Embed Size (px)
Citation preview

Molecular basis for recognition of methylated andspecific DNA sequences by the zinc finger protein KaisoBethany A. Buck-Koehntopa, Robyn L. Stanfielda, Damian C. Ekierta, Maria A. Martinez-Yamouta, H. Jane Dysona,Ian A. Wilsona,b, and Peter E. Wrighta,b,1
aDepartment of Molecular Biology and bSkaggs Institute for Chemical Biology, The Scripps Research Institute, La Jolla, CA 92037
Contributed by Peter E. Wright, August 7, 2012 (sent for review July 19, 2012)
Methylation of CpG dinucleotides in DNA is a common epigeneticmodification in eukaryotes that plays a central role in maintenanceof genome stability, gene silencing, genomic imprinting, develop-ment, and disease. Kaiso, a bifunctional Cys2His2 zinc finger proteinimplicated in tumor-cell proliferation, binds to bothmethylated CpG(mCpG) sites and a specific nonmethylated DNA motif (TCCTGCNA)and represses transcription by recruiting chromatin remodeling co-repression machinery to target genes. Here we report structures ofthe Kaiso zinc finger DNA-binding domain in complex with its non-methylated, sequence-specificDNA target (KBS) andwitha symmet-rically methylated DNA sequence derived from the promoter regionof E-cadherin. Recognition of specific bases in the major groove ofthe core KBS and mCpG sites is accomplished through both classicaland methyl CH···O hydrogen-bonding interactions with residues inthe first two zinc fingers, whereas residues in the C-terminal exten-sion following the third zinc finger bind in the opposing minorgroove and are required for high-affinity binding. The C-terminalregion is disordered in the free protein and adopts an ordered struc-ture upon binding to DNA. The structures of these Kaiso complexesprovide insights into the mechanism by which a zinc finger proteincan recognize mCpG sites as well as a specific, nonmethylated reg-ulatory DNA sequence.
protein–DNA interaction | NMR spectroscopy | X-ray crystallography |folding upon binding | intrinsic disorder
In eukaryotes, DNA methylation is a common epigenetic mod-ification that is central to the maintenance of genome stability,
gene silencing, genomic imprinting, development, and disease(1, 2). Methyl-CpG–binding proteins (MBPs) mediate these pro-cesses by binding to methylated DNA signals and recruitingchromatin remodeling corepressor complexes, resulting in com-paction of chromatin into its transcriptionally inactive state (3).To date, three classes of MBPs that recognize 5-methyl cytosine(5mC) have been identified. The SRA domain family has speci-ficity for hemimethylated sites and is required for maintenancemethylation during DNA replication (4). In contrast, the methylCpG-binding domain (MBD) and Kaiso families of MBPs functionas essential mediators of epigenetically controlled gene silencingby recognizing symmetrically methylated CpG sites (5). Kaiso is aMBP belonging to the BTB/POZ (broad complex, tramtrak, bric àbrac/pox virus and zinc finger) subfamily of transcription factors thatrecognize cognate DNA sequences through a C-terminal zinc fingerdomain; the N-terminal BTB/POZ domain mediates protein–pro-tein interactions (6). Kaiso is a POZ protein that participates inboth methyl-dependent and sequence-specific transcriptional re-pression, using its three Cys2His2 zinc fingers to recognize either twoconsecutive symmetrically methylated CpG dinucleotides (mCpG)(7, 8) or a TCCTGCNA consensus (termed “KBS”) (9). Althoughoriginally it was reported that Kaiso only requires zinc fingers 2 and3 for DNA recognition (9), we determined that Kaiso in factrequires all three zinc fingers plus adjacent regions for structuralstability and high-affinity binding to both sequence classes (10).Transcriptional repression by Kaiso has implications in both
development and cancer (11–13). Kaiso binds to and silencesaberrantly methylated tumor-suppressor and DNA-repair genes
in cancer cells (14). Specific Kaiso-binding sites are found in thepromoters of several target genes (matrilysin, cyclin-D1, siamois,c-myc, and Wnt11) regulated by the Wnt signaling pathway,which plays a critical role in early development and tumorprogression (9, 15, 16). The cytoplasmic armadillo repeat pro-tein p120 catenin (p120ctn) binds Kaiso and relieves Kaiso-mediated transcriptional repression (6, 17, 18). The interactionof Kaiso with p120ctn has been implicated in regulation of ca-nonical and noncanonical Wnt signaling pathways througha mechanism that involves KBS recognition (15, 16). Addi-tionally, there is increasing evidence that Kaiso expression lev-els are elevated in tumorigenesis, indicating that Kaiso may playa direct role in modulating cancer through transcriptionalcontrol of aberrantly methylated and KBS-containing genes (14,19, 20). It still is unclear whether the transcriptional regulationof the two distinctly different DNA sequences is synergistic innature or mutually exclusive.Although structures of the SRA (UHRF1) (21–23) and MBD
(MBD1, MeCP2, and MBD2) (24–26) domains in complex withmethylated DNA have been determined, the structural basis forrecognition of both mCpG and KBS sites by the Kaiso family isunknown. Here we report crystal and solution structures of theKaiso zinc finger domain in complex with its nonmethylated, se-quence-specific DNA site (KBS) (Fig. 1A) and the crystal struc-ture of the complex with an oligonucleotide derived from thepromoter region of E-cadherin, a known Kaiso-binding site (7, 27)that contains two sequential, symmetrically methylated CpG sites(MeECad) (Fig. 1B). In normal cells, E-cadherin is an importantmediator of cell–cell adhesion, but hypermethylation of the pro-moter in several cancers results in down-regulation of E-cadherinexpression and has been correlated with increasing invasion andmetastasis (28). The current studies provide insights into themechanism by which Kaiso recognizes both methylated DNA andthe unmethylated KBS DNA sequence. Remarkably, the molec-ular interactions that mediate binding to these distinct DNAtarget sites are very similar.
Results and DiscussionOverall Structure of the Kaiso:DNA Complexes. The crystal structureof the Kaiso:KBS complex was determined at 2.4-Å resolutionusing single-wavelength anomalous dispersion from the zinc, andthe Kaiso:MeECad complex then was solved by molecular re-placement at 2.8-Å resolution as detailed in SI Methods. The
Author contributions: B.A.B.-K., M.A.M.-Y. and P.E.W. designed research; B.A.B.-K., R.L.S.,D.C.E., andM.A.M.-Y. performed research; B.A.B.-K., R.L.S., D.C.E., H.J.D., I.A.W., and P.E.W.analyzed data; and B.A.B.-K., H.J.D., I.A.W., and P.E.W. wrote the paper.
The authors declare no conflict of interest.
Data deposition: Atomic coordinates and structure factors have been deposited in ProteinData Bank, www.pdb.org [PDB ID codes 4F6M (crystal structure of the KBS complex), 4F6N(crystal structure of the MeECad complex), and 2LT7 (NMR structures)] and resonanceassignments in the Biological Magnetic Resonance Bank, http://www.bmrb.wisc.edu[accession no. 18462 (NMR assignments)].1To whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1213726109/-/DCSupplemental.
www.pnas.org/cgi/doi/10.1073/pnas.1213726109 PNAS | September 18, 2012 | vol. 109 | no. 38 | 15229–15234
BIOCH
EMISTR
Y
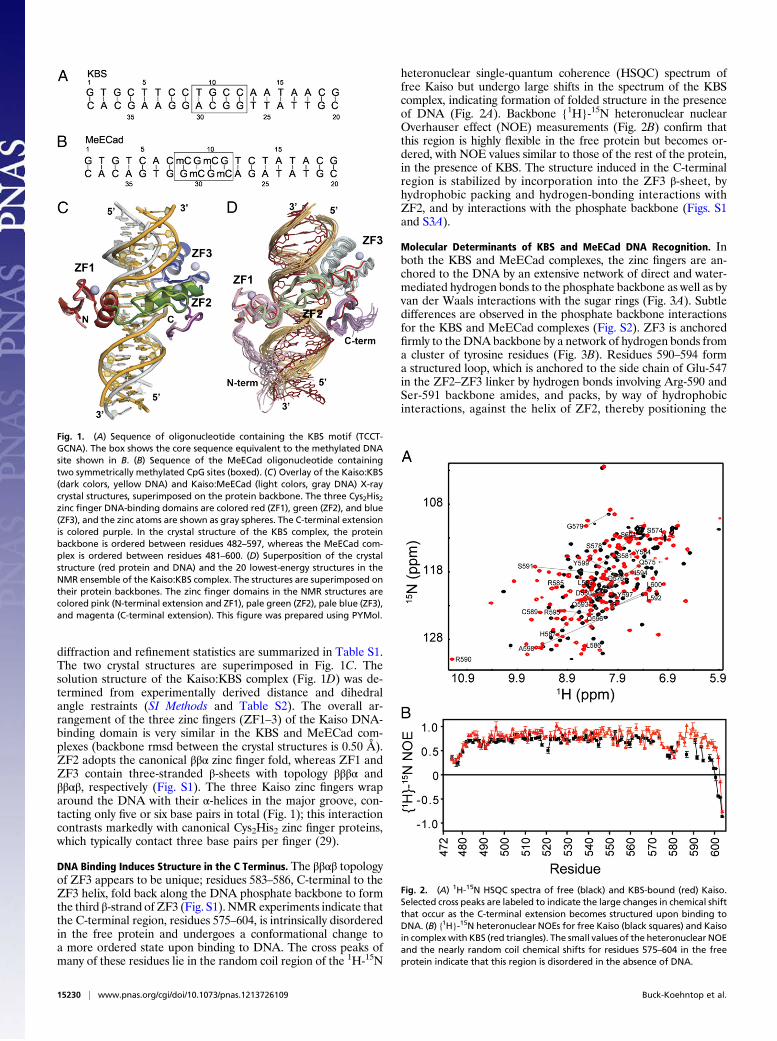
diffraction and refinement statistics are summarized in Table S1.The two crystal structures are superimposed in Fig. 1C. Thesolution structure of the Kaiso:KBS complex (Fig. 1D) was de-termined from experimentally derived distance and dihedralangle restraints (SI Methods and Table S2). The overall ar-rangement of the three zinc fingers (ZF1–3) of the Kaiso DNA-binding domain is very similar in the KBS and MeECad com-plexes (backbone rmsd between the crystal structures is 0.50 Å).ZF2 adopts the canonical ββα zinc finger fold, whereas ZF1 andZF3 contain three-stranded β-sheets with topology βββα andββαβ, respectively (Fig. S1). The three Kaiso zinc fingers wraparound the DNA with their α-helices in the major groove, con-tacting only five or six base pairs in total (Fig. 1); this interactioncontrasts markedly with canonical Cys2His2 zinc finger proteins,which typically contact three base pairs per finger (29).
DNA Binding Induces Structure in the C Terminus. The ββαβ topologyof ZF3 appears to be unique; residues 583–586, C-terminal to theZF3 helix, fold back along the DNA phosphate backbone to formthe third β-strand of ZF3 (Fig. S1). NMR experiments indicate thatthe C-terminal region, residues 575–604, is intrinsically disorderedin the free protein and undergoes a conformational change toa more ordered state upon binding to DNA. The cross peaks ofmany of these residues lie in the random coil region of the 1H-15N
heteronuclear single-quantum coherence (HSQC) spectrum offree Kaiso but undergo large shifts in the spectrum of the KBScomplex, indicating formation of folded structure in the presenceof DNA (Fig. 2A). Backbone {1H}-15N heteronuclear nuclearOverhauser effect (NOE) measurements (Fig. 2B) confirm thatthis region is highly flexible in the free protein but becomes or-dered, with NOE values similar to those of the rest of the protein,in the presence of KBS. The structure induced in the C-terminalregion is stabilized by incorporation into the ZF3 β-sheet, byhydrophobic packing and hydrogen-bonding interactions withZF2, and by interactions with the phosphate backbone (Figs. S1and S3A).
Molecular Determinants of KBS and MeECad DNA Recognition. Inboth the KBS and MeECad complexes, the zinc fingers are an-chored to the DNA by an extensive network of direct and water-mediated hydrogen bonds to the phosphate backbone as well as byvan der Waals interactions with the sugar rings (Fig. 3A). Subtledifferences are observed in the phosphate backbone interactionsfor the KBS and MeECad complexes (Fig. S2). ZF3 is anchoredfirmly to the DNA backbone by a network of hydrogen bonds froma cluster of tyrosine residues (Fig. 3B). Residues 590–594 forma structured loop, which is anchored to the side chain of Glu-547in the ZF2–ZF3 linker by hydrogen bonds involving Arg-590 andSer-591 backbone amides, and packs, by way of hydrophobicinteractions, against the helix of ZF2, thereby positioning the
Fig. 1. (A) Sequence of oligonucleotide containing the KBS motif (TCCT-GCNA). The box shows the core sequence equivalent to the methylated DNAsite shown in B. (B) Sequence of the MeECad oligonucleotide containingtwo symmetrically methylated CpG sites (boxed). (C) Overlay of the Kaiso:KBS(dark colors, yellow DNA) and Kaiso:MeECad (light colors, gray DNA) X-raycrystal structures, superimposed on the protein backbone. The three Cys2His2zinc finger DNA-binding domains are colored red (ZF1), green (ZF2), and blue(ZF3), and the zinc atoms are shown as gray spheres. The C-terminal extensionis colored purple. In the crystal structure of the KBS complex, the proteinbackbone is ordered between residues 482–597, whereas the MeECad com-plex is ordered between residues 481–600. (D) Superposition of the crystalstructure (red protein and DNA) and the 20 lowest-energy structures in theNMR ensemble of the Kaiso:KBS complex. The structures are superimposed ontheir protein backbones. The zinc finger domains in the NMR structures arecolored pink (N-terminal extension and ZF1), pale green (ZF2), pale blue (ZF3),and magenta (C-terminal extension). This figure was prepared using PYMol.
Fig. 2. (A) 1H-15N HSQC spectra of free (black) and KBS-bound (red) Kaiso.Selected cross peaks are labeled to indicate the large changes in chemical shiftthat occur as the C-terminal extension becomes structured upon binding toDNA. (B) {1H}-15N heteronuclear NOEs for free Kaiso (black squares) and Kaisoin complex with KBS (red triangles). The small values of the heteronuclear NOEand the nearly random coil chemical shifts for residues 575–604 in the freeprotein indicate that this region is disordered in the absence of DNA.
15230 | www.pnas.org/cgi/doi/10.1073/pnas.1213726109 Buck-Koehntop et al.

C-terminal region of the polypeptide chain to enter the minorgroove (Fig. 3B and Fig. S3A). A notable difference between theX-ray and NMR structures is observed at the N terminus. There isno electron density for residues preceding the N-terminal β-strandof ZF1 in either crystal structure. The conformation of this regionalso is disordered in the NMR structural ensemble, but weak in-termolecular NOEs from the Arg-475 and Lys-477 side chains in-dicate transient interactions with the DNA backbone across theminor groove (Fig. 1D).
The C-terminal extension to ZF3, which is intrinsically disor-dered in the absence of DNA, is absolutely essential for high-affinity DNA binding. Truncation of the zinc finger construct atGly-579, following the ZF3 helix, severely impairs KBS binding(10). The structures show that this impairment results from theelimination of DNA backbone contacts (from Tyr-584 and Leu-586) and the disruption of an extensive network of minor grooveinteractions. In the crystal structures of both complexes, the Arg-595 side chain is inserted deeply into the minor groove oppositethe core-recognition site, where it forms hydrogen bonds with theO2 of 5mC28 and the N3 of A27 in MeECad (Fig. 4A) and withthe O2 of T26 and the N3 of G27 in KBS (Fig. 4B). In theMeECad complex, the aromatic rings of Tyr-597 and Tyr-599 areinserted deeply into the narrow minor groove at the core mCpGsite, where they pack tightly against the sugars of T12, C13, andG29. The OηH atom of the Tyr-597 side chain forms a hydrogenbond to the N2 amino group of G11, an interaction that is but-tressed by an additional hydrogen bond to the Arg-595 guani-dinium group. The Ala-598 amide hydrogen bonds to thephosphate backbone of C13, helping anchor the neighboring Tyrresidues in the minor groove. In the crystal structure of the KBScomplex, the location of the two Tyr side chains is indeterminate,because no electron density is observed for residues beyondposition 597, and no side-chain density is observed for Gln-596or Tyr-597. In solution, however, intense intermolecular NOEsare observed between the ribose protons of A13/G28 and thearomatic protons of Tyr-597/Tyr-599, showing that these sidechains do in fact bind in the minor groove of the KBS DNA (Fig.4C and Fig. S3B). We conclude that the C-terminal extensionassumes essentially the same conformation in the two complexes,making extensive minor groove contacts that are essential forhigh-affinity binding to both the mCpG and the KBS sites.Base-specific interactions are dominated by side chains in the
N-terminal regions of the ZF1 and ZF2 helices, whereas ZF3interacts primarily with the phosphate backbone and makes no(MeECad) or few (KBS) base-specific contacts. In both thecrystal and NMR structures of the KBS complex (Fig. 1), devi-ations from regular B-form geometry in the core-recognition site(6TCCTGCCA13) allow the ZF3 helix to penetrate the majorgroove so that the side chain of Gln-563 can form direct andwater-mediated hydrogen bonds to the bases of G32 and C7(Table S3). In contrast, because of differences in DNA confor-mation, the ZF3 helix is unable to extend deeply enough into themajor groove of the MeECad DNA to form base-specific con-tacts (Fig. 1A). Instead, Nε2 of the Gln-563 side chain formsa hydrogen bond to the phosphate backbone of C5 (Table S3).Overall, the area of the major groove contacted by ZF3 issmaller in the MeECad complex than in the complex with KBS.Recognition of the KBS and mCpG sites by Kaiso is accom-
plished through base-specific hydrogen bonding and hydrophobicinteractions with the C5-methyl groups of thymines (KBS) ormethylated cytosines (MeECad) within the major groove (Fig.5A). In the MeECad complex, residues in the N-terminal regionsof the ZF1 and ZF2 helices provide hydrophobic environmentsfor the C5-methyls of 5mC10/5mC28 and 5mC8/5mC30, re-spectively. The Cys-505 side chain and the Cγ-methyl of Thr-507form a hydrophobic pocket that accommodates the C5-methylgroup of 5mC10 (Fig. 5B); if the cytosine were unmethylated, thedistance between Cys-505 and the base edge would be greater,and the effectiveness of these hydrophobic interactions would bediminished. The Thr-507 Cγ-methyl also packs against the C5-methyl of 5mC28 (Fig. 5B). Similar interactions are observed inthe KBS complex, through a change in the side-chain rotamerthat places the Thr-507 Cγ-methyl in contact with the T26 C5-methyl and leads to the formation of a water-mediated hydrogenbond between Thr-507 Oγ and the N4-amino of C11 (Fig. 5C). Afurther contribution to specificity comes from hydrophobicinteractions with the Cδ2-methyl of Leu-533, which lies close to
Fig. 3. (A) DNA backbone contacts between ZF1 (magenta) and ZF2 (green)and the coding strand of the KBS complex. Cyan spheres indicate water mol-ecules that mediate hydrogen bonding to the phosphate backbone. (B)Interactions of ZF2 (green) and ZF3 (blue)with the backboneof the noncodingstrand in the MeECad complex. ZF3 is anchored to the DNA backbone byanetworkof hydrogenbonds froma cluster of tyrosines (Tyr-550, Tyr-562, andTyr-584) and from theArg-549guanidiniumgroup. The side chains ofArg-595,Tyr-597, and Tyr-599 in the C-terminal extension (pink) project deeply intothe narrowed minor groove. This figure was prepared using PyMol.
Buck-Koehntop et al. PNAS | September 18, 2012 | vol. 109 | no. 38 | 15231
BIOCH
EMISTR
Y

the C5-methyl groups of 5mC8 and T9 in the MeECad and KBScomplexes, respectively.There are marked similarities in the hydrogen-bonding inter-
actions with bases in the major groove of the core MeECad andKBS sites, despite differences in sequence and cytosine methyl-ation. These interactions are mediated almost entirely by the sidechains of Glu-535 (ZF2) and Arg-511 (ZF1). These residues areinvariant in Kaiso sequences from different species and in theKaiso-like zinc finger proteins ZBTB4 and ZBTB38 (30), sug-gesting that these proteins all share a common mechanism ofDNA recognition. The Glu-535 side chain plays a critical role inthe recognition of the symmetrically methylated mCpG site inMeECad by forming CH···O hydrogen bonds with the C5-methyls of these bases (Fig. 5D and Fig. S4). The marked de-crease in binding affinity observed when all of the 5mC bases arereplaced with nonmethylated cytosines (7, 10) confirms the im-portance of the methyl CH···O hydrogen bonds in specific rec-ognition of mCpG sites by Kaiso. Methyl CH···O hydrogen bondsappear to be common in 5-mC recognition, because they havebeen observed in complexes of MeCP2 and UHRF1 withmethylated or hemimethylated DNA, respectively (22, 23, 25).Interestingly, the position of the Glu-535 side chain is virtuallyidentical in the KBS complex of Kaiso, where it forms a hydro-gen bond with the N4-amino group of C29 as well as a methylCH···O hydrogen bond with the C5-methyl of T9 (Fig. 5D andFig. S4). The Glu-535 Oε2 atom is surprisingly close (2.7 Å) tothe O4 atom of T9, suggesting formation of a hydrogen bondthrough protonation of the carboxylic side chain. A comparisonof the NMR spectrum of Kaiso in complex with the two differentDNA sequences (Fig. S5) shows a large difference in the 15Namide chemical shifts of Glu-535 and Tyr-536, likely reflectinga change in the Glu-535 protonation state between the twocomplexes. The hydrogen-bonding interactions of Arg-511 andGlu-535 are poorly defined in the NMR ensemble because ofa lack of intermolecular NOEs between their side chains andthe DNA.Base-specific interactions involving the guanidinium of Arg-511
also are similar in the two complexes (Fig. 5E). In the MeECadcomplex, Arg-511 forms asymmetrical hydrogen-bonding contactsacross the major groove with the guanines of the central two mC:G base pairs (O6 atoms of G9 and G29). This observation pro-vides insights into why Kaiso prefers two consecutive mCpG basepairs at the center of the recognition site or, at the very least,
requires that a guanine occupy both positions, irrespective of themethylation status of the base-paired cytosine. Although Arg-511cannot distinguish 5mC from C, it clearly discriminates againstApT or TpA steps and thus plays a key role in recognition of thecentral G9:mC30/mC10:G29 base pairs. In the KBS complex,Arg-511 forms similar side-chain hydrogen bonds to the O6/N7 ofG10 and the O6 of G28 (Fig. 5E); substitution of either guanineby thymine greatly impairs binding (9). The importance of thesehydrogen bonds for all DNA-binding interactions is demonstratedby mutagenesis (to alanine) of the equivalent Arg residue (Arg-326) in the Kaiso homolog ZBTB4, which abrogates binding toboth methylated and unmethylated DNA (31). Comparison of theKBS and MeECad complexes shows that as long as the Arg-511/guanine hydrogen bonds are maintained, Kaiso can toleratethymine substitutions at the 5mC8 position (equivalent to T9 inKBS) and likely at 5mC28, although substitution of either 5mC10or 5mC30 with a thymine would greatly impair binding (9).
Recognition of Hemimethylated DNA. To assess the extent to whicheach 5mC site contributes to recognition, we examined the bindingof Kaiso to hemimethylated and single-site symmetrically meth-ylated DNA. Kaiso binds equally well to MeECad and to a probemethylated only on the coding strand (5mC8/5mC10), whereasmethylation on only the noncoding strand (5mC28/5mC30)reduces the affinity slightly (Fig. S6 A and B). Additionally, sym-metric methylation at a single site decreases the binding affinityonly slightly, by ∼1.5-fold for the 5′-site (5mC8/5mC30) and∼twofold for the 5mC10/5mC28 site (Fig. S6C). Thus, Kaiso bindswith significant affinity to hemimethylated DNA and to a singlemCpG site within the context of the ECad sequence. Binding tohemimethylated DNA appears to be asymmetric, with a slightpreference for methylation on the coding strand. Similar behaviorhas been observed for the Kaiso-like protein ZBTB4 (24, 31),suggesting a common mechanism by which Kaiso family membersread and interpret the methylation signal.
ConclusionsOur work has shown how the three Cys2His2 zinc fingers andflanking regions of Kaiso are uniquely adapted for recognition ofboth mCpG and specific nonmethylated DNA sequences. Al-though the mode of DNA binding, with three zinc fingers in themajor groove, is generally similar to other Cys2His2 zinc fingerproteins, Kaiso differs markedly in the length of its core-
Fig. 4. Minor groove interactions. (A) View of minor groove protein:DNA contacts for the MeECad complex, showing hydrogen bonding (dashed red lines)between the Arg-595 and Tyr-597 side chains and the bases of A27, 5mC28 (orange), and G11. A buttressing hydrogen bond between Tyr-597 and Arg-595 isshown in green. The protein backbone is cut away for clarity. (B) View of minor groove contacts in the KBS complex showing hydrogen bonding of the Arg-595 side chain to T26 and G27. (C) DNA contacts made by C-terminal extension [purple backbone with nitrogen (red) and oxygen (blue) molecules] in NMRstructures of the Kaiso:KBS complex, showing the insertion of Arg-595, Tyr-597, and Tyr-599 side chains into the minor groove (DNA in beige with nitrogen inblue and oxygen in red). This figure was prepared using PyMol.
15232 | www.pnas.org/cgi/doi/10.1073/pnas.1213726109 Buck-Koehntop et al.

recognition site. Canonical three-zinc finger proteins “read” thesequence in the major groove through hydrogen bonding and hy-drophobic contacts with nine or 10 base pairs (29). In contrast,Kaiso has evolved to recognize a highly localized site, containingonly four or five base pairs, through hydrogen-bonding interactionsin both the major and minor grooves. Like Kaiso, theMBD familyproteinsMBD1,MeCP2, andMBD2bind to a highly localized site,makingminimal major groove contacts in the immediate vicinity of
a single symmetrical mCpG site (24–26). Kaiso is anchored to theDNA by a plethora of phosphate backbone contacts that un-doubtedly contribute to binding affinity. Major groove contactswith the four or five base pairs in the core DNA sequences aremediated entirely by side chains within ZF1 and ZF2. Thesequences of these two zinc fingers are strongly conserved betweenKaiso and its ZBTB4 and ZBTB38 homologs, but the sequence ofZF3 has diverged considerably (30), suggesting that the third zincfinger of the various family members may interact differently withDNA or with other proteins. The minor groove interactions, madeby residues in the C-terminal extension following ZF3, are re-quired for high-affinity binding to both KBS and methylated DNA(10). The present structural analysis reveals the mechanism ofDNA recognition by the Kaiso zinc finger domain and representsa first step toward a molecular-level description of the complexpathways involved in methylation-dependent silencing of tumor-suppressor genes and in the regulation of the Wnt signalingpathway by Kaiso (14, 32). Once bound to DNA, Kaiso repressesthe target gene by recruiting an N-CoR/histone deacetylase com-plex through its N-terminal BTB/POZ domain (33). Furtherstudies will be required to elucidate the complex network ofdownstream protein–protein interactions through which Kaisoregulates gene expression in development and carcinogenesis.
MethodsSample Preparation. Kaiso-ZF123(472-604) uniformly labeled with 15N, 15N/13C,or 15N/13C/2H was expressed and purified, and protein–DNA complexes wereprepared as described previously (10). Methylated and unmethylated oligo-nucleotides were prepared synthetically (IDT Inc.) and dissolved in 10 mM Tris(pH 7.0). Equimolar amounts of complementary oligonucleotides were heatedto 90 °C and annealed by slow cooling to room temperature to form duplexDNA.All samples of the complexwere prepared in argon-saturated 10mMd11-Tris buffer (pH 7.0) containing 1 mM Tris(2-carboxyethyl)phosphine, 0.005%(wt/vol) NaN3, and 5% (vol/vol) D2O (10). Complete formation of the 1:1 Kaiso:MeECad and Kaiso:KBS complexes used for NMR and crystallography wasverified from 1D NMR spectra of the DNA imino proton resonances and 2D15N-1H HSQC spectra of protein amide resonances.
EMSA. EMSA experiments were run on 5% nondenaturing acrylamide gelsusing 5′-end 32P-labeled duplex DNA sequences. Experimental details aregiven in SI Methods.
X-Ray Structure Determination. Crystals of the Kaiso:DNA complexes wereobtained using sitting-drop vapor diffusion. Crystallographic data werecollected on cryo-cooled crystals at the Advanced Photon Source (APS)beamline 23ID-D and the Stanford Synchrotron Radiation Lightsource (SSRL)beamline 9-2 for the KBS andMeECad complexes, respectively. The Kaiso:KBScomplex crystal structure was solved by zinc single-wavelength anomalousdispersion and refined to 2.4-Å resolution. The crystal structure of the Kaiso:MeECad complex was solved by molecular replacement using the Kaiso fromthe Kaiso:KBS crystal structure as a search model and was refined to 2.8 Å. Amore comprehensive description of the methods and refinement procedurecan be found in SI Methods. The crystallographic and refinement statisticsare summarized in Table S1.
NMR Spectroscopy. NMR spectra of free Kaiso and of the KBS and MeECadcomplexes were recorded at 298K on Bruker 600, 750, 800, and 900 MHzspectrometers. Assignments of backbone and side-chain resonances weremade using standard multidimensional NMR experiments. Structures weredetermined using experimentally derived distance, dihedral angle, and re-sidual dipolar coupling restraints. Initial structures were generated using theprogram CYANA (34) and were refined using restrained molecular dynamicssimulated annealing using the Amber 10 software package (35). The ex-perimental restraints and refinement statistics for the NMR structure en-semble are summarized in Table S2. Details of the assignment procedure andstructure calculations are given in the SI Methods.
ACKNOWLEDGMENTS. We thank Joel Gottesfeld for assistance with EMSAexperiments, Gerard Kroon for assistance with NMR experiments, and BrianM. Lee, Brendan Borin, Chul Won Lee, and Christine Beuck for helpful dis-cussions. X-ray data for 4F6M were collected at the Advanced Photon SourceGeneral Medicine/Cancer (GM/CA) beamline 23ID-D. The GM/CA Collaborative
Fig. 5. Kaiso:KBS and Kaiso:MeECad base-specific interactions. (A) Summaryof major groove interactions. Recognition of MeECad is mediated by majorgroove interactions with only the four 5mC:G base pairs; additional minorgroove hydrogen bonds extend the binding site to A27. The contact surfacefor KBS is larger because of the bending of the DNA around the core recog-nition sequence. The dashed box highlights the core contact site in the twoDNA sequences. Residues are colored red, green, blue, and purple to denotetheir location in ZF1, ZF2, ZF3, and the C-terminal extension, respectively.Residues in the C-terminal extension (purple) make minor groove contacts.Black arrows denote hydrophobic interactions, pink arrows indicated classicalhydrogen bonds, and dashed blue arrows indicate water-mediated hydrogenbonds. A complete summary of all DNA base and backbone contacts is shownin Fig. S2 and Table S3. (B) Cys-505 and Thr-507 form a hydrophobic pocketthat accommodates the methyl groups (orange spheres) of 5mC10 and 5mC28.The 5-methylcytosines are highlighted in orange. (C) Base-specific interactionsof Thr-507 in the KBS complex. The red lines show water-mediated hydrogenbonds to the N4-amino of C11. The water is shown as a cyan sphere. (D) Hy-drogen-bonding interactions involving Glu-535 in the core mCpG- (gray) andKBS- (orange) binding sites. (E) Hydrogen-bonding interactions involving Arg-511 in the core mCpG- (gray) and KBS- (orange) binding sites. The Kaiso colorscheme is as in Fig. 1C. This figure was prepared using PyMol.
Buck-Koehntop et al. PNAS | September 18, 2012 | vol. 109 | no. 38 | 15233
BIOCH
EMISTR
Y

Access Team has been funded in whole or in part with federal funds throughNational Cancer Institute Grant Y1-CO-1020 and National Institute of GeneralMedical Science Grant Y1-GM-1104. Use of the Advanced Photon Source wassupported by the US Department of Energy (DOE), Basic Energy Sciences,Office of Science, under Contract DE-AC02-06CH11357. X-ray data for 4F6Nwere collected at Beamline 9-2 of the Stanford Synchrotron Radiation Light-source (SSRL), a Directorate of the Stanford Linear Accelerator CenterNational Accelerator Laboratory and an Office of Science User Facility
operated for the DOE Office of Science by Stanford University. The SSRLStructural Molecular Biology Program is supported by the DOE Office of Bi-ological and Environmental Research, by the National Institutes of Health(NIH), National Center for Research Resources, Biomedical Technology Pro-gram Grant P41RR001209, and by the National Institute of General MedicalSciences. This work was supported by the NIH Grant GM036643 and theSkaggs Institute of Chemical Biology. B.A.B.-K. was supported by Grant PF-07-124-01-GMC from the American Cancer Society.
1. Miranda TB, Jones PA (2007) DNA methylation: The nuts and bolts of repression. J CellPhysiol 213:384–390.
2. Robertson KD (2005) DNA methylation and human disease. Nat Rev Genet 6:597–610.3. Clouaire T, Stancheva I (2008) Methyl-CpG binding proteins: Specialized transcrip-
tional repressors or structural components of chromatin? Cell Mol Life Sci 65:1509–1522.
4. Bostick M, et al. (2007) UHRF1 plays a role in maintaining DNA methylation inmammalian cells. Science 317:1760–1764.
5. Klose RJ, Bird AP (2006) Genomic DNA methylation: The mark and its mediators.Trends Biochem Sci 31:89–97.
6. Daniel JM, Reynolds AB (1999) The catenin p120(ctn) interacts with Kaiso, a novel BTB/POZ domain zinc finger transcription factor. Mol Cell Biol 19:3614–3623.
7. Prokhortchouk A, et al. (2001) The p120 catenin partner Kaiso is a DNA methylation-dependent transcriptional repressor. Genes Dev 15:1613–1618.
8. Prokhortchouk AV, Aitkhozhina DS, Sablina AA, Ruzov AS, Prokhortchouk EB (2001)Kaiso, a new protein of the BTB/POZ family, specifically binds to methylated DNAsequences. Russ J Genet 37(6):603–609.
9. Daniel JM, Spring CM, Crawford HC, Reynolds AB, Baig A (2002) The p120(ctn)-bindingpartner Kaiso is a bi-modal DNA-binding protein that recognizes both a sequence-specific consensus and methylated CpG dinucleotides. Nucleic Acids Res 30:2911–2919.
10. Buck-Koehntop BA, Martinez-Yamout MA, Dyson HJ, Wright PE (2012) Kaiso uses allthree zinc fingers and adjacent sequence motifs for high affinity binding to sequence-specific and methyl-CpG DNA targets. FEBS Lett 586:734–739.
11. Daniel JM (2007) Dancing in and out of the nucleus: p120(ctn) and the transcriptionfactor Kaiso. Biochim Biophys Acta 1773:59–68.
12. Kelly KF, Daniel JM (2006) POZ for effect—POZ-ZF transcription factors in cancer anddevelopment. Trends Cell Biol 16:578–587.
13. van Roy FM, McCrea PD (2005) A role for Kaiso-p120ctn complexes in cancer? Nat RevCancer 5:956–964.
14. Lopes EC, et al. (2008) Kaiso contributes to DNA methylation-dependent silencing oftumor suppressor genes in colon cancer cell lines. Cancer Res 68:7258–7263.
15. Kim SW, et al. (2004) Non-canonical Wnt signals are modulated by the Kaiso tran-scriptional repressor and p120-catenin. Nat Cell Biol 6:1212–1220.
16. Park JI, et al. (2005) Kaiso/p120-catenin and TCF/β-catenin complexes coordinatelyregulate canonical Wnt gene targets. Dev Cell 8:843–854.
17. Kelly KF, Spring CM, Otchere AA, Daniel JM (2004) NLS-dependent nuclear localiza-tion of p120ctn is necessary to relieve Kaiso-mediated transcriptional repression. J CellSci 117:2675–2686.
18. Spring CM, et al. (2005) The catenin p120ctn inhibits Kaiso-mediated transcriptionalrepression of the beta-catenin/TCF target gene matrilysin. Exp Cell Res 305:253–265.
19. Prokhortchouk A, et al. (2006) Kaiso-deficient mice show resistance to intestinal
cancer. Mol Cell Biol 26:199–208.20. Dai SD, et al. (2011) Upregulation of δ-catenin is associated with poor prognosis and
enhances transcriptional activity through Kaiso in non-small-cell lung cancer. Cancer
Sci 102:95–103.21. Hashimoto H, et al. (2008) The SRA domain of UHRF1 flips 5-methylcytosine out of the
DNA helix. Nature 455:826–829.22. Avvakumov GV, et al. (2008) Structural basis for recognition of hemi-methylated DNA
by the SRA domain of human UHRF1. Nature 455:822–825.23. Arita K, Ariyoshi M, Tochio H, Nakamura Y, Shirakawa M (2008) Recognition of hemi-
methylated DNA by the SRA protein UHRF1 by a base-flipping mechanism. Nature
455:818–821.24. Ohki I, et al. (2001) Solution structure of the methyl-CpG binding domain of human
MBD1 in complex with methylated DNA. Cell 105:487–497.25. Ho KL, et al. (2008) MeCP2 binding to DNA depends upon hydration at methyl-CpG.
Mol Cell 29:525–531.26. Scarsdale JN, Webb HD, Ginder GD, Williams DC, Jr. (2011) Solution structure and
dynamic analysis of chicken MBD2 methyl binding domain bound to a target-meth-
ylated DNA sequence. Nucleic Acids Res 39:6741–6752.27. Zhang BZ, Gu LK, Deng DJ (2007) [Methylation specific binding activity of zinc finger
protein Kaiso]. Zhonghua Yu Fang Yi Xue Za Zhi, 41(Suppl):43–46. Chinese.28. Hirohashi S, Kanai Y (2003) Cell adhesion system and human cancer morphogenesis.
Cancer Sci 94:575–581.29. Wolfe SA, Nekludova L, Pabo CO (2000) DNA recognition by Cys2His2 zinc finger
proteins. Annu Rev Biophys Biomol Struct 29:183–212.30. Filion GJP, et al. (2006) A family of human zinc finger proteins that bind methylated
DNA and repress transcription. Mol Cell Biol 26:169–181.31. Sasai N, Nakao M, Defossez PA (2010) Sequence-specific recognition of methylated
DNA by human zinc-finger proteins. Nucleic Acids Res 38:5015–5022.32. Del Valle-Pérez B, et al. (2011) Wnt controls the transcriptional activity of Kaiso
through CK1ε-dependent phosphorylation of p120-catenin. J Cell Sci 124:2298–2309.33. Yoon HG, Chan DW, Reynolds AB, Qin J, Wong J (2003) N-CoR mediates DNA meth-
ylation-dependent repression through a methyl CpG binding protein Kaiso. Mol Cell
12:723–734.34. Güntert P (2004) Automated NMR structure calculation with CYANA. Methods Mol
Biol 278:353–378.35. Case DA, et al. (2005) The Amber biomolecular simulation programs. J Comput Chem
26:1668–1688.
15234 | www.pnas.org/cgi/doi/10.1073/pnas.1213726109 Buck-Koehntop et al.