followed by genotypes 22861, Toria Sathi and Toria-A(Fig. 4). Similar trends of growth pattern were also recorded for root dry weight (Fig. 6).
328 GENETIKA, Vol. 48, No.1, 323-338, 2016
Figure 3. Effect of salinity on shoot fresh weight
Figure 4. Effect of salinity on root fresh weight
S .A..JAN et al: SALT TOLERANCE OF Brassica rapa SUB SPECIES 329
Figure 5. Effect of salinity on shoot dry weight
Figure 6. Effect of salinity on root dry weight
330 GENETIKA, Vol. 48, No.1, 323-338, 2016
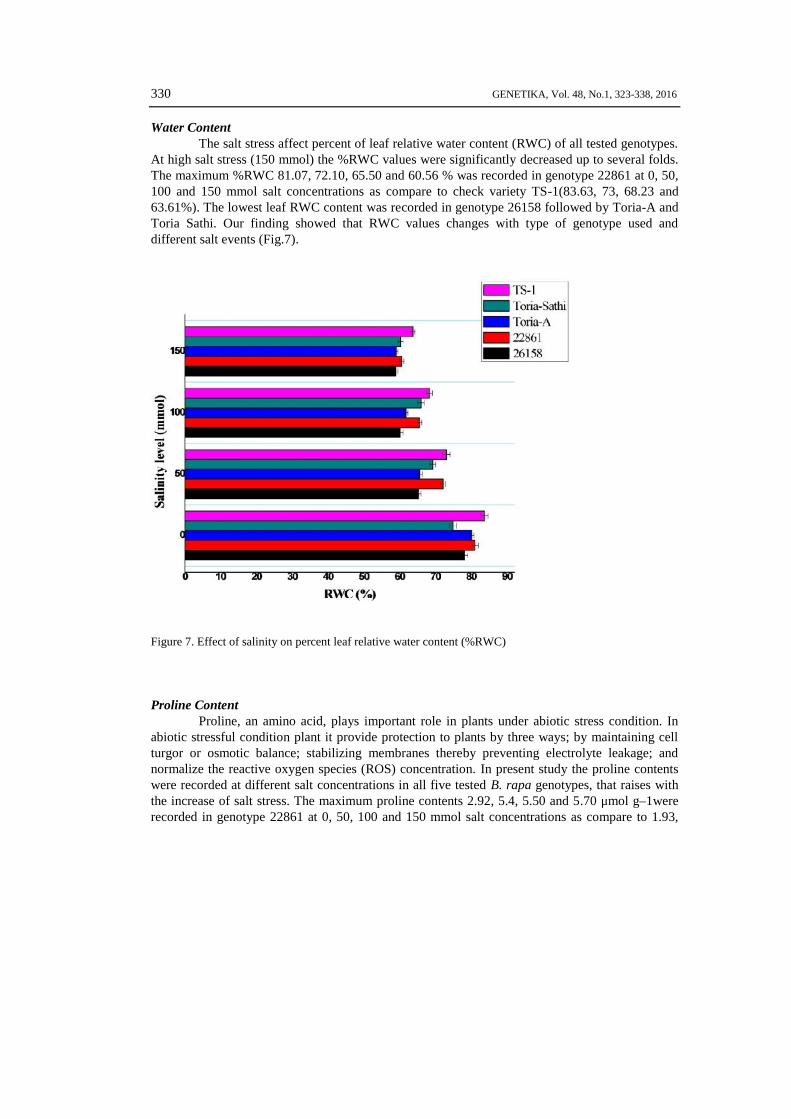
Water ContentThe salt stress affect percent of leaf relative water content (RWC) of all tested genotypes.
At high salt stress (150 mmol) the %RWC values were significantly decreased up to several folds.The maximum %RWC 81.07, 72.10, 65.50 and 60.56 % was recorded in genotype 22861 at 0, 50,100 and 150 mmol salt concentrations as compare to check variety TS-1(83.63, 73, 68.23 and63.61%). The lowest leaf RWC content was recorded in genotype 26158 followed by Toria-A andToria Sathi. Our finding showed that RWC values changes with type of genotype used anddifferent salt events (Fig.7).
Figure 7. Effect of salinity on percent leaf relative water content (%RWC)
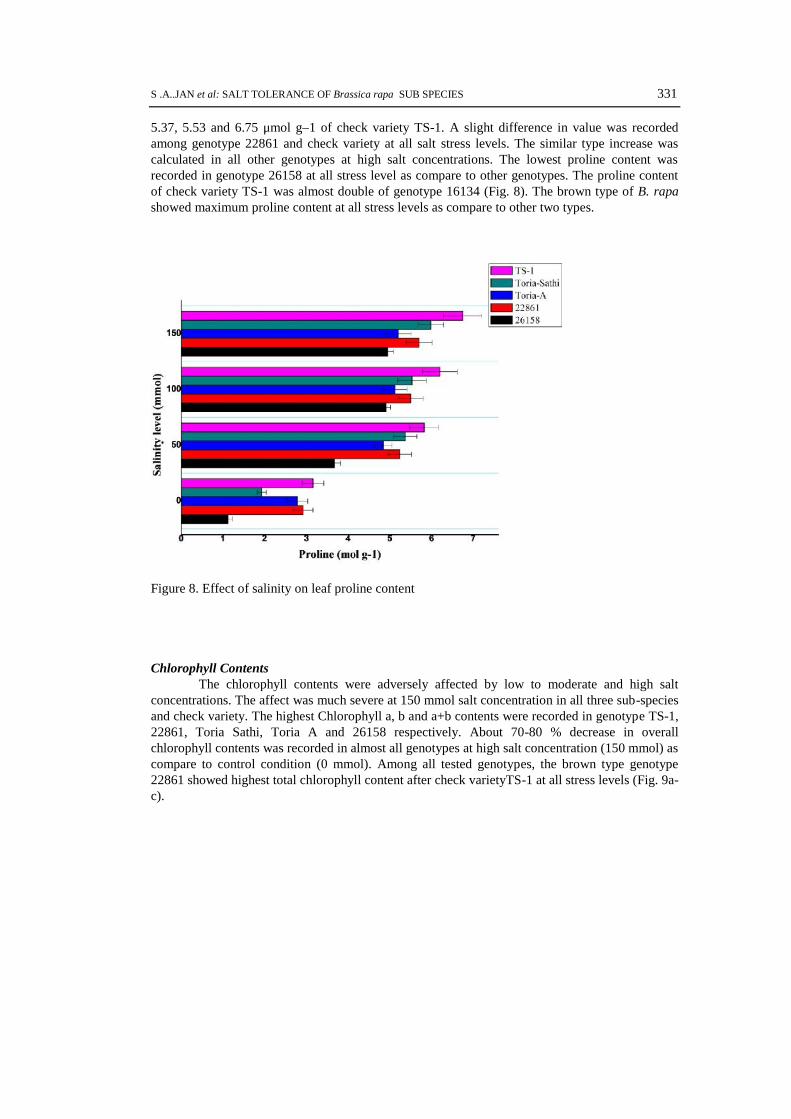
Proline ContentProline, an amino acid, plays important role in plants under abiotic stress condition. In
abiotic stressful condition plant it provide protection to plants by three ways; by maintaining cellturgor or osmotic balance; stabilizing membranes thereby preventing electrolyte leakage; andnormalize the reactive oxygen species (ROS) concentration. In present study the proline contentswere recorded at different salt concentrations in all five tested B. rapa genotypes, that raises withthe increase of salt stress. The maximum proline contents 2.92, 5.4, 5.50 and 5.70 μmol g–1wererecorded in genotype 22861 at 0, 50, 100 and 150 mmol salt concentrations as compare to 1.93,
S .A..JAN et al: SALT TOLERANCE OF Brassica rapa SUB SPECIES 331
5.37, 5.53 and 6.75 μmol g–1 of check variety TS-1. A slight difference in value was recordedamong genotype 22861 and check variety at all salt stress levels. The similar type increase wascalculated in all other genotypes at high salt concentrations. The lowest proline content wasrecorded in genotype 26158 at all stress level as compare to other genotypes. The proline contentof check variety TS-1 was almost double of genotype 16134 (Fig. 8). The brown type of B. rapashowed maximum proline content at all stress levels as compare to other two types.
Figure 8. Effect of salinity on leaf proline content
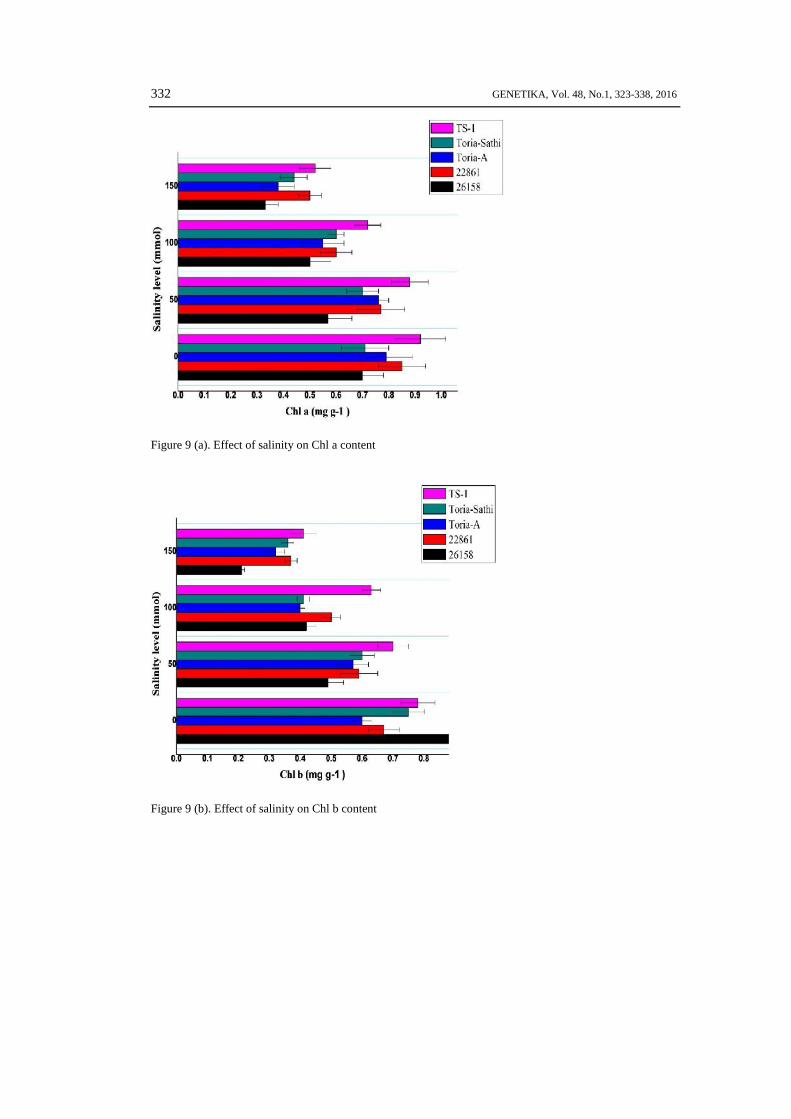
Chlorophyll ContentsThe chlorophyll contents were adversely affected by low to moderate and high salt
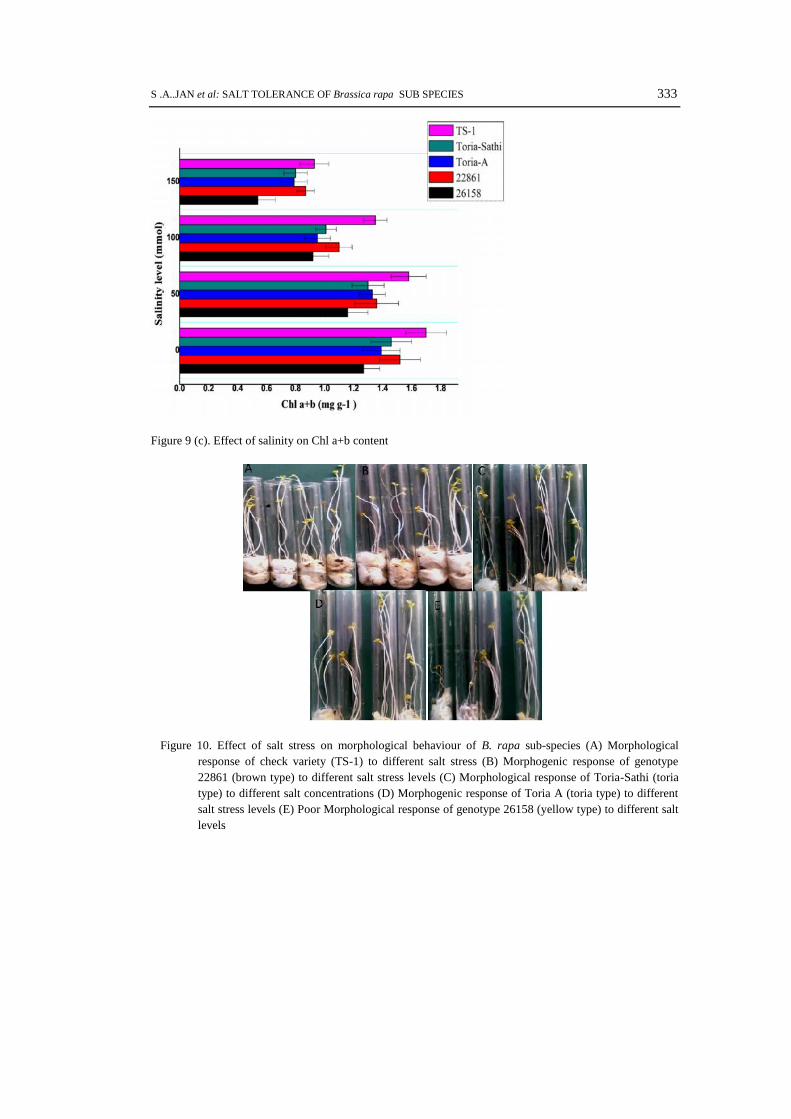
concentrations. The affect was much severe at 150 mmol salt concentration in all three sub-speciesand check variety. The highest Chlorophyll a, b and a+b contents were recorded in genotype TS-1,22861, Toria Sathi, Toria A and 26158 respectively. About 70-80 % decrease in overallchlorophyll contents was recorded in almost all genotypes at high salt concentration (150 mmol) ascompare to control condition (0 mmol). Among all tested genotypes, the brown type genotype22861 showed highest total chlorophyll content after check varietyTS-1 at all stress levels (Fig. 9a-c).
332 GENETIKA, Vol. 48, No.1, 323-338, 2016
Figure 9 (a). Effect of salinity on Chl a content
Figure 9 (b). Effect of salinity on Chl b content
S .A..JAN et al: SALT TOLERANCE OF Brassica rapa SUB SPECIES 333
Figure 9 (c). Effect of salinity on Chl a+b content
Figure 10. Effect of salt stress on morphological behaviour of B. rapa sub-species (A) Morphologicalresponse of check variety (TS-1) to different salt stress (B) Morphogenic response of genotype22861 (brown type) to different salt stress levels (C) Morphological response of Toria-Sathi (toriatype) to different salt concentrations (D) Morphogenic response of Toria A (toria type) to differentsalt stress levels (E) Poor Morphological response of genotype 26158 (yellow type) to different saltlevels

334 GENETIKA, Vol. 48, No.1, 323-338, 2016
DISCUSSIONSalt stress affects the morpho-physiological, biochemical and other molecular processes
of B. rapa sub-species. Therefore, proffer screening of improved salt tolerant genotypes isimportant against this environmental extreme. In present study the effect of various salt stresslevels on growth performance and other morpho-biochemical processes of B. rapa subspeciesbrown, yellow and toria was studied. Our finding shows significantly different types of variationamong all three types, at low to moderate and high salt levels. Some genotype showed almostsimilar performance as check variety while, others gave very poor performance even in low saltstress, and that shows that salt tolerance ability vary among genotypes and with different NaClconcentrations. Abiotic stresses affect the yield and other performance of many brassica speciesand the responses among genotypes varies with different salt concentrations (SHAHBAZI et al.,2011, UMAR et al., 2011, ALAM et al., 2014).
Various morphological parameters were studied under three different salt stress levels(50, 100 and 150 mmol) and at control condition (0 mmol) of four new genotypes (26158, 22861,Toria-A and Toria Sathi) along with a check variety TS-1. The salt stress significantly decreasedthe growth of shoot length, root length, shoot fresh, dry weight and root fresh, dry weight up toseveral fold. The genotype 22861 showed excellent growth rate followed by Toria Sathi and ToriaA at all stress levels as compare to check variety. The morphological performance of genotype22861 is almost similar to check variety, that was opposite for genotype 26158. The salt stressaffects the morphological characters of many important B. napus species. The effect was moresevere when the salts stress rising from 50 to 150 mmol (SHAHBAZI et al., 2011). Our finding arealso in line with (ASHRAF and HARRIS, 2004; MUNNS and TESTER, 2008) those studied that low tohigh salt concentrations in soil affect both morpho-biochemical processes of plants that negativelyinhibit the growth and development. The high salt concentration of NaCl (7 and 9 dS m-1) effectimportant morphological characters such as germination rate, root and shoot length, fresh and dryroot weight, fresh and shoot weights of all tested pumpkins varieties (DADASHPOUR, 2012).According to VITAL et al. (2008) high salt stress disturb normal ion-balance uptake process,produce some toxic substances and decrease the availability of important essential nutrients. UMARet al. (2011) noted different sorts of performance at different salinity levels in different Brassicacampestris genotypes. The morpho-biochemical responses vary with genotypes and type of saltstress used (MUNNS and TESTER, 2008; ARSHI et al., 2005; MUNNS et al., 2006).
The percent leaf relative water content (%RWC) values decreases with the increase of saltstress level from 50 to 150 mmol. The genotype 22861 gave maximum %RWC value at all stresslevel as compare to other tested genotypes. The %RWC value was lowest for genotype 26158. The%RWC values decrease up to 60-70% in almost all genotypes at 150 mmol salt stress level (Fig.2). The %RWC values decreases with increase of drought stress in many important B. napus, B.campestris and B. juncea cultivars (ALAM et al. 2014). Our findings are also in line with that ofGUPTA et al. (2001) and MEENA et al. (2003). KHALATBARI et al. (2013) reported that high NaClconcentrations (50 and 100 mmol) effect the morpho-physiological characters of wild (Columbia-0) and vte4 mutant Arabidopsis plants. The growth, development and relative water content wassignificantly decreased with high salt concentration. The cold stress decrease % RWC values, upto many fold in many important tomato cultivars SHAH et al. (2014).
When plants are subjected to different abiotic stress the proline increases with theincrease, that maintain normal osmotic pressure, help in signal transduction/gene expression andperform antioxidant activities too (BATES et al., 1973; NAHAR et al., 2013). From Fig. 8 we noted

S .A..JAN et al: SALT TOLERANCE OF Brassica rapa SUB SPECIES 335
that proline content almost double in many genotypes when the salt stress increase from 50 mmolto 150 mmol. The increase of proline contents help plant to survive in such adverse condition. Themaximum proline content was recorded in brown type 22861 while it is lowest for yellowtype26158 as compare to check variety TS-1. The leaf proline contents were maximum with theincrease level of drought stress in B. campestris (ALAM et al. 2014). The increase level of prolinecontent was noted at high drought stress condition in many diverse brassica species (DEEPAK et al.,1995). These results are also in agreement with previous studies (ALI and ASHRAF, 2011; NOUNJANet al., 2012).
The low to moderate or high salt stress also affect the photosynthetic rate via decreasingthe chlorophyll contents of many important B. rapa sub-species. The chlophyl a, b and a+bcontents were decreased many folds in all three B. rapa sub-species. The brown type 22861showed excellent chlorophyll contents followed by Toria Sathi Toria-A and 26158 respectively ascompare to check variety. A decrease amount of total chlorophyll content (70-80%) was recordedat 150 mmol salt stress as compare to controlled condition. Abiotic stresses such as drought,salinity and frost decrease the photosynthetic rate via alternation of many important photosyntheticpigments (ANJUM et al., 2011; PANDEY et al., 2012). The extreme drought and salt stresses inhibitthe chlorophyll content of several important plant species (CHAVES et al., 2003, RENOLDS et al.,2005). The salt stress decreases the amounts of Chlorophyll a, b and a+b up to several folds inmany brassica species such as B. napus, B. juncea and B. campestris (ALAM et al. 2014). Theseresults are also in commitment with KAUSER et al. (2006).
Received October 24th, 2015Accepted January 25th, 2016
REFERENCESALAM, M.M., K. NAHAR, M. HASANUZZAMAN and M. FUJITA (2014): Trehalose-induced drought stress tolerance: A
comparative study among different Brassica species. Plant Omics J., 7(4):271-283.ALI, Q. and M. ASHRAF (2011): Induction of drought tolerance in maize (Zea mays L.) due to exogenous application of
trehalose: Growth, photosynthesis, water relations and oxidative defence mechanism. J. Agron. Crop Sci.,197:258–271.
ALMODARES, A., M.R. HADI and B. DOSTI (2007): Effects of salt stress on germination percentage and seedling growth insweet sorghum cultivars. J. Biol. Sci., 7: 1492–1495.
ANALYTICAL and SOFTWARE (2005): Statistix version 8.1: User’s manual. Analytical Software, Tallahassee, Florida.ANJUM, S.A., X. XIE, M. FAROOQ, L. WANG, L. XUE, M. SHAHBAZ and J. SALHAB (2011): Effect of exogenous methyl
jasmonate on growth, gas exchange and chlorophyll contents of soybean subjected to drought. Afr. J.Biotechnol., 10: 9640-9646.
ASHRAF, M. and P.J.C. HARRIS (2004): Potential biochemical indicators of salinity tolerance in plants. Plant Sci., 3(16):166.ARNON, D. I. (1949): Copper, enzymes in isolated chloroplasts Poly phenoloxidase in Beta Vulgarus. Plant Physiol.,
63:1143-1148.ARSHI, A., M.Z. ABDIN and M. IQBAL (2005): Ameliorative effects of CaCl2 on growth, ionic relations and proline content ofsenna under salinity stress. J. Plant Nutr., 28: 101-125.ASHRAF, M. and T. MCNEILLY (2004): Salinity tolerance in Brassica Oilseeds. Critical Reviews in Plant Sciences, 2(23):
157-174.BARRS, H.D. and P.E. WEATHERLEY (1962): A re-examination of the relative turgidity technique for estimating water deficits
in leaves. Aust J Biol Sci., 15:413–428.BATES, L.S., R. P. WALDREN and I. D. TEARE (1973): Rapid determination of free proline for water-stress studies. Plant Soil.,
39: 205-207.

336 GENETIKA, Vol. 48, No.1, 323-338, 2016
CUIN, T.A. and S. SHABALA (2007): Compatible solutes reduce ROS-induced potassium efflux In Arabidopsis roots. PlantCell Env., 230: 875-885.
CHAVES, M. M., J. P. MAROCO and J. S. PEREIRA (2003): Understanding Plant response to drought: from genes to the wholeplant. Func. Plant Biol., 30:239-264.
DADASHPOUR, A. (2012): Impacts of different salt source and concentrations on germination and seedling growth of manypumpkin seeds used as rootstoch in Iran. Genetika., 44 (2):235 - 250.
DEEPAK, M., P. N. WATTAL and D. MATHUE (1995): Influence of water stress on seed yield of Canadian mustard at floweringand role of metabolic factors. Plant Physiol. Bioch., 22:115-118.
GARTHWAITE. A.J., R.V. BOTHMER and T.D. COLMER (2005): Salt tolerance in wild Hordeum species is associated withrestricted entry of Na+ and Cl– into the shoots.J. Exp. Bot., 56:2365-2378.
GUPTA, N.K., S. GUPTA and A. KUMAR (2001): Effect of water stress on physiological attributes and their relationship withgrowth and yield of wheat genotypes at different stages. J. Agron. Crop Sci. Ger., 186:52-62.
ISLAM, M.M. and M.A. KARIM (2010): Evaluation of rice genotypes at germination and early seedling stage for theirtolerance to salinity. Agriculturists, 8:57–65.
KAUSER, R., H.R. ATHAR and M. ASHRAF (2006): Chlorophyll Fluorescence: A potential Indicator for rapid assessment ofwater stress tolerance in canola (Brassica Napus L.). Pak. J. Bot., 38:1501-1509.
KHALATBARI, A.A., H. Z.E. JAFFAR, M. MAHMOOD, R. OTHMAN and A.M. KHALATBARI (2013): Effects of salt stress on wildtype and vte4 mutant Arabidopsis thaliana………..a: model plant to engineer tolerance towards salinity.Genetika, 45(3):777-791.
KHAYAT, P.N., S. JAMAATI-E-SOMARIN, R. ZABIHI-E-MAHMOODABAD, A. YARI, M. KHAYATNEZHAD and R. GHOLAMIN
(2010): Screening of salt tolerance canola cultivars (Brassica napus L.) World Appl. Sci. J., 10:817–820.KUMAR, G., R.S. PURTY, M.P. SHARMA, S.L. SINGLA-PAREEK and A. PAREEK (2009): Physiological responses among Brassica
species under salinity stress show strong correlation with transcript abundance for SOS pathway-related genes.J. Plant Physiol., 166:507–520.
KUMAR, D. (1995): Salt tolerance in oilseed brassicas-present status and future prospects. Plant Breed., 65(10):1439-1477.MASOOD, S., Y. SEIJI, Z. K. SHINWARI and R. ANWAR (2005): Mapping quantitative trait loci QTLs) for salt tolerance in rice
(Oryza sativa) using RFLPs. Pak. J. Bot., 36(4):825 -834.MARTINEZ-BELTRAN J. and C.L. MANZUR (2005): Overview of salinity problems in the world and FAO strategies to address
the problem, Proc. Intl. Sal. For., Riverside, California.MEENA, S.K., N.K. GUPTA, S. GUPTA, S.R. KHANDELWALANDE and V.D. SHASTRY (2003): Effect of sodium chloride on the
growth and gas exchange of young. ziziphus seedling root stocks. J. Hort. Sci. Biotech., 78:454-457.MUNIR, S., E. HUSSAIN, K.H. BHATTI, K. NAWAZ, K. HUSSAIN, R. RASHID and I. HUSSAIN (2013): Assessment of inter-cultivar
variations for salinity tolerance in winter radish (Raphanus sativus L.) using photosynthetic attributes aseffective selection criteria. World Appl. Sci. J., 21:384–388.
MUNNS, R. and M. TESTER (2008): Mechanisms of salinity tolerance. Annu. Rev. Plant Biol., 59:651-681.MUNNS, R., R.A. JAMES and A. LAUCHLI (2006): Approaches to increasing the salt tolerance of wheat and other cereals. J.
Exp. Bot., 57:1025-1043.NAHAR, K., M. HASANUZZAMAN, M.M. ALAM and M. FUJITA (2013): Exogenous glutathione-induced drought stress tolerance
in Vigna radiata seedlings through enhanced antioxidant defense and methylglyoxal system. Interdrought IVConference September 02-September 09, Perth, Australia.
NOUNJAN, N, P.T. NGHIA and P. THEERA KULPISUT (2012): Exogenous proline and trehalose promote recovery of riceseedlings from salt-stress and differentially modulate antioxidant enzymes and expression of related genes. JPlant Physiol., 169:596– 604.
PANDEY, H.C., M.J. BAIG and R.K. BHATT (2012): Effect of moisture stress on chlorophyll accumulation and nitrate reductaseactivity at vegetative and flowering stage in Avena species. Agric Sci Res., 2:111–118.

S .A..JAN et al: SALT TOLERANCE OF Brassica rapa SUB SPECIES 337
QUESADA, V., S. GARCIA-MARTINEZ, P. PIQUERAS, M.R. PONCE and J.L. MICOL (2002): Genetic architecture of NaCl tolerancein Arabidopsis. Plant Physiol., 130: 951–963.
SHAH, S.H., S. ALI, S.A. JAN, JALAL-U-UDDIN and G.M. ALI (2014): Piercing and incubation method of in plantatransformation producing stable transgenic plants by over expressing DREB1A gene in tomato (Solanumlycopersicum Mill.). Plant. Cell. Tiss. Org Cult., DOI 10.1007/s11240-014-0670-6.
SHAHBAZI, E., A. ARZANI and G SAEIDI (2011): Effects of NaCl treatments on seed germination and antioxidant activity ofcanola (Brassica napus l.) cultivars. Bang J. Bot., 41(1):67-73.
UMAR, S., I. DIVA, A. NASER, ANJUM, M. IQBAL, I. AHMAD and E. PEREIRA (2011): Potassium-induced alleviation of salinitystress in Brassica campestris L. Cent. Eur. J. Biol., 6(6):1054-1063.
SHINWARI, Z.K., K. NAKASHIMA, S. MIURA, K. YAMAGUCHI-SHINOZAKI and K.C. SHINOZAKI (1998): An Arabidopsis genefamily encoding DRE/CRT binding protein involved in low temperature responsive gene expression. Biochem.Biophys. Res. Comm., 250:161-170.
REYNOLDS, M.P., A.M. KAZI and M. SAWKINS (2005): Prospects for utilizing plant adaptive mechanisms to improve wheatand other crops in drought and salinity prone environments. Annals. Appl. Biol., 146:239-259.
ULFAT, M., H.R. ATHAR, M. ASHRAF, N.A. AKRAM and A. JAMIL (2007): Appraisal of physiological and biochemical selectioncriteria for evaluation of salt tolerance in canola (Brassica napus L.). Pak. J. Bot., 39:1593–1608.
VITAL, S.A., R.W. FOWLER, A. VIRGEN, D.R. GOSSETT, S.W. BANKS and J. RODRIGUEZ (2008): Opposing roles forsuperoxideand nitric oxide in the NaCl stress induce regulation of antioxidant enzyme activity in cotton callus tissue. Env.Exp. Bot., 62: 60-68.
ZAMANI, Z., M.T. NEZAMI, D. HABIBI and M.B. KHORSHIDI (2010): Effect of quantitative and qualitative performance of fourcanola cultivars (Brassica napus L.) to salinity conditions. Adv. Env Biol., 4(3):422-427.

338 GENETIKA, Vol. 48, No.1, 323-338, 2016
MORFO – BIOHEMIJSKA EVALUACIJA Brassica rapa sub – SPECIESA NATOLERANTNOST PREMA ZASLANJENOSTI
Sohail Ahmad JAN1*, Zabta Khan SHINWARI1, Malik Ashiq RABBANI2
1Odeljenje za biothenologiju, Quaid-i- Azam Univerzitet, Islamabad, Pakistan2Institut za biljne genetičke resurse, Nacionalni istraživački poljoprivredni centar, Islamabad,
Pakistan
IzvodVršena su ispitivanja efekta različitih koncentracija NaCl (0, 50,100 I 150 mmol) na
morfološke i biohemijske osobineon u cilju identifikacije tolerantnih genotipova na soli međusemđim, žutim iI uz dodavanje četiri nivoa NaCl toria B. rapa sub-speciesa. (0, 50,100 and 150mmol). Određivani su efekat zaslanjenosti na dužinu korena biljke, prinos sveže i suve supstancebiljke i korena, relativnog sadržaja vode (RWC), sadržaja prolina, i hlorofila a, b, I a+b hlorofila.Genotip 22861 (smeđi tip) je pokazao odlične morfološke i biohmijske osobine u svim uslovimastresa a slede Toria–Sathi, i Toria–A u poređenju sa kontrolnim TS-1genotipom. Genotip 26158(žut tip) je imao vrlo siromašne osobine i porast. Vrednosti RWC % i hlorofila a, b i sadržaj a+bhlorofila su bile smanjene nekoliko puta sa povećanjem koncentracije soli. Povećanje sadržajaprolina je pratilo povećanje koncentracij soli. Smeđi i Toria tipovi su pokazali maksimalnutolerantnost prema solima u ranoj fazi klijanja, kada se uporede sa žutim tipovima.. Ovaistraživanja će poslužiti kao model za brzi razvoj tolerantnih genotipova.
Primljeno 24. X. 2015.Odobreno 25. I. 2016.