Embed Size (px)
Citation preview

Modeling the Electromobility of Type-I Collagen Molecules in the
Electrochemical Fabrication of Dense and Aligned Tissue Constructs
JORGE ALFREDO UQUILLAS1 and OZAN AKKUS
1,2,3,4
1Weldon School of Biomedical Engineering, Purdue University, 206 S. Martin Jischke Drive, West Lafayette, IN, USA;2Mechanical and Aerospace Engineering, Case Western Reserve University, 10900 Euclid Avenue, Cleveland, OH, USA;
3Biomedical Engineering, Case Western Reserve University, 10900 Euclid Avenue, Cleveland, OH, USA; and 4Department ofOrthopaedics, Case Western Reserve University, 11100 Euclid Avenue, Cleveland, OH, USA
(Received 27 September 2011; accepted 28 January 2012; published online 8 February 2012)
Associate Editor Laura Suggs oversaw the review of this article.
Abstract—Isoelectric focusing (IEF) of type-I collagen mol-ecules is a technology with proven efficacy to produce denseand aligned collagen-based biomaterials. The forces andmechanisms during IEF of collagen molecules in carrierampholyte-free environments remain unknown. This studypresents theoretical framework describing the congregationof collagen molecules along the isoelectric point (pI). A singlemolecule was modeled as a rod-like particle, distributedhomogeneously between parallel electrodes. Upon applica-tion of electrical current, molecules migrated to the pI. Theresults showed that self-aggregation of collagen moleculesalong the pI occurred due to formation of a non-linear pHgradient that rendered the anodic side acidic, and thecathodic side basic. This pH profile and the amphotericnature of collagen resulted in positively charged molecules atthe anode and negatively charged molecules at the cathode.Therefore, repulsive electrostatic forces aided self-aggrega-tion of molecules along the pI. The model could effectivelyvalidate the pI of collagen, the pI location, and predict thatthe instantaneous velocity acting on a molecule at the anodewas higher than those velocities at the cathode. Thisfundamental information represents the baseline theory uponwhich we can expand our knowledge to the production ofbiomaterials to engineer soft tissues.
Keywords—Mathematical model, Experimental validation,
Non-linear pH profile, Viscosity gradient, Electrostatic force,
Drag force, Biomaterials, Soft tissue engineering.
INTRODUCTION
Recently, research groups have focus their attentionto transform collagen solutions to condensed solid
state biomaterials based on the principles of isoelectricfocusing (IEF).1,9,14,21,23,39 While IEF is routinely usedfor protein identification and separation, its utilizationfor fabricating materials is novel and significant. Thesignificance of electrochemical fabrication is in itspotential to generate collagenous biomaterials whosefabric is as dense, as aligned and as strong as collagen-rich tissues such as tendons and ligaments.9,21,39 Spe-cifically, ELAC threads have shown mechanicalproperties converging to those of native tendon,9 haveshown to incorporate decorin-mimic molecules withinits structure,21 and have exhibited a periodic staggeredconformation in the form of d-banding pattern.39
Electrochemically aligned collagen threads have goodin vivo biocompatibility.22 Additionally, ELAC threadshave shown tenogenic differentiation of human mes-enchymal stem cells only by direct contact, without theneed of growth factor supplementation.20 The highlyaligned fabric orientation makes this material ideal forcontact guiding of cells and other groups have recentlypicked up this fabrication method to fabricate scaf-folds for directional neurite guidance.1 The methodinvolves the application of electric current to desaltedaqueous collagen solutions which in turn induces a pHgradient. Amphoteric collagen molecules assumevarying charges with changing pH, resulting in theelectromigration of molecules to the isoelectric point(pI). Unlike a typical IEF experiment where a stablepH gradient is created by electrophoretic mobility ofcarrier ampholytes to fixed zones,31 the pH gradient iscreated by the electrolysis of an aqueous environmentin collagen solutions. Therefore, it becomes difficult topredict the pH gradient profile. In the presence of anelectric field, the charged molecules acquire electro-phoretic mobility as dictated by the protein charge,
Address correspondence to Ozan Akkus, Mechanical and Aero-
space Engineering, Case Western Reserve University, 10900 Euclid
Avenue, Cleveland, OH, USA. Electronic mail: ozan.akkus@case.
edu, [email protected]
Annals of Biomedical Engineering, Vol. 40, No. 8, August 2012 (� 2012) pp. 1641–1653
DOI: 10.1007/s10439-012-0528-1
0090-6964/12/0800-1641/0 � 2012 Biomedical Engineering Society
1641
the direction of the electric field, and the drag forceimposed by the solution. However, the driving forcesbehind the focusing phenomena are not fullyunderstood. That is, the order of magnitude of elec-trostatic forces driving collagen molecules and theviscous drag forces countering the migration areunknown.
Computer simulations of IEF of proteins havedemonstrated significant importance as a researchtool.7,26,35 The complexity when dealing with macro-molecules arises with the lack of mathematical expres-sions to describe their ionization. The large number ofionisable groups in type-I collagen generates a consid-erable number of local net charges at a given pH thatneeds to be accounted for systematically. Additionally,most of the simulation work has dealt with low-molecular-mass species. However, several studies havesimulated the IEF of proteins. Mosher et al.28 predictedthe electrophoretic mobility of albumin which wasvalidated by others using transport computer algo-rithms.34 Modeling of two proteins with ten chargedstates under an electric field of 30,000 V/m within a pHrange of 6–9 was done by Shim et al.33 IEF of threepeptides: leucine enkephalin, angiotensin II, and apeptide from the human proteome digestion weremodeled in an IPG gel with pH gradient.24 The peptidecharge slope at the isoelectric point was determined tobe an important parameter in focusing dynamics todetermine the optimal separation-well width for bestfocusing.
Despite the promise of ELAC for repair of multipletissues, little is known about the mechanisms by whichthe congregation of molecules take place under theeffect of electrochemical forces. The pH, charge ofmolecules, electrostatic forces and viscous forces act inunison; however, their relative roles are unclear.Understanding the effects of process variables on theelectrochemical alignment process would allowimproving the fabrication method. For instance, col-lagen mobility, thus, the speed of IEF will be a func-tion of the electrode separation and the appliedvoltage. However, these are not simple linear rela-tionships and models predicting these relations wouldhelp reduce experimental work.
The aims of the current study were: (a) to estimatethe changes in the charge distribution of type-I colla-gen molecules at various pH values, (b) to define thepH profile during IEF and clarify its role duringmolecular migration and aggregation along the iso-electric point (pI), (c) to calculate the variation in theelectrostatic force acting on a collagen molecule usingcharge calculations, and (d) to predict the effects ofvoltage amplitude on the migration time of collagenmolecules.
MATERIALS AND METHODS
Electrochemical Cell Layout for IEF of Type-ICollagen Molecules
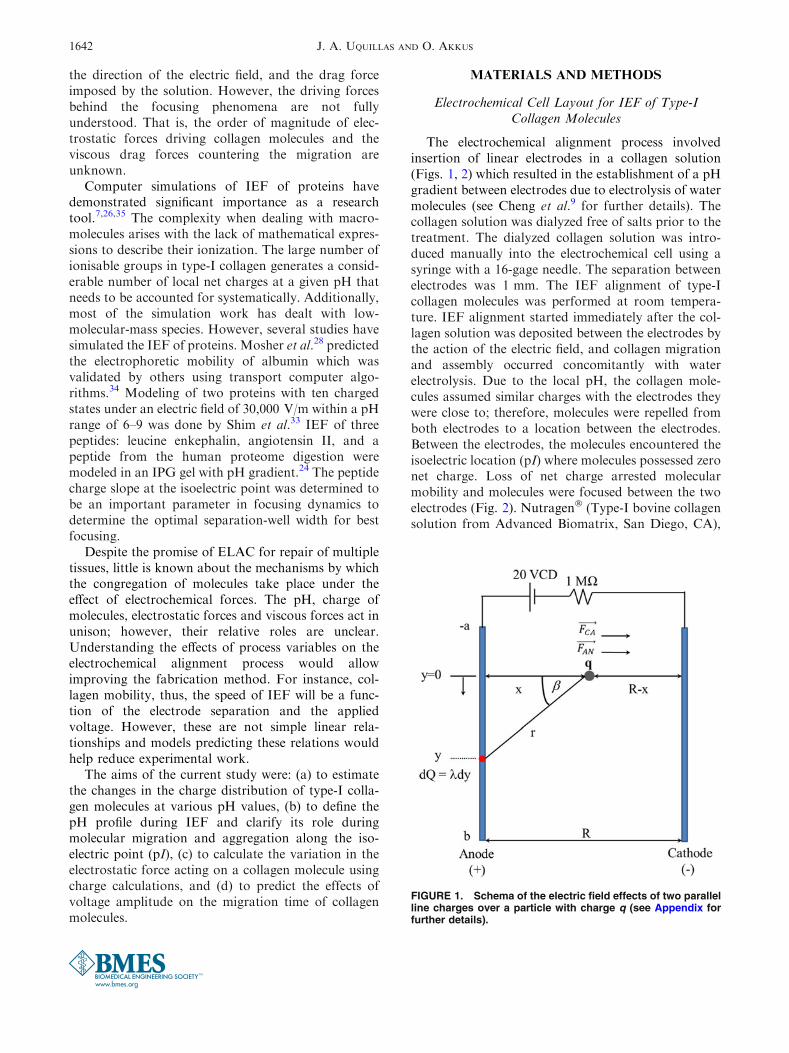
The electrochemical alignment process involvedinsertion of linear electrodes in a collagen solution(Figs. 1, 2) which resulted in the establishment of a pHgradient between electrodes due to electrolysis of watermolecules (see Cheng et al.9 for further details). Thecollagen solution was dialyzed free of salts prior to thetreatment. The dialyzed collagen solution was intro-duced manually into the electrochemical cell using asyringe with a 16-gage needle. The separation betweenelectrodes was 1 mm. The IEF alignment of type-Icollagen molecules was performed at room tempera-ture. IEF alignment started immediately after the col-lagen solution was deposited between the electrodes bythe action of the electric field, and collagen migrationand assembly occurred concomitantly with waterelectrolysis. Due to the local pH, the collagen mole-cules assumed similar charges with the electrodes theywere close to; therefore, molecules were repelled fromboth electrodes to a location between the electrodes.Between the electrodes, the molecules encountered theisoelectric location (pI) where molecules possessed zeronet charge. Loss of net charge arrested molecularmobility and molecules were focused between the twoelectrodes (Fig. 2). Nutragen� (Type-I bovine collagensolution from Advanced Biomatrix, San Diego, CA),
FIGURE 1. Schema of the electric field effects of two parallelline charges over a particle with charge q (see Appendix forfurther details).
J. A. UQUILLAS AND O. AKKUS1642

a pepsin-digested form of collagen having the telopep-tide regions cleaved, was used in all the experiments.
Net Charge of the Type-I Collagen Moleculesas a Function of pH
The net electrostatic force acting over a molecule isa function of the net charge of the molecule, which is in
turn a function of the pH of its environment. The netcharge of the molecule is the sum of the charges ofamino acid side chains. Amino acid sequences foralpha11 (a11), and alpha21 (a21) chains making up thecollagen molecule were obtained from the Uniprotdatabase.37,38 Using a custom-written code (Matlab,The MathWorks Inc., Natick, MA) the amino acidsequences in letters were converted to integer values toobtain a matrix of three columns, with each columncorresponding to one of the three collagen alpha chains[a11, a21, a11] (Fig. 3). The chains were staggered toalign glycine residues with the appropriate residues inneighboring chains. The charge in each amino acid wascalculated using the ionization constants (pKa)(Table 1) at a given pH value to obtain the chargedistribution along the molecule’s length. The net elec-trical charge of the molecule (q) was calculated by
FIGURE 2. Compensated polarized optical image of a colla-gen solution placed between two electrodes to achieve IEF.Collagen was congregated and assembled along a fixed band(double arrow). The blue color indicates that molecules areoriented parallel to the slow axis of the polarizer which is setparallel to the electrodes. dpI represents the distance betweenthe anode to the isoelectric line (pI). Scale bar 5 250 lm.
FIGURE 3. General approach to estimate the charge distribution along a type-I collagen molecule at pH 3.5. Alpha1 chain is 1053residues long, and alpha2 chain is 1020 residues long.
TABLE 1. List of ionization constants (pKa) of amino acidspresent in pepsin-treated type-I collagen molecules with
ionisable side chain.
Amino acid type pKa
Aspartic acid 3.9
Glutamic acid 4.3
Histidine 6.0
Cysteine 8.3
Tyrosine 10.07
Lysine 10.53
Arginine 12.48
Modeling the Electromobility of Type-I Collagen Molecules 1643

adding individual amino acid charges along the lengthof the molecule.
Forces Acting on the Type-I Collagen Molecule
The electrostatic force (FE) is dependent on theelectric field E and charge q of the particle in the field.The voltage DV between the electrodes needs to begreater than 1.7 VDC to enable electrolysis of watermolecules and the generation of a pH gradient. Theelectrode separation R was 1 mm, resulting in an elec-tric field of >1700 V/m.9 The migration of molecules,in bulk, was monitored using compensated polarizedimaging. In this method, the electrochemical cell wasplaced on the microscope stage (Olympus BX 51) andilluminated in transmission mode. The incoming lightwas polarized, passed through the sample, a gypsumwave plate and an analyzer before being captured in aCCD camera. The net effect of the compensated opticswas such that the molecules aligned along the fast axisof the wave plate assumed a blue color, enabling thevisualization of molecules in bulk.
The electric force (FE) acting on a particle due to acontinuously distributed line charge can be found bysuperimposing the point charge fields of infinitesimalcharge elements (see the Appendix for the derivation).
FE�!¼ Q�q
4�p�e�L�
8
>
>
<
>
>
:
b
x�ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
x2þb2ð Þp þ a
x�ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
x2þa2ð Þp
" #
þ b
R�xð Þffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
R�xð Þ2þb2� �
r þ a
R�xð Þffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
R�xð Þ2þa2� �
r
2
6
6
4
3
7
7
5
9
>
>
=
>
>
;
ð1Þ
Here e is the permittivity of the collagen mediumprovided in the literature27,36 at 5 times the electricconstant (e0). The force depends on the charge in theelectrodes (Q), the separation (x) between the elec-trodes and the collagen molecule, the separationbetween electrodes (R), the length of the electrodes (L),and the parameters a and b which represent locationsalong the electrodes.
The instantaneous velocity (vi, lm/s) of the collagenmolecule was calculated as17:
vi ¼FE
c� 1� e �t=sð Þ� �
ð2aÞ
where FE (pN) is the electrostatic force acting on thecollagen molecule, and s is the time constant at whichthe velocity of the particle approaches the instanta-neous velocity, and is the ratio between the mass of thecollagen molecule (300 kDa) and the drag coefficient(c). Molecules of small mass have very low time con-stants (~m/c) in the range of femtoseconds in this
particular case. Therefore, the molecule reaches theterminal velocity instantaneously when the electro-static force is applied and the instantaneous velocity atany given location becomes:
vi ¼FE
cð2bÞ
It then becomes possible to calculate the time it takesfor the molecule to traverse a certain distance by inte-grating the velocity over distance. Overall, thisapproach utilizes a physically correct velocity profilethat diminishes to zero and that is calculable fromelectrostatic force. It also enables the prediction of thetime it takes for the molecule to cover a certain distance.
The drag coefficient (c, nN s/m) for a linear mole-cule in a viscous medium is calculated as:
c ¼ 2pg0l
ln l2 �R
� �
� 0:20ð3Þ
Here, g0 is the dynamic viscosity (Pa s) of the col-lagen solution, l the length of a collagen molecule(300 nm), and �R the radius of a collagen molecule(1.5 nm). The form of equation of the drag coefficient(Eq. 3) describes the motion of the particle parallel tothe plane surface of the electrochemical cell. This formof drag coefficient accounts for the proximity of mol-ecules to a plane surface.17
Besides the electrostatic force, the drag (Fd) forceacts on the molecule. The drag force (Fd) depends onthe geometry of the molecule, the drag coefficient (c),and the instantaneous velocity (vi) of the collagenmolecule in the medium.17
Fd ¼ � c � við Þ ð4Þ
Finally, the collagen molecule experiments a totalnet force, that is equivalent to the summation of theelectrostatic and drag forces (FE + Fd).
Determination of pH Gradient
A pH indicator dye (Universal indicator pH 3–11,Riedel-de-Haen, Seelze, Germany) was added to thedialyzed collagen solution between the electrodes andimages were obtained at the steady state. The pH wasdetermined from colorimetric image analysis of the dyeas a function of the inter-electrode distance. These pHvalues were used to calculate the charge induction intype-I collagen molecules.
Assessment of Viscosity as a Function of pH
The viscosity of collagenous medium is dependenton pH11,13; therefore, the drag force is a function ofdistance between the electrodes. The pH of dialyzed
J. A. UQUILLAS AND O. AKKUS1644

collagen solutions (Nutragen�, 6 mg/mL, AdvancedBiomatrix, San Diego, CA) were adjusted to variousvalues in the range of 3 and 11 using 0.1 N HCl or0.1 N NaOH solutions. Calculations of g0 were doneusing a method adopted from Bailey et al.3 with slightmodifications. Apparent viscosities of dialyzed colla-gen solutions at different pH values were measuredusing a rheometer (AR2000, TA Instruments, NewCastle, DE) with cone-plate geometry (40 mm, 2� coneangle). Viscosities were measured at shear rates of 1 to100 Hz at 25 �C. Apparent viscosity can be extrapo-lated to zero shear rate and reported as g0½ �D�0 whereD ~ 0 Hz represents the zero shear rate.2 Theapproximation to zero shear rates was done using theAR2000 rheometer at D � 1 s21. Dynamic viscositywas analytically derived using the Kelvin–Voigt vis-coelastic model.8
Statistical Analysis
The results of the dynamic viscosity study (n = 3measurements per pH value) were analyzed with oneway ANOVA for repeated measures with Tukeypost hoc comparisons. Statistical significance thresholdwas set at 0.05 for all tests (p< 0.05, Minitab 16, StateCollege, PA).
RESULTS
Dependence of the Charge Distribution of a Type-ICollagen Molecule With pH
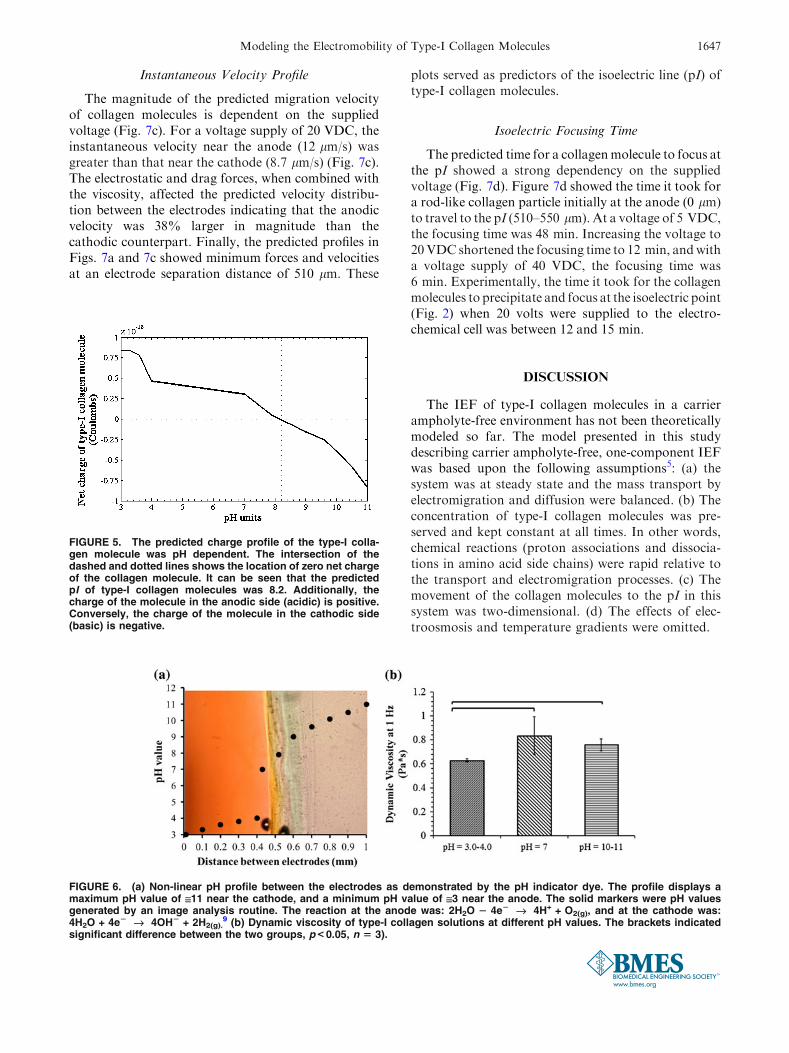
The predicted charge distribution profile along thetype-I collagen molecule showed a strong dependencyon pH (Fig. 4). When the molecule was close to theanode (acidic pH), the net charge resulting by thesummation of all zwitterions was mainly positive(~0.8 9 10216 C; Fig. 4a). Amino acids with positiveand negative electrical charge at different locations arecalled zwitterions. In the charge profile along a rod-likecollagen molecule, ionisable amino acids exhibiteddifferent electrical charges depending on the pH of themedium. When the molecule was at a region of slightlybasic pH, the predicted net charge along the moleculewas equally positive and negative (Fig. 4b). This meantthat the average net charge along the moleculeapproached zero. When the molecule was close to thecathode (very basic pH), the predicted absolute netcharge was mainly negative (~20.8 9 10216 C,Fig. 4c). The predicted net charge along the type-Icollagen molecules as predicted by the model wasplotted against the pH (Fig. 5). The predicted pI valuewhere zero net charge was attained was close topH 8.2.
Isoelectric Focusing in a Nonlinear, ExperimentallyMeasured pH Gradient
The s-shaped pH profile was found to be non-linear(Fig. 6a). Thus, the effect of pH on the electrostatic,drag, and total forces, as well as on the velocity of atype-I collagen molecule was calculated using thisexperimentally observed non-linear pH gradient.
Isoelectric Focusing in a pH-DependentViscous Medium
The dynamic viscosity of a type-I collagen solutionmeasured with a rheometer showed a close dependencyon the pH (Fig. 6b). We assume that the measureddynamic viscosity values at different pH are represen-tative of the trends in relative viscosities of the collagensolution at different pH values between the electrodes.Near the anode the environment was highly acidic,which would result in a less viscous collagen solution(0.63 Pa s). At a neutral pH the viscosity would besignificantly higher (0.84 Pa s) than that at the anode.The dynamic viscosity at the basic reservoir near thecathode would be significantly higher (0.76 Pa s) thanthat at the anode, but not significantly different thanregions where the pH was neutral.
Electrostatic Force and Drag Coefficient Profiles
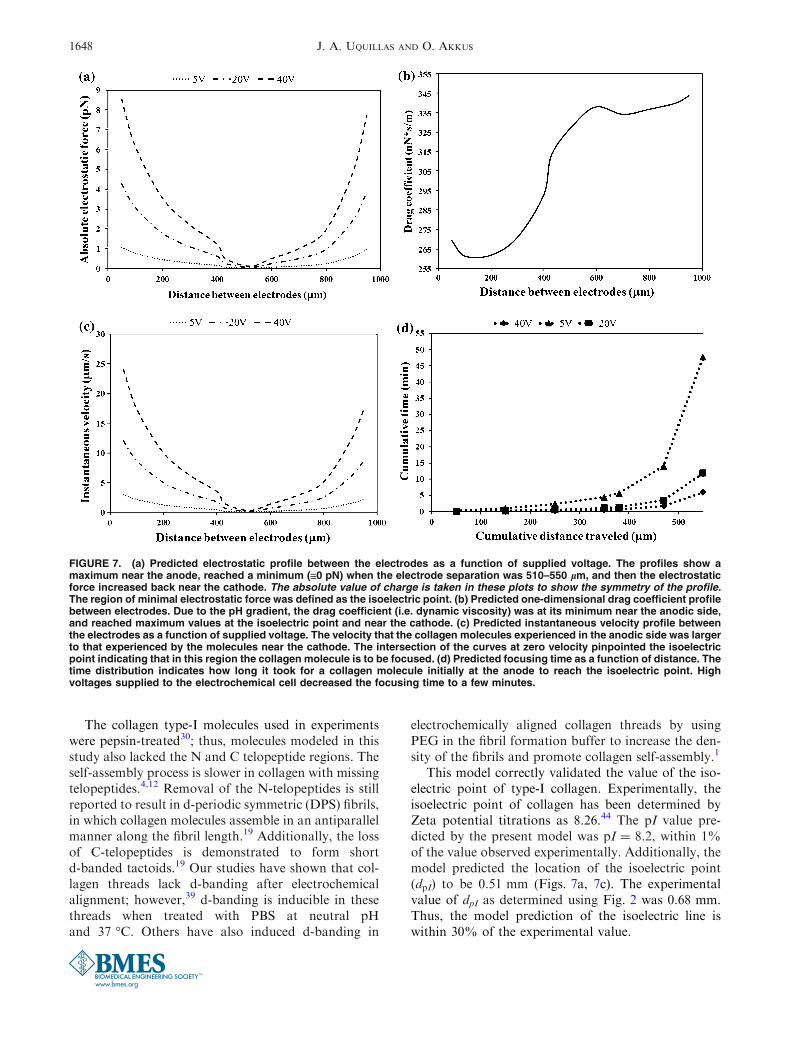
The predicted U-shaped absolute electrostatic forceprofiles showed high electrostatic forces near the anodeand the cathode (Fig. 7a). The three curves showelectrostatic force profiles for three voltages at 5, 20,and 40 VDC. As the supplied voltage increases, theelectrostatic force increases. However, the minimum ofall three profiles is at the isoelectric point (510 lm).For a supplied voltage of 20 VDC, the electrostaticforce near the cathode (3.9 pN) was similar to thatnear the anode (4.23 pN) since the collagen moleculeexhibited the same charge magnitude (0.8 9 10216 C)near both electrodes (Fig. 5) but with opposite charges.The predicted drag coefficient profile indicated amonotonic increase of the viscosity in the regionbetween the electrodes (Fig. 7b). This monotonicincrease was at its maximum near the cathode, indicatingcollagen gelation at basic pH. The anodic (acidic)region was less viscous than the cathodic (basic)counterpart. The drag force profile (not shown) thatcounteracted the electrostatic force and prevented themigration of type-I collagen molecules to the pI wassimilar in magnitude to the electrostatic force. Thisindicated that the net force acting on the collagenmolecule was very close to zero. Thus, the moleculehad no acceleration during molecular migration but afinite velocity equal to the instantaneous velocity (vi).
Modeling the Electromobility of Type-I Collagen Molecules 1645
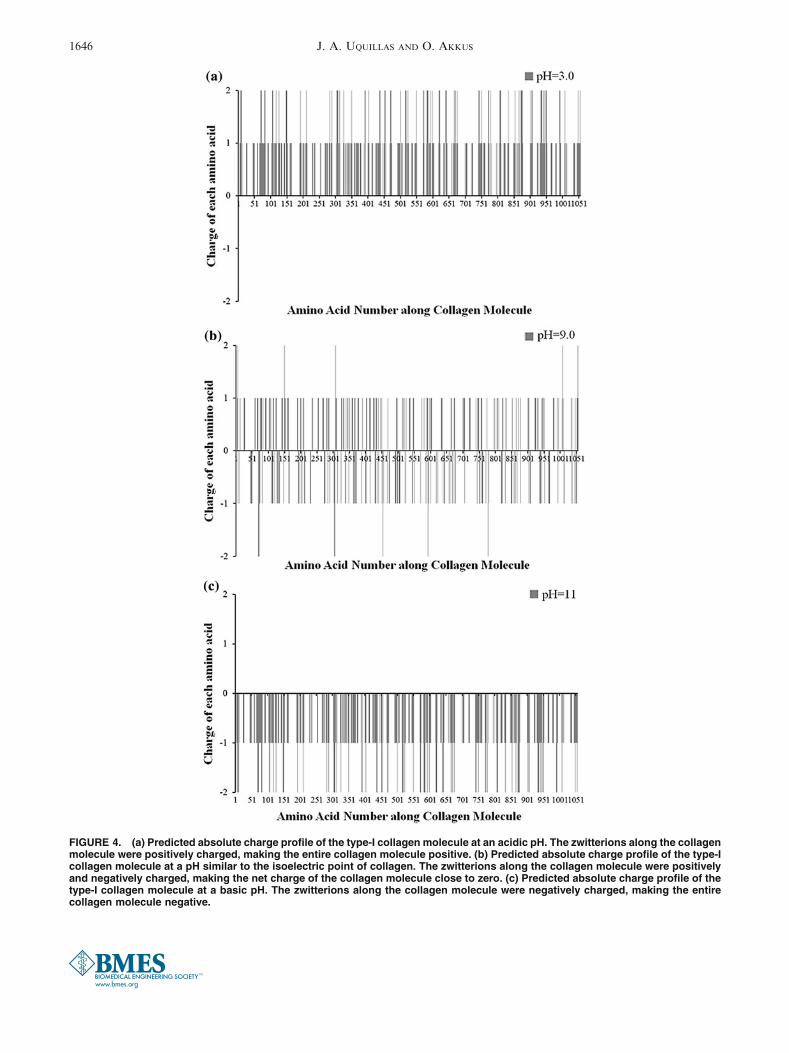
FIGURE 4. (a) Predicted absolute charge profile of the type-I collagen molecule at an acidic pH. The zwitterions along the collagenmolecule were positively charged, making the entire collagen molecule positive. (b) Predicted absolute charge profile of the type-Icollagen molecule at a pH similar to the isoelectric point of collagen. The zwitterions along the collagen molecule were positivelyand negatively charged, making the net charge of the collagen molecule close to zero. (c) Predicted absolute charge profile of thetype-I collagen molecule at a basic pH. The zwitterions along the collagen molecule were negatively charged, making the entirecollagen molecule negative.
J. A. UQUILLAS AND O. AKKUS1646
Instantaneous Velocity Profile
The magnitude of the predicted migration velocityof collagen molecules is dependent on the suppliedvoltage (Fig. 7c). For a voltage supply of 20 VDC, theinstantaneous velocity near the anode (12 lm/s) wasgreater than that near the cathode (8.7 lm/s) (Fig. 7c).The electrostatic and drag forces, when combined withthe viscosity, affected the predicted velocity distribu-tion between the electrodes indicating that the anodicvelocity was 38% larger in magnitude than thecathodic counterpart. Finally, the predicted profiles inFigs. 7a and 7c showed minimum forces and velocitiesat an electrode separation distance of 510 lm. These
plots served as predictors of the isoelectric line (pI) oftype-I collagen molecules.
Isoelectric Focusing Time
The predicted time for a collagenmolecule to focus atthe pI showed a strong dependency on the suppliedvoltage (Fig. 7d). Figure 7d showed the time it took fora rod-like collagen particle initially at the anode (0 lm)to travel to the pI (510–550 lm). At a voltage of 5 VDC,the focusing time was 48 min. Increasing the voltage to20VDC shortened the focusing time to 12 min, andwitha voltage supply of 40 VDC, the focusing time was6 min. Experimentally, the time it took for the collagenmolecules to precipitate and focus at the isoelectric point(Fig. 2) when 20 volts were supplied to the electro-chemical cell was between 12 and 15 min.
DISCUSSION
The IEF of type-I collagen molecules in a carrierampholyte-free environment has not been theoreticallymodeled so far. The model presented in this studydescribing carrier ampholyte-free, one-component IEFwas based upon the following assumptions5: (a) thesystem was at steady state and the mass transport byelectromigration and diffusion were balanced. (b) Theconcentration of type-I collagen molecules was pre-served and kept constant at all times. In other words,chemical reactions (proton associations and dissocia-tions in amino acid side chains) were rapid relative tothe transport and electromigration processes. (c) Themovement of the collagen molecules to the pI in thissystem was two-dimensional. (d) The effects of elec-troosmosis and temperature gradients were omitted.
FIGURE 5. The predicted charge profile of the type-I colla-gen molecule was pH dependent. The intersection of thedashed and dotted lines shows the location of zero net chargeof the collagen molecule. It can be seen that the predictedpI of type-I collagen molecules was 8.2. Additionally, thecharge of the molecule in the anodic side (acidic) is positive.Conversely, the charge of the molecule in the cathodic side(basic) is negative.
FIGURE 6. (a) Non-linear pH profile between the electrodes as demonstrated by the pH indicator dye. The profile displays amaximum pH value of @11 near the cathode, and a minimum pH value of @3 near the anode. The solid markers were pH valuesgenerated by an image analysis routine. The reaction at the anode was: 2H2O 2 4e2 fi 4H+ + O2(g), and at the cathode was:4H2O + 4e2 fi 4OH2 + 2H2(g).
9 (b) Dynamic viscosity of type-I collagen solutions at different pH values. The brackets indicatedsignificant difference between the two groups, p < 0.05, n 5 3).
Modeling the Electromobility of Type-I Collagen Molecules 1647
The collagen type-I molecules used in experimentswere pepsin-treated30; thus, molecules modeled in thisstudy also lacked the N and C telopeptide regions. Theself-assembly process is slower in collagen with missingtelopeptides.4,12 Removal of the N-telopeptides is stillreported to result in d-periodic symmetric (DPS) fibrils,in which collagen molecules assemble in an antiparallelmanner along the fibril length.19 Additionally, the lossof C-telopeptides is demonstrated to form shortd-banded tactoids.19 Our studies have shown that col-lagen threads lack d-banding after electrochemicalalignment; however,39 d-banding is inducible in thesethreads when treated with PBS at neutral pHand 37 �C. Others have also induced d-banding in
electrochemically aligned collagen threads by usingPEG in the fibril formation buffer to increase the den-sity of the fibrils and promote collagen self-assembly.1
This model correctly validated the value of the iso-electric point of type-I collagen. Experimentally, theisoelectric point of collagen has been determined byZeta potential titrations as 8.26.44 The pI value pre-dicted by the present model was pI = 8.2, within 1%of the value observed experimentally. Additionally, themodel predicted the location of the isoelectric point(dpI) to be 0.51 mm (Figs. 7a, 7c). The experimentalvalue of dpI as determined using Fig. 2 was 0.68 mm.Thus, the model prediction of the isoelectric line iswithin 30% of the experimental value.
FIGURE 7. (a) Predicted electrostatic profile between the electrodes as a function of supplied voltage. The profiles show amaximum near the anode, reached a minimum (@0 pN) when the electrode separation was 510–550 lm, and then the electrostaticforce increased back near the cathode. The absolute value of charge is taken in these plots to show the symmetry of the profile.The region of minimal electrostatic force was defined as the isoelectric point. (b) Predicted one-dimensional drag coefficient profilebetween electrodes. Due to the pH gradient, the drag coefficient (i.e. dynamic viscosity) was at its minimum near the anodic side,and reached maximum values at the isoelectric point and near the cathode. (c) Predicted instantaneous velocity profile betweenthe electrodes as a function of supplied voltage. The velocity that the collagen molecules experienced in the anodic side was largerto that experienced by the molecules near the cathode. The intersection of the curves at zero velocity pinpointed the isoelectricpoint indicating that in this region the collagen molecule is to be focused. (d) Predicted focusing time as a function of distance. Thetime distribution indicates how long it took for a collagen molecule initially at the anode to reach the isoelectric point. Highvoltages supplied to the electrochemical cell decreased the focusing time to a few minutes.
J. A. UQUILLAS AND O. AKKUS1648

The experimentally observed non-linear pH profilewas used in the model as a realistic depiction of IEF oftype-I collagen molecules. The electrostatic force atequidistant points from the anode and the cathode washigher at the anode due to lower viscosity of collage atacidic pH. Specifically, the electrostatic force (Fig. 7a)at 20 VDC indicated that at x = 100 lm from theanode the force was 3.01 (±0.166) pN. The corre-sponding force at x = 900 lm from the anode was 2.45(±0.093) pN. Additionally, the total force atx = 200 lm from the anode was 1.77 (±0.102) pN,and the corresponding force at x = 800 lm from theanode was 0.97 (±0.042) pN. Greater repulsion forcesat the anode than in the cathode agrees with theexperimental observation which indicated the migra-tion to be faster on the anode side. Besides differentialviscosity which is lower at the anode, it appears thatthe charge of the molecules near the anode gives forthto greater repulsion. These predictions confirm that thedeceleration of molecular mobility near the cathodewas, in part, due to viscous effects.
Several parameters affected the sensitivity of themodel more than others. The separation betweenelectrodes (R) affected the strength of the electric field,influencing the predicted electrostatic force and veloc-ity profiles. Previous experiments have demonstratedthat the alignment of collagen diminishes as the elec-trode separation increases.9 Additionally, parameterslike dynamic viscosity and voltage (Figs. 7a, 7c) sup-plied to the electrochemical cell greatly influenced themolecular velocity at the anode and cathode. Forexample, assuming a collagen medium with twice theviscosity distribution as shown in Fig. 6b, the time ittook to reach from x = 0 mm to x = 0.05 mm was6.61 s on the anode side. On the more viscous cathodicside, the time it took to travel from x = 1 mm tox = 0.95 mm was 9.39 s for 20 VDC. If the dynamicviscosity were reduced two fold, then the correspond-ing times were predicted as 3.13 s from x = 0 mm tox = 0.05 mm and 4.17 s from x = 1 mm tox = 0.95 mm for 20 VDC.
The resistance to flow depends on the relative shearor slippage of contiguous molecules within the mate-rial. In protein solutions, the resistance of molecules toslip past each other is determined by intramolecularinteractions such as physical molecular entanglement,electrostatic forces (i.e. hydrogen bonding), hydro-philic, and hydrophobic interactions.6 Our resultssuggested that the intramolecular interactions betweentype-I collagen molecules in the anode region were lesssignificant than those in the cathode. A highly pro-tonated collagen molecule in the anodic region mayexperience less intramolecular interactions with sur-rounding molecules (low formation of aggregates),thus decreasing the viscosity of the medium (as seen in
Fig. 6b). In a low viscosity medium, the effect of theelectrostatic force would increase the mobility, migra-tion, and velocity of single collagen molecules that donot form aggregates. This may be the reason for highermolecular velocities in the anode region. Highly ener-getic collagen molecules migrated faster from theanode region, and self-aggregate along the pI, where thecharge of the collagen molecules was at its minimum.Additionally, the self-aggregation phenomena alongthe pI of neutrally charged collagen molecules mayexplain why type-I collagen threads formed using thismodified isoelectric method lacked the typicald-banding pattern of natural collagenous structures.39
Based on the neutral charge of collagen moleculesalong the pI, it can be inferred that the molecules willaggregate in a direction parallel the electrodes with nopreferential staggered order that would contribute tothe d-banding pattern. The lack of d-banding in freshlyaligned fibers teased from ELAC threads has beenexperimentally observed by Cheng et al.9 However,treatment of aligned collagen in phosphate bufferedsaline at neutral pH and incubation at 37 �C result inthe induction of d-bands as we have shown earlierusing SAXS.9,39
In vitro, the self-assembly process of type-I collagenmolecules can be initiated from cold acid conditions(4 �C, pH 3) to warm neutral conditions (34–37 �C, pH7.4).41,42 This self-assembly process reveals a lag phase,with no increase in turbidity, followed by a sigmoidalgrowth with concomitant turbidity increase, and a finalplateau when precipitation of collagen occurs.16,29,43
The pH of the self-assembly process is an importantparameter to obtain d-banded collagenous structures;that is, slightly acidic to slightly neutral pH values(7.1–8) are essential to form mature d-banded collagentype-I fibers.15,18,25,42 The non-linear pH profile pre-sented here brackets the optimal pH value for fibrilformation; however, the temperature and presence ofsalts are also essential for the formation of d-bandedstructures. Since the electrochemical alignment processis inhibited by the presence of salts, we induced-banding after the alignment by incubating alignedthreads to in PBS at 37 �C.39
The non-linear pH and viscosity gradients describedin this study elucidated the mechanisms of type-I col-lagen self-aggregation along the isoelectric line. Thealignment and self-aggregation of type-I collagen isoriented parallel to the electrodes in the electrochemi-cal cell. The focusing mechanism can be as follows:first, there was a steeper pH gradient (5.2 pH units)over a 0.50–0.55 mm electrode separation from theanode to the pI. The pH gradient in this region con-centrated positively charged collagen molecules at theanode side with greater repelling forces and fastermigration velocities. Second, the less steep pH gradient
Modeling the Electromobility of Type-I Collagen Molecules 1649

between the pI and the cathodic side (2.8 pH units)over a 0.5–0.45 mm electrode separation resulted in aconcentration of negatively charged collagen mole-cules. The negatively charged molecules experimentedlower repelling forces at the cathodic region, resultingin molecules moving with slower migration velocities.Finally, neutral and basic pH values between the pIand the cathode favored the gelation of type-I colla-gen,10,32,43 which in turn increased the dynamic vis-cosity of the type-I collagen solution.
The analysis of the time constant (s) of a collagenmolecule to reach terminal velocity upon applicationof electrostatic force indicated that the collagen mol-ecule reached its local instantaneous velocity extremelyquickly (1–2 fs). In comparison, the duration formolecules to migrate to the pI was in the order ofminutes (Fig. 7d). Therefore, the model assumed thatat any given point, molecules reached the terminalvelocity instantaneously. As the molecule traveledfurther away from the electrodes, the electrostaticrepulsion reduced and the molecule decelerated to zerovelocity at the pI where net charge; thus, net repulsion,was zero.
The migration and self-aggregation was explained byIEF as an electrophysical phenomenon. Aggregates ofneutrally-charged type-I collagen molecules are com-pressed from both sides to the pI, causing the rod-likemolecules to align perpendicular to the electric field.Thus, by using computational modeling, we have con-firmed for the first time that the self-aggregationmechanism along the pI depended on the pH and vis-cosity gradients. Also, we quantified the electrostaticand drag forces acting on the collagen molecule ofampholytic nature. The experimental validation of themigration velocity predicted by the model can improvein several aspects. The experimental measurement ofthe duration it took for molecules to travel to the pIwasconducted by visualizing the first emergence of collagenmolecules in the pI region using compensated polarizedimaging. A more rigorous and direct quantification ofthe velocity of a single collagen molecule may be utili-zation of fluorescently labeled molecules. Also, themodel utilizes pKa values of isolated amino acids anddoes not account for the dependence of pKa on theadjacent amino acids.40 However, it appears that theutilized pKa values do not diverge substantially fromthe actually pKa values because the collagen pI has beencorrectly estimated and validated by the model (Fig. 5)using the pKa values provided in Table 1. Finally, theanalysis has been done for a single, rod-like collagenmolecule without accounting for the intermolecularforces. Further work will focus on the interaction ofgroups of type-I collagen molecules during migrationand self-aggregation along the pI during IEF in anampholyte-free viscous medium.
CONCLUSIONS
The present study investigated the charge distributionalong a type-I collagen molecule during IEF; demon-strated the role of the pH gradient in collagen migrationto the pI; and predicted the variation in the electrostaticforce, drag coefficient, and velocity profiles in the inter-electrode space. The collagen molecule was computa-tionally modeled as a rod initially randomly distributedbetween the inter-electrode space that accumulated andprecipitatedat thepI. Thekeyfindingsof this studywere1:the predicted chargeof the collagenmoleculewas positivenear the anode and negative near the cathode2; the pre-dicted isoelectric line of collagen was computationallyestimated as pH 8.23; experimentally, a nonlinear pHprofile was observed4; the experimentally measured vis-cosity of the collagen solution near the anode was sig-nificantly less than the viscosity near the cathode5; thefocusing time predicted by the model was in generalagreement with the experimental observations.
In conclusion, the major forces andmechanisms for theIEF of type-I collagen molecules in a carrier ampholyte-free environment have been elucidated. This model hasexplained the origin of the charge distribution of a type-Icollagenmoleculewhen itmigrates to the pI in a non-linearpH gradient. Also, the model has confirmed that theobserved pH gradient was sufficient to induce electrostaticforces, resulting in electrophoretic migration. Therefore,the theoretical and experimental material presented hereexplains clearly the fundamentals of IEF under uniformelectric fields, and represent the baseline framework toexpand this IEF process in the production of multicom-ponent collagen-basedbiomaterials to engineer soft tissues.
APPENDIX
Derivation of the Electrostatic Force Expression
The electric field was derived to estimate the elec-trostatic force acting on a charged particle that was ata distance x from the anode. The electric field gener-ated by a charged line source was estimated bysuperimposing the point charge fields of infinitesimalcharge elements dQ (Fig. 1).
The electric field by dQ was defined as:
dE ¼ kdQ
r2ðA1Þ
The horizontal component of the field at the loca-tion of the charged particle q was given by:
dEðxÞ ¼ kkdyr2
cos b ¼ kkr2
x
rdy ¼ kkx
y2 þ x2ð Þ3=2dy ðA2Þ
where k was the charge per unit length (Q/L, C/m), Lthe length of the electrodes (m), and k was the
J. A. UQUILLAS AND O. AKKUS1650

Coulomb’s constant (N m2/C2). The integral to obtainthe field expression was:
E xð Þ ¼ kkxZ
b
�a
dy
y2 þ x2ð Þ3=2¼ kk
x
yffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
y2 þ x2ð Þp
" #b
�a
ðA3Þ
The electric field effect due to the anode was:
EANx ¼ kk
x
bffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
x2 þ b2ð Þp þ a
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
x2 þ a2ð Þp
" #
ðA4Þ
And the electric field expression due to the cathodewas:
ECAx ¼
kkR� xð Þ
bffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
R� xð Þ2þb2� �
r þ affiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
R� xð Þ2þa2� �
r
2
6
6
4
3
7
7
5
ðA5Þ
Charge per unit length (k) and the Coulomb’s con-stant (k) were:
k ¼ 1
4peðA6aÞ
k ¼ Q
LðA6bÞ
where Q was the charge in the electrodes. e was thepermittivity of collagen27,36 which is equal to:
e ¼ 5e0; e0 ¼ 4:4270939085� 10�11 A s/Vmð Þ ðA7Þ
A particle of charge q positioned in this electric fieldexperienced an electrostatic force equal to:
F!¼ Ex
�!q ðA8Þ
The net electrostatic force FE�!
acting on the chargedparticle between two parallel wires of opposite chargeswas:
FE�! ¼ FAN
��!þ FCA��! ¼ q EAN
x
��!þ ECA
x
��!� �
ðA9Þ
where FAN��!
and FCA��!
indicated the forces applied by theanode and cathode. The corresponding net force theygenerate became:
FE�!¼kkq
8
>
>
<
>
>
:
b
xffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
x2þb2ð Þp þ a
xffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
x2þa2ð Þp
" #
þ b
R�xð Þffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
R�xð Þ2þb2� �
r þ a
R�xð Þffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
R�xð Þ2þa2� �
r
2
6
6
4
3
7
7
5
9
>
>
=
>
>
;
ðA10Þ
And substituting Eqs. (A6a) and (A6b) in Eq. (A10)we obtained:
FE�!¼ Qq
4peL
8
>
>
<
>
>
:
b
xffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
x2þb2ð Þp þ a
xffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
x2þa2ð Þp
" #
þ b
R�xð Þffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
R�xð Þ2þb2� �
r þ a
R�xð Þffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
R�xð Þ2þa2� �
r
2
6
6
4
3
7
7
5
9
>
>
=
>
>
;
ðA11Þ
The field expression in the vertical direction E yð Þ hasbeen calculated to be close to zero. This was the casedue to the symmetry of the system.
Derivation of the Charge Q at the Wires
The charge Q has been calculated at the anode. Itwas assumed that the cathode carried the same chargebut with different sign. In this case, the variables a, andb in Eq. (A4) have been assumed to be +L/2. Thus, theelectric field equation at the anode was:
EANx ¼ k � k � L
x
1ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
x2 þ L=2ð Þ2h i
r
2
6
6
4
3
7
7
5
ðA12Þ
Multiplying both sides of the equation with dx gave:
EANx dx ¼ k � k � L
x
1ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
x2 þ L=2ð Þ2h i
r
2
6
6
4
3
7
7
5
dx ¼ �dV
Here dV was the differential voltage along the sep-aration R between the wires. Integrating both sidesyielded:
�Z
VCA
VAN
dV ¼ kkLZ
XCA
XAN
1
x �ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
x2 þ L=2ð Þ2h i
r
2
6
6
4
3
7
7
5
dx
� VCA�VANð Þ¼� kkLð Þ 2
Lln
L=2þffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
X2CAþ L=2ð Þ2
� �
r
XCA
0
B
B
@
1
C
C
A
2
6
6
4
� 2
Lln
L=2þffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
X2ANþ L=2ð Þ2
� �
r
XAN
0
B
B
@
1
C
C
A
3
7
7
5
DV ¼ 2kk ln
L=2þffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
X2CAþ L=2ð Þ2ð Þ
p
XCA
L=2þffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
X2ANþ L=2ð Þ2ð Þ
p
XAN
0
B
@
1
C
A
2
6
4
3
7
5
Modeling the Electromobility of Type-I Collagen Molecules 1651

DV ¼ 2kk ln
XAN L=2þffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
X2CA þ L=2ð Þ2
� �
r�
XCA L=2þffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
X2AN þ L=2ð Þ2
� �
r�
0
B
B
B
@
1
C
C
C
A
2
6
6
6
4
3
7
7
7
5
We defined the term between brackets in the aboveequation as the shape factor (a):
a ¼ ln
XAN L=2þffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
X2CA þ L=2ð Þ2
� �
r�
XCA L=2þffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
X2AN þ L=2ð Þ2
� �
r�
0
B
B
B
@
1
C
C
C
A
ðA13Þ
The voltage drop across the wires then became:
DV ¼ 2kka ðA14Þ
Substituting Eqs. (A6a) and (A6b) in Eq. (A14), weobtained the expression of the charge in the wires as:
Q ¼ 2pDVLea
ðA15Þ
ACKNOWLEDGMENTS
This study was funded by the National ScienceFoundation (CBET-0754442). Any opinions, findings,conclusions, and/or recommendations expressed in thisstudy are those of the authors and do not necessarilyreflect the views of the National Science Foundation.
CONFLICT OF INTEREST
The authors have no conflict of interests to declare.
REFERENCES
1Abu-Rub, M. T., K. L. Billiar, M. H. van Es, A. Knight,B. J. Rodriguez, D. I. Zeugolis, S. McMahon, A. J.Windebank, and A. Pandit. Nano-textured self-assembledaligned collagen hydrogels promote directional neuriteguidance and overcome inhibition by myelin associatedglycoprotein. Soft Matter 7(6):2770–2781, 2011.2Ananthanarayanan, S., and A. Veis. The molecularparameters of monomeric and acid-soluble collagens. Lowshear gradient viscosity and electric birefringence. Bio-polymers 11(7):1365–1377, 1972.3Bailey, J. L., P. J. Critser, C. Whittington, J. L. Kuske,M. C. Yoder, and S. L. Voytik-Harbin. Collagen oligomersmodulate physical and biological properties of three-dimensional self-assembled matrices. Biopolymers95(2):77–93, 2011.4Berg, R. A., D. E. Birk, and F. H. Silver. Physical char-acterization of type-I procollagen in solution—evidencethat the propeptides limit self-assembly. Int. J. Biol. Mac-romol. 8(3):177–182, 1986.
5Bier, M., R. A. Mosher, and O. A. Palusinski. Computer-simulation and experimental validation of isoelectric-focusing in ampholine-free systems. J. Chromatogr. 211(3):313–335, 1981.6Bos, M. A., and T. van Vliet. Interfacial rheologicalproperties of adsorbed protein layers and surfactants: areview. Adv. Colloid Interface Sci. 91(3):437–471, 2001.7Cabrera, C. R., B. Finlayson, and P. Yager. Formation ofnatural pH gradients in a microfluidic device under flowconditions: model and experimental validation. Anal.Chem. 73(3):658–666, 2001.8Chan, R. W., and I. R. Titze. Viscosities of implantablebiomaterials in vocal fold augmentation surgery. Laryn-goscope 108(5):725–731, 1998.9Cheng, X., U. A. Gurkan, C. J. Dehen, M. P. Tate, H. W.Hillhouse, G. J. Simpson, and O. Akkus. An electro-chemical fabrication process for the assembly of aniso-tropically oriented collagen bundles. Biomaterials29(22):3278–3288, 2008.
10Christiansen, D. L., E. K. Huang, and F. H. Silver.Assembly of type I collagen: fusion of fibril subunits andthe influence of fibril diameter on mechanical properties.Matrix Biol. 19(5):409–420, 2000.
11de Paula, M., G. Goissis, and V. C. Martins. Rheologicalbehavior of anionic collagen injectable gels in the presenceof rhamsan for plastic surgery applications. J. Mater. Sci.Mater. Med. 18(9):1683–1690, 2007.
12Fratzl, P. Collagen: Structure and Mechanism (1st ed.).New York: Springer, 2008.
13Friess, W., and M. Schlapp. Effects of processing condi-tions on the rheological behavior of collagen dispersions.Eur. J. Pharm. Biopharm. 51(3):259–265, 2001.
14Gurkan, U. A., X. Cheng, V. Kishore, J. A. Uquillas, andO. Akkus. Comparison of morphology, orientation, andmigration of tendon derived fibroblasts and bone marrowstromal cells on electrochemically aligned collagen con-structs. J. Biomed. Mater. Res. A 94(4):1070–1079, 2010.
15Harris, J. R., and A. Reiber. Influence of saline and pH oncollagen type I fibrillogenesis in vitro: Fibrit polymorphismand colloidal gold labelling. Micron 38(5):513–521, 2007.
16Holmes, D. F., M. J. Capaldi, and J. A. Chapman.Reconstitution of collagen fibrils invitro—the assemblyprocess depends on the initiating procedure. Int. J. Biol.Macromol. 8(3):161–166, 1986.
17Howard, J. Mechanics of Motor Proteins and the Cyto-skeleton. Sunderland, MA: Sinauer Associates, Inc., 2001.
18Jiang, F. Z., H. Horber, J. Howard, and D. J. Muller.Assembly of collagen into microribbons: effects of pH andelectrolytes. J. Struct. Biol. 148(3):268–278, 2004.
19Kadler, K. E., D. F. Holmes, J. A. Trotter, and J. A.Chapman. Collagen fibril formation. Biochem. J. 316(Pt 1):1–11, 1996.
20Kishore, V., W. Bullock, X. Sun, W. S. Van Dyke, and O.Akkus. Tenogenic differentiation of human MSCs inducedby the topography of electrochemically aligned collagenthreads. Biomaterials 33(7):2137–2144, 2012.
21Kishore, V., J. E. Paderi, A. Akkus, K. M. Smith, D.Balachandran, S. Beaudoin, A. Panitch, and O. Akkus.Incorporation of a decorin biomimetic enhances themechanical properties of electrochemically aligned collagenthreads. Acta Biomater. 7(6):2428–2436, 2011.
22Kishore, V., J. A. Uquillas, A. Dubikovsky, M. Alshehabat,P. Snyder, G. Breur, and O. Akkus. In vivo response toelectrochemically aligned collagen bioscaffolds. J. Biomed.Mater. Res. B., 2012. doi:10.1002/jbm.b.31962.
J. A. UQUILLAS AND O. AKKUS1652

23Kumar, M. R., E. F. Merschrod, and K. M. Poduska.Correlating mechanical properties with aggregation pro-cesses in electrochemically fabricated collagen membranes.Biomacromolecules 10(7):1970–1975, 2009.
24Lam, H. T., J. Josserand, N. Lion, and H. H. Girault.Modeling the isoelectric focusing of peptides in anOFFGELmulticompartment cell. J. Proteome Res. 6(5):1666–1676,2007.
25Li, Y. P., A. Asadi, M. R. Monroe, and E. P. Douglas. pHeffects on collagen fibrillogenesis in vitro: electrostaticinteractions and phosphate binding. Mater. Sci. Eng. C29(5):1643–1649, 2009.
26Macounova, K., C. R. Cabrera, and P. Yager. Concen-tration and separation of proteins in microfluidic channelson the basis of transverse IEF. Anal. Chem. 73(7):1627–1633, 2001.
27Minary-Jolandan, M., and M. F. Yu. Nanoscale charac-terization of isolated individual type I collagen fibrils:polarization and piezoelectricity. Nanotechnology 20(8):085706, 2009.
28Mosher, R. A., D. Dewey, W. Thormann, D. A. Saville,and M. Bier. Computer-simulation and experimental vali-dation of the electrophoretic behavior of proteins. Anal.Chem. 61(4):362–366, 1989.
29Na, G. C., L. J. Butz, and R. J. Carroll. Mechanism ofinvitro collagen fibril assembly—kinetic and morphologi-cal-studies. J. Biol. Chem. 261(26):2290–2299, 1986.
30Nalinanon, S., S. Benjakul, W. Visessanguan, and H.Kishimura. Use of pepsin for collagen extraction from theskin of bigeye snapper (Priacanthus tayenus). Food Chem.104(2):593–601, 2007.
31Palusinski, O. A., T. T. Allgyer, R. A. Mosher, M. Bier,and D. A. Saville. Mathematical modeling and com-puter simulation of isoelectric focusing with electrochemi-cally defined ampholytes. Biophys. Chem. 13(3):193–202,1981.
32Roeder, B. A., K. Kokini, J. E. Sturgis, J. P. Robinson, andS. L. Voytik-Harbin. Tensile mechanical properties ofthree-dimensional type I collagen extracellular matriceswith varied microstructure. J. Biomech. Eng. 124(2):214–222, 2002.
33Shim, J., P. Dutta, and C. F. Ivory. Effects of ampholyteconcentration on protein behavior in on-chip isoelectricfocusing. Electrophoresis 29(5):1026–1035, 2008.
34Sounart, T. L., and J. C. Baygents. Simulation of electro-phoretic separations by the flux-corrected transport method.J. Chromatogr. A 890(2):321–336, 2000.
35Thormann, W., T. M. Huang, J. Pawliszyn, and R. A.Mosher. High-resolution computer simulation of thedynamics of isoelectric focusing of proteins. Electrophoresis25(2):324–337, 2004.
36Tomasell, V. P., and M. H. Shamos. Electrical properties ofhydrated collagen. 1. Dielectric properties. Biopolymers12(2):353–366, 1973.
37Uniprot.org. P02453[162-1215], Collagen alpha-1(I) chain,Bos taurus, 2010.
38Uniprot.org. P02465[80-1100], Collagen alpha-2(I) chain,Bos taurus, 2010.
39Uquillas, J. A., V. Kishore, and O. Akkus. Effects ofphosphate-buffered saline concentration and incubationtime on the mechanical and structural properties of elec-trochemically aligned collagen threads. Biomed. Mater.6(3):035008, 2011.
40van Vlijmen, H. W. T., M. Shaefer, and M. Karplus.Improving the accuracy of protein pKa calculations: con-formational averaging versus the average structure. Pro-teins 33(2):145–158, 1998.
41Ward, N. P., D. J. S. Hulmes, and J. A. Chapman. Col-lagen self-assembly invitro—electron-microscopy of initialaggregates formed during the lag phase. J. Mol. Biol.190(1):107–112, 1986.
42Williams, B. R., R. A. Gelman, D. C. Poppke, and K. A. Piez.Collagen fibril formation—optimal invitro conditions and pre-liminary kinetic results. J. Biol. Chem. 253(18):6578–6585, 1978.
43Wood, G. C., and M. K. Keech. The formation of fibrilsfrom collagen solutions. 1. The effect of experimentalconditions: kinetic and electron-microscope studies. Bio-chem. J. 75:588–598, 1960.
44Zhang, Z. K., G. Y. Li, and B. Shi. Physicochemicalproperties of collagen, gelatin and collagen hydrolysatederived from bovine limed split wastes. J. Soc. Leath. Tech.Ch. 90(1):23–28, 2006.
Modeling the Electromobility of Type-I Collagen Molecules 1653





























