Embed Size (px)
Citation preview

ORIGINAL ARTICLE
Molecular cloning and biochemical characterization of threeConcord grape (Vitis labrusca) flavonol 7-O-glucosyltransferases
Dawn Hall • Kyung Hee Kim • Vincenzo De Luca
Received: 18 May 2011 / Accepted: 24 June 2011 / Published online: 14 July 2011
� Springer-Verlag 2011
Abstract Grapes berries produce and accumulate many
reactive secondary metabolites, and encounter a wide range
of pathogen- and human-derived xenobiotic compounds.
The enzymatic glucosylation of these metabolites changes
their reactivity, stability and subcellular location. Two ESTs
with more than 90% nucleotide sequence identity to three
full-length glucosyltransferases are expressed in several
grape tissues. The full-length clones have more than 60%
amino acid sequence similarity to previously characterized
flavonoid 7-O-glucosyltransferases, catechin O-glucos-
yltransferases and anthocyanin 5-O-glucosyltransferases.
In vitro, these enzymes glucosylate flavonols and the
xenobiotic 2,4,5-trichlorophenol (TCP). Kinetic analysis
indicates that TCP is the preferred substrate for these
enzymes, while expression analysis reveals variable tran-
scription of these genes in grape leaves, flowers and berry
tissues. The in vivo role of these Vitis labrusca glucos-
yltransferases is discussed.
Keywords Concord grape � Detoxification mechanism �Flavonol � Glucosyltransferase � Xenobiotic
Abbreviations
AF After flowering
DFCI Dana Farber Cancer Institute
GTs Glucosyltransferases
PMSF Phenylmethyl sulfonyl fluoride
PSPG Plant secondary product
glucosyltransferases
PVP-10 or PVPP Polyvinylpyrrolidone
Q3G Quercetin 3-O-glucoside
TCP Trichlorophenol
UDPG Uridine 50-diphosphoglucose
VLOGT Vitis labrusca O-glucosyltransferase
VLRSGT Vitis labrusca resveratrol
glucosyltransferase
Introduction
Plants encounter a wide range of external stimuli and envi-
ronmental conditions that trigger the production of diverse
secondary metabolites. These compounds accumulate within
the vacuole, in specialized cell types, or outside the plant in
cell walls, on the leaf or in the rhizosphere surrounding plant
roots. Enzymatic modification by hydroxylation, methylation,
acylation and glycosylation changes the physiological and
biochemical properties of these molecules and alters their role
and location within the plant.
Grapes have been cultivated for thousands of years for
the production of wine, and they remain culturally, eco-
nomically and agronomically important in many areas
around the world. The grape berry can be divided into the
seed, mesocarp (flesh), and exocarp (skin) tissues that
differ in their enzymatic and metabolite profiles throughout
development. A key stage of grape berry development,
Electronic supplementary material The online version of thisarticle (doi:10.1007/s00425-011-1474-0) contains supplementarymaterial, which is available to authorized users.
D. Hall � K. H. Kim � V. De Luca (&)
Department of Biological Sciences, Brock University,
500 Glenridge Ave, St. Catharines, ON L2S 3A1, Canada
e-mail: [email protected]
Present Address:D. Hall
Michael Smith Laboratories, The University of British
Columbia, #301, 2185 East Mall, Vancouver, BC V6T 1Z4,
Canada
123
Planta (2011) 234:1201–1214
DOI 10.1007/s00425-011-1474-0

veraison, is characterized by berry softening, the rapid accu-
mulation of total soluble solids (�Brix) and the accumulation
of anthocyanins in the exocarp to produce red/blue colors.
Many of the berry-derived compounds contribute to the
organoleptic characteristics (color, flavour, astringency) of
wine (Polaskova et al. 2008), and their regular consumption is
associated with numerous human health benefits (Leifert and
Abeywardena 2008; Huntley 2007).
Glucosylated metabolites are detected in many grape-
vine tissues. Grape berries accumulate the 3-O-mono-
glucosides and the 3,5-O-diglucosides of the colorful
anthocyanins (Munoz-Espada et al. 2004; Wu and Prior
2005), the 3-O- and 7-O-glucosides of the UV-protectant
flavonols (Castillo-Munoz et al. 2007, 2009) and the 3-O/
40-O-glucosides of health beneficial stilbenes (Ali and
Strommer 2003). Glucosylated anthocyanins, flavonols
and stilbenes have also been identified in grape leaves and
flowers (Langcake and Pryce 1977; Moore and Giannasi
1994; Schneider et al. 2008).
Glucosyltransferases (GTs) catalyze the transfer of
glucose from uridine 50- diphosphoglucose (UDPG) to
acceptor molecules that include plant-derived and xenobi-
otic metabolites (Vogt and Jones 2000). In plants, GTs
exist as large, multigene families (Bowles 2002) and the
Arabidopsis genome currently has 121 Group 1 plant sec-
ondary product glucosyltransferases (PSPG; CaZy—
http://www.cazy.org/) which can be subdivided into 12
classes (A–N) (Lim et al. 2003). Although more than 150
GTs have been cloned from a number of plant species, only
4 GTs have been functionally identified in grape. These
Vitis GTs catalyze the glucosylation of the 3-O- position of
anthocyanins and flavonols (Ford et al. 1998), the 5-O-
positions of anthocyanin-3-O-glucosides (Janvary et al.
2009), and produce either the glucose esters of hydroxy-
cinnamic acids or stilbene diglucosides (Hall and De Luca
2007), however, GTs which glucosylate the 7-O-position of
flavonols have not been identified.
The present study details the homology-based molecular
cloning and biochemical characterization of three full-
length members of the 7GT-family from Vitis labrusca cv.
Concord that O-glucosylate flavonols, and a xenobiotic in
vitro. Substrate specificity, kinetic analysis, RT-PCR gene
expression studies, and in vivo enzyme activity profiles
suggest putative roles for these GTs in the modification of
xenobiotics and flavonols.
Materials and methods
Plant material
Grape berries (Vitis labrusca cv. Concord) were harvested
weekly from June until October, 2003–2006 at the 3rd St.
site of the G & H Wiley vineyard (St. Catharines, ON,
Canada). Vitis vinifera cv. Pinot Noir and Vitis vinifera cv.
Pinot Noir Droit were harvested from the Chateau des
Charmes Vineyard (Niagara on the Lake, ON, Canada) in
September of 2004. All stages of grape berry maturity are
reported as weeks after flowering (AF), and physiological
parameters of grape berry development were determined as
reported previously (Hall and De Luca 2007). Concord pre-
bloom flower buds were harvested June 12, 2006, grape
flowers were harvested June 16, 2006 and grapevine leaves
and stems were harvested June 21, 2006. All plant material
was stored at -80�C for future use. To separate grape berry
exocarp from mesocarp tissue, berries were briefly thawed;
the exocarp was removed by hand, scraped to remove
residual mesocarp tissue, and was quick frozen in liquid
nitrogen until future use.
Chemicals
All chemicals were purchased from Sigma (http://
www.sigmaaldrich.com) (Oakville, ON, Canada) or Indo-
fine Chemical (http://www.indofinechemical.com) (Hills-
borough, NJ, USA) and were prepared as reported previously
(Hall and De Luca 2007). Kaempferol, quercetin, and isorh-
amnetin 3-O-glucosides and quercetin 7-O-glucoside were
prepared enzymatically using the recombinant UDP-glucose:
flavonoid 3-O-glucosyltransferase (VL3GT) or the recombi-
nant UDP-glucose: resveratrol/hydroxycinnamic acid gluco-
syltransferase (VLRSGT), respectively (Hall and De Luca
2007).
Crude protein extraction and profiling of GT enzyme
activity throughout development
Vitis labrusca cv. Concord pre-bloom flower bud (1 g), leaf
(0.5 g), flower (1 g), week 1 whole berry (1 g), weeks 3, 6,
8, 10, 12, 14, and 16 AF and post-veraison (week 14 AF)
Vitis vinifera cv. Pinot Noir and cv. Pinot Noir Droit
exocarp (1 g) and mesocarp (2 g) tissue was ground to a
fine powder in a mortar and pestle with liquid nitrogen and
extracted in 4 ml of grape extraction buffer [500 mM Tris–
HCl, pH 8.0, 0.1% b-mercaptoethanol, 1% PVP-10 (poly-
vinylpyrrolidone), 5 mM sodium metabisulfite (Na2S2O5),
10% glycerol (v/v), 1 mM phenylmethyl sulfonyl fluoride
(PMSF), and 10% insoluble polyvinylpyrrolidone (PVPP)].
The extracts were centrifuged at 21,000g for 10 min at
4�C, and the supernatant was desalted on a PD-10 Sepha-
dex G-25 column (http://www.gehealthcare.com/caen), as
per the manufacturer’s instructions. For each enzyme
assay, 100 ll of protein was used with 100 lM quercetin
or cyanidin 3-O-glucoside and 9 mM UDPG in a final
reaction volume of 125 ll. The assays were incubated for
90 min at 30�C, stopped with 100% methanol, and
1202 Planta (2011) 234:1201–1214
123

processed prior to HPLC analysis as described below. All
specific activities are reported as picomoles of glucoside
produced in 1 s by 1 mg of total protein.
High performance liquid chromatography (HPLC)
analysis of enzyme assays
Enzyme assays were performed and analyzed by HPLC as
described previously (Hall and De Luca 2007). Addition-
ally, the O-glucosides of luteolin (350 nm), eriodictyol
(290 nm), myricetin (370 nm), isorhamnetin (370 nm),
dihydroquercetin (290 nm) and 3,6,20,30-tetrahydroxyflav-
one (320 nm) were monitored using a 2996 photodiode
array detector (Waters, Milford, MA, USA; http://www.
waters.com).
Flavonoid and anthocyanin analysis of mature grape
exocarp and mesocarp
The exocarp and mesocarp (with seeds removed) of mature
week 16 AF Concord grapes were harvested, dried, defatted
with 100% hexane and dried to produce 1 g dry weight of 3
biological replicates. Individual samples were extracted
three times in 10 ml of 15% acetic acid in methanol and the
combined supernatants from each extraction were centri-
fuged (7,250g, 4�C, 10 min) and the supernatants were
concentrated to dryness using a rotary evaporator at 40�C.
The acidic methanolic extracts were dissolved in 10 and
30 ml of water for mesocarp and exocarp fractions,
respectively, and then partitioned three times with 20 and
50 ml of ethylacetate for mesocarp and exocarp fractions,
respectively. After drying the combined organic fractions
using a rotary evaporator at 40�C, samples were reconsti-
tuted in 1 and 6 ml of 50% methanol for mesocarp and
exocarp fractions, respectively. The injection volume for
UPLC–MS analysis was 5 and 3 ll for mesocarp and exo-
carp fractions, respectively and analyses were performed
using an Aquity UPLC BEH C18 column with a particle size
of 1.7 lm and column dimensions of 1.0 9 50 mm and the
following parameters (Fig. S1, Table S1).
RNA extraction
RNA was extracted from grape berry exocarp and meso-
carp tissue as described previously (Hall and De Luca
2007), except that 1 ll of linear acrylamide (5 lg/ll) was
added to the RNA as a co-precipitant and the extracted
RNA was resuspended on ice. RNA (0.5–1 lg) was reverse
transcribed using either Superscript III reverse transcriptase
(Invitrogen) or the Takara RNA PCR kit version 3.0
(Fisher Scientific, http://www.fishersci.ca/) as per the
manufacturer’s instructions, and was diluted fivefold with
water. For RACE-ready cDNA, 5 lg of Vitis labrusca
week 12 AF berry exocarp RNA was extracted as above,
and prepared using the GeneRacer Kit (Invitrogen) as per
the manufacturer’s instructions.
Cloning of putative glucosyltransferase (GT) genes
from Vitis labrusca
At the time of cloning glucosyltransferases, the grape gen-
ome had yet to be released and 50 RACE was used to obtain
full length of glucosyltransferase sequences. A BLAST
search of Dana-Farber Cancer Institute (DFCI) Vitis vinifera
grape gene index EST database (http://compbio.dfci.
harvard.edu/tgi/cgi-bin/tgi/gimain.pl?gudb=grape) with the
(PSPG) box (WAPQVEVLAHPAVGCFVTHCGWNS-
TLESISAGVPMVAWPFFADQ) (Vogt and Jones 2000;
Jones and Vogt 2001) identified 141 expressed sequence tags
(ESTs) with sequence identity (E value \10-5) to GTs.
Three ESTs (TC75303, TC91700, NP9526770) with at least
60% sequence amino acid similarity to previously charac-
terized flavonoid GTs were identified, and their corre-
sponding full-length clones were obtained through two
rounds of 50 RACE.
Primers were designed to amplify the corresponding
full-length genes from Vitis labrusca week 12 AF exocarp-
enriched cDNA. VLOGT1 (EF533704) and VLOGT2
(EF533705) corresponding to the Vitis vinifera ESTs
NP9526770 and TC91700, respectively, were amplified
using FP3 (50 ATGGACAAACATCACTTCCTCTTAC 30)and RP6 (50 CTCAATGAGTCCTGACTTCTAAGC 30),and VLOGT3 (EF533606) corresponding to the EST
TC75303 was amplified with the primers followed by FP4
(50 GATCAGCACCCGGAAACGGAGAA 30) and RP14
(50 CTAAAAAATACTGCCTTGTCCCA 30). The prod-
ucts of these PCR reactions were cloned into the pGEM-T
easy TA-vector (Promega, http://www.promega.com) and
sequence verified.
Expression of recombinant VLOGT1, VLOGT2
and VLOGT3
Full-length VLOGT1, VLOGT2 and VLOGT3 were sub-
cloned into the pGEX-4T-1 GST-fusion protein expression
vector (GE Healthcare) and were transformed into Esche-
richia coli (E. coli) cells for recombinant protein expression.
Several E. coli strains, different induction conditions [(0.1 or
1 mM IPTG) (Bioshop, http://www.bioshopcanada.com)],
three growth temperatures following induction (16–18, 25,
or 37�C), and concurrent expression of the groES-groEL
chaperone proteins (to assist in GT folding) were tested
to obtain maximal amounts of soluble protein. For each
condition, soluble protein was extracted, assayed for GT
activity and 5–10 lg of protein was subjected to SDS-PAGE
Planta (2011) 234:1201–1214 1203
123

analysis to determine the optimal growth conditions for these
enzymes.
Expression in the pGEX-4T-1 N-terminal GST-fusion
vector in DE3 E. coli cells expressing chaperone protein,
induced with 0.1 mM IPTG and grown for 20 h at 16–18�C
was determined to yield the most active, soluble protein
and these conditions were used for all future analysis. The
soluble protein fraction was either affinity purified by
glutathione Sepharose 4B, or was desalted prior to use in
enzyme assays. One 50 ml bacterial culture produced
25–150 lg of purified recombinant (r)VLOGT2, 1–5 lg of
purified recombinant (r)VLOGT1 or 1–10 lg of purified
recombinant (r)VLOGT3. For this reason, only rVLOGT2
was purified prior to enzyme assays, while the desalted
soluble fraction was used in enzyme assays for rVLOGT3
and rVLOGT1 with the exception that rVLOGT3 was
purified prior to performing kinetic analyses.
RT-PCR analysis of VLOGT gene expression
Gene expression was monitored using the gene-specific
primers: VLOGT1,2—FP: 50 GCCCTTGGACACTATAT
GAATG 30, RP: 50 GGCGATGACTTTCCATCAGTC 30;VLOGT3—FP: 50 GGCGCTACAA GCTGTGGATAAGG
30, RP: 50 CTGCTGCTGTGTAGCAAAGC 30; VL3GT—
FP: 50TGCAGGGCCTAACTCACTCT 30, RP: 50 GCAGT
CGCCTTAGGTAGCAC 30; as well as the gene-specific
primers used in RT-PCR analysis for VLRSGT and VLActin
as reported previously (Hall and De Luca 2007).
VLOGT1,2; VLOGT3; and VLActin gene expression anal-
ysis was performed in duplicate and was quantified using
Multigauge ver 3.0 (Fujifilm, Tokyo, Japan; http://www.
fujifilm.ca), and the mean values were divided by the mean
actin gene expression value to obtain a relative value for
gene expression (% expression) in these tissues.
Template generated from RNA without reverse tran-
scription was used to check for genomic DNA contami-
nation. Tissue specificity of the RNA extracts was verified
using Vitis labrusca UDP-glucose: flavonoid 3-O-GT
(VL3GT) primers (which amplified a product from exocarp,
but not mesocarp tissue) and VLRSGT primers (which
amplified a product from mesocarp tissue but not exocarp
tissue) (Hall and De Luca 2007).
Gene specificity of the primers was verified by in-silico
analysis of the DFCI grape ESTs to the best of our
knowledge at the time (prior to release of the grape gen-
ome). Additionally, primer specificity was evaluated based
on amplification of PCR products from vectors containing
five Vitis labrusca GTs (VLOGT1, VLOGT2, VLOGT3,
VLRSGT, VL3GT). PCR using VLOGT1,2 and VLOGT3
gene-specific primers with identical amounts of plasmid
template and identical PCR cycling conditions generated
nearly identical amounts of amplified product.
In vivo biocatalysis of GT reactions
E. coli cultures expressing Vitis labrusca VLOGT1,
VLOGT2 and VLOGT3 proteins were grown as described
above. Concurrent with the induction of GT gene expres-
sion, substrates were added to a final concentration of
100 lM and the induced bacterial culture was grown for
20 h at 16�C, at which time the cultures were harvested by
centrifugation at 3,600g for 5 min. The supernatant was
decanted, acidified and extracted three times with ethyl
acetate. The ethyl acetate fractions were pooled, evapo-
rated to dryness, dissolved in 1 ml of methanol, and sub-
jected to HPLC.
Recombinant GT enzyme activity assays
Recombinant protein (rVLOGT1, rVLOGT2, rVLOGT3)
was isolated and rVLOGT2 was further purified by glu-
tathione Sepharose 4B (GE Healthcare) affinity chroma-
tography (as described in Hall and De Luca 2007). The
enzymes were assayed in 100 mM Tris, pH 8.0, with
0.1% b-mercaptoethanol in a final reaction volume of
125 ll, with 9 mM UDPG, 100 lM acceptor substrate
and 2–8 lg of purified rVLOGT2, 140–283 lg of desal-
ted total soluble rVLOGT1 protein or 119–242 lg of
desalted total soluble rVLOGT3 protein. Assays were
incubated for 2 h at 30�C, were stopped with an equal
volume of methanol or 0.12 N HCl in methanol (antho-
cyanidin substrates), were centrifuged, filtered and sub-
jected to HPLC analysis as described previously (Hall
and De Luca 2007).
The acceptor substrates quercetin, 2,4,5-tricholorphenol
(TCP), phenol, benzyl alcohol and phenylethanol were
assayed using the radioactive assay in a 100 ll reaction
volume containing 100 mM Tris–HCl, pH 8.0, 0.1% b-
mercaptoethanol, 100 lM (quercetin, TCP, phenol) or
20 mM (benzyl alcohol, phenylethanol) acceptor substrate,
25 mM gluconic acid lactone, 196 lM 14C-UDPG (specific
activity = 7.4 GBq/mmol), 804 lM cold UDPG and pro-
tein as described above. The assays were incubated for
30–120 min at 30�C, stopped with 10 ll of 1 N HCl,
extracted with 500 ll of ethyl acetate, centrifuged at
21,000g for 10 min and the organic phase was evaporated
to dryness. The products were resuspended in 10 ll of
methanol, spotted on a Polygram Sil/UV254 (Machery-
Nagel; Fisher Scientific) TLC plate and developed in the
solvent systems ethyl acetate:glacial acetic acid:formic
acid:water (100:11:11:20, by vol) (quercetin); ethyl ace-
tate:methanol:water (60:35:8, by vol) (TCP, phenol) or
ethyl acetate:acetone:dichloromenthane:methanol:water
(40:30:12:10:8, by vol) (benzyl alcohol, phenethyl alcohol
by vol) (Jones et al. 1999). The TLC plate was dried, and
exposed to a storage phosphor screen (GE Healthcare) for
1204 Planta (2011) 234:1201–1214
123

16–48 h prior to analysis with a Phosporimager FLA-3000
(Fujifilm) and MultiGuage ver 3.0.
Characterization of rVLOGT1, rVLOGT2
and rVLOGT3
Determination of pH optima, substrate specificities and
kinetic analysis were completed as described previously
(Hall and De Luca 2007) with the exceptions that the
typical buffer system used was 100 mM Tris–HCl, pH 8.0,
0.1% b-mercaptoethanol, and that the assay incubation
time was 2 h. To determine kinetic parameters with TCP, a
50 ll final reaction volume (100 mM Tris–HCl, pH 8.0,
0.1% b-mercaptoethanol), 50 lM C14UDPG, 250 lM of
cold UDPG 0.04 lg of purified rVLOGT2 and varying
concentrations of TCP were incubated for 90 min at 30�C,
were stopped with the addition of 5 ll of 0.12 N HCl and
extracted with 500 ll ethyl acetate, which was dried and
processed as described above. Non radioactive assays were
performed in order to determine the types of quercetin
glucosides produced by each GT reaction and reaction
products were analyzed and identified by HPLC–mass
spectrometry (HPLC-negative-ESI/MS) on an Agilent
1100 HPLC system equipped with a DAD detector and a
Bruker HCT ? ESI/MS.
The anthocyanin substrates cyanidin, cyanidin-3-O-
glucoside, cyanidin-3,5-O-diglucoside, peonidin, peonidin-
3-O glucoside, delphinidin, and malvidin; the stilbenes
trans-resveratrol, cis-resveratrol and trans-piceatannol; as
well as sapogenin, coumaric acid, sinapic acid, caffeic
acid, cinnamic acid, ferulic acid, chlorogenic acid, para-
hydroxybenzoic acid, benzoic acid, gallic acid, vanillin,
benzyl alcohol, phenylethanol, esculetin, catechin, epicat-
echin, phenol, abscisic acid, indole-3-acetic acid, salicylic
acid and UDP-glucuronic acid were not accepted by any of
the recombinant enzymes. UDP-galactose was not accepted
by rVLOGT2 with luteolin as an acceptor substrate, eri-
odictyol was not accepted by rVLOGT1 and 3, 6, 20, 30
tetrahydroxyflavone was not accepted by rVLOGT3.
In silico and phylogenetic analysis of Vitis labrusca
GTs
A BLAST search of the NCBI grape genome (Vitis vinifera
cv. Pinot Noir) database (http://www.ncbi.nlm.nih.gov/
genome/seq/BlastGen/BlastGen.cgi?taxid=29760) (PSPG)
box (WAPQVEVLAHPAVGCFVTHCGWNSTLESISAG
VPMVAWPFFADQ) (Vogt and Jones 2000) as a query
identified 273 sequences (e-5) with sequence identity
(E value \10-5) to plant secondary product glucos-
yltransferases. An additional BLAST search of the Vitis
vinifera genome with the nucleotide sequences of VLOGT1,
VLOGT2 and VLOGT3 identified the orthologous Vitis
vinifera sequences (LOC100248120; LOC100242998 and
LOC100246377, respectively). Sequence alignment and
phylogenetic analysis of the Vitis sp. GTs and other func-
tionally characterized GTs, were performed and visualized
using the Clustal algorithm of the CLC Bio Main Workbench
software (v.5.1).
Statistical analysis of the results
The data provided in Figs. 1, 2, 3 represents the mean of
3 or more biological replicates, while the relative gene
expression results provided in Fig. 4 represent the mean
real time RT-PCR values of two biological replicates.
The data provided in Fig. S1 represents the mean of 3
biological replicates. The data provided in Table S3 for
kinetic parameters of rVLOGT2 and rVLOGT3 repre-
sent mean values from at least three independent
experiments.
Results
Detection of cyanidin-3-O-glucoside-5-O-GT (Cy5OGT),
quercetin-30-O-GT (Q30OGT) and quercetin-7-O-GT
(Q7OGT) activities during grape berry development and in
several grape tissues.
Grape exocarp and mesocarp tissues prepared from
different stages of grape development were extracted and
desalted to produce crude protein extracts that were
assayed for Q30OGT and Q7OGT activity with quercetin as
glucose acceptor and for Cy5OGT activity using cyanidin
3-O-glucoside as glucose acceptor (Fig. 1). The different
quercetin glucosides including those substituted in posi-
tions 3, 7, 30 and 40 could be differentiated based on their
mass, HPLC retention times and absorbance maxima
(Table S2; Jurd 1962; Harborne 1967). Analyses by HPLC
of reaction products showed variable glucosylating activity
of each substrate depending on the tissue and stage of
development being analyzed (Fig. 1). While high levels of
Q7GT activity were clearly detected within grape meso-
carp (Fig. 1a) during grape ripening (veraison) between
weeks 12 and 16 AF, little of this activity could be detected
within grape exocarp in any stage of development
(Fig. 1b). Interestingly, there were low levels of Q30GT
activity in grape mesocarp but not in grape exocarp during
weeks 1–3 AF that disappeared between weeks 4–8 and
then reappeared during veraison between weeks 10–16 AF.
In contrast to these results, Cy5OGT activity was exclu-
sively found in grape exocarp, that it is the main site of
anthocyanin biosynthesis during grape development and
only with the onset of veraison between weeks 8–16
(Fig. 1b).
Planta (2011) 234:1201–1214 1205
123
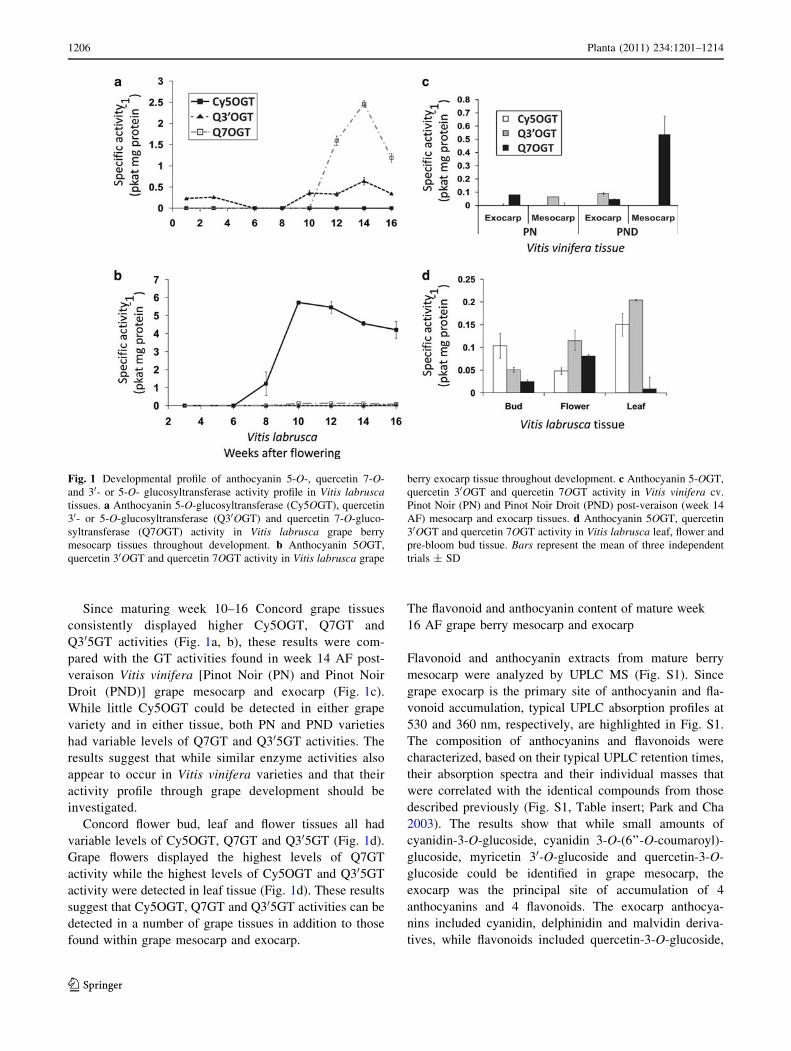
Since maturing week 10–16 Concord grape tissues
consistently displayed higher Cy5OGT, Q7GT and
Q305GT activities (Fig. 1a, b), these results were com-
pared with the GT activities found in week 14 AF post-
veraison Vitis vinifera [Pinot Noir (PN) and Pinot Noir
Droit (PND)] grape mesocarp and exocarp (Fig. 1c).
While little Cy5OGT could be detected in either grape
variety and in either tissue, both PN and PND varieties
had variable levels of Q7GT and Q305GT activities. The
results suggest that while similar enzyme activities also
appear to occur in Vitis vinifera varieties and that their
activity profile through grape development should be
investigated.
Concord flower bud, leaf and flower tissues all had
variable levels of Cy5OGT, Q7GT and Q305GT (Fig. 1d).
Grape flowers displayed the highest levels of Q7GT
activity while the highest levels of Cy5OGT and Q305GT
activity were detected in leaf tissue (Fig. 1d). These results
suggest that Cy5OGT, Q7GT and Q305GT activities can be
detected in a number of grape tissues in addition to those
found within grape mesocarp and exocarp.
The flavonoid and anthocyanin content of mature week
16 AF grape berry mesocarp and exocarp
Flavonoid and anthocyanin extracts from mature berry
mesocarp were analyzed by UPLC MS (Fig. S1). Since
grape exocarp is the primary site of anthocyanin and fla-
vonoid accumulation, typical UPLC absorption profiles at
530 and 360 nm, respectively, are highlighted in Fig. S1.
The composition of anthocyanins and flavonoids were
characterized, based on their typical UPLC retention times,
their absorption spectra and their individual masses that
were correlated with the identical compounds from those
described previously (Fig. S1, Table insert; Park and Cha
2003). The results show that while small amounts of
cyanidin-3-O-glucoside, cyanidin 3-O-(6’’-O-coumaroyl)-
glucoside, myricetin 30-O-glucoside and quercetin-3-O-
glucoside could be identified in grape mesocarp, the
exocarp was the principal site of accumulation of 4
anthocyanins and 4 flavonoids. The exocarp anthocya-
nins included cyanidin, delphinidin and malvidin deriva-
tives, while flavonoids included quercetin-3-O-glucoside,
Fig. 1 Developmental profile of anthocyanin 5-O-, quercetin 7-O-
and 30- or 5-O- glucosyltransferase activity profile in Vitis labruscatissues. a Anthocyanin 5-O-glucosyltransferase (Cy5OGT), quercetin
30- or 5-O-glucosyltransferase (Q30OGT) and quercetin 7-O-gluco-
syltransferase (Q7OGT) activity in Vitis labrusca grape berry
mesocarp tissues throughout development. b Anthocyanin 5OGT,
quercetin 30OGT and quercetin 7OGT activity in Vitis labrusca grape
berry exocarp tissue throughout development. c Anthocyanin 5-OGT,
quercetin 30OGT and quercetin 7OGT activity in Vitis vinifera cv.
Pinot Noir (PN) and Pinot Noir Droit (PND) post-veraison (week 14
AF) mesocarp and exocarp tissues. d Anthocyanin 5OGT, quercetin
30OGT and quercetin 7OGT activity in Vitis labrusca leaf, flower and
pre-bloom bud tissue. Bars represent the mean of three independent
trials ± SD
1206 Planta (2011) 234:1201–1214
123

quercetin-3-O-glucuronide, myricetin-3-O-glucuronide,
myricetin 30-O-glucoside. It is important to note that
quercetin and myricetin 7-O-glucosides were not detected
in either exocarp or mesocarp tissues.
Identification and molecular cloning
of glucosyltransferase-like genes from grape berry
exocarp-enriched tissue
Initially, the corresponding full-length clones were isolated
from Vitis labrusca week 12 AF exocarp-enriched cDNA
using multiple rounds of 50RACE. During the course of
research, the grape genome was released, confirming the
presence of these genes within the Vitis vinifera genome.
A BLAST search of the DFCI grape index EST database with
the plant secondary product glucosyltransferase (PSPG) box
consensus sequence (Vogt and Jones 2000) identified 141
expressed Vitis sequences (E value \10e-5). Three Vitis
vinifera ESTs (TC75303, TC91700, NP9526770) with at
least 60% amino acid sequence similarity to previously
characterized flavonoid GTs were identified and pursued
based on their expression profile in several grape tissues,
including berries, flowers, and leaves, which glucosylate
quercetin at the 7-O-position (Fig. 1).
An alignment of TC75303, TC91700 and NP9526770
with the 273 Vitis vinifera genomic glucosyltransferases
suggested that these ESTs corresponded to three unique
genomic glucosyltransferase sequences located on chro-
mosome 5. TC75303 had 97% nucleotide sequence identity
to LOC100246377, TC91700 had 99% nucleotide
sequence identity to LOC100242998 and NP9526770 had
99% nucleotide sequence identity to LOC100248120.
Primers based on the predicted open reading frame of
these genes were used with Vitis labrusca week 12 AF
exocarp cDNA to amplify the corresponding transcripts
from Concord grape. The Concord clones (VLOGT1,
VLOGT2, VLOGT3) had 98–99% nucleotide sequence
identity and 97–98% amino acid sequence identity to their
Vitis vinifera equivalents (NP9526770, TC91700,
TC75303). VLOGT1 has 93 and 62% nucleotide sequence
identity to VLOGT2 and VLOGT3, respectively (1,344 bp)
and the three genes encode putative proteins of 447, 448
and 464 aa, respectively, with corresponding theoretical
molecular weights of 49.8, 49.7 and 51.3 kDa.
The presence of the PSPG box in the C-terminus of the
three Vitis labrusca clones (Fig. S2) placed these proteins
with more than 100 functionally characterized family 1 GTs
(Fig. S3). A BLAST search with VLOGT1, VLOGT2 and
VLOGT3 protein sequences revealed at least 60% amino
acid similarity to a Nicotiana tabacum GT (NtGT2) that
glucosylates the 3-O- position of coumarins and the 7-O-
position of flavonols (Taguchi et al. 2003; BAB88935), to a
Lycium barbarium catechin 40-O-GT which also glucosy-
lates the 7-O-position of flavonols (Noguchi et al. 2007;
BAG80544) to a Eucalyptus perriniana monoterpene O-GT
(Nagashima et al. 2004; BAD90934), to an anthocyanin
5-O-glucosyltransferase from Petunia x hybrida (Yamazaki
et al. 2002 (BAA89009), and to two flavonoid 7-O-GTs
(7GT)s from Maclura pomifera (Tian et al. 2006;
ABL85474) and Pyrus communis (Fischer et al. 2007;
AAY27090) (Fig. S3).
Biofermentation of kaempferol by rVLOGT1,
rVLOGT2, and rVLOGT3
Bacterial cultures (50 ml) expressing rVLOGT1, rVLOGT2,
rVLOGT3 or the pGEX empty vector were incubated with
cyanidin 3-O-glucoside (anthocyanin), kaempferol (flavonol)
and sapogenin (saponin) (Cascado and Heredia 1999) as
potential substrates for glucosylation. Cultures expressing the
empty vector did not glucosylate any of the substrates;
whereas cultures expressing rVLOGT1, rVLOGT2, and
Fig. 2 Effect of assay pH on rVLOGT2 and rVLOGT3 activity in
vitro. a Effect of assay pH on the production of quercetin 7-O-
monoglucoside (Q 7OMG), quercetin 30 or 5-O-monoglucoside
(Q 30OMG), and quercetin 40-O-monoglucoside (Q 40OMG) by
rVLOGT2 in vitro. b Effect of assay pH on the production of
isorhamnetin 7-O-monoglucoside (IsoR7OMG) by rVLOGT3 in
vitro. Each point represents the mean of three independent
trials ± SD
Planta (2011) 234:1201–1214 1207
123
rVLOGT3 produced 40.5 ± 11.6, 112.5 ± 14.0 and
26.8 ± 2.6 ng of kaempferol 7-O-glucoside/ml of cell culture
media [identified by co-chromatography with authentic stan-
dard and by UV spectral analysis (Hall and De Luca 2007)],
respectively, but none of the cultures were able to glucosylate
sapogenin or cyanidin-3-O-glucoside. The product of ka-
empferol glucosylation was identified as kaempferol 7-O-
glucoside by co-chromatography and UV spectral analysis
(Hall and De Luca 2007). These results confirmed that
rVLOGT1, rVLOGT2, and rVLOGT3 expressing lines were
biochemically active for further characterization using a cell-
free system.
Preliminary in vitro characterization of recombinant
GTs
E. coli cells expressing VLOGT1, VLOGT2 and VLOGT3
were induced for protein expression, and bacterial pellets
were harvested and extracted. SDS-PAGE analysis indi-
cated that approximately 95% of the GST-tagged recom-
binant proteins (molecular weight, 80 kDa) were insoluble.
Attempts to solubilise and refold an active protein from the
insoluble protein fractions of VLOGT1, VLOGT2 and
VLOGT3 were unsuccessful. When soluble active enzymes
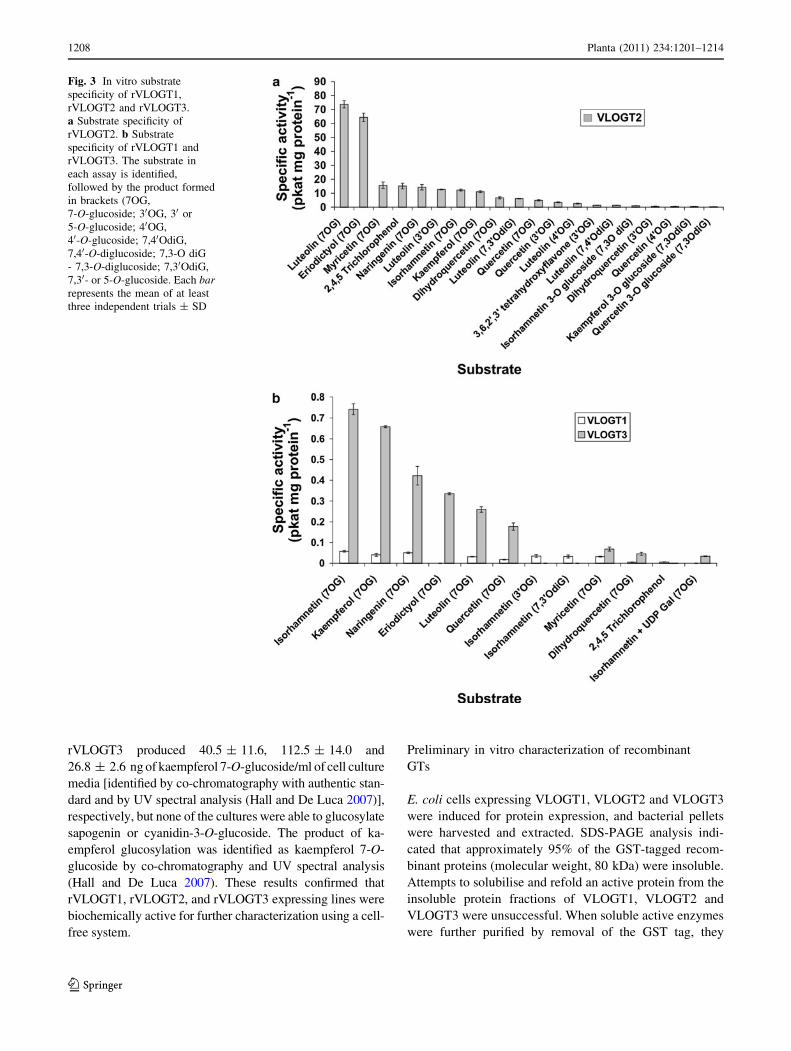
were further purified by removal of the GST tag, they
Fig. 3 In vitro substrate
specificity of rVLOGT1,
rVLOGT2 and rVLOGT3.
a Substrate specificity of
rVLOGT2. b Substrate
specificity of rVLOGT1 and
rVLOGT3. The substrate in
each assay is identified,
followed by the product formed
in brackets (7OG,
7-O-glucoside; 30OG, 30 or
5-O-glucoside; 40OG,
40-O-glucoside; 7,40OdiG,
7,40-O-diglucoside; 7,3-O diG
- 7,3-O-diglucoside; 7,30OdiG,
7,30- or 5-O-glucoside. Each barrepresents the mean of at least
three independent trials ± SD
1208 Planta (2011) 234:1201–1214
123

displayed the same activity when they were GST tagged.
Crude desalted protein extracts of rVLOGT1 and
rVLOGT3 as well as GST purified rVLOGT2 were tested
with UDPG and the substrates quercetin, isorhamnetin,
dihydroquercetin, sinapic acid, cyanidin, cyanidin-3-O-
glucoside, para-hydroxybenzoic acid, trans-resveratrol,
and esculetin. Remarkably rVLOGT1 and rVLOGT3 con-
verted quercetin and isorhamnetin to monoglucosides as
determined by HPLC, while rVLOGT2 converted querce-
tin to three separate monoglucosides and kaempferol,
isorhamnetin and dihydroquercetin to monoglucosides. No
reaction products were formed with boiled enzymes or with
bacterial extracts expressing empty vector.
Since rVLOGT1 and rVLOGT3 seemed to have simi-
lar substrate specificities, further studies were performed
with rVLOGT2 and rVLOGT3 using quercetin to deter-
mine the pH optima of these 2 enzymes as described
previously (Hall and De Luca 2007). The pH optima of
rVLOGT2 was between pH 8.5 and 9.0 (Fig. 2a) while
that of rVLOGT3 was pH 8.5 (Fig. 2b). However
rVLOGT2 converted quercetin to one major (quercetin
7-O-monoglucoside; no hypsochromic shift of UV max
from 370 nm; Harborne 1967; Vogt et al. 1997; Kramer
et al. 2003; Hall and De Luca 2007) and 2 minor (quer-
cetin 30-O-monoglucoside and quercetin 40-O-monog-
lucoside; Jurd 1962; Vogt et al. 1997; Kramer et al. 2003)
products that were identified based on their different
HPLC retention times, their different absorbance maxima
and their identical masses (Table S2). In contrast,
rVLOGT3 converted quercetin to a single monoglucoside
that had the same HPLC, absorbance maxima and mass
properties of quercetin 7-O-glucoside.
Substrate specificity of rVLOGT1, rVLOGT2
and rVLOGT3
Substrate specificity of the recombinant proteins was
determined in vitro with UDP-glucose (UDPG) as a donor
substrate and 34 different acceptor substrates for
rVLOGT1, while 45 different acceptor substrates for tested
for rVLOGT2 and rVLOGT3. Recombinant rVLOGT2
produced the 7-O-glucoside of several flavonoids and
glucosylated the non-flavonoid model xenobiotic substrate
2,4,5-trichlorophenol (TCP). The 7-O-position of luteolin
Fig. 4 Relative gene expression of VlOGT1,2, and VlOGT3 as
compared to VLActin in berry mesocarp (a) and berry exocarp
(b) tissues of Vitis labrusca (VL) and in berry mesocarp and exocarp
tissues of Vitis vinifera cvs. Pinot Noir (PN) and Pinot Noir Droit
(PND) (c). d Relative gene expression of VlOGT1,2, and VlOGT3compared to VLActin gene expression in Vitis labrusca leaf, flower
bud and flower tissue. All values represent the mean of two
independent trials ± SD
Planta (2011) 234:1201–1214 1209
123

and eriodictyol were glucosylated by rVLOGT2 with the
highest specific activities (Fig. 3a) while the 30-O-gluco-
sides of luteolin and 3,6,20,30-tetrahydroxyflavone and the
40-O-glucosides of luteolin and quercetin were also pro-
duced at low levels. Recombinant VLOGT2 also converted
luteolin to 7,30-O-diglucoside and 7,40-O-diglucosides in
small amounts, while isorhamnetin and kaempferol could
also be converted to the corresponding 3, 7-O-diglucosides
(Fig. 3a).
Incubation of rVLOGT1 with isorhamnetin and UDPG
produced isorhamnetin 7-O-glucoside as the major product
and isorhamnetin 30-O- or 5-O-glucoside and isorhamnetin
7,30-O-diglucoside as minor products. rVLOGT1 also
glucosylated the 7-O -position of the flavonoids naringenin,
kaempferol, myricetin, luteolin, quercetin and dihydroqu-
eretin, while it was also able to glucosylate TCP at low
levels (Fig. 3b). rVLOGT3 glucosylated TCP and pro-
duced single products, identified as the 7-O-glucosides of
isorhamnetin, kaempferol, naringenin, eriodictyol, luteolin,
quercetin, myricetin and dihydroquercetin T (Fig. 3b).
Interestingly, VLOGT3 was able to use UDP-galactose
(UDP-Gal) as a donor with isorhamnetin as an acceptor at
low levels (Fig. 3b). Substrates which were not accepted by
any of the recombinant proteins are listed in ‘‘Materials and
methods.’’
Michaelis-Menton kinetic analysis of rVLOGT2
and rVLOGT3
Since recombinant enzymes appear to accept a broad range
of flavonoids as substrates, kinetic analyses were per-
formed with myricetin (for rVLOGT2) and isorhamnetin
(for rVLOGT3) as well as with TCP for both enzymes.
Kinetic analysis of rVLOGT2 calculated a 3.8-fold higher
affinity (Km) for myricetin than for TCP (48.4 and
184.5 lM) (Table S3). The turnover number (Kcat) with
TCP as a substrate for rVLOGT2 (Kcat = 105.7 s-1) is
more than 10,000-fold higher than with myricetin
(Kcat = 2.4 9 E-4 s-1) and the calculated catalytic effi-
ciency (Kcat/Km) is more than 100,000-fold higher with
TCP (Kcat/Km = 0.573 lM-1 s-1) than with myricetin
(Kcat/Km = 4.96 E-6 lM-1 s-1) (Table S3).
Michaelis-Menton kinetic analysis of rVLOGT3 with
varying concentrations of isorhamnetin yielded a Km of
30.2 lM, a Kcat of 8.11 E-7 s-1, and a Kcat/Km of
2.69 E-8 lM-1 s-1 (Table S3), similar to the kinetic
characteristics of rVLOGT2 with myricetin. Kinetic anal-
ysis of UDPG for rVLOGT2 (100 lM myricetin) and
rVLOGT3 (100 lM isorhamnetin) yielded Kms of 940,
and 886 lM, respectively; Kcats of 2.0 E-4 s-1 and
7.67 E-6 s-1, respectively, and Kcat/Kms of 2.13
E-7 lM-1 s-1 and 8.66 E-9 lM-1 s-1, respectively
(Table S3).
Gene expression profiling of VLOGT1 and VLOGT2
(VLOGT1,2) and VLOGT3 in grape berry, vegetative
and reproductive tissues
RNA was extracted from grape tissues and was screened
with one pair of primers to amplify both VLOGT1 and
VLOGT2, and a second pair of VLOGT3-specific primers.
Amplification with gene-specific primers corresponding to
the Vitis vinifera actin transcript was used to determine
baseline gene expression. The relative expression of the
VLOGT genes was analyzed in berry mesocarp (Fig. 4a)
and exocarp (Fig. 4b) to show that all 3 genes appeared to
be expressed to various extents throughout grape devel-
opment., with VLOGT1, 2 being expressed at higher levels
than VLOGT3 throughout development (Fig. 4a, b). While
the VLOGT3 transcript was variably detected throughout
grape development in both tissues, no clear peaks of gene
expression could be established to correlate with the 7- and
30/5-OGT enzyme activities observed throughout exocarp
and mesocarp grape development (Fig. 1a, b). In contrast
the relative expression of VLOGT1, 2 transcripts increases
in both mesocarp (Fig. 4a) and exocarp (Fig. 4b) tissues
with the onset of grape ripening from week 8 onwards that
may tentatively be correlated with the observed increases
of 7-OGT activities observed in mesocarp tissues (Fig. 1a).
VLOGT1, 2 also appeared to be expressed in both week
14 AF berry exocarp and mesocarp tissues of Vitis vinifera
cv. Pinot Noir and cv. Pinot Noir Droit, with lower relative
expression levels of the VLOGT3 transcript being detected
in berry exocarp tissue and no transcript being detected in
berry mesocarp tissue (Fig. 4c). Conversely, both
VLOGT1, 2 and VLOGT3 transcripts were detected in Vitis
labrusca leaf, flower bud and flower tissues (Fig. 4d).
These expression profiles are consistent with the corre-
sponding Vitis vinifera ESTs expression summaries from
the DFCI EST database in which these genes are expressed
in several tissues including flowers, pre-veraison berries,
and roots.
Phylogenetic analysis places VLOGT1, VLOGT2
and VLOGT3 in a clade with flavonoid 7GTs
A BLAST search of the NCBI grape genome database
(http://www.ncbi.nlm.nih.gov/genome/seq/BlastGen/Blast
Gen.cgi?taxid=29760) with the three VLOGT1, VLOGT2,
and VLOGT3 queries identified three corresponding geno-
mic sequences from Vitis vinifera cv. Pinot Noir (LOC
100248120, LOC100242998 and LOC100246377, respec-
tively) located on chromosome 5.
A phylogenetic tree places the Vitis sp. GTs identified in
this study in a clade with enzymes which glucosylate
terpenes, catechins, the 5-O-position of anthocyanins and
the 7-O-position of flavonols (Fig. S3; Yamazaki et al. 2002;
1210 Planta (2011) 234:1201–1214
123

Taguchi et al. 2003; Nagashima et al. 2004; Tian et al. 2006;
Fischer et al. 2007; Noguchi et al. 2007). GTs which accept
TCP as a substrate (Loutre et al. 2003; Meßner et al. 2003),
other 7GTs (Hirotani et al. 2000; Kramer et al. 2003; Jones
et al. 2003; Willits et al. 2004) and other 5GTs (Imayama et al.
2004; Ogata et al. 2005; Tohge et al. 2005) are located
throughout the phylogenetic tree (Fig. S3).
Discussion
The Vitis vinifera genome contains more than 140
genes with identity to glucosyltransferases
More than 150 plant group 1 glucosyltransferases have
been cloned from various plant sources, expressed in
recombinant organisms and functionally characterized in
vitro. This includes the recombinant expression of 107 GTs
from Arabidopsis which have been screened for activity
towards terpenes (Caputi et al. 2008), cytokinins (Hou et al.
2004), ABA (Lim et al. 2005), and quercetin (Lim et al.
2003). A BLAST search of the Vitis vinifera cv. Pinot noir
genome sequence with the PSPG box (Vogt and Jones
2000) identified 243 group 1 glucosyltransferases and a
BLAST search of the Vitis vinifera DFCI grape gene index
with the PSPG box as a query identified 141 Vitis sp. group
1 glucosyltransferases, which are expressed in grape, sug-
gesting biological roles for many of these enzymes.
Despite the prevalence of expressed GT sequences, only
six of the Vitis sp. enzymes have been functionally char-
acterized. Glucosyltransferases involved in the biosynthe-
sis of the viticulturally important anthocyanins (Ford et al.
1998; Janvary et al. 2009), a glycosyltransferase involved
in formation of flavonol 3-glucoronate (Ono et al. 2010), a
bifunctional glycosyltransferase involved in the formation
of flavonol-3-glucoside or galactoside (Ono et al. 2010)
and a bifuncional grape GT, which glucosylates hydroxy-
cinnamic acids and the human health beneficial stilbenes
(Hall and De Luca 2007) have characterized at the
molecular and biochemical level. In addition to anthocya-
nins, hydroxycinnamic acids and stilbenes, grapes biosyn-
thesize and accumulate the 3-O-, 7-O-, and 30O-glucosides
of many flavonols including quercetin, kaempferol, my-
ricetin and isorhamnetin that may involve GTs, which have
yet to be characterized. For example, recent studies (Park
and Cha 2003) using spot densities of flavonoids separated
by cellulose TLC combined with HPLC analysis identified
Q3G as a major flavonoid in grape exocarps while Q3G
and Q3G-7-O-glucuronide were the major flavonoids of
grape leaves and isorhamnetin-3-O–G and kaempferol
3,7-O–G were found in minor quantities.
The current study describes the homology-based cloning
and biochemical characterization of three full-length
(VLOGT1, VLOGT2, VLOGT3) members of the Vitis
labrusca cv. Concord flavonoid 7 GT gene family. The
deduced protein sequences of these genes are highly
identical to the 7 GTs from other plant species and group
phylogenetically with GTs that glucosylate monoterpenes,
phenolic acids, flavonols and dihydroflavonols in vitro
(Fig. S3; Lim et al. 2002; Taguchi et al. 2003; Nagashima
et al. 2004; Tian et al. 2006; Fischer et al. 2007; Noguchi
et al. 2007).
rVLOGT1, rVLOGT2, and rVLOGT3 glucosylate
flavonoids and the xenobiotic TCP
Biocatalysis with bacterial cultures expressing rVLOGT1,
rVLOGT2, and rVLOGT3 converted kaempferol to the
7-O-glucoside. In vitro cell-free extracts of these enzymes
glucosylated the model xenobiotic TCP as well as several
flavonoids to produce mono- and di-glucosides (Fig. 3).
rVLOGT2 predominantly glucosylated the 7-O-position in
vitro, with the flavone luteolin and the flavanone eri-
odictyol (differing only in B-ring stereochemistry) yielding
the two highest specific activities. Enzyme assays with
luteolin yielded 3 mono- and 2 di-glucosylated products,
whereas only the 7-O-glucoside was detected with eri-
odictyol as a substrate, suggesting that B-ring orientation is
essential for secondary and tertiary glucosylations. This
broad regio-specificity is consistent with previously char-
acterized flavonoid 7GTs, which can glucosylate multiple
positions on several flavonoid substrates in vitro (Kramer
et al. 2003; Tian et al. 2006).
VLRGT 1, 2, 3 gene expression as it relates to GT
activities and flavonoid/anthocyanin profiles
The predominant anthocyanins within Concord grape berry
are anthocyanidin 3-O-glucosides and quercetin 3-O-
glucosides (Park and Cha 2003; Wu and Prior 2005),
respectively. While the present analysis of mature Concord
berry tissues (Fig. S1) confirms the prevalence in the
exocarp of the 3-O-glucosides of different anthocyanidins-
and of quercetin, this tissue also contains the considerable
amounts of quercetin 3-O-glucuronide, myricetin 3-O-
glucoside and myricetin 3-O-glucuronide. The exocarp
tissue of several grape cultivars (Park and Cha 2003;
Amico et al. 2008; Castillo-Munoz et al. 2009) have also
been shown to accumulate quercetin 3-O-rhamnoglucoside
as well as the 3-O-galactosides, 3-O-glucuronides and 3-O-
glucosides of quercetin, kaempferol, myricetin, isorham-
netin, laricitrin and syringetin. Consistent with the preva-
lence of 3-O-glucosylated metabolites and the lack of 7-O-;
30-O-; or 40-O-glucosides in grape berries, enzyme assays
for quercetin-3-OGT showed up to 100-fold higher specific
activity (250 pkat/mg protein) than those obtained for
Planta (2011) 234:1201–1214 1211
123

Q7GT (2.5 pkat/mg protein) or Q30GT (0.5 pkat/mg pro-
tein) (Fig. 1) found in grape berry tissues. Low levels of
Q30GT activity were detected biphasically in Concord
berry mesocarp tissue, whereas both Q30GT and Q7GT
activity were detected during veraison in Vitis labrusca and
Vitis vinifera exocarp tissue (Fig. 1), consistent with
VLOGT1,2 and VLOGT3 gene expression in pre- and post-
veraison mesocarp and exocarp (Fig. 4). While the low
reaction rates observed in vitro (rVLOGT2 Kcat/Km
(myricetin) = 4.96 M-1 s-1; rVLOGT3 Kcat/Km(isorham-
netin) = 0.0269 M-1 s-1) are consistent with the lack of
accumulation of these metabolites within the grape berry,
these observations do not preclude a role for these genes in
flavonol modification in planta. However, these low reac-
tion rates may also suggest that the correct substrates for
VLOGT1,2,3 remain to be determined as well as the
pathways that they are involved with.
Within Vitis labrusca flower and bud tissue, Q7GT
activity is detected (Fig. 1c), and VLOGT1,2 and VLOGT3
are expressed (Fig. 4d). In Vitis labrusca leaves, the
expression of VLOGT1,2 and VLOGT3 (Fig. 4c) and the
detection of Q7GT activity is consistent with the accu-
mulation of kaempferol 3,7-O-diglycoside (Park and Cha
2003) and luteolin 7-O-glucoside (Moore and Giannasi
1994).
Broad substrate specificity and kinetic analysis suggests
a possible biological role for VLOGT1, VLOGT2
and VLOGT3 in the plant stress response to xenobiotics
In plants, GTs exist as large multigene families (Li et al.
2001; Bowles 2002; Lim et al. 2003) and have broad, but
regio-selective substrate specificity (Vogt and Jones 2000),
allowing the plant to glucosylate a wide range of natural
products and xenobiotics. Although in vitro characteriza-
tion of GTs may not reflect their in vivo function, over-
expression in heterologous systems facilitates the discovery
of several catalytic roles. The glucosylation of multiple
positions and functional groups on many metabolites in
addition to glucosylation of the endogenous substrate has
been demonstrated (Vogt et al. 1999; Kramer et al. 2003;
Nagashima et al. 2004; Tian et al. 2006; Hall and De Luca
2007; Brazier-Hicks et al. 2007).
Broad substrate specificity of GTs is a proposed mech-
anism that plants use to respond to their constantly
changing environment, and serves as a mechanism to
detoxify and compartmentalize xenobiotics (Jones and
Vogt 2001; Brazier-Hicks et al. 2007). The 7GTs from
Maclura pomifera, Oryza sativa, Allium cepa and Scutel-
laria baicalensis are not regio-selective since they produce
different mono- and di-glucosides with various flavonoid
substrates (Kramer et al. 2003; Jones et al. 2003; Tian et al.
2006). Similarly, rVLOGT1, rVLOGT2 and rVLOGT3
exhibit broad substrate specificity and VLOGT2 also
demonstrates broad regio-specificity with flavonol sub-
strates (Fig. 3). The recombinant Vitis labrusca GTs
characterized in this study glucosylate the xenobiotic TCP
in vitro (Fig. 3), and kinetic analysis of rVLOGT2 reveals
that TCP is the preferred substrate for this enzyme (Kcat/Km
for TCP is 105-fold higher than with myricetin) (Table S3),
consistent with a possible role for these enzymes in
xenobiotic detoxification.
Many Arabidopsis thaliana GTs glucosylate plant
metabolites and are also active in glucosylating xenobiotics
with varying affinities and catalytic efficiencies (Meßner
et al. 2003). Similarly, many GTs with broad substrate
specificity (including 7GTs) are often rapidly induced by
the application of herbicide safeners (Loutre et al. 2003)
and by wounding (Hirotani et al. 2000). The induction of
GTs by treatment with methyl jasmonate (Imanishi et al.
1998; Taguchi et al. 2003) or with salicylic acid (Hirotani
et al. 2000; Taguchi et al. 2003) suggests a proposed
downstream role in systemic acquired resistance and the
plant defense response (Dong 2004). Simultaneous initia-
tion of the plant defense response and an increase of GT
gene expression would prepare the plant for the concomi-
tant increase of secondary metabolites produced by the
plant (which are glucosylated and stored in vacuoles) to
combat the pathogen, as well as to prepare the plant for
exposure to pathogen-derived metabolites that can be
detoxified by glucosylation. The 7GT-like gene family of
Vitis labrusca likely contributes to this model of plant
defense, since these genes are expressed spatially and
temporally in tissues frequently subjected to pathogen
attack, such as post-veraison berry exocarp, leaf, flower
and pre-bloom bud tissue (Fig. 4).
Conclusion
This study has used homology-based cloning to identify
three full-length members of the Vitis labrusca cv. Concord
7GT gene family that correlate with homologous genes
found in the Vitis vinifera genome. Biocatalysis and sub-
strate specificity studies indicate that the Vitis labrusca
GTs can glucosylate flavonols as well as the xenobiotic
TCP in vitro. While enzyme activity profiles with crude
extracts from Vitis labrusca show that both 7- and 30-OGT
activities can be found in grape berries, in leaves and in
flower tissues, relevant 7- or 30-O-glucosides (Fig. S1, apart
from myricetin 30-O-glucoside) do not appear to accumu-
late in grape berries. The 100-fold lower levels of 7- and 30-OGT activities observed compared with those Q3GT may
suggest that VLOGTs 1, 2 & 3 play very minor roles in
flavonoid-O-glucosylation during grape development.
Kinetic analysis indicates that TCP is the preferred
1212 Planta (2011) 234:1201–1214
123

substrate for this class of enzymes, and gene expression of
VLOGT1,2 and VLOGT3 in tissues which are commonly
exposed to pathogen attack and xenobiotics suggests a
possible second role for the Vitis labrusca 7GT gene family
in plant defense.
Acknowledgments This research was funded by a Natural Sciences
and Engineering Research Council of Canada Discovery grant to
V.D.L. and a Tier 1 Canada Research Chair in Plant Biotechnology to
V.D.L., as well as an NSERC post-graduate fellowship to D.H. We
thank G&H Wiley Ltd (St, Catharine’s, Ontario, Canada) for allowing
us to collect Concord grapes during the growing season. We thank
Tim Jones for performing HPLC-mass spectrometry on selected fla-
vonoid reaction products.
References
Ali A, Strommer J (2003) A simple extraction and chromatographic
system for the simultaneous analysis of anthocyanins and
stilbenes of Vitis species. J Agric Food Chem 51:7246–7251
Amico V, Chillemi R, Mangiafico S, Spatafora C, Tringali C (2008)
Polyphenol-enriched fractions from Sicilian grape pomace:
HPLC-DAD analysis and antioxidant activity. Bioresource
Technol 99:5960–5966
Bowles D (2002) A multigene family of glycosyltransferases in
a model plant, Arabidopsis thaliana. Biochem Soc Trans
30:301–306
Brazier-Hicks M, Edwards LA, Edwards R (2007) Selection of plants
for roles in phytoremediation: the importance of glucosylation.
Plant Biotechnol J 5:627–635
Caputi L, Lim E, Bowles D (2008) Discovery of new biocatalysts
for the glucosylation of terpenoid scaffolds. Chem Eur J
14:6656–6662
Cascado CG, Heredia A (1999) Structure and dynamics of reconsti-
tuted cuticular waxes of grape berry cuticle (Vitis vinifera L.).
J Exp Bot 50:175–182
Castillo-Munoz N, Gomez-Alonso S, Garcıa-Romero E, Hermosın-
Gutierrez I (2007) Flavonol profiles of Vitis vinifera red grapes
and their single cultivar wines. J Agric Food Chem 55:992–1002
Castillo-Munoz N, Gomez-Alonso S, Garcıa-Romero E, Gomez V,
Velders A, Hermosın-Gutierrez I (2009) Flavonol 3-O-glyco-
sides series of Vitis vinifera cv. Petit Verdot red wine grapes.
J Agr Food Chem 57:209–219
Dong X (2004) NPR1, all things considered. Curr Opin Plant Biol
7:547–552
Fischer T, Gosch C, Pfeiffer J, Halbwirth H, Halle C, Stich K,
Forkmann G (2007) Flavonoid genes of pear (Pyrus communis).
Trees 21:521–529
Ford CM, Boss PK, Høj PB (1998) Cloning and characterization of
Vitis vinifera UDP-Glucose:flavonoid 3-O-glucosyltransferase, a
homologue of the enzyme encoded by the maize Bronze-1 locus
that may primarily serve to glucosylate anthocyanidins in vivo.
J Biol Chem 273:9224–9233
Hall D, De Luca V (2007) Mesocarp localization of a bi-functional
resveratrol/hydroxycinnamic acid glucosyltransferase of Con-
cord grape (Vitis labrusca). Plant J 49:579–591
Harborne JB (1967) Comparative biochemistry of the flavonoids.
Academic Press, London
Hirotani M, Kuroda R, Suzuki H, Yoshikawa T (2000) Cloning and
expression of UDP-glucose: flavonoid 7-O-glucosyltransferase
from hairy root cultures of Scutellaria baicalensis. Planta
210:1006–1013
Hou B, Lim E, Higgins G, Bowles D (2004) N-Glucosylation of
cytokinins by glucosyltransferases of Arabidopsis thaliana.
J Biol Chem 279:47822–47832
Huntley A (2007) Grape flavonoids and menopausal health. Meno-
pause Int 13:165–169
Imanishi S, Hashizume K, Kojima H, Ichihara A, Nakamura K (1998)
An mRNA of tobacco cell, which is rapidly inducible by methyl
jasmonate in the presence of cycloheximide, codes for a putative
glycosyltransferases. Plant Cell Physiol 39:202–211
Imayama T, Yoshihara N, Fukuchi-Mizutani M, Tanaka Y, Ino I,
Yabuya T (2004) Isolation and characterization of a cDNA clone
of UDP-glucose: anthocyanin 5-O-glucosyltransferase in Irishollandica. Plant Sci 167:1243–1248
Janvary L, Hoffmann T, Pfeiffer J, Hausmann L, Topfer R, Fischer T,
Schwab W (2009) A double mutation in the anthocyanin 5-O-
glucosyltranferase gene disrupts enzymatic activity in Vitisvinifera L. J Agr Food Chem 57:3512–3518
Jones P, Vogt T (2001) Glycosyltransferases in secondary plant
metabolism: tranquilizers and stimulant controllers. Planta
213:164–174
Jones P, Møller B, Høj P (1999) The UDP-glucose:p-hydroxymand-
elonitrile-O-glucosyltransferase that catalyzes the last step in
synthesis of the cyanogenic glucoside dhurrin in Sorghumbicolor. J Biol Chem 274:35483–35491
Jones P, Meßner B, Nakajima JI, Schaffner AR, Saito K (2003)
UGT73C6 and UGT78D1, glycosyltransferases involved in
flavonol glycoside biosynthesis in Arabidopsis thaliana. J Biol
Chem 278:43910–43918
Jurd L (1962) Spectral properties of flavonoid compounds. In:
Geissmann TA (ed) The chemistry of flavonoid compounds.
Pergamon Press Inc, Oxford, pp 107–155
Kramer CM, Prata RTN, Willits MG, De Luca V, Steffens JC, Graser G
(2003) Cloning and regiospecificity studies of two flavonoid glucos-
yltransferases from Allium cepa. Phytochemistry 64:1069–1076
Langcake P, Pryce RJ (1977) A new class of phytoalexins from
grapevine. Experientia 33:151–152
Leifert W, Abeywardena M (2008) Cardioprotective actions of grape
polyphenols. Nutr Res 28:729–737
Li Y, Baldauf S, Lim EK, Bowles DJ (2001) Phylogenetic analysis of
the UDP-glycosyltransferase multigene family of Arabidopsisthaliana. J Biol Chem 276:4338–4343
Lim E, Doucet CJ, Li Y, Elias L, Worrall D, Spencer SP, Ross J,
Bowles DJ (2002) The activity of Arabidopsis glycosyltransfer-
ases toward salicylic acid, 4-hydroxybenzoic acid and other
benzoates. J Biol Chem 277:586–592
Lim E, Baldauf S, Li Y, Elias L, Worrall D, Spencer SP, Jackson RG,
Taguchi G, Ross J, Bowles DJ (2003) Evolution of substrate
recognition across a multigene family of glycosyltransferases in
Arabidopsis. Glycobiology 13:139–145
Lim E, Doucet C, Hou B, Jackson R, Abrams S, Bowles D (2005)
Resolution of (?)-abscisic acid using an Arabidopsis glucosyl-
transferase. Tetrahedron Assymetr 16:143–147
Loutre C, Dixon DP, Brazier M, Slater M, Cole DJ, Edwards R (2003)
Isolation of a glucosyltransferase from Arabidopsis thalianaactive in the metabolism of the persistent pollutant 3, 4-dichlo-
roaniline. Plant J 34:485–493
Meßner B, Thulke O, Schaffner AR (2003) Arabidopsis glucos-
yltransferases with activities toward both endogenous and
xenobiotic substrates. Planta 217:138–146
Moore MO, Giannasi DE (1994) Foliar flavonoids of eastern North
American Vitis (Vitaceae) north of Mexico. Plant Syst Evol
193:21–36
Munoz-Espada A, Wood K, Bordelon B, Watkins B (2004) Antho-
cyanin quantification and radical scavenging capacity of Con-
cord, Norton, and Marechal Foch grapes and wines. J Agr Food
Chem 52:6779–6786
Planta (2011) 234:1201–1214 1213
123

Nagashima S, Tomo S, Orihara Y, Yoshikawa T (2004) Cloning and
characterization of glucosyltransferase cDNA from Eucalyptusperriniana cultured cells. Plant Biotech 21:343–348
Noguchi A, Saito A, Homma Y, Nakao M, Sasaki N, Nishino T,
Takahashi S, Nakayama T (2007) A UDP-glucose: isoflavone
7-O-glucosyltransferase from the roots of soybean (Glycine max)
seedlings. J Biol Chem 282:23581–23590
Ogata J, Kanno Y, Itoh Y, Tsugawa H, Suzuki M (2005) Plant
biochemistry: anthocyanin biosynthesis in roses. Nature
435:757–758
Ono E, Homma Y, Horikawa M, Kunikane-Doi S, Imai H, Takahashi
S, Kawai Y, Ishiguro M, Fukui Y, Nakayama T (2010)
Functional differentiation of the glycosyltransferases that con-
tribute to the chemical diversity of bioactive flavonol glycosides
in grapevines (Vitis vinifera). Plant Cell 22:2856–2871
Park HJ, Cha HC (2003) Flavonoids from leaves and exocarps of the
grape Kyoho. Korean J Biol Sci 7:327–330
Polaskova P, Herszage J, Ebeler S (2008) Wine flavor: chemistry in a
glass. Chem Soc Rev 37:2478–2489
Schneider E, von der Heydt H, Esperester A (2008) Evaluation of
polyphenol composition in red leaves from different varieties of
Vitis vinifera. Plant Med 74:565–572
Taguchi G, Ubukata T, Nobuaki H, Yamamoto H, Okazaki M (2003)
Cloning and characterization of a glucosyltransferase that reacts
on the 7-hydroxyl group of flavonol and the 3-hydroxyl group
of coumarin from tobacco cells. Arch Biochem Biophys
420:95–102
Tian L, Blount JW, Dixon RA (2006) Phenylpropanoid glycosyl-
transferases from osage orange (Maclura pomifera) fruit. FEBS
Lett 580:6915–6920
Tohge T, Nishiyama Y, Hirai MY, Yano M, Nakajima J, Awazuhara
M, Inoue E, Takahashi H, Goodenow DB, Kitayama M, Noji M,
Yamazaki M, Saito K (2005) Functional genomics by integrated
analysis of metabolome and transcriptome of Arabidopsisplants over-expressing an MYB transcription factor. Plant J
42:218–235
Vogt T, Jones P (2000) Glycosyltransferases in plant natural product
synthesis: characterization of a supergene family. Trends Plant
Sci 5:380–388
Vogt T, Zimmermann E, Grimm R, Meyer M, Strack D (1997) Are
the characteristics of betanidin glucosyltransferases from cell-
suspension cultures of Dorotheanthus bellidiformis indicative of
their phylogenetic relationship with flavonoid glucosyltransfer-
ase? Planta 203:349–361
Vogt T, Grimm R, Strack D (1999) Cloning and expression of a
cDNA encoding betanidin 5-O-glucosyltransferase, a betanidin-
and flavonoid-specific enzyme with high homology to inducible
glucosyltransferases from the Solanaceae. Plant J 19:509–519
Willits MG, Giovanni M, Prata RTN, Kramer CM, De Luca V,
Steffens JC, Graser G (2004) Bio-fermentation of modified
flavonoids: an example of in vivo diversification of secondary
metabolites. Phytochemistry 65:31–41
Wu X, Prior RL (2005) Systematic identification and characterization of
anthocyanins by HPLC–ESI–MS/MS in common foods in the
United States: fruits and berries. J Agric Food Chem 53:2589–2599
Yamazaki M, Yamagishi E, Gong Z, Fukuchi-Mizutani M, Fukui Y,
Tanaka Y, Kusumi T, Yamaguchi M, Saito K (2002) Two
flavonoid glucosyltransferases from Petunia hybrida: molecular
cloning, biochemical properties and developmentally regulated
expression. Plant Mol Biol 48:401–411
1214 Planta (2011) 234:1201–1214
123







![New Reported Flavonol Characterized by NMR from the Petals ... · IR and MS of both flavonol glycosides do not allow differentiate the structures of the last mentioned compounds [10]](https://img.pdfslide.us/doc/110x75/5c9b590e09d3f28d6a8bb242/new-reported-flavonol-characterized-by-nmr-from-the-petals-ir-and-ms-of.jpg)





















