Embed Size (px)
Citation preview

Proc. Nati. Acad. Sci. USAVol. 88, pp. 5993-5997, July 1991Immunology
Molecular mimicry by Trypanosoma cruzi: the Fl-160 epitope thatmimics mammalian nerve can be mapped to a 12-amino acid peptide
(autoimmunity/Chagas disease/antigenic mimicry/flagellar protein)
WESLEY C. VAN VOORHIS*, LYNN SCHLEKEWY*, AND HAI LE TRONGtDepartments of *Medicine and tBiochemistry, Mailstop SJ-10, University of Washington, Seattle, WA 98195
Communicated by Seymour J. Klebanoff, April 8, 1991 (receivedfor review January 25, 1991)
ABSTRACT Antigenic mimicry by Trypanosoma cruzi an-tigens that share epitopes with mammalian tissues may driveautoreactive B- or T-cell clones to expand and cause autoim-mune pathogenesis. We have been studying one of theseantigens, FI-160, a 160-kDa protein on the surface of T. cruzithat antigenically mimics a 48-kDa protein found in mamma-lian axonal and myenteric plexus cells. The FI-160 antigen hasbeen characterized by cloning and expression of T. cruzi DNAencoding Fl-160 in Escherichia coli. Recombinant peptidesfrom various regions of the Fl-160 gene were expressed andused to compete with affinity-purified polyclonal anti-FI-160antibodies binding to nerve. Recombinant 48-amino acid pep-tide (48X) derived from expression of base pairs 611-761 of theDNA sequence completely inhibited anti-FI-160 binding tonerve. Recombinant peptides expressed from DNA lacking thisregion did not inhibit anti-FI-160 binding to nerve. Threepeptides were synthesized to encompass the 48X peptide, a12-amino acid peptide and two 18-amino acid peptides. The12-amino acid peptide TPQRKTTEDRPQ (12X), correspond-ing to bases 615-651, completely inhibited the binding ofanti-Fl-160 antibodies to nerve at a concentration of 80 ng/ml(30 ,M). The two 18-residue peptides did not inhibit, even at10 ,ug/ml. Thus, the epitope of Fl-160 crossreactive withnervous tissue can be mapped to a 12-amino acid peptide. Somehumans with T. cruzi infection make antibodies to Fl-160 andto the 48X and 12X peptides. Control sera from uninfectedpersons did not react with these antigens. Anti-48X antibodies,immunoselected from human serum with 48X peptide, bind tohuman nerve axons. This demonstrates that some individualsinfected with T. cruzi make antibodies to the Fl-160 epitopecrossreactive with nervous tissues.
Trypanosoma cruzi, a protozoan hemoflagellate, causes Cha-gas disease (i.e., American trypanosomiasis). Chagas diseasehas an acute stage and a chronic stage, the latter of whichoccurs many years after initial infection. The acute stage isgenerally characterized by a mild influenza-like syndrome,but rarely myocarditis or meningoencephalitis occurs. Dur-ing the chronic stage, fatal cardiomyopathy and arrhythmiasoccur, and damage to the peripheral nervous system occurs,leading to megacolon, megaesophagus, and neuropathies (1).Nonetheless, "80% of chronically infected individuals re-main asymptomatic. The acute-phase pathology correlateswith damage by parasites in tissues. However, parasites arerare in tissues of chronically infected animals and humans,despite abundant inflammatory infiltrates (2). Inflammatorydestruction of the myenteric (Auerbach's) plexi of gut cor-relates with megagastrointestinal manifestations. Inflamma-tory destruction ofcardiac conductive tissues and myocardialcells correlates with arrhythmias and cardiomyopathy.
Increasing evidence suggests that chronic-phase pathologyis autoimmune in nature. Autoreactive antibodies and T cellsdirected to target organs have been described in experimentaland natural chronic T. cruzi infection (3-14). There are atleast two nonexclusive explanations for the generation ofautoimmunity in Chagas disease: (i) infection with the par-asite perturbs immunoregulation, leading to loss of tolerancefor self-antigens; (ii) immune recognition of T. cruzi antigensis crossreactive with selected mammalian antigens, leading toautoimmunity (molecular mimicry).Molecular mimicry by T. cruzi is evident by the existence
of crossreactive antibodies and T cells to T. cruzi andmammalian antigens. Antibodies to endocardium, vascula-ture, and interstitium of striated muscle (EVI) and Schwanncells ofperipheral nerve have been found in acute and chroniccases of Chagas disease (3, 4). Antibodies to EVI have beenfound to be directed to carbohydrate on mammalian laminin,apparently stimulated by crossreactive carbohydrate epi-topes present in T. cruzi (5, 6). Selected monoclonal anti-bodies generated against nervous tissue or T. cruzi arecrossreactive with parasite and nervous-tissue antigens (7-11). Van Voorhis and Eisen (12) reported cloning the gene fora 160-kDa flagellum-associated surface protein (Fl-160) oftrypomastigotes, the motile mammalian form of T. cruzi. Thisgene codes for a protein that molecularly mimics a 48-kDaprotein found exclusively in nervous tissues. The 48-kDaprotein is found in neuronal axons and in the myentericplexus of gut tissues. Inflammatory destruction in Chagasdisease often localizes in the myenteric plexus (leading tomegagastrointestinal disease) and occasionally in the periph-eral nervous system (1). Thus, Fl-160 is a candidate antigenfor directing autoimmunity via molecular mimicry of a pro-tein found in key targets ofchronic pathology. We report herethat the epitope of Fl-160 that is crossreactive with nervoustissues has been mapped to a 12-amino acid peptideA
MATERIALS AND METHODSProtein Expression and Antibody Techniques. The strain of
T. cruzi from which Fl-160 was derived was a subclone of theCL strain (15). The Fl-160 clone was derived from a randomlysheared genomic DNA library in Agtll (12) and subclonedinto the Bluescribe (Stratagene) plasmid EcoRI site. Thisclone was expressed in Escherichia coli DH5-1 as a fusionwith 8-galactosidase in the Bluescribe vector or with Schis-tosoma japonicum glutathione S-transferase (GST) in thepGEX1 vector (16). After induction with 0.1 mM isopropyl,3-D-thiogalactopyranoside for 2 hr during the logarithmicphase of growth, lysates were prepared by resuspendingbacteria at 10% (vol/vol) in 10 mM Tris (pH 7.4) with 1 mMphenylmethylsulfonyl fluoride and 25 ,g of lysozyme per ml,
Abbreviations: GPI, glycosyl-phosphatidylinositol; GST, glu-tathione S-transferase; ORF, open reading frame.*The sequence reported in this paper has been deposited in theGenBank data base (accession no. M65032).
5993
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.

5994 Immunology: Van Voorhis et al.
freezing and thawing twice, and sonicating. GST was ex-pressed from pGEX1 without insert (16). Anti-FI-160 anti-bodies were affinity-purified from rabbit immune sera asdescribed (12). Antibodies to the 48-amino acid peptide 48Xwere affinity-purified from serum from a person with Chagasdisease by removal of nonspecific antibodies by exposure toGST-Sepharose, followed by adsorption to 48X-GST fusionprotein linked to CNBr-activated Sepharose and elution ofantibodies with 3 M isothiocyanate (12). ELISA was byantibody capture on wells coated with antigen (10 pug/ml)(12). Sera from subjects with T. cruzi infection and endemiccontrols were obtained by Martin Cetron, in collaborationwith J. N. Paes and F. P. Basilio, Universidade de Ceara,Forteleza, Brazil. Subjects gave informed consent to partic-ipate in a protocol approved by the University ofWashingtonand Universidade de Ceara Human Subjects InstitutionalReview Boards.DNA Sequencing. Restriction endonuclease fragments of
the Fl-160 clone were subcloned into the Bluescribe plasmidand sequenced on both strands. Sites used for subcloningwere Rsa I (positions 161 and 401), HindIII (279), HincII(736), Ava I (740 and 1053), and Nae I (1513). Single-strandDNA was prepared by the phagemid procedure as describedby the manufacturer (Stratagene). DNA sequencing wasperformed by the dideoxynucleotide method using the Se-quenase kit (United States Biochemical) as described by themanufacturer.Polymerase Chain Reaction (PCR) Expression Cloning. Se-
quence information was used to generate oligonucleotideprimers for PCR amplification of subclones of Fl-160 plasmidDNA (17). The oligonucleotides (synthesized on an AppliedBiosystems DNA synthesizer) were as follows (identifyingnumber is where restriction cut occurs): (i) 5' sense-strandprimers with a BamHI site (71, GTTGGGATCCCACAG-CATCTCGC; 458, CGGGATCCTCATTGGCTCCAAC;611, GAGGCGGTTGGATCCACTCCGC; and 750,GGAGAGGATCCACACAAATCCCC) and (it) 3' antisense-strand with an EcoRI site (488, CTGAATTCTCCCGCTGT-TGGAGCC; 641, CGGAATTCAGTGGTTTTCCTCT-GCGG; 761, CACGGCAGTGTGGGGAATTCTG; 926,TTGAATTCCCCAGGCGTTGTGCGC; and 1098, CA-GAATTCCGCGGTGCCCCACAG). PCR was performed in100 1.d with 1 ng of Fl-160 plasmid, 0.1 gmol of each primer,and 0.2 tumol of each dNTP in 10 mM Tris, pH 8.3/50 mMKCl/1.5 mM MgCl2/0.01% gelatin. Thermocycling was car-ried out for 30 cycles of 1 min at 940C, 2 min at 550C, and 3min at 72TC. The cassettes-were cut with EcoRI and BamHI,ligated into BamHI- and EcoRI-cut pGEX2T vector (ref. 16;see Fig. 2) and transformed into E. coli DH5-1. Each cassettewas expressed as a fusion protein with S. japonicum GST,and lysates were prepared as described above and analyzedby SDS/10% PAGE (12). The 48X-GST fusion protein waspurified from bacterial lysates by adsorption to glutathione-agarose (Sigma) (16) and elution with 5 mM free glutathione;yield was 50 mg/liter of bacterial culture. The 48X-GSTprotein formed a single band at 33 kDa as shown byCoomassie staining after SDS/PAGE. The 48X-GST fusionprotein was dialyzed and the 48X peptide was cleaved fromGST by thrombin (Sigma) (16). The 48X peptide was purifiedfrom uncut and free GST by repeat passage through glu-tathione-agarose. The resultant peptide gave a single peak onreverse-phase high-pressure liquid chromatography (HPLC)that was different from the peaks seen in control runs withGST and 48X-GST. Amino acid analysis was consistent withthe predicted amino acid content of the 48X peptide.
Competition of Binding of Anti-FI-160 to Nerve. The bacteriallysate was used to compete with affinity-purified rabbit anti-Fl-160 antibodies (12) for binding to nerve. Anti-Fl-160 anti-bodies (5 tzg/ml) were mixed with an equal volume of bacteriallysate, incubated on ice for 1 hr, and exposed to a 5-gm frozen
section of uninfected mouse (C3H/He) sciatic nerve. Forpeptide competitions, dilutions ofpeptide were substituted forbacterial lysates. For human sciatic nerve, affinity-selectedhuman anti-48X antibodies were used at a final concentrationof 0.3 ug/ml. After 1 hr, slides were rinsed in phosphate-buffered saline and developed with biotinylated anti-rabbit oranti-human IgG antibodies, avidin-biotin-peroxidase conju-gates, and 3,3'-diaminobenzidine (Sigma) according to themanufacturer's protocol (Vector Laboratories). Human sci-atic nerve was obtained at autopsy from an uninfected personwith liver failure due to Wilson disease.
Analysis of Peptides. Peptides were synthesized with anApplied Biosystems peptide synthesizer, model 430. Aminoacid analysis was performed on a Waters Pico-Tag system(18). HPLC was performed on C18 columns, using trichloro-acetic acid/acetonitrile reverse-phase elution. Amino acidsequencing was performed using an Applied Biosystemsmodel 470 gas-phase sequenator equipped with an on-linemodel 120A phenylthiohydantoin analyzer.
RESULTSSequence Analysis. The open reading frame (ORF) of the
genomic clone of FI-160 was sequenced on both strands byrestriction-site subcloning (Fig. 1). The 160-kDa size ofFl-160 protein suggests that the 1104-base-pair (bp) ORF isonly about one-third of the total Fl-160 ORF. Consistent withthis, the Fl-160 DNA hybridizes with a 4-kilobase message on
GAATTCGATTATGAAGATGAGGATTACGAGGGCTATAAGAGCTTTTATGA 50E F D Y E D E D Y E G Y K S F Y DCAGGTCACAAAAACTGTTGAGCTCCCACAGCATCTCGCACTTTTACATTG 100R S Q K L L S S H S I S H F Y I G
GCGGGGACGGCAAGAGCAGCTCGGACAACATTCATGTGACGGTGAGCAAC 150G D G K S S S D N I H V T V S N
GTCCTGCTGTACAACCGCTTATTGCAAGATGATGAGCTCAACCCACTAAT 200V L L Y N R L L Q D D E L N P L MGAAGACGAAGGCGGCCGCTGCTCCGGAGGCGGAGGTGTCGGCCCCGGAAG 250K T K A A A A P E A E V S A P E G
GTGCGCCTCAAAACAGACATTTGGGACAAGCTTCTGAAAAAGACACCACA 300A P Q N R H L G Q A S E K D T T
CCATCACCACAAAATCAAGGATTATCACCGGAAAAATCAAAAAATGAAAA 350P S P Q N Q G L S P E K S K N E KGTATTCAGCAGGTTCCGGGCAGACAACTTCTGCTGATTTCACTGGCTTGT 400Y S A G S G Q T T S A D F T G L Y
ACACCTCAGCCGCCGAGGGGGAAGTGGAAGAAGTGGCATCCAGCAGTGTT 450T S A A E G E V E E V A S S S V
GGCTCGGCGTCCTCATTGGCTCCAACAGCGGGAGAAGGCAGTCCTCAAAA 500G S A S S L A P T A G E G S P Q KAGCACCTGAAACCGATGTGCTTGCCACTGAGGATCACTTTGAACGTGAAC 550A P E T D V L A T E D H F E R E QAGGAGCACTCGTCTCTCAGCGTGGTTCAGCCGATGACGGAGCAGGCTGAA 600
E H S S L S V V Q P M T E Q A EGAGGCGGTTGTCGCCACTCCGCAGAGGAAAACCACTGAAGATCGGCCGCA 650E A V V A T P Q R K T T E D R P QGCACAGCACTCTATCTGATGCTTCAGAGGACATGGAAGAGTCGTCGTCCC 700H S T L S D A S E D M E E S S S H
ATTCCGCCACGCTCACCAGCGACGAACAAACCGTTGACCCCGAGGAGAGG 750S A T L T S D E Q T V D P E E R
AAGGACACAAATCCCCACACTGCCGTGGGTGCGAGCAGCGGCTCCGACTC 800K D T N P H T A V G A S S G S D SATCCCACAGCACTGAGGTGGCGTCCGTGGACGGTGCCACAGACCCGGCAA 850S H S T E V A S V D G A T D P A T
CTGCTCAAGGCCATGATGAAGTGTTGGATGGCGGTGATGCCGCAAAGGAT 900A Q G H D E V L D G G D A A K D
TATTTGCGCACAACGCCTGGGGAAACCAAGATCCCATCGGAGTCCAACGC 950Y L R T T P G E T K I P S E S N ACACATCGCTCTCGGACCATGACATTTTGCTTGAGCACGGGCATTTGAGCG 1000T S L S D H D I L L E H G E L S D
ATTTGGCGTCCATGGCTCTAATTGGTGACAGCACCGTGCATGGGTGTGTG 1050L A S M A L I G D S T V H G C V
TCTCGGGTGTTGTTGCTGCTGCTTCTGGGGCTGTGGGGCACCGCGGCTCT 1100S R V L L L L L L G L W G T A A LCTGCTGAGGCGTGAGGAAGGAGTGTAATTGCGGCTTTTTGTGGAACAACA 1150C *
ACTGTCAGCTCCCCAGCACTACTCACTCAACTTTTTCAAGGATACTTCTT 1200TTTTTTTTTTTTTTTTGCGTCTCTTAAGTTTTTTTTCTTTTCATTTTTT 1250TCCTGTGTCCCCTTGTGGCTATGATTTTGCTTGTTTCTTTTTTTTCCTTT 1300TTTGTTTTTATTGTTTTTGTTGTTTTTTTTTTTTTTCTGTTTTCTTTTT 1350ATTGCTGGTGCTCTGCTGAAGTAATCAGATTCCTGTTTGTGTGTTTTTCC 1400CCAAAAAAACGGTCGCTTGCTAAACACCGTCACGAGTGACGTGCTTCCCC 1450
TGTTGCTTTTTTTGCCCACACGCGGTGCCA 1487
FIG. 1. Nucleic acid and amino acid sequence of FI-160 genomicclone.
Proc. Natl. Acad. Sci. USA 88 (1991)
Proc. Natl. Acad. Sci. USA 88 (1991) 5995
Northern analysis (12). The protein predicted from the ORFhas a hydrophobic carboxyl terminus with a sequence con-sistent with glycolipid anchoring (19, 20). The 16 carboxyl-terminal amino acids are highly hydrophobic with no identi-fiable cytoplasmic domain (polar, charged amino acids). Thesequence Gly-Asp-Ser, predicted 27-25 amino acids from thecarboxyl terminus, is the most likely glycosyl-phosphatidyl-inositol (GPI) anchoring site, though other possibilities exist.Mapping the Crossreactive Epitope by PCR Subcloning.
Anti-FI-160 antibodies bind to mouse nerve axons, demon-strating a crossreactive epitope of FI-160 and nerve (12).Lysates of bacteria expressing the 1104-bp ORF of the FI-160clone completely inhibited the binding of anti-FI-160 anti-bodies to nerve. Complete inhibition occurred when Fl-160was expressed as a fusion protein with either GST or /3-ga-lactosidase. Purified Fl-160-GST fusion protein also com-pletely inhibited binding of anti-FI-160 to nerve. Controlbacterial lysates with GST expression alone gave no inhibi-tion of anti-FI-160 binding to nerve (data not shown). Thus,competition with recombinant protein could be used todemonstrate which region of FI-160 encodes the crossreac-tive epitope with nervous tissue.PCR subcloning of segments of the Fl-160 DNA ORF was
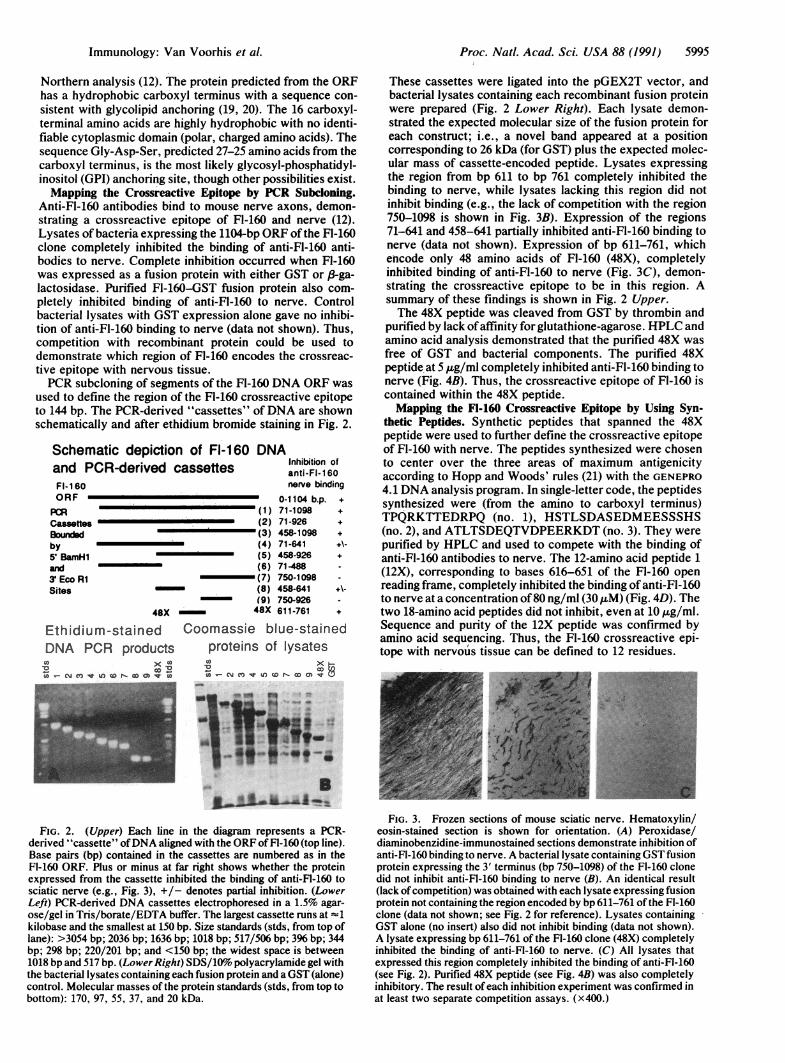
used to define the region of the FI-160 crossreactive epitopeto 144 bp. The PCR-derived "cassettes" ofDNA are shownschematically and after ethidium bromide staining in Fig. 2.
Schematic depiction of Fl-160 DNAand PCR-derived cassettes anti-FI-160FI- 1 60 nerve bindingORF 0-1104 b.p. +
PCRz (1) 71-1098 +Cassettes (2) 71-926 +Bourm~ed (3) 458-1098 +by (4) 71-641 +\-5' BamrH1 (5) 458-926 +and (6) 71-4883EcoR1 (7) 750-1098Sites (8) 458-641 +\
(9) 750-92648X 48X 611-761 +
Ethidium-stainedDNA PCR productsu) x (a
U)co
- 0u)_CMCX st ) @ rs X o stGo
_wv m ~_ _ .
Coomassie blue-stainedproteins of lysates0
OJ C 'tm 0 CD r-. w n14
These cassettes were ligated into the pGEX2T vector, andbacterial lysates containing each recombinant fusion proteinwere prepared (Fig. 2 Lower Right). Each lysate demon-strated the expected molecular size of the fusion protein foreach construct; i.e., a novel band appeared at a positioncorresponding to 26 kDa (for GST) plus the expected molec-ular mass of cassette-encoded peptide. Lysates expressingthe region from bp 611 to bp 761 completely inhibited thebinding to nerve, while lysates lacking this region did notinhibit binding (e.g., the lack of competition with the region750-1098 is shown in Fig. 3B). Expression of the regions71-641 and 458-641 partially inhibited anti-FI-160 binding tonerve (data not shown). Expression of bp 611-761, whichencode only 48 amino acids of Fl-160 (48X), completelyinhibited binding of anti-FI-160 to nerve (Fig. 3C), demon-strating the crossreactive epitope to be in this region. Asummary of these findings is shown in Fig. 2 Upper.The 48X peptide was cleaved from GST by thrombin and
purified by lack of affinity for glutathione-agarose. HPLC andamino acid analysis demonstrated that the purified 48X wasfree of GST and bacterial components. The purified 48Xpeptide at 5 ,ug/ml completely inhibited anti-FI-160 binding tonerve (Fig. 4B). Thus, the crossreactive epitope of FI-160 iscontained within the 48X peptide.Mapping the Fl-160 Crossreactive Epitope by Using Syn-
thetic Peptides. Synthetic peptides that spanned the 48Xpeptide were used to further define the crossreactive epitopeof Fl-160 with nerve. The peptides synthesized were chosento center over the three areas of maximum antigenicityaccording to Hopp and Woods' rules (21) with the GENEPRO4.1 DNA analysis program. In single-letter code, the peptidessynthesized were (from the amino to carboxyl terminus)TPQRKTTEDRPQ (no. 1), HSTLSDASEDMEESSSHS(no. 2), and ATLTSDEQTVDPEERKDT (no. 3). They werepurified by HPLC and used to compete with the binding ofanti-FI-160 antibodies to nerve. The 12-amino acid peptide 1(12X), corresponding to bases 616-651 of the Fl-160 openreading frame, completely inhibited the binding of anti-FI-160to nerve at a concentration of 80 ng/ml (30 ,tM) (Fig. 4D). Thetwo 18-amino acid peptides did not inhibit, even at 10 ,g/ml.Sequence and purity of the 12X peptide was confirmed byamino acid sequencing. Thus, the Fl-160 crossreactive epi-tope with nervous tissue can be defined to 12 residues.
a
L%|
FIG. 2. (Upper) Each line in the diagram represents a PCR-derived "cassette" ofDNA aligned with the ORF of Fl-160 (top line).Base pairs (bp) contained in the cassettes are numbered as in theFl-160 ORF. Plus or minus at far right shows whether the proteinexpressed from the cassette inhibited the binding of anti-FI-160 tosciatic nerve (e.g., Fig. 3), +/- denotes partial inhibition. (LowerLeft) PCR-derived DNA cassettes electrophoresed in a 1.5% agar-ose/gel in Tris/borate/EDTA buffer. The largest cassette runs at -1
kilobase and the smallest at 150 bp. Size standards (stds, from top oflane): >3054 bp; 2036 bp; 1636 bp; 1018 bp; 517/506 bp; 396 bp; 344bp; 298 bp; 220/201 bp; and <150 bp; the widest space is between1018 bp and 517 bp. (Lower Right) SDS/10%o polyacrylamide gel withthe bacterial lysates containing each fusion protein and aGST (alone)control. Molecular masses of the protein standards (stds, from top tobottom): 170, 97, 55, 37, and 20 kDa.
FIG. 3. Frozen sections of mouse sciatic nerve. Hematoxylin/eosin-stained section is shown for orientation. (A) Peroxidase/diaminobenzidine-immunostained sections demonstrate inhibition ofanti-FI-160 binding to nerve. A bacterial lysate containing GST fusionprotein expressing the 3' terminus (bp 750-1098) of the FI-160 clonedid not inhibit anti-FI-160 binding to nerve (B). An identical result(lack ofcompetition) was obtained with each lysate expressing fusionprotein not containing the region encoded by bp 611-761 ofthe FI-160clone (data not shown; see Fig. 2 for reference). Lysates containingGST alone (no insert) also did not inhibit binding (data not shown).A lysate expressing bp 611-761 of the Fl-160 clone (48X) completelyinhibited the binding of anti-FI-160 to nerve. (C) All lysates thatexpressed this region completely inhibited the binding of anti-FI-160(see Fig. 2). Purified 48X peptide (see Fig. 4B) was also completelyinhibitory. The result ofeach inhibition experiment was confirmed inat least two separate competition assays. (x400.)
Immunology: Van Voorhis et al.

5996 Immunology: Van Voorhis et al.
Serologic Studies of Humans Infected with T. cruzi. Serafrom persons infected with T. cruzi were screened for ser-oreactivity to Fl-160. Forty-four percent of persons infectedwith T. cruzi (serum bank from northeastern Brazil) werefound to have detectable antibodies to Fl-160 in the antibody-capture ELISA, as defined by absorbances > 2 SD over themean of the uninfected controls (M. C. Cetron, R. Hoff, S.Kahn, H. Eisen, and W.C.V.V., unpublished work). Twoinfected persons' sera, chosen to be reactive with Fl-160,have been examined in more detail (Table 1). Absorbances inthe ELISA with Fl-160, 48X peptide, and 12X peptide were>2 SD above the mean of the absorbances obtained from thethree control sera. This was true at serum dilutions of 1:200(Table 1) as well as 1:100 and 1:400 (data not shown). Thisdemonstrates that humans infected with T. cruzi developantibodies to the crossreactive epitope of Fl-160.
Anti-48X antibodies were affinity-purified from serum of aperson with Chagas disease (subject 2, Table 1) by removalof nonspecific antibodies by exposure to GST-Sepharose,followed by adsorption to 48X-GST-Sepharose and elution ofantibodies. Nonspecific antibodies, eluted from GST-Sepharose, did not bind human nerve (Fig. 5). Anti-48Xantibodies bound human sciatic nerve (Fig. 5). This bindingwas inhibited by 12X peptide (Fig. 5) but not by the two18-residue peptides (data not shown). Similar binding ofanti-48X to mouse nerve axons was observed, and thisbinding was inhibited with 12X peptide (data not shown).
DISCUSSIONFI-160 is an antigen expressed by T. cruzi trypomastigotes thatmimics a protein of mammalian nervous tissue (12). Thisnervous-tissue protein is found in myenteric plexus cells,which are destroyed by inflammatory infiltrates in chronicChagas disease, leading to megacolon and megaesophagus.Thus, Fl-160 is a possible stimulus for the autoimmune re-sponse hypothesized to cause pathology in chronic Chagasdisease. We report here the sequence for the portion of Fl-160that has been cloned, and we have mapped the epitope that iscrossreactive with nervous tissues to a 12-amino acid region ofFl-160. Knowledge of the molecular nature of crossreactiveepitopes of T. cruzi and mammalian tissues will aid in ourunderstanding the generation of autoimmunity in chronic T.cruzi infection and may help us to specifically interrupt theautoimmune pathophysiology of chronic Chagas disease.The current genomic clone includes 1104 bp at the 3' end
of the F1-160 ORF. The sequence encodes regions consistentwith the protein being anchored by GPI. T. cruzi has a
-46,% 47A40 f ,
D
FIG. 4. Frozen sections of mouse sciatic nerve were immuno-peroxidase-stained with anti-FI-160 (5 .ug/ml) that had been prein-cubated with peptides. Staining with anti-FI-160 without competingpeptide is shown in A. The recombinant 48X peptide (5 ,g/ml),purified from GST, completely inhibited staining (B). Neither theamino-terminal 18-residue peptide 3 (10 ,ug/ml) (C) nor the central18-residue peptide 2 (data not shown) inhibited anti-FI-160 binding tonerve. The carboxyl-terminal peptide 1 (12X) completely inhibitedthe binding of anti-Fl-160 to nerve (peptide concentration, 80 ng/ml).(D) The result of each inhibition experiment was confirmed in threeseparate competition assays over a wide range of peptide concen-trations (80 ng/ml to 10 jig/ml). (x630.)
'~~~~~~~~~~l I "'e.YI ..
Table 1. Human sera react with the Fl-160 crossreactive epitopeAbsorbance units x 103
48X 12XSubject(s) Fl-160 peptide peptide
Indeterminate 476 326 429subject 1
Cardiomyopathy 801 710 709subject 2
Negative control 205 ± 70 198 ± 57 168 ± 85(n = 3)Antibody-capture ELISA was performed using 1:200 dilutions of
sera from two subjects infected with T. cruzi and three negativecontrol sera. The absorbances are shown in milliunits, with the meanand SD shown for the negative controls. The negative controlsincluded two individuals from the endemic area in Brazil (one withidiopathic congestive heart failure, one with multiple myeloma) andone normal U.S. citizen.
phospholipase C, which cleaves molecules linked by GPIfrom the surface of the parasite (22). Furthermore, exchangeof GPI-linked molecules from the surface of T. cruzi to otherlipid bilayers can occur, as has been shown with transfer ofGPI-linked variant surface glycoproteins from Trypanosomabrucei to erythrocytes (23). This would explain how theFl-160 protein is shed from the parasite and occurs in thecytoplasm and on the surface of parasitized and nonparasit-ized cells (12). The major surface glycoprotein of T. cruzi, the85-kDa surface antigen SA85, also is most likely anchored byGPI and is shed into cytoplasm and fluid phase (S. Kahn andH. Eisen, personal communication). That these proteins arefound on uninfected cells may extend the immune responseagainst T. cruzi to destroy uninfected cells.The crossreactive protein of nervous tissue has been found
to be 48 kDa by Western immunoblotting (12). It is found inaxons of peripheral nerve, cell bodies of myenteric plexuscells, and filamentous structures in brain. No significantsequence homology of Fl-160 or the 12X peptide to anyknown proteins was found by searching GenBank (Release65) or Protein Identification Resource (Release 25) databases. This, combined with the molecular mass and distri-bution of the protein, suggests that this protein has not beendescribed. Since the Fl-160 antigen is large, many of theanti-Fl-160 antibodies probably bind to epitopes other thanthe crossreactive epitope. The affinity-purified anti-48X an-tibodies will be useful to immunoselect cDNAs expressingthis neuronal protein for further characterization.Human subjects infected with T. cruzi make antibodies that
recognize the Fl-160 epitope crossreactive with nerve anti-gen. This was shown by ELISA and immunoselection tech-niques. Anti-FI-160 antibodies bind to human sciatic nerve
b WS~~~!P-- ._X
FIG. 5 .Frozen sections of human sciatic nerve were immuno-peroxidase stained with immunoselected human anti-48X antibodies(0.3 Ag/ml). Nonspecific antibodies selected on GST-Sepharose didnot react with nerve axons (A). Anti-48X antibodies stained nerveaxons (B) in a similar pattern to that obtained with rabbit anti-FI-160antibodies. Binding of anti-48X was blocked by peptide 12X (400ng/ml) (C) but not the two 18-residue peptides (tested up to 10 A~g/ml;data not shown) (x630.)
Proc. Natl. Acad. Sci. USA 88 (1991)

Proc. Natl. Acad. Sci. USA 88 (1991) 5997
(12). Antibodies selected from serum of a person with Chagasdisease with the crossreactive epitope of Fl-160 react withhuman sciatic nerve. This, combined with the ability of 12Xpeptide to block this binding and binding to mouse sciaticnerve, suggests that the crossreactive nervous-tissue epitopeis similar in humans and mice. Forty-four percent of personsinfected with T. cruzi react with Fl-160 recombinant antigen(M. C. Cetron, R. Hoff, S. Kahn, H. Eisen, and W.C.V.V.,unpublished work). Though the ability ofhuman sera to reactwith FI-160 did correlate with T. cruzi infection, it did notcorrelate with clinical disease. Screening for reactivity to the12X synthetic peptide, representing the crossreactive epitopeof FI-160, might reveal a better correlation of antibodyreactivity with clinical disease.
It is possible that clinical disease correlates better with theT-cell response to molecular mimicry antigens. In the mousemodel, transfer of T cells derived from chronically infectedmice to naive mice led to cardiac damage and nervous tissuedamage (13, 14). T cells from chronically infected micerecognize Fl-160 recombinant ag (W.C.V.V. and H. Eisen,unpublished data). The Fl-160 expression subclones used inthis report to map the serologic crossreactive epitope shouldalso prove useful to map the T-cell crossreactive epitope(s).W.C.V.V. is a Pew Scholar in the Biomedical Sciences and a
Searle Scholar. This work was supported by the Pew CharitableTrust and the Searle Foundation/Chicago Community Trust. H.L.T.was supported by National Institutes of Health Grant GM15731 toKenneth A. Walsh. We gratefully acknowledge Drs. Harry Char-bonneau and Kenneth A. Walsh, University of Washington Bio-chemistry Department, for helpful discussions and use ofequipment;Dr. Harvey Eisen, Fred Hutchinson Cancer Research Center, inwhose laboratory the sequencing of Fl-160 was initiated; the excel-lent technical assistance of Molly Weaver; Dr. Martin 'Cetron,University of Washington, who provided the sera from persons fromBrazil, and the collaborators who referred the subjects, Dr. J. N.Paes and Dr. F. P. Basilio, Universidade de Ceara, Brazil; and Dr.N. Samaras and G. Mitchell, Walter and Eliza Hall Institute andAmgen Corp., who provided the pGEX vectors.
1. Koeberle, F. (1968) Adv. Parasitol. 6, 63-116.2. Tafuri, W. L. (1979) in Biology ofKinetoplastia, eds. Lumsden,
W. H. R. & Evans, D. A. (Academic, London), Vol. 2, p. 547.
3. Cossio, P. M., Diez, C., Szarfman, A., Kreutzer, E., Candiolo,B. & Arana, R. M. (1974) Circulation 49, 13-21.
4. Khoury, E. L., Ritacco, V., Cossio, P. M., Laguens, R. P.,Szarfman, A., Diez, C. & Arana, R. M. (1979) Clin. Exp.Immunol. 36, 8-15.
5. Szarfman, A., Terranova, V. P., Rennard, S. I., Foidart,J. M., Lima, M. D. F., Scheinman, J. I. & Martin, G. B. (1982)J. Exp. Med. 155, 1161-1171.
6. Towbin, H., Rosenfelder, G., Wieslander, J., Avila, J. L.,Rojas, M., Szarfman, A., Esser, K., Nowack, H. & Timpl, R.(1987) J. Exp. Med. 166, 419-432.
7. Wood, J. N., Hudson, L., Jessell, T. M. & Yamamoto, M.(1982) Nature (London) 296, 34-38.
8. Snary, D., Flint, J. E., Wood, J. N., Scott, M. T., Chapman,M. D., Dodd, J., Jessell, T. M. & Miles, M. A. (1983) Clin.Exp. Immunol. 54, 617-621.
9. Petry, K., Voison, P. & Baltz, T. (1987) Acta Trop. (Basel) 44,381-386.
10. Petry, K., Voison, P., Baltz, T. & Labouesse, J. (1987) J.Neuroimmunol. 16, 237-252.
11. Petry, K., Nudelman, E., Eisen, H. & Hakimori, S. (1988) Mol.Biochem. Parasitol. 30, 113-121.
12. Van Voorhis, W. C. & Eisen, H. (1989) J. Exp. Med. 169,641-652.
13. Laguens, R. P., Meckert, P. C., Chambo, G. & Gelpi, R. J.(1981) Medicina (Buenos Aires) 41, 40-43.
14. Hontebeyrie-Joskowicz, M., Said, G., Milon, G., Marchal, G.& Eisen, H. (1987) Eur. J. Immunol. 17, 1027-1033.
15. Plata, F., Pons, F. G. & Eisen, H. (1984) Eur. J. Immunol. 14,392-399.
16. Smith, D. B. & Johnson, K. S. (1988) Gene 67, 31-40.17. Scharf, S. J. (1990) in PCR Protocols: A Guide to Methods and
Applications, eds. Innis, M. A., Gelfand, D. H., Sninsky, J. J.& White, T. J. (Academic, New York), pp. 84-91.
18. Bidlingmeyer, B. A., Cohen, S. A. & Tarvin, T. L. (1984) J.Chromatogr. 336, 93-104.
19. Ferguson, M. A. J. & Williams, A. F. (1988) Annu. Rev. Bio-chem. 57, 285-320.
20. Cross, G. A. M. (1990) Annu. Rev. Cell Biol. 6, 1-39.21. Hopp, T. P. & Woods, K. R. (1981) Proc. Natl. Acad. Sci.
USA 78, 3824-3828.22. Andrews, N. W., Robbins, E. S., Ley, V., Hong, K. S. &
Nussenzweig, V. (1988) J. Exp. Med. 167, 300-314.23. Riflin, M. R. & Landsberger, F. R. (1990) Proc. Natl. Acad.
Sci. USA 87, 801-805.
Immunology: Van Voorhis et al.





























