Embed Size (px)
Citation preview

8/12/2019 mocropaleo crusman
http://slidepdf.com/reader/full/mocropaleo-crusman 1/19
Diversity of Polar Monothalamous Foraminifera –
Morphological and Molecular Approach
Anna Sabbatini
Department of Marine Sciences, Polytechnic University of Marche,Via Brecce Bianche, 60131, Ancona, Italy
Introduction
In many marine areas, Foraminifera are a dominant meiofaunal group in terms of both numerical
abundance and biomass. Monothalamous (single-chambered) species with either agglutinated or
organic-walled shells (‘allogromiids’ in the broad sense) are an important component of marine
foraminiferal assemblages, particularly in fine-grained sediments, for example in the deep sea and in
high latitudes regions (Gooday, 2002). Monothalamous Foraminifera also occur in freshwater
environment (Holzmann et al. 2002, 2003) and have even been discovered to live in a damp terrestrial
habitat (Meisterfeld et al. 2001). These delicate Foraminifera often account for 10-20% of individualsand species in deep-sea samples (Gooday, 2002). In Polar coastal regions, influenced by turbid glacial
melt water, and in some estuaries, allogromiids (including saccamminids) represent an even higher
proportion of live Foraminifera, in some cases >90% (Gooday, 2002).
Despite of their high abundance and ecological importance, the diversity of monothalamous
Foraminifera is very poorly known. Relatively few species have been described and most of these come
from easily accessible, intertidal environments. Deeper sea samples routinely yield ten or more
undescribed species (Gooday et al. 1998). Several new morphotypes of allogromiids have also been
reported from Antarctic coastal waters (Gooday et al. 1996). Several new species of polar and subpolar
monothalamous Foraminifera have been described (Gooday et al. 1995; DeLaca et al. 2002; Wilding2002) or redescribed (Bowser et al. 2002) during the past few years. Because the morphological
characters are not always sufficient to establish the phylogenetic relationships among allogromiids,
many recent species descriptions include the analysis of molecular data (Pawlowski et al. 2002a;
Cedhagen and Pawlowski, 2002; Gooday et al. in press; Gooday and Pawlowski, submitted; Sabbatini
et al., 2004).
Molecular phylogenetic studies challenged the traditional, morphology-based classification of
Foraminifera, prompting a profound revision of higher-level taxonomy and species identification
(Pawlowski 2000, Pawlowski and Holzmann 2002). Phylogenetic analysis of molecular data, based
mainly the small subunit ribosomal RNA and actin gene sequences, revealed that the Foraminifera
include not only testate marine species but also the naked freshwater amoeboid protists (Pawlowski et
al. 1999). Molecular studies also show that there is no clear boundary between allogromiids in the
‘traditional’ sense (i.e. organic-walled species) and single-chambered agglutinated Foraminifera,
traditionally considered as astrorhizids, suggesting that all of them form a paraphyletic group of
monothalamous Foraminifera (Pawlowski et al., 2002a, 2003). A particularly high taxonomic diversity

8/12/2019 mocropaleo crusman
http://slidepdf.com/reader/full/mocropaleo-crusman 2/19
of allogromiid Foraminifera, represented in majority by undescribed species, was revealed by molecular
study of Antarctic Foraminifera (Pawlowski et al. 2002b).
At the beginning of project the main objective was to describe new Antarctic monothalamous
Foraminifera. Many of these Foraminifera were isolated and molecularly characterized in preceding
study (Pawlowski et al. 2002c) but could not be attributed to any known species. During the year there
was possibility to collect other samples but in a different site and environment (Porcupine Abyssal
Plain, NE Atlantic Ocean). The morphological analyses carried out on board on superficial sediment
samples revealed an interesting monothalamous foraminiferal fauna composed especially by specimens
belonging to Komokiacea group. These specimens are really poor known and the most recent studies
are by Schroder et al. (1989), Gooday (1982, 1983), Gooday & Cook (1984), Tendal (1972, 1979),
Tendal & Hessler (1977), Kamenskaya (1990, 1993).
Since there are no sequences available of Komokiacea yet, and several samples of them were available
also from Antarctica to study, new aim was describe and identify a larger number of komokiacean
specimens from these two sites and where possible correlate them. Important goals are 1) molecular
data of Komokiacea group from the Atlantic Ocean in order to reveal them taxonomic diversity and
phylogenetic position within the Foraminifera 2) a comparison of species present in the Antarctic and
NE-Atlantic areas of study 3) morphological description of new species. I focused on the above points
1 and 2.
Komokiacea (Textulariina, Foraminiferida, Protista) are agglutinated Foraminifera with test consisting
of a complex system of fine, branching tubules of even diameter. The test wall is simple, with
argillaceous particles. Stercomata accumulate within the tubules. The group has worldwide distribution,
particularly in the abyssal zone. The greatest relative abundances have been found in abyssal
oligotrophic areas and in hadal trenches. Tendal & Hessler (1977) divided Superfamily Komokiaceain two families Komokiidae (Tendal & Hessler, 1977) and Bacullelidae (Tendal & Hessler, 1977). First
have test bushy or arborescent constructing widely spaced branching cylindrical tubules and second test
variously shaped, sparsely branched or dense clump. Branching is the basic feature controlling body
form. In most cases branching is dichotomous, although tri- or polychotomy also exists. In the tube wall
two layers can be distinguished: the inner layer is generally very thin and frequently laminated while
the outer part is composed by agglutinated sediment. Komokiacea are devoid of real apertures. The
pseudopodia must penetrate the tube wall through minute pores having possibly temporary character
(Tendal & Hessler, 1977). The Komokiacea vary in according to the amount and type of particles in the
agglutinated layer.
Material and Methods
Specimen collection
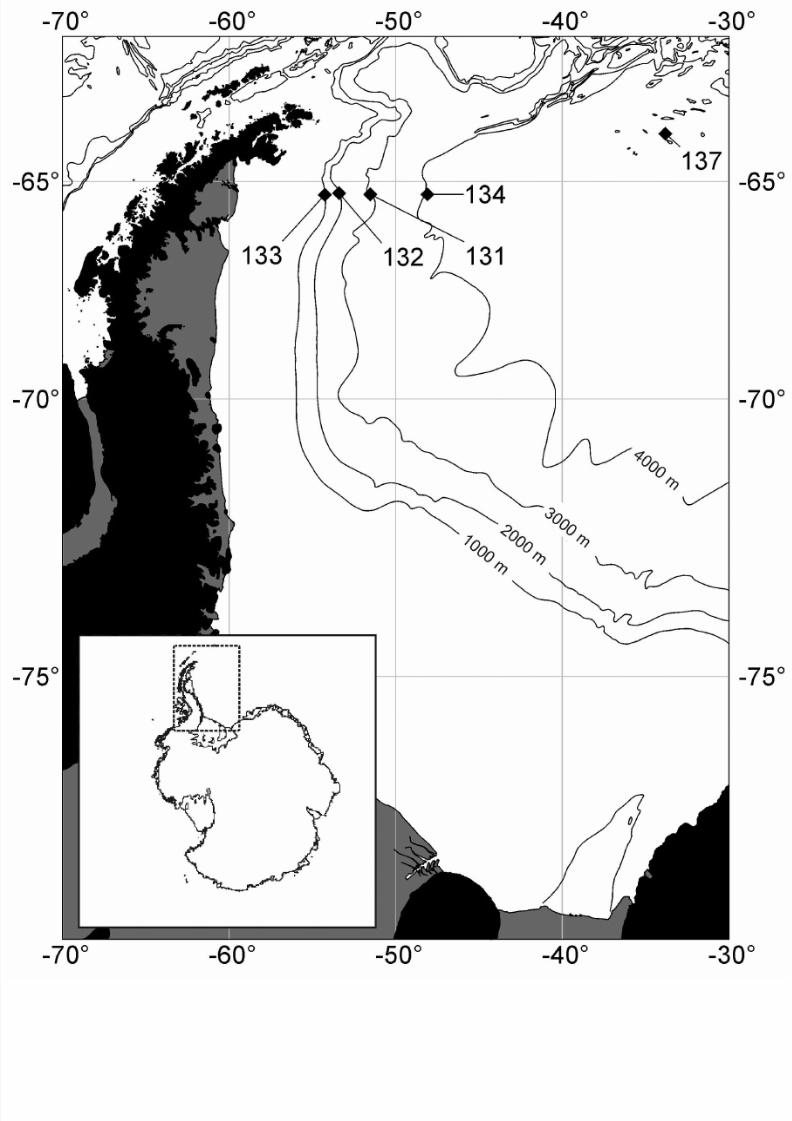
The material for this study was collected in the Weddel Sea (Antartica) during R/V Polarstern cruise
ANT-XIX/4 (ANDEEP II, February 28th to April 1st, 2002) (Figure 1).

8/12/2019 mocropaleo crusman
http://slidepdf.com/reader/full/mocropaleo-crusman 3/19
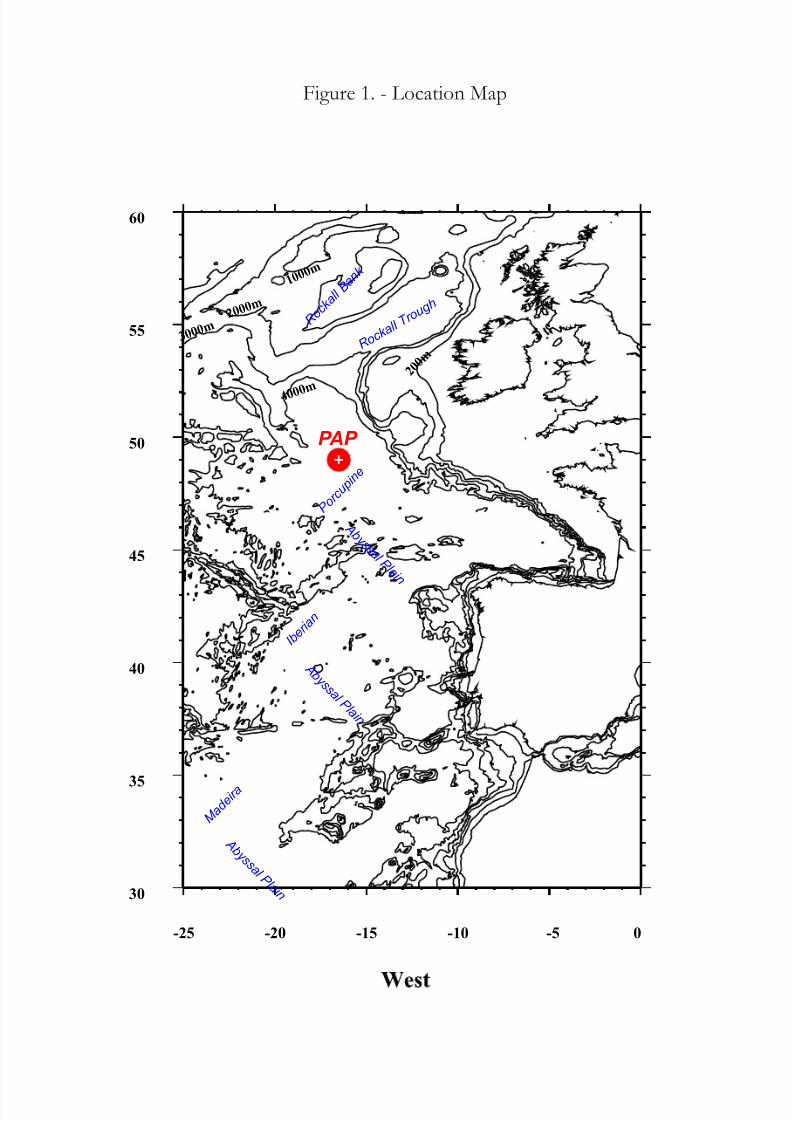
The other material was collected in the Porcupine Abyssal Plain (NE – Atlantic Ocean) in June 2004
(Figure 2). The Foraminifera (not only Komokiacea for a total of 80 samples) for molecular work have
been isolated and stored in guanidine. Sediment samples for morphological studies were sieved on 300
micron mesh and preserved in formalin 10%. All monothalamous soft-walled Foraminifera (organic
and agglutinated) will be sorted by hand, under a stereoscopic microscope, from the floated residue.
Individuals will be placed in glycerol on a cavity slide, described, photographed and classified.
DNA extraction, amplification and sequencing
DNA was extracted from single or several cells using the guanidine method as described in Tkach
& Pawlowski (1999). PCR amplifications were performed in a total volume of 50 l with an
amplification profile consisting of 40 cycles of 30s at 94C, 30s at 48C and 120s at 72C, followed by
5 min at 72C for final extension. A fragment of the SSU rRNA gene was amplified by PCR with the
primer pair s14F3 (5’ACG CA(AC) GTG TGA AAC TTG) and sB (5’ TGA TCC TTC TGC AGG TTC
ACC TAC). When the first PCR was unsuccessful, the PCR products were re-amplified using the nested
primer s14F1 (5' AAG GGC ACC ACA AGA ACG C), with an amplification profile consisting of 25
cycles and 52C for annealing time. A second series of analyses were carried out using the primer pair
s14F3 (5’ACG CA(AC) GTG TGA AAC TTG) and s17 (5’CGG TCA CGT TCG TTG C) for the PCR
and the same nested primer (5' AAG GGC ACC ACA AGA ACG C) for the re-amplification. The
amplified PCR products were purified using High Pure PCR Purification Kit (Roche Diagnostics), then
either sequenced directly or ligated into pGEM-T Vector system (Promega) and cloned in XL-2
Ultracompetent Cells (Stratagene). Sequencing reactions were prepared by using ABI-PRISM Big Dye
Terminator Cycle Sequencing Kit and analysed with an ABI 3100 DNA sequencer (Perkin-Elmer), all
according to the manufacturer’s instructions.
Sequence analysis
Sequences were aligned manually to the large database of foraminiferal sequences, using theSeaview software of Galtier et al. (1996) and Bioedit program (2004). A fragment of 462 sites was used
for the analysis with sB primer and a fragment of 100 sites was used for analysis with s17 primer.
Phylogenetic analyses were performed with the neighbor joining method (NJ) using Kimura 2 distance
algorithm.
Result Morphological work
Here I present a short taxonomic survey of some of the must abundant larger (>300 micron) rhizopod
species, representing a wide range of taxa, from populations on both the Porcupine Abyssal Plain and
Weddel Sea.The species and morphotypes from both study sites were divided into taxonomic categories in most
cases genera such as Ipoa spp., Komokia spp., Lana spp., Normanina spp. and Septuma spp. but also
indeterminate groups, e.g. Chain spp.
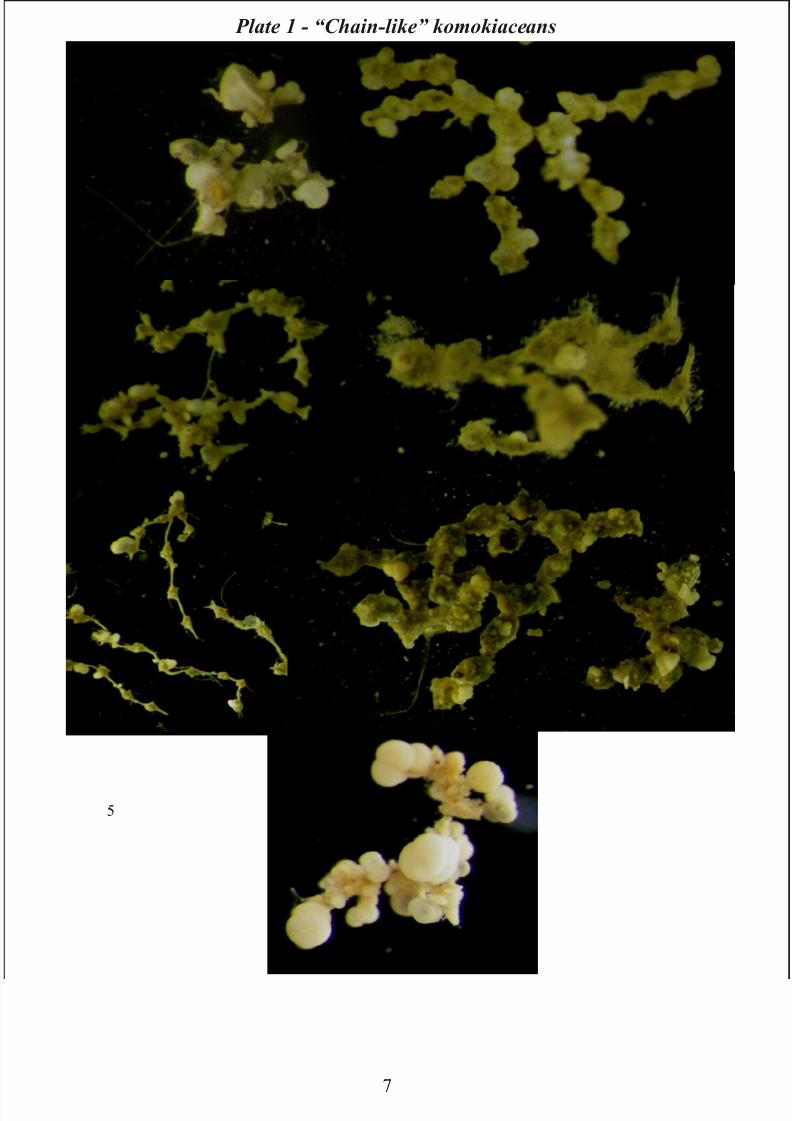
“Chain-like” komokiaceans:
These are common in the Porcupine Abyssal Plain and it is possible distinguish 7 morphotypes. The
“chains” often consist of a branched or occasionally branching chain of small spherical chambers

8/12/2019 mocropaleo crusman
http://slidepdf.com/reader/full/mocropaleo-crusman 4/19
connected by long, narrow, gently tapering necks. Occasionally globigerinacean tests of varying size
are incorporated within the chamber wall. Other types are organized as a rigid chain of irregularly-
shaped, elongate chambers connected by narrow necks. The chambers are angular; long narrow tubules
of even width may extend from the end of the chamber. The test wall consists of globigerinacean tests
of variable size. Similar morphotypes are distinguished from these forms by the irregularly-shaped
chambers as well as by chambers separated by rigid, short, narrow necks. Other “Chain-like” forms
consist of a series of spherical/ cylindrical chambers which adjoin directly; the chambers have a bumpy
exterior surface, incorporating globicerinacean tests of varying size incorporated. Occasional long,
single filaments may extend from the surface. Sometimes irregularly shaped chains of chambers are
densely encrusted with globicerinacean tests which obscure the basically organic test wall and make
individual chambers difficult to distinguish. Also in this case occasional short, fine, transparent fibres,
and long, white filaments, extend from the surface. In other species, a branching chain of triangular,
claw or arrow-head shaped chambers joined together at their pointed ends to produce a chain with an
elegant appearance. Finally, a distinctive morphotype comprises pear-shaped chambers joined to form
a branching chain which undergoes characteristic angular changes of direction. The chambers are joined
head-to-toe (Plate 1).
All chain-like morphotypes are quite delicate, they break easily and are typically found in a fragment
state.
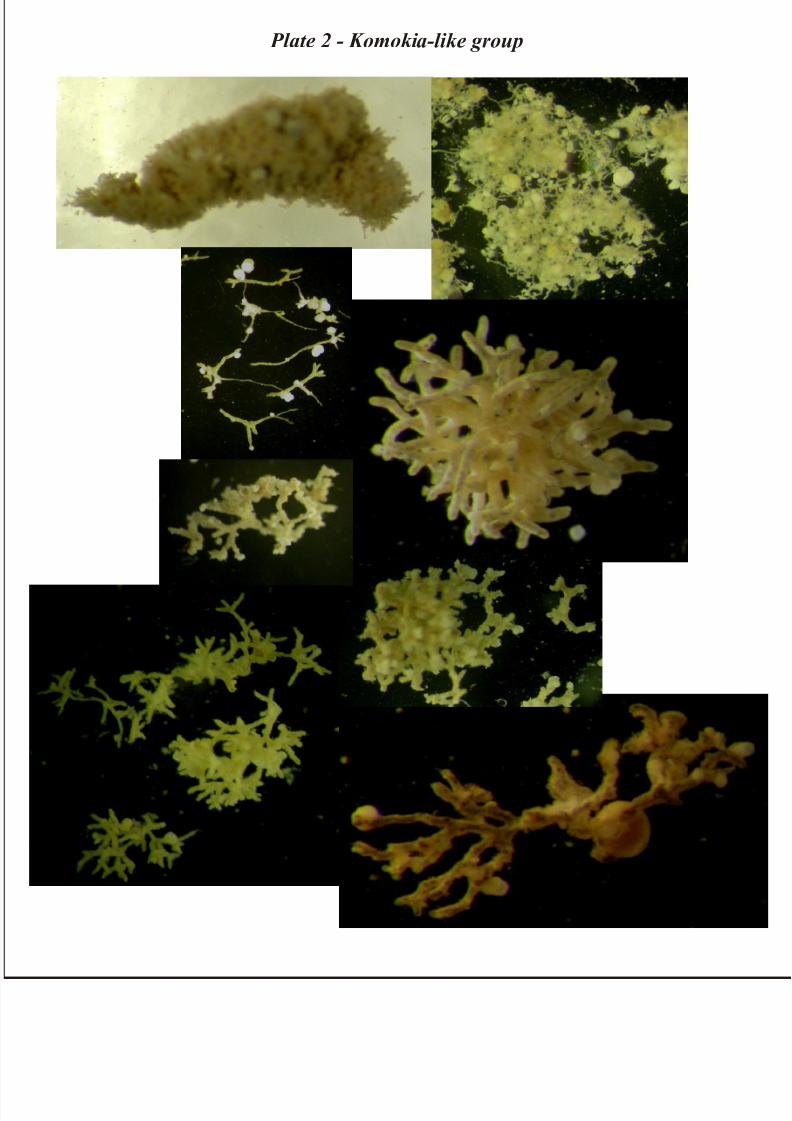
Komokia-like group
This group is dominant in samples from Porcupine Abyssal Plain area; some specimens Komoki-like
were found also in the Antarctic study area.
According to Tendal & Hessler (1977), Komokia morphotypes have generally bush –like body
consisting of mostly dichotomously branching tubules without anastomoses, the branching points
occurring at any place along the tubules. Ramification increases toward periphery of body. All tubulesare the same diameter (more less), and are nonseptate (Tendal & Hessler, 1977).
In our material the interstices between the tubules are sometimes filled with abundant sediment and the
general body form is elongate, pear or club-shaped body. Long, umbranched, empty filaments of
varying length, frequently extend from the surface. Specimens often differ from classical representatives
of the genus is consisting of only a few tubules. Globigerinacean tests are occasionally attached to
branching tubules (Plate 2).
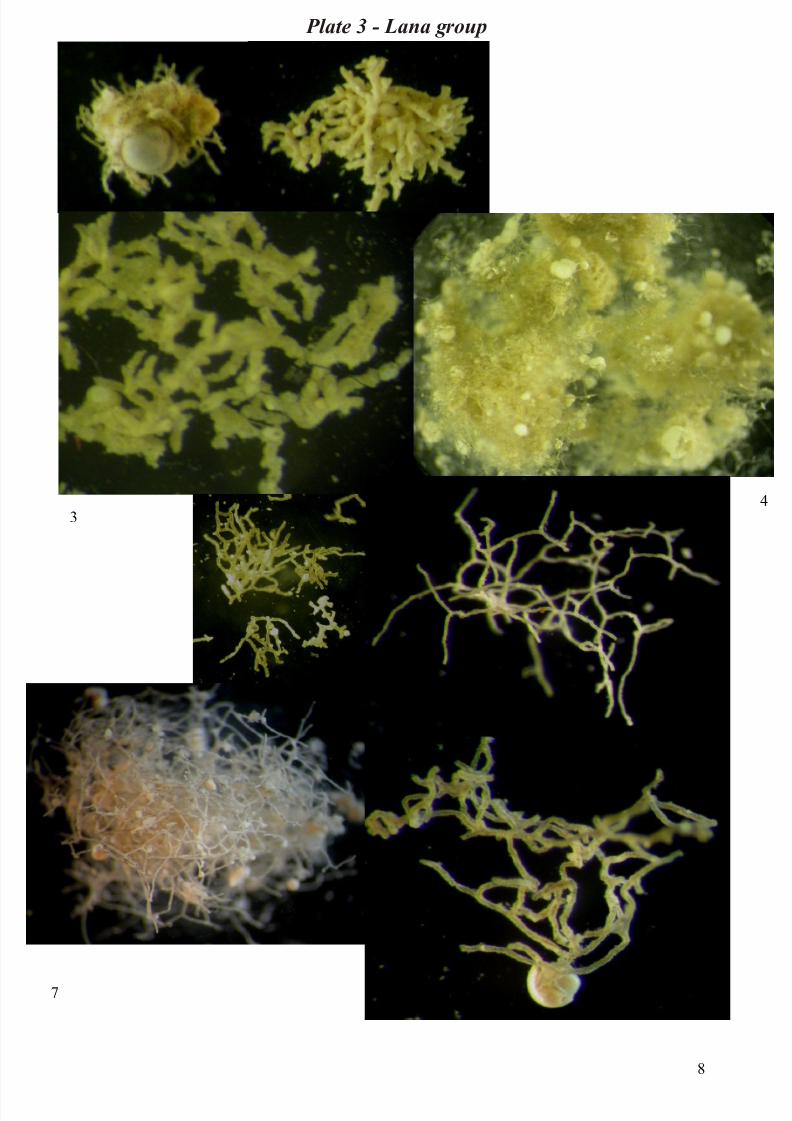
Genus Lana and Reticulum
Species of the closely-related genera Lana are common in both study areas. Lana species have the formof a loose mass of branching and anastomosing tubules with no centre of organization or pattern of
growth. The tubules are parallel sided and non-septate (Tendal & Hessler, 1977). In Reticulum the
tubules anastomose more extensively to than in the case of Lana. It is possible to recognize a large
number of morphotypes and classification is based on form of the clump, on the frequency of
anastomoses, and on the degree of filling of interstices with sediments (Gooday & Cook, 1984).
The species of Lana and Reticulum studied vary from irregularly-shaped clump of fine, or very fine,

8/12/2019 mocropaleo crusman
http://slidepdf.com/reader/full/mocropaleo-crusman 5/19
branching and anastomosing tubules of even diameter to a pear-shaped, tightly packed clump of tubules
of even diameter. There are also morphotypes represented by coherent, spongy, irregularly to
spherically-shaped masses of regularly anastomosing tubules or loose network of branching tubules
(sometimes fine) anastomosing, and not always of even diameter. The interstices of the Lana/Reticulum
tubule system are either devoid of sediment or sediment-filled; the tubules may be transparent and
tipically contain dark masses of stercomata. Globigerinacean tests are often incorporated within the
tubules network or throughout tubule clump (Plate 3).
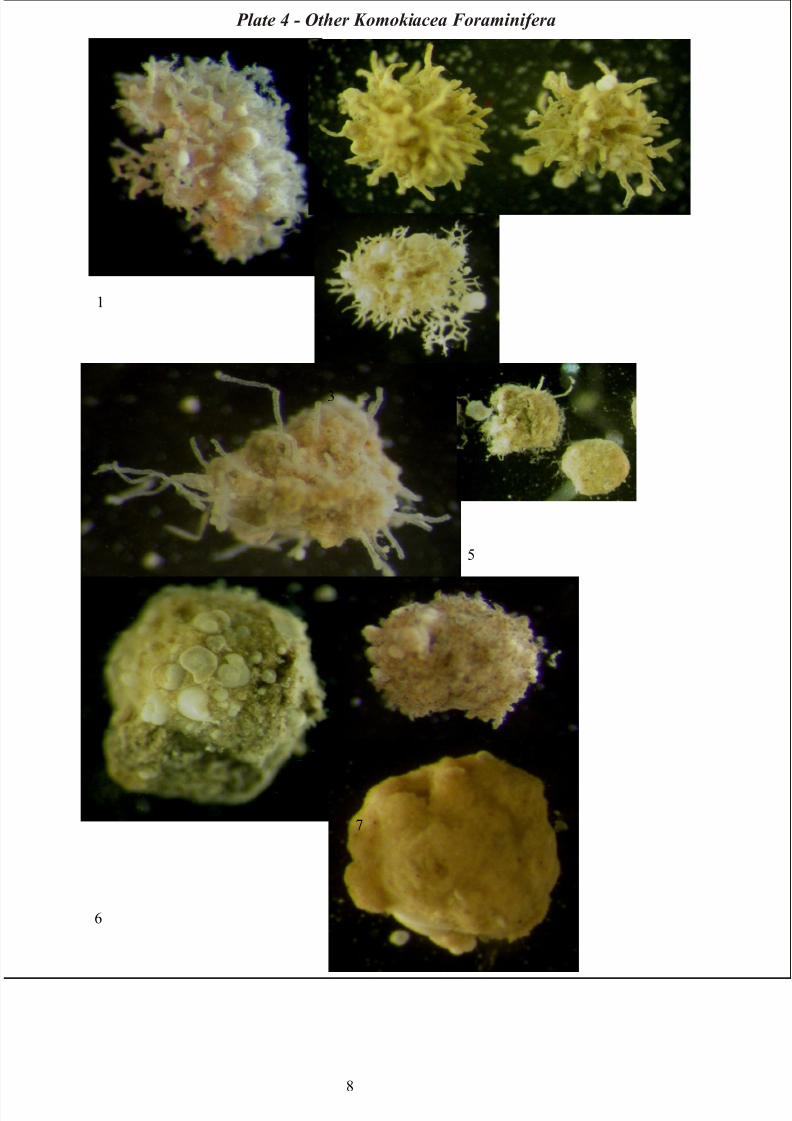
Ipoa group
Morphotypes belonging to Ipoa group are better represented in the Antarctic environment.
Most specimens are tree-like fragments treelike with a basal stem followed by burst of tightly spaced
multiple branching; diameter of tubules decreases with each branching. Tubules are avoid of septa
(Tendal & Hessler, 1977) (Plate 4).
Normanina group
Also specimens belonging to this group are also more common in the Antarctic although there are some
PAP representatives.
They have a base or center composed of tubules branching in complex, irregular pattern. Tubules radiate
outward from this base and terminate in globular or clublike chambers (Tendal & Hessler, 1977) (Plate
4).
Septuma group
Species of Septuma group in the Antarctic samples are represented by specimens with bushlike body,
sparse branching, dichotomous and nonastomosing at base. All tubules, as suggested by the group name,
are divided internally by septa with foramina (Tendal & Hessler, 1977) (Plate 4).
Edgertonia spp. and Komoki mudball spp. these are the dominant benthic Foraminifera in the Porcupine
Abyssal Plain samples. In addition to the above mentioned Komokiacea, a number of other
monothalamous Foraminifera (Muddy Tube spp., Lagenammina spp., Rhizammina spp.) and
polythalamous (Reophax spp.) Foraminifera were studied in the Antarctic samples.
Result Molecular work
Sixty-seven samples from Porcupine Abyssal Plain (NE Atlantic Ocean) and 24 samples from Weddel
Sea (Antarctica) were analysed using molecular approach. It was possible obtain only DNA sequencesfrom fragment of the SSU rRNA gene of monothalamous agglutinated Foraminifera (Astrorhiizina) and
polithalamous agglutinated and calcareous Foraminifera (Textulariina and Rotaliina). Only 7 DNA
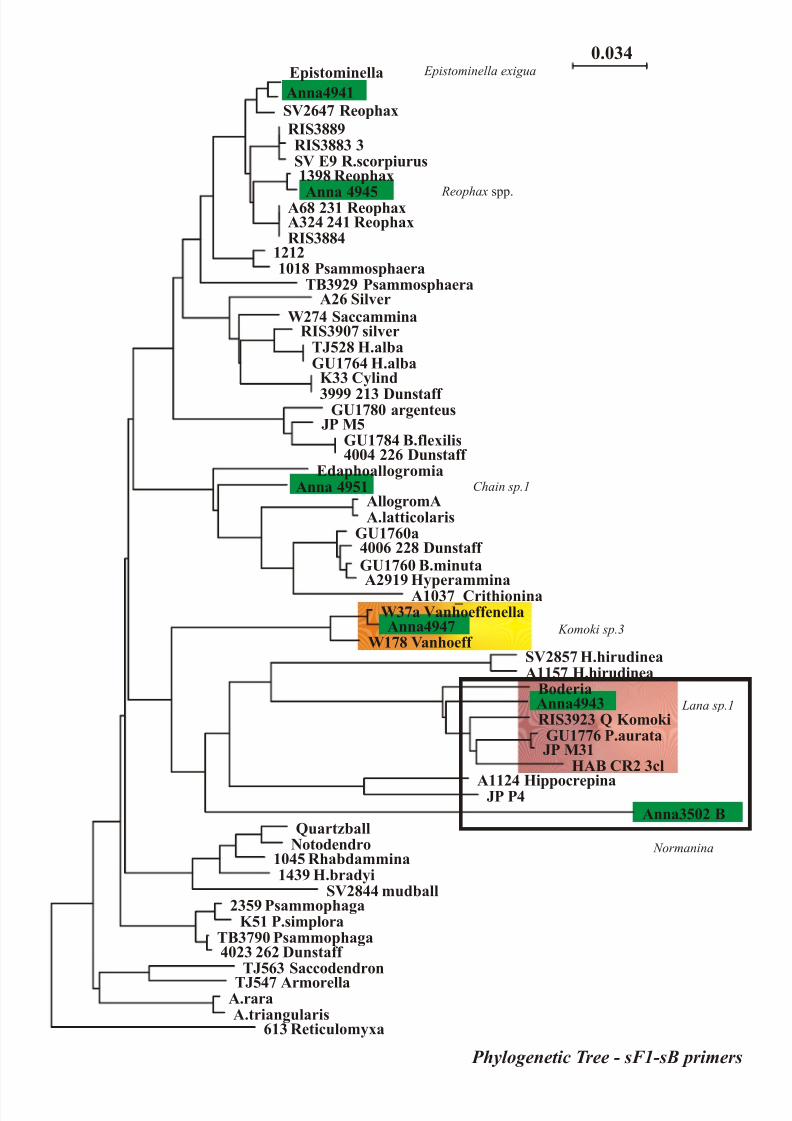
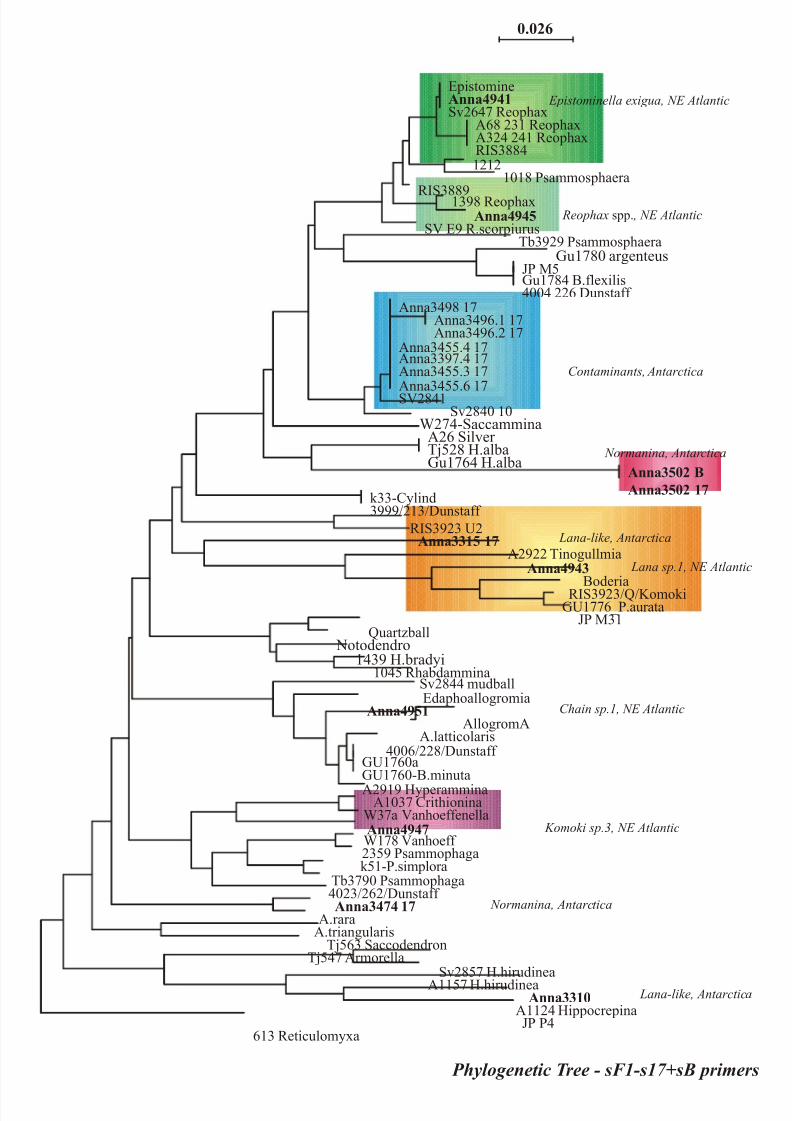
sequences from komokiacean morphotypes were obtained and results are represented in figure 3 (a-b).
We obtained also DNA sequences of monothalamous and polithalamous agglutinated Foraminfera
(Lagenammina spp., Reophax spp. and Rhizammina spp.) from the PAP; this is an important goal
because all data of this genera come from shallow water or polar settings. In this case we have the first

8/12/2019 mocropaleo crusman
http://slidepdf.com/reader/full/mocropaleo-crusman 6/19
DNA sequences from deep Atlantic Ocean.
Discussion and Conclusion
Komokiacea are an important group of macrofaunal sized agglutinated Foraminifera. This group
of protists eluded, until recently, description because fragility of many species means that they were
often lost during sample processing or their identification and classification were difficult. Even when
present in sample residues, they are difficult to recognize as organisms (Tendal, 1979), looking instead
like non descript organic detritus, poorly washed balls of sediment, or minor fragments from the surface
of some unidentified larger organisms (Tendal & Hessler, 1977) (Gooday & Cook, 1984).
Very few observations have been made on live cells of Komokiacea although they are classified
within the Foraminifera, granulo-reticulate pseudopodia have not been observed in any true
komokiacean organisms, causing some workers to doubt their assignment to this group. The group has
non clear ultrastructural identity and is distinguished by the unusual gross morphology consisting of a
system of branching of tubules.
Usually foraminiferal species are identified exclusively on the basis of morphological characters,
such as the number and form of the chambers, the form of test periphery and the type of ornament.
However, the paucity of morphological characters and their relatively large variability renders species
identification in Foraminifera and in Komokiacea particularly difficult. In this project we therefore
made the first attempts to investigate what Komokiacea are and their phylogenetic position between
Foraminifera group based on molecular data. Our results are based principally on analysis of ribosomal
DNA sequences.
During the course of this project we met problems with PCR analyses and on primers use. PCR
is a tool of unrivalled power, but as is so often the case, this power is linked to unrivalled complexity.
The source of this complexity is the PCR reaction itself-a myriad of ionic interactions, kinetic constants,and enzymatic activities, all taking place repeatedly and, hopefully, perfectly, within the space of a few
hours. Because each DNA (template) and each primer pair is different, and because molecular
systematics tend to use a wide variety of taxa or primers in a single research program, PCR reactions
need to be carefully optimised. This means that a certain amount of trial and error is an integral part of
the PCR experience. Apart from PCR conditions attention has been focused on the primers and how
strongly they anneal to the template DNA. A first series of analyses have been made using foraminiferal
primers and in particular primers with monothalamous foraminiferal “target”. We obtained few results
associated to specimens belonging to polythalamous agglutinated (Reophax) and calcareous
(Epistominella) Foraminifera. A second series of analyses were made using universal eukaryotic primers obtaining only contaminant sequences of animal and fungi. Other studies will be made using
other foraminiferal primers. There is no evidence yet, but the possibility exists that these results could
lead us to not consider Komokiacea as Foraminifera.
On the other hand morphological features, wall composition and test structure all suggest that
Komokiacea are Foraminifera.
The test of Komokiacea are partially formed organically but it also include a varying proportion

8/12/2019 mocropaleo crusman
http://slidepdf.com/reader/full/mocropaleo-crusman 7/19
of extraneous matter as the typical foraminiferal agglutinated tests. The foreign particles utilized in test
construction could reflect to some extent the local environment; material commonly utilized includes
quarts grains, various heavy minerals, clay or carbonate fragments or grains, and organic debris,
including tests of smaller Foraminifera, radiolarians, coccoliths, fragments of molluscan shells, and
sponge spicules (Loeblich & Tappan, 1964). However Komokiacea are known to accumulate very fine
particles within tubule lumen where they are presumably digested (Gooday & Cook,1984). Many
aspects of the ecology and biology of the Komokiacea are poorly understood: protoplasmic body,
nucleus, pseudopodia, gamets, cysts and in particular symbionts and parasites life history and feeding
habits are still unknown.
Little is known also about the distribution of komokiacean species (Schroder et al., 1989; Gooday
& Cook, 1984; Kamenskaya, 1993).
Systematic tree obtained from molecular data (figure 3) revealed the distribution of several
“komokiacean DNA sequences” with environmental DNA sequences or “not common foraminiferal
DNA sequences” in a clade of monothalamous agglutinated Foraminifera from several habitats. These
DNA sequences could belong to parasite organisms living in the sediment particles agglutinated by
Komokiacea. This strategy is common to several protozoans and from the literature we know that the
coccidian Trophosphaera planorbulinae (LE CALVEZ) (Loeblich and Tappan, 1964) lives in the larger
chambers of Elphidium crispum extending throughout the test and destroying it eventually. Nematode
worms may also be parasitic on them. An other condition is represented by bryozoan colonies that inter
grow as networks with the komokiacean tubules (Lana and Edgertonia genera) (Gooday & Cook, 1984)
forming a symbiotic relation between ctenostome peristomes of Bryozoa and the tubule mass of
Komokiacea. These kinds of relationships complicate the study of komokiacean molecular
identification.
Our findings suggest the following conclusions 1) to find DNA sequences of Komokiacea isnecessary perform the PCR analyses with new foraminiferal primers or other universal primers; 2) we
need to improve our knowledge of ultrastructural morphology in order to demonstrate that Komokiacea
are really Foraminifera; 3) we need more information about the natural history of Komokiacea in
general.
Figure Captions
Fig. 1 – Location Map Antarctic Samples
Fig. 2 – Location Map NE Atlantic Samples
Fig. 3 – Molecular data a) 462 sites were used for the analysis with sB primer; phylogenetic
analyses were performed with the neighbor joining method (NJ) using Kimura 2 distance algorithm;
b) 100 sites were used for analysis with s17 primer; phylogenetic analyses were performed with the
neighbor joining method (NJ) using Kimura 2 distance algorithm.

8/12/2019 mocropaleo crusman
http://slidepdf.com/reader/full/mocropaleo-crusman 8/19
Plates of Komokiacean Foraminifera from NE-Atlantic
Plate 1 – “Chain-like” komokiaceans: 1) Chain sp.1 - 2) Chain sp.2 - 3) Chain sp.3 – 4) Chain
sp.4 – 5) Chain sp.5 – 6) Chain sp.6 – 7) Chain sp.7
Plate 2 - Komokia-like group: 1) Komoki sp.1 – 2) Komoki sp.2 – 3) Komoki sp.3 – 4) Komoki
sp.4 – 5) Komoki sp.5 – 6) Komoki sp.5-bis – 7) Komoki sp.6 – 8) Komoki sp.7
Plate 3 - Lana group: 1) Lana mudball – 2) Lana sp.1 or Cerebrum (Schroder) – 3) Lana sp.2 –
4) Lana sp.3 – 5) Lana sp.4 – 6) Lana sp.5 – 7) Lana sp.5a – 8) Lana sp.6
Plate 4 – Other Komokiacean Foraminifera: 1) Ipoa spp. – 2) Septuma spp. – 3) Reticulum sp.1
– 4) Edgertonia argillispherula – 5) Edgertonia floccula – 6) Mudball Komoki sp.2 – 7) Mudball
Komoki sp.3 – 8) Mudwall Crithionina spp
Plate of Komokiacean Foraminifera from Weddel Sea
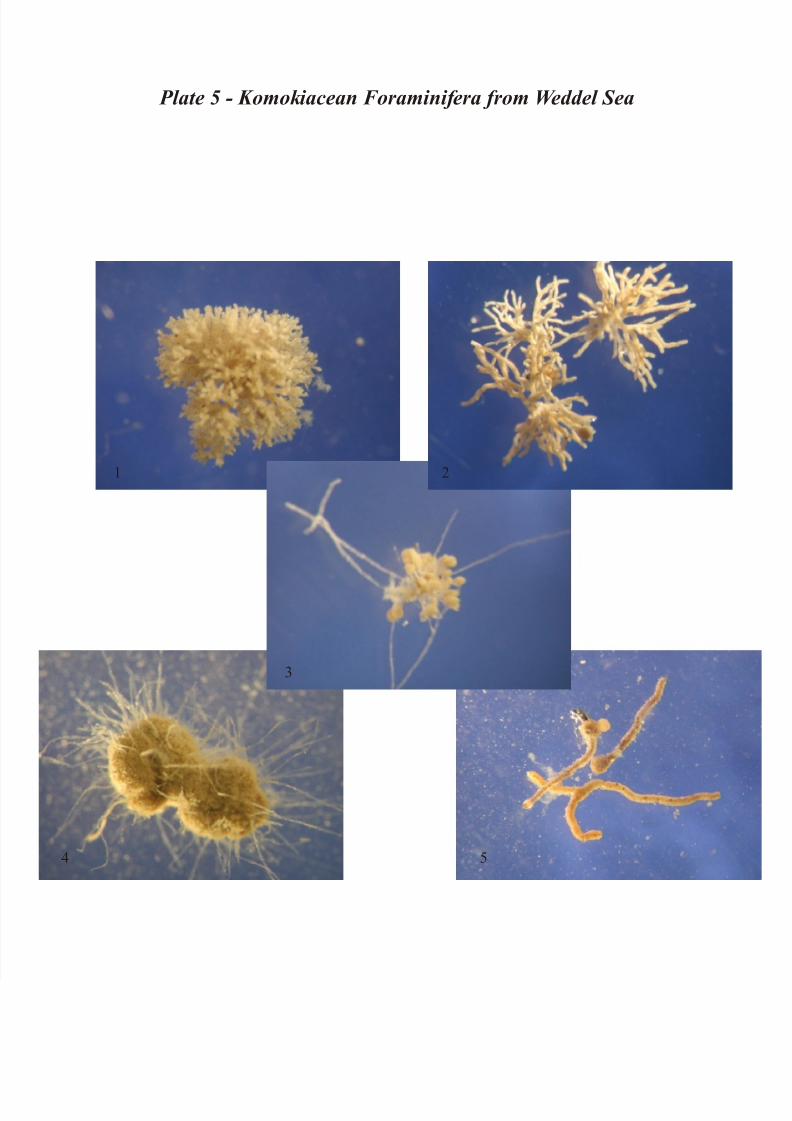
Plate 5 – Other Komokiacea Foraminifera: 1) Ipoa spp. – 2) Septuma spp. – 3) Normanina spp. –
4) Edgertonia spp.– 5) Saccorhiza spp.
R EFERENCES
BOWSER , S.S., BERNHARD, J.M., HABURA, A., GOODAY, A.J. (2002) Structure, Taxonomy and
Ecology of Astrammina triangularis (Earland), an allogromiid-like agglutinated foraminifer from
Explorers Cove, Antarctica. Journal of Foraminiferal Research 32 (2): 364-374
CEDHAGEN, T. & PAWLOWSKI J. (2002) Toxisarcon synsuicidica n.gen, n.sp., a large
monothalamous Foraminiferan from the west coast of Sweden. Journal of Foraminiferal Research
32 (2):351-357
DE LACA, T.E., BERNHARD, J.M., R EILLY, A., BOWSER , S.S. (2002) Notodendrodeshyalinosphaira (sp. nov.): structure and autoecology of an allogromiid-like agglutinated
foraminifer. Journal of Foraminiferal Research 32 (2): 177-187
GALTIER , N. & GOUY, M. (1996) SEAVIEW and PHYLO_WIN: two graphic tools for
sequence alignment and molecular phylogeny. Computer Application in the Bioscences 12: 543-548
GOODAY, A.J. (1983) Primitive Foraminifera and Xenophyophorea in IOS epibenthic sledge
samples from the Northeast Atlantic. Institute of Oceanographic Science, Report no. 156: 1-33

8/12/2019 mocropaleo crusman
http://slidepdf.com/reader/full/mocropaleo-crusman 9/19
GOODAY, A.J. (2002) Organic-walled allogromiids: aspects of their occurrence, diversity and
ecology in marine habitats. Journal of Foraminiferal Research 32
GOODAY, A.J. & NOTT, J.A. (1982) Intracellular barite crystals in two xenophyophores,
Aschemonella ramuliformis and Galatheammina sp. (Protozoa: Rhizopoda) with comments on the
taxonomy of A. ramuliformis. Journal of the marine biological association U.K . 62: 595-605
GOODAY, A.J. & COOK , P.L. (1984) An association between komokiacean foraminiferans
(Protozoa) and paludicelline ctenostomes (Bryozoa) from the abyssal northeast Atlantic. Journal
of Natural History 18: 765-784
GOODAY, A.J., NOTT, J.A., DAVIS, S., MANN, S. (1995) Apatite particles in the test wall of the
large agglutinated foraminifer Bathysiphon Major (Protista). Journal of the marine biological
association 75:469-481
GOODAY, A.J., BOWSER , S.S., BERNHARD, J.M. (1996) Benthic foraminiferal assemblages in
Explorers Cove, Antarctica: a shallow-water site with deep-sea characteristics. Progress in
Oceanography 37: 117-166
GOODAY, A.J., BETT, B.J., SHIRES, R. AND LAMBSHEAD, P.J.D. (1998) Deep-sea benthic
Foraminiferal species diversity in the NE Atlantic and NW Arabian Sea: a synthesis. Deep-Sea
Research II, 45: 165-201.
GOODAY, A.J. & PAWLOWSKI, J. (2004) Conqueria laevis gen. & sp. nov., a new soft-walled,
monothalamous foraminiferan from the deep Weddell Sea. Journal of the Marine Biological
Association of the United Kingdom, 84: 919-924.
GOODAY, A.J., HOLZMANN, M., GUIARD, M., CORNELIUS, N., & JAN PAWLOWSKI, (2004) A
new monothalamous foraminiferan from 1000-6300 m water depth in the Weddell Sea:
morphological and molecular characterisation. Deep-Sea Research II, in press.
HOLZMANN, M. & PAWLOWSKI, J. (2002) Freshwater Foraminiferans from Lake Geneva: pastand present. Journal of Foraminiferal Research 32:344-350
HOLZMANN, M., HABURA, A., GILES, H., BOWSER , S.S., & PAWLOWSKI J. (2003) Freshwater
Foraminiferans revealed by analysis of environmental DNA samples. Journal of Eukaryotic
Microbiology 50: 135-139.
MEISTERFELD, R., HOLZMANN, M., PAWLOWSKI, J. (2001) Morphological and molecular
characterization of a new terrestrial allogromiid species: Edaphoallogromia australica gen. et spec.
nov. (Foraminifera) from Northern Queensland (Australia). Protist 152: 185-192.
PAWLOWSKI, J. (2000) Introduction to the molecular systematics of Foraminifera.
Micropaleontology, 46, supplement no.1, pp.1-12.PAWLOWSKI, J., BOLIVAR , I., FAHRNI, J., DE VARGAS, C., & BOWSER , S.S. (1999) Molecular
evidence that Reticulomyxa filosa is a freshwater naked foraminifer. Journal of Eukaryotic
Microbiology 46: 612-617.
PAWLOWSKI, J., HOLZMANN, M., BERNEY, C., FAHRNI, J., CEDHAGEN, T., BOWSER , S.S.
(2002a) Phylogeny of allogromiid Foraminifera inferred from SSU rRNA gene sequences. Journal
of Foraminiferal Research 32:334-343

8/12/2019 mocropaleo crusman
http://slidepdf.com/reader/full/mocropaleo-crusman 10/19
PAWLOWSKI, J., FAHRNI, J.F., BRYKCZYNSKA , U., HABURA, A., BOWSER , S.S. (2002b)
Molecular data reveal high taxonomic diversity of allogromiid Foraminifera in Explorers Cove
(McMurdo Sound, Antarctica). Polar Biology 25: 96-105.
PAWLOWSKI, J., FAHRNI, J., BOWSER , S.S. (2002c) Phylogenetic analysis and genetic diversity
of Notodendrodes hyalinosphaira. Journal of Foraminiferal Research 32: 173-176.
PAWLOWSKI, J. & HOLZMANN, M. (2002) Molecular phylogeny of Foraminifera - a review.
European Journal of Protistology 38: 1-10.
PAWLOWSKI, J., HOLZMANN, M., BERNEY, C., FAHRNI, J., GOODAY, A.J., CEDHAGEN, T.,
HABURA, A., BOWSER , S.S. (2003) The evolution of early Foraminifera. Proc. Nat. Acad. Sci. USA
(in press)
SABBATINI, A., PAWLOWSKI, J., GOODAY, A.J., PIRAINO, S., BOWSER , S.S., MORIGI, C.,
NEGRI, A. (2004) Vellaria antarctica n.sp., a new small monothalamous foraminifer from Terra
Nova Bay, Antarctica. Antarctic Science 16(3): 307-312.
SCHRODER , C.J., MEDIOLI, F.S., SCOTT, D.B. (1989) Fragile abyssal foraminifera (including
Komokiacea) from the Nares Abyssal Plain. Micropaleontology 35: 10-48
TKACH, V. & PAWLOWSKI J. (1999) A new method of DNA extraction from ethanol-fixed
parasitic worms. Acta Parasitologica 44: 147-148
TENDAL, O.S. (1972) A monograph of the Xenophyophoria (Rhizopoda, Protozoa). Galathea
Report 12: 7-99, pls 1-17
TENDAL, O.S. & HESSLER , R.R. (1977) An introduction to the biology and systematics of
komokiacea (Textulariina, Foraminiferida). Galathea Report 14: 165-194, pls 9-26
TENDAL, O.S. (1979) Aspects of the biology of Komokiacea and Xenophyophoria.Sarsia 64:
13-17.
WILDING, T. Taxonomy and ecology of Toxisarcon alba sp.nov. from Loch Linnhe, westcoast of Scotland, UK. Journal of Foraminiferal Research 32: 358-363.
K AMENSKAYA, O.E. (1990) Preliminary date on the komoki (Komokioidea, Foraminifera)
from the transatlantic section (31ºS - 36ºS). Transactions of the Shirshov Institute of Oceanology
126: 62-66.
K AMENSKAYA, O.E. (1993) Komokiacea from the region of the underwater seamount
Valdivia (South-Eastern Atlantic). Transactions of the Shirshov Institute of Oceanology, 127: 72-
81.
8/12/2019 mocropaleo crusman
http://slidepdf.com/reader/full/mocropaleo-crusman 11/19
8/12/2019 mocropaleo crusman
http://slidepdf.com/reader/full/mocropaleo-crusman 12/19
-25 -20 -15 -10 -5 0
30
35
40
45
50
55
60
2 0 0 m
4 0 0 0 m
3 0 0 0 m
2 0 0 0 m
1 0 0 0 m
R o c k a l l
B a n k
R o c k a
l l T r o u
g h
P o r c u
p i n e
A b y s s a l P
l a i n
PAP
M a d e i r a
A b y s s a l P
l a i n
I b e r i a
n
A b y s s a l P
l a i n
Figure 1. - Location Map
8/12/2019 mocropaleo crusman
http://slidepdf.com/reader/full/mocropaleo-crusman 13/19
Phylogenetic Tree - sF1-sB primers
0.034Epistominella
Anna4941
SV2647 Reophax
RIS3889RIS3883 3SV E9 R.scorpiurus1398 Reophax
Anna 4945A68 231 ReophaxA324 241 ReophaxRIS3884
12121018 Psammosphaera
TB3929 PsammosphaeraA26 Silver
W274 SaccamminaRIS3907 silver
TJ528 H.albaGU1764 H.alba
K33 Cylind
3999 213 Dunstaff GU1780 argenteus
JP M5GU1784 B.flexilis4004 226 Dunstaff
EdaphoallogromiaAnna 4951
AllogromAA.latticolaris
GU1760a4006 228 Dunstaff GU1760 B.minutaA2919 Hyperammina
A1037_CrithioninaW37a VanhoeffenellaAnna4947
W178 Vanhoeff SV2857 H.hirudineaA1157 H.hirudinea
BoderiaAnna4943RIS3923 Q Komoki
GU1776 P.aurataJP M31
HAB CR2 3cl
A1124 HippocrepinaJP P4Anna3502 B
QuartzballNotodendro
1045 Rhabdammina1439 H.bradyi
SV2844 mudball2359 Psammophaga
K51 P.simploraTB3790 Psammophaga4023 262 Dunstaff
TJ563 Saccodendron
TJ547 ArmorellaA.raraA.triangularis
613 Reticulomyxa
Lana sp.1
Komoki sp.3
Normanina
Chain sp.1
Reophax spp.
Epistominella exigua
8/12/2019 mocropaleo crusman
http://slidepdf.com/reader/full/mocropaleo-crusman 14/19
0.026
EpistomineAnna4941
Sv2647 ReophaxA68 231 ReophaxA324 241 ReophaxRIS38841212
1018 Psammosphaera
RIS38891398 ReophaxAnna4945
SV E9 R.scorpiurusTb3929 Psammosphaera
Gu1780 argenteusJP M5Gu1784 B.flexilis4004 226 Dunstaff
Anna3498 17Anna3496.1 17Anna3496.2 17
Anna3455.4 17Anna3397.4 17Anna3455.3 17
Anna3455.6 17SV2841Sv2840 10
W274-SaccamminaA26 Silver Tj528 H.albaGu1764 H.alba
Anna3502 B
Anna3502 17k33-Cylind3999/213/Dunstaff
RIS3923 U2Anna3315 17
A2922 TinogullmiaAnna4943
BoderiaRIS3923/Q/Komoki
GU1776_P.aurataJP M31
Quartzball Notodendro
1439 H.bradyi1045 Rhabdammina
Sv2844 mudballEdaphoallogromia
Anna4951
AllogromAA.latticolaris
4006/228/Dunstaff GU1760aGU1760-B.minutaA2919 Hyperammina
A1037 CrithioninaW37a VanhoeffenellaAnna4947W178 Vanhoeff 2359 Psammophagak51-P.simplora
Tb3790 Psammophaga4023/262/Dunstaff Anna3474 17
A.raraA.triangularis
Tj563 SaccodendronTj547 Armorella
Sv2857 H.hirudineaA1157 H.hirudinea
Anna3310
A1124 HippocrepinaJP P4
613 Reticulomyxa
Lana-like, Antarctica
Contaminants, Antarctica
Lana sp.1, NE Atlantic
Normanina, Antarctica
Chain sp.1, NE Atlantic
Komoki sp.3, NE Atlantic
Normanina, Antarctica
Lana-like, Antarctica
Phylogenetic Tree - sF1-s17+sB primers
Epistominella exigua, NE Atlantic
Reophax spp. , NE Atlantic
8/12/2019 mocropaleo crusman
http://slidepdf.com/reader/full/mocropaleo-crusman 15/19
1 2
3 4
5 6
7
Plate 1 - “Chain-like” komokiaceans
8/12/2019 mocropaleo crusman
http://slidepdf.com/reader/full/mocropaleo-crusman 16/19
4
3
5
6
7
8
Plate 2 - Komokia-like group
8/12/2019 mocropaleo crusman
http://slidepdf.com/reader/full/mocropaleo-crusman 17/19
1 2
3
4
5 6
7
8
Plate 3 - Lana group
8/12/2019 mocropaleo crusman
http://slidepdf.com/reader/full/mocropaleo-crusman 18/19
1
2
3
4
5
6
7
8
Plate 4 - Other Komokiacea Foraminifera
8/12/2019 mocropaleo crusman
http://slidepdf.com/reader/full/mocropaleo-crusman 19/19
Plate 5 - Komokiacean Foraminifera from Weddel Sea
1 2
3
4 5





























