-
Breast Cancer Research and Treatment 69: 2938, 2001. 2001 Kluwer
Academic Publishers. Printed in the Netherlands.
Report
Modulation of expression of ribosomal protein L7a (rpL7a) by
ethanol inhuman breast cancer cells
Yunfeng Zhu, Hong Lin, Zheng Li, Mei Wang, and Jia LuoDepartment
of Anatomy, West Virginia University School of Medicine,
Morgantown, WV, USA
Key words: alcohol, differential display reverse transcription
PCR, metastasis, northern blot, stress response,transcriptional
regulation
Summary
Epidemiological studies indicate that there is a positive
correlation between alcohol consumption and the risk ofbreast
cancer. Experimental results demonstrate that ethanol is a tumor
promoter and chronic ethanol exposureenhances metastasis and growth
of breast cancer. The present study used an in vitro model to
investigate themolecular mechanism(s) underlying tumor promoting
effects of ethanol. With differential display reverse
tran-scription polymerase chain reaction, we demonstrated that
human ribosomal large subunit protein L7a (rpL7a) wasan
ethanol-responsive factor in T47D breast cancer cells. The results
of northern blot hybridization revealed thatthe effect of ethanol
on L7a expression was duration- and concentration-dependent.
Initial exposure resulted in a2-fold increase in rpL7a level,
whereas a longer exposure period produced a down-regulation.
Ethanol had littleeffect on the stability of rpL7a mRNA; however,
the transcription rate of rpL7a was significantly increased
byethanol. Ethanol-induced up-regulation of rpL7a was not a simple
stress response, because other stress inducers,such as heat shock,
did not affect the expression of rpL7a. Furthermore, breast cancer
cells expressed higherlevel of rpL7a than normal mammary epithelial
cells. Ribosomal proteins are known to play an important role
intranslational regulation, and they have been implicated in the
control of cellular transformation, tumor growth,aggressiveness and
metastasis. Specially, rpL7a activates the trk oncogene by
contributing an amino-terminal-activating sequence to the receptor
kinase domain of trk. Thus, ethanol-induced alteration of rpL7a
expression maymediate the promoting effects of ethanol on breast
cancer development.
Introduction
Breast cancer is a leading cause of morbidity andmortality in
women [1]. The cause of most breastcancer remains elusive and
considerable research at-tention has focused on identifying the
endogenous andenvironmental factors that contribute to its
etiology.Alcohol, tobacco and diet are the three major humancancer
risk factors [2, 3]. High consumption of alco-holic beverages
increases the risk of certain cancers[4]. Ethanol as a risk factor
for breast cancer was firstsuggested by Williams and Horm in 1977
[5]. Today,over 50 epidemiologic studies have examined the
re-lationship between ethanol consumption and breastcancer.
Although some controversy remains, most ofthe studies agree that
there is a positive correlation
between ethanol intake and risk of breast cancer [seereview
69].
The positive association between ethanol con-sumption and breast
cancer is highlighted by severalrecent studies [e.g., 1015].
Findings are summarizedas follows:
(a) Consuming 560 g of alcohol per day increases therisk of
breast cancer by 1570%.
(b) Risk is directly correlated with the duration ofdrinking,
that is, the greater the total number ofyears of drinking, the
greater the risk.
(c) Alcohol consumption is strongly associated withadvanced and
invasive breast tumors.
This suggests that alcohol exposure promotes tumorgrowth and
metastasis. Despite the documentation
-
30 Y Zhu et al.
of the epidemiology of alcohol and breast cancer,little
information regarding the mechanism of ethanol-induced
tumorogenesis is known. Therefore, isolationof specific
ethanol-responsive genes (ERGs) in breastcancer cells is important,
because identification ofsuch genes will not only clarify the
targets of eth-anol but also provide an insight into the
molecularmechanisms of cancer development.
T47D human breast cancer cells were used as anin vitro model.
These cells are derived from humanmammary duct carcinoma and
express estrogen re-ceptors [16]. We have previously demonstrated
thatethanol enhanced migration of T47D cells in culture[17]. The
current study uses differential display re-verse transcription
polymerase chain reaction (DDRT-PCR) to identify ERGs in breast
cancer cells. Herein,we demonstrate that ethanol exposure modulates
theexpression of ribosomal protein L7a (rpL7a) in T47Dcells.
Materials and methods
Cell culture
Human mammary epithelial cells (MCF-10A), andbreast cancer cells
(T47D, MCF-7 and MDA-MB-231) were obtained from American Type
Culture Col-lection (ATCC, Rockville, MD). MCF-10A cells weregrown
in a mammary growth medium specified byATCC, supplemented with 100
ng/ml cholera toxin(Calbiochem. La Jolla, CA). T47D cells were
grownin RPMI1640 medium supplemented with 2 mM L-glutamine, 4.5 g/l
glucose, 10 mM HEPES, 1 mM so-dium pyruvate, 1.5 g/l sodium
bicarbonate, 7.2 mg/linsulin, 100g/ml Penicilin/Strepmycin, and
10%fetal bovine serum. MCF-7 and MDA-MB-231 cellswere maintained
Eagles minimal essential medium(EMEM) with the same supplement as
that for T47Dcells. Cells were incubated at 37C in an
atmospherecontaining 5.0% CO2 and saturating humidity. Themedium
was changed every 23 days.
Ethanol exposure and heat shock treatment
Because of ethanols volatility, a method utilizingsealed
containers [18, 19] was used to maintain eth-anol levels in the
culture medium. With this method,ethanol was added directly to the
culture medium intissue culture dishes, and the dishes then placed
insealed containers with an ethanol-containing waterbath in the
bottom. The concentration of ethanol in
the bath was the same as that in the culture medium.Ethanol from
the bath evaporates into the air of thesealed container and
maintains the ethanol concentra-tion in the culture medium. A small
volume of CO2was injected into the container prior to sealing.
Theethanol bath was changed daily to maintain the
ethanolconcentration. Control cultures were maintained
con-currently in separate sealed containers; however, thewater bath
in the control groups contained no ethanol.All containers were
incubated at 37C. Cells were ex-posed to ethanol at 100400 mg/dl.
The concentrationsapplied are physiologically relevant to blood
ethanollevels observed in alcoholics. Generally higher
ethanolconcentration is required in vitro to replicate
similareffects in vivo [20, 21]. In previous studies, we
haveconfirmed that this sealed container method accuratelymaintains
ethanol concentrations in the culture me-dium [19]. The procedure
for heat shock treatmentwas similar to that previously described
[22]. Briefly,cells grown in a 175 cm2 flask were heated at 45C
for30 min. After heat shock, cells were incubated at 37Cfor the
time indicated prior to RNA extraction.
DDRT-PCR
DDRT-PCR was first developed in 1992 to identifydifferentially
expressed genes, and it has been suc-cessfully applied to isolate
specifically induced genesin cancer [23]. Total RNA was isolated
using a TRIREAGENT kit (Molecular Research Center, INC,
Cin-cinnati, Ohio). The concentration of RNA was de-termined by the
absorbance at 260 nm, and the puritywas determined by the 260/280
ratio with a BioPho-tometer (Eppendorf, Westbury, NY). DDRT-PCR
wascarried out using a commercially available kit,
DeltatmDifferential Display Kit (Cat. # K1810-1, Clontech,Palo Ato,
CA). The protocol consists of two stages:cDNA synthesis and
differential display PCR. First-strand cDNA was synthesized using
each of the RNAsof interest as a template and oligo (dT) as a
primer.One of the advantages of this protocol is that it re-quires
only a single cDNA synthesis reaction for eachRNA sample, in
contrast to 912 syntheses requiredfor each RNA sample in other
differential display pro-tocols. The manufacturers instructions
were followed.In brief, 2g of total RNA was reverse transcribedwith
oligo-dT at 42C for 1 h. cDNA was ampli-fied by PCR reaction.
Amplification was performedin the presence of specific primers and
[-33P]dATP.The PCR products were separated on 6.0% dena-turing
polyacrylamide/urea gel for 24 h. Gels were
-
Ethanol alters the expression of ribosomal protein 31dried and
exposed to Kodak X-Omat film overnightat 70C. Differentially
displayed cDNAs were re-covered from the gel and re-amplified with
the sameprimers.
DNA cloning and sequence analysis
Differentially displayed PCR products were ligatedinto a
pCR4-TOPO, TA vector (Invitrogen, Carls-bad, CA). Competent
bacterial cells were transformedwith the ligated construct. Plasmid
DNA subcolonescontaining inserts were purified using QIAprep
SpinMiniprep Kit (Qiagen, Stanford Valencia, CA). Se-quence
analysis on the inserts was performed by Re-search Genetics, INC
(Huntsville, AL). Sequencesthus derived were compared for homology
to the se-quences available in the current GenBank databasethrough
the National Center for Biotechology Inform-ation using the BLAST
program [24].
Northern blot hybridization
The cDNA probes were derived from differential dis-play PCR as
described above. The probes were labeledwith [32P]dCTP by PCR
reaction. The reaction mix-ture contained the following components:
[32P]dCTP(3000 Ci/mM, ICN), templates (plasmid subclone,20 ng/l),
50 dNTP (10 mM of dATP, dGTP, anddTTP, and 100M of dCTP), Taq DNA
polymerase(5 U/l), and primers (same as for DDRT-PCR).
Amp-lification was performed at 95C for 3 min for denatur-ing,
followed by 30 cycles of additional reaction (94Cfor 30 s, 55C for
30 s, and 72C for 2 min). Labeledprobes were then purified by G-50
column (Promega,Madison, WI).
Ten micrograms of total RNA per lane were separ-ated by
electrophoresis on 1.0% agarose/formaldehydegel. The RNA was
transferred to Nytran filters with aBio-RAD Vacuum Blotter Model
785 and ultraviolet-cross-linked. The hybridization was performed
withthe following procedure. Pre-hybridization occurredby
incubating filters with a hybridization buffer(ULTRAhybTM Buffer,
Austin, TX) at 42C for30 min. After pre-hybridization, the filters
were hy-bridized with cDNA probes at a final concentration of1 106
cpm/ml in the same buffer (42C, overnight).The filters were washed
three times (5 min each) at42C in 2X SSC containing 0.1% SDS. This
was fol-lowed by three additional washes (15 min each) at42C in
0.1X SSC containing 0.1% SDS. The signalswere processed with a
PhosphorImager.
mRNA stability
The stability of the mRNA was determined by amethod described by
Dani et al. [25]. Briefly, totalRNA was extracted from T47D cells
before and atvarious times (30 min8 h) after treatment with
actino-mycin D (5g/ml). Actinomycin D at this concentra-tion blocks
essentially all transcriptional activity. RNAsamples were then
subject to northern blot analysis asdescribed above. The signals
were processed with aPhosphorImager
Nuclear run-off assayNuclei were isolated by sucrose gradient
centrifu-gation [26] and stored at 70C. For analysis
oftranscription rate of specific gene, 250l of nuc-lei suspension
(4 107 nuclei/100l) was incubatedwith 100Ci of [-32P] UTP and 60l
of 5X run-off buffer (25 mM Tris-HCl, pH 8.0, 12.5 mM MgCl2,750 mM
KCl, 1.25 mM of ATP, CTP, and GTP) for30 min at 30C. After
reaction, the labeled RNA wasisolated using Nick columns (Pharmacia
Biotech, Pis-cataway, NJ). The RNA transcripts were
partiallyhydrolyzed with 200 mM NaOH at 0C for 10 min,then
neutralized with HEPES acid. Nytran filters(Schleicher &
Schuell, Keene, NH) were coated with1g of denatured cDNAs for rpL7a
or cyclophilin byslot-blotting. Then the filters were UV
cross-linkedat 254 nm (Spectro Linkertm, Spectronics Corpora-tion,
West Tbury, NY), and pre-hybridized with ahybridization buffer (50%
formamide, 250 mM so-dium phosphate, pH 7.2, 1 mM EDTA, 250 mM
NaCl,100 mg/ml ssDNA, 40g/ml tRNA and 7.0% SDS) at55C for 4 h.
Hybridization in the same solution con-taining 2 106 cpm of the
32P-RNA was performed at55C for 48 h. Filters were washed four
times (20 mineach) in 2X SSC, followed by a 15 min wash in 2XSSC
containing 5g/ml RNAse A, 100g/ml RNAseT1 at 65C. Finally, the
filters were rinsed in 0.3XSSC containing 1.0% SDS for 1 h. The
hybridizationsignals were visualized using a PhosphorImager.
Statistical analysis
Differences among treatment groups were tested us-ing a one-way
analysis of variance (ANOVA). Dif-ferences in which p< 0.05 were
considered statistic-ally significant. In cases where significant
differenceswere detected, specific post-hoc comparisons
amongtreatment groups were performed using a Student-Newman-Keuls
test.
-
32 Y Zhu et al.
Results
Identification of ethanol-responsive gene
With DDRT-PCR, eleven cDNA PCR products thatwere differentially
expressed in T47D cells betweencontrols and ethanol-treated cells
were identified. Thedifferentially expressed cDNA fragments were
thenexcised from the polyacramide gel and re-amplifiedwith the same
primers as used in DDRT-PCR. Amongthe eleven cDNA products
recovered, three of themwere verified by northern blots as
differentially dis-played fragments. These three candidate
ethanol-responsive genes (ERGs) were purified and ligatedinto a
PCR4-TOPO, TA cloning vector. The differ-entially expressed
candidate ERG cDNA inserts weresequenced and compared for homology
to entries inthe existing nucleic acid database available
throughthe National Center for Biotechology Information.
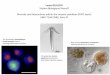
Figure 1. Differential display of mRNA in control (Ct) and
ethanol(Et)-treated breast cancer cells. T47D cells were exposed to
ethanol(400 mg/dl) for 2 weeks. Total RNA was isolated and a sample
of2g of total RNA was used for DDRT-PCR. Lanes 1 and 3 wereloaded
with the cDNA product diluted at 1:10. Lanes 2 and 4 wereloaded
with cDNA product diluted with 1:40. Arrow points to
thedifferentially displayed product, rpL7a.
Figure 2. Northern blot analysis of rpL7a expression in control
(Ct)and ethanol (Et)-treated breast cancer cells. T47D cells were
ex-posed to ethanol (400 mg/dl) for various durations (6 h, 24 h, 4
daysand 2 weeks), and total RNA was isolated. Top panel: Ten
micro-grams of total RNA was subject to northern blot analysis
using aprobe prepared from the product of DDRT-PCR. Bottom panel:
Thesame blot was stripped off cDNA probe for rpL7a, and
re-hybridizedwith a probe for the housekeeping gene
cyclophilin.
Figure 3. Time sequence for ethanol-mediated rpL7a
expression.T47D cells were exposed to ethanol (400 mg/dl) for
various dura-tions (6 h2 weeks). rpL7a expression was examined with
northernblot hybridization (see Figure 2). The relative amount of
rpL7a levelwas determined with a PhosphorImager and expressed as
percentageof control. The expression of rpL7a was normalized with
the levelof cyclophilin. denotes a statistically significant
difference fromcontrol. The result was the mean S.E.M. of four
replicates.
Two cDNA fragments of candidate ERGs displayedno homology to any
identified genes. One PCRproduct (512 bp) exhibited 100% homology
to hu-man ribosomal large subunit protein rpL7a (accession#:
NM000972). As determined by DDRT-PCR (Fig-ure 1), ethanol exposure
for 2 weeks suppressed theexpression of this gene.
Expression of rpL7a mRNA
The expression of rpL7a mRNA was examined bynorthern blot
hybridization using a specific cDNAprobe for rpL7a. The probe
hybridized a mRNA bandaround 1 kb. Time sequence analysis of rpL7a
expres-sion showed that the effect of ethanol on rpL7a was
-
Ethanol alters the expression of ribosomal protein 33
Figure 4. Concentration-dependent effect of ethanol on rpL7a
expression. A: T47D cells were exposed to ethanol (0400 mg/dl) for
either one(Left panel) or 4 days (Right panel), and the total RNA
was isolated. Top panel: the expression of rpL7a was examined with
Northern blothybridization using a probe prepared from the product
of DDRT-PCR. Bottom panel: the same blot was stripped off cDNA
probe for rpL7a,and re-hybridized with a probe for the housekeeping
gene cyclophilin. B: The relative amount of rpL7a was determined
with a PhosphorImager.The expression of rpL7a was normalized with
the level of cyclophilin. Left panel: the expression of rpL7a in
the cells that were exposed toethanol for 1 day. Right panel: the
expression of rpL7a in cells that were treated with ethanol for 4
days. The result was the mean S.E.M. offour replicates. denotes a
statistically significant difference from control.
specific and duration-dependent (Figure 2). Ethanolspecifically
altered the expression of rpL7a mRNA,but had little effect on
cyclophilin expression. Theexpression of rpL7a and cyclophilin was
quantifiedwith a PhosphorImager and the relative amount ofrpL7a was
normalized with cyclophilin (Figure 3).Ethanol produced an initial
increase in rpL7a ex-pression with the amount of rpL7a transcript
doub-ling after 24 h of ethanol exposure. Then, the effectof
ethanol was diminished thereafter. Although theethanol-mediated
increase of rpL7a expression wasreduced, the up-regulation was
still evident after 4days of exposure. In contrast to initial
increases,long-term exposure to ethanol (2 weeks)
significantlyreduced the transcript of rpL7a by more than
60%(Figure 3). Although ethanol did induce up-regulationof rpL7a in
other breast cancer cell lines (MCF-7 andMDA-MB231) to various
extents (data not shown),T47D cells displayed the most dramatic and
consistentresponse.
To examine the effect of ethanol at pharmacologic-ally relevant
concentrations, T47D cells were exposedto a range of ethanol
concentrations. As shown inFigure 4, ethanol-induced up-regulation
of rpL7a wasconcentration-dependent. Expression of rpL7a
wassignificantly increased following 1-day of exposureat ethanol
concentrations as low as 100 mg/dl, andprogressively higher ethanol
concentrations producedenhanced up-regulation. Although the effect
of ethanoldiminished, a statistically significant up-regulationwas
still observed following 4-days of exposure toethanol at
concentrations of 200 and 400 mg/dl.
Figure 5. Effect of heat shock on the expression of rpL7a.
T47Dcells received heat shock treatment as described in Materials
andmethods. Total RNA was isolated at various time points post
heatshock. rpL7a expression was examined with northern blot
hybrid-ization. The relative amount of rpL7a level was determined
with aPhosphorImager and expressed as percentage of control. The
ex-pression of rpL7a was normalized with the level of cyclophilin.
Theamount of rpL7a transcript was not signigicantly altered by
heatshock treatment at any of the times studied (2, 6, 12, 24 h).
Theresult was the mean S.E.M. of four replicates.
Ethanol is known to evoke cellular stress response.Ethanol
exposure, and other stress inducers, such asheat shock, induces the
expression of stress-relatedproteins (e.g., heat shock proteins and
Alu)[22, 2729]. To determine whether rpL7a is a
stress-responsiveprotein, we applied heat shock to T47D cells and
ex-amined the expression of rpL7a following heat shock.The
expression of heat shock protein 27 and 70(HSP27 and HSP70) was
up-regulated following thistreatment (data not shown). As shown in
Figure 5, heat
-
34 Y Zhu et al.
Figure 6. Expression of rpL7a in mammary epithelial cells
andbreast cancer cells. Total RNA was extracted from various cell
linesand equal amount of RNA was subject to northern blot
hybridiza-tion. The expression of rpL7a was compared among normal
mam-mary epithelial cells (MCF-10A) and various breast cancer
cells(T47D, MCF-7 and MDA-MB-231). (1) T47D; (2) MDA-MB-231;(3)
MCF-7; and (4) MCF-10A.
Figure 7. Stability of rpL7a mRNA. T47D cells were exposed
toethanol (0, or 400 mg/dl) for 24 h. Total RNA was extracted
fromT47D cells before and at various times (30 min8 h) after
treatmentwith actinomycin D (5g/ml). RNA samples were then subject
tonorthern blot analysis as described in the Materials and
methods.The signals were processed with a PhosphorImager and
expressedas percentage of the amount prior to application of
actinomycin D.The result was the mean S.E.M. of four
replicates.
shock treatment had little effect on the expression ofrpL7a mRNA
in T47D cells.
To determine whether rpL7a is over-expressed inbreast cancer
cells, mRNA level of rpL7a was com-pared among normal mammary
epithelial cells andvarious breast cancer cells using northern blot
hybrid-ization. MCF-10A is a normal mammary epithelialcell line.
T47D and MCF-7 are estrogen receptor-positive human breast cancer
cells, and MDA-MB-231is an estrogen receptor-negative breast cancer
cell line.As shown in Figure 6, the mRNA of rpL7a in all
threebreast cancer cell lines (T47D, MCF-7 and MDA-MB-231) was
significantly higher than normal mammaryepithelial cells (MCF-10A)
and estrogen receptor-positive breast cancer cell lines displayed
extremelyhigh level of rpL7a.
mRNA stability and transcription of rpL7a
The ethanol-induced up-regulation of mRNA of rpL7acould result
from an alteration in the rate of gene tran-scription or the
stability of mRNA, or both. To distin-guish between these
possibilities, analysis of mRNAstability and nuclear run-off assay
were conducted incontrol and ethanol-exposed cells. T47D cells
wereexposed to ethanol at 200 and 400 mg/dl for 24 h. Ana-lysis of
mRNA stability and nuclear run-off assay wereperformed as described
in the Materials and methods.As shown in Figure 7, ethanol had
little effect on thedegradation rate of rpL7a transcript. In
contrast, eth-anol at 400 mg/dl almost doubled the transcription
rateof rpL7a gene, which was determined by the ability ofnuclei to
produce mRNA of rpL7a (Figure 8). As ex-pected, the transcription
rate of cyclophilin gene wasnot changed following ethanol
exposure.
Discussion
DDRT-PCR revealed that ethanol exposure alters theexpression of
the ribosomal large subunit proteinrpL7a. Subsequent study with
northern blot hybrid-ization indicated that ethanol modulates
expressionof rpL7a in a duration- and
concentration-dependentmanner. Ethanol induces an initial
up-regulation ofrpL7a expression followed by a suppression of
thisgene. The acute effect of ethanol may better mimic thein vivo
situation. On the other hand, down-regulationof rpL7a at late time
point may reflect a desensitizationor compensatory response to the
chronic exposure.Ethanol at pharmacologically relevant
concentration(e.g., 100 mg/dl) significantly up-regulates rpL7a
ex-pression, and the higher the ethanol concentrationapplied, the
stronger up-regulation observed. Ethanol-mediated increase in rpL7a
expression results fromenhanced transcriptional activity. In
addition, ethanol-induced up-regulation of rpL7a is not a simple
stressresponse, because other stress inducers, such as heatshock,
did not affect the expression of rpL7a.
Ribosomes are catalyzers of protein synthesis. Eu-karyotic
ribosomes are composed of two subunits, 40Sand 60S, according to
their sedimentation coefficients.During protein synthesis, mRNA and
tRNA are boundto the small subunit, while the peptide bond
form-ation is catalyzed by the large subunit. The smallsubunit
consists of an 18S ribosomal RNA (rRNA)molecule and about 33
proteins. The large subunit iscomposed of 5S, 5.8S, and 28S rRNA,
together with
-
Ethanol alters the expression of ribosomal protein 35
Figure 8. Transcription rate analysis of rpL7a from control (Ct)
and ethanol-treated (Et) T47D cells. A: Cells were exposed to
ethanol (0,200, and 400 mg/dl) for 24 h. Nuclei were isolated and
subject to nuclear run-off assay as described in Materials and
methods. A constantcDNA concentration (1g) was used in each lane.
B: The relative transcription rate was determined with a
PhosphorImager and expressed aspercentage of control. The result
was the mean S.E.M. of four independent experiments. p< 0.05
versus control values.
approximately 49 proteins. Although a large numberof ribosomal
proteins exist, many of them do not ap-pear to be essential for
ribosome function. Ribosomalproteins participate in various
extra-ribosomal activit-ies [3032]. The specific function for many
ribosomalproteins remains unknown. Cumulative evidence sug-gests
that ribosomal proteins are involved in the ma-lignant
transformation, growth of cancer cells, tumoraggressiveness and
metastasis [3339]. For example,over-expression of several ribosomal
proteins has beenreported in colon [36, 40, 41], liver [39], brain
[42],breast [43] and prostate [44] carcinomas.
The ribosomal protein rpL7a is DNA-damage in-ducible. Ben-Ishai
et al. [45] have shown that ul-traviolet irradiation and other
DNA-damaging agentsspecifically induce transient increases of rpL7a
tran-scripts. UV irradiation and DNA damage act as both atumor
initiator and as a tumor promoter [4649]. Thissuggests rpL7a may be
involved in carcinogenesis anddevelopment of tumors. The
involvement of rpL7ain tumorogenesis is supported by the finding
thatrpL7a activates the trk (tyrosine receptor kinase) on-cogene by
contributing the amino-terminal-activatingsequence to the receptor
kinase domain of trk [50].Furthermore, over-expression of rpL7a has
been ob-served in prostate, colon and brain tumors [36, 42, 44].Our
results suggest that rpL7a is over-expressed inbreast cancer cells.
Elevated expression of rpL7a in tu-mor cells suggests that it plays
a role in the malignanttransformation and the growth of cancer
cells. rpL7ais believed to be involved in cell growth and
differenti-ation [51]. For example, thyrotropin (TSH) is a
growthfactor for thyroid follicular cells, and TSH stimula-
tion of thyroid follicular cell leads to cellular
growth,replication and differentiation. Treatment of thyroidcells
with TSH results in a significant up-regulationof rpL7a [51].
It has been suggested that control of cell growthmay be
regulated, at least partially, by rpL7as interac-tion with nuclear
hormone receptors. rpL7a specific-ally interacts with human thyroid
hormone receptor(THR) and retinoic acid receptor (RAR) which in
turninhibits transactivation of the two nuclear hormone re-ceptors
[52]. Retinoic acid (RA) and thyroid hormone(TH) play an important
role in regulation of cell pro-liferation, differentiation and
death. RA and TH areknown to inhibit the proliferation of various
cancercells including breast cancer cells [53, 54]. Lack
ofsensitivity of advanced lung and breast cancer to RAis a
suggested mechanism for abnormal growth of can-cer cells [53].
rpL7a is a co-repressor for RAR andTHR by inhibiting RAR and
THR-mediated transcrip-tion of target genes [52]. Up-regulation of
rpL7a byethanol may diminish the sensitivity of breast cancercells
to RA and THR and thereby result in disruptionof growth
control.
The precise mechanism(s) by which ethanol mod-ulates the
expression of rpL7a is not clear. Ethanolunder certain
circumstances induces DNA damage[5557]. It is likely that ethanol
activation of rpL7ais triggered by DNA damage. Alternatively,
ethanolexposure alters the levels of circulating estrogen
andup-regulate the expression of estrogen receptors (ER)in breast
cancer cells [5860]. It is noted that the5-flanking sequence of
rpL7a gene contains severalimperfect estrogen responsive elements
(ERE). Recent
-
36 Y Zhu et al.
study suggests that ER may interact with these im-perfect or
half-sites of ERE and subsequently activatethe regulated genes
[61]. Therefore, ethanol may in-directly regulate the transcription
of rpL7a gene byaltering ER levels.
Although the exact mechanisms by which chronicalcohol
consumption stimulates carcinogenesis and tu-mor development are
not known, experimental studiesin animals support the concept that
ethanol alone isnot a carcinogen, but a co-carcinogen and/or a
tumorpromoter [62, 63]. Ethanol exposure is shown to en-hance the
metastasis and aggressiveness of mammarytumor in rat [64]. Recent
studies using cultured hu-man breast cancer cells confirm that
ethanol promotesproliferation, migration and invasion of tumor
cells invitro [17, 60, 65]. The current study identifies
thatribosomal protein rpL7a is an ethanol-regulated genein breast
cancer cells. rpL7a has been suggested tobe involved in
tumorogenesis and growth of cancercells. Therefore, modulation of
rpL7a expression maybe a mechanism underlying the promoting effect
ofethanol on cancer development. Further exploration ofrpL7a
function in malignant transformation and can-cer progression should
provide important insights intonot only the target of ethanol
toxicity but also themechanisms of carcinogenesis.
Acknowledgements
We thank Drs. Frank Reilly and Michael Miller fortheir critical
reading of this manuscript. This researchwas supported by grants
from the National Institutesof Health (AA12968 and CA 90385).
References
1. Kelsey J, Horn-Ross PL: Breast cancer: magnitude of
theproblem and descriptive epidemiology. Epidemiol Rev 15:716,
1993
2. Doll R, Peto R: The causes of cancer: quantitative
estimatesof avoidable risk of cancer in the United States today. J
NatlCancer Inst 66: 11921309, 1981
3. Mufti SI: International society for biomedical research on
al-coholism: relationship of cell necrosis and proliferation,
freeradicals and other agents to alcohol-related cancers. J
CancerRes Clin Oncol 119: 304305, 1993
4. Tnnesen H, Mller H, Andersen JR, Jensen E, Juel K:
Cancermorbidity in alcohol abusers. British J Cancer 69:
327332,1994
5. Willam RR, Horm JW: Association of cancer sites with to-bacco
and alcohol consumption and socioeconomic status of
patient: interview study from Third National Cancer Survey.
JNatl Cancer Inst 58: 12981305, 1977
6. Hiatt R: Alcohol consumption and breast cancer. Med
OncolTumor Pharmacother 7: 143151, 1990
7. Plant ML: Alcohol and breast cancer: a review. Intl J
Addict27: 107128, 1992
8. Rosenberg L, Metzger LS, Palmer JR: Alcohol consumptionand
risk of breast cancer: a review of epidemiologic evidence.Epidemiol
Rev 15: 133144, 1993
9. Longnecker M: Alcohol beverage consumption in relation torisk
of breast cancer: meta-analysis and review. Cancer CausesControl 5:
7382, 1994
10. Weiss HA, Brinton LA, Brogan D, Coates RJ, Gammon MD,Malone
KE, Schoenberg JB, Swanson CA: Epidemiology ofin situ and invasive
breast cancer in women aged under 45.British J Cancer 73: 12981305,
1996
11. Bowlin SJ, Leske MC, Varma A, Nasca P, Weinstein A,Caplan L:
Breast cancer risk and alcohol consumption: resultsfrom a large
case-control study. Intl J Epidemiol 26: 915923,1997
12. Swanson CA, Coates RJ, Malone KE, Gammon MD, Schoen-berg JB,
Brogan DJ, McAdams M, Potischman N, HooverRN, Brinton LA: Alcohol
consumption and breast cancer riskamong women under age 45 years.
Epidemiol 8: 231237,1997
13. Smith-Warner SA, Spiegelman D, Yaun SS, Vandenbrandt
PA,Folsom AR, Goldbohm RA, Graham S, Holmberg L, HoweGR, Marshall
JR, Miller AB, Potter JD, Speizer FE, WillettWC, Wolk A, Hunter DJ:
Alcohol and breast cancer in women a pooled analysis of cohort
studies. JAMA 279: 535540,1998
14. Vaeth PAC, Schlessinger WA: Alcohol consumption andbreast
cancer stage at diagnosis. Alcohol Clin Exp Res 22:928934, 1998
15. Kuper H, Ye W, Weiderpass E, Ekbom A, Trichopoulos D,Nyren
O, Adami HO: Alcohol and breast cancer risk: thealcoholism paradox.
Br J Cancer 83: 949951, 2000
16. Keydar I, Chen L, Karby S, Weiss FR, Delarea J, Radu
M,Brenner HJ: Establishment and characterization of a cell lineof
human breast carcinoma origin. Eur J Cancer 15: 650670,1979
17. Luo J, Miller MW: Ethanol enhances erbB-mediated migra-tion
of human breast cancer cells in culture. Breast Cancer ResTreat 63:
6169, 2000
18. Adickes ED, Mollner TJ, Lockwood SK: Closed chambersystem
for delivery of ethanol to cell culture. Alcohol 23:377381,
1988
19. Luo J, Miller MW: Ethanol inhibits basic fibroblast
growthfactor-mediated proliferation of C6 astrocytoma cells.
JNeurochem 67: 14481456, 1996
20. Miller MW, Nowakowski RS: Effect of prenatal exposure
toethanol on the cell cycle kinetics and growth fraction in
theproliferative zones of fetal rat cerebral cortex. Alcohol
ClinExp Res 15: 229232, 1991
21. Luo J, Miller MW: Basic fibroblast growth factor- and
platelet-derived growth factor-mediated cell proliferation in
B104neuroblastoma cells: effects of ethanol on cell cycle
kinetics.Brain Res 770: 139150, 1997
22. Liu W-M, Chu W-M, Choudary PV, Schmid CW: Cell stressand
translational inhibitors transiently increase the abundanceof
mammalian SINE transcripts. Nucleic Acids Res 23: 17581765,
1995
23. Liang P, Averboukh L, Keyomarsi K, Sager R, Pardee
AB:Differential display and cloning of messenger RNAs from hu-
-
Ethanol alters the expression of ribosomal protein 37man breast
cancer versus mammary epithelial cells. CancerRes 52: 69666968,
1992
24. Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ:
Basiclocal alignment search tool. J Mol Biol 215: 403410, 1990
25. Dani C, Blanchard JM, Piechaczyk M, EL Sabouty S, MartyL,
Jeanteur PH: Extreme instability of myc mRNA in normaland
transformed human cells. Proc Natl Acad Sci USA 81:70467050,
1984
26. Ausubel FM, Brent R, Kingston RE, Moore DD, Smith JA,Seidman
JS, Struhl K (eds): Current Protocols in MolecularBiology. Wiley,
New York, 1994
27. Hahn GM, Shiu EC, Auger EA: Mammalian stress proteinsHSP70
and HSP28 co-induced by nicotine and either ethanolor heat. Mol
Cell Biol 11: 60346040, 1991
28. Su CY, Chong KY, Owen OE, Dillmann WH, Chang C, LaiCC:
Constitutive and inducible hsp70s are involved in oxidat-ive
resistance evoked by heat shock and ethanol. J Mol CellCardiol 30:
587598, 1998
29. Li T-H, Spearow J, Rubin CM, Schmid CW:
Physiologicalstresses increase mouse short interspersed element
(SINE)RNA expression in vivo. Gene 239: 367372, 1999
30. Wool IG: Extraribosomal functions of ribosomal
proteins.Trends Biochem Sci 21: 164165, 1996
31. Frodin M, Gammeltoft S: Role and regulation of 90 kDa
ri-bosomal S6 kinase (RSK) in signal transduction. Mol
CellEndocrinol 151: 6577, 1999
32. Noara H, Naora H: Involvement of ribosomal proteins in
regu-lating cell growth and apoptosis: translational modulation
orrecruitment for extraribosomal activity? Immunol Cell Biol77:
197205, 1999
33. Chiao PJ, Shin DM, Sacks PG, Hong WK, Tainsky MA: El-evated
expression of the ribosomal protein S2 gene in humantumors. Mol
Carcinogenesis 5: 219231, 1992
34. Barnard GF, Staniunas RJ, Mori M, Puder M, Jessup MJ,Steele
GD Jr, Chen LB: Gastric and hepatocellular carcinomasdo not
over-express the same ribosomal protein messengerRNAs as colonic
carcinoma. Cancer Res 53: 40484052, 1993
35. Wong JM, Mafune K, Yow H, Rivers EN, Ravikumar TS,Steele GD
Jr, Chen LB: Ubiquitin-ribosomal-protein S27agene over expressed in
human colorectal carcinoma is an earlygrowth-responsive gene.
Cancer Res 53: 19161920, 1993
36. Wang Y, Cheong D, Chan S, Hooi SC: Ribosomal protein L7agene
is up-regulated but not fused to the tyrosine kinase re-ceptor as
chimeric trk oncogen in human colorectal carcinoma.Int J Oncol 16:
757762, 2000
37. Kondoh N, Schweinfest CW, Henderson KW, Papas TS:
Dif-ferential expression of S19 ribosomal protein,
laminin-bindingprotein, and human lymphocyte antigen class I
messengerRNAs associated with colon carcinoma progression and
dif-ferentiation. Cancer Res 52: 791796, 1992
38. Clausse N, Jackers P, Jares P, Joris B, Sobel ME,
CastronovoV: Identification of the active gene coding for the
metastasis-associated 37LRP/p40 multifunctional protein. DNA Cell
Biol15: 10091023, 1996
39. Barnard GF, Staniunas RJ, Bao S, Mafune K, Steele GDJr,
Gollan JL, Chen LB: Increased expression of human ri-bosomal
phosphoprotein P0 messenger RNA in hepatocellularcarcinoma and
colon carcinoma. Cancer Res 52: 30673072,1992
40. Chester KA, Robson L, Begent RHJ, Talbot IC, Pringle
JH,Primrose L, Macpherson AJ, Boxer G, Southall P, MalcolmAD:
Identification of a human ribosomal protein mRNA withincreased
expression in colorectal tumors. Biochim BiophysActa 1009: 297300,
1989
41. Pogue-geile K, Geiser JR, Shu M, Miller C, Wool IG,Meisler
AI, Pipas JM: Ribosomal genes are over-expressed incolorectal
cancer: isolation of a cDNA encoding human S3ribosomal protein. Mol
Cell Biol 11: 38423849, 1991
42. Kroes RA, Jastrow A, Mclone MG, Yamamoto H, Colley P,Kersey
DS, Yong VW, Mkrdichian E, Cerullo L, Leestma J,Moskal JR: The
identification of novel therapeutic targets forthe treatment of
malignant brain tumors. Cancer Lett 156:191198, 2000
43. Henry JL, Coggin DL, King CR: High expression of the
ri-bosomal protein L19 in human breast tumors that
over-expresserbB-2. Cancer Res 53: 14031408, 1993
44. Vaarala MH, Porvari KS, Kyllnen AP, Mustonen MVJ,Lukkarinen
O, Vihko P: Several genes encoding ribosomalproteins are
over-expressed in prostate-cancer cell lines: con-firmation of L7a
and L37 over-expression in prostate-cancertissue samples. Int J
Cancer 78: 2732, 1998
45. Ben-Ishai R, Scharf R, Sharon R, Kapten L: A human cel-lular
sequence implicated in trk oncogene activation is DNAdamage
inducible. Proc Natl Acad Sci USA 87: 60396043,1990
46. Strickland PT: Photocarcinogenesis by near-ultraviolet
(UVA)radiation in Sencar mice. J Invest Dermatol 87: 272275,
1986
47. Noonan FP, Otsuka T, Bang S, Anver MR, Merlino G:
Acceler-ated ultraviolet radiation-induced carcinogenesis in
hepatocytegrowth factor/scatter factor transgenic mice. Cancer Res
60:37383743, 2000
48. Moustacchi E: DNA damage and repair: consequences
ondose-responses. Mutat Res 464: 3540, 2000
49. Gupta RC, Lutz WK: Background DNA damage for endo-genous and
unavoidable exogenous carcinogens: a basis forspontaneous cancer
incidence? Mutat Res 4: 18, 1999
50. Kozma SC, Redmond SMS, Xiao-Chang F, Saurer SM, GronerB,
Hynes NE: Activation of the receptor kinase domain ofthe trk
oncogene by recombination with two different cellularsequences.
EMBO J 7: 147154, 1988
51. Chazenbalk GD, Wadsworth HL, Rapoport B: Tyrotropin-induced
expression of a gene for a ribosomal protein relatedto the trk
oncogen. Mol Cell Endocrinol 68: 2530, 1990
52. Burris T, Nawaz Z, Tsal M-J, OMalley BW: A nuclear hor-mone
receptor-associate protein that inhibits transactivationby the
thyroid hormone and retinoic acid receptors. Proc NatlAcad Sci USA
92: 95259529, 1995
53. Zhang XK, Liu Y, Lee MO: Retinoid receptors in human lungand
breast cancer. Mutat Res 350: 267277, 1996
54. Martinez MB, Ruan M, Fitzpatrick LA: Altered response
tothyroid hormones by prostate and breast cancer cells.
CancerChemother Pharmacol 45: 93102, 2000
55. Brook PJ: DNA damage, DNA repair, and alcohol toxicity
areview. Alcohol Clin Exp Res 21: 10731082, 1997
56. Wright RM, McManaman JL, Repine JE: Alcohol-inducedbreast
cancer: a proposed mechanism. Free Radic Biol Med26: 348354,
1999
57. Navasumrit P, Ward TH, Dodd NJ, OConnor PJ: Ethanol-induced
free radicals and hepatic DNA strand breaks areprevented in vivo by
antioxidants: effects of acute and chronicethanol exposure.
Carcinogenesis 21: 9399, 2000
58. Purohit V: Moderate alcohol consumption and estrogen
levelsin postmenopausal women: a review. Alcohol Clin Exp Res22:
994997, 1998
59. Fan S, Meng Q, Gao B, Grossman J, Yadegarri M, GoldbergID,
Rosen EM: Alcohol stimulates estrogen receptor signalingin human
breast cancer cells lines. Cancer Res 60: 56355639,2000
-
38 Y Zhu et al.
60. Singletary K, Frey R, Yan W: Effect of ethanol on
proliferationand estrogen receptor-alpha expression in human breast
cancercells. Cancer Lett 165: 131137, 2001
61. Petit FG, Metivier R, Valotaire Y, Pakdel F:
Synergismbetween a half-site and imperfect estrogen-responsive
ele-ment, and cooperation with COUP-TFI are required for estro-gen
receptor I (ER) to activate a maximal estrogen-stimulationof
rainbow trout ER gene. Eur J Biochem 259: 385395, 1999
62. Singletary K: Ethanol and experimental breast cancer:
areview. Alcohol Clin Exp Res 21: 334339, 1997
63. Seitz HL, Pschl G, Simanowski UA: Alcohol and cancer.
In:Galanter (ed) The Consequence of Alcoholism. Plenum Press,New
York, Vol 14, 1998, pp 395
64. Yirmya R, Ben-Eliyahu S, Gale R, Shavit Y, Liebeskind
J,Taylor A: Ethanol increases tumor progression in rats: pos-sible
involvement of natural killer cells. Brain Behav Immun6: 7486,
1992
65. Meng Q, Gao B, Goldberg ID, Rosen EM, Fan S: Stimulationof
cell invasion and migration by alcohol in breast cancercells.
Biochem Biophys Res Commun 273: 448453, 2000
Address for offprints and correspondence: Jia Luo, Ph.D.,
Depart-ment of Anatomy, West Virginia University School of
Medicine,Robert C. Byrd Health Science Center, Morgantown, WV
26506-9128, USA; Tel.: 304-293-0608; Fax: 304-293-8159;
E-mail:[email protected]