Embed Size (px)
Citation preview

1
1
Mesenchymal Stem Cell and Chondrocyte Fates in a Multi-Shear Microdevice are
Regulated by Yes-Associated Protein
Weiliang Zhonga, Kang Tian
a, Linan Li
a, Weiguo Zhang
a*, Shouyu Wang
a*, Jianhua Qin
b
a Department of Orthopaedics, First Affiliated Hospital of Dalian Medical University, 222 Zhongshan Road,
Dalian, P.R. China
b Department of Biotechnology, Dalian Institute of Chemical Physics, CAS, 457 Zhongshan Road, Dalian,
P.R. China
*Corresponding to Prof. Weiguo Zhang and Shouyu Wang
Tel/Fax: +86- 411-83632383
E-mail: [email protected]
Running Title: YAP and Mesenchymal Stem Cell and Chondrocyte Fates
Page 1 of 35
Stem
Cel
ls a
nd D
evel
opm
ent
Mes
ench
ymal
Ste
m C
ell a
nd C
hond
rocy
te F
ates
in a
Mul
ti-Sh
ear
Mic
rode
vice
are
Reg
ulat
ed b
y Y
es-A
ssoc
iate
d Pr
otei
n (d
oi: 1
0.10
89/s
cd.2
012.
0685
)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

2
2
Abstract
Mechanical cues exert considerable influence on the fates of stem cells and terminally
differentiated chondrocytes. The elucidation of the interactions between cell fate and mechanical
cues in nuclear mechanotransduction will provide new clues to modulate tissue homeostasis and
regeneration. In this study, we used an integrated microfluidic perfusion device to simultaneously
generate multiple-parameter fluid shear stresses to investigate the role of fluid flow stimuli in the
regulation of YAP expression and the fates of MSCs and primary chondrocytes. YAP expression
was regulated by the level of fluid flow stimulus in both MSCs and chondrocytes. An increase in
the magnitude of stimulation enhanced the expression of YAP, ultimately resulting in an increase
in osteogenesis and a decrease in adipogenesis for MSCs, and initiating dedifferentiation for
chondrocytes. Cytochalasin D not only repressed nuclear YAP accumulation in the flow state but
also abrogated flow-induced effects on MSC differentiation and the chondrocyte phenotype,
resulting in MSC adipogenesis and the maintenance of the chondrocyte phenotype. Our findings
reveal the connection between YAP and MSC/chondrocyte fates in a fluid flow-induced
mechanical microenvironment and provide new insights into the mechanisms by which
mechanical cues regulates the cell fates of MSCs and chondrocytes.
Keywords: YAP; Microfluidics; Fluid shear stress; Mesenchymal stem cell; Chondrocyte
Page 2 of 35
Stem
Cel
ls a
nd D
evel
opm
ent
Mes
ench
ymal
Ste
m C
ell a
nd C
hond
rocy
te F
ates
in a
Mul
ti-Sh
ear
Mic
rode
vice
are
Reg
ulat
ed b
y Y
es-A
ssoc
iate
d Pr
otei
n (d
oi: 1
0.10
89/s
cd.2
012.
0685
)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

3
3
1. Introduction
The regulation of stem cell differentiation and the chondrocyte phenotype remains a
challenge in tissue engineering and regenerative medicine. When the balance between the
adipogenesis and osteogenesis of mesenchymal stem cells (MSCs) and the maintenance of
chondrogenic phenotype are disrupted, bone and cartilage disorders such as osteoporosis and
osteoarthritis may arise [1]. During development, regeneration and homeostasis in the body,
mechanical cues play important role in regulating the fate and lineage commitment of stem cells
[2,3]. Therefore, it is important to elucidate the intricate reciprocal molecular interactions
between mechanical microenvironment and stem cells or chondrocytes in the musculoskeletal
system. Under physiological loading conditions, mechanical cues are recognized as an essential
component in the maintenance of articular cartilage matrix and healthy bone tissue homeostasis.
The interstitial flow driven by dynamic loading through the cartilage matrix or the canalicular
network of bone is an essential mechanical signal for the survival of chondrocyte or osteocyte
[4,5]. Increasing evidence has demonstrated that fluid flow plays an important role in regulating
cell behaviors and functions in the musculoskeletal system [6,7].
The cells perceive and transmit mechanical cues via mechanotransduction mechanisms. It
has been well established that the actin cytoskeleton plays a significant role in mechanosensing
and mechanotransduction [8,9]. The lineage commitment of MSCs can be predicted based on cell
geometry and cytoskeletal organization [10,11]. For example, flattened morphologies, stiffer
Page 3 of 35
Stem
Cel
ls a
nd D
evel
opm
ent
Mes
ench
ymal
Ste
m C
ell a
nd C
hond
rocy
te F
ates
in a
Mul
ti-Sh
ear
Mic
rode
vice
are
Reg
ulat
ed b
y Y
es-A
ssoc
iate
d Pr
otei
n (d
oi: 1
0.10
89/s
cd.2
012.
0685
)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

4
4
substrates and tensile stress result in high cytoskeletal tension, which promotes osteogenesis. In
contrast, rounded morphologies, soft substrates and a lack of extrinsic mechanical force
contribute to adipogenesis [2,12]. Similarly, chondrocytes with a rounded shape or exposed to
soft substrates can maintain their phenotype; otherwise, these cells are likely to dedifferentiate
[13,14].
YAP (Yes-associated protein) plays crucial roles in cell proliferation, survival, differentiation,
tissue regeneration and organ size control, and these effects are mediated through the Hippo
pathway [15-17]. Recently, it has been confirmed that YAP is a regulator of the nuclear
transduction of mechanical cues [18]. The activity of YAP is regulated by intrinsically local cues,
such as cell shape [18,19], cell-cell contact [17,20], extracellular matrix stiffness [18] and
cytoskeleton tension [18,19,21], which have a distinct effect on the fate decision that occur
during stem cell differentiation [16,22-24]. However, the responses of YAP to extrinsic
mechanical force stimuli such as pressure stress, tensile stress and fluid shear stress have not been
well established. In particular, the relationships between YAP and the lineage commitment of
MSCs and chondrocyte phenotype in response to passive mechanical force remain incompletely
understood.
Currently, microfluidic-based microdevices are attracting much attention in the field of
cellular biomechanics, and their major advantage is the reduced time and resources they require
due to their increased experimental throughput and the minimized size of the experimental
platform compared with conventional way [25,26]. In this study, we employed a novel integrated
Page 4 of 35
Stem
Cel
ls a
nd D
evel
opm
ent
Mes
ench
ymal
Ste
m C
ell a
nd C
hond
rocy
te F
ates
in a
Mul
ti-Sh
ear
Mic
rode
vice
are
Reg
ulat
ed b
y Y
es-A
ssoc
iate
d Pr
otei
n (d
oi: 1
0.10
89/s
cd.2
012.
0685
)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

5
5
microfluidic device that can simultaneously generate multiple shear stresses to investigate the
role of fluid flow stimuli in the regulation of YAP expression and the lineage commitment of
MSCs and chondrocytes.
2. Materials and methods
2.1. Microfluidic device design and fabrication
The design and operation of the microfluidic device were presented in Fig. 1A. The device
had one inlet, one outlet, and four cell culture chambers connected to resistance channels of
different dimensions. Each chamber was 100 µm in height, 1.2 mm in width, and 6 mm in length.
All devices were fabricated using conventional microfabrication techniques involving SU-8
(Microchem, Newton, MA, USA) photolithography and polydimethylsiloxane (PDMS) (Sylgard
184, Dow Corning, Midland, MI, USA) soft lithography [27]. Briefly, the mask was designed in
AutoCAD 2007 (Autodesk) and printed on transparencies with a 4000 dpi resolution. The
transparency mask was used in 1:1 contact photolithography with SU-8 photoresist to yield a
negative master that was constructed from a photoresist relief on a silicon wafer. The PDMS was
poured over the patterned wafer to completely cover the pattern, and the sample was placed in an
oven at 80°C for 60 min. The PDMS replica and a clean glass substrate were irreversibly sealed
using oxygen plasma (2 torr, 100 W) for 1 min.
Page 5 of 35
Stem
Cel
ls a
nd D
evel
opm
ent
Mes
ench
ymal
Ste
m C
ell a
nd C
hond
rocy
te F
ates
in a
Mul
ti-Sh
ear
Mic
rode
vice
are
Reg
ulat
ed b
y Y
es-A
ssoc
iate
d Pr
otei
n (d
oi: 1
0.10
89/s
cd.2
012.
0685
)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

6
6
2.2. Numerical modeling
To evaluate the local fluid shear stress distribution in the chamber, we simulated the 3D flow
field in the chamber using a finite volume method (FVM) -based CFD code in FLUENT 6.3
(ANSYS, Inc., Lebanon, NH, USA). We used a model in which the flow was assumed to be
laminar, viscous, and incompressible, and we designed the microfluidic networks based on the
electric circuit analogy [28]. Using this analogy, individual channel sections were treated as
resistances within the flow circuit. A precise solution for the hydraulic resistance of the 3D
rectangular channel was derived via Fourier series expansions. To avoid the computational rigor
required to solve Fourier series expansions, we used an approximate version in algebraic form
[29]:
3
12 1
1 0.63( / )
LR
h w h w
(1)
where is the fluid viscosity, w is the channel width and h is the channel height, for h w .
For a square microchannel, the resistance can be calculated by
4
128.4R L
h (2)
A constant pressure drop p results in a constant flow rate Q . This result can be summarized
using the Hagen–Poiseuille equation, as follows [30]:
Hp QR (3)
Using simple algebraic manipulation, we determined the pressure values at the inlets and outlets
of the individual culture chambers. These pressure values were then used as the inlet and outlet
Page 6 of 35
Stem
Cel
ls a
nd D
evel
opm
ent
Mes
ench
ymal
Ste
m C
ell a
nd C
hond
rocy
te F
ates
in a
Mul
ti-Sh
ear
Mic
rode
vice
are
Reg
ulat
ed b
y Y
es-A
ssoc
iate
d Pr
otei
n (d
oi: 1
0.10
89/s
cd.2
012.
0685
)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

7
7
pressure conditions to simulate the 3D flow field in each culture chamber using the CFD method.
The incompressible Navier-Stokes equations were used to model the steady-state flow field in the
culture chambers. It is worth noting that we did not consider the effect of the cells on the flows in
the present simulations. The computational domain was discretized using approximately 52,000
hexahedral meshes and solved using FVM along with the aforementioned inlet/outlet pressure
conditions and no-slip boundary conditions at the chamber walls. The density of the perfusion
medium was 993.2 kg/m3, and its viscosity was 7.85×10
-4 Pa s at 37°C.
2.3. The isolation and culture of MSCs and chondrocytes
All experimental procedures were approved by the Committee on Animal Use and Care of
Dalian Medical University. Articular cartilage chondrocytes were isolated from the
humeral heads, femoral heads and femoral condyles of male Sprague Dawley rats weighing
80-120 g, as previously described [31]. Briefly, chondrocytes were isolated by digestion with
0.15% type II collagenase for 16 h and resuspended in Dulbecco’s modified Eagle’s
medium/F-12 (Hyclone, USA) containing 10% fetal bovine serum (FBS, Hyclone, USA), 50
mg/ml ascorbic acid-2-phosphate (Sigma, USA), and 100 units/ml penicillin-streptomycin. The
primary chondrocytes were used in the subsequent experiments. Primary rat mesenchymal stem
cells (MSCs) were isolated from the bilateral femurs and tibias of the same rats. The distal ends
of the bone were cut open, and the marrow cavities were lavaged with sterile phosphate-buffered
saline (PBS). The cells were resuspended in low-glucose Dulbecco’s modified Eagle medium
Page 7 of 35
Stem
Cel
ls a
nd D
evel
opm
ent
Mes
ench
ymal
Ste
m C
ell a
nd C
hond
rocy
te F
ates
in a
Mul
ti-Sh
ear
Mic
rode
vice
are
Reg
ulat
ed b
y Y
es-A
ssoc
iate
d Pr
otei
n (d
oi: 1
0.10
89/s
cd.2
012.
0685
)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

8
8
(GIBCO Invitrogen, USA) containing 10% FBS (Hyclone, USA) and 100 units/ml
penicillin-streptomycin. After 48 h of incubation at 37°C in 5% CO2, the medium was changed to
remove the non-adherent cells. After two passages, the attached MSCs were devoid of any
non-adhering cells and used in the following experiments.
The microdevice was sterilized in an autoclave and then air-dried on a clean bench. The cell
culture chambers were coated with 100 μg/ml fibronectin (Sigma, USA) for 1 h at room
temperature. Then, the chambers were washed with PBS. MSC suspension of 0.5×105 cells/ml
and a chondrocyte suspension of 1×105 cells/ml were individually injected into chambers through
the outlet, and the device was incubated at 37°C for 12 h to allow cell attachment. Then, the inlet
of the device was connected to a peristaltic pump (Longer Pump BT100-2J, China), and the outlet
was connected to a reservoir. MSCs were exposed to adipogenic-osteogenic co-induction medium
during perfusion culture. The mixed induction medium contained 1:1 adipogenic
induction:osteogenic differentiation media (Cyagen Biosciences Inc, Sunnyvale, CA, USA), as
previously described [12]. The cells were exposed to 1 µM cytochalasin D (CytoD) (Sigma, USA)
for 1 h to disrupt stress fibers before the application of the flow stimulus.
2.4. Immunofluorescence staining
Samples in the device were washed with PBS, fixed with 4% paraformaldehyde at room
temperature for 15 min and permeabilized with 0.1% Triton X-100 for 10 min. After washing
with PBS 3 times, the samples were blocked with normal goat serum at room temperature for 30
Page 8 of 35
Stem
Cel
ls a
nd D
evel
opm
ent
Mes
ench
ymal
Ste
m C
ell a
nd C
hond
rocy
te F
ates
in a
Mul
ti-Sh
ear
Mic
rode
vice
are
Reg
ulat
ed b
y Y
es-A
ssoc
iate
d Pr
otei
n (d
oi: 1
0.10
89/s
cd.2
012.
0685
)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

9
9
min, incubated with primary antibodies against YAP (Santa Cruz, USA), PPAR (Santa Cruz,
USA), Runx2 (Santa Cruz, USA), Sox9 (Santa Cruz, USA), collagen II (Sigma, USA), collagen I
(Sigma, USA) at 4°C overnight; and then incubated with FITC-conjugated goat anti-rabbit IgG or
TRITC-conjugated goat anti-rabbit secondary antibodies (Zhongshan, China) at room
temperature for 1 h. The nucleus was stained with 4,6-diamino-2-phenyl indole (DAPI)
(Invitrogen, USA) for 10 min. After staining, the devices were washed with PBS 2-3 times and
imaged using fluorescence microscopy (Olympus IX71). The fluorescence intensities of collagen
I and collagen II were determined from the fluorescence photographs (N=10) using Image-Pro
Plus 6.0 software to obtain relative fluorescence intensity (RFI) values.
2.5. Cell staining
After 5 days of perfusion, the cells were stained with oil red O and alkaline phosphatase
(ALP) to determine adipogenic differentiation and osteogenic differentiation, respectively. Briefly,
the cells were fixed with 4% paraformaldehyde and then stained with Fast Blue RR/naphthol
(Sigma, USA) to visualize ALP. After washing with PBS, the cells were stained with 30 mg/ml
oil red O (Sigma, USA) in 60% isopropanol to visualize the lipid droplets and then rinsed in PBS.
2.6. Statistics and analysis
All experiments were performed at least in triplicate with different batches of devices.
Differences among three groups were analyzed using one-way ANOVA. Comparisons of two
Page 9 of 35
Stem
Cel
ls a
nd D
evel
opm
ent
Mes
ench
ymal
Ste
m C
ell a
nd C
hond
rocy
te F
ates
in a
Mul
ti-Sh
ear
Mic
rode
vice
are
Reg
ulat
ed b
y Y
es-A
ssoc
iate
d Pr
otei
n (d
oi: 1
0.10
89/s
cd.2
012.
0685
)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

10
10
groups were performed by using the two-sample t-test. P<0.05 was considered statistically
significant.
3. Results
3.1. Computational simulation of the fluid dynamics in the microchambers
In this study, a microdevice was designed based on the principles of the Electric Circuit
Analogy. This device was composed of one inlet, one outlet, resistance channels and four cell
culture chambers. The flow of fluid through the microdevice was typically driven by a peristaltic
pump. Thus, the pressure-driven fluid flow through the chamber could be defined as steady-state
flow and modeled as Poiseuille flow. The fluid shear stress on the cells could be assumed to be
equal to the wall shear stress at the bottom of chamber. When the perfusion flow rate of 30
µl/min was applied to the microdevice, different levels of fluid flow stimulus could be generated.
The central area of these chambers exhibited a uniform distribution for the wall shear stress (Fig.
1B). The results demonstrated that the majority of the bottom of the chamber experienced
uniform shear stress except the areas near the corners, the inlet, the outlet, and the side walls. To
quantify the local distribution of shear stress in detail, we computed the wall shear stress along
the vertical and horizontal center axes of each microchamber. This observation was further
confirmed by the wall shear stress curves along the vertical center axis (Fig. 1C) and horizontal
center axis (Fig. 1D). The average bottom wall shear stresses in Chamber 1 to Chamber 4 in these
Page 10 of 35
Stem
Cel
ls a
nd D
evel
opm
ent
Mes
ench
ymal
Ste
m C
ell a
nd C
hond
rocy
te F
ates
in a
Mul
ti-Sh
ear
Mic
rode
vice
are
Reg
ulat
ed b
y Y
es-A
ssoc
iate
d Pr
otei
n (d
oi: 1
0.10
89/s
cd.2
012.
0685
)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

11
11
uniform regions were 1.089 dyne/cm2, 0.231 dyne/cm
2, 0.055 dyne/cm
2, and 0.009 dyne/cm
2,
respectively.
3.2. The proliferation rates of MSCs and chondrocytes exposed to different shear stresses
To examine the effects of different shear stresses on cell proliferation, the proliferation rates
of MSCs and chondrocytes were analyzed by counting the number of cells present in the
chambers under each condition. As shown in Fig. 2A, the proliferation rate of MSCs increased
significantly in response to shear stress at 0.231 and 1.089 dyne/cm2 after 24 h and 48 h of
perfusion compared to the minimum stimulus (0.009 dyne/cm2). For chondrocytes, the
proliferation rate increased significantly in response to shear stress at 1.089 dyne/cm2 after 24 h
and shear stress at 0.231 and 1.089 dyne/cm2 after 48 h. These findings suggested that the
proliferation rates of both MSCs and chondrocytes increased with an increasing magnitude of the
fluid shear stress.
3.3. The changes in YAP expression in response to different shear stresses
To investigate the effects of different fluid shear stresses on YAP expression in MSCs and
chondrocytes, the cells in the microdevices were stained for YAP after 5 days of perfusion for
MSCs and 2 days of perfusion for chondrocytes. As shown in Fig. 2B, the distribution of YAP in
MSCs was mainly in the nucleus with low level in the cytoplasm. Although the increase in fluid
shear stress did not alter YAP localization, the level of YAP in the nucleus tended to increase (Fig.
Page 11 of 35
Stem
Cel
ls a
nd D
evel
opm
ent
Mes
ench
ymal
Ste
m C
ell a
nd C
hond
rocy
te F
ates
in a
Mul
ti-Sh
ear
Mic
rode
vice
are
Reg
ulat
ed b
y Y
es-A
ssoc
iate
d Pr
otei
n (d
oi: 1
0.10
89/s
cd.2
012.
0685
)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

12
12
2B). In addition, we found that in primary chondrocytes YAP was predominantly cytoplasmic at
an extremely low level of shear stress. However, increased nuclear YAP accumulation was
observed with an increasing magnitude of stimulation. At higher stimulus levels (0.231 and 1.089
dyne/cm2), the nuclear localization of YAP was markedly increased (Fig. 2B). Conversely, MSCs
and chondrocytes treated with CytoD developed a more rounded morphology and exhibited YAP
diffusion into the cytoplasm after exposure to the fluid flow. The quantification of nuclear YAP in
MSCs and chondrocytes confirmed these findings (Fig. 2C). The percentage of nuclear YAP in
these two types of cells was significantly higher under shear stresses of 0.231 and 1.089 dyne/cm2
than under the lowest level of shear stress (0.009 dyne/cm2). CytoD treatment was associated
with a significant decrease in nuclear YAP relative to the lowest stimulus level.
3.4. The fate of MSCs changes under different shear stress conditions
To evaluate the effects of different fluid shear stresses on MSC differentiation, cells in the
microdevices were stained for ALP and intracellular lipid droplets after 5 days of perfusion. As
shown in Fig. 3A, cells exposed to an extremely low level of shear stress tended to undergo
adipogenic differentiation, but with an increasing magnitude of shear stress, increased cells
underwent osteogenic differentiation. Especially at the higher shear stress level of 1.089
dyne/cm2, cells tended to become committed to the osteogenic lineage. The ratio of osteogenic
commitment to adipogenic commitment reflected a significant mechanical effect of different fluid
shear stresses (Fig. 3B). However, CytoD-treated cells exposed to the fluid flow stimulus
Page 12 of 35
Stem
Cel
ls a
nd D
evel
opm
ent
Mes
ench
ymal
Ste
m C
ell a
nd C
hond
rocy
te F
ates
in a
Mul
ti-Sh
ear
Mic
rode
vice
are
Reg
ulat
ed b
y Y
es-A
ssoc
iate
d Pr
otei
n (d
oi: 1
0.10
89/s
cd.2
012.
0685
)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

13
13
appeared to lose the ability to undergo osteogenic differentiation regardless of the level of flow.
Interestingly, the pattern of adipogenic and osteogenic differentiation were consistent with the
pattern of YAP distribution in the cytoplasm and nucleus.
Next, we examined the expression of more lineage-specific regulators such as PPAR,
Runx2, and Sox9 in MSCs. As shown in Fig. 3C, PPAR that promotes adipocyte differentiation
in MSCs was predominantly localized in the nucleus, but a gradual decline in PPAR expression
was observed with an increasing magnitude of flow stimulus. In contrast, Runx2, a key regulator
of osteogenic differentiation, displayed a gradually increasing expression in the cells, especially
in the nucleus with an increasing magnitude of flow stimulus. However, CytoD-treated cells
displayed high expression level of PPAR but low expression level of Runx2 regardless of flow
stimulus. For Sox9, an essential regulator of chondrogenesis, no obvious expression was detected
under different fluid shear stress even when treated with CytoD. Taken together, we found that
the changes in the expression of PPAR and Runx2 were in accordance with the variation in lipid
droplets and ALP.
3.5. The phenotypic variations of chondrocytes in response to different shear stresses
After 2 days of fluid flow stimulation, the phenotypic changes in primary chondrocytes were
investigated by immunofluorescence staining for collagen I and collagen II. As shown in Fig. 4A,
primary chondrocytes were nearly negative for collagen I and positive for collagen II when
exposed to extremely low level of fluid flow. However, with an increasing magnitude of shear
Page 13 of 35
Stem
Cel
ls a
nd D
evel
opm
ent
Mes
ench
ymal
Ste
m C
ell a
nd C
hond
rocy
te F
ates
in a
Mul
ti-Sh
ear
Mic
rode
vice
are
Reg
ulat
ed b
y Y
es-A
ssoc
iate
d Pr
otei
n (d
oi: 1
0.10
89/s
cd.2
012.
0685
)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

14
14
stress, the cells began to synthesize collagen I, whereas the expression of collagen II diminished.
When the cells were treated with CytoD before the exposure to the fluid flow stimulus, the
hyaline cartilage phenotype of the cells was rescued, with low expression of collagen I. To further
quantify the variation in the expression levels of collagen II and collagen I, the relative
fluorescence intensities were plotted (Fig. 4B and C). We found that from 0.055 to 1.089
dyne/cm2, RFI values were significantly higher for collagen I and lower for collagen II than the
values at the minimum level of stimulus. As shown in Fig. 2 and Fig. 4, we found that
chondrocyte dedifferentiation was concomitant with YAP accumulation in the nucleus, indicating
that mechanical cue of fluid flow leads to increased YAP expression in dedifferentiated
chondrocytes.
4. Discussion
YAP transcriptional coactivator has emerged as a key mediator of the regulation of cell
behaviors such as proliferation, survival, apoptosis and differentiation [16,17,21]. In this study,
we evaluated the change in YAP expression and the consequent biological responses to
flow-induced mechanical signals of different magnitudes. Our results suggest that YAP may
mediate the effects of fluid flow-induced shear stress on the fates of MSCs and chondrocytes.
We characterized the changes in YAP expression along with the fates of MSCs and
chondrocytes in response to different levels of flow stimulus. Our data showed that YAP
Page 14 of 35
Stem
Cel
ls a
nd D
evel
opm
ent
Mes
ench
ymal
Ste
m C
ell a
nd C
hond
rocy
te F
ates
in a
Mul
ti-Sh
ear
Mic
rode
vice
are
Reg
ulat
ed b
y Y
es-A
ssoc
iate
d Pr
otei
n (d
oi: 1
0.10
89/s
cd.2
012.
0685
)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

15
15
expression in MSCs was increased with an increasing magnitude of shear stress. Furthermore, we
demonstrated that in chondrocytes YAP was translocated into the nucleus in response to fluid
flow stimulus. The transduction of local mechanostimuli into biochemical signals occurs through
several signaling pathways. Recently, the transcriptional regulators YAP/TAZ have been shown
to act as not only sensors but also mediators of physical signals, shuttling between the cytoplasm
and the nucleus [22]. As the downstream effectors of mechanotransduction, YAP function
depends on the tension of the actomyosin cytoskeleton and Rho GTPase activity because proper
cytoskeleton tension is necessary to maintain YAP transcriptional activity. Moreover, increasing
evidence has shown that shear stress can cause cytoskeleton reorganization in mechanically
sensitive cells. Thus, the flow-dependent changes in YAP expression are likely due to an increase
in cytoskeleton tension. When the flow shear force is transmitted to the intracellular space, the
cells re-organize the cytoskeleton to gradually increase the strength of attachment with an
increasing magnitude of stimulation. Meanwhile, cytoskeletal cues can mediate YAP nuclear
localization to ultimately affect gene expression. It is well known that the actin cytoskeleton is
disrupted when cells are treated with CytoD, thereby abrogating the ability of cells to sense
external mechanical cues. In this study, the treatment of cells with CytoD resulted in YAP
downregulation, and the flow-dependent response was abolished, further suggesting that shear
flow plays a role in the regulation of YAP and the integrity of the actomyosin cytoskeleton is
vital for shear stress-based regulation of YAP expression. Additionally, the effects of fluid shear
stress on cells involve other pathways that may communicate with YAP or Hippo pathway. The
Page 15 of 35
Stem
Cel
ls a
nd D
evel
opm
ent
Mes
ench
ymal
Ste
m C
ell a
nd C
hond
rocy
te F
ates
in a
Mul
ti-Sh
ear
Mic
rode
vice
are
Reg
ulat
ed b
y Y
es-A
ssoc
iate
d Pr
otei
n (d
oi: 1
0.10
89/s
cd.2
012.
0685
)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

16
16
cell-cell contact or a high cell density inhibits YAP activity by activating the Hippo pathway [20].
To avoid such an influence, a low seeding density was employed in our microdevice experiments.
We assumed that this may limit the activation of Hippo pathway and better investigate the effects
of fluid shear stress on YAP. Consequently, we found that an increasing magnitude of fluid shear
stress could promote cell proliferation and concomitantly enhance YAP expression in the nuclei.
Shear stress-induced proliferation of osteocytes, MSCs, as well as chondrocytes has been
reported [32-34], involving several mechanisms such as ERK pathway, MAP kinase pathway,
NO/cGMP/PKG and calcium signaling [35]. In addition,mechanical signaling through the
cytoskeleton linkage between focal adhesion and regulators of cellular contractility contribute to
the regulation of cell proliferation [36]. YAP has been shown to act as a transcriptional
co-activator of TEAD transcription factors to promote cell proliferation and survival in many
tissues [37]. Here, we propose that fluid flow upregulates YAP, which then promotes cell
proliferation by binding to the TEAD family of transcription factors. Therefore, we conclude that
nuclear YAP expression is a key regulator that has a correlation with mechanical signal-induced
proliferation. Further investigations are required to determine whether these known pathways
cooperate with the Hippo/YAP pathway to promote cell proliferation.
Control of the balance between adipogenesis and osteogenesis during MSC differentiation is
necessary to maintain bone homeostasis. Stem cells are highly sensitive to mechanical cues and
can convert mechanical stimuli into biochemical signals via mechanotransduction systems. In this
study, we characterized the effects of distinct levels of flow stimulus on the regulation of MSC
Page 16 of 35
Stem
Cel
ls a
nd D
evel
opm
ent
Mes
ench
ymal
Ste
m C
ell a
nd C
hond
rocy
te F
ates
in a
Mul
ti-Sh
ear
Mic
rode
vice
are
Reg
ulat
ed b
y Y
es-A
ssoc
iate
d Pr
otei
n (d
oi: 1
0.10
89/s
cd.2
012.
0685
)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

17
17
fate. We found that the expression level of YAP is correlated with the intensity of the stimulus
experienced by the cells and the fates of the MSCs. Increased YAP expression is in accordance
with the increase in the expression of Runx2 and ALP, indicating that increased YAP expression
may contribute to osteogenesis in MSCs. Conversely, decreased YAP expression is concomitant
with the increase in lipid droplets and the expression of PPAR, which indicate adipogenesis in
MSCs. It has been reported that YAP interacts with Runx2 and PPARγ to regulate
adipocyte/osteocyte gene expression [38]. Moreover, YAP modulated Runx2 and PPARγ activity
by altering the activation of Wnt/β-catenin signaling [39]. It is therefore possible that fluid shear
stress regulates the choice between adipogenesis and osteogenesis in MSCs by controlling the
expression of YAP. Growing evidence has shown that the enhancement of mechanical or
cytoskeletal cues would increase osteogenic differentiation and decrease adipogenic
differentiation. In addition, cell density is an important factor that regulates the adipogenic and
osteogenic differentiation of MSCs [12,40].
In a multilineage differentiation experiment, MSCs differentiated into adipocytes in
response to induction medium containing 10-6
M dexamethasone but differentiated into osteocytes
in response to induction medium containing 10-7
M dexamethasone [41]. In this study, the mixed
differentiation medium contained a relatively high concentration of dexamethasone because the
two types of induction medium were mixed in equal proportions, thus causing an increase in the
proportion of cells undergoing adipogenic differentiation. However, with increases in the shear
stress magnitude, the proportions of cells undergoing adipogenic and osteogenic differentiation
Page 17 of 35
Stem
Cel
ls a
nd D
evel
opm
ent
Mes
ench
ymal
Ste
m C
ell a
nd C
hond
rocy
te F
ates
in a
Mul
ti-Sh
ear
Mic
rode
vice
are
Reg
ulat
ed b
y Y
es-A
ssoc
iate
d Pr
otei
n (d
oi: 1
0.10
89/s
cd.2
012.
0685
)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

18
18
were reversed due to the upregulation of YAP expression, leading to an increase in osteogenesis
and a decline in adipogenesis. Thus, by analyzing YAP expression level, we can preliminarily
estimate the lineage to which the MSCs have committed or will commit and determine which
flow stimulus condition is appropriate for target differentiation.
Chondrocytes differentiate from mesenchymal cells during development and are apt to
dedifferentiate in in vitro cultures [42]. Excess fluid flow stimulus has been shown to be
detrimental to the maintenance of chondrocyte function [43-45]. We first investigated the
expression of YAP in response to distinct levels of fluid flow in primary chondrocytes and then
monitored the changes in the chondrocyte phenotype that are associated with the changes in YAP
localization. We found that increasing flow stimulus resulted in nuclear YAP accumulation and
the loss of chondrocyte properties. When primary chondrocytes were exposed to extremely low
stimulus, no phenotypic variation was observed compared to the higher stimulus, suggesting that
such a condition is not enough to induce chondrocyte dedifferentiation. Instead, this condition
apparently mimicked the flow of interstitial fluid in the cartilage and played an important role in
nutrient transport. A previous study confirmed that YAP expression was suppressed by treatment
with CytoD [19]. In our study, after the disruption of the cytoskeleton with CytoD, the cells
undergoing YAP cytoplasmic translocation failed to respond to the flow stimulation but better
maintained their phenotype, suggesting that the process of regulating the chondrocyte phenotype
is related to YAP expression. Taken together, our findings indicate that the maintenance of the
phenotype of primary chondrocytes is associated with the exclusion of YAP from the nucleus,
Page 18 of 35
Stem
Cel
ls a
nd D
evel
opm
ent
Mes
ench
ymal
Ste
m C
ell a
nd C
hond
rocy
te F
ates
in a
Mul
ti-Sh
ear
Mic
rode
vice
are
Reg
ulat
ed b
y Y
es-A
ssoc
iate
d Pr
otei
n (d
oi: 1
0.10
89/s
cd.2
012.
0685
)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

19
19
which may involve mechanical and cytoskeletal cues or the Hippo pathway. For instance, both
pellet or monolayer culture with high cell density and soft substrate with low cytoskeleton
tension can better maintain chondrocyte phenotype in vitro. Such a high cell density induces the
activation of Hippo signaling to suppress the expression of YAP [20], and low cytoskeleton
tension also causes YAP inactivation [18]. The detailed biochemical mechanisms underlying
these phenomena await further characterization. Notably, based on the localization of YAP in
chondrocytes, we can estimate the stimulus intensity experienced by cells and evaluate whether
chondrocytes would dedifferentiate when exposed to a particular magnitudes of shear flow.
Microtechnology can be used to regulate biological stimuli at the cellular and subcellular
scales, enabling us to decipher the roles of specific cues and mechanisms in the cells [46]. In our
study, we combined the electric circuit theory with CFD analysis to design a lab-on-a-chip and
thus solve the problems related to complex microfluidic network-based devices. Using a physical
analogy, the systematic networking of the microchannels from the inlet to outlet ports could
readily be used to establish relatively simple model. Thus, multiple shear stresses could be
obtained through the different inlet channel resistances of the different chambers. Furthermore,
this microdevice allowed the control of multiple shear stress values in a particular range by
modifying the inlet resistance of each microchamber and the inlet flow rate. In addition, this
microdevice provided a simple platform to rapidly screen cellular responses to different levels of
flow stimulus and minimize the sample volume to the micron scale.
Page 19 of 35
Stem
Cel
ls a
nd D
evel
opm
ent
Mes
ench
ymal
Ste
m C
ell a
nd C
hond
rocy
te F
ates
in a
Mul
ti-Sh
ear
Mic
rode
vice
are
Reg
ulat
ed b
y Y
es-A
ssoc
iate
d Pr
otei
n (d
oi: 1
0.10
89/s
cd.2
012.
0685
)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

20
20
5. Conclusions
Using microfluidic techniques, we developed a novel integrated microdevice that
simultaneously produces multiple fluid shear stresses comparable to or stronger than that of
interstitial flow. Using this device, the roles of the fluid flow stimulus in the regulation of YAP
expression and the fates of MSC and chondrocytes were investigated. Our results suggest that
YAP expression in MSCs and chondrocytes is regulated by fluid shear stress and YAP mediates
the fate determination of MSCs and chondrocytes in response to fluid flow stimulus. Therefore,
the modulation of YAP expression could be exploited to manipulate stem cell fate and increase
the regenerative potential of terminally differentiated organs with poor intrinsic regenerative
capacity.
Acknowledgements
This work was supported by the National Nature Science Foundation of China (No.
81171464 and No. 81270052). These authors have no conflict of interest.
Author Disclosure Statement
Page 20 of 35
Stem
Cel
ls a
nd D
evel
opm
ent
Mes
ench
ymal
Ste
m C
ell a
nd C
hond
rocy
te F
ates
in a
Mul
ti-Sh
ear
Mic
rode
vice
are
Reg
ulat
ed b
y Y
es-A
ssoc
iate
d Pr
otei
n (d
oi: 1
0.10
89/s
cd.2
012.
0685
)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

21
21
No competing financial interests exist in this study.
References:
1. Hoshiba T, N Kawazoe and G Chen. (2012). The balance of osteogenic and adipogenic
differentiation in human mesenchymal stem cells by matrices that mimic stepwise tissue
development. Biomaterials 33:2025-31.
2. Guilak F, DM Cohen, BT Estes, JM Gimble, W Liedtke and CS Chen. (2009). Control of
stem cell fate by physical interactions with the extracellular matrix. Cell Stem Cell
5:17-26.
3. Sun Y, CS Chen and J Fu. (2012). Forcing stem cells to behave: a biophysical perspective
of the cellular microenvironment. Annu Rev Biophys 41:519-42.
4. Rutkowski JM and MA Swartz. (2007). A driving force for change: interstitial flow as a
morphoregulator. Trends Cell Biol 17:44-50.
5. Jaalouk DE and J Lammerding. (2009). Mechanotransduction gone awry. Nature Reviews
Molecular Cell Biology 10:63-73.
6. Swartz MA and ME Fleury. (2007). Interstitial flow and its effects in soft tissues. Annu
Rev Biomed Eng 9:229-56.
7. Arnsdorf EJ, P Tummala, RY Kwon and CR Jacobs. (2009). Mechanically induced
osteogenic differentiation--the role of RhoA, ROCKII and cytoskeletal dynamics. Journal
Page 21 of 35
Stem
Cel
ls a
nd D
evel
opm
ent
Mes
ench
ymal
Ste
m C
ell a
nd C
hond
rocy
te F
ates
in a
Mul
ti-Sh
ear
Mic
rode
vice
are
Reg
ulat
ed b
y Y
es-A
ssoc
iate
d Pr
otei
n (d
oi: 1
0.10
89/s
cd.2
012.
0685
)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

22
22
of Cell Science 122:546-53.
8. Mammoto A and DE Ingber. (2009). Cytoskeletal control of growth and cell fate
switching. Current Opinion in Cell Biology 21:864-70.
9. Vogel V and M Sheetz. (2006). Local force and geometry sensing regulate cell functions.
Nature Reviews Molecular Cell Biology 7:265-75.
10. Treiser MD, EH Yang, S Gordonov, DM Cohen, IP Androulakis, J Kohn, CS Chen and PV
Moghe. (2010). Cytoskeleton-based forecasting of stem cell lineage fates. Proc Natl Acad
Sci U S A 107:610-5.
11. Kilian KA, B Bugarija, BT Lahn and M Mrksich. (2010). Geometric cues for directing the
differentiation of mesenchymal stem cells. Proc Natl Acad Sci U S A 107:4872-7.
12. McBeath R, DM Pirone, CM Nelson, K Bhadriraju and CS Chen. (2004). Cell shape,
cytoskeletal tension, and RhoA regulate stem cell lineage commitment. Dev Cell
6:483-95.
13. Allen JL, ME Cooke and T Alliston. (2012). ECM stiffness primes the TGFbeta pathway
to promote chondrocyte differentiation. Mol Biol Cell 23:3731-42.
14. Schuh E, J Kramer, J Rohwedel, H Notbohm, R Muller, T Gutsmann and N Rotter. (2010).
Effect of matrix elasticity on the maintenance of the chondrogenic phenotype. Tissue
Engineering Part A 16:1281-90.
15. Halder G and RL Johnson. (2011). Hippo signaling: growth control and beyond.
Development 138:9-22.
Page 22 of 35
Stem
Cel
ls a
nd D
evel
opm
ent
Mes
ench
ymal
Ste
m C
ell a
nd C
hond
rocy
te F
ates
in a
Mul
ti-Sh
ear
Mic
rode
vice
are
Reg
ulat
ed b
y Y
es-A
ssoc
iate
d Pr
otei
n (d
oi: 1
0.10
89/s
cd.2
012.
0685
)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

23
23
16. Lian I, J Kim, H Okazawa, J Zhao, B Zhao, J Yu, A Chinnaiyan, MA Israel, LS Goldstein,
R Abujarour, S Ding and KL Guan. (2010). The role of YAP transcription coactivator in
regulating stem cell self-renewal and differentiation. Genes Dev 24:1106-18.
17. Zhao B, K Tumaneng and KL Guan. (2011). The Hippo pathway in organ size control,
tissue regeneration and stem cell self-renewal. Nature Cell Biology 13:877-83.
18. Dupont S, L Morsut, M Aragona, E Enzo, S Giulitti, M Cordenonsi, F Zanconato, J Le
Digabel, M Forcato, S Bicciato, N Elvassore and S Piccolo. (2011). Role of YAP/TAZ in
mechanotransduction. Nature 474:179-83.
19. Wada K, K Itoga, T Okano, S Yonemura and H Sasaki. (2011). Hippo pathway regulation
by cell morphology and stress fibers. Development 138:3907-14.
20. Zhao B, X Wei, W Li, RS Udan, Q Yang, J Kim, J Xie, T Ikenoue, J Yu, L Li, P Zheng, K
Ye, A Chinnaiyan, G Halder, ZC Lai and KL Guan. (2007). Inactivation of YAP
oncoprotein by the Hippo pathway is involved in cell contact inhibition and tissue growth
control. Genes Dev 21:2747-61.
21. Zhao B, L Li, L Wang, CY Wang, J Yu and KL Guan. (2012). Cell detachment activates
the Hippo pathway via cytoskeleton reorganization to induce anoikis. Genes Dev
26:54-68.
22. Halder G, S Dupont and S Piccolo. (2012). Transduction of mechanical and cytoskeletal
cues by YAP and TAZ. Nature Reviews Molecular Cell Biology 13:591-600.
23. Zhang H, M Deo, RC Thompson, MD Uhler and DL Turner. (2012). Negative regulation
Page 23 of 35
Stem
Cel
ls a
nd D
evel
opm
ent
Mes
ench
ymal
Ste
m C
ell a
nd C
hond
rocy
te F
ates
in a
Mul
ti-Sh
ear
Mic
rode
vice
are
Reg
ulat
ed b
y Y
es-A
ssoc
iate
d Pr
otei
n (d
oi: 1
0.10
89/s
cd.2
012.
0685
)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

24
24
of Yap during neuronal differentiation. Developmental Biology 361:103-15.
24. Hong JH, ES Hwang, MT McManus, A Amsterdam, Y Tian, R Kalmukova, E Mueller, T
Benjamin, BM Spiegelman, PA Sharp, N Hopkins and MB Yaffe. (2005). TAZ, a
transcriptional modulator of mesenchymal stem cell differentiation. Science 309:1074-8.
25. Moraes C, Y Sun and CA Simmons. (2011). (Micro)managing the mechanical
microenvironment. Integr Biol (Camb) 3:959-71.
26. Ghafar-Zadeh E, JR Waldeisen and LP Lee. (2011). Engineered approaches to the stem
cell microenvironment for cardiac tissue regeneration. Lab Chip 11:3031-48.
27. McDonald JC, DC Duffy, JR Anderson, DT Chiu, H Wu, OJ Schueller and GM
Whitesides. (2000). Fabrication of microfluidic systems in poly(dimethylsiloxane).
Electrophoresis 21:27-40.
28. Oh KW, K Lee, B Ahn and EP Furlani. (2012). Design of pressure-driven microfluidic
networks using electric circuit analogy. Lab Chip 12:515-545.
29. Beebe DJ, GA Mensing and GM Walker. (2002). Physics and applications of
microfluidics in biology. Annu Rev Biomed Eng 4:261-286.
30. Bruus H. (2011). Acoustofluidics 1: Governing equations in microfluidics. Lab Chip
11:3742-3751.
31. Gosset M, F Berenbaum, S Thirion and C Jacques. (2008). Primary culture and
phenotyping of murine chondrocytes. Nature Protocols 3:1253-60.
32. Kapur S, DJ Baylink and KH Lau. (2003). Fluid flow shear stress stimulates human
Page 24 of 35
Stem
Cel
ls a
nd D
evel
opm
ent
Mes
ench
ymal
Ste
m C
ell a
nd C
hond
rocy
te F
ates
in a
Mul
ti-Sh
ear
Mic
rode
vice
are
Reg
ulat
ed b
y Y
es-A
ssoc
iate
d Pr
otei
n (d
oi: 1
0.10
89/s
cd.2
012.
0685
)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

25
25
osteoblast proliferation and differentiation through multiple interacting and competing
signal transduction pathways. Bone 32:241-51.
33. Riddle RC, AF Taylor, DC Genetos and HJ Donahue. (2006). MAP kinase and calcium
signaling mediate fluid flow-induced human mesenchymal stem cell proliferation. Am J
Physiol Cell Physiol 290:C776-84.
34. Malaviya P and RM Nerem. (2002). Fluid-induced shear stress stimulates chondrocyte
proliferation partially mediated via TGF-beta1. Tissue Engineering 8:581-90.
35. Liu L, W Yuan and J Wang. (2010). Mechanisms for osteogenic differentiation of human
mesenchymal stem cells induced by fluid shear stress. Biomechanics and Modeling in
Mechanobiology 9:659-70.
36. Provenzano PP and PJ Keely. (2011). Mechanical signaling through the cytoskeleton
regulates cell proliferation by coordinated focal adhesion and Rho GTPase signaling.
Journal of Cell Science 124:1195-205.
37. Zhang L, F Ren, Q Zhang, Y Chen, B Wang and J Jiang. (2008). The TEAD/TEF family
of transcription factor Scalloped mediates Hippo signaling in organ size control. Dev Cell
14:377-87.
38. Hiemer SE and X Varelas. (2012). Stem cell regulation by the Hippo pathway. Biochimica
et Biophysica Acta.
39. Imajo M, K Miyatake, A Iimura, A Miyamoto and E Nishida. (2012). A molecular
mechanism that links Hippo signalling to the inhibition of Wnt/beta-catenin signalling.
Page 25 of 35
Stem
Cel
ls a
nd D
evel
opm
ent
Mes
ench
ymal
Ste
m C
ell a
nd C
hond
rocy
te F
ates
in a
Mul
ti-Sh
ear
Mic
rode
vice
are
Reg
ulat
ed b
y Y
es-A
ssoc
iate
d Pr
otei
n (d
oi: 1
0.10
89/s
cd.2
012.
0685
)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

26
26
EMBO Journal 31:1109-22.
40. Seo CH, K Furukawa, K Montagne, H Jeong and T Ushida. (2011). The effect of substrate
microtopography on focal adhesion maturation and actin organization via the
RhoA/ROCK pathway. Biomaterials 32:9568-75.
41. Zhu H, ZK Guo, XX Jiang, H Li, XY Wang, HY Yao, Y Zhang and N Mao. (2010). A
protocol for isolation and culture of mesenchymal stem cells from mouse compact bone.
Nature Protocols 5:550-60.
42. Yoon YM, SJ Kim, CD Oh, JW Ju, WK Song, YJ Yoo, TL Huh and JS Chun. (2002).
Maintenance of differentiated phenotype of articular chondrocytes by protein kinase C
and extracellular signal-regulated protein kinase. Journal of Biological Chemistry
277:8412-20.
43. Lee MS, MC Trindade, T Ikenoue, SB Goodman, DJ Schurman and RL Smith. (2003).
Regulation of nitric oxide and bcl-2 expression by shear stress in human osteoarthritic
chondrocytes in vitro. Journal of Cellular Biochemistry 90:80-6.
44. Healy ZR, NH Lee, X Gao, MB Goldring, P Talalay, TW Kensler and K Konstantopoulos.
(2005). Divergent responses of chondrocytes and endothelial cells to shear stress:
cross-talk among COX-2, the phase 2 response, and apoptosis. Proc Natl Acad Sci U S A
102:14010-5.
45. Zhu F, P Wang, NH Lee, MB Goldring and K Konstantopoulos. (2010). Prolonged
application of high fluid shear to chondrocytes recapitulates gene expression profiles
Page 26 of 35
Stem
Cel
ls a
nd D
evel
opm
ent
Mes
ench
ymal
Ste
m C
ell a
nd C
hond
rocy
te F
ates
in a
Mul
ti-Sh
ear
Mic
rode
vice
are
Reg
ulat
ed b
y Y
es-A
ssoc
iate
d Pr
otei
n (d
oi: 1
0.10
89/s
cd.2
012.
0685
)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

27
27
associated with osteoarthritis. PLoS One 5:e15174.
46. Park JY, S Takayama and SH Lee. (2010). Regulating microenvironmental stimuli for
stem cells and cancer cells using microsystems. Integr Biol (Camb) 2:229-40.
Figure Legends
Page 27 of 35
Stem
Cel
ls a
nd D
evel
opm
ent
Mes
ench
ymal
Ste
m C
ell a
nd C
hond
rocy
te F
ates
in a
Mul
ti-Sh
ear
Mic
rode
vice
are
Reg
ulat
ed b
y Y
es-A
ssoc
iate
d Pr
otei
n (d
oi: 1
0.10
89/s
cd.2
012.
0685
)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

28
28
Page 28 of 35
Stem
Cel
ls a
nd D
evel
opm
ent
Mes
ench
ymal
Ste
m C
ell a
nd C
hond
rocy
te F
ates
in a
Mul
ti-Sh
ear
Mic
rode
vice
are
Reg
ulat
ed b
y Y
es-A
ssoc
iate
d Pr
otei
n (d
oi: 1
0.10
89/s
cd.2
012.
0685
)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

29
29
Fig. 1. A: Schematic diagram of the multi-shear microfluidic device and a photograph of a
prototype microdevice. In the schematic diagram, Q is the flow rate in the microchamber, w is the
width of the microchamber and h is the height of the microchamber. B-D: The numerical
simulation results for the cell culture chambers based on CFD analysis. B: Contours of the wall
shear stress distribution in the microchambers of the device. C: Variation in the wall shear stress
along the vertical center axis of each microchamber. D: Variation in the wall shear stress along
the horizontal center axis of each microchamber.
Page 29 of 35
Stem
Cel
ls a
nd D
evel
opm
ent
Mes
ench
ymal
Ste
m C
ell a
nd C
hond
rocy
te F
ates
in a
Mul
ti-Sh
ear
Mic
rode
vice
are
Reg
ulat
ed b
y Y
es-A
ssoc
iate
d Pr
otei
n (d
oi: 1
0.10
89/s
cd.2
012.
0685
)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

30
30
Page 30 of 35
Stem
Cel
ls a
nd D
evel
opm
ent
Mes
ench
ymal
Ste
m C
ell a
nd C
hond
rocy
te F
ates
in a
Mul
ti-Sh
ear
Mic
rode
vice
are
Reg
ulat
ed b
y Y
es-A
ssoc
iate
d Pr
otei
n (d
oi: 1
0.10
89/s
cd.2
012.
0685
)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

31
31
Fig. 2. A: The proliferation of MSCs and chondrocytes in the presence of different shear stresses.
(i): Histogram of MSC proliferation rates after 24 and 48 h. (ii): Histogram of chondrocyte
proliferation rates after 24 and 48 h. The values are the mean ± SD. * and # represent 24 and 48
hours, respectively. #
and *, P<0.05; ##
and **, P<0.01 versus 0.009 dyne/cm2. B-C: The changes
in YAP expression in cells subjected to different shear stresses. MSCs and chondrocytes were
immunostained for YAP (green) after 5 and 2 days of perfusion, respectively. B: The YAP
distribution in MSCs and chondrocytes subjected to different shear stresses. Scale bar: 20 μm. C:
Graphs indicating the percentage of cells with nuclear YAP under different conditions. The values
are the mean ± SD. * and # represent MSCs and chondrocytes, respectively.
# and *, P<0.05;
###
and ***, P<0.001 versus 0.009 dyne/cm2.
Page 31 of 35
Stem
Cel
ls a
nd D
evel
opm
ent
Mes
ench
ymal
Ste
m C
ell a
nd C
hond
rocy
te F
ates
in a
Mul
ti-Sh
ear
Mic
rode
vice
are
Reg
ulat
ed b
y Y
es-A
ssoc
iate
d Pr
otei
n (d
oi: 1
0.10
89/s
cd.2
012.
0685
)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.
32
32
Page 32 of 35
Stem
Cel
ls a
nd D
evel
opm
ent
Mes
ench
ymal
Ste
m C
ell a
nd C
hond
rocy
te F
ates
in a
Mul
ti-Sh
ear
Mic
rode
vice
are
Reg
ulat
ed b
y Y
es-A
ssoc
iate
d Pr
otei
n (d
oi: 1
0.10
89/s
cd.2
012.
0685
)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

33
33
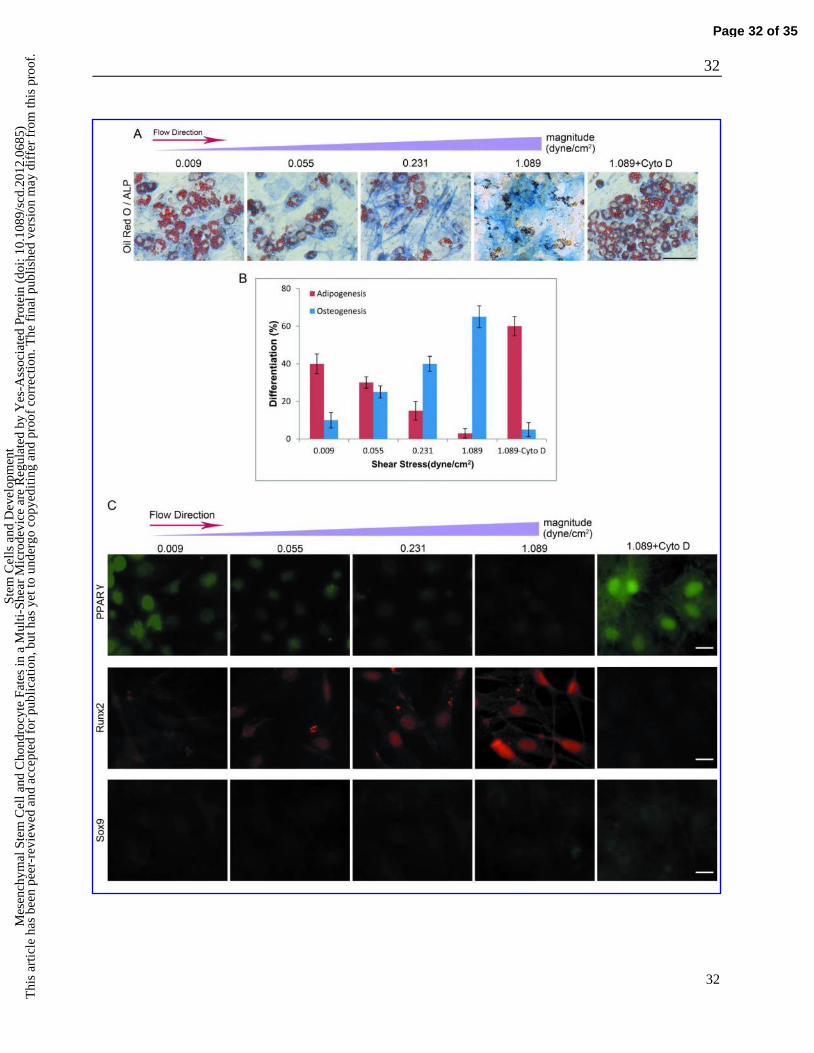
Fig. 3. MSC commitment varies with changes in the flow stimulus. A: Brightfield images of the
adipogenic and osteogenic differentiation of MSCs after 5 days of exposure to different induction
conditions. The cells were stained with Fast Blue RR/naphthol and oil Red O to visualize ALP
(blue) and lipid droplets (red) as indicators of osteogenesis and adipogenesis, respectively. Scale
bar: 100 μm. B: Percentage of osteogenic and adipogenic differentiation of MSCs after exposure
to the mixed induction medium for 5 days. The error bars are mean ± SD. C: Comparison of the
expressions of lineage-specific regulators (PPAR, Runx2 and Sox9) in MSCs under different
induction conditions. Fluorescence images of PPAR, Runx2 and Sox9 staining in cells exposed
to different conditions for 5 days. Scale bar: 20 μm.
Page 33 of 35
Stem
Cel
ls a
nd D
evel
opm
ent
Mes
ench
ymal
Ste
m C
ell a
nd C
hond
rocy
te F
ates
in a
Mul
ti-Sh
ear
Mic
rode
vice
are
Reg
ulat
ed b
y Y
es-A
ssoc
iate
d Pr
otei
n (d
oi: 1
0.10
89/s
cd.2
012.
0685
)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

34
34
Fig. 4. The phenotypic variations of chondrocytes in response to different shear stresses.
Chondrocytes were immunostained for collagen I (green), collagen II (red), and DAPI (blue) after
2 days of perfusion. A: Fluorescence images of collagen I and collagen II staining in cells
exposed to different conditions. Scale bar: 50 μm. B: Quantitative analysis of the relative
fluorescence intensities of collagen II and collagen I under different conditions. The error bars are
the mean ± SD. *P < 0.05 versus 0.009 dyne/cm2.
Page 34 of 35
Stem
Cel
ls a
nd D
evel
opm
ent
Mes
ench
ymal
Ste
m C
ell a
nd C
hond
rocy
te F
ates
in a
Mul
ti-Sh
ear
Mic
rode
vice
are
Reg
ulat
ed b
y Y
es-A
ssoc
iate
d Pr
otei
n (d
oi: 1
0.10
89/s
cd.2
012.
0685
)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

35
35
Page 35 of 35
Stem
Cel
ls a
nd D
evel
opm
ent
Mes
ench
ymal
Ste
m C
ell a
nd C
hond
rocy
te F
ates
in a
Mul
ti-Sh
ear
Mic
rode
vice
are
Reg
ulat
ed b
y Y
es-A
ssoc
iate
d Pr
otei
n (d
oi: 1
0.10
89/s
cd.2
012.
0685
)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.





























