Embed Size (px)
Citation preview

Mimicking the Plant Cell Interior under WaterStress by Macromolecular Crowding: DisorderedDehydrin Proteins Are Highly Resistant toStructural Collapse1[W]
Jean-Marie Mouillon2, Sylvia K. Eriksson, and Pia Harryson*
Umea Plant Science Center, Department of Plant Physiology, Umea University, S–901 87 Umea, Sweden(J.-M.M.); and Department of Biochemistry and Biophysics, Arrhenius Laboratories for Natural Sciences,Stockholm University, S–106 91 Stockholm, Sweden (S.K.E., P.H.)
The dehydrins are a class of drought-induced proteins in plants that lack a fixed three-dimensional structure. Their specificmolecular action, aswell as the reason for their disordered character, is as yet poorly understood. It has been speculated, however,that the dehydrins are tuned to acquire a biologically active structure only under the conditions in which they normally function(i.e. upon dehydration). To test this hypothesis, we here investigate the effect of reduced water content and macromolecularcrowding on three dehydrins from Arabidopsis (Arabidopsis thaliana). As a simplistic model for mimicking cellular dehydration,we used polyethylene glycol, glycerol, and sugars that plants naturally employ as compatible solutes (i.e. sucrose and glucose).Macromolecular crowding was induced by the large polysaccharides Ficoll and dextran. The results show that the dehydrins areremarkably stable in their disordered state and are only modestly affected by the solvent alterations. A notable exception is thedehydrinCor47,which shows a small, intrinsic increase in helical structure at high concentrations of osmolytes.We also examinedthe effect of phosphorylation but found no evidence that such posttranslational modifications of the dehydrin sequencesmodulate their structural response to osmolytes and crowding agents. These results suggest that the dehydrins are highlyspecializedproteins that have evolved tomaintain their disordered character under conditions inwhichunfolded states of severalglobular proteins would tend to collapse.
The loss of water from plant cells during droughtand freezing triggers a series of adaptation processes.One of these is the production of osmolytes such asSuc, Pro, and betaine (Hasegawa et al., 2000; Wanget al., 2003). As a consequence, the already crowdedinterior of the cell gets even denser and the physio-logical processes need to adjust to elevated back-grounds of nonreacting macromolecules and stericconfinement. In some cases, the increase in macromo-lecular density can be radical, going from 300 to 400 gL21 under ideal growth conditions to.900 g L21 uponsevere desiccation (Ellis, 2001a, 2001b; Bryant et al.,2005). Such cellular crowding not only presents achallenge in terms of modulating diffusion-basedchemical reactions (Ellis, 2001a; Minton, 2005a, 2005b,
2006) but will also affect structural integrity. Crowd-ing promotes protein assembly by favoring compactconformations over extended ones, and the reductionin bulk water will directly influence membrane to-pology and dynamics. The molecular conditions inwater-starved plant cells, therefore, are very differentfrom those normally used to characterize biochemicalprocesses in vitro. It has been put forward that insuch situations the biological function of proteinscould be regulated through crowding-induced con-formational changes (Hall, 2006; Hall and Dobson,2006). Under these extreme conditions, the plant cellsrespond by expressing a characteristic class of stressproteins, the dehydrins (Close, 1997; Garay-Arroyoet al., 2000; Fig. 1).
The function of dehydrins is as yet unknown. Acomplicating factor, from a biochemical perspective, isthat they seem to lack ordered structure (Lisse et al.,1996; Soulages et al., 2003; Mouillon et al., 2006). Evenso, all dehydrins contain, by definition, at least onecopy of a highly conserved sequence segment, theK-seg, and may or may not include other conservedsequences called the S-seg, the Y-seg (Close, 1996), andthe charged peptide 1 (ChP-1; Mouillon et al., 2006).Since the dehydrins accumulate at very high concen-trations inside cells (Bartels and Salamini, 2001), theyare unlikely to be signal molecules or conventionalenzymes. The role of the dehydrins is probably of a
1 This work was supported by the Swedish Research Council forthe Environment, Agricultural Sciences, and Spatial Planning (P.H.),the Carl Tryggers Foundation (P.H.), and the Lawski Foundation(S.K.E.).
2 Present address: Fluxome Sciences A/S, Diplomvej 378, DK–2800 Lyngby, Denmark.
* Corresponding author; e-mail [email protected] author responsible for distribution of materials integral to the
findings presented in this article in accordance with the policydescribed in the Instructions for Authors (www.plantphysiol.org) is:Pia Harryson ([email protected]).
[W] The online version of this article contains Web-only data.www.plantphysiol.org/cgi/doi/10.1104/pp.108.124099
Plant Physiology, December 2008, Vol. 148, pp. 1925–1937, www.plantphysiol.org � 2008 American Society of Plant Biologists 1925 www.plantphysiol.orgon April 11, 2019 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

more general nature. For example, they may stabilizemembranes, act as chaperones, or by other meansbuffer the altered solvent properties inside water-stressed cells (Close, 1996; Garay-Arroyo et al., 2000;Boudet et al., 2006). As a clue to how the stressresponse is orchestrated molecularly, it has beensuggested that the functional structure of the dehy-drins is induced by their hydration status (Boudetet al., 2006). Accordingly, the disordered appearance ofthe dehydrins may be converted into active, three-dimensional structures, provided that the conditionsinside a drought-stressed plant cell are sufficientlywell reproduced. The disordered conformations seenunder dilute conditions would then represent thedenatured state and, consequently, tell very little aboutthe dehydrin’s actual biological function. This idea issupported by the observations that water can modu-late ionization potentials, pKA values, and protein-binding potentials (Kornblatt and Kornblatt, 2002).Other classes of disordered proteins have been foundto participate in molecular recognition, regulation, andcell signaling (Dunker et al., 2002; Uversky et al., 2005).Function, then, is induced by folding of the disorderedprotein into a structured state, where the functionalgroups are placed at the right locations for interac-tions. Folding can be initiated (or templated) by bind-ing to a biological partner such as other proteins,DNA, RNA, or metals (Dunker et al., 2002; Tompa andCsermely, 2004).
Of particular interest with respect to the dehydrinsis that the conformational changes are sometimesconditional (i.e. the protein structure responds tochanges in the environment, such as temperature[McNulty et al., 2006a], pH [Tornroth-Horsefieldet al., 2006], availability of water [Luo and Baldwin,1997], or macromolecular crowding [Minton, 2005a,
2005b; Hall, 2006]). Disorder is used to allow thefunction to be switched on and off (e.g. a 10% changein the concentration of intracellular proteins can leadto changes up to a factor 10 in the activity of molecularregulatory species; Al-Habori, 2001). Computationalstudies have recently demonstrated that the effect ofcrowding, despite its underlying complexity, can besimply mimicked by encapsulation of the proteins inspherical pores (Klimov et al., 2002; Cheung andThirumalai, 2007). This freedom in how the crowdingpressure can be applied suggests that simplified ex-periments in which crowding effects are tested by theaddition of large branched polymers, such as Ficolland dextran, could produce valid approximations ofthe in vivo condition. In fact, it was recently verified byNMR that the effect of such crowding agents in vitroreproduces the intracellular conditions fairly well(Selenko et al., 2006).
In this study, we examine whether the addition ofmacromolecular crowding agents in vitro can force thedehydrins to fold. The progressive substitution ofwater with various polymers is performed as a simplemodel of how the cellular interior changes upondrought/desiccation. As representative crowding agents,we chose a series of natural and synthetic osmolytes,the effects of which are well characterized in othersystems. To distinguish the response of the full-lengthdehydrins from that of their constituent K-seg andChP-1 segments, the latter have been analyzed sepa-rately in the form of peptides. The results show thatboth the full-length dehydrins and the isolated K-segand ChP-1 segments retain their disordered characterat extremely high levels of osmolytes. Moreover, thepropensity of the dehydrins to undergo unspecificcollapse (compare with amorphous glass) appearseven lower than that of denatured states of globularproteins. Taken together, these observations suggestthat the dehydrin sequence is highly evolved andadapted to remain disordered under conditions ofsevere dehydration. In this respect, the dehydrins aredifferent from the class of disordered proteins that relyon folding to become functional. The function of thedehydrins, therefore, is likely to lie in the interactionsof the conserved segments with their specific biolog-ical targets.
RESULTS
Analysis of Structural Transitions Using
Circular Dichroism
Circular dichroism (CD) is a common spectroscopicmethod for detecting secondary structure in proteins,where a-helices, b-sheets, and random coils displaycharacteristic spectra at wavelengths of 190 to 250 nm(Fig. 2). Accordingly, the CD spectrum of a typicalfolded protein is a characteristic structural fingerprintmade up by the sum of its constituent secondary-structure elements. In physiological buffer, however,
Figure 1. Distribution of the conserved segments in the sequences ofthe dehydrins Cor47, Lti29, and Lti30 from Arabidopsis. The dehydrinsconstitute group 2 of the large family of the hydrophilic LEA proteins(Close, 1996; Bartels and Salamini, 2001). The amino acid sequencesare as follows: Cor47, MAEEYKNNVPEHETPTVATEESPATTTEVTDRGL-FDFLGKKEEEVKPQETTTLESEFDHKAGISEPELAAEHEEVKENKITLLEEL-GEKTEEDEENKPSVIEKLHRSNSSSSSSSDEEGEEKKEKKKKIVEGEEDKK-GLVEKIKEKLPGHHDKTAEDDVPVSTTIPVPVSESVVEHDHPEIKEKLPG-YHAKTTEEEVKKEKESDD; Lti29, MAEEYKNTYPEGETPKVATEESSAPE-IKERGMFDFLKKKEEVKPQETTTLASEFEHKTGISEPESFVAKHEEEEHKPT-LLEGLHQKHEEEEENKPSLLDKLHRSNSSSSSSSDEEGEDGEKPEEEEKK-GFMDKIKEKLPGHSKKPEDSGVVNTTPLVETATPIADIPEEKKGFMDKIK-EKLPGYHAKTTGEEEKKEKVSD; and Lti30, MNSHQNQTGVQKKG-ITEKIMEKLPGHHGPTNTGVVHHEKKGMTEKVMEQLPGHHGATGT-GGVHHEKKGMTEKVMEQLPGHHGSHQTGTNTTYGTTNTGGVH-HEKKSVTEKVMEKLPGHHGSHQTGTNTAYGTNTNVVHHEKKGIAEK-IKEGLPGHHGTHKTGTTTSYGNTGVVHHENKST.
Mouillon et al.
1926 Plant Physiol. Vol. 148, 2008 www.plantphysiol.orgon April 11, 2019 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.
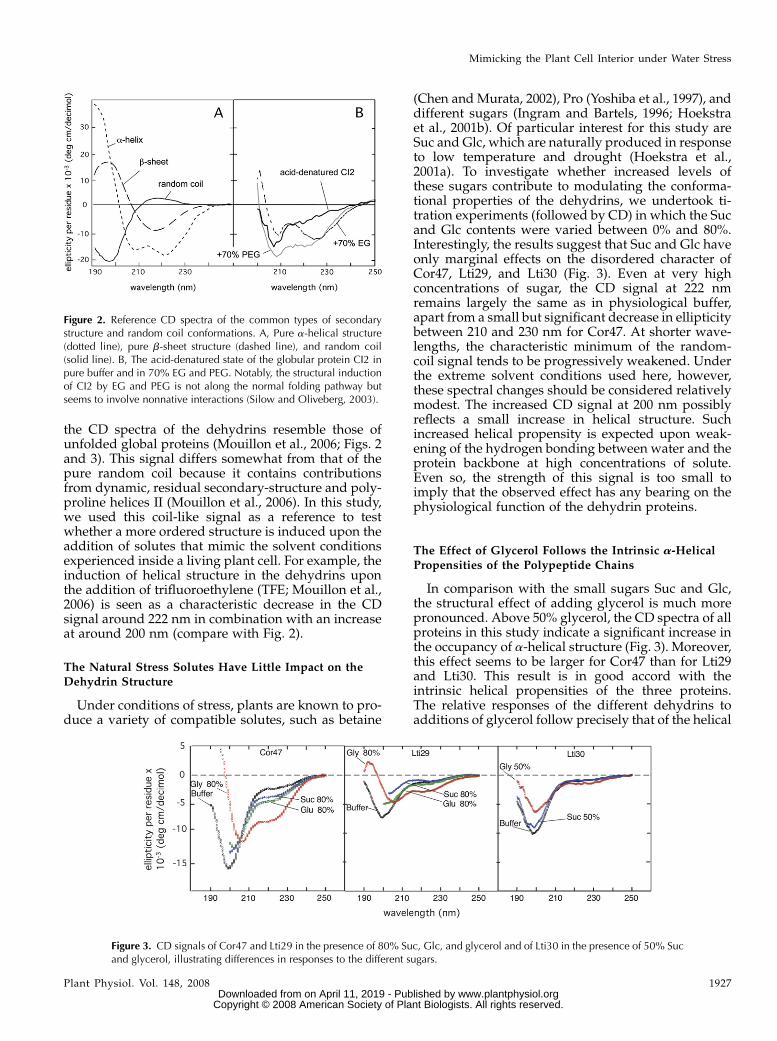
the CD spectra of the dehydrins resemble those ofunfolded global proteins (Mouillon et al., 2006; Figs. 2and 3). This signal differs somewhat from that of thepure random coil because it contains contributionsfrom dynamic, residual secondary-structure and poly-proline helices II (Mouillon et al., 2006). In this study,we used this coil-like signal as a reference to testwhether a more ordered structure is induced upon theaddition of solutes that mimic the solvent conditionsexperienced inside a living plant cell. For example, theinduction of helical structure in the dehydrins uponthe addition of trifluoroethylene (TFE; Mouillon et al.,2006) is seen as a characteristic decrease in the CDsignal around 222 nm in combination with an increaseat around 200 nm (compare with Fig. 2).
The Natural Stress Solutes Have Little Impact on theDehydrin Structure
Under conditions of stress, plants are known to pro-duce a variety of compatible solutes, such as betaine
(Chen and Murata, 2002), Pro (Yoshiba et al., 1997), anddifferent sugars (Ingram and Bartels, 1996; Hoekstraet al., 2001b). Of particular interest for this study areSuc and Glc, which are naturally produced in responseto low temperature and drought (Hoekstra et al.,2001a). To investigate whether increased levels ofthese sugars contribute to modulating the conforma-tional properties of the dehydrins, we undertook ti-tration experiments (followed by CD) in which the Sucand Glc contents were varied between 0% and 80%.Interestingly, the results suggest that Suc and Glc haveonly marginal effects on the disordered character ofCor47, Lti29, and Lti30 (Fig. 3). Even at very highconcentrations of sugar, the CD signal at 222 nmremains largely the same as in physiological buffer,apart from a small but significant decrease in ellipticitybetween 210 and 230 nm for Cor47. At shorter wave-lengths, the characteristic minimum of the random-coil signal tends to be progressively weakened. Underthe extreme solvent conditions used here, however,these spectral changes should be considered relativelymodest. The increased CD signal at 200 nm possiblyreflects a small increase in helical structure. Suchincreased helical propensity is expected upon weak-ening of the hydrogen bonding between water and theprotein backbone at high concentrations of solute.Even so, the strength of this signal is too small toimply that the observed effect has any bearing on thephysiological function of the dehydrin proteins.
The Effect of Glycerol Follows the Intrinsic a-Helical
Propensities of the Polypeptide Chains
In comparison with the small sugars Suc and Glc,the structural effect of adding glycerol is much morepronounced. Above 50% glycerol, the CD spectra of allproteins in this study indicate a significant increase inthe occupancy of a-helical structure (Fig. 3). Moreover,this effect seems to be larger for Cor47 than for Lti29and Lti30. This result is in good accord with theintrinsic helical propensities of the three proteins.The relative responses of the different dehydrins toadditions of glycerol follow precisely that of the helical
Figure 2. Reference CD spectra of the common types of secondarystructure and random coil conformations. A, Pure a-helical structure(dotted line), pure b-sheet structure (dashed line), and random coil(solid line). B, The acid-denatured state of the globular protein CI2 inpure buffer and in 70% EG and PEG. Notably, the structural inductionof CI2 by EG and PEG is not along the normal folding pathway butseems to involve nonnative interactions (Silow and Oliveberg, 2003).
Figure 3. CD signals of Cor47 and Lti29 in the presence of 80% Suc, Glc, and glycerol and of Lti30 in the presence of 50% Sucand glycerol, illustrating differences in responses to the different sugars.
Mimicking the Plant Cell Interior under Water Stress
Plant Physiol. Vol. 148, 2008 1927 www.plantphysiol.orgon April 11, 2019 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.
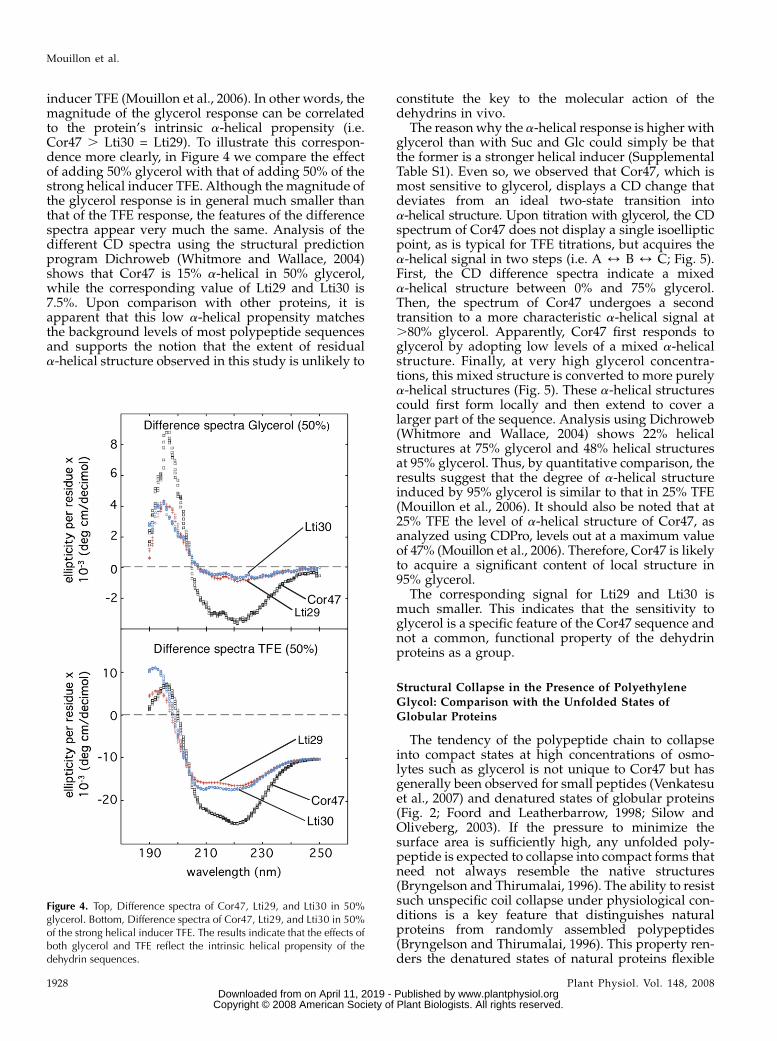
inducer TFE (Mouillon et al., 2006). In other words, themagnitude of the glycerol response can be correlatedto the protein’s intrinsic a-helical propensity (i.e.Cor47 . Lti30 = Lti29). To illustrate this correspon-dence more clearly, in Figure 4 we compare the effectof adding 50% glycerol with that of adding 50% of thestrong helical inducer TFE. Although the magnitude ofthe glycerol response is in general much smaller thanthat of the TFE response, the features of the differencespectra appear very much the same. Analysis of thedifferent CD spectra using the structural predictionprogram Dichroweb (Whitmore and Wallace, 2004)shows that Cor47 is 15% a-helical in 50% glycerol,while the corresponding value of Lti29 and Lti30 is7.5%. Upon comparison with other proteins, it isapparent that this low a-helical propensity matchesthe background levels of most polypeptide sequencesand supports the notion that the extent of residuala-helical structure observed in this study is unlikely to
constitute the key to the molecular action of thedehydrins in vivo.
The reasonwhy the a-helical response is higher withglycerol than with Suc and Glc could simply be thatthe former is a stronger helical inducer (SupplementalTable S1). Even so, we observed that Cor47, which ismost sensitive to glycerol, displays a CD change thatdeviates from an ideal two-state transition intoa-helical structure. Upon titration with glycerol, the CDspectrum of Cor47 does not display a single isoellipticpoint, as is typical for TFE titrations, but acquires thea-helical signal in two steps (i.e. A 4 B 4 C; Fig. 5).First, the CD difference spectra indicate a mixeda-helical structure between 0% and 75% glycerol.Then, the spectrum of Cor47 undergoes a secondtransition to a more characteristic a-helical signal at.80% glycerol. Apparently, Cor47 first responds toglycerol by adopting low levels of a mixed a-helicalstructure. Finally, at very high glycerol concentra-tions, this mixed structure is converted to more purelya-helical structures (Fig. 5). These a-helical structurescould first form locally and then extend to cover alarger part of the sequence. Analysis using Dichroweb(Whitmore and Wallace, 2004) shows 22% helicalstructures at 75% glycerol and 48% helical structuresat 95% glycerol. Thus, by quantitative comparison, theresults suggest that the degree of a-helical structureinduced by 95% glycerol is similar to that in 25% TFE(Mouillon et al., 2006). It should also be noted that at25% TFE the level of a-helical structure of Cor47, asanalyzed using CDPro, levels out at a maximum valueof 47% (Mouillon et al., 2006). Therefore, Cor47 is likelyto acquire a significant content of local structure in95% glycerol.
The corresponding signal for Lti29 and Lti30 ismuch smaller. This indicates that the sensitivity toglycerol is a specific feature of the Cor47 sequence andnot a common, functional property of the dehydrinproteins as a group.
Structural Collapse in the Presence of Polyethylene
Glycol: Comparison with the Unfolded States ofGlobular Proteins
The tendency of the polypeptide chain to collapseinto compact states at high concentrations of osmo-lytes such as glycerol is not unique to Cor47 but hasgenerally been observed for small peptides (Venkatesuet al., 2007) and denatured states of globular proteins(Fig. 2; Foord and Leatherbarrow, 1998; Silow andOliveberg, 2003). If the pressure to minimize thesurface area is sufficiently high, any unfolded poly-peptide is expected to collapse into compact forms thatneed not always resemble the native structures(Bryngelson and Thirumalai, 1996). The ability to resistsuch unspecific coil collapse under physiological con-ditions is a key feature that distinguishes naturalproteins from randomly assembled polypeptides(Bryngelson and Thirumalai, 1996). This property ren-ders the denatured states of natural proteins flexible
Figure 4. Top, Difference spectra of Cor47, Lti29, and Lti30 in 50%glycerol. Bottom, Difference spectra of Cor47, Lti29, and Lti30 in 50%of the strong helical inducer TFE. The results indicate that the effects ofboth glycerol and TFE reflect the intrinsic helical propensity of thedehydrin sequences.
Mouillon et al.
1928 Plant Physiol. Vol. 148, 2008 www.plantphysiol.orgon April 11, 2019 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.
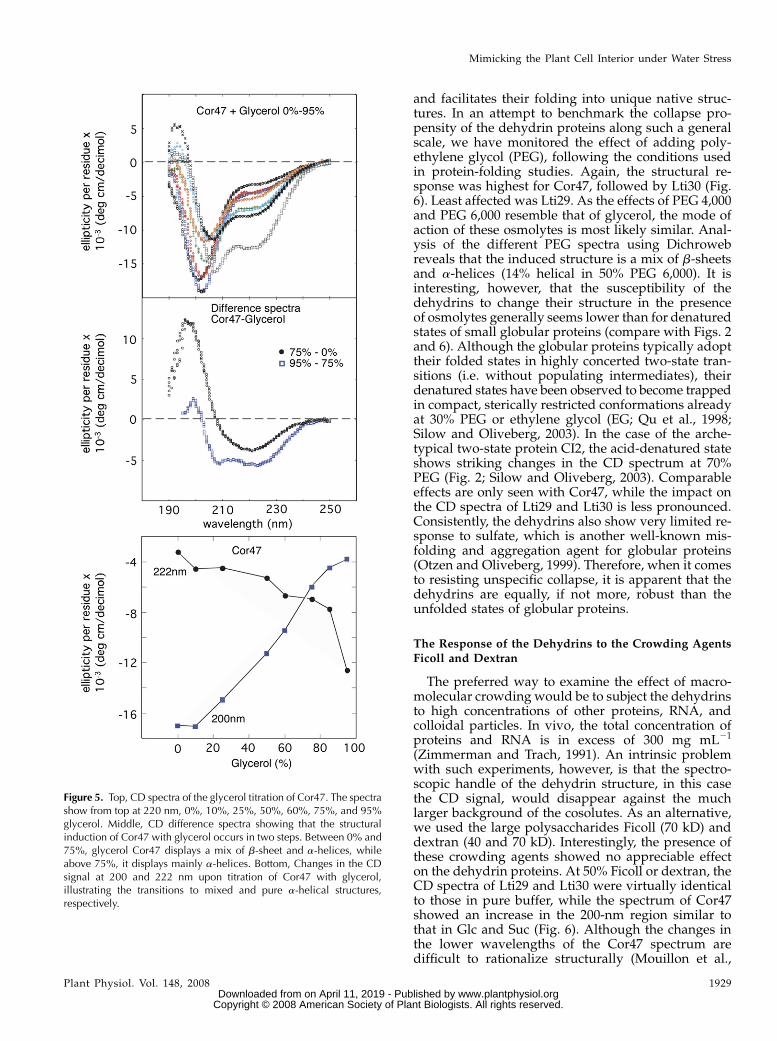
and facilitates their folding into unique native struc-tures. In an attempt to benchmark the collapse pro-pensity of the dehydrin proteins along such a generalscale, we have monitored the effect of adding poly-ethylene glycol (PEG), following the conditions usedin protein-folding studies. Again, the structural re-sponse was highest for Cor47, followed by Lti30 (Fig.6). Least affected was Lti29. As the effects of PEG 4,000and PEG 6,000 resemble that of glycerol, the mode ofaction of these osmolytes is most likely similar. Anal-ysis of the different PEG spectra using Dichrowebreveals that the induced structure is a mix of b-sheetsand a-helices (14% helical in 50% PEG 6,000). It isinteresting, however, that the susceptibility of thedehydrins to change their structure in the presenceof osmolytes generally seems lower than for denaturedstates of small globular proteins (compare with Figs. 2and 6). Although the globular proteins typically adopttheir folded states in highly concerted two-state tran-sitions (i.e. without populating intermediates), theirdenatured states have been observed to become trappedin compact, sterically restricted conformations alreadyat 30% PEG or ethylene glycol (EG; Qu et al., 1998;Silow and Oliveberg, 2003). In the case of the arche-typical two-state protein CI2, the acid-denatured stateshows striking changes in the CD spectrum at 70%PEG (Fig. 2; Silow and Oliveberg, 2003). Comparableeffects are only seen with Cor47, while the impact onthe CD spectra of Lti29 and Lti30 is less pronounced.Consistently, the dehydrins also show very limited re-sponse to sulfate, which is another well-known mis-folding and aggregation agent for globular proteins(Otzen and Oliveberg, 1999). Therefore, when it comesto resisting unspecific collapse, it is apparent that thedehydrins are equally, if not more, robust than theunfolded states of globular proteins.
The Response of the Dehydrins to the Crowding AgentsFicoll and Dextran
The preferred way to examine the effect of macro-molecular crowding would be to subject the dehydrinsto high concentrations of other proteins, RNA, andcolloidal particles. In vivo, the total concentration ofproteins and RNA is in excess of 300 mg mL21
(Zimmerman and Trach, 1991). An intrinsic problemwith such experiments, however, is that the spectro-scopic handle of the dehydrin structure, in this casethe CD signal, would disappear against the muchlarger background of the cosolutes. As an alternative,we used the large polysaccharides Ficoll (70 kD) anddextran (40 and 70 kD). Interestingly, the presence ofthese crowding agents showed no appreciable effecton the dehydrin proteins. At 50% Ficoll or dextran, theCD spectra of Lti29 and Lti30 were virtually identicalto those in pure buffer, while the spectrum of Cor47showed an increase in the 200-nm region similar tothat in Glc and Suc (Fig. 6). Although the changes inthe lower wavelengths of the Cor47 spectrum aredifficult to rationalize structurally (Mouillon et al.,
Figure 5. Top, CD spectra of the glycerol titration of Cor47. The spectrashow from top at 220 nm, 0%, 10%, 25%, 50%, 60%, 75%, and 95%glycerol. Middle, CD difference spectra showing that the structuralinduction of Cor47 with glycerol occurs in two steps. Between 0% and75%, glycerol Cor47 displays a mix of b-sheet and a-helices, whileabove 75%, it displays mainly a-helices. Bottom, Changes in the CDsignal at 200 and 222 nm upon titration of Cor47 with glycerol,illustrating the transitions to mixed and pure a-helical structures,respectively.
Mimicking the Plant Cell Interior under Water Stress
Plant Physiol. Vol. 148, 2008 1929 www.plantphysiol.orgon April 11, 2019 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.
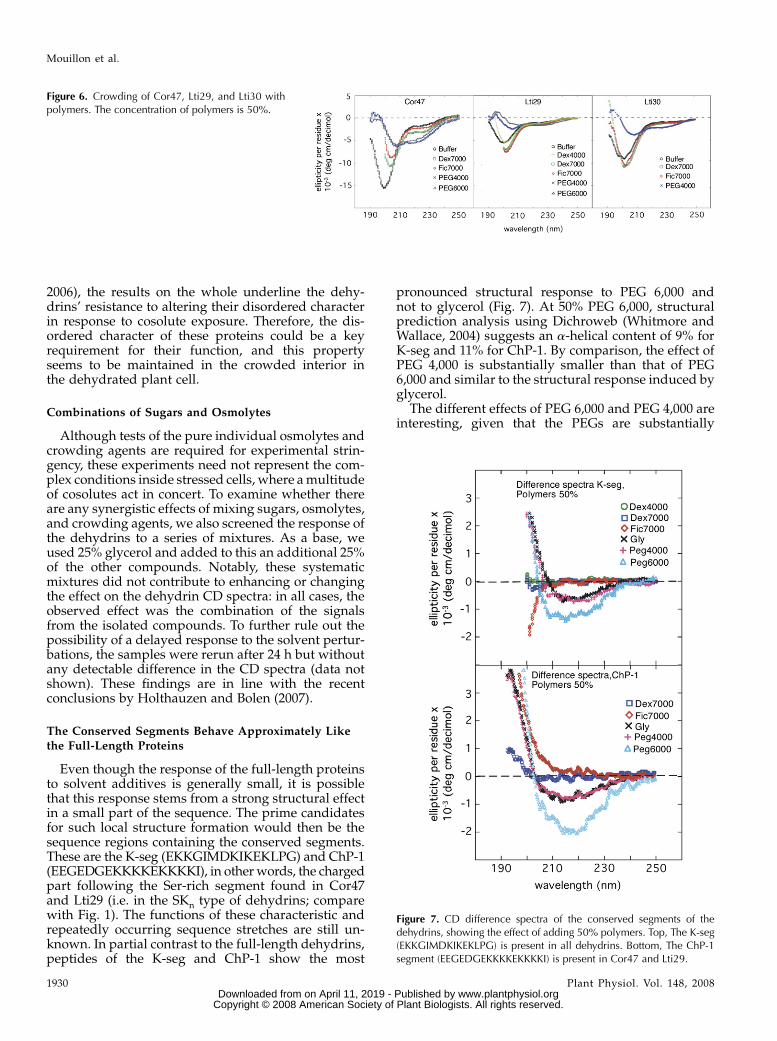
2006), the results on the whole underline the dehy-drins’ resistance to altering their disordered characterin response to cosolute exposure. Therefore, the dis-ordered character of these proteins could be a keyrequirement for their function, and this propertyseems to be maintained in the crowded interior inthe dehydrated plant cell.
Combinations of Sugars and Osmolytes
Although tests of the pure individual osmolytes andcrowding agents are required for experimental strin-gency, these experiments need not represent the com-plex conditions inside stressed cells, where amultitudeof cosolutes act in concert. To examine whether thereare any synergistic effects of mixing sugars, osmolytes,and crowding agents, we also screened the response ofthe dehydrins to a series of mixtures. As a base, weused 25% glycerol and added to this an additional 25%of the other compounds. Notably, these systematicmixtures did not contribute to enhancing or changingthe effect on the dehydrin CD spectra: in all cases, theobserved effect was the combination of the signalsfrom the isolated compounds. To further rule out thepossibility of a delayed response to the solvent pertur-bations, the samples were rerun after 24 h but withoutany detectable difference in the CD spectra (data notshown). These findings are in line with the recentconclusions by Holthauzen and Bolen (2007).
The Conserved Segments Behave Approximately Like
the Full-Length Proteins
Even though the response of the full-length proteinsto solvent additives is generally small, it is possiblethat this response stems from a strong structural effectin a small part of the sequence. The prime candidatesfor such local structure formation would then be thesequence regions containing the conserved segments.These are the K-seg (EKKGIMDKIKEKLPG) and ChP-1(EEGEDGEKKKKEKKKKI), in otherwords, the chargedpart following the Ser-rich segment found in Cor47and Lti29 (i.e. in the SKn type of dehydrins; comparewith Fig. 1). The functions of these characteristic andrepeatedly occurring sequence stretches are still un-known. In partial contrast to the full-length dehydrins,peptides of the K-seg and ChP-1 show the most
pronounced structural response to PEG 6,000 andnot to glycerol (Fig. 7). At 50% PEG 6,000, structuralprediction analysis using Dichroweb (Whitmore andWallace, 2004) suggests an a-helical content of 9% forK-seg and 11% for ChP-1. By comparison, the effect ofPEG 4,000 is substantially smaller than that of PEG6,000 and similar to the structural response induced byglycerol.
The different effects of PEG 6,000 and PEG 4,000 areinteresting, given that the PEGs are substantially
Figure 6. Crowding of Cor47, Lti29, and Lti30 withpolymers. The concentration of polymers is 50%.
Figure 7. CD difference spectra of the conserved segments of thedehydrins, showing the effect of adding 50% polymers. Top, The K-seg(EKKGIMDKIKEKLPG) is present in all dehydrins. Bottom, The ChP-1segment (EEGEDGEKKKKEKKKKI) is present in Cor47 and Lti29.
Mouillon et al.
1930 Plant Physiol. Vol. 148, 2008 www.plantphysiol.orgon April 11, 2019 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.
larger than the peptide fragments and the full-lengthdehydrins and would, accordingly, be expected toaffect these in an equivalent way. Ficoll and dextranare also considerably larger than either peptide butgive as low a response as with the full-length dehy-drins. Upon titration with glycerol, the K-seg and ChP-1both show an approximately linear decrease of the CDsignal at 222 nm. Therefore, K-seg and ChP-1 behavevery much like the full-length proteins Lti29 and Lti30and lack the characteristic late transition of Cor47 at95% glycerol (data not shown).
Phosphorylation Has Only a Marginal Effect on the
Dehydrin Structures
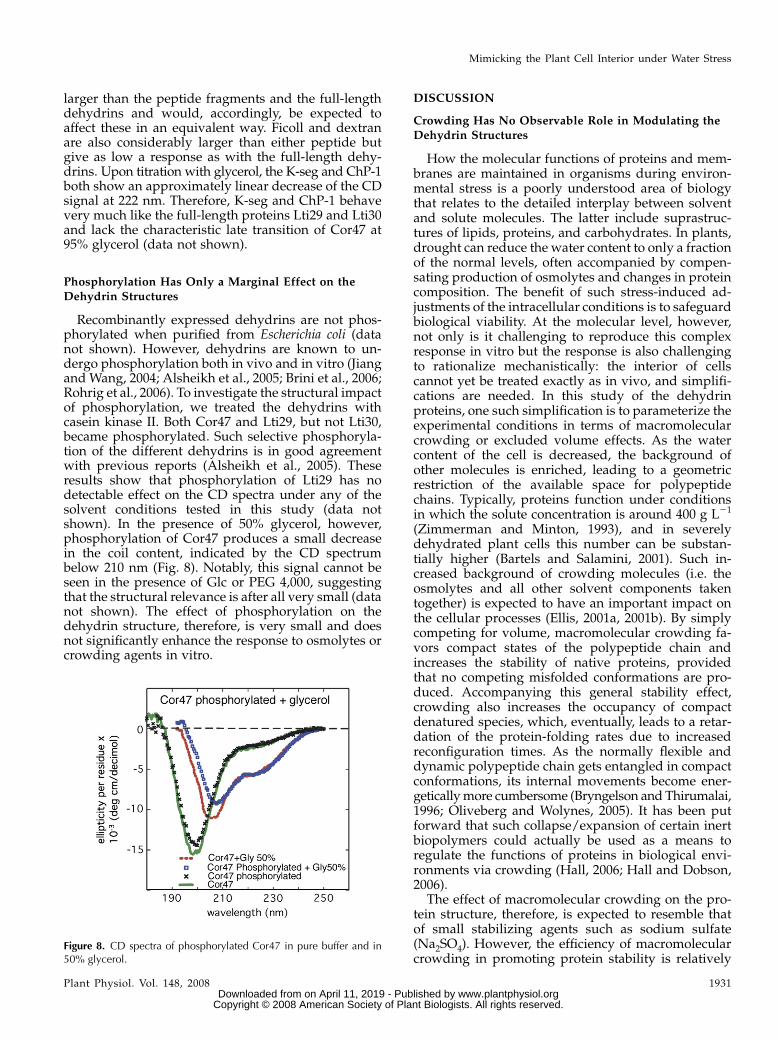
Recombinantly expressed dehydrins are not phos-phorylated when purified from Escherichia coli (datanot shown). However, dehydrins are known to un-dergo phosphorylation both in vivo and in vitro (Jiangand Wang, 2004; Alsheikh et al., 2005; Brini et al., 2006;Rohrig et al., 2006). To investigate the structural impactof phosphorylation, we treated the dehydrins withcasein kinase II. Both Cor47 and Lti29, but not Lti30,became phosphorylated. Such selective phosphoryla-tion of the different dehydrins is in good agreementwith previous reports (Alsheikh et al., 2005). Theseresults show that phosphorylation of Lti29 has nodetectable effect on the CD spectra under any of thesolvent conditions tested in this study (data notshown). In the presence of 50% glycerol, however,phosphorylation of Cor47 produces a small decreasein the coil content, indicated by the CD spectrumbelow 210 nm (Fig. 8). Notably, this signal cannot beseen in the presence of Glc or PEG 4,000, suggestingthat the structural relevance is after all very small (datanot shown). The effect of phosphorylation on thedehydrin structure, therefore, is very small and doesnot significantly enhance the response to osmolytes orcrowding agents in vitro.
DISCUSSION
Crowding Has No Observable Role in Modulating theDehydrin Structures
How the molecular functions of proteins and mem-branes are maintained in organisms during environ-mental stress is a poorly understood area of biologythat relates to the detailed interplay between solventand solute molecules. The latter include suprastruc-tures of lipids, proteins, and carbohydrates. In plants,drought can reduce the water content to only a fractionof the normal levels, often accompanied by compen-sating production of osmolytes and changes in proteincomposition. The benefit of such stress-induced ad-justments of the intracellular conditions is to safeguardbiological viability. At the molecular level, however,not only is it challenging to reproduce this complexresponse in vitro but the response is also challengingto rationalize mechanistically: the interior of cellscannot yet be treated exactly as in vivo, and simplifi-cations are needed. In this study of the dehydrinproteins, one such simplification is to parameterize theexperimental conditions in terms of macromolecularcrowding or excluded volume effects. As the watercontent of the cell is decreased, the background ofother molecules is enriched, leading to a geometricrestriction of the available space for polypeptidechains. Typically, proteins function under conditionsin which the solute concentration is around 400 g L21
(Zimmerman and Minton, 1993), and in severelydehydrated plant cells this number can be substan-tially higher (Bartels and Salamini, 2001). Such in-creased background of crowding molecules (i.e. theosmolytes and all other solvent components takentogether) is expected to have an important impact onthe cellular processes (Ellis, 2001a, 2001b). By simplycompeting for volume, macromolecular crowding fa-vors compact states of the polypeptide chain andincreases the stability of native proteins, providedthat no competing misfolded conformations are pro-duced. Accompanying this general stability effect,crowding also increases the occupancy of compactdenatured species, which, eventually, leads to a retar-dation of the protein-folding rates due to increasedreconfiguration times. As the normally flexible anddynamic polypeptide chain gets entangled in compactconformations, its internal movements become ener-geticallymore cumbersome (Bryngelson and Thirumalai,1996; Oliveberg and Wolynes, 2005). It has been putforward that such collapse/expansion of certain inertbiopolymers could actually be used as a means toregulate the functions of proteins in biological envi-ronments via crowding (Hall, 2006; Hall and Dobson,2006).
The effect of macromolecular crowding on the pro-tein structure, therefore, is expected to resemble thatof small stabilizing agents such as sodium sulfate(Na2SO4). However, the efficiency of macromolecularcrowding in promoting protein stability is relatively
Figure 8. CD spectra of phosphorylated Cor47 in pure buffer and in50% glycerol.
Mimicking the Plant Cell Interior under Water Stress
Plant Physiol. Vol. 148, 2008 1931 www.plantphysiol.orgon April 11, 2019 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

modest and is substantially smaller than that ofNa2SO4. The equilibrium constant for RNAse T1 wasonly shifted 7.5-fold toward the native state in 400 mgmL21 dextran 70 (Qu and Bolen, 2002), while thecorresponding effect of Na2SO4 on the protein L23 isseveral times larger (Hedberg and Oliveberg, 2004).Consistent with the earlier observation that Na2SO4has no appreciable impact on the dehydrin structures(Mouillon et al., 2006), we could not discern anycompelling effect upon crowding by the largebranched sugars Ficoll and dextran (Fig. 6). The onlyoutcome is a small change in the low-wavelengthregion of the CD spectra. This change could possiblyreflect a relaxation of PII helices toward more flexiblecoil conformations. By comparison, induction of aclassic secondary structure, such as a-helices orb-sheets, would lead to a reduction in the 200-nm signalwith a simultaneous increase in the minimum around220 nm (compare with Fig. 2). However, it is necessaryto consider also the size of the tested crowding agentin relation to that of the dehydrins. At a first approx-imation, a small crowding agent is expected to getcloser to the protein and, hence, to occupy volumemore efficiently compared with a large crowdingagent. Even so, the dehydrin structures are equallyindifferent to the smaller sugars Glc and Suc (Figs. 3and 4).
We could not distinguish any correspondence be-tween changes in CD spectra and the size of any of theadded solutes in this study. The disordered nature ofthe dehydrin polypeptide seems to persist also whenthe available space is geometrically restricted. A plau-sible explanation for this result is that the dehydrinsdo not easily adapt compact conformations, either inthe form of uniquely defined folded states, as has beenobserved for several other disordered proteins (Dawsonet al., 2003; Love et al., 2004; Fink, 2005), or in the formof unspecifically collapsed conformations that aresometimes observed in the denatured states of globu-lar proteins (Lattman et al., 1994; Otzen and Oliveberg,2002; Ittah et al., 2004). The behavior is in goodagreement with earlier reports on the disorderedc-Fos and p27Kip1 proteins, in which crowding withdifferently sized dextrans (9, 37.5, or 77 kD) andFicoll 70 at concentrations of up to 250 g L21 was notsufficient to induce an ordered structure (Flaughand Lumb, 2001). The dehydrins resemble also theParkinson-related a-synuclein that remains disor-dered in the crowded interior of E. coli cells and athigh concentrations of bovine serum albumin (McNultyet al., 2006b). In contrast, the transcription factor regu-lator FlgM partly gains structure under the correspond-ing conditions (Dedmon et al., 2002), and FlgM alsogains structure upon binding to its biological target(Daughdrill et al., 1997). These differences have beentaken to exemplify two classes of disordered proteins:those that are functionally tuned to remain disorderedin the crowded interior of the cell (Flaugh and Lumb,2001) and those that are not. According to the data inFigures 3 to 8, the dehydrins belong to the former
category. Taken together, the effect of macromolecularcrowding on disordered protein conformations seemsto be variable and needs to be tested in each separatecase. A brief summary of the crowding effects onother disordered proteins and denatured globularproteins is given in Supplemental Table S1. Anothereffect of water loss is increased ionic strength. Wedemonstrated earlier, however, that increased ionicstrength or elevated levels of metal ions has noappreciable impact on the dehydrin structure in vitro(Mouillon et al., 2006).
Preferential Protein Solute Exclusion Increases theSampling of the Local a-Helical Structure
The conformational transitions underlying the al-tered CD signals of Cor47 in the presence of glyceroland PEGs, therefore, are likely to have a molecularorigin different from pure crowding. From a functionalperspective, it is reasonable to examine whether thesemolecular factors are somehow coupled to changes inthe accessibility of water. For example, they may bemanifested in altered structures of the protein hydra-tion shells. However, such water-mediated effects areintrinsically difficult to mechanistically rationalize be-cause of the complexity of aqueous solutions. A sim-plified view is offered by weak interaction models.These models assume that the solute molecules, typ-ically osmolytes, are either accumulated or excludedfrom the protein-solvent interface. In the event that theosmolyte is preferentially attracted to the proteinsurface, this will favor extended conformations andpromote destabilization according to mass action(Schellman, 2002). Examples of such destabilizingosmolytes are the denaturants urea and guanidiumchloride. Conversely, if the osmolyte is preferentiallyexcluded from the protein surface, this will lead tominimization of the protein-solvent interface area andprotein stabilization (Bolen and Baskakov, 2001). Suchstabilizing, or protective, osmolytes are sugars, freeamino acids, and glycerol. The reason for the “affinitydifferences” between the osmolytes seems partly re-lated to the degree to which they can form favorablepolar interactions with the polypeptide backbone(Street et al., 2006). Computational (Street et al., 2006)and experimental (Qu and Bolen, 2002) analyses sug-gest that the order of stabilizing capacity of theosmolytes used in this study is Suc . sorbitol . Pro .glycerol. From this perspective, it is interesting that theosmolyte that most efficiently perturbed the CD spec-tra of the dehydrin Cor47 is glycerol (Fig. 5). The effectof 80% Suc, on the other hand, is still relatively modest.An explanation for this reversed order could lie in thesomewhat unusual amino acid composition of thedehydrin sequences.
The computational ranking of the osmolyte effect isobtained from backbone interactions alone (Streetet al., 2006), which is in good accord with the stabilityeffect on RNAse T1 (Qu and Bolen, 2002) with ahydrophobic side chain content typical of globular
Mouillon et al.
1932 Plant Physiol. Vol. 148, 2008 www.plantphysiol.orgon April 11, 2019 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

proteins (compare with pdf structure 1bu4 in the RCSBProtein Data Bank). In contrast, the dehydrin se-quences contain a relatively high proportion ofcharged and polar side chains, presenting a higherdegree of hydrogen-bonding opportunities with thesurrounding solution molecules. This peculiaritycould also explain why a-synuclein, in contrast tothe dehydrins, collapses upon addition of Glc (Moraret al., 2001): a-synuclein contains approximately 40%hydrophobic residues, while the dehydrins containjust 15% (Lti30) to 30% (Cor47 and Lti29).Even so, and regardless of the mechanistic origin of
the accentuated glycerol effect of the dehydrins, it isapparent from the CD data that the structural impactof adding osmolytes is only small at physiologicallyrelevant concentrations (Figs. 3 and 5). The same istrue for the fragments of their conserved segments(Fig. 7). The glycerol effect is not clearly manifestedunless the glycerol content is increased above 30%, alevel that is likely to greatly exceed the physiologicalconcentrations. By comparison, the highest reportedlevel of Suc in vivo is 40% of the dry weight, and this ina resurrection plant that more or less tolerates com-plete desiccation (Ingram and Bartels, 1996). More-over, the structural transition observed at extremeconcentrations of glycerol corresponds mainly to anincreased sampling of the a-helical structure, which isin good accord with the intrinsic a-helical propensityof the different dehydrin proteins (Fig. 1). The largesteffect was observed for Cor47, followed by Lti29 andLti30, which precisely matched the gain of thea-helical structure in TFE titrations (Mouillon et al.,2006). Consistently, a similar structural response wasobserved upon the addition of PEG 4,000 and PEG6,000. These polymers are well-known protein stabi-lizers (Kornblatt and Kornblatt, 2002; Kozer andSchreiber, 2004; Ren et al., 2006) that have also beenobserved to unspecifically collapse the denaturedstates of globular proteins (Silow and Oliveberg,2003). On this basis, we conclude that the exertion ofpressure to minimize the solvent-accessible surfacearea of the dehydrin chains has no appreciable effectson their tertiary structures. They generally remaindisordered. The effect on the local a-helical contentmerely follows the intrinsic a-helical propensity of thepolypeptide chains. The latter, however, is more likelyto reflect the generic properties of the polypeptidechain than specific function traits.
Possible Interactions between Dehydrins and
Lipid Membranes
A frequently proposed function of dehydrins andother LEA proteins is as membrane stabilizers. Inessence, the dehydrins would interact with mem-branes to modulate their topology and phase transi-tions under stress, and in this process the dehydrinscould also gain ordered structure. This idea is supportedby several findings of interactions between lipid ves-icles and dehydrins and other LEA proteins (Soulages
et al., 2002, 2003; Koag et al., 2003; Tolleter et al., 2007;Kovacs et al., 2008). In some cases, as with the maize(Zea mays) dehydrin DHN1, the membrane interactionleads to conformational changes in the protein’s sec-ondary structure (i.e. an increase in the a-helicalcontent; Koag et al., 2003). However, it is not clear towhat extent these effects can be transferred to othermembers of the protein family, as, in contrast, theinteraction between Lti29 and negatively chargedlipids did not result in any conformational changes(Kovacs et al., 2008). Also, the presence of lipids failedto induce any structural effect on the soybean (Glycinemax) LEA protein GmD-19 and dehydrin GmDHN1(Soulages et al., 2002, 2003). Although the involvementof the dehydrins in safeguarding membrane structureduring stress is indeed an attractive possibility, itsgenerality remains to be established.
Phosphorylation Does Not Generate Any SignificantStructural Changes of Dehydrins
Phosphorylation is a posttranslational modificationthat is essential for the regulation of cellular func-tions. It was recently shown that most phosphoryla-tion sites, 86%, are within disordered regions ofproteins (Collins et al., 2008). Even so, there are fewstudies on the effect of phosphorylation on the struc-ture of disordered proteins. Of particular interest forthis study is the fact that phosphorylation of thedehydrins occurs in vivo and has been reported tomodulate the coordination of calcium ions (Jiang andWang, 2004; Alsheikh et al., 2005; Brini et al., 2006;Rohrig et al., 2006). In this study, only Cor47 andLti29 could be phosphorylated by casein kinase II.The result of this phosphorylation, however, had nosignificant bearing on the structural behavior ofCor47 and Lti29. Recent NMR studies of a-synucleinhave shown that phosphorylation at Ser-129 by caseinkinase II increases the disordered character of theprotein. It was further shown that this structuralalteration increased the protein’s tendency to self-associate (Sasakawa et al., 2007). As seen in Figure 8,the structure of Cor47 is left unaltered upon phosphor-ylation and also shows no tendency to aggregate.Crowding by glycerol makes no difference. Eventhough Cor47 and Lti29 seem to be structurally un-changed, it is important to note that phosphorylationcan still play a key role as a regulatory factor for theinteraction with other cellular targets.
CONCLUSION
From a physiological standpoint, the separate testsof crowding agents and osmolytes constitute a fairlycrude representation of the intracellular conditionsin which spatial confinement and the presence ofcomplex mixtures of background molecules act inconcert on the dehydrin structures. However, wefailed to observe any synergistic structural effects in
Mimicking the Plant Cell Interior under Water Stress
Plant Physiol. Vol. 148, 2008 1933 www.plantphysiol.orgon April 11, 2019 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

mixtures of glycerol and the other solutes used in thisstudy, lending further support to the idea that thedehydrin structures are generally insensitive tochanging solvent conditions. Based on previous re-sults, the structural insensitivity also includeschanges of ionic strength and levels of metal ions(Mouillon et al., 2006). This suggests that the dehy-drin sequences are evolved to remain flexible alsounder severe stress conditions and that this propertyis an integral requirement of their physiologicalfunction. Possibly, the high content of charged andpolar residues of the dehydrins effectively preventsthe structural collapse due to the accompanyingdesolvation penalties of ionic moieties and hydrogenbond partners. Such transitions into compact glass-like species are otherwise characteristic of randompolymers (Bryngelson and Thirumalai, 1996) and tosome extent also of denatured states of globularproteins (Silow and Oliveberg, 2003). In both in-stances, the affected sequences generally containhigher fractions of hydrophobic side chains thanobserved for the dehydrins.
Additional indications that the dehydrins are tunedto remain flexible under in vivo conditions come fromtheir high solubility and resistance to precipitation(Mouillon et al., 2006). As a contrasting example,a-synuclein readily assembles in ordered fibrillar ag-gregates at elevated protein concentrations or uponthe addition of PEG/osmolytes (Goers et al., 2003;Fink, 2006). In discussions about the biological func-tion of the dehydrins, it has been pointed out that theirunfolded nature would effectively maximize the coor-dination of water. According to NMR experiments,this water is more tightly bound than in other disor-dered proteins (Bokor et al., 2005). Some of the waterseems tied up in bridging interactions within PIIhelices (Mouillon et al., 2006). Upon removal of thecoordinated water by evaporation, other proteins inthe LEA family have been observed to undergo struc-tural rearrangements (Wolkers et al., 2001; Goyal et al.,2003, 2005). For example, the LEA protein AavLea1increases its content of a-helical structure (Goyal et al.,2003). Could the dehydrins then act as water reservoirsthat expel coordinated water to the surrounding me-dium (Boudet et al., 2006)? Considering the findingthat the in vivo concentration of dehydrins does notseem to exceed 2% to 4% of cytoplasmic proteins,however, the benefit of such water expulsion is likelyto be marginal (Roberts et al., 1993; Ceccardi et al.,1994; Close, 1996), at least in moderate drought. Morelikely, the molecular action of the dehydrins is morespecific. One possibility is that the dehydrins providestructural support, either spatially by cross-linkingwith other macromolecular constituents (Abu-Abiedet al., 2006) or laterally by adhering to membranesurfaces (Tolleter et al., 2007). Desiccation of the cellwill not only alter the molecular composition of theaqueous compartments of the cell but also distort itsphysical dimensions, including the shape and prox-imity of membrane structures.
It is easy to envisage that such global alterationscould have a pronounced effect on the colloidal pro-cesses, for example by promoting phase transitions,topological frustration, and altered dynamics of mem-brane fusion and budding. The role of the dehydrinsmay be to intervene with these processes by filling out,and perhaps topologically organizing, crevices inbulging membranes and thereby safeguarding theirstructural integrity. Alternatively, the dehydrins couldsupport macromolecular structures of proteins, carbo-hydrates, or nucleotides. In any case, the elements ofrecognition with the cellular target are likely to be theconserved segments. The remarkable capacity of thedehydrins to remain coil like suggests that the highflexibility of the sequence regions connecting thesesegments is an important part of their function (Fig. 1):the conserved segments are organized as “hooks on astring” (Mouillon et al., 2006). Notably, this arrange-ment not only restricts the spatial location of theconserved elements but also increases their local con-centration.
MATERIALS AND METHODS
Expression and Heat Fractionation
Expression, purification, and identification of the recombinant Arabidopsis
(Arabidopsis thaliana) dehydrin proteins were as described previously (Svensson
et al., 2000). Glycerol stocks of the different Escherichia coli strains were made,
and 200 mL was spread on Luria agar plates with 150 mg of ampicillin. The
plates were kept at 37�C and grown overnight. The cells were suspended and
added to 1 L of Luria-Bertani medium containing 50 mg mL21 ampicillin.
Expression was induced at an optical density of 0.3 by adding isopropyl b-D-
thiogalactopyranoside to a final concentration of 1 mM. The suspensions were
kept at 37�C until grown to an optical density of 0.8 to 1.0 (i.e. 2–4 h). Cells
were harvested by centrifugation at 4,000g for 45 min, and the pellet was
stored at –20�C. The thawed cells from 1-L cultures were resuspended in 25
mL of 20 mM Na2HPO4, pH 7.2, and 150 mM NaCl. To this was added 1 mM
(final concentration) phenylmethylsulfonyl fluoride and 0.1 mg mL21 lyso-
syme, and the suspension was left for 30 min on ice. Lysated cells were
sonicated six times for 15 s each and centrifuged at 9,000g for 30 min. The
supernatants were placed in a water bath at 70�C for 20 min. To precipitate
heat-denatured proteins, the sample was centrifuged for 30 min at 9,000g, and
the supernatant was stored at –80�C.
Purification of Dehydrins
Purification of dehydrins by immobilized metal ion affinity chromatogra-
phy and the following step by ion-exchange chromatography was according
to Svensson et al. (2000). The supernatants from heat precipitation were
diluted 1:2 with 20 mM NaHPO4, pH 7.2, 1.85 M NaCl, and 1 mM phenyl-
methylsulfonyl fluoride. The samples were loaded on a 5-mL HiTrap IDA-
Sepharose column (Pharmacia) connected to a FPLC system (Pharmacia) and
chargedwith 2mL of 3mgmL21 CuSO4. Absorbancewas read at 280 nm. Before
applying the sample, five volumes of 20 mM Na2HPO4, pH 7.2, and 1.0 M NaCl
were used to equilibrate the column. The same buffer (40 volumes) was used
to wash off unbound sample from the column. Fractions of 5 mL were
collected for analysis during the whole run. Elution was performed with 2 M
NH4Cl in 20 mM Na2HPO4, pH 7.2, and 1.0 M NaCl in one step. The column
was then equilibrated with 10 volumes of 20 mM Na2HPO4, pH 7.2, followed
by elution of the copper with 10 mM EDTA in 20 mM Na2HPO4, pH 7.2.
Precipitation of protein was done with 80% (NH4)2SO4, and proteins were
collected by centrifugation at 18,000g for 30 min. The different dehydrins were
resuspended in 2 mL of the following buffers: Lti29 and Cor47 in 20 mM
BisTris, pH 6.0, Rab18 in 20 mM Tris-HCl, pH 8.0, and Lti30 in 50 mM Gly, pH
9.0. The dehydrins were desalted on a PD-10 column (Pharmacia) by the
Mouillon et al.
1934 Plant Physiol. Vol. 148, 2008 www.plantphysiol.orgon April 11, 2019 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

resuspension buffers. The achieved 3-mL fractions were put on an anion-
exchange column (1-mL Mono QHR 5/5; Pharmacia) in the case of Cor47 and
Lti29 and on a cation-exchange column (1-mL Mono S HR 5/5; Pharmacia)
connected to an FPLC system in the case of Lti30, and absorbance was read at
280 nm. The columns were equilibrated with the resuspension buffers and
elution by a NaCl gradient from 0 to 0.5 M in the respective buffer over 30
volumes. Fractions of 0.5 mL were collected during the runs for analysis. The
purity was tested by SDS-PAGE, and without exception the purified material
was run as untreated dehydrins from crude cell extracts (data not shown). An
extensive identification of the recombinant dehydrins has been published by
Svensson et al. (2000).
Analysis by CD
CDmeasurements were carried out using a JASCO J-810 spectropolarimeter,
and the results are presented as mean ellipticity per residue. The scan rate was
20 nm min21 at a band pass of 0.2 nm, with 20 mdeg sensitivity. All samples
were mixed 1 h prior to the CD analysis and centrifuged at 12,000g for 2 min
before filling the cuvettes. Protein concentration was 1 mg mL21 in 20 mM MES
(0.2-mm cuvette). Unless stated otherwise, all runs were performed at 25�C.
Preparation of Samples
Unless otherwise stated, concentrations are given as percentage referring
to grams per liter. Dextrans (average molecular mass of 40 or 70 kD; Sigma),
Ficolls (average molecular mass of 70 kD; Sigma), and PEG (averagemolecular
mass of 4,000 or 6,000 D; Sigma) were used in concentrations of up to 500 to
800 g L21. All osmolytes, sugars, and crowding agents were titrated in steps of
10% to 25%. Each titration series was repeated at least twice. The exception
was Lti30, for which the titration steps were 15% to 25% and titration ended at
50% because of the low amount of high-purity Lti30 protein.
Phosphorylation of Dehydrins
The dehydrins were phosphorylated by 5.4 units mL21 casein kinase II
(Sigma) in 1 mM ATP, 20 mM Tris-HCl, 50 mM KCl, and 10 mM MgCl2, pH 7.5,
for 10 h. The phosphorylation status of the dehydrins was tested by Pro-Q
Diamond Phosphoprotein Gel stain (Invitrogen) on 15% Tris-HCl gels (Bio-
Rad), and the background levels of protein were measured by SYPRO Ruby
Gel stain (Invitrogen). Stained gels were visualized with an FLA-3000G (Fuji
Photo Film) and a UV transilluminator. The phosphorylation status was
measured on three different batches of dehydrins, with identical results. For
the structural analysis, the casein kinase II was separated from the dehydrins
by precipitation at 70�C, followed by centrifugation. The effect of solvent
additives on the phosphorylated dehydrins was measured as described above.
Sequence data from this article can be found in the GenBank/EMBL data
libraries under accession numbers P42758, P42759, and CAA48178.
Supplemental Data
The following materials are available in the online version of this article.
Supplemental Figure S1. Phosphorylation of Lti29 and Cor47 by casein
kinase II.
Supplemental Figure S2. CD spectra of phosphorylated Lti29 in the
presence of 50% glycerol.
Supplemental Table S1. Effects of various crowding agents on intrinsically
disordered proteins and denatured globular proteins.
ACKNOWLEDGMENT
We are most grateful to Professor Mikael Oliveberg (Stockholm Univer-
sity) for the data regarding CI2 and for helpful discussions.
Received July 18, 2008; accepted October 1, 2008; published October 10, 2008.
LITERATURE CITED
Abu-Abied M, Golomb L, Belausov E, Huang S, Geiger B, Kam Z, Staiger
CJ, Sadot E (2006) Identification of plant cytoskeleton-interacting pro-
teins by screening for actin stress fiber association in mammalian
fibroblasts. Plant J 48: 367–379
Al-Habori M (2001) Macromolecular crowding and its role as intracellular
signalling of cell volume regulation. Int J Biochem Cell Biol 33: 844–864
Alsheikh MK, Svensson JT, Randall SK (2005) Phosphorylation regulated
ion-binding is a property shared by the acidic subclass dehydrins. Plant
Cell Environ 28: 1114–1122
Bartels D, Salamini F (2001) Desiccation tolerance in the resurrection plant
Craterostigma plantagineum: a contribution to the study of drought
tolerance at the molecular level. Plant Physiol 127: 1346–1353
Bokor M, Csizmok V, Kovacs D, Banki P, Friedrich P, Tompa P, Tompa K
(2005) NMR relaxation studies on the hydrate layer of intrinsically
unstructured proteins. Biophys J 88: 2030–2037
Bolen DW, Baskakov IV (2001) The osmophobic effect: natural selection of
a thermodynamic force in protein folding. J Mol Biol 310: 955–963
Boudet J, Buitink J, Hoekstra FA, Rogniaux H, Larre C, Satour P, Leprince
O (2006) Comparative analysis of the heat stable proteome of radicles of
Medicago truncatula seeds during germination identifies late embryo-
genesis abundant proteins associated with desiccation tolerance. Plant
Physiol 140 1418–1436
Brini F, Hanin M, Lumbreras V, Irar S, Pages M, Masmoudi K (2006)
Functional characterization of DHN-5, a dehydrin showing a differen-
tial phosphorylation pattern in two Tunisian durum wheat (Triticum
durum Desf.) varieties with marked difference in salt and drought
tolerance. Plant Sci 172: 20–28
Bryant JE, Lecomte JT, Lee AL, Young GB, Pielak GJ (2005) Protein
dynamics in living cells. Biochemistry 44: 9275–9279
Bryngelson JD, Thirumalai D (1996) Internal constraints induce localiza-
tion in an isolated polymer molecule. Phys Rev Lett 76: 542–545
Ceccardi TL, Meyer NC, Close TJ (1994) Purification of a maize dehydrin.
Protein Expr Purif 5: 266–269
Chen TH, Murata N (2002) Enhancement of tolerance of abiotic stress by
metabolic engineering of betaines and other compatible solutes. Curr
Opin Plant Biol 5: 250–257
Cheung MS, Thirumalai D (2007) Effects of crowding and confinement on
the structures of the transition state ensemble in proteins. J Phys Chem B
111: 8250–8257
Close TJ (1996) Dehydrins: emergence of a biochemical role of a family of
plant dehydration proteins. Physiol Plant 97: 795–803
Close TJ (1997) Dehydrins: a commonality in the response of plants to
dehydration and low temperature. Physiol Plant 100: 291–296
Collins MO, Yu L, Campuzano I, Grant SG, Choudhary JS (2008)
Phosphoproteomic analysis of the mouse brain cytosol reveals a pre-
dominance of protein phosphorylation in regions of intrinsic sequence
disorder. Mol Cell Proteomics 7: 1331–1348
Daughdrill GW, Chadsey MS, Karlinsey JE, Hughes KT, Dahlquist FW
(1997) The C-terminal half of the anti-sigma factor, FlgM, becomes
structured when bound to its target, sigma 28. Nat Struct Biol 4: 285–291
Dawson R, Muller L, Dehner A, Klein C, Kessler H, Buchner J (2003)
The N-terminal domain of p53 is natively unfolded. J Mol Biol 332:
1131–1141
Dedmon MM, Patel CN, Young GB, Pielak GJ (2002) FlgM gains structure
in living cells. Proc Natl Acad Sci USA 99: 12681–12684
Dunker AK, Brown CJ, Lawson JD, Iakoucheva LM, Obradovic Z (2002)
Intrinsic disorder and protein function. Biochemistry 41: 6573–6582
Ellis RJ (2001a) Macromolecular crowding: an important but neglected
aspect of the intracellular environment. Curr Opin Struct Biol 11:
114–119
Ellis RJ (2001b) Macromolecular crowding: obvious but underappreciated.
Trends Biochem Sci 26: 597–604
Fink AL (2005) Natively unfolded proteins. Curr Opin Struct Biol 15: 35–41
Fink AL (2006) The aggregation and fibrillation of alpha-synuclein. Acc
Chem Res 39: 628–634
Flaugh SL, Lumb KJ (2001) Effects of macromolecular crowding on the
intrinsically disordered proteins c-Fos and p27(Kip1). Biomacromole-
cules 2: 538–540
Foord RL, Leatherbarrow RJ (1998) Effect of osmolytes on the exchange
rates of backbone amide protons in proteins. Biochemistry 37: 2969–2978
Garay-Arroyo A, Colmenero-Flores JM, Garciarrubio A, Covarrubias AA
(2000) Highly hydrophilic proteins in prokaryotes and eukaryotes are
common during conditions of water deficit. J Biol Chem 275: 5668–5674
Goers J, Uversky VN, Fink AL (2003) Polycation-induced oligomerization
Mimicking the Plant Cell Interior under Water Stress
Plant Physiol. Vol. 148, 2008 1935 www.plantphysiol.orgon April 11, 2019 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

and accelerated fibrillation of human alpha-synuclein in vitro. Protein
Sci 12: 702–707
Goyal K, Pinelli C, Maslen SL, Rastogi RK, Stephens E, Tunnacliffe A
(2005) Dehydration-regulated processing of late embryogenesis abundant
protein in a desiccation-tolerant nematode. FEBS Lett 579: 4093–4098
Goyal K, Tisi L, Basran A, Browne J, Burnell A, Zurdo J, Tunnacliffe A
(2003) Transition from natively unfolded to folded state induced by
desiccation in an anhydrobiotic nematode protein. J Biol Chem 278:
12977–84
Hall D (2006) Protein self-association in the cell: a mechanism for fine
tuning the level of macromolecular crowding? Eur Biophys J 35: 276–280
Hall D, Dobson CM (2006) Expanding to fill the gap: a possible role for
inert biopolymers in regulating the extent of the ‘macromolecular
crowding’ effect. FEBS Lett 580: 2584–2590
Hasegawa PM, Bressan RA, Zhu JK, Bohnert HJ (2000) Plant cellular and
molecular responses to high salinity. Annu Rev Plant Physiol Plant Mol
Biol 51: 463–499
Hedberg L, Oliveberg M (2004) Scattered Hammond plots reveal second
level of site-specific information in protein folding: phi# (beta++). ProcNatl Acad Sci USA 101: 7606–7611
Hoekstra FA, Golovina EA, Buitink J (2001a) Mechanisms of plant des-
iccation tolerance. Trends Plant Sci 6: 431–438
Hoekstra FA, Golovina EA, Tetteroo FA, Wolkers WF (2001b) Induction of
desiccation tolerance in plant somatic embryos: how exclusive is the
protective role of sugars? Cryobiology 43: 140–150
Holthauzen LM, Bolen DW (2007) Mixed osmolytes: the degree to which
one osmolyte affects the protein stabilizing ability of another. Protein Sci
16: 293–298
Ingram J, Bartels D (1996) The molecular basis of dehydration tolerance in
plants. Annu Rev Plant Physiol Plant Mol Biol 47: 377–403
Ittah V, Kahana E, Amir D, Haas E (2004) Applications of time-resolved
resonance energy transfer measurements in studies of the molecular
crowding effect. J Mol Recognit 17: 448–455
Jiang X, Wang Y (2004) Beta-elimination coupled with tandem mass
spectrometry for the identification of in vivo and in vitro phosphory-
lation sites in maize dehydrin DHN1 protein. Biochemistry 43: 15567–
15576
Klimov DK, Newfield D, Thirumalai D (2002) Simulations of beta-hairpin
folding confined to spherical pores using distributed computing. Proc
Natl Acad Sci USA 99: 8019–8024
Koag MC, Fenton RD, Wilkens S, Close TJ (2003) The binding of maize
DHN1 to lipid vesicles: gain of structure and lipid specificity. Plant
Physiol 131: 309–316
Kornblatt JA, Kornblatt MJ (2002) The effects of osmotic and hydrostatic
pressures on macromolecular systems. Biochim Biophys Acta 1595:
30–47
Kovacs D, Kalmar E, Torok Z, Tompa P (2008) Chaperone activity of ERD10
and ERD14, two disordered stress-related plant proteins. Plant Physiol
147: 381–390
Kozer N, Schreiber G (2004) Effect of crowding on protein-protein asso-
ciation rates: fundamental differences between low and high mass
crowding agents. J Mol Biol 336: 763–774
Lattman EE, Fiebig KM, Dill KA (1994) Modeling compact denatured
states of proteins. Biochemistry 33: 6158–6166
Lisse T, Bartels D, Kalbitzer HR, Jaenicke R (1996) The recombinant
dehydrin-like desiccation stress protein from the resurrection plant
Craterostigma plantagineum displays no defined three-dimensional
structure in its native state. Biol Chem 377: 555–561
Love JJ, Li X, Chung J, Dyson HJ, Wright PE (2004) The LEF-1 high-
mobility group domain undergoes a disorder-to-order transition upon
formation of a complex with cognate DNA. Biochemistry 43: 8725–8734
Luo P, Baldwin RL (1997) Mechanism of helix induction by trifluoroetha-
nol: a framework for extrapolating the helix-forming properties of
peptides from trifluoroethanol/water mixtures back to water. Biochem-
istry 36: 8413–8421
McNulty BC, Tripathy A, Young GB, Charlton LM, Orans J, Pielak GJ
(2006a) Temperature-induced reversible conformational change in the
first 100 residues of alpha-synuclein. Protein Sci 15: 602–608
McNulty BC, Young GB, Pielak GJ (2006b) Macromolecular crowding in
the Escherichia coli periplasm maintains alpha-synuclein disorder.
J Mol Biol 355: 893–897
Minton AP (2005a) Influence of macromolecular crowding upon the
stability and state of association of proteins: predictions and observa-
tions. J Pharm Sci 94: 1668–1675
Minton AP (2005b) Models for excluded volume interaction between an
unfolded protein and rigid macromolecular cosolutes: macromolecular
crowding and protein stability revisited. Biophys J 88: 971–985
Minton AP (2006) Macromolecular crowding. Curr Biol 16: R269–271
Morar AS, Olteanu A, Young GB, Pielak GJ (2001) Solvent-induced
collapse of alpha-synuclein and acid-denatured cytochrome c. Protein
Sci 10: 2195–2199
Mouillon JM, Gustafsson P, Harryson P (2006) Structural investigation of
disordered stress proteins: comparison of full-length dehydrins with
isolated peptides of their conserved segments. Plant Physiol 141: 638–650
Oliveberg M, Wolynes PG (2005) The experimental survey of protein-
folding energy landscapes. Q Rev Biophys 38: 245–288
Otzen DE, Oliveberg M (1999) Salt-induced detour through compact
regions of the protein folding landscape. Proc Natl Acad Sci USA 96:
11746–11751
Otzen DE, Oliveberg M (2002) Conformational plasticity in folding of the
split beta-alpha-beta protein S6: evidence for burst-phase disruption of
the native state. J Mol Biol 317: 613–627
Qu Y, Bolen CL, Bolen DW (1998) Osmolyte-driven contraction of a
random coil protein. Proc Natl Acad Sci USA 95: 9268–9273
Qu Y, Bolen DW (2002) Efficacy of macromolecular crowding in forcing
proteins to fold. Biophys Chem 101–102: 155–165
Ren X, Yang Z, Kuang T (2006) Solvent-induced changes in photochemical
activity and conformation of photosystem I particles by glycerol. Biol
Chem 387: 23–29
Roberts JK, DeSimone NA, Lingle WL, Dure L III (1993) Cellular con-
centrations and uniformity of cell-type accumulation of two lea proteins
in cotton embryos. Plant Cell 5: 769–780
Rohrig H, Schmidt J, Colby T, Brautigam A, Hufnagel P, Bartels D (2006)
Desiccation of the resurrection plant Craterostigma plantagineum in-
duces dynamic changes in protein phosphorylation. Plant Cell Environ
29: 1606–1617
Sasakawa H, Sakata E, Yamaguchi Y, Masuda M, Mori T, Kurimoto E,
Iguchi T, Hisanaga S, Iwatsubo T, Hasegawa M, et al (2007) Ultra-high
field NMR studies of antibody binding and site-specific phosphoryla-
tion of alpha-synuclein. Biochem Biophys Res Commun 363: 795–799
Schellman JA (2002) Fifty years of solvent denaturation. Biophys Chem 96:
91–101
Selenko P, Serber Z, Gadea B, Ruderman J, Wagner G (2006) Quantitative
NMR analysis of the protein G B1 domain in Xenopus laevis egg extracts
and intact oocytes. Proc Natl Acad Sci USA 103: 11904–11909
Silow M, Oliveberg M (2003) High concentrations of viscogens decrease
the protein folding rate constant by prematurely collapsing the coil. J
Mol Biol 326: 263–271
Soulages JL, Kim K, Arrese EL, Walters C, Cushman JC (2003) Confor-
mation of a group 2 late embryogenesis abundant protein from soybean:
evidence of poly (L-proline)-type II structure. Plant Physiol 131: 963–975
Soulages JL, Kim K, Walters C, Cushman JC (2002) Temperature-induced
extended helix/random coil transitions in a group 1 late embryogenesis-
abundant protein from soybean. Plant Physiol 128: 822–832
Street TO, Bolen DW, Rose GD (2006) A molecular mechanism for
osmolyte-induced protein stability. Proc Natl Acad Sci USA 103:
13997–14002
Svensson J, Palva ET, Welin B (2000) Purification of recombinant Arabi-
dopsis thaliana dehydrins by metal ion affinity chromatography. Protein
Expr Purif 20: 169–178
Tolleter D, Jaquinod M, Mangavel C, Passirani C, Saulnier P, Manon S,
Teyssier E, Payet N, Avelange-Macherel MH, Macherel D (2007)
Structure and function of a mitochondrial late embryogenesis abundant
protein are revealed by desiccation. Plant Cell 19: 1580–1589
Tompa P, Csermely P (2004) The role of structural disorder in the function
of RNA and protein chaperones. FASEB J 18: 1169–1175
Tornroth-Horsefield S, Wang Y, Hedfalk K, Johanson U, Karlsson M,
Tajkhorshid E, Neutze R, Kjellbom P (2006) Structural mechanism of
plant aquaporin gating. Nature 439: 688–694
Uversky VN, Oldfield CJ, Dunker AK (2005) Showing your ID: intrinsic
disorder as an ID for recognition, regulation and cell signaling. J Mol
Recognit 18: 343–384
Venkatesu P, Lee MJ, Lin HM (2007) Thermodynamic characterization of
the osmolyte effect on protein stability and the effect of GdnHCl on the
protein denatured state. J Phys Chem B 111: 9045–9056
Mouillon et al.
1936 Plant Physiol. Vol. 148, 2008 www.plantphysiol.orgon April 11, 2019 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

Wang W, Vinocur B, Altman A (2003) Plant responses to drought, salinity
and extreme temperatures: towards genetic engineering for stress tol-
erance. Planta 218: 1–14
Whitmore L, Wallace BA (2004) DICHROWEB, an online server for protein
secondary structure analyses from circular dichroism spectroscopic
data. Nucleic Acids Res 32: W668–W673
Wolkers WF, McCready S, Brandt WF, Lindsey GG, Hoekstra FA (2001)
Isolation and characterization of a D-7 LEA protein from pollen that
stabilizes glasses in vitro. Biochim Biophys Acta 1544: 196–206
Yoshiba Y, Kiyosue T, Nakashima K, Yamaguchi-Shinozaki K, Shinozaki
K (1997) Regulation of levels of proline as an osmolyte in plants under
water stress. Plant Cell Physiol 38: 1095–1102
Zimmerman SB, Minton AP (1993) Macromolecular crowding: biochem-
ical, biophysical, and physiological consequences. Annu Rev Biophys
Biomol Struct 22: 27–65
Zimmerman SB, Trach SO (1991) Estimation of macromolecule concentra-
tions and excluded volume effects for the cytoplasm of Escherichia coli.
J Mol Biol 222: 599–620
Mimicking the Plant Cell Interior under Water Stress
Plant Physiol. Vol. 148, 2008 1937 www.plantphysiol.orgon April 11, 2019 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.





























