Embed Size (px)
Citation preview

Journal of Medical Virology 81:826–835 (2009)
Molecular Characterization and Functional Analysisof Occult Hepatitis B Virus Infection in ChinesePatients Infected With Genotype C
Yong Fang,1 Xu Teng,1 Wei-Zhen Xu,1 Di Li,1 Hong-Wei Zhao,2 Li-Juan Fu,2 Feng-Min Zhang,1
and Hong-Xi Gu1*1Department of Microbiology, Harbin Medical University, Harbin, China2Department of Infectious Diseases, Heilongjiang Provincial Hospital, Harbin, China
Occult HBV infection is defined as the persistenceof HBV DNA in individuals negative for HBVsurface antigen (HBsAg), and many differentmechanisms have been reported in differentcountries. However, in China, one of the endemicareas for HBV infection, no reports have beenpublished on occult HBV infection. The presentstudy investigated the virological features andthe mechanism of occult HBV infection in China.Full-length HBV DNA from eight patients withoccult HBV infection (S1–S8) and three HBsAg-positive cases (SWT1–SWT3) was cloned andsequenced. Additionally, four entire linear HBVgenomes from occult cases were transfectedtransiently into HepG2 cells. The sequencingresults showed two major mutations in patientswith occult HBV infection as follows: deletions inthe pre-S1 (S3, S4, and S7) and X (S1, S2, and S5)regions. Such deletions covered the S promoterand the basal core promoter (BCP), and functionanalysis of these variants also showed a decreasein DNA replication and antigen expression. Twopatients with occult infection (S6 and S8) had nomutations capable of interfering with viral repli-cation and gene expression in the major viralpopulation. Thus, the deletions in the S promoterand the BCP regions that disable the regulatoryelements may be the reason for the absence ofHBsAg, and multiple mechanisms may beresponsible for occult HBV infection. J. Med.Virol. 81:826–835, 2009. � 2009 Wiley-Liss, Inc.
KEY WORDS: hepatitis B virus; occult infec-tion; mutation
INTRODUCTION
Occult HBV infection is defined as the presence ofHBV DNA in blood or liver, in the absence of detectableserum HBV surface antigen (HBsAg), and has beenclassified into seropositive and seronegative infection
depending on the presence for antibody to the HBV coreantigen (anti-HBc) and/or HBsAg (anti-HBs) [Brechotet al., 2001; Conjeevaram and Lok, 2001]. Occult HBVinfection has its own risks of disease transmission andmay contribute to acute exacerbation and developmentof HBV-associated disease such as hepatic inflamma-tion, cirrhosis, and hepatocellular carcinoma (HCC)[Pollicino et al., 2004; Chevrier et al., 2007; Kannangaiet al., 2007]. However, the virological features and themechanism of occult HBV infection have not beenelucidated. Many earlier studies have supported theview that rearrangements in the S gene lead to synthesisof an antigenically modified S protein undetectable bythe commercially available HBsAg assays [Hou et al.,1995; Minuk et al., 2005], but research on HBVvariability has generated conflicting results. Recently,a few studies have extended sequence analysis to thefull-length viral genome, to detect mutations outside theS gene capable of interfering with its expression and/orwith viral replication, but with differencing results[Chaudhuri et al., 2004; Pollicino et al., 2007; Zahn et al.,2008], and regional differences were identified. In Chinathere have been no related reports. The present studyattempted to amplify the full-length HBV genomeisolated from Chinese patients, and examined the viralgenomic variability of isolates, and then evaluated
Additional Supporting Information may be found in the onlineversion of this article.
Yong Fang and Xu Teng contributed equally in this work.
Grant sponsor: Ph.D. Foundation of Education Ministry, China;Grant number: 20050226002; Grant sponsor: Doctor Foundationof Harbin Medical University, China; Grant sponsor: YouthFoundation of Heilongjiang Province, China; Grant number:QC060061.
*Correspondence to: Hong-Xi Gu, Department of Microbiology,Harbin Medical University, 157 Baojian Road, Nangang District,Harbin, Heilongjiang Province 150081, China.E-mail: [email protected]
Accepted 13 January 2009
DOI 10.1002/jmv.21463
Published online in Wiley InterScience(www.interscience.wiley.com)
� 2009 WILEY-LISS, INC.

replication capacity and antigen expression of typicaloccult HBV isolates in vitro.
MATERIALS AND METHODS
Patients and Clinical Data
Between January 2006 and December 2008, theprevalence of occult HBV infection in a group of patientswith chronic liver disease without any known cause(such as alcohol abuse, hepatotoxic drug, Wilson’sdisease, biliary tract disease, venous outflow obstruc-tion, iron overload, or autoimmune disease) was inves-tigated. None of the patients had been treated withantiviral or immunosuppressive drugs and none wereco-infected with HIV, CMV, HCV, HDV, HAV, or HEV.The diagnosis of chronic liver disease was made byconventional clinical, biochemical, imaging, and endo-scopic criteria. Cryptogenic chronic liver disease wasdiagnosed when all known identifiable causes wereexcluded by relevant investigations.
Occult HBV infection in patients with cryptogenicliver disease was diagnosed when PCR detected HBVDNA in sera for at least two different regions in theabsence of detectable HBsAg [Conjeevaram and Lok,2001]. Patients with occult HBV infection were includedin the study. Serum samples were selected at randomfrom the patients with occult HBV infection, forcomplete genome analysis. The three HBsAg-positivepatients that served as a control group were diagnosedwith chronic hepatitis B. This study was approved by theHarbin Medical University Committee on ClinicalInvestigation and informed consent was obtained fromall the patients.
Serological Assays
Blood samples were centrifuged at 700g and serumsamples were used for serological tests within 4 hr, andthe remainder of the same serum samples were stored at�808C. HBsAg, anti-HBs, HBV e antigen (HBeAg), anti-HBe, anti-HBc and anti-HCV were measured by ELISAusing a diagnostic kit (Kehua Biotechnology, Shanghai,China). According to the instructions, the sensitivity ofthe assay for HBsAg was 0.05 ng/ml.
Sera from patients negative for all the above testswere investigated for the presence of HBV DNA by PCR.Patients who were negative for HBsAg and othercausative factors implicated in the cause of chronic liverdisease, but positive for HBV DNA by PCR, were labeledas occult HBV infection as per the diagnostic criteriadescribed earlier [Conjeevaram and Lok, 2001]. Serumsamples from these patients were retested for thepresence of HBsAg by another commercial kit (WantaiBiotechnology, Beijing, China) to confirm the absence ofHBsAg in the serum.
Isolation and Detection of HBV DNA From Serum
Viral DNA was extracted from 250 ml of eachserum sample by the AxyPrep kit (Axygen Biosciences,
Hangzhou, China) according to the manufacturer’sinstructions.
For the HBsAg-negative cases, HBV genome detec-tion was performed by nested PCR using three primersets, each specific for preS–S, precore–core, and X viralgenomic regions, respectively [Goshi et al., 2000]. Withthis approach, only the cases in whom HBV DNA wasdetected using at least two different sets of primers werediagnosed as occult HBV infection and considered foramplification of the full-length genomes.
In order to prevent carryover contamination duringPCR, each step of the procedure was performed in aseparate room with dedicated equipment and direc-tional flow from the beginning of the procedure tothe end. All samples were tested in duplicate. Controlsamples included normal sera, HBV-positive sera, andnegative controls. Positive and negative controls wereincluded in the extraction steps and in both rounds ofamplification.
Amplification of Full-LengthHBV Genomes
HBV genomes were full-length amplified using differ-ent PCR methods. HBV sequences from HBsAg-positivepatients were obtained by one-step full-length genomeamplification using a single set of primers: sense primerP1 (50-CCGGAAAGCTTGAGCTCTTCTTTTTCACCTC-TGCCTAATCA-30 1821–1841) and the antisense primerP2 (50-CCGGAAAGCTTGAGCTCTTCAAAAAGTTGC-ATGGTGCTGG-30 1806–1825), as in the method usedby Gunther et al. [1995]. On the other hand, HBV DNAfrom patients with occult HBV infection was full-lengthamplified by a two-step PCR approach. The first-roundPCR yield was very faint on 0.7% agarose gel afterelectrophoresis using the primers P1/P2 to amplify theentire genome from occult HBV cases. Therefore,the second-round PCR was performed with differentprimer pairs: sense primer P3 (50-GAGCTCTTCTTTTT-CACCTCTGCCTAATCA-30 1821–1841) and the anti-sense primer P4 (50-GAGCTCTTCAAAAAGTTGCATG-GTGCTGG-30 1806–1825). The nucleotide sequences ofprimers P3 and P4 differed from those of primers P1 andP2 because of the absence of the heterologous nucleo-tides that contained the HindIII restriction enzyme site.The first-round PCR contained 5 ml 10�LA PCR buffer,400 mM each dNTP, 10 mM each primer, 2.5 U LA Taqpolymerase (Takara Biotechnology, Dalian, China) and10 ml template DNA preparation per 50 ml reaction.Amplification was performed for 40 cycles with denatu-ration at 948C for 1 min, annealing at 608C for 1 min,and elongation at 688C for 3 min, with an increment of1 min after 10 cycles; cycles were followed by an extraincubation at 728C for 10 min to ensure full extension ofthe products. Subsequently, 5 ml (10%) of the first PCRproduct was re-amplified for 30 cycles in 50-ml assaymixtures with primer set P3 and P4. The second-roundPCR conditions were similar to those of the first-roundPCR, except annealing was done at 658C.
J. Med. Virol. DOI 10.1002/jmv
Molecular Characterization of Occult HBV 827

Cloning and Sequencing of theAmplified HBV Genome
The amplicons were recovered from agarose gel withthe AxyPrep DNA Gel Extraction Kit (Axygen Bio-sciences) and cloned into the pMD18-T Vector (TakaraBiotechnology), following the manufacturers’ instruc-tions. For each sample, three to four clones weresequenced by an automated DNA sequencer (ABI3100-Avant Genetic Analyze; Applied Biosystems,Carlsbad, CA) with a set of primers that encompassedthe whole HBV genome plus universal M13 primers.HBV primers are described in detail in SupplementaryTable SI.
Nucleotide and Amino Acid Analysis
Nucleotide and amino acid sequences of HBV DNAisolates from patients with occult HBV infection andHBsAg-positive controls were aligned with 50 HBVsequences from GenBank using the CLUSTAL Wprogram. The conventional EcoRI position was takenas nucleotide position 1. Genotyping of HBV DNAsequences was performed by phylogenetic comparisonwith the representative genotypes (HBVADW2 X02763,genotype A) [Valenzuela et al., 1980], HPBADW1(D00329, genotype B), HPBADRA (M12906, genotype C),XXHEPAV (X02496, genotype D), HHVBE4 (X75664,genotype E), and HHVBF (X75663, genotype F) over theentire genome.
Phylogenetic and molecular evolutionary analyseswere conducted using MEGA version 3.1 [Kumar et al.,2004] by the neighbor-joining algorithm based onKimura two-parameter distance estimation.
Cell Culture and Transfection
HepG2 cells were cultured in Dulbecco’s modifiedEagle’s medium, supplemented with 10% fetal calfserum, penicillin (100 U/ml) and streptomycin (100 mg/ml) at 378C in a humidified incubator with 5% CO2. Fourdifferent constructs (S1.3, S3.3, S4.1, and S7.2) thatcontained full-length HBV genomes isolated from occultHBV patients S1, S3, S4, and S7, respectively, were usedto perform transient transfection. Each isolate repre-sented the dominant strain of the HBV population thatinfected the liver of each patient. Linear HBV monomerswere released from S1.3, S3.3, S4.1, and S7.2 constructsand from the wild-type (SWT3.3) control plasmid bycleavage with SapI restriction enzyme (New EnglandBiolabs, Ipswich, MA). Full-length HBV genome DNA(4 mg) from each clone was used to transfect HepG2 cellsin six-well culture plates using Lipofection 2000 reagent(Invitrogen, Carlsbad, CA) following the manufacturer’sguidelines. Triple plates were used for all samples, and1 mg reporter plasmid expressing enhanced greenfluorescence protein was co-transfected into each cul-ture as an internal control to normalize the transfectionefficiency between plates. Cells were collected 72 hrafter transfection.
Real-Time PCR Analysis of HBV DNA
HBV DNA was extracted from the intracellular andextracellular core particles using Axygen DNA Mini kit(Axygen Biosciences) at 72 hr post-transfection, andthen assayed by real-time PCR using a DiagnosticsLightCycler (Roche, Mannheim, Germany), followingthe manufacturer’s guidelines. According to the instruc-tions, an HBV DNA level �5.0�102 copies/ml (2.70 logcopies/ml) was considered as a positive response.
HBsAg and HBeAg Assays
At 72 hr post-transfection, the culture supernatantwas collected, centrifuged at 1,000g for 5 min to removecellular debris, and transferred to a clean tube forfurther analysis. The expression levels of HBsAg andHBeAg were separately assayed using an enzymeimmunoassay kit (Kehua Biology, Shanghai, China).According to the instructions, a ratio of sample/negative(S/N)�2.1 was considered as a positive response toHBsAg or HBeAg.
Statistical Analysis
Data were expressed as mean�SD. Data werecompared using the F test and one-way ANOVA. Allstatistical analysis was performed using SPSS 12.0software (SPSS, Inc., Chicago, IL). P<0.05 was consid-ered statistically significant.
RESULTS
Patients
During the period 2006 to 2008, 159 patients werediagnosed with cryptogenic chronic liver disease, all ofwhom had chronic hepatitis. HBV DNA was detected in45 (28.3%) of these patients. HCV RNA was negative inall of the patients. Seventy-five of the 159 patients,including the 45 HBV-DNA-positive cryptogenic chronicliver disease patients, were positive for anti-HBc IgG.Anti-HBe was not detected in any of these patients. The45 patients positive for anti-HBc IgG and HBV DNAwere categorized as having occult HBV infection. Serafrom eight of patients with occult HBV infection wereselected randomly for analysis of complete HBV genome(S1–S8). The sera of these eight patients were testedfor the presence of anti-HBsAg by using a commercialELISA kit (Kehua Biotechnology) and were found tobe negative for anti-HBsAg. Sera from three HBsAg-positive patients with confirmed chronic hepatitis wereused as controls (SWT1–SWT3). Complete genomicanalysis of the HBV DNA was also taken in the controlgroup for comparison with any mutation. The clinicaldata of these patients are shown in Table I. The‘‘infection history’’ means the length of HBV infection,and the beginning of infection was defined as the firsttime for finding abnormal liver function in patients withcryptogenic chronic liver disease, or HBsAg in chronicHBV infecion patients.
J. Med. Virol. DOI 10.1002/jmv
828 Fang et al.

The HBV full-length genomes from eight patients withoccult HBV infection were amplified and cloned success-fully, and three or four clones from each patientwere sequenced. For the HBsAg-positive patients,three clones of each were sequenced. All 11 patientswere infected with HBV genotype C strains, which is thepredominant genotype in China (Genebank Accession-Numbers: S1–S8 EU916204–EU916232; SWT1–SWT3EU916233–EU916241) (Fig. 1).
Analysis of the Pre-S–S Genomic Region
In the pre-S1 region, long-stretch deletions of aa 58–118 (nt 3019–3201) were observed in most isolates fromthree patients (S3, S4, and S7) with occult HBV infection(Fig. 2). In addition, patients S4.4 and S7.4 presentedwith deletions of aa 1–6, which abolished the pre-S1start codon. The B-cell antigenic epitope in pre-S1protein (aa 19–26) [Maeng et al., 2000] was conserved.Within the pre-S2 region, the start codon was abolishedin three isolates (S2.3, S7.1, and S7.2) by a pointmutation (ATG!ATA), and the mutation of codon 3introducing a stop signal terminated the pre-S2 gene inall of the isolates from patient S3 (Fig. 2).
The analysis of the S region showed that threevariants (S4.2, S4.3, and S4.4) of patient S4 had threeamino acid changes within the putative human leuko-cyte antigen class I-restricted cytotoxic T lymphocyte(CTL) epitope (aa 28–51) of HBsAg [Tai et al., 1997].Patients S1, S2, S5, S7, and S8 only had one variant withone or two amino acid changes, and the other patientshad no mutations (Table II). Substitutions within themajor hydrophilic region of HBsAg (aa 104–172) werefound in most patients (one or two amino acids perpatient), except patients S4 and S8 (Table II). Theimmunodominant a determinant (aa 124–147) [Feitel-son et al., 1981] of HBsAg had no variation in any of theisolates, except variants S2.2 (A128V), S 2.3 (I126T), andS4.4 (I126T), all the substitutions were within the firsthydrophilic loop of theadeterminant (Table II). The stopcodon mutations in variant S1.1, S7.4, and S8.1 wereobserved in the coding sequence of HBsAg.
Analysis of the Pre-C–C Genomic Region
Within the pre-core region, the nucleotide mutationG1896A, introducing a stop signal at codon 28 andpreventing HBeAg synthesis, was found in five isolates(S3.1, S3.3, S3.4, S4.2, and S7.1), and the rest of theisolates had a wild-type pre-core region.
Within the core region, amino acid changes weredistributed at random, and different immunogenicantigenic epitopes were observed. Three different aminoacid changes (A58V, I59S, and L60V) were detectedwithin the CD4þT-cell epitope (aa 48–69) [Ferrariet al., 1991], but only a single amino acid substitution(L60V) was present in most of the HBV isolates.Concerning the core protein CD8þCTL epitopes, onlyvariant S7.3 had three amino acid substitutions (P20A,F23L, and I27V) within the epitope 18–27. CTL epitope141–151 was conserved in all isolates, except variants
J. Med. Virol. DOI 10.1002/jmv
TA
BL
EI.
Cli
nic
al
Man
ifes
tati
ons
an
dL
abor
ato
ryD
ata
ofO
ccu
ltIn
fect
ion
Pati
ents
(S1
–S
8)
an
dH
BsA
g-P
osit
ive
Case
s(S
WT
1-S
WT
3)
Pati
ents
(IU
/L)a
Age
(yea
rs)
Sex
(F/M
)D
iagn
osis
Vir
al
DN
Alo
ad
(cop
ies/
ml)
Ala
nin
etr
an
sam
inase
Asp
art
ate
tran
sam
inase
(IU
/L)a
HB
sAg
An
ti-H
Be
An
ti-H
Bs
HB
eAg
An
ti-H
Bs
Infe
ctio
nh
isto
ry(y
ears
)
S1
59
MC
H6.4
15�
10
4169
115
��
þ�
�2
S2
63
MC
H2.9
72�
10
5210
232
��
þ�
�2
S3
52
MC
H9.3
47�
10
5149
128
��
þ�
�1
S4
56
FC
H2.5
53�
10
4186
160
��
þ�
�1
S5
65
FC
H4.2
38�
10
4170
153
��
þ�
�2
S6
65
MC
H3.0
74�
10
5205
203
��
þ�
�1
S7
66
FC
H6.7
62�
10
4230
195
��
þ�
�2
S8
72
MC
H3.9
62�
10
5173
169
��
þ�
�1
SW
T1
58
MC
H6.8
95�
10
7150
115
þ�
þþ
�5
SW
T2
49
MC
H7.8
82�
10
7189
201
þ�
þþ
�4
SW
T3
63
MC
H8.7
00�
10
7175
198
þ�
þ�
þ6
CH
,ch
ron
ich
epati
tis.
aA
lan
ine
tran
sam
inase
(AL
T)
an
dasp
art
ate
tran
sam
inase
(AS
T)
nor
mal
valu
es:0
–40
IU/L
;�
,n
egati
ve;
þ,p
osit
ive.
Molecular Characterization of Occult HBV 829

S4.1 (E145G) and S5.2 (V148I). Two point mutationswere found in the arginine-rich motif of the C-terminaldomain of the core protein of variants S5.4 (R164C)and S7.2 (R167W). In addition, long-fragment deletionmutations were found in patients with occult infection,S1 (S1.1, aa 81–109; and S1.2, aa 78–106) and S5 (S5.1,aa 65–104; S5.3 and S5.4, aa 84–116), which resulted ina truncated core protein.
Analysis of the Pol Genomic Region
In the terminal protein domain of the polymerase, notypical mutation that may induce occult infection wasfound. The internal deletions of the pre-S1 regioncoincided with the spacer region of the polymerase gene,which was tolerant to deletions and substitutions. TheL180M substitution inside the conserved YMDD motif of
J. Med. Virol. DOI 10.1002/jmv
Fig. 1. Phylogenetic analysis of variants of S1–S8 and SWT1–SWT3 with genotype C isolates fromGenebank.
830 Fang et al.

the reverse transcriptase domain of the Pol gene wasfound only in variant S7.3. This mutation is usuallyselected during lamivudine treatment, but none of thepatients had been treated previously with antivirals.The RNase H domain was conserved in all isolates.
Analysis of the X Genomic Region
The typical change in the X region was a deletionmutation, the deletion of aa 126–133/134 were observedin patient S2, and deletion of aa 127–133 was observedin patients S1 and S5. Double nucleotide mutations at
positions 1762/1764 that resulted in amino acid changesK130M/V131I were found in variants S5.1 and S5.4.
Analysis of Transcription Regulatory Regions
The main regulatory elements were also analyzed inall viral isolates from the 11 studied patients to detectmutations potentially implicated in the inhibition ofviral replication and/or gene expression. The pre-S1promoter had a wild-type sequence in all but twopatients (S1 and S8), whereas the same 183 nt deletions(nt 3019–3201) were detected in the S promoter region
J. Med. Virol. DOI 10.1002/jmv
Fig. 2. Schematic representation showing types of variations in the pre-S1–S2 region affecting the Spromoter. Top, the essential S-promoter region (&), transcription factor binding sites (&), start sites of thepre S1/S2/S ATG (*). Regions corresponding to wild-type in all variants are shown by a continuous line,deletions are shown by a textured pattern box, and point mutation are shown by triangle.
TABLE II. Amino Acid Changes in HBV S Protein in Isolates S1–S8
Patientisolate
Upstream changes a determinant Downsteam changes
1–27CTL epitope
28–51 52–103MHR
104–1231st loop124–138
2st loop139–147
MHR148–172 173–226
S1 S34W L94S P111L R160G W182stop;Y225C
S2 L21S; R24K Q51P I126T; A128V S154PS3 N3Sa D99G T116A V168AS4 G44Ea; P46Ha;
T47Va; Q51RQ101R I126T
S5 I28T L77Q T1231S6 E164D;
W165RL205P
S7 N3S; R24K L42stop; G44E W165L F220LS8 E2R; N3H; I25V A45T; P46T D33stop
aMutant at this position exists as a majority population in all the clones sequenced of the particular isolate.
Molecular Characterization of Occult HBV 831

in three occult infection patients (S3, S4, and S7) (Fig. 2).The deletion covered the CCAAT element, which isknown to bind nuclear factor-g transcription factorand is necessary for S promoter activity. The analysisof enhancer I/X promoter indicated only several pointmutations, but three occult infection patients (S3, S4,and S6) showed the same mutation (A951T/G) locatedwithin the C/EBP binding site of enhancer I. EnhancerII analysis showed only one unique point mutationG1661A (located in the HNF4-binding site) was found inpatient S1 (Fig. 3). Analysis of the basal core promoter(BCP) showed the presence of deletion mutations inoccult infection patients S1, S2, and S5 (Fig. 3). Thedouble A1762T and G1764A nucleotide exchanges,which have been reported to cause a decrease in HBeAgexpression and enhanced viral replication, were foundin isolates S5.1 and S5.4.
Functional Analysis of Occult HBV Genomes
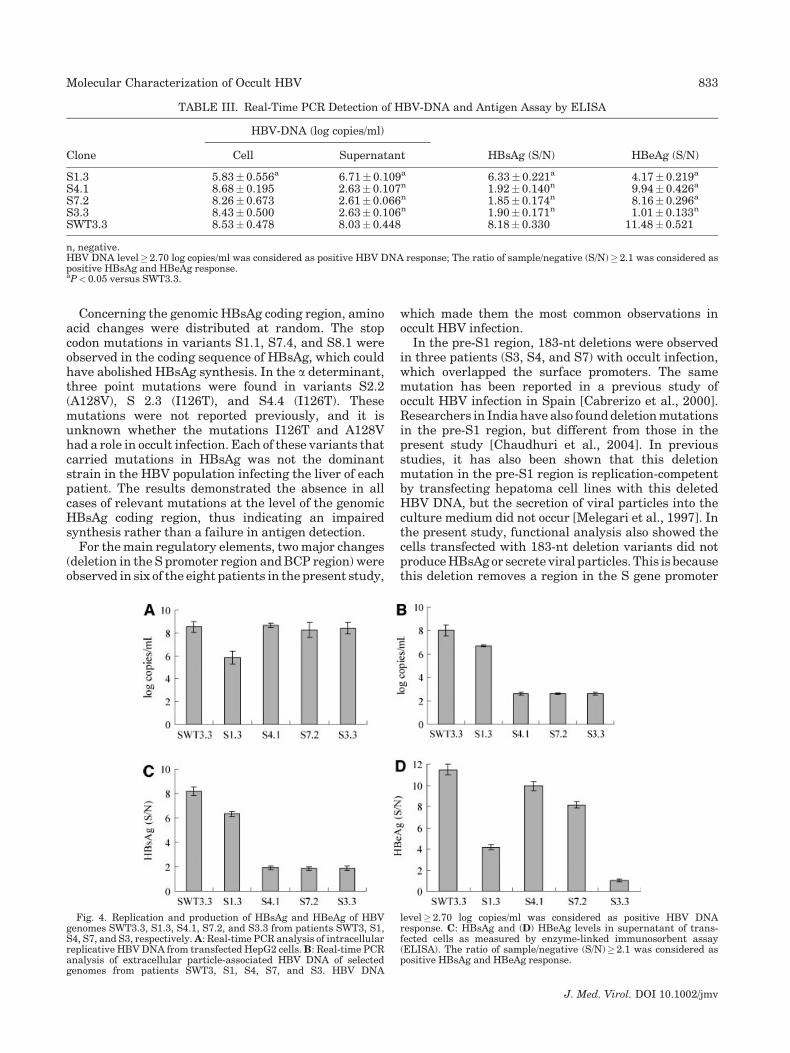
Frequent deletions were observed in the BCP region(patients S1, S2, and S5) and in the S promoter region(patients S3, S4, and S7), compared with wild-typeisolates. To investigate the possibility of altered generegulation caused by these altered promoter regions,functional assays of these mutants were performed, andwild-type isolate (SWT3.3) was used as control. S1, S2,and S5 had a similar deletion, and therefore, a func-tional assay of S1.3 isolate was performed. S4.1 only hada deletion mutation in the S promoter, and S7.2 carriedthe preS2 start codon mutation and deletion in theS promoter. S3.3 had both the pre-core stop codonmutation and the stop codon mutation in the pre-S2region, and the deletion in the S promoter region. At72 hr post-transfection, HBV DNA was isolated from cellsupernatants and cell lysates, and real-time PCR wasperformed. As shown in Table III and Figure 4A, exceptvariant S1.3 (P<0.05), HBV-DNA expression levels ofthe other three HBVs were similar to those of wild-type
strains SWT3.3 in HepG2 cells. In the culture super-natants, HBV DNA was not found in mutant HBVwith S promoter deletions, and in variant S1.3, it wassignificantly lower than that in wild-type strain SWT3.3(Table III and Fig. 4B). Cells transfected withS-promoter-deleted variant genomes (S4.1, S7.2, andS3.3) were negative for HBsAg secretion, and HBsAgsecretion from S1.3 was significantly lower than thatfrom wild-type HBV-transfected cells (P<0.05)(Table III and Fig. 4C). HBeAg expression levels for allvariant genomes were lower than those of the controls(P<0.05), and HBeAg was negative in the supernatantof S3.3-transfected cells because S3.3 carried the pre-core stop codon (Table III and Fig. 4D).
DISCUSSION
Occult HBV infection is a problem in many parts of theworld and affects blood banks, the diagnosis of chronichepatitis, and immunization. In the present study,multiple clones of full-length HBV genomes wereobtained from eight patients with occult HBV infectionwith genotype C and compared with the wild-type HBVgenome prevalent in China. The patients were nottreated with antiviral drugs or co-infected with otherviruses such as HCV and HIV, which can be a feature ofHBV infection. There were two possible explanations forthe HBsAg-negative HBV infection: the virus producedan antigenically modified S protein undetectable byroutine tests [Yamamoto et al., 1994; Chiou et al., 1997;Jeantet et al., 2004]; or the virus carried mutationscapable of inhibiting S gene expression and/or viralreplication [Blum et al., 1991; Pollicino et al., 2007].In the present study, entire viral genomes from eightoccult HBV carriers were examined and comparedwith isolates from HBsAg-positive individuals. Then,the capacity of replication and antigen expression ofsome occult isolates were verified in vitro.
J. Med. Virol. DOI 10.1002/jmv
Fig. 3. Schematic representation showing types of variations in the enhancer II and BCP region affectingthe basal core promoter. Top, the essential S-promoter region (&), transcription factor binding sites (&).Regions corresponding to wild-type in all variants are shown by a continuous line, deletions are shown by atextured pattern box, and point mutation are shown by triangle.
832 Fang et al.
Concerning the genomic HBsAg coding region, aminoacid changes were distributed at random. The stopcodon mutations in variants S1.1, S7.4, and S8.1 wereobserved in the coding sequence of HBsAg, which couldhave abolished HBsAg synthesis. In the a determinant,three point mutations were found in variants S2.2(A128V), S 2.3 (I126T), and S4.4 (I126T). Thesemutations were not reported previously, and it isunknown whether the mutations I126T and A128Vhad a role in occult infection. Each of these variants thatcarried mutations in HBsAg was not the dominantstrain in the HBV population infecting the liver of eachpatient. The results demonstrated the absence in allcases of relevant mutations at the level of the genomicHBsAg coding region, thus indicating an impairedsynthesis rather than a failure in antigen detection.
For the main regulatory elements, two major changes(deletion in the S promoter region and BCP region) wereobserved in six of the eight patients in the present study,
which made them the most common observations inoccult HBV infection.
In the pre-S1 region, 183-nt deletions were observedin three patients (S3, S4, and S7) with occult infection,which overlapped the surface promoters. The samemutation has been reported in a previous study ofoccult HBV infection in Spain [Cabrerizo et al., 2000].Researchers in India have also found deletion mutationsin the pre-S1 region, but different from those in thepresent study [Chaudhuri et al., 2004]. In previousstudies, it has also been shown that this deletionmutation in the pre-S1 region is replication-competentby transfecting hepatoma cell lines with this deletedHBV DNA, but the secretion of viral particles into theculture medium did not occur [Melegari et al., 1997]. Inthe present study, functional analysis also showed thecells transfected with 183-nt deletion variants did notproduce HBsAg or secrete viral particles. This is becausethis deletion removes a region in the S gene promoter
J. Med. Virol. DOI 10.1002/jmv
TABLE III. Real-Time PCR Detection of HBV-DNA and Antigen Assay by ELISA
Clone
HBV-DNA (log copies/ml)
HBsAg (S/N) HBeAg (S/N)Cell Supernatant
S1.3 5.83� 0.556a 6.71� 0.109a 6.33� 0.221a 4.17� 0.219a
S4.1 8.68� 0.195 2.63� 0.107n 1.92� 0.140n 9.94� 0.426a
S7.2 8.26� 0.673 2.61� 0.066n 1.85� 0.174n 8.16� 0.296a
S3.3 8.43� 0.500 2.63� 0.106n 1.90� 0.171n 1.01� 0.133n
SWT3.3 8.53� 0.478 8.03� 0.448 8.18� 0.330 11.48� 0.521
n, negative.HBV DNA level� 2.70 log copies/ml was considered as positive HBV DNA response; The ratio of sample/negative (S/N)� 2.1 was considered aspositive HBsAg and HBeAg response.aP< 0.05 versus SWT3.3.
Fig. 4. Replication and production of HBsAg and HBeAg of HBVgenomes SWT3.3, S1.3, S4.1, S7.2, and S3.3 from patients SWT3, S1,S4, S7, and S3, respectively. A: Real-time PCR analysis of intracellularreplicative HBV DNA from transfected HepG2 cells. B: Real-time PCRanalysis of extracellular particle-associated HBV DNA of selectedgenomes from patients SWT3, S1, S4, S7, and S3. HBV DNA
level� 2.70 log copies/ml was considered as positive HBV DNAresponse. C: HBsAg and (D) HBeAg levels in supernatant of trans-fected cells as measured by enzyme-linked immunosorbent assay(ELISA). The ratio of sample/negative (S/N)� 2.1 was considered aspositive HBsAg and HBeAg response.
Molecular Characterization of Occult HBV 833

that is a binding site for the transcription factor, SP1,thus causing a drastic reduction in HBsAg synthesis.For assembly of the envelope particles, a specific ratio ofmajor (small) HBsAg and large HBsAg is required. Ifthis ratio is altered, it may alter the HBsAg assemblyand secretion [Bock et al., 1997; Gerner et al., 2003] incomparison with that of the wild phenotype, which leadsto a decrease in HBsAg and HBV in the serum. If thevariants are obtained from serum, this indicates that theHBsAg must be provided in trans by a wild-type virus forthe secretion of viral particles that contain mutantgenomes. In this sense, the deletion mutant may beaccompanied by the wild-type virus in the pre-S1 region.
Further deletion mutations were found in the BCPregion in three patients with occult infection, andfunctional analysis indicated that the cells transfectedwith the BCP deletion variant showed a decrease inDNA replication and antigen expression. The BCP wassufficient for accurate initiation of both pre-core mRNAand pregenomic (pg) RNA transcription. It contains fourAT-rich regions and the initiators for pre-core mRNAand pgRNA transcription. The AT-rich regions can bindTATA-binding protein, which is important for pre-coremRNA and pgRNA transcription. In the present survey,there were four deletion patterns of 18 or 20 bp, whichencompassed TA1, TA2, and TA3.One deletion pattern(nt 1753–1772) has been found in occult HBV in Japan[Saito et al., 1999]. These deletion mutations that resultin decreased HBV replication caused by a decrease inpgRNA may lead to a decrease in HBV load and HBsAgexpression in serum. At another level, all deletionswithin the BCP result in a frameshift mutation andtruncation of the X protein at its C-terminal end, whichinfluence the expression and function of the X protein.This usually affects the transactivating function of theX protein, and loss of this activity may be an additionalreason for the decreased viral replication and reducedantigen production seen in patients with occult infec-tion.
HBsAg detection by immunoassay is the mainstay ofdiagnosis of HBV infection. This is dependent on excessHBsAg in the blood. The regulatory region has much tocontribute towards this excess production, as does therelative quantity of the major and large HBsAg. Theanalysis of mutated promoters observed in the presentstudy showed a decrease in HBsAg expression andviral replication. However, there were occult infectionpatients who had no mutations capable of interferingwith viral replication and gene expression. Thus, itappears that the non-detectability of HBsAg in serummay arise from several mechanisms, caused by alter-ations in the structural, functional and regulatoryregions of the HBV genome. The host immune systemis likely to play a critical role, and future research shouldbe carried out.
REFERENCES
Blum H, Galun E, Liang TJ, von Weizsacker F, Wands JR. 1991.Naturally occurring missense mutation in the polymerase geneterminating hepatitis B virus replication. J Virol 65:1836–1842.
Bock CT, Tillmann HL, Maschek HJ, Manns MP, Trautwein C. 1997. ApreS mutation isolated from a patient with chronic hepatitis Binfection leads to virus retention and misassembly. Gastroenterol-ogy 113:1976–1982.
Brechot C, Thiers V, Kremsdorf D, Nalpas B, Pol S, Paterlini-Brechot P.2001. Persistent hepatitis B virus infection in subjects withouthepatitis B sur-face antigen: Clinically significant or purely ‘occult’?Hepatology 34:194–203.
Cabrerizo M, Bartolome J, Caramelo C, Barril G, Carreno V. 2000.Molecular analysis of hepatitis B virus DNA in serum andperipheral blood mononuclear cells from hepatitis B surfaceantigen-negative cases. Hepatology 32:116–123.
Chaudhuri V, Tayal R, Nayak B, Acharya SK, Panda SK. 2004. Occulthepatitis B virus infection in chronic liver disease: Full-lengthgenome and analysis of mutant surface promoter. Gastroenterology127:1356–1371.
Chevrier MC, St-Louis M, Perreault J, Caron B, Castilloux C, LarocheJ, Delage G. 2007. Detection and characterization of hepatitis Bvirus of anti-hepatitis B core antigen-reactive blood donors inQuebec with an in-house nucleic acid testing assay. Transfusion47:1794–1802.
Chiou HL, Lee TS, Kuo J, Mau YC, Ho MS. 1997. Altered antigenicity of‘a’ determinant variants of hepatitis B virus. J Gen Virol 78:2639–2645.
Conjeevaram HS, Lok AS. 2001. Occult hepatitis B virus infection: Ahidden menace. Hepatology 34:204–206.
Feitelson MA, Marion PL, Robinson WS. 1981. Antigenic andstructural relationships of the surface antigens of hepatitis B virus,ground squirrel hepatitis virus, and woodchuck hepatitis virus.J Virol 39:447–454.
Ferrari C, Bertoletti A, Penna A, Cavalli A, Valli A, Missale G, Pilli M,Fowler P, Giuberti T, Chisari FV. 1991. Identification of immuno-dominant T cell epitopes of the hepatitis B virus nucleocapsidantigen. J Clin Invest 88:214–222.
Gerner P, Schafer HM, Prange R, Pravitt D, Wirth S. 2003. Functionalanalysis of a rare HBV deletion mutant in chronically infectedchildren. Pediatr Res 53:891–897.
Gunther S, Li BC, Miska S, Kruger DH, Meisel H, Will H. 1995. A novelmethod for efficient amplification of whole hepatitis B virusgenomes permits rapid functional analysis and reveals deletionmutants in immunosuppressed patients. J Virol 69:5437–5444.
Hou J, Karayiannis P, Walters J, Luo K, Liang C, Thomas H. 1995. Aunique insertion in the S gene of surface antigen-negative hepatitisB virus Chinese carriers. Hepatology 21:273–278.
Jeantet D, Chemin I, Mandrand B, Tran A, Zoulim F, Merle P, Trepo C,Kay A. 2004. Cloning and expression of surface antigens from occultchronic hepatitis B virus infections and their recognition bycommercial detection assays. J Med Virol 73:508–515.
Kannangai R, Vivekanandan P, Netski D, Mehta S, Kirk GD, ThomasDL, Torbenson M. 2007. Liver enzyme flares and occult hepatitis Bin persons with chronic hepatitis C infection. J Clin Virol 39:101–105.
Kumar S, Tamura K, Nei M. 2004. MEG A3: Integrated software formolecular evolutionary genetics analysis and sequence alignment.Briefings Bioinformatics 5:150–163.
Maeng CY, Ryu CJ, Gripon P, Guguen-Guillouzo C, Hong HJ. 2000.Fine mapping of virus-neutralizing epitopes on hepatitis B virusPreS1. Virology 270:9–16.
Melegari M, Scaglioni PP, Wands JR. 1997. The small envelope proteinis required for secretion of a naturally occurring hepatitis B virusmutant with pre-S1 deleted. J Virol 71:5449–5454.
Minuk GY, Sun DF, Uhanova J, Zhang M, Caouette S, Nicolle LE,Gutkin A, Doucette K, Martin B, Giulivi A. 2005. Occult hepatitis Bvirus infection in a North American community-based population.J Hepatol 42:480–485.
Pollicino T, Squadrito G, Cerenzia G, Cacciola I, Raffa G, Craxi A,Farinati F, Missale G, Smedile A, Tiribelli C, Villa E, Raimondo G.2004. Hepatitis B virus maintains its pro-oncogenic properties inthe case of occult HBV infection. Gastroenterology 126:102–110.
Pollicino T, Raffa G, Costantino L, Lisa A, Campello C, Squadrito G,Levrero M, Raimondo G. 2007. Molecular and functional analysis ofoccult hepatitis B virus isolates from patients with hepatocellularcarcinoma. Hepatology 45:277–285.
Saito T, Shinzawa H, Uchida T, Kawamata O, Honma S, Watanabe H,Shao L, Saito K, Togashi H, Takahashi T. 1999. Quantitative DNAanalysis of low-level hepatitis B viremia in two patients with
J. Med. Virol. DOI 10.1002/jmv
834 Fang et al.

serologically negative chronic hepatitis B. J Med Virol 58:325–331.
Shiota G, Oyama K, Ungawa A, Tanaka K, Nomi T, Kitamura A,Tsutsumi A, Noguchi N, Takano Y, Yashima K, Kishimoto Y, SuouT, Kawasaki H. 2000. Occult hepatitis B virus infection in HBsantigen-negative hepatocellular carcinoma in a Japanese popula-tion: Involvement of HBx and p53. J Med Virol 62:151–158.
Tai P, Banik D, Lin G, Pai S, Pai K, Lin MH, Yuoh G, Che S, Hsu SH,Chen TC, Kuo TT, Lee CS, Yang CS, Shih C. 1997. Novel andfrequent mutations of hepatitis B virus coincide with a majorhistocompatibility complex class I-restricted T-cell epitope of thesurface antigen. J Virol 71:4852–4856.
Valenzuela P, Quiroga M, Zalvidar J, Gray P, Rutter W. 1980. Thenucleo-tide sequence of the hepatitis B viral genome and the identification ofthe major viral genes. In: Fields BN, Jaenisch R, Fox CF, editors.Animal virus genetics. New York: Academic Press. pp 57–70.
Yamamoto K, Horikita M, Tsuda F, Itoh K, Akahane Y, Yotsumoto S,Okamoto H, Miyakawa Y, Mayumi M. 1994. Naturally occurringescape mutants of hepatitis B virus with various mutations in the Sgene in carriers seropositive for antibody to hepatitis B surfaceantigen. J Virol 68:2671–2676.
Zahn A, Li C, Danso K, Candotti D, Owusu-Ofori S, Temple J, Allain JP.2008. Molecular characterization of occult hepatitis B virus ingenotype E-infected subjects. J Gen Virol 89:409–418.
J. Med. Virol. DOI 10.1002/jmv
Molecular Characterization of Occult HBV 835





























