Embed Size (px)
Citation preview

Met induces mammary tumors with diversehistologies and is associated with pooroutcome and human basal breast cancerMarisa G. Ponzoa,b,1, Robert Lesurfa,c,1, Stephanie Petkiewicza,b, Frances P. O’Malleyd,e, Dushanthi Pinnaduwagef,Irene L. Andrulisg,h, Shelley B. Bullf,i, Naila Chughtaia, Dongmei Zuoa, Margarita Souleimanovaa, David Germaina,Atilla Omerogluj, Robert D. Cardiffk, Michael Halletta,c, and Morag Parka,b,l,2
aGoodman Cancer Centre and bDepartment of Experimental Medicine, McGill University, Montreal, QC, Canada H3A 1A3; cMcGill Centre for Bioinformaticsand jDepartment of Pathology, McGill University, Montreal, QC, Canada H3A 2B4; dDepartment of Laboratory Medicine and Pathobiology, University ofToronto, Toronto, ON, Canada M5G 1L5; eDepartment of Pathology and Laboratory Medicine and gFred A. Litwin Centre for Cancer Genetics, SamuelLunenfeld Research Institute, Mount Sinai Hospital, Toronto, ON, Canada M5G 1X5; fProsserman Centre for Health Research, Samuel Lunenfeld ResearchInstitute, Mount Sinai Hospital, Toronto, ON, Canada M5T 3L9; hDepartment of Molecular Genetics, University of Toronto, Toronto, ON, Canada M5S 1A8;iDalla Lana School of Public Health, University of Toronto, Toronto, ON, M5T 3M7, Canada; kCentre for Comparative Medicine, University of California,Davis, CA 95616; and lDepartments of Oncology and Biochemistry, McGill University, Montreal, QC, Canada H2W 1S6
Edited by Joan S. Brugge, Harvard Medical School, Boston, MA, and approved March 2, 2009 (received for review October 20, 2008)
Elevated MET receptor tyrosine kinase correlates with poor out-come in breast cancer, yet the reasons for this are poorly under-stood. We thus generated a transgenic mouse model targetingexpression of an oncogenic Met receptor (Metmt) to the mammaryepithelium. We show that Metmt induces mammary tumors withmultiple phenotypes. These reflect tumor subtypes with geneexpression and immunostaining profiles sharing similarities tohuman basal and luminal breast cancers. Within the basal subtype,Metmt induces tumors with signatures of WNT and epithelial tomesenchymal transition (EMT). Among human breast cancers, METis primarily elevated in basal and ERBB2-positive subtypes withpoor prognosis, and we show that MET, together with EMT marker,SNAIL, are highly predictive of poor prognosis in lymph node-negative patients. By generating a unique mouse model in whichthe Met receptor tyrosine kinase is expressed in the mammaryepithelium, along with the examination of MET expression inhuman breast cancer, we have established a specific link betweenMET and basal breast cancer. This work identifies basal breastcancers and, additionally, poor-outcome breast cancers, as thosethat may benefit from anti-MET receptor therapies.
gene expression profiling � mouse models �epithelial to mesenchymal transition
Breast cancer is a heterogeneous disease that comprises distinctbiological entities that are correlated with diverse clinical
outcomes and responses to treatment. Gene expression profilingand molecular pathology have revealed that breast cancers naturallydivide into the luminal, ERBB2-positive, and basal-like subtypes (1,2). These subtypes were named to reflect gene expression patternsof the 2 principal cell types of the differentiated breast, luminalepithelial cells lining the duct and lobule, and myoepithelial cellsthat form a single layer surrounding the luminal cells. The luminalsubtype comprises �60% of breast cancers, is estrogen receptor(ESR1)-positive, and expresses ESR1-responsive genes and luminalmarkers such as keratin 8/18. Up to 25% of breast cancers areidentified with overexpression/amplification of the ERBB2 recep-tor tyrosine kinase, and these tumors are generally ESR1/progesterone receptor (PGR)-negative. The basal group is charac-terized as ESR1/PGR/ERBB2-negative and is frequently positivefor basal keratins 5/6 (3, 4). Breast cancers within the luminalsubtype receive antiestrogen therapies and tend to have a goodprognosis. Because of the lack of treatment options, patients withinthe basal subtype historically have a poor prognosis (1). Hence, anunderstanding of the signaling pathways active in these tumors iscrucial for the generation of targeted therapies.
The MET receptor tyrosine kinase, which is the receptor forhepatocyte growth factor/scatter factor (HGF/SF), is expressed at
elevated levels in 15–20% of human breast cancers (5), and is aprognostic factor for poor outcome (6, 7). High levels of the METreceptor ligand HGF/SF in the serum of breast cancer patients isalso correlated with a shorter disease-free interval after surgery (8)and a higher tumor/lymph node/metastasis score (9).
Although several transgenic mouse models have examined thetumorigenic capacity of Met receptor signaling, none have targetedthe expression of Met specifically to the mammary epithelium.When either HGF or an activated Met receptor is expressed underthe constitutively active metallothionein promoter, transgenic micedevelop malignancies, including mammary tumors (10–12). Addi-tionally, multiparous transgenic mice expressing HGF under themammary-specific whey acidic protein (WAP) promoter developmammary tumors (13). These transgenic mice highlight the sus-ceptibility of the mammary epithelium to transformation by anenhanced Met/HGF signal.
Here, we describe a murine model of breast cancer generated bythe expression of weakly transforming mutants of the Met receptortyrosine kinase in the mammary epithelium. We demonstrate thatMetmt induces mammary tumors with diverse histology, which,based on immunohistochemistry and expression profiling, includestumors with basal and luminal characteristics. By performingmicroarray analyses, we reveal that Metmt-induced basal tumorsshow expression of basal keratins, enrichment for markers of theWnt pathway, display features indicative of epithelial to mesenchy-mal transition (EMT), and cluster with human basal breast cancerand murine models of basal-like breast cancer. In human breastcancer, MET shows consistent elevated expression in the basalsubtype and identifies patients with poor outcome. Our transgenicmouse model, coupled with human breast cancer data, identifiesMET as a rational therapeutic target for both basal and aggressivebreast cancer.
Author contributions: M.G.P., R.L., and M.P. designed research; M.G.P., R.L., S.P., F.P.O.,D.P., I.L.A., S.B.B., N.C., D.Z., M.S., and D.G. performed research; M.G.P., R.L., and M.H.contributed new reagents/analytic tools; M.G.P., R.L., F.P.O., D.P., I.L.A., S.B.B., A.O., R.D.C.,M.H., and M.P. analyzed data; and M.G.P., R.L., I.L.A., S.B.B., and M.P. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
Data deposition: The data reported in this paper have been deposited in the GeneExpression Omnibus (GEO) database, www.ncbi.nlm.nih.gov/geo (accession no. GSE10450).
1M.G.P. and R.L. contributed equally to this work.
2To whom correspondence should be addressed at: Goodman Cancer Centre, McGillUniversity, 1160 Avenue Des Pins Ouest, Cancer Pavilion, Room 511, Montreal, QC, CanadaH3A 1A3. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/cgi/content/full/0810402106/DCSupplemental.
www.pnas.org�cgi�doi�10.1073�pnas.0810402106 PNAS � August 4, 2009 � vol. 106 � no. 31 � 12903–12908
MED
ICA
LSC
IEN
CES
Dow
nloa
ded
by g
uest
on
Apr
il 5,
202
0

ResultsMMTV/Metmt Induces Tumors with Heterogeneous Histology. We havegenerated a transgenic mouse model that expresses the Met recep-tor in mammary epithelium driven by the mouse mammary tumorvirus (MMTV) promoter/enhancer. Mice transgenic for wild-typeand oncogenic variants of the Met receptor (M1248T, Y1003F/M1248T), hereafter called Metmt (12, 14), develop tumors withmoderate penetrance (9–40%) and long latency [381–475 days;supporting information (SI) Table S1] (15). MMTV expression isgreatly enhanced during puberty, pregnancy, and involution (16),and, consistent with this, tumor induction was observed predomi-nantly in multiparous mice (Table S1). Within this cohort of mice,�50% produced carcinomas with solid nodular histopathology(�50%) that are most common to MMTV/Neu and MMTV/PyMTmodels (16, 17). Interestingly, the remaining 50% induced tumorswith either papillary, scirrhous, adenosquamous, or spindle-cellphenotypes (Fig. 1A, Table S2). In general, each animal producedtumors with similar pathology, although a minority of animalsproduced multiple tumors each with distinct pathology (Table S2).
To confirm that mammary tumors in Metmt mice resulted fromintegration of the transgene, we first performed PCR analysis onDNA from tumors and matched-normal tissue and demonstratedthe presence of the Met transgene in both (Fig. S1A). By using anantibody specific for the transgene, Metmt protein was detected onlyin tumor tissue and at variable levels, but was undetectable intumors possessing spindle-cell pathology (5482T, Fig. 1A and Fig.S1B). This is consistent with previous observations demonstrating
suppression of the MMTV promoter in spindle-cell tumors (18). Toaddress the utility of the Metmt transgenic mice as a preclinicalmodel, we examined the ability of epithelial cell lines derived fromMetmt tumors to invade (Fig. S1 C–F). Using a specific small-molecule inhibitor (PHA-665752) and siRNA for Met, we demon-strate that elevated Akt and Erk1 and -2 activity depends on Metactivity (Fig. S1 E and F). In addition, the invasive (Fig. S1 C andD) capacity of these cell lines also depends on Met, which providesstrong support that tumors derived from the Metmt transgenic miceare indeed dependent on Met for a biological response, a criterionrequired for effective preclinical models.
To better understand the molecular characteristics associatedwith the distinct histologies found in Metmt tumors, we performedgene expression analyses using Agilent whole-genome mouse ar-rays. Epithelial tissue from different tumor histologies andmatched-normal were analyzed. Unsupervised hierarchical cluster-ing revealed that the arrayed samples naturally fell into 3 distinctclasses. The first major division within the data separated all normalsamples (‘‘normal epithelium,’’ green) from tumor samples (Fig.1B). Within the group of tumor samples, 2 subclusters formed; thefirst subcluster consisted of tumors with a solid phenotype (‘‘solid,’’purple), and the second consisted of tumors with papillary, scir-rhous, adenosquamous, and spindle cell pathologies (‘‘mixed-pathology,’’ red) (Fig. 1B).
To understand the differences between these tumor types, genesdifferentially expressed between clusters were analyzed for over-representation of biological pathways (Tables S3–S8). Genes over-expressed in Metmt mixed-pathology tumors (red, Fig. 1B) com-pared with the Metmt solid tumors (purple, Fig. 1B) were related togene ontology (GO) categories involving epithelial and mesenchy-mal cell differentiation, including genes linked to EMT (Table S3).Overrepresentation of categories involved in tissue remodeling,collagen production, cytokine signaling, cell migration, angiogen-esis, inflammatory response, and Wnt, integrin, and TGFß signalingpathways were also observed (Table S3). In contrast, Metmt solidtumors (purple, Fig. 1B) showed enrichment for GO terms relatedto apoptosis, cell adhesion, and small GTPase signal transduction(Table S4), supporting the interpretation that distinct tumorpathology is associated with gene expression profiles represent-ing different biological processes. Similar results were obtainedby using GSEA and KEGG (Tables S5–S8).
Metmt Mixed-Pathology Tumors Cluster with Basal-Like Murine Mam-mary Tumors. Transgenic mouse models of breast cancer haverecently been analyzed at the gene expression level and were foundto correlate with the subtypes defined in human breast tumors (19).To determine whether Metmt-induced tumors resemble a specificsubtype of mammary tumors, we used the ‘‘866 intrinsic gene set’’found to be representative of murine mammary tumors (19). Oursamples were combined with this dataset, containing 13 othermouse mammary tumor models, and samples were hierarchicallyclustered over the intrinsic gene set. This analysis further confirmedthat the 2 subtypes of Metmt tumors represent distinct molecularlydefined groups (Fig. 2). Metmt solid tumors clustered closelytogether with mouse models exhibiting solid adenocarcinoma-likepathology, including MMTV/Neu and MMTV/PyMT models (Fig.2). In contrast, the majority of mixed-pathology Metmt tumorsclustered within a group that included p53�/� transplants, p53�/�
irradiated (IR), C3(1)/Tag, and WAP/T121 tumors, which expressbasal and mesenchymal related genes (Fig. 2) (19). These modelsare consistent with our observation that EMT and mesenchymalpathways are enriched in Metmt mixed-pathology tumors (Fig. 4A).
To assess whether Metmt tumors have large-scale transcriptionalsimilarities to human breast cancers, we developed a hierarchicalcluster of Metmt tumors with 172 human breast tumors using across-species intrinsic gene set (19). The results support the inter-pretation that Metmt mixed-pathology tumors reflect a basal sub-type, because these tumors clustered with human basal breast
B
0.0
0.5
1.0
1.5
2.0
6696
N55
9N46
91N
3229
N56
2N50
33N
4164
N51
54N
5156
N55
9T41
64T
562T
5156
T66
96T
4691
T50
33T
5482
T51
54T
3229
T50
33-4
T46
95T
567T
5612
T44
25T
10095
70
93
100100
80
95
Normal Epithelium Solid Tumors Mixed Pathology Tumors
70 80
Hei
ght
AS
olid
nod
ular
car
cino
ma P
apillary adenocarcinom
aA
denosquamous
carcinomaS
cirr
hous
ad
enoc
arci
nom
a
Spindle cell tum
or
Ade
nom
yoep
ithel
ial
carc
inom
a
5488 5482
32295484
4691 5385
Fig. 1. Metmt mice develop mammary tumors with diverse phenotypes thatcluster into 2 groups by microarray analysis. (A) Metmt transgenic mice show aspectrum of histological phenotypes. (Scale bars, 50 �m.) (B) Class discoveryanalysis of Metmt tumor and normal epithelium segregate samples into 3 mainclusters: normal (green), solid tumors (purple), and mixed pathology tumors(red). The significance is calculated by using multiscale bootstrap resamplingand is provided for both approximately unbiased (brown) and bootstrapprobability (gray) P values. The height of the dendrogram represents thedistance between cluster centroids.
12904 � www.pnas.org�cgi�doi�10.1073�pnas.0810402106 Ponzo et al.
Dow
nloa
ded
by g
uest
on
Apr
il 5,
202
0
cancers, whereas Metmt solid tumors clustered among humanluminal tumors (Fig. 3). To further explore these relationships, thecorrelation between each Metmt murine tumor and each centroidfor the 3 human subtypes was quantified (Fig. S2). Consistent withhierarchical clustering results, the Metmt mixed-pathology mousetumors correlated highest with the human basal subtype, whereasthe Metmt solid mouse tumors correlated highest with the humanluminal subtype (Fig. S2).
Metmt Mixed-Pathology Tumors Express Markers of EMT and Basal-Like Breast Cancer. Tumors with solid histology are most commonlyobserved when receptor tyrosine kinases, such as ErbB2, or onco-proteins that activate the Ras pathway are driven by the MMTV
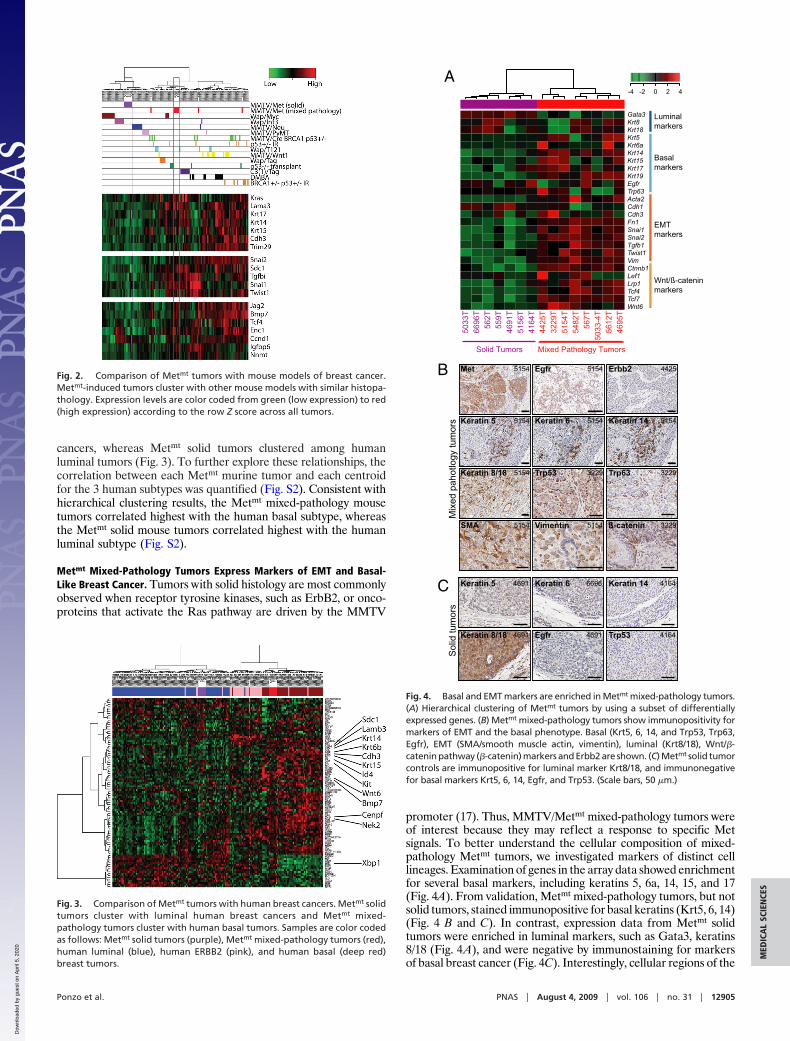
promoter (17). Thus, MMTV/Metmt mixed-pathology tumors wereof interest because they may reflect a response to specific Metsignals. To better understand the cellular composition of mixed-pathology Metmt tumors, we investigated markers of distinct celllineages. Examination of genes in the array data showed enrichmentfor several basal markers, including keratins 5, 6a, 14, 15, and 17(Fig. 4A). From validation, Metmt mixed-pathology tumors, but notsolid tumors, stained immunopositive for basal keratins (Krt5, 6, 14)(Fig. 4 B and C). In contrast, expression data from Metmt solidtumors were enriched in luminal markers, such as Gata3, keratins8/18 (Fig. 4A), and were negative by immunostaining for markersof basal breast cancer (Fig. 4C). Interestingly, cellular regions of the
Fig. 2. Comparison of Metmt tumors with mouse models of breast cancer.Metmt-induced tumors cluster with other mouse models with similar histopa-thology. Expression levels are color coded from green (low expression) to red(high expression) according to the row Z score across all tumors.
Fig. 3. Comparison of Metmt tumors with human breast cancers. Metmt solidtumors cluster with luminal human breast cancers and Metmt mixed-pathology tumors cluster with human basal tumors. Samples are color codedas follows: Metmt solid tumors (purple), Metmt mixed-pathology tumors (red),human luminal (blue), human ERBB2 (pink), and human basal (deep red)breast tumors.
A0 2 4-2-4
Solid Tumors Mixed Pathology Tumors
559T
4164
T
562T
5156
T
6696
T
4691
T
5033
T
5154
T
5033
-4T
4695
T
567T
5612
T
4425
T
5482
T
3229
T
Gata3Krt8
Snai1
Acta2Trp63Egfr
Cdh3
Krt17Krt15Krt14Krt6aKrt5Krt18
Ctnnb1Lef1Lrp1Tcf4Tcf7Wnt6
Vim
Krt19
Fn1
Twist1
Snai2
Cdh1
Tgfb1
Luminal markers
Basal markers
EMTmarkers
Wnt/ß-cateninmarkers
B
Trp53 3229 Trp63 3229
5154Met 4425Erbb25154Egfr
5154Keratin 145154Keratin 65154Keratin 5
5154SMA 3229ß-catenin5154Vimentin
5154Keratin 8/18
Sol
id tu
mor
s
C
Keratin 8/18
Keratin 5 Keratin 6 Keratin 14
Trp53Egfr 4691 4164
4691 6696 4164
4691
Mix
ed p
ahot
logy
tum
ors
Fig. 4. Basal and EMT markers are enriched in Metmt mixed-pathology tumors.(A) Hierarchical clustering of Metmt tumors by using a subset of differentiallyexpressed genes. (B) Metmt mixed-pathology tumors show immunopositivity formarkers of EMT and the basal phenotype. Basal (Krt5, 6, 14, and Trp53, Trp63,Egfr), EMT (SMA/smooth muscle actin, vimentin), luminal (Krt8/18), Wnt/�-catenin pathway (�-catenin) markers and Erbb2 are shown. (C) Metmt solid tumorcontrols are immunopositive for luminal marker Krt8/18, and immunonegativefor basal markers Krt5, 6, 14, Egfr, and Trp53. (Scale bars, 50 �m.)
Ponzo et al. PNAS � August 4, 2009 � vol. 106 � no. 31 � 12905
MED
ICA
LSC
IEN
CES
Dow
nloa
ded
by g
uest
on
Apr
il 5,
202
0
mixed-pathology Metmt tumors showed coexpression of Metmt
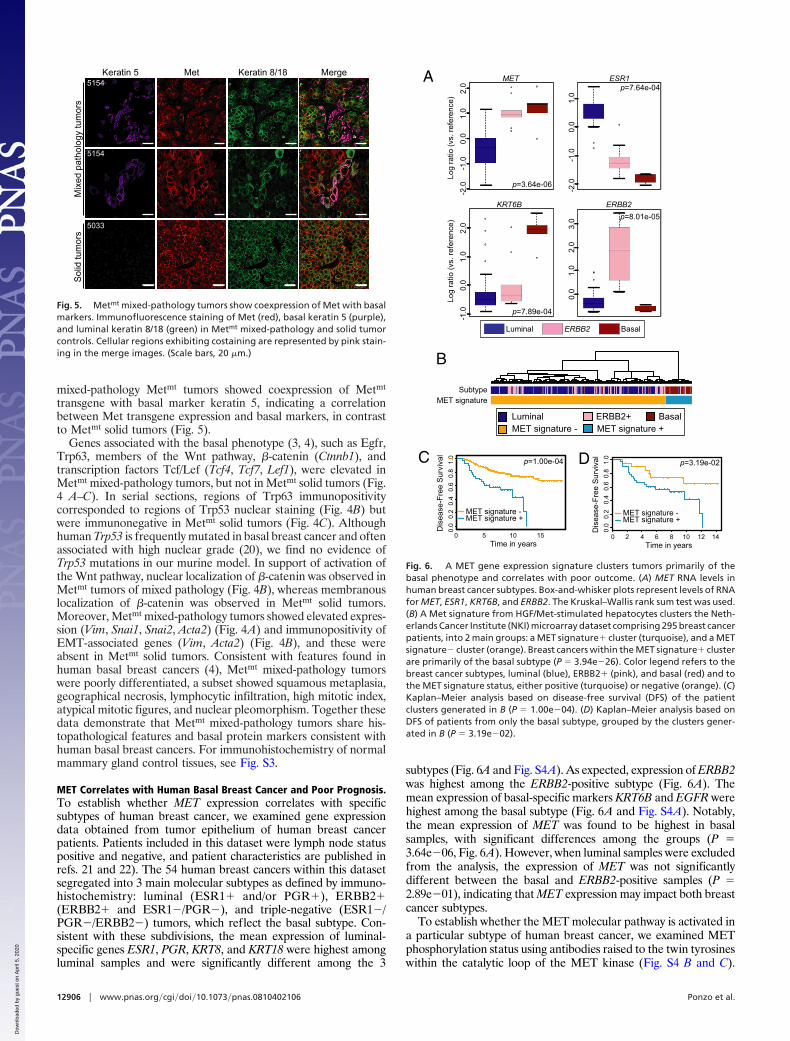
transgene with basal marker keratin 5, indicating a correlationbetween Met transgene expression and basal markers, in contrastto Metmt solid tumors (Fig. 5).
Genes associated with the basal phenotype (3, 4), such as Egfr,Trp63, members of the Wnt pathway, �-catenin (Ctnnb1), andtranscription factors Tcf/Lef (Tcf4, Tcf7, Lef1), were elevated inMetmt mixed-pathology tumors, but not in Metmt solid tumors (Fig.4 A–C). In serial sections, regions of Trp63 immunopositivitycorresponded to regions of Trp53 nuclear staining (Fig. 4B) butwere immunonegative in Metmt solid tumors (Fig. 4C). Althoughhuman Trp53 is frequently mutated in basal breast cancer and oftenassociated with high nuclear grade (20), we find no evidence ofTrp53 mutations in our murine model. In support of activation ofthe Wnt pathway, nuclear localization of �-catenin was observed inMetmt tumors of mixed pathology (Fig. 4B), whereas membranouslocalization of �-catenin was observed in Metmt solid tumors.Moreover, Metmt mixed-pathology tumors showed elevated expres-sion (Vim, Snai1, Snai2, Acta2) (Fig. 4A) and immunopositivity ofEMT-associated genes (Vim, Acta2) (Fig. 4B), and these wereabsent in Metmt solid tumors. Consistent with features found inhuman basal breast cancers (4), Metmt mixed-pathology tumorswere poorly differentiated, a subset showed squamous metaplasia,geographical necrosis, lymphocytic infiltration, high mitotic index,atypical mitotic figures, and nuclear pleomorphism. Together thesedata demonstrate that Metmt mixed-pathology tumors share his-topathological features and basal protein markers consistent withhuman basal breast cancers. For immunohistochemistry of normalmammary gland control tissues, see Fig. S3.
MET Correlates with Human Basal Breast Cancer and Poor Prognosis.To establish whether MET expression correlates with specificsubtypes of human breast cancer, we examined gene expressiondata obtained from tumor epithelium of human breast cancerpatients. Patients included in this dataset were lymph node statuspositive and negative, and patient characteristics are published inrefs. 21 and 22). The 54 human breast cancers within this datasetsegregated into 3 main molecular subtypes as defined by immuno-histochemistry: luminal (ESR1� and/or PGR�), ERBB2�(ERBB2� and ESR1�/PGR�), and triple-negative (ESR1�/PGR�/ERBB2�) tumors, which reflect the basal subtype. Con-sistent with these subdivisions, the mean expression of luminal-specific genes ESR1, PGR, KRT8, and KRT18 were highest amongluminal samples and were significantly different among the 3
subtypes (Fig. 6A and Fig. S4A). As expected, expression of ERBB2was highest among the ERBB2-positive subtype (Fig. 6A). Themean expression of basal-specific markers KRT6B and EGFR werehighest among the basal subtype (Fig. 6A and Fig. S4A). Notably,the mean expression of MET was found to be highest in basalsamples, with significant differences among the groups (P �3.64e�06, Fig. 6A). However, when luminal samples were excludedfrom the analysis, the expression of MET was not significantlydifferent between the basal and ERBB2-positive samples (P �2.89e�01), indicating that MET expression may impact both breastcancer subtypes.
To establish whether the MET molecular pathway is activated ina particular subtype of human breast cancer, we examined METphosphorylation status using antibodies raised to the twin tyrosineswithin the catalytic loop of the MET kinase (Fig. S4 B and C).
MergeMetKeratin 55154
5154
5033
Keratin 8/18S
olid
tum
ors
Mix
ed p
atho
logy
tum
ors
Fig. 5. Metmt mixed-pathology tumors show coexpression of Met with basalmarkers. Immunofluorescence staining of Met (red), basal keratin 5 (purple),and luminal keratin 8/18 (green) in Metmt mixed-pathology and solid tumorcontrols. Cellular regions exhibiting costaining are represented by pink stain-ing in the merge images. (Scale bars, 20 �m.)
A
Log
ratio
(vs.
refe
renc
e)0.
01.
02.
0-2
.0-1
.0
MET
p=3.64e-06
0.0
1.0
-2.0
-1.0
ESR1p=7.64e-04
0.0
1.0
2.0
3.0
ERBB2p=8.01e-05
0.0
1.0
2.0
-1.0
KRT6B
p=7.89e-04
Log
ratio
(vs.
refe
renc
e)
Luminal BasalERBB2
MET signatureSubtype
Luminal ERBB2+ BasalMET signature - MET signature +
CD
isea
se-F
ree
Sur
viva
l
Time in years
MET signature -MET signature +
p=1.00e-04
10
0.0
0.2
0.4
0.6
0.8
1.0
1550
D
B
Dis
ease
-Fre
e S
urvi
val
0.0
0.2
0.4
0.6
0.8
1.0
0 2 4 6 8 10 12 14
p=3.19e-02
MET signature -MET signature +
Time in years
Fig. 6. A MET gene expression signature clusters tumors primarily of thebasal phenotype and correlates with poor outcome. (A) MET RNA levels inhuman breast cancer subtypes. Box-and-whisker plots represent levels of RNAfor MET, ESR1, KRT6B, and ERBB2. The Kruskal–Wallis rank sum test was used.(B) A Met signature from HGF/Met-stimulated hepatocytes clusters the Neth-erlands Cancer Institute (NKI) microarray dataset comprising 295 breast cancerpatients, into 2 main groups: a MET signature� cluster (turquoise), and a METsignature� cluster (orange). Breast cancers within the MET signature� clusterare primarily of the basal subtype (P � 3.94e�26). Color legend refers to thebreast cancer subtypes, luminal (blue), ERBB2� (pink), and basal (red) and tothe MET signature status, either positive (turquoise) or negative (orange). (C)Kaplan–Meier analysis based on disease-free survival (DFS) of the patientclusters generated in B (P � 1.00e�04). (D) Kaplan–Meier analysis based onDFS of patients from only the basal subtype, grouped by the clusters gener-ated in B (P � 3.19e�02).
12906 � www.pnas.org�cgi�doi�10.1073�pnas.0810402106 Ponzo et al.
Dow
nloa
ded
by g
uest
on
Apr
il 5,
202
0

Notably, triple-negative breast tumors with high MET RNA (Fig.6A) displayed high MET immunostaining and stained positive withMET phosphospecific antibodies (Fig. S4B), consistent with METactivation and signaling in human basal breast cancers. In contrast,luminal tumors with low MET RNA (Fig. 6A) showed no detectableMET or phosphospecific MET immunostaining (Fig. S4C). More-over, when a MET transcriptional signature (23) was applied to anindependent human breast cancer dataset (NKI) (24), the METsignature clearly separated human breast tumors into 2 clusters(Fig. 6B), with the smaller cluster (turquoise) representing aninduction of the MET transcriptional response when comparedwith the larger cluster (orange). Genes expected to be overex-pressed in MET activation were significantly higher in the turquoisecluster (P � 2.20e�16), whereas genes expected to be repressed byMET activation were significantly lower in this cluster (P �1.90e�09). Notably, tumors belonging to the human basal subtypewere overrepresented in the turquoise cluster identified by theMET transcriptional signature (P � 2.99e�24), supporting theassociation of MET with the basal group. The cluster of tumors witha MET-activated transcriptional signature (turquoise) had a signif-icantly worse overall prognosis (Fig. 6C, P � 1.00e�04). Further-more, the MET signature identified poor outcome within the basalsubtype (Fig. 6D, P � 3.19e�02). Unfortunately, there wereinsufficient luminal and ERBB2-positive samples in the MET-activated group (n � 3 and n � 2, respectively) to assess whetherthe MET signature was associated with poor prognosis in thesesubtypes. In cDNA microarray analyses conducted in a prospec-tively accrued cohort of women with axillary lymph node-negative(ANN) breast cancer (25), we identified MET as differentiallyexpressed between tumors from 48 women who experienced dis-ease recurrence within 4 years of diagnosis when compared with41 who remained disease-free for �10 years (Table S9; P �1.33e�02 by standard t test, P � 8.70e�03 by nonparametricWilcoxon rank-sum test in univariate analysis), an associationthat was independent of traditional clinicopathological param-eters (SI Text, P � 3.10e�02).
To further investigate the correlation between transcriptionalactivation of MET and the basal subtype, we stained for METprotein in a cohort of 668 ANN human breast cancer cases (26).High MET immunostaining was observed in all subtypes, but asignificantly greater proportion of basal subtype tumors were METpositive (‘‘high MET,’’ 65%) as opposed to MET negative (‘‘lowMET,’’ 35%), when compared with the ERBB2-positive and lumi-nal subtypes (Fig. 7A, P � 6.51e�03). Consistent with previousresults, the Kaplan–Meier plot demonstrates that MET-positivetumor status correlates with poor disease-free survival (DFS)outcome among ANN patients (Fig. 7A, log-rank P � 3.94e�02).Additionally, MET-positive protein status is associated with pooroutcome in univariate analyses irrespective of breast cancer subtype(SI Text, P � 6.31e�01 by Cox model test for differences betweenbasal and nonbasal, Table S10 and Fig. S4D), which is consistentwith MET expression being causally related to an aggressivephenotype.
Because we observed a strong induction of EMT genes includingSnail (Snai1) in the murine Metmt mixed-pathology tumors (Fig.4A) as well as the induction of SNAIL (SNAI1) mRNA afterstimulation of epithelial cells with HGF (Fig. S4E) we investigatedcoexpression of MET with the downstream EMT target, SNAIL.Notably, tumors exhibiting coexpression of MET with SNAILshowed a significant correlation with poor outcome when com-pared with those negative for either MET and/or SNAIL (Fig. 7B,log-rank P � 7.40e�03), demonstrating that the combination ofMET with an EMT signal, SNAIL, is a strong predictor of pooroutcome. This association persisted in multivariate analysis afteradjustment for traditional histopathologic prognostic factors(Tables S11 and S12) and for basal/nonbasal status (SI Text andTable S13). Together, these results strongly support a role for MET
signaling both in human basal breast cancers and breast cancers withpoor outcome.
DiscussionUnderstanding the oncogenic pathways that distinguish subsets ofhuman cancer is critical for the development of new therapies.Here, we show that mammary-specific expression of weakly onco-genic forms of the Met receptor promotes the formation ofmammary carcinomas with histology, gene expression, and immu-nohistochemical profiles similar to human basal breast cancers.
Although elevated MET protein levels have been associated withpoor outcome in human breast cancer, the role for the METreceptor tyrosine kinase in the induction and development of breastcancer is poorly understood. Mammary-specific expression ofMMTV/Metmt surprisingly induced tumors with distinct histologi-cal phenotypes (solid and mixed pathology, Fig. 1). The expressionprofile derived from Metmt mixed-pathology tumors clusters thesewith human basal breast cancers and murine models of basal tumors(Figs. 2 and 3). This is consistent with findings that Metmt mixed-pathology tumors display histology similar to the human basalsubtype and express a series of known basal markers (Fig. 4).
When compared with other murine models, Metmt mixed-pathology tumors clustered with murine tumors having basal char-acteristics including MMTV/Cre;p53�/�, TgWAP/T121, TgWAP/Tag, and TgC3(1)/Tag tumors (Fig. 2). Within this class, Metmt
mixed-pathology tumors shared expression and immunopositivityof EMT-associated genes, including Tgfbi, Snai1, Snai2, and Twist1,Sdc1 (18), in addition to basal-like markers including Krt 5, 6a, 14,and 17 as well as Kras (Figs. 2 and 4). Although a detailed analysisof previous Met/HGF transgenic mouse models is lacking, modelswith constitutively activated Met, such as Tpr-Met or WAP/HGF(11, 13, 27), also produce mammary tumors with heterogeneoushistological phenotypes. In some cases, tumors were shown to bekeratin 6-positive (27) with nuclear accumulation of �-catenin (13),
A
0 12 36 60 84 108 132 156 180
0.6
0.7
0.8
0.9
1.0
MET-MET+
p=3.94e-02
Dis
ease
-Fre
e S
urvi
val
Time in Months
Basal (n=60) 35 65ERBB2 (n=41) 54 46Luminal (n=408) 57 43p=6.51e-03
Low MET (%) (0-3)
High MET (%) (4-8)Subtype
Dis
ease
-Fre
e S
urvi
val
0 12 36 60 84 108 132 156 180
0.6
0.7
0.8
0.9
1.0
p=7.40e-03
Time in Months
MET-/SNAIL-MET-/SNAIL+MET+/SNAIL-MET+/SNAIL+
B
Fig. 7. Elevated MET protein correlates with poor-outcome human basalbreast tumors and MET/SNAIL coexpression correlates with poor outcome. (A)Kaplan–Meier analysis shows the relationship between MET protein level andDFS. (B) Kaplan–Meier analysis shows the relationship between MET andSNAIL proteins and DFS.
Ponzo et al. PNAS � August 4, 2009 � vol. 106 � no. 31 � 12907
MED
ICA
LSC
IEN
CES
Dow
nloa
ded
by g
uest
on
Apr
il 5,
202
0

consistent with data observed in our Metmt mixed-pathologytumors (Fig. 4B).
The observation that Metmt-induced tumors with solid patholo-gies resemble MMTV/Neu, MMTV/PyMT, and MMTV/ras tumorswithin a luminal subtype (Figs. 2 and 4A) may reflect the ability ofthe Metmt carrying the Y1003F mutation to promote enhancedactivity of the ras pathway (14). The presence of both luminal andbasal-like tumor subtypes raises the possibility that Met may targeta multipotent progenitor cell(s) with the capacity to differentiatetoward each lineage. In support of this, coexpression of Met withbasal and luminal markers, keratin 5 and 8/18 respectively, wasobserved in cells within Metmt mixed-pathology basal-like tumorsbut not in solid luminal-like tumors (Fig. 5). Moreover, cellsimmunopositive for nuclear Trp63 or Trp53 in Metmt basal-liketumors (Fig. 4B) may be indicative of a stem/progenitor cellpopulation (28).
We show that elevated MET RNA, protein, and transcriptionalresponse are associated with the human basal subtype (Figs. 6 and7A) and that when elevated, MET immunopositive sections stainwith MET phosphospecific sera indicative of MET activation (Fig.S4 B and C). Moreover, in patients who are lymph node-negativeat presentation, elevated MET protein is associated with pooroutcome (Fig. 7A). Specifically, in the node-negative cohort, ele-vated MET protein is significantly correlated with the basal subtype(P � 6.51e�03, Fig. 7A). Among the biological processes high-lighted in gene expression studies with the Metmt basal tumors aregenes involved in EMT (Fig. 4A and Table S3). Within EMT-related genes, one of the most differentially regulated genes inMetmt mixed-pathology tumors is the Snail transcription factor,which regulates EMT during development. Strikingly, elevatedlevels of both MET and SNAIL in tumors is significantly correlatedwith poor outcome in lymph node-negative patients whencompared with either protein alone (log-rank P � 7.40e�03and Fig. 7B).
This is a first report of an MMTV-driven receptor tyrosine kinasemouse model that produces tumors resembling human basal breastcancer. Significantly, an adjoining manuscript demonstrates a sim-ilar basal phenotype induced by a different oncogenically activated
Met receptor tyrosine kinase driven from its endogenous promoter(29). Together, this provides strong support that activation of Metsignaling pathways play an important role in the induction of tumorsof the basal subtype. Because human basal breast cancer is espe-cially difficult to treat due to a lack of understanding of the genesand processes involved in its induction, developing a model for thedissection of this subtype is of utmost importance. Here, weestablish that the MET receptor is casually associated with basaland poor outcome breast cancer. Elevated MET RNA, protein anda MET transcriptional profile correlates with human basal breastcancer indicating that MET may prove a useful therapeutic targetin the management of these patients.
MethodsTransgenic Mice. MMTV/Met mice were generated as described (15). The 8.7-kbfragment containing the Met cDNA, MMTV promoter, and SV40 polyA wasinjected into the pronuclei of FVB/N zygotes and implanted into FVB/N hosts bythe McIntyre Transgenic Core Facility (Montreal, Canada). Mice were housed inaccordance with McGill University Animal Ethics Committee guidelines.
Mouse and Human Tissue Processing. Processing of human and mouse tissueshave been described (21, 30). For human ANN breast cancer cases, a prospectivelyascertained consecutive series was enrolled, as previously described (25). Consentwas obtained from patients in this Institutional Review Board-approved study.
Immunohistochemical Analysis. Tissue sections were stained by using the ABCElite and MOM reagents (Vector Laboratories). Detailed procedures are providedin SI Text.
Gene Expression Microarray Data. RNA was prepared for hybridization, and datawere analyzed as previously described (21). Detailed procedures are provided inSI Text.
ACKNOWLEDGMENTS. We thank Anie Monast for animal assistance; KarimKhetani for reviewing H&E slides; and Peter Siegel, William Muller, and AlainNepveu for helpful discussions. M.G.P. was supported by the Canadian Institutesof Health Research (CIHR) MD/PhD Studentship, and a Richard H. TomlinsonDoctoral Fellowship. R.L. was supported by the Natural Sciences and EngineeringResearch Council, McGill School of Computer Science, and the Andre Courte-manche Fellowship for Bioinformatics. S.B.B. held a CIHR Senior InvestigatorAward (2002–2007). M.P. holds the Diane and Sal Guerrera Chair in CancerGenetics at McGill University. This work was supported by grants from theNational Cancer Institute of Canada with funds from the Terry Fox Foundation.
1. Sorlie T, et al. (2001) Gene expression patterns of breast carcinomas distinguish tumorsubclasses with clinical implications. Proc Natl Acad Sci USA 98(19):10869–10874.
2. Da Silva L, Clarke C, Lakhani SR (2007) Demystifying basal-like breast carcinomas. J ClinPathol 60(12):1328–1332.
3. Nielsen TO, et al. (2004) Immunohistochemical and clinical characterization of thebasal-like subtype of invasive breast carcinoma. Clin Cancer Res 10(16):5367–5374.
4. Livasy CA, et al. (2006) Phenotypic evaluation of the basal-like subtype of invasivebreast carcinoma. Mod Pathol 19(2):264–271.
5. Camp RL, Rimm EB, Rimm DL (1999) Met expression is associated with poor outcome inpatients with axillary lymph node negative breast carcinoma. Cancer 86(11):2259–2265.
6. Lengyel E, et al. (2005) C-Met overexpression in node-positive breast cancer identifiespatients with poor clinical outcome independent of Her2/neu. Int J Cancer 113(4):678–682.
7. Ghoussoub RA, et al. (1998) Expression of c-met is a strong independent prognosticfactor in breast carcinoma. Cancer 82(8):1513–1520.
8. Yamashita J, et al. (1994) Immunoreactive hepatocyte growth factor is a strong andindependent predictor of recurrence and survival in human breast cancer. Cancer Res54(7):1630–1633.
9. Taniguchi T, et al. (1995) Serum concentrations of hepatocyte growth factor in breastcancer patients. Clin Cancer Res 1(9):1031–1034.
10. Takayama H, et al. (1997) Diverse tumorigenesis associated with aberrant developmentin mice overexpressing hepatocyte growth factor/scatter factor. Proc Natl Acad Sci USA94(2):701–706.
11. Liang TJ, Reid AE, Xavier R, Cardiff RD, Wang TC (1996) Transgenic expression oftpr-met oncogene leads to development of mammary hyperplasia and tumors. J ClinInvest 97(12):2872–2877.
12. Jeffers M, et al. (1998) The mutationally activated Met receptor mediates motility andmetastasis. Proc Natl Acad Sci USA 95(24):14417–14422.
13. Gallego MI, Bierie B, Hennighausen L (2003) Targeted expression of HGF/SF in mousemammary epithelium leads to metastatic adenosquamous carcinomas through theactivation of multiple signal transduction pathways. Oncogene 22(52):8498–8508.
14. Abella JV, et al. (2005) Met/Hepatocyte growth factor receptor ubiquitination sup-presses transformation and is required for Hrs phosphorylation. Mol Cell Biol25(21):9632–9645.
15. Petkiewicz SL (2007) The Met receptor tyrosine kinase in mammary gland tumorigen-esis and development. PhD Thesis (McGill University, Montreal).
16. Cardiff RD, Muller WJ (1993) Transgenic mouse models of mammary tumorigenesis.Cancer Surv 16:97–113.
17. Rosner A, et al. (2002) Pathway pathology: Histological differences between ErbB/Rasand Wnt pathway transgenic mammary tumors. Am J Pathol 161(3):1087–1097.
18. Damonte P, Gregg JP, Borowsky AD, Keister BA, Cardiff RD (2007) EMT tumorigenesisin the mouse mammary gland. Lab Invest 87(12):1218–1226.
19. Herschkowitz JI, et al. (2007) Identification of conserved gene expression featuresbetween murine mammary carcinoma models and human breast tumors. Genome Biol8(5):R76.
20. Rakha EA, et al. (2006) Morphological and immunophenotypic analysis of breastcarcinomas with basal and myoepithelial differentiation. J Pathol 208(4):495–506.
21. Finak G, et al. (2006) Gene expression signatures of morphologically normal breasttissue identify basal-like tumors. Breast Cancer Res 8(5):R58.
22. Finak G, et al. (2008) Stromal gene expression predicts clinical outcome in breast cancer.Nat Med 14(5):518–527.
23. Kaposi-Novak P, et al. (2006) Met-regulated expression signature defines a subset ofhuman hepatocellular carcinomas with poor prognosis and aggressive phenotype.J Clin Invest 116(6):1582–1595.
24. van de Vijver MJ, et al. (2002) A gene-expression signature as a predictor of survival inbreast cancer. N Engl J Med 347(25):1999–2009.
25. Andrulis IL, et al. (1998) neu/erbB-2 amplification identifies a poor-prognosis group ofwomen with node-negative breast cancer. Toronto Breast Cancer Study Group. J ClinOncol 16(4):1340–1349.
26. Mulligan AM, Pinnaduwage D, Bull SB, O’Malley FP, Andrulis IL (2008) Prognostic effectof basal-like breast cancers is time dependent: Evidence from tissue microarray studieson a lymph node-negative cohort. Clin Cancer Res 14(13):4168–4174.
27. Welm AL, Kim S, Welm BE, Bishop JM (2005) MET and MYC cooperate in mammarytumorigenesis. Proc Natl Acad Sci USA 102(12):4324–4329.
28. DiRenzo J, et al. (2002) Growth factor requirements and basal phenotype of animmortalized mammary epithelial cell line. Cancer Res 62(1):89–98.
29. Graveel CR, et al. (2009) Met induces diverse breast carcinomas in mice and is associatedwith human basal breast cancer 10.1073/pnas.0810403106.
30. Ursini-Siegel J, et al. (2008) ShcA signalling is essential for tumour progression in mousemodels of human breast cancer. EMBO J 27(6):910–920.
12908 � www.pnas.org�cgi�doi�10.1073�pnas.0810402106 Ponzo et al.
Dow
nloa
ded
by g
uest
on
Apr
il 5,
202
0