Embed Size (px)
Citation preview

Molecular BiologyFourth Edition
Chapter 10
Eukaryotic RNA Polymerases and Their Promoters
Lecture PowerPoint to accompany
Robert F. Weaver
Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display.

10-2
10.1 Multiple Forms of Eukaryotic RNA Polymerase
• There are at least two RNA polymerases operating in eukaryotic nuclei– One transcribes major ribosomal RNA genes– One or more to transcribe rest of nuclear genes
• Ribosomal genes are different from other nuclear genes– Different base composition from other nuclear genes– Unusually repetitive– Found in different compartment, the nucleolus

10-3
Separation of the Three Nuclear Polymerases
• Eukaryotic nuclei contain three RNA polymerases– These can be separated by ion-exchange
chromatography
• RNA polymerase I found in nucleolus– Location suggests in transcribes rRNA genes
• RNA polymerases II and III are found in the nucleoplasm

10-4
Roles of the Three RNA Polymerases
• Polymerase I makes large rRNA precursor
• Polymerase II makes – Heterogeneous
nuclear RNA (hnRNA)– Small nuclear RNA
• Polymerase III makes precursors to tRNAs, 5S rRNA and other small RNA

10-5
RNA Polymerase Subunit Structures

10-6
Polymerase II Structure
• For enzymes like eukaryotic RNA polymerases, can be difficult to tell: – Which polypeptides copurify with polymerase
activity – Which are actually subunits of the enzyme
• Technique to help determine whether a polypeptide copurifies or is a subunit is called epitope tagging

10-7
Epitope Tagging
• Add an extra domain to one subunit
• Other subunits normal• Polymerase labeled
by growing in labeled amino acids
• Purify with antibody• Denature with
detergent and separate on a gel

10-8
Polymerase II
Original 10 subunits are placed in 3 groups:
• Core – related in structure and function to bacterial core subunits
• Common – found in all 3 nuclear RNA polymerases
• Nonessential subunits – conditionally dispensable for enzymatic activity

10-9
Core Subunits
• Three polypeptides, Rpb1, Rpb2, Rpb3 are absolutely required for enzyme activity
• These are homologous to ’-, -, and -subunits• Both Rpb1 and ’-subunit binds DNA• Rpb2 and -subunit are at or near the
nucleotide-joining active site• Rpb3 does not resemble -subunit
– There is one 20-amino acid subunit of great similarity– 2 subunits are about same size, same stoichiometry– 2 monomers per holoenzyme

10-10
Common Subunits• There are five common subunits
– Rpb5– Rpb6– Rpb8– Rpb10– Rpb12
• Little known about function• They are all found in all 3 polymerases• Suggests play roles fundamental in
transcription

10-11
Subunits Nonessential for Elongation
• Rpb4 and Rpb7 – Dissociate fairly easily from polymerase– Found in substoichiometric quantities– Might shuttle from one polymerase II to another– Rpb4 may help anchor Rpb7 to the enzyme– Mutants without Rpb4 and Rpb7 transcribes well, but
cannot initiate at a real promoter
• Rpb7 is an essential subunit, so must not be completely absent in the mutant

10-12
Heterogeneity of the Rpb1 Subunit
• RPB1 gene product is subunit II• Subunit IIa is the primary product in yeast
– Can be converted to IIb by proteolytic removal of the carboxyl-terminal domain (CTD) which is 7-peptide repeated over and over
– Converts to IIo by phosphorylating 2 ser in the repeating heptad of the CTD
– Enzyme with IIa binds to the promoter– Enzyme with IIo is involved in transcript
elongation

10-13
The Three-Dimensional Structure of RNA Polymerase II
• Structure of yeast polymerase II (specifically pol II 4/7) at atomic resolution reveals a deep cleft that accepts a linear DNA template from one end to another
• Catalytic center lies at the bottom of the cleft and contains a Mg2+ ion
• A second Mg2+ ion present in low concentrations• Geometry allows enough space for:
– TFIID to bind at the TATA box of the promoter– TFIIB to link the polymerase to TFIID– Places polymerase correctly to initiate transcription

10-14
3-D Structure - RNA Polymerase II in an Elongation Complex
• Structure of polymerase II bound to DNA template and RNA product in an elongation complex has been determined
• When nucleic acids are present, the clamp region of the polymerase has shifted closed over the DNA and RNA– Closed clamp ensures that transcription is
processive – able to transcribe a whole gene without falling off and terminating prematurely

10-15
Position of Nucleic Acids in the Transcription Bubble
• DNA template strand is shown in blue
• DNA nontemplate strand shown in green
• RNA is shown in red

10-16
Position of Critical Elements in the Transcription Bubble
Three loops of the transcription bubble are:
– Lid: maintains DNA dissociation
– Rudder: initiating DNA dissociation
– Zipper: maintaining dissociation of template DNA

10-17
Proposed Translocation Mechanism
• The active center of the enzyme lies at the end of pore 1• Pore 1 also appears to be the conduit for:
– Nucleotides to enter the enzyme– RNA to exit the enzyme during backtracking
• Bridge helix lies next to the active center– Flexing this helix may function in translocation during
transcription

10-18
3-D Structure - RNA Polymerase II in the Posttranslocation State
• X-ray crystallography has shown the lid of Rpb1 interacts with the DNA-RNA hybrid to force the hybrid open after base pair -8
• The lid then interacts with bases of the nascent RNA to keep the hybrid melted beyond base pair -8
• The rudder of Rpb1 collaborates with lid to keep the hybrid melted by interacting with bases -9 and -10
• Fork loop 1 of Rpb2 interacts with bases -5, -6, and -7 of the RNA to keep the RNA-DNA hybrid together

10-19
Structural Basis of Nucleotide Selection
• Moving through the entry pore toward the active site of RNA polymerase II, incoming nucleotide first encounters the E (entry) site– E site is inverted relative to its position in the A site
(active) where phosphodiester bonds form– E and A sites partially overlap– Rotation of nucleotide between the E and A sites may
play a role in base and sugar specificity
• Two metal ions (Mg2+ or Mn2+) are present at the active site– One is permanently bound to the enzyme– The other enters the active site complexed to the
incoming nucleotide

10-20
The Role of Rpb4 and Rpb7
• Structure of the 12-subunit RNA polymerase II reveals that, with Rpb4/7 in place, clamp is forced shut
• Initiation occurs, with its clamp shut, it appears that the promoter DNA must melt to permit template DNA strand to enter the active site
• The Rpb4/7 extends the dock region of the polymerase, which makes binding of transcription factors easier

10-21
10.2 Promoters
• Three eukaryotic RNA polymerases have:– Different structures– Transcribe different classes of genes
• Expect that the 3 polymerases would recognize different promoters

10-22
Class II Promoters
• Promoters recognized by RNA polymerase II (class II promoters) are similar to prokaryotic promoters
• Considered to have two parts:– Core promoter having
4 elements– Upstream promoter
element

10-23
Core Promoter Elements – TATA Box
• TATA box – Found on the nontemplate strand– Very similar to the prokaryotic -10 box– There are frequently TATA-less promoters
• Housekeeping genes that are constitutively active in nearly all cells as they control common biochemical pathways
• Developmentally regulated genes

10-24
Linker Scanning
• Systematically substitute a 10-bp linker for 10-bp sequences throughout the promoter
• Found that mutations within the TATA box destroyed promoter activity

10-25
Core Promoter Elements• In addition to TATA box, core promoters are:
– TFIIB recognition element (BRE)– Initiator (Inr)– Downstream promoter element (DPE)
• At least one of the four core elements is missing in most promoters
• TATA-less promoters tend to have DPEs• Promoters for highly specialized genes tend to
have TATA boxes • Promoters for housekeeping genes tend to lack
them

10-26
Upstream Elements
• Upstream promoter elements are usually found upstream of class II core promoters
• Differ from core promoters in binding to relatively gene-specific transcription factors– GC boxes bind transcription factor Sp1– CCAAT boxes bind CTF (CCAAT-binding
transcription factor)
• Upstream promoter elements can be orientation-independent, yet are relatively position-dependent

10-27
Class I Promoters
• Class I promoters are not well conserved in sequence across species
• General architecture of the promoter is well conserved – two elements:– Core element surrounding transcription start
site– Upstream promoter element (UPE) 100 bp
farther upstream– Spacing between these elements is important

10-28
Class III Promoters
• RNA polymerase III transcribes a set of short genes
• These have promoters that lie wholly within the genes
• There are 3 types of these promoters
10-29
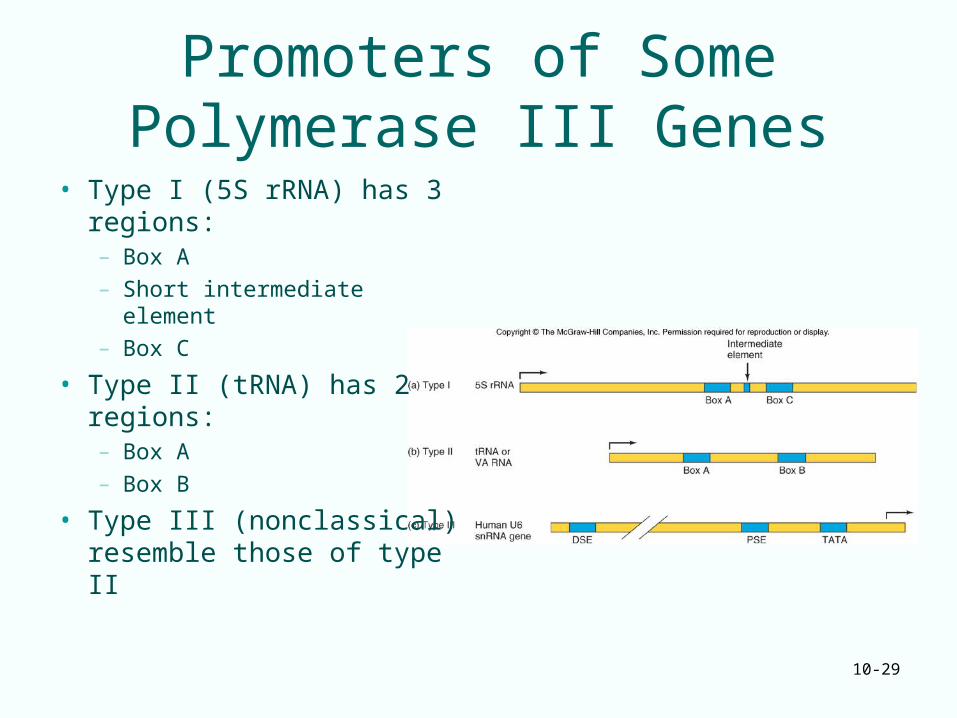
Promoters of Some Polymerase III Genes
• Type I (5S rRNA) has 3 regions:– Box A– Short intermediate element– Box C
• Type II (tRNA) has 2 regions:– Box A – Box B
• Type III (nonclassical) resemble those of type II

10-30
10.3 Enhancers and Silencers
• These are position- and orientation-independent DNA elements that stimulate or depress, respectively, transcription of associated genes
• Are often tissue-specific in that they rely on tissue-specific DNA-binding proteins for their activities
• Some DNA elements can act either as enhancer or silencer depending on what is bound to it