Embed Size (px)
Citation preview

Modern interpretation of the results of psychophysical studies of the threshold of visualperception of light
A. S. Tibilova)
NPK S. I. Vavilov State Optical Institute, St. Petersburg
Yu. E. Shelepin
I. P. Pavlov Institute of Physiology, Russian Academy of Sciences, St. Petersburg
(Submitted July 6, 2011)Opticheskiı̆ Zhurnal 78, 42–49 (December 2011)
This paper shows that there currently are difficulties with the traditional interpretation ofpsychophysical experiments for determining the energy threshold of light perception. Thefunctioning of the retina in the threshold regime is analyzed using models that give a newinterpretation of these tests and that take into account the achievements of physiological studiesof threshold vision. c© 2011 Optical Society of America.
INTRODUCTION
A period of intensive psychophysical studies of thresholdvisual perception occurred between the 1930s and the 1950s.1)
Since the 1970s, other, new methods (electrophysiological,genetic, biochemical, etc.) have become the fundamentalmethods for studying this still interesting problem.2,3 As timepassed, the model within which the results of psychophysicaltests were interpreted started to conflict with the data obtainedby these new methods and certain new data on vision. Theknown methodological problem makes it hard to accuratelycorrect this model. For obvious reasons, the main object ofthe new studies was the retina of animals rather than thehuman retina. However, it is difficult to organize behavioralexperiments with animals similar to psychophysical tests atsuch low intensities. In this article, we ignore this method-ological problem to some extent but depend on establishedconcepts that the characteristics of the human retina are similarto those of primates. We assume two hypothetical models ofthe threshold operating regime of the retina, in which, besidesthe results of psychophysical experiments, we use the resultsof measurements of the electrophysiological characteristics ofthe retinal receptors of primates for quantitative estimates.The purpose of developing these models is not only “to makeboth ends meet” in the modern interpretation of the results ofthe psychophysical tests but also to increase the role of theseresults when experiments in the physiology of threshold visionare being discussed and planned.
PSYCHOPHYSICAL TESTS ON DETERMINING THE ENERGYTHRESHOLD OF THE PERCEPTION OF LIGHT
Before explaining the essence of the proposed models,it is necessary, considering the wide range of readers ofOpticheskiı̆ Zhurnal, to give a brief, simplified review ofthe results of psychophysical studies of threshold vision.Here we shall rely on two classical papers: the most oftencited experimental study, carried out by Hecht et al. in1942,4 and the theoretical interpretation of the results of thepsychophysical studies, given by Barlow in 1956.5
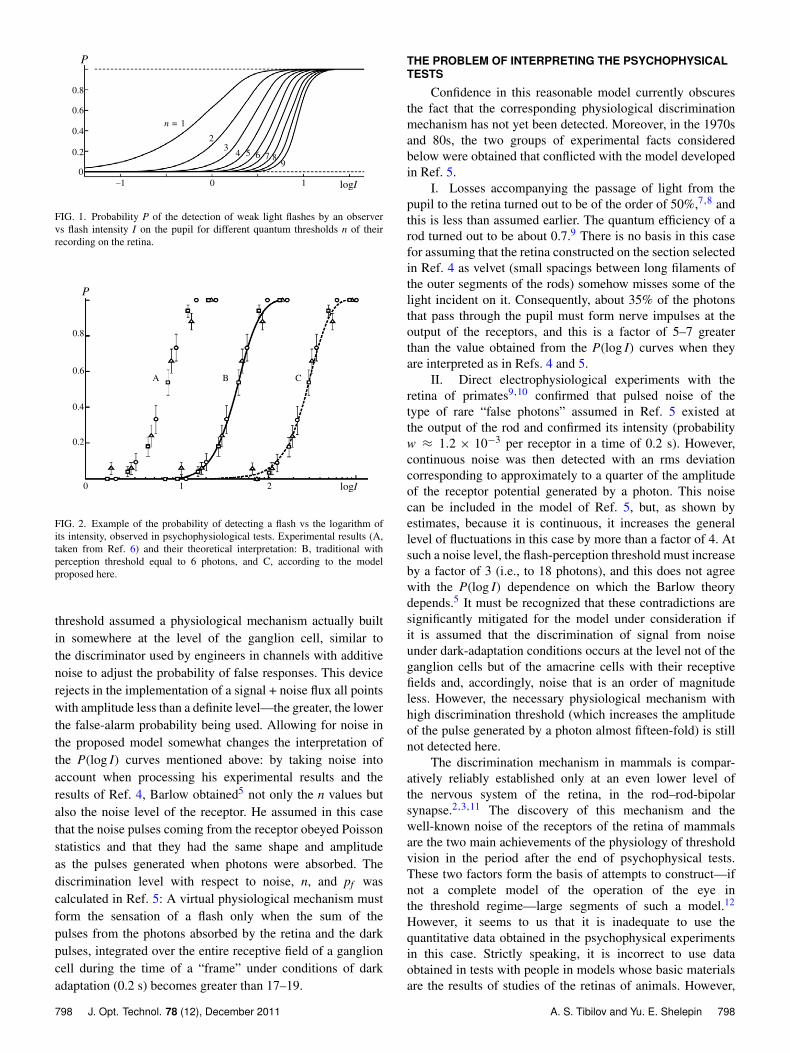
It was reliably established rather early that the energythreshold of detecting a light flash corresponds to an energyof about 100 photons at the input to the observer’s pupil.However, concepts that light losses in the eye were very highprevailed at that time (even though there was no reliable ex-perimental evidence of this because it was difficult to make thecorresponding measurements), and the idea arose of detectingthe recording of separate photons at the retinal level. Assumingthat the photons obeyed Poisson statistics, an elegant methodwas developed for determining the light-perception thresholddirectly on the retina, for which it is sufficient to plot howthe probability P that the observer will detect a light flashdepends on the logarithm of the energy I of the flash at theinput to the pupil. The shape of such a curve depends onlyon the threshold value of the number n of photons that causethe observer to have the sensation of a flash. Figure 1, takenfrom Ref. 4, shows a set of these logistic curves for variousvalues of n. Losses when light passes through the eye haveno effect on the shape of the curve but can be determinedfrom its position relative to the vertical axis. This methodwas successfully developed by various groups of investigators,reaching a high level of perfection, including in Ref. 4, whereindividual values of n from 5 to 7 were obtained for differentobservers (an example of experimental data from Ref. 6 isshown in Fig. 2, curve A). This result agreed with the results ofmost other investigations of that time. Three features intrinsicto the experiments in Ref. 4 are important in what follows:
• The mean threshold energy I0 of the light pulse at the inputinto the pupil equals 112 photons.• The light from the flash was focused on a section of the
retina that contains about 500 rods.• The probability pf of the observer incorrectly recording a
flash when it is omitted without the observer’s knowledgeequals 0.005.
The finite value of the probability of a false flash wasevidence that dark noise is present in the visual system.The model proposed in Ref. 5 for forming the perception
797 J. Opt. Technol. 78 (12), December 2011 1070-9762/2011/120797-06/$15.00 c© 2011 Optical Society of America 797
0.8
0.6
0.4
0.2
0–1
n = 1
23
4 5 6 7 89
P
logI
FIG. 1. Probability P of the detection of weak light flashes by an observervs flash intensity I on the pupil for different quantum thresholds n of theirrecording on the retina.
0.2
210
0.4
0.6
0.8
0.2
P
logI
A B C
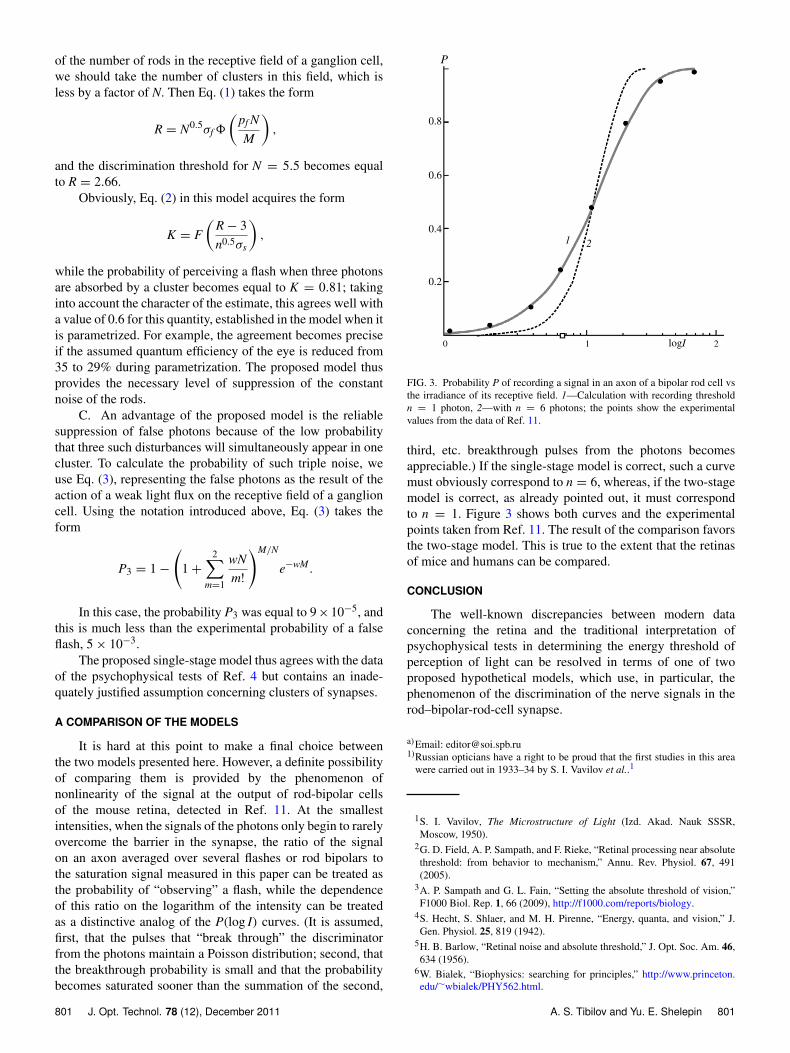
FIG. 2. Example of the probability of detecting a flash vs the logarithm ofits intensity, observed in psychophysiological tests. Experimental results (A,taken from Ref. 6) and their theoretical interpretation: B, traditional withperception threshold equal to 6 photons, and C, according to the modelproposed here.
threshold assumed a physiological mechanism actually builtin somewhere at the level of the ganglion cell, similar tothe discriminator used by engineers in channels with additivenoise to adjust the probability of false responses. This devicerejects in the implementation of a signal + noise flux all pointswith amplitude less than a definite level—the greater, the lowerthe false-alarm probability being used. Allowing for noise inthe proposed model somewhat changes the interpretation ofthe P(log I) curves mentioned above: by taking noise intoaccount when processing his experimental results and theresults of Ref. 4, Barlow obtained5 not only the n values butalso the noise level of the receptor. He assumed in this casethat the noise pulses coming from the receptor obeyed Poissonstatistics and that they had the same shape and amplitudeas the pulses generated when photons were absorbed. Thediscrimination level with respect to noise, n, and pf wascalculated in Ref. 5: A virtual physiological mechanism mustform the sensation of a flash only when the sum of thepulses from the photons absorbed by the retina and the darkpulses, integrated over the entire receptive field of a ganglioncell during the time of a “frame” under conditions of darkadaptation (0.2 s) becomes greater than 17–19.
THE PROBLEM OF INTERPRETING THE PSYCHOPHYSICALTESTS
Confidence in this reasonable model currently obscuresthe fact that the corresponding physiological discriminationmechanism has not yet been detected. Moreover, in the 1970sand 80s, the two groups of experimental facts consideredbelow were obtained that conflicted with the model developedin Ref. 5.
I. Losses accompanying the passage of light from thepupil to the retina turned out to be of the order of 50%,7,8 andthis is less than assumed earlier. The quantum efficiency of arod turned out to be about 0.7.9 There is no basis in this casefor assuming that the retina constructed on the section selectedin Ref. 4 as velvet (small spacings between long filaments ofthe outer segments of the rods) somehow misses some of thelight incident on it. Consequently, about 35% of the photonsthat pass through the pupil must form nerve impulses at theoutput of the receptors, and this is a factor of 5–7 greaterthan the value obtained from the P(log I) curves when theyare interpreted as in Refs. 4 and 5.
II. Direct electrophysiological experiments with theretina of primates9,10 confirmed that pulsed noise of thetype of rare “false photons” assumed in Ref. 5 existed atthe output of the rod and confirmed its intensity (probabilityw ≈ 1.2 × 10−3 per receptor in a time of 0.2 s). However,continuous noise was then detected with an rms deviationcorresponding to approximately to a quarter of the amplitudeof the receptor potential generated by a photon. This noisecan be included in the model of Ref. 5, but, as shown byestimates, because it is continuous, it increases the generallevel of fluctuations in this case by more than a factor of 4. Atsuch a noise level, the flash-perception threshold must increaseby a factor of 3 (i.e., to 18 photons), and this does not agreewith the P(log I) dependence on which the Barlow theorydepends.5 It must be recognized that these contradictions aresignificantly mitigated for the model under consideration ifit is assumed that the discrimination of signal from noiseunder dark-adaptation conditions occurs at the level not of theganglion cells but of the amacrine cells with their receptivefields and, accordingly, noise that is an order of magnitudeless. However, the necessary physiological mechanism withhigh discrimination threshold (which increases the amplitudeof the pulse generated by a photon almost fifteen-fold) is stillnot detected here.
The discrimination mechanism in mammals is compar-atively reliably established only at an even lower level ofthe nervous system of the retina, in the rod–rod-bipolarsynapse.2,3,11 The discovery of this mechanism and thewell-known noise of the receptors of the retina of mammalsare the two main achievements of the physiology of thresholdvision in the period after the end of psychophysical tests.These two factors form the basis of attempts to construct—ifnot a complete model of the operation of the eye inthe threshold regime—large segments of such a model.12
However, it seems to us that it is inadequate to use thequantitative data obtained in the psychophysical experimentsin this case. Strictly speaking, it is incorrect to use dataobtained in tests with people in models whose basic materialsare the results of studies of the retinas of animals. However,
798 J. Opt. Technol. 78 (12), December 2011 A. S. Tibilov and Yu. E. Shelepin 798

this is not such a great sin even if it is taken into accountthat not many measurements of the noise of the receptors ofthe human retina conformed well to the noise structure of thereceptors of primates. There is even less of a basis for doubtingthe presence of a discrimination barrier in the rod–rod-bipolarsynapse in man. Therefore, taking the electrophysiologicaldata on the noise of the receptors in primates from a singlereliable source,9 we propose below two versions of theuse of the phenomenon of signal discrimination when thesignal is transmitted through the rod–rod-bipolar synapse toexplain, most importantly, the results of the psychophysicalexperiments.
THE TWO-STAGE MODEL
In the first model, we start from an estimate of how muchnoise discrimination is needed in the synapse to reduce thefalse-signal probability to the value obtained in Ref. 4. Takingthe mean signal amplitude as unity and initially using thefalse-flash probability pf (5×10−3), the rms deviation σf of thenoise (0.25), and the number M of receptors in the receptivefield of the ganglion cell (104), we estimate the discriminationthreshold from
R = σf8(pf
M
), (1)
where8 is the normal dispersion corresponding to probabilitypf /M. It turned out that R = 1.22. Now, knowing the rmsdeviation σs for the signal (0.22) and using the probabilityintegral F, it is possible to use R to estimate the probabilityK that the signal will exceed the discrimination threshold,
K = F
(R− 1σs
). (2)
In this case, K turned out to be equal to 0.16, and thisguarantees that, on the average, every sixth photon from thoseabsorbed by the retina after a flash will exceed the barrier. Ifwe assume that this is the photon that causes the sensation ofa flash, two problems arise.
First, the “false-photon” problem is not solved with thiskind of discrimination. Actually, the probability of one rodgenerating such a photon in time 0.2 s is w = 1.2× 10−3 s−1,and there are twelve false flashes indistinguishable fromphoton signals in the entire receptive field of the ganglion cellduring this time. The discriminator considered above reducesthe number of signals from that number to two, but this is400 times as great as the experimentally determined false-flashprobability.4
Second, with such a simple mechanism, the P(log I) curvemust correspond to n = 1, and not n = (5–7), as in Ref. 4.(The photons absorbed by the receptors but not transmittedthrough the barrier in the synapse do not differ at all, fromthe viewpoint of the interpretation of the P(log I) curves, fromthe photons lost in the other parts of the eye: They only shiftthe curve without affecting its shape). Thus, it is not possibleto create a model that corresponds to the entire collection ofresults of the psychophysical experiments on the basis of onediscriminator of the type under consideration.
It is possible to escape this situation by combining thecontinuous-noise-filtering mechanism discussed here with theold model5 of just one discriminator operating at higher levelsof the retina. Let us consider what parameters must appear insuch a system in the second discriminators that operate at thelevel of the outputs of the ganglion (G), amacrine (A), andbipolar (rod) (B) cells. Initially assuming that the false photonsthat get through the first discrimination barrier have a Poissondistribution, we determine the discrimination level md neededfor the observed false-signal probability pf = 5 × 10−3 fromthe equation
pf = k∞∑md
(KwMk
)mm!
exp(−
KwM
k
),
where k is the ratio of the size of the corresponding receptivefield to the size of the receptive field of the ganglion cell.
Knowing md, it is possible to estimate the flash-perceptionthreshold np with probability ps = 0.6 from the formula
ps =
∞∑md
(KwMk + np
)mm!
exp[−
(KwM
k+ np
)].
When k = 1 (the case in which the discriminator islocated at the output of a ganglion cell), this gives md = 7and np = 5.5 (i.e., five or six photons); when k = 20 (the caseof an amacrine cell), md = np = 3, and, finally, for k = 400(the case of a bipolar cell), md = np = 2.
It is possible to estimate from np what quantum efficiencyJ of the retina corresponds to each of the three cases underconsideration of the location of the second discriminator.Since, in Hecht’s experiments,4 a section of the retina wasilluminated corresponding to the size of the receptive fieldof an amacrine cell, for the first two cases it is sufficient todivide np by the probability of a signal from a photon passingthrough the first discriminator (0.16) and the threshold energyof a light pulse at the input of the pupil measured in Ref. 4 (112photons). For the third case (when the second discriminator islocated at the output of a bipolar), one must have recourse tothe Brindley formula15 [see Eq. (3)]. The corresponding valuesof J turned out to equal JG = 0.31, JA = 0.17, and JB = 0.32.
These results show that, among the possible locationsof the second discriminator considered above, the case inwhich this discriminator is located somewhere at the outputof a ganglion cell or outside the retina correspond best to theresults of the psychophysical studies and to modern conceptsconcerning the energy efficiency of the retina. The values of npequal to 3 and 2 obtained in the second and third cases differtoo strongly from the value of the perception threshold of theretina of 5–7 photons obtained in Ref. 4.
It should be recognized that this conclusion contradictsthe result obtained in Ref. 13 when investigating how thebackground illumination affects the perception threshold ofweak flashes. It is shown in that paper that the mechanismthat increases the threshold as the illumination increases islocalized between the rod bipolar and the amacrine cell.The flux of “false photons” does not differ from the flux ofillumination photons, and its effect on the flash-perception
799 J. Opt. Technol. 78 (12), December 2011 A. S. Tibilov and Yu. E. Shelepin 799

threshold must be adjusted by the same mechanism and in thesame place, for which np = 2 in our model.
In connection with this discrepancy, it is appropriateto recall one phenomenon that was not explained earlier.1,5
When a red light point that fixes the observer’s glance in thepsychophysical tests is transferred either far from the sectionof the retina illuminated by the flash1,14 or altogether to theother eye,15 the perception threshold decreases sharply, tonp = 2. Now, when it is known that the output of the amacrinecell is directly associated with the output of a rod bipolar, itis inappropriate to locate the red point close to the section ofthe retina illuminated by the flash in Ref. 4 and in most otherpapers. Vavilov’s appeal to check this technique1 remainscrucial.
Comparing the characteristics obtained for the seconddiscriminator (we assume at the level of the ganglion cell) withthe characteristics of the discriminator considered in Ref. 5, weshould point out the following favorable differences that arisefrom the former discriminator:
• It is now protected from the continuous noise of thereceptors.
• The discrimination threshold needed to combat falsephotons has been reduced by almost a factor of 3.
• The losses when light passes through the eye assumed bythis model have come into agreement with the actual losses.
Although the hierarchy of the discriminators somewhatcorresponds to the hierarchical structure of the nervous systemof the retina itself, by adopting it, we in some sense turn back60 years, assuming an unknown physiological mechanism forthe second discriminator. This motivates us to propose yetanother threshold-perception model in terms of which a singlediscrimination mechanism in the rod–rod-bipolar synapse canbe adequate to explain the results of the psychophysical tests.
THE SINGLE-STAGE MODEL
The first step in constructing the second model was toreject the paradigm assumed earlier by most investigatorswhen interpreting P(log I) curves, according to which the for-mation of the flash-perception signal, including distinguishingit from noise, occurs at the level of a receptive field that isequal to or exceeds the section of the retina illuminated bythe focused flash. Brindley’s broader calculational model16
was considered, which interprets these logistic curves on theassumption that the decision that a signal is present is madeat the level of a receptive field s that constitutes only a part ofthe region S of the retina illuminated by the flash. Accordingto Ref. 16, the probability Pn of observing a flash depends onthe amount of light I (in photons) in the flash, “converted” bythe retina into receptor signals, as
Pn = 1−
(1+
n−1∑m=1
(Is/S)m
m!
)S/s
e−I, (3)
where n is now the threshold number of photons that haveexcited the receptors and cause the sensation of a flash whenthey simultaneously fall on one of the rods s.
In the new, broad interpretation, the shape of the P(log I)curves depends not only on n, but also on the ratio s/S,with the interpretation of the P(log I) curves used in Refs. 4and 5 being a particular case of the model of Ref. 16 whens/S = 1. By varying the values of s/S within reasonable limits1 > s/S > 1/500), it is possible to determine for which nthe P(log I) curves calculated from Eq. (3) agree well in shapewith the experimental P(log I) curves from Ref. 4. It turned outthat, for n equal to 3, 4, and 5, it is possible to select a region ofS/s values that provides such agreement and is characterized,respectively, by S/s > 50, S/s < 5, and S/s < 2. Eachof these cases corresponds to its own threshold value of In
thr(corresponding to Pn = 0.6): I3
thr > 27, I4thr < 11, and I5
thr < 8photons. The modern requirement of high quantum efficiencyof the eye (35%, see above) thus corresponds only to the casen = 3, S/s = 90, and I3
thr = 39 (see Fig. 2, curve B).The ratio S/s = 90 corresponds to a group s composed
of 5–6 rods (a mean of N = 5.5). Assuming that this is afairly compact group, we get the following concept concerningthe process of threshold perception of a flash focused on theretina: The sensation of a flash occurs when three or moresignals from absorbed photons are generated on some compactsection composed of 5–6 receptors of the receptive field of arod bipolar. The result of this can be shown to be strange if theestablished concepts are not kept in mind concerning clustersof receptors of just the size which, because of the electric andchemical interaction of the closely spaced receptors, integratetheir signals and noise even at the input of the bipolar.16 Mostof the papers devoted to this phenomenon discuss clustersformed as a result of the appearance of temporary linksbetween the receptors. This process apparently has a complexand unstable character and never takes in the entire set ofreceptors. It is obvious that clusters of this type are not suitablefor the proposed model. A diffusion mechanism is more likelyto form the necessary type of clusters at the level not of the re-ceptors themselves but of their synapses. Clusters of synapseshave been detected in the brain cells17 and are explained by theintegrating property of free diffusion of the glutamate that fillsthe intercellular space and the somewhat closely spaced synap-tic clefts. As applied to our case, integration in the first approx-imation should be understood as equality of the glutamate con-centrations in all the synaptic clefts of the receptors of the clus-ter. This common concentration of glutamate will fall underthe action of light until, when three photons are simultaneouslyincident on the cluster, it reaches a critical value that removesthe inhibition in all the parts of this collective synapse.
Assuming this hypothesis concerning the nature of theclusters until the appropriate experimental data appear, weshall show that the model is effective from the viewpoint ofthe problem discussed here.
A. The model is acceptable when the quantum efficiencyis greater than 0.24; i.e., when it matches the modern conceptof optical losses in the eye.
B. In the same way as in the preceding model, weestimate the discrimination threshold from the false-signalprobability, taking into account that the rms deviation in thenoise of a “collective receptor” composed of N rods witha normal distribution is a factor of N0.5 larger than for anindividual receptor. Moreover, in integrating the noise, instead
800 J. Opt. Technol. 78 (12), December 2011 A. S. Tibilov and Yu. E. Shelepin 800
of the number of rods in the receptive field of a ganglion cell,we should take the number of clusters in this field, which isless by a factor of N. Then Eq. (1) takes the form
R = N0.5σf8
(pf N
M
),
and the discrimination threshold for N = 5.5 becomes equalto R = 2.66.
Obviously, Eq. (2) in this model acquires the form
K = F
(R− 3
n0.5σs
),
while the probability of perceiving a flash when three photonsare absorbed by a cluster becomes equal to K = 0.81; takinginto account the character of the estimate, this agrees well witha value of 0.6 for this quantity, established in the model when itis parametrized. For example, the agreement becomes preciseif the assumed quantum efficiency of the eye is reduced from35 to 29% during parametrization. The proposed model thusprovides the necessary level of suppression of the constantnoise of the rods.
C. An advantage of the proposed model is the reliablesuppression of false photons because of the low probabilitythat three such disturbances will simultaneously appear in onecluster. To calculate the probability of such triple noise, weuse Eq. (3), representing the false photons as the result of theaction of a weak light flux on the receptive field of a ganglioncell. Using the notation introduced above, Eq. (3) takes theform
P3 = 1−
(1+
2∑m=1
wN
m!
)M/N
e−wM.
In this case, the probability P3 was equal to 9×10−5, andthis is much less than the experimental probability of a falseflash, 5× 10−3.
The proposed single-stage model thus agrees with the dataof the psychophysical tests of Ref. 4 but contains an inade-quately justified assumption concerning clusters of synapses.
A COMPARISON OF THE MODELS
It is hard at this point to make a final choice betweenthe two models presented here. However, a definite possibilityof comparing them is provided by the phenomenon ofnonlinearity of the signal at the output of rod-bipolar cellsof the mouse retina, detected in Ref. 11. At the smallestintensities, when the signals of the photons only begin to rarelyovercome the barrier in the synapse, the ratio of the signalon an axon averaged over several flashes or rod bipolars tothe saturation signal measured in this paper can be treated asthe probability of “observing” a flash, while the dependenceof this ratio on the logarithm of the intensity can be treatedas a distinctive analog of the P(log I) curves. (It is assumed,first, that the pulses that “break through” the discriminatorfrom the photons maintain a Poisson distribution; second, thatthe breakthrough probability is small and that the probabilitybecomes saturated sooner than the summation of the second,
210
0.8
0.6
0.4
0.2
P
1 2
logI
FIG. 3. Probability P of recording a signal in an axon of a bipolar rod cell vsthe irradiance of its receptive field. 1—Calculation with recording thresholdn = 1 photon, 2—with n = 6 photons; the points show the experimentalvalues from the data of Ref. 11.
third, etc. breakthrough pulses from the photons becomesappreciable.) If the single-stage model is correct, such a curvemust obviously correspond to n = 6, whereas, if the two-stagemodel is correct, as already pointed out, it must correspondto n = 1. Figure 3 shows both curves and the experimentalpoints taken from Ref. 11. The result of the comparison favorsthe two-stage model. This is true to the extent that the retinasof mice and humans can be compared.
CONCLUSION
The well-known discrepancies between modern dataconcerning the retina and the traditional interpretation ofpsychophysical tests in determining the energy threshold ofperception of light can be resolved in terms of one of twoproposed hypothetical models, which use, in particular, thephenomenon of the discrimination of the nerve signals in therod–bipolar-rod-cell synapse.
a)Email: [email protected])Russian opticians have a right to be proud that the first studies in this area
were carried out in 1933–34 by S. I. Vavilov et al..1
1S. I. Vavilov, The Microstructure of Light (Izd. Akad. Nauk SSSR,Moscow, 1950).
2G. D. Field, A. P. Sampath, and F. Rieke, “Retinal processing near absolutethreshold: from behavior to mechanism,” Annu. Rev. Physiol. 67, 491(2005).
3A. P. Sampath and G. L. Fain, “Setting the absolute threshold of vision,”F1000 Biol. Rep. 1, 66 (2009), http://f1000.com/reports/biology.
4S. Hecht, S. Shlaer, and M. H. Pirenne, “Energy, quanta, and vision,” J.Gen. Physiol. 25, 819 (1942).
5H. B. Barlow, “Retinal noise and absolute threshold,” J. Opt. Soc. Am. 46,634 (1956).
6W. Bialek, “Biophysics: searching for principles,” http://www.princeton.edu/∼wbialek/PHY562.html.
801 J. Opt. Technol. 78 (12), December 2011 A. S. Tibilov and Yu. E. Shelepin 801

7E. A. Boettner and J. R. Wolter, “Transmission of the ocular media,” Invest.Ophthalmol. 1, 776 (1962).
8J. Van de Kraats and D. Van Norren, “Optical density of the aging humanocular media in the visible and the UV,” J. Opt. Soc. Am. A 24, 1842(2007).
9D. A. Baylor, B. J. Nunn, and J. L. Schnapf, “The photocurrent, noise andspectral sensitivity of rods of the monkey Macaca fascicularis,” J. Physiol.357, 575 (1984).
10G. D. Field and F. Rieke, “Mechanisms regulating variability of thesingle-photon responses of mammalian rod photoreceptors,” Neuron 35,733 (2002).
11G. D. Field and F. Rieke, “Nonlinear signal transfer from mouse rodsto bipolar cells and implications for visual sensitivity,” Neuron 34, 773(2002).
12M. C. Van Rossum and R. G. Smith, “Noise removal at the rod synapse ofmammalian retina,” Visual Neurosci. 15, 809 (1998).
13A. D. Felice, T. Doan, P. S. Alapakkam, and F. Rieke, “Controlling thegain of rod-mediated signals in the mammalian retina,” J. Neurosci. 26,3959 (2006).
14S. I. Vavilov, “Visual observation of light fluctuations,” Izv. Akad. NaukSSSR (Fiz. Ser.) No. 1/2, 176 (1936).
15H. A. Van der Velden, “The number of quanta necessary for the perceptionof light of the human eye,” Ophthalmologica 111, 321 (1946).
16G. S. Brindley, “The order of coincidence required for visual threshold,”Proc. Phys. Soc. London, Sect. B 67, 673 (1954).
17E. P. Hornstein, J. Verweij, P. H. Li, and J. L. Schnapf, “Gap-junctionalcoupling and absolute sensitivity of photoreceptors in macaque retina,” J.Neurosci. 25, 11201 (2005).
18A. Scimemi, A. Fine, D. M. Kullmann, and D. A. Rusakov, “NR2B-containing receptors mediate cross talk among hippocampal synapses,” J.Neurosci. 24, 4767 (2004).
802 J. Opt. Technol. 78 (12), December 2011 A. S. Tibilov and Yu. E. Shelepin 802





























