Embed Size (px)
Citation preview

Mutation Research, 283 (1992) 145-156 145 © 1992 Elsevier Science Publishers B.V. All rights reserved 0165-7992/92/$05.00
MUTLET 0715
Mechanism of mutagenicity by 5-hydroperoxymethyl-2'-deoxyuridine, an intermediate product of ionizing radiation, in bacteria
HPMdU bacterial mutagenicity and oxidation of D N A bases
Umesh Patel ~'*, Ramesh Bhimani a and Krystyna Frenkel a,b,c New York University Medical Center, Departments of ~ Environmental Medicine and b Pathology and ~ Comprehensive Kaplan Cancer
Center, 550 First Avenue, New York, NY 10016-6451, USA
(Received 19 February 1992) (Revision received 26 May 1992)
(Accepted 13 June 1992)
Keywords: Oxidative DNA modification; Nucleoside hydroperoxide; Chelating agents; Free-radical scavengers; Inhibitors
Summary
The specific objective was to find what processes are responsible for the mutagenicity of 5-hydroper- oxymethyl-2'-deoxyuridine (HPMdU), which is a product of ionizing radiation, and what role transition metal ions play in those processes. We found that HPMdU is a more potent mutagen than its decomposition products 5-hydroxymethyl-2'-deoxyuridine (HMdU) and 5-formyl-2'-deoxyuridine (FdU) in the Salmonella typhimurium strains tested, with the TA100 strain being the most sensitive. HMdU exerted intermediate mutagenicity and FdU was the weakest of the three compounds. At 50 nmoles/ plate, HPMdU increased the number of revertants by 4-fold, whereas 1000 nmoles HMdU was required to enhance the number of revertants by 5-fold. Pretreatment of TA100 with o-phenanthroline, a membrane-permeable Fe and Cu chelator, caused an increase in mutagenicity of the low HPMdU doses but inhibited that of the 50 nmoles HPMdU/plate, while desferal, a membrane-impermeable Fe chelator, had virtually no effect. Azide (a catalase inhibitor) enhanced HPMdU mutagenicity, whereas 3-amino-l,2,4-triazole (a catalase and peroxidase inhibitor) and ammonium formate (a hydroxyl radical scavenger) were protective. Preincubation of TA100 cells with 20 and 40 nM HPMdU caused dose-de-
* Present address: Department of Molecular Biology, The Cleveland Clinic Foundation, 9500 Euclid Avenue, Cleve- land, OH 44195, USA.
Correspondence: Dr. Krystyna Frenkel, Department of Envi- ronmental Medicine, PHL 802, New York University Medical Center, 550 First Avenue, New York, NY 10016-645l, USA. Tel. (212) 263-6610; Fax (212) 263-6649.
Abbreviations: Reactive oxygen species (ROS); superoxide an- ion radicals ( . 0 2 ); hydrogen peroxide (H202); hydroxyl radi-
cals (. OH); polymorphonuclear leukocytes (PMNs); cis-6-hy- droperoxy-5-hydroxy-5,6-dihydrothymine (6-TOOH); cis-5-hy- droperoxy-6-hydroxy-5,6-dihydrothymine (5-TOOH); 5-hydro- peroxymethyl uracil (HPMU); 5-hydroxymethyl uracil (HMU); cis-5(6)-hydroperoxy-6(5)-hydroxy-5,6-dihydrothymidine (dTOOH); 5-hydroperoxymethyl-2'-deoxyuridine (HPMdU); 5-hydroxymethyl-2'-deoxyuridine (HMdU); 5-formyl-2'-de- oxyuridine (FdU); 8-hydroxyl-2'-deoxyguanosine (8-OHdG); o-phenanthroline (OP); acetic anhydride (Ac20); 3-amino- 1,2,4-triazole (AT); ammonium formate (AF); plaque-forming units (pfu).

146
pendent formation of the oxidized DNA base derivatives HMdU, thymidine glycol and 8-hydroxyl-2'-de- oxyguanosine (8-OHdG), known hydroxyl radical-mediated oxidation products. Cumulatively, these results suggest that the genetic effects of HPMdU are due to its hydroperoxide moiety, which upon reacting with Fe generates hydroxyl radicals that in turn oxidize neighboring bases in cellular DNA. This also may be a mechanism by which ionizing radiation exerts its long-term effects.
Reactive oxygen species (ROS), which include superoxide anion radicals ( . 0 2), hydrogen perox- ide (H202), hydroxyl radicals ( .OH) and singlet oxygen, are generated inside the cells as a result of normal electron transport or respiration (Cerutti, 1985; Vuillaume, 1987). These ROS can also be generated by the action of ionizing radia- tion (Scholes, 1983; Hutchinson, 1985; T6oule, 1987), oxidative burst by polymorphonuclear leukocytes (PMNs) (Badwey and Karnovsky, 1980; Klebanoff, 1980; Babior, 1984; Frenkel and Chrzan, 1987a,b; Zhong et al., 1990), as well as during metabolism of various xenobiotics (Lesko and Lorentzen, 1985; Frenkel et al., 1988). Exces- sively generated ROS induce various types of damage in cells (Weitzman and Gordon, 1990; Frenkel, 1992), one of which is oxidative modifi- cation of bases in DNA (Troll et al., 1984; Lewis and Adams, 1985; Frenkel et al., 1986; Frenkel, 1989; Floyd, 1990). Qualitatively the same oxi- dized bases are formed by cellularly-produced as by radiogenically-generated ROS (Frenkel et al., 1986). Among the DNA bases, thymine is consid- ered the most susceptible to the damaging effects of ionizing radiation, as exemplified by formation of 13 hydroperoxides (Cadet and T6oule, 1978; Scholes, 1983; Simic and Jovanovic, 1986). Of these, the major products are cis-6-hydroperoxy- 5-hydroxy-5,6-dihydrothymine (6-TOOH) and its isomer cis-5-hydroperoxy-6-hydroxy-5,6-dihydro- thymine (5-TOOH), whereas 5-hydroperoxymeth- yl uracil (HPMU) is the minor product. These hydroperoxides gradually undergo secondary re- actions to form chemically more stable oxy- genated derivatives. The decomposition of both cis-TOOH isomers results in the formation of the same products, (+ ) and ( - ) cis-thymine glycol, whereas that of HPMU results in the formation of 5-hydroxymethyl uracil (HMU) and 5-formyl uracil (Cadet and T6oule, 1975a,b).
Both types of thymine hydroperoxides (TOOH and HPMU) are mutagenic and this effect of TOOH was found to be enhanced by transition metal ions (Thomas et al., 1976; Wang et al., 1979). HMU, which is the decomposition product of HPMU, is weakly mutagenic in the TA100 and TA102 strains of S. typhimurium (Shirnam6-Mor6 et al., 1987), while the ( + ) and ( - ) isomers of thymine glycol, the decomposition products of TOOH, are not mutagenic (Cadet et al., 1981). The 2'-deoxyribonucleosides of TOOH and HPMU [dTOOH and HPMdU (5-hydroper- oxymethyl-2'-deoxyuridine)] are more potent mutagens than the base hydroperoxides TOOH and HPMU. Similarly, the 2'-deoxyribonucleoside of HMU [5-hydroxymethyl-2'-deoxyuridine (HMdU)], which is a strong mutagen considering that it is a base analog, is much more effective than its base HMU (Shirnam6-Mor6 et al., 1987). Decomposition of HPMdU in the presence of transition metal ions leads to the formation of HMdU and/or FdU (5-formyl-2'-deoxyuridine) in various proportions, depending on the type of metal ions and the chelators used (Tofigh and Frenkel, 1989). Although FdU is also mutagenic, it is much less potent than HMdU (Kasai et al., 1990; Bhimani et al., 1992). In addition to its mutagenic potential, HMdU is also cytotoxic and cytostatic (Waschke et al., 1975; Kahilainen et al., 1985; Meldrum et al., 1985), whereas FdU monophosphate is an inhibitor of thymidine syn- thetas¢ (Wataya et al., 1977; Balzarini et al., 1982).
HPMdU formed in the DNA through the ac- tion of y-radiation may be subject to spontaneous or metal ion-induced decomposition, with the products of that decomposition constituting a fur- ther threat to the normal functioning of the cells. Studies on HPMdU degradation in vitro suggest that, in addition to HMdU and FdU, also -OH

can be formed from HPMdU during interaction of this hydroperoxide with transition metal ions (Tofigh and Frenkel, 1989). Since these radi- cals are known to cause damage to DNA bases, their release from HPMdU could lead to the additional oxidative DNA modification. Hence HPMdU, which is a product of the free-radical attack on thymidine (free or in DNA), potentially can become a source of radicals and propagate the damage.
Until now, the published mutagenicity studies were carried out on the HPMdU-derived decom- position products HMdU and FdU (Bilimoria and Gupta, 1986; Shirnam6-Mor6 et al., 1987; Kasai et al., 1990) rather than on the hydroperoxide itself, except for one meeting presentation (Cadet et al., 1981). The aims of this investigation were to answer the following questions: (1) Is HPMdU mutagenic in bacteria, and if so are these effects due to its decomposition products HMdU and/or FdU? (2) Are iron ions and /o r formation of H20 2 necessary for the mutagenic activity of HPMdU? (3) Are .OH released from HPMdU and do they cause oxidative modification of cellu- lar DNA?
Materials and methods
Chemicals 5-Hydroxymethyl-2'-deoxyuridine (HMdU),
sodium azide, o-phenanthroline (OP) and cata- lase were obtained from Sigma Chemical Com- pany (St. Louis, MO), and desferal from Ciba Pharmaceutical Company (Summit, NJ). [3H]- Acetic anhydride {[3H]-Ac20} with a specific activity of 50 mCi/mmole was purchased from New England Nuclear (DuPont, Boston, MA). HPMdU and FdU were synthesized and purified in our laboratory according to the previously de- scribed procedures using HMdU as the substrate (Cline et al., 1959; Tofigh and Frenkel, 1989).
Briefly, to obtain HPMdU, HMdU (5 rag) was treated with 30% H20 2 (750 ~l) in the presence of concentrated HCI (12.5 ~1). The reaction mix- ture was stirred at room temperature for 40 h and, after adjusting the pH to 6.0 with ammo- nium hydroxide, was purified by HPLC on an Altex ODS column (1.0 × 25 cm i.d.) with water as the eluent. The concentration of HPMdU in
147
the combined fractions was usually about 1 mM, as determined by the absorbance at 265 nm and the extinction coefficient • = 12300. HPMdU was stored in polypropylene tubes as an aqueous solu- tion at 4°C. Under these conditions HPMdU is stable; however, any attempt to concentrate it results in partial or complete decomposition (un- published results).
To obtain FdU, HMdU (20 mg) was dissolved in 0.2 N HC1 (0.8 ml) and treated with MnO 2 (80 mg) at room temperature. The reaction mixture was stirred for 48 h, and the FdU was purified by HPLC on an ODS column, lyophilized and stored at 4°C.
Bacterial strains Tester strains TA97a, TA98, TA100 and
TA102, which are S. typhimurium LP2 histidine- deficient auxotrophs, were kindly provided by Dr. Bruce N. Ames (Dept. of Biochemistry, Univer- sity of California, Berkeley, CA). These strains were chosen because they can be converted to prototrophy by agents that cause frame-shift mu- tations (TA97 and TA98) or base-pair substitu- tions at the GC site (TA100) or the AT site (TA102) (Ames et al., 1975; McCann et al., 1975; Maron and Ames, 1983). All of these strains carry the pKM101 plasmid, with TA102 additionally containing a multicopy plasmid pAQ1, which is the carrier of its histidine mutation (Levin et al., 1982; Levin and Ames, 1986). These strains were used in the current study because the presence of plasmid pK101 was required for the expression of mutagenicity by HMdU (Shirnam~-Mor6 et al., 1987).
Ames test Cultures were grown overnight in Oxoid broth
to a density of 1-2 × 109 cells/ml, as previously described (Ames et ai., 1975; Maron and Ames, 1983). The test agent and 0.1-ml aliquot of bacte- ria were mixed together with 2 ml of molten top agar containing 0.05 mM histidine and biotin, and the mixture was poured onto minimal agar plates with 2% glucose. In some experiments, bacteria were washed with phosphate-buffered saline, pre-incubated with iron chelators or catalase in- hibitors at 37°C for 30 min with shaking, and washed again prior to mixing with the test agent

148
and molten agar. The plates were incubated at 37°C for 48 h and scored for his + revertants. The results are expressed as rever tants /p la te and are mean values from 2-4 Expts.+_ SE. All of the agents used for the pretreatment of bacteria were non-toxic to TA100 at the highest concentrations used. The number of spontaneous colonies were at least at the same level, if not higher (azide), as in their absence. Only formate decreased the number of spontaneous colonies, suggesting that the background mutagenesis may be caused by • OH. Since it is not possible to count cells grown as a lawn in the bacterial mutagenesis assays, only non-toxic ( < 50 nmoles /p la te ) concentrations of HPMdU were used in the experiments, as deter- mined by thinning of the lawn (Maron and Ames, 1983).
Analysis of DNA S. typhimurium TA100 cells (100 ml) were
grown overnight as described above, pelleted and resuspended into the half volume of double- strength Luria broth. HPMdU (sterilized by fil- tration through a 0.2-1zm membrane) was added to the cells to a final concentration of 20 and 40 nM, and the volume was adjusted to 100 ml. The cells were incubated at 37°C with mixing for 2 h, harvested and suspended into 2 ml of Tr is- sucrose buffer (pH 8.5) containing 5 mg /ml lysozyme. After incubation at 37°C for 30 min, the cells were lysed by adding 5 ml of 10% SDS. The control preparation was grown in the ab- sence of HPMdU and lysed the same way as the HPMdU-trea ted bacteria. After lysis, DNA was extracted and purified using A.S.A.P T M columns from Boehringer-Mannheim according to the procedure of the manufacturer. Purified DNA was enzymatically digested to nucleosides at 37°C for 2.5 h with DNAase I ( + M g 2+) at pH 7.4, followed by nuclease P1 ( + Z n 2+) at pH 5.1 and alkaline phosphatase at pH 7.4, as described by Frenkel et al. (1991). Oxidized nucleosides were separated from normal nucleosides by HPLC on the ODS column (HPLC 1), using water as the eluent for 40 min at 2 ml /min , followed by 2.5% acetonitrile in water for 40 min, and 5% and 10% acetonitrile for 20 min each. All fractions after the first 30 ml, except those containing normal nucleosides were combined, dried and acetylated
with [3H]Ac20 in the presence of a catalyst (Frenkel et al., 1991). Tritium-containing nucleo- side acetates were spiked with marker acetates synthesized in our laboratory and chromato- graphed on the ODS column (HPLC 2). Quanti- tation of the oxidized nucleosides was based on the known specific activity of [3H]Ac20 used for the acetylation, the level of tritium-containing material present in the peaks coeluting with marker acetates (HPLC 2) and the amount of normal nucleosides, the levels of which were quantitated by integration of their peak areas on HPLC 1 chromatograms (Frenkel et al., 1991; Wei and Frenkel, 1991). The following formula was used for quantitation:
Net cpm of a specific oxidized nucleoside acetate
Efficiency × conversion factor × number of acetylated groups
where the efficiency of the liquid-scintillation counter was 30% (0.3). The conversion factor is 11.1, when radioactive:non-radioactive Ac20 is 1 : 10 and the specific activity of [3H]Ac20 is 3300 cpm/nmole acetate. The number of acetyl groups was determined by mass spectroscopy of marker acetates (Frenkel et al., 1991).
Results
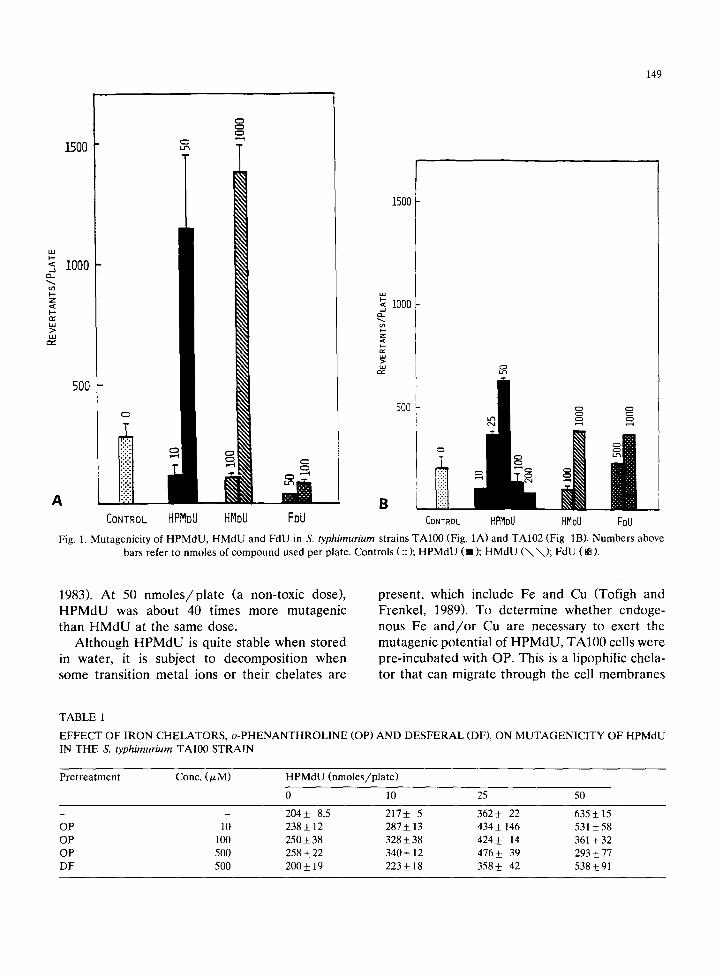
Both HPMdU and HMdU were mutagenic in TA100, with the former compound being more potent than the latter, whereas FdU was virtually inactive (Fig. 1A). In TA102, HPMdU was again the most effective, followed by HMdU and FdU, which were about equipotent (Fig. 1B). HPMdU and HMdU were approximately 2- and 3.5-fold more mutagenic in TA100 than in TA102. All 3 agents were inactive in TA97 and only HPMdU was weakly mutagenic in TA98 (not shown). Since TA100 was the most sensitive to the mutagenic effects of HPMdU, this S. typhimurium strain was used in all other experiments.
Fig. 2 shows that the number of TA100 rever- tants was enhanced with the increased doses of HPMdU and HMdU but those induced by hy- droperoxide were very effective even at low doses. However, at the higher doses (100 nmoles /p la te) , HPMdU was already toxic as visualized by thin- ning of the background lawn (Maron and Ames,
149
I - z
I.-
>
A
1500
1500
1000
1000
5OO
5OO
° i
B CONTROL HPMDU HMDU FDU CONTROL HPMDU HMDU FDU
Fig. I. Mutagenicity ol HPMdU, HMdU and FdU in S, typhimurium strains TAI00 (Fig. IA) and TA]02 (Fig. IB). Numbers above bars refer to nmoles of compound used per plate. Controls (::); HPMdU (•); HMdU ( \ \ ) ; FdU (BB).
1983). At 50 nmoles /p la t e (a non-toxic dose), HPMdU was about 40 times more mutagenic than HMdU at the same dose.
Although HPMdU is quite stable when stored in water, it is subject to decomposition when some transition metal ions or their chelates are
present, which include Fe and Cu (Tofigh and Frenkel, 1989). To determine whether endoge- nous Fe a n d / o r Cu are necessary to exert the mutagenic potential of HPMdU, TA100 cells were pre-incubated with OP. This is a lipophilic chela- tor that can migrate through the cell membranes
TABLE 1
E F F E C T OF IRON C H E L A T O R S , o - P H E N A N T H R O L I N E (OP) A N D D E S F E R A L (DF), ON M U T A G E N I C I T Y OF H P M d U IN THE S. typhimurium TA100 STRAIN
Pretreatment Conc. (IzM) H P M d U (nmoles /p la te )
0 10 25 50
- 204+ 8.5 217+ 5 362_+ 22 OP 10 238 + 12 287 -+ 13 434 -+ 146 OP 100 250-+38 328-+38 424_+ 14 OP 500 258 -+ 22 340-+ 12 476-+ 39 DF 500 200-+ 19 223 + 18 358-+ 42
635 + 15 531 _+ 58 361 _+ 32 293 _+ 77 538__+91

1 5 0
2C00 I
} 1000"
i
5
- T / /
~0 100 1000
N~L/P~ATE
Fig. 2. Mutagenicity of HPMdU (o) , HMdU (e) and FdU ( • ) in S. typhimurium TA100 strain: dose-response curves.
and bind both Fe and Cu (Freeman and Crapo, 1982; de Mello Filho and Meneghini, 1985). The presence of 10, 100 and 500 p,M OP elevated the mutagenicity of 10 nmoles HPMdU/plate by 32, 51 and 57%, and of 25 nmoles HPMdU/plate by 20, 17 and 31%, respectively (Table 1). However, OP inhibited the mutagenicity when the cells
were incubated with 50 nmoles HPMdU. Those decreases are substantial, and OP dose-depen- dent at 32%, 74% and 92% for cells incubated with 10, 100 and 500 /xM OP, respectively. Des- feral (500 ~xM), an Fe chelator that does not migrate through cell membranes (Aust et al., 1985; Halliwell and Gutteridge, 1986), especially when incubated for such a short period of time (30 min), had no effect when followed by 10 and 25 nmoles HPMdU and slightly inhibited (22%) HPMdU-mediated his + reversion when 50 nmoles hydroperoxide were used.
Preincubation of TA100 with 0.1 mM azide, a catalase inhibitor, increased the mutagenic po- tency of 1 and 5 nmoles HPMdU/plate by about 15%, but some toxicity was observed when 25 and 50 nmoles HPMdU/plate followed the azide. 1 mM azide enhanced the mutagenicity of 1 nmole HPMdU by 63% but was already toxic when the latter was used at 5 nmoles/plate (Table 2), and there was no bacterial growth at 25 and 50 nmoles/plate. Interestingly, 1 mM azide also en- hanced the spontaneous reversion by 45%, which points to H20 2 as being a possible intermediate
TABLE 2
EFFECTS OF AZIDE AND 3-AMINO-1,2,4-TRIAZOLE (AT) ON MUTAGENICITY OF HPMdU IN THE S. typhimurium TA100 STRAIN
Pre- Conc. treat- (/xM) ment
HPMdU (nmoles/plate)
0 1 5 25 50
- - 410_+ 14 330 + 14 299 +_ 1 573 _+ 13 940_+ 60 Azide 0.1 423 _+ 27 385 +_ 15 425 +- 25 272 _+ 52 135 + 5 a Azide 1.0 596_+24 538_+ 2 215_+10 ~ NG NG AT 0.1 237 +- 17 250 290 +- 10 380 _+ 20 930 _+ 70
Some toxicity observed. NG, no growth.
TABLE 3
EFFECTS OF AMMONIUM FORMATE (AF) ON HPMdU-INDUCED MUTAGENICITY IN THE S. typhimurium TA100
STRAIN
Pretreatment Conc. (Mm) HPMdU (nmoles plate)
0 5 25 50
- - 410+-20 410+- 2 481 +-56 AF 0.1 386 _+ 24 365 +_ 15 408_+ 5 AF 1.0 141 _+ 16 139+- 6 151 _+25
788+ 108 488_+ 13 148+ 57
151
1 0
. . . .
0 - - ' f - - 50 75 125 150 175
%LUME (ML)
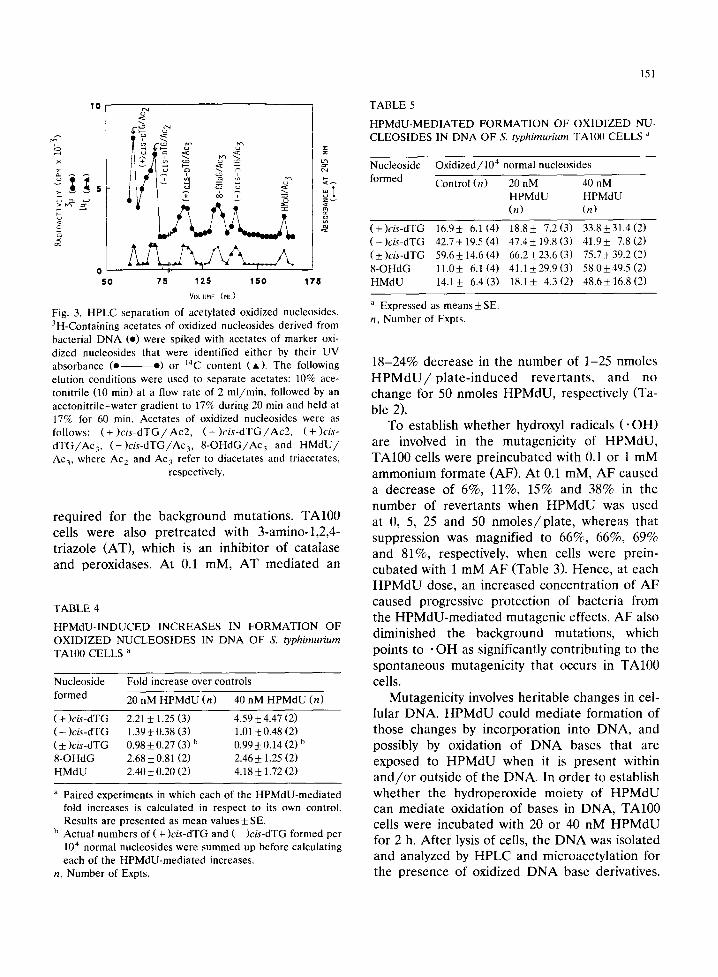
Fig. 3. HPLC separation of acetylated oxidized nucleosides. 3H-Containing acetates of oxidized nucleosides derived from bacterial D N A (e) were spiked with acetates of marker oxi- dized nucleosides that were identified either by their UV absorbance (e e) or 14C content (A). The following elution conditions were used to separate acetates: 10% ace- tonitrile (10 min) at a flow rate of 2 m l / m i n , followed by an acetoni tr i le-water gradient to 17% during 20 min and held at 17% for 60 min. Acetates of oxidized nucleosides were as follows: ( + )cis-dTG / Ac2, ( - )cis-dTG / Ac2, ( + )cis- d T G / A c 3 , (-)cis-dTG/Ac3, 8 - O H d G / A c 3 and H M d U / Ac3, where Ac 2 and Ac 3 refer to diacetates and triacetates,
respectively.
required for the background mutations. TA100 cells were also pretreated with 3-amino-l,2,4- triazole (AT), which is an inhibitor of catalase and peroxidases. At 0.1 mM, AT mediated an
T A B L E 4
H P M d U q N D U C E D INCREASES IN F O R M A T I O N OF O X I D I Z E D NUCLEOSIDE S IN D N A OF S. typhimurium TA100 CELLS a
Nucleoside formed
Fold increase over controls
20 nM H P M d U (n) 40 nM H P M d U (n)
( + )cis-dTG 2.21 _+ 1.25 (3) 4.59 _+ 4.47 (2) ( - )cis-dTG 1.39 _+ 0.38 (3) 1.01 _+ 0.48 (2) (_+)cis-dTG 0.98+0.27(3) b 0.99_+0.14(2) b 8-OHdG 2.68 -+ 0.81 (2) 2.46 -+ 1.25 (2) H M d U 2.40 _+ 0.20 (2) 4.18 _+ 1.72 (2)
a Paired experiments in which each of the HPMdU-media ted fold increases is calculated in respect to its own control. Results are presented as mean values_+ SE.
~' Actual numbers of ( + )cis-dTG and ( - )cis-dTG formed per 104 normal nucleosides were summed up before calculating each of the HPMdU-media ted increases.
n, Number of Expts.
TABLE 5
H P M d U - M E D I A T E D F O R M A T I O N OF O X I D I Z E D NU- CLEOSIDES IN DNA OF S. typhirnurium TA100 CELLS ~
Nucleoside formed
Oxidized/104 normal nucleosides
Control (n) 20 nM 40 nM H P M d U H P M d U (n) (n)
(+)cis-dTG 16.9_+ 6.1(4) 18.8_+ 7.2(3) 33.8_+31.4(2) (-)cis-dTG 42.7-+19.5(4) 47.4-+19.8(3) 41.9_+ 7.8(2) (+_)cis-dTG 59.6-+14.6(4) 66.2-+23.6(3) 75.7-+39.2(2) 8-OHdG 11.0-+ 6.1(4) 41.1_+29.9(3) 58 .0+49.5(2) H M d U 14.1_+ 6.4(3) 18.1_+ 4.3(2) 48.6+16.8(2)
Expressed as means_+ SE. n, Number of Expts.
18-24% decrease in the number of 1-25 nmoles HPMdU/p la t e - induced revertants, and no change for 50 nmoles HPMdU, respectively (Ta- ble 2).
To establish whether hydroxyl radicals (. OH) are involved in the mutagenicity of HPMdU, TA100 cells were preincubated with 0.1 or 1 mM ammonium formate (AF). At 0.1 mM, AF caused a decrease of 6%, 11%, 15% and 38% in the number of revertants when HPMdU was used at 0, 5, 25 and 50 nmoles/plate, whereas that suppression was magnified to 66%, 66%, 69% and 81%, respectively, when cells were prein- cubated with 1 mM AF (Table 3). Hence, at each HPMdU dose, an increased concentration of AF caused progressive protection of bacteria from the HPMdU-mediated mutagenic effects. AF also diminished the background mutations, which points to • OH as significantly contributing to the spontaneous mutagenicity that occurs in TA100 ceils.
Mutagenicity involves heritable changes in cel- lular DNA. HPMdU could mediate formation of those changes by incorporation into DNA, and possibly by oxidation of DNA bases that are exposed to HPMdU when it is present within and /or outside of the DNA. In order to establish whether the hydroperoxide moiety o f HPMdU can mediate oxidation of bases in DNA, TA100 cells were incubated with 20 or 40 nM HPMdU for 2 h. After lysis of cells, the DNA was isolated and analyzed by HPLC and microacetylation for the presence of oxidized DNA base derivatives.
152
10
5 <
Z
CONTROL HMDU (+)C,_LS_-DTG (-)C.j,_E-DTG 8-0HDG
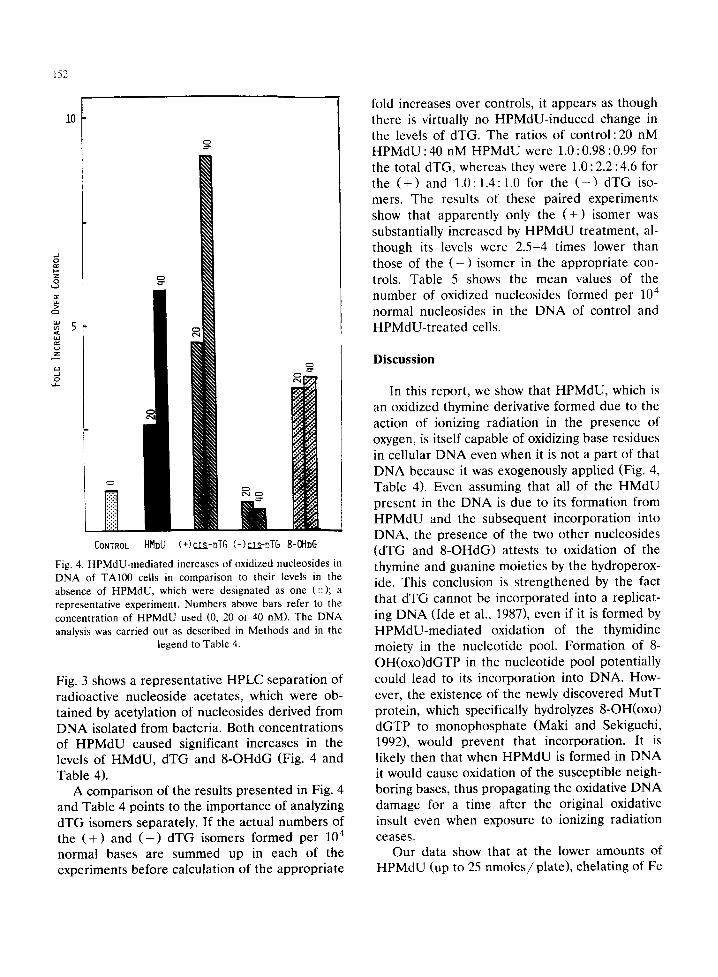
Fig. 4. HPMdU-mediated increases of oxidized nucleosides in DNA of TA100 cells in comparison to their levels in the absence of HPMdU, which were designated as one (::); a representative experiment. Numbers above bars refer to the concentration of HPMdU used (0, 20 or 40 nM). The DNA analysis was carried out as described in Methods and in the
legend to Table 4.
Fig. 3 shows a representative HPLC separation of radioactive nucleoside acetates, which were ob- tained by acetylation of nucleosides derived from D N A isolated from bacteria. Both concentrations of H P M d U caused significant increases in the levels of HMdU, dTG and 8 -OHdG (Fig. 4 and Table 4).
A comparison of the results presented in Fig. 4 and Table 4 points to the importance of analyzing dTG isomers separately. If the actual numbers of the ( + ) and ( - ) dTG isomers formed per 10 4
normal bases are summed up in each of the experiments before calculation of the appropriate
fold increases over controls, it appears as though there is virtually no HPMdU-induced change in the levels of dTG. The ratios of control :20 nM H P M d U : 40 nM HPMdU were 1.0 : 0.98 : 0.99 for the total dTG, whereas they were 1.0 : 2.2 : 4.6 for the ( + ) and 1.0:1.4:1.0 for the ( - ) dTG iso- mers. The results of these paired experiments show that apparently only the ( + ) isomer was substantially increased by HPMdU treatment, al- though its levels were 2.5-4 times lower than those of the ( - ) isomer in the appropriate con- trols. Table 5 shows the mean values of the number of oxidized nucleosides formed per 104 normal nucleosides in the DNA of control and HPMdU-t rea ted cells.
Discussion
In this report, we show that HPMdU, which is an oxidized thymine derivative formed due to the action of ionizing radiation in the presence of oxygen, is itself capable of oxidizing base residues in cellular D N A even when it is not a part of that D N A because it was exogenously applied (Fig. 4, Table 4). Even assuming that all of the H M d U present in the D N A is due to its formation from H P M d U and the subsequent incorporation into DNA, the presence of the two other nucleosides (dTG and 8-OHdG) attests to oxidation of the thymine and guanine moieties by the hydroperox- ide. This conclusion is strengthened by the fact that dTG cannot be incorporated into a replicat- ing D N A (Ide et al., 1987), even if it is formed by HPMdU-media ted oxidation of the thymidine moiety in the nucleotide pool. Formation of 8- OH(oxo)dGTP in the nucleotide pool potentially could lead to its incorporation into DNA. How- ever, the existence of the newly discovered MutT protein, which specifically hydrolyzes 8-OH(oxo) dGTP to monophosphate (Maki and Sekiguchi, 1992), would prevent that incorporation. It is likely then that when H P M d U is formed in D N A it would cause oxidation of the susceptible neigh- boring bases, thus propagating the oxidative D N A damage for a time after the original oxidative insult even when exposure to ionizing radiation ceases.
Our data show that at the lower amounts of HPMdU (up to 25 nmoles /p la te ) , chelating of Fe

153
a n d / o r Cu ions by pre-incubation of TA100 with OP (a lipophilic chelator) enhances mutagenicity of HPMdU, but decreases it at 50 nmoles /p la t e (Table 1). These findings suggest that, when pres- ent in smaller amounts, HPMdU is partially de- composed en route to the DNA by Fe. This would lead to a lower apparent mutagenicity, unless OP is also present and chelates that Fe. However, at higher amounts, more intact HP- MdU can reach the DNA and, in the presence of Fe bound to DNA, exert its mutagenic effects. In this case, OP would remove the DNA-bound Fe that apparently is needed for mutagenicity of HPMdU. This explanation seems possible be- cause a lower concentration of OP (10 ~M) only caused a 32% decline, whereas higher concentra- tions (100 /.~M and 500 p~M) caused 74% and 92% inhibition in the mutagenicity of 50 nmoles H P M d U / p l a t e , respectively. An OP-mediated increase in background mutation in TA100 points to the possibility that Cu is involved in the spon- taneous mutagenic events of this bacterial strain. This would be because the C u - O P complex is more reactive than Cu alone, whereas chelating of Fe by OP diminishes Fe-mediated reactions (Aust et al., 1985; Halliwell and Gutteridge, 1986). Desferal, which is an Fe chelator that cannot cross the cellular membranes within a short expo- sure time, has virtually no effect on HPMdU mutagenicity.
Although pretreatment of bacteria with AT resulted in only minimal apparent inhibition of HPMdU-mediated mutagenicity, it caused a sig- nificant (40%) decrease in spontaneous reversion (Table 2). Using that number of revertants as a control shows that in actuality the AT-mediated inhibition of catalase a n d / o r peroxidase causes a dose-dependent enhancement in the mutagenicity of HPMdU. The AT-mediated increases ranged from 5%, through 22%, 60% and 290% for 1, 5, 25 and 50 nmoles H P M d U / p l a t e , respectively. Whereas in the absence of AT, mutagenicity of 25 and 50 nmoles H P M d U / p l a t e was enhanced only by 25% and 130% over the unexposed con- trol, respectively. All of these results suggest that it is the intact HP MdU that is required for its mutagenicity, while the cellular antioxidant en- zymes (catalase a n d / o r peroxidase) protect these cells from undergoing mutational changes. How-
ever, it is also likely that it is a peroxidase that mediates at least some of the spontaneous muta- genicity in the TA100 strain because AT de- creased it, while azide (a catalase inhibitor) en- hanced it, probably due to the accumulation of U202.
Cumulatively, the results presented point to the intact hydroperoxide as being responsible for the mutagenic potential of HPMdU and also sug- gest that Fe is involved in that process. Moreover, it is likely that • OH are formed as a result of Fe interaction with this hydroperoxide because AF (an -OH scavenger) strongly (up to 85%) sup- presses the mutagenic effects of HPMdU (Table 3). The spontaneous mutagenicity is also signifi- cantly (65%) decreased by AF, which points to the involvement of • OH in the background muta- genicity of this bacterial strain. These conclusions are strengthened by our finding that treatment of TA100 cells with HPMdU causes oxidation of bases in the bacterial DNA (Table 4), a process known to be mediated by • OH (Klein et al., 1991; Frenkel, 1992).
The extent of the mutagenic changes for HPMdU and HMdU are 2- and 3.5-fold lower, respectively, in the TA102 than in the TA100 strain (Fig. 1). The reasons why HPMdU is more mutagenic in the TA100 than in the TA102 strain are not known. However, it potentially can oxi- dize the guanine residue present at the GC locus, thus resulting in formation of 8-OHdG (known to be mutagenic) (Kuchino et al., 1987; Wood et al., 1990; Shibutani et al., 1991; Cheng et al., 1992), or of another type of oxidized G a n d / o r C derivative. Although HPMdU was expected to be more potent in TA102, because this particular strain is sensitive to the action of oxidizing muta- gens, this did not occur. The lower mutagenicity in TA102 could be due to the following: (a) The HPMdU did not get incorporated in the place of T as an intact hydroperoxide or HMdU; (b) Even if HPMdU or HMdU was incorporated, it still might direct the insertion of A into the comple- mentary DNA strand, as was shown for HMdU (Levy and Teebor, 1990) and therefore would not be miscoding; a n d / o r (c) If dTG is formed at the AT locus by • OH liberated from HPMdU, it may constitute a replicative block (Ide et al., 1985; Rouet and Essigmann, 1985) rather than a muta-

154
genic lesion (Basu et al., 1989). The levels of HMdU-induced mutagenicity in this study are comparable to those obtained previously (Shir- nam6-Mor6 et al., 1987).
Cumulatively, these results prove that the genotoxicity and mutagenicity of HPMdU is not mediated by HMdU and/or FdU, which are the decomposition products of hydroperoxide. More- over, taken together with the results of pretreat- ment with various scavengers, formation of dTG and 8-OHdG in cellular DNA proves that this type of oxidation is mediated by • OH generated by the hydroperoxide moiety of HPMdU interact- ing with Fe ions bound to DNA or to nearby proteins. These results also point to the possible mechanism of the long-term effects induced by ionizing radiation, since formation of HPMdU in DNA allows site-specific oxidative modification of genetic material long after radiation exposure.
Acknowledgments
This research was supported in part by grant Nos. CA 49798 and CA 13343 from the National Cancer Institute (its contents are solely the re- sponsibility of the authors and do not necessarily represent the official views of the National Can- cer Institute), by ES 00260 from the National Institute of Environmental Health Sciences, and by SIG-9 from the American Cancer Society.
References
Ames, B.N., J. McCann and E. Yamasaki (1975) Methods for detecting carcinogens and mutagens with Salmonella/ mammalian-microsome mutagenicity test, Mutation Res., 31,347-364.
Aust, S.D., L.A. Morehouse and C.E. Thomas (1985) Role of metals in oxygen radical reactions, J. Free Rad. Biol. Med., 1, 3-25.
Babior, B.M. (1984) Oxidants from phagocytes: Agents of defense and destruction, Blood, 64, 959-966.
Badwey, J.A., and M.L. Karnovsky (1980) Active oxygen species and the functions of phagocytic leukocytes, Annu. Rev. Biochem., 49, 695-726.
Balzarini, J., E. De Clercq, M.P. Mertes and P.F. Torrence (1982) 5-Substituted 2'-deoxyuridines: Correlation be- tween inhibition of tumor cell growth and inhibition of thymidine kinase and thymidylate synthetase, Biochem. PharmacoL, 31, 3673-3682.
Basu, A.K., E.L. Loechler, S.A. Leadon and J.M. Essigman (1989) Genetic effects of thymine glycol: Site-specific mu-
tagenesis and molecular modeling studies, Proc. Natl. Acad. Sci. (U.S.A.), 86, 7677-7681.
Bhimani, R., U. Patel and K. Frenkel (1992) Mutagenicity of 5-hydroperoxymethyl-2'-deoxyuridine (HPMdU), an inter- mediate formed in DNA by y-radiation, Proc. Am. Assoc. Cancer Res., 33, in press.
Bilimoria, M.K., and S.V. Gupta (1986) Comparison of the mutagenic activity of 5-hydroxymethyldeoxyuridine with 5-substituted 2'-deoxyuridine analogs in the Ames Salmonella/microsome test, Mutation Res., 169, 123-127.
Cadet, J., and R. T6oule (1975a) Radiolyse gamma de la thymidine en solution aqueuse a6re& I. Identification de hydroxyhydroperoxydes, Bull. Soc. Chim. Fr., 3-4, 879- 884.
Cadet, J., and R. T6oule (1975b) Radiolyse gamma de ta thymidine en solution aqueuse a6re6. II. Characterization des produits stables, Bull. Soc. Chim. Fr., 3-4, 885-890.
Cadet, J., and R. T6oule (1978) Comparative study of oxida- tion of nucleic acid components by hydroxyl radicals, sin- glet oxygen and superoxide anion radicals, Photochem. Photobiol., 28, 661-667.
Cadet, J., L. Voituriez, J.L. Seed, E. Bueding, T. Nagamatsu and S.Y. Wang (1981) Preparation and mutagenic activi- ties of thymidine hydroperoxides and of their degradation products, in: International Round Table on Nucleosides, Nucleotides and Their Biological Applications, Belgium/ Antwerp, pp. 13-14.
Cerutti, P.A. (1985) Prooxidant state and tumor promotion, Science, 22, 375-381.
Cheng, K.C., D.S. Cahill, H. Kasai, S. Nishimura and L.A. Loeb (1992) 8-Hydroxyguanine, an abundant form of ox- idative DNA damage, causes G ~ T and A ~ C substitu- tions, J. Biol. Chem., 267, 166-172.
Cline, R.E., R.M. Fink and K. Fink (1959) Synthesis of 5-substituted pyrimidines via formaldehyde addition, J. Am. Chem. Soc., 81, 2521-2527.
de Metlo Filho, A.C., and R. Meneghini (1985) Protection of mammalian cells by o-phenanthroline from lethal and DNA-damaging effects produced by active oxygen species, Biochim. Biophys. Acta, 847, 82-89.
Floyd, R.A. (1990) The role of 8-hydroxyguanine in carcino- genesis, Carcinogenesis, 11, 1447-1450.
Freeman, B.A., and J.D. Crapo (1982) Biology of disease, Free radicals and tissue injury, Lab. Invest., 47, 412-426.
Frenkel, K. (1989) Oxidation of DNA bases by tumor pro- moter-activated processes, Environ. Health Perspect., 81, 45-54.
Frenkel, K. (1992) Carcinogen-mediated oxidant formation and oxidative DNA damage, Pharmacol. Therapeut., in press.
Frenkel, K., and K. Chrzan (1987a) H20 2 formation and DNA base modification by tumor promoter activated poly- morphonuclear leukocytes, Carcinogenesis, 8, 455-460.
Frenkel, K., and K. Chrzan (1987b) Radiation-like modifica- tion of DNA and H20 2 formation by activated human polymorphonuclear leukocytes (PMNs), in: P.A. Cerutti, O.F. Nygaard and M.G. Simic (Eds.), Anticarcinogenesis and Radiation Protection, Plenum, New York, pp. 97-102.

Frenkel, K., K. Chrzan, W. Troll, G.W. Teebor and J.J. Steinberg (1986) Radiation-like modification of bases in DNA exposed to tumor promoter-activated polymor- phonuclear leukocytes, Cancer Res., 46, 5533-5540.
Frenkel, K., J.M. Donahue and S. Banerjee (1988) Benzo(a)pyrene-induced oxidative DNA damage: A possi- ble mechanism for promotion by complete carcinogens, in: P. Cerutti, |. Fridovich and J. McCord (Eds.), Oxy-radicals in Molecular Biology and Pathology, UCLA Symposia on Molecular and Cellular Biology, New Series, Vol. 82, Liss, New York, pp. 509-524.
Frenkel, K., Z. Zhong, H. Wei, J. Karkoszka, U. Patel, K. Rashid, M. Georgescu and J.J. Solomon (1991) Quantita- tive high-performance liquid chromatography analysis of DNA oxidized in vitro and in vivo, Anal. Biochem., 196, 126-136.
Halliwell, B., and J.M.C. Gutteridge (1986) Oxygen free radi- cals in relation to biology and medicine: Some problems and concepts, Arch. Biochem. Biophys., 246, 501-514.
Hutchinson, F. (1985) Chemical changes induced in DNA by ionizing radiation, Prog. Nucleic Acid Res. Mol. Biol., 32, 115-154.
lde, H., Y.W. Kow and S.S. Wallace (1985) Thymine glycol and urea residues in M13 template constitute replicative blocks in vitro, Nucleic Acids Res., 13, 8035-8052.
Ide, H., R.J. Melamede and S.S. Wallace (1987) Synthesis of dihydrothymidine and thymidine glycol 5'-triphosphates and their ability to serve as substrates for Escherichia coli DNA polymerase I, Biochemistry, 26, 964-969.
Kahilainen, L.I., D.E. Bergstrom and J.A. Vilpo (1985) 5-Hy- droxymetyl-2'-deoxyuridine, cytotoxicity and DNA incor- poration studies by using a novel (2-14C) derivative with normal and leukemic human hematopoietic cells, Acta Chem. Scand. B, 39, 477-484.
Kasai, H., A. Iida, Z. Yamaizumi, S. Nishimura and H. Tanooka (1990) 5-Formyldeoxyuridine: a new type of DNA damage induced by ionizing radiation and its mutagenicity to Salmonella strain TAI02, Mutation Res., 243, 249-253.
Klebanoff, S.J. (1980) Oxygen metabolism and the toxic prop- erties of phagocytes, Ann. Intern. Med., 93, 480-489.
Klein, C.B., K. Frenkel and M. Costa (1991) The role of oxidative processes in metal carcinogenesis, Chem. Res. Toxicol., 4, 592-604.
Kuchino, Y., F. Mori, H. Kasai, H. Inoue, S. lwai, K. Miura, E. Ohtsuka and S. Nishimura (1987) Misreading of DNA templates containing 8-hydroxydeoxyguanosine at the modified base and at adjacent residues, Nature (London), 327, 77-79.
Lesko, S.A., and R.J. Lorentzen (1985) Benzo(a)pyrene dione-benzo(a)pyrene diol oxidation-reduction couples; involvement in DNA damage, cellular toxicity, and car- cinogenesis, J. Toxicol. Env. Health, 16, 679-691.
Levin, D.E., and B.N. Ames (1986) Classifying mutagens as to their specificity in causing six possible transitions and transversions: A simple analysis using the Salmonella mu- tagenicity assay, Environ. Mutagen., 8, 9-28.
Levin, D.E., M. Hollstein, M. Christman, E. Schwiers and B.N. Ames (1982) A new Salmonella tester strain (TA102)
155
with A-T base pairs at the site of mutation detects oxida- tive mutagens, Proc. Natl. Acad. Sci. (U.S.A.), 79, 7445- 7449.
Levy, D., and G.W. Teebor (1991) Site directed substitution of 5-hydroxymethyluracil for thymine in replicating phi-X- 174am3 DNA via synthesis of 5-hydroxymethyl-2'-de- oxyuridine-5'-triphosphate, Nucleic Acids Res., 19, 3337- 3343.
Lewis, J.G., and D.O. Adams (1985) Induction of 5,6-ring saturated thymine bases in NIH-3T3 cells by phorbol es- ter-stimulated macrophages: role of reactive oxygen inter- mediates, Cancer Res., 45, 1270-1275.
Maki, H., and M. Sekiguchi (1992) MutT protein specifically hydrolyzes a potent mutagenic substrate for DNA synthe- sis, Nature (London), 355, 273-275.
Maron, D.M., and B.N. Ames (1983) Revised methods for the Salmonella mutagenicity test, Mutation Res., 113, 173-215.
McCann, J., N.E. Springarn,, J. Kobori and B.N. Ames (1975) Detection of carcinogens as mutagens: Bacterial tester strains with R factor plasmids, Proc. Natl. Acad. Sci. (U.S.A.), 72, 979-983.
Meldrum, J.B., V.S. Gupta, N.R. Lowes and A.R.P. Peterson (1985) Toxicologic and antitumor studies on 5-hydroxy- methyl deoxyuridine, Toxicol. Appl. Pharmacol., 79, 423- 435.
Rouet, P., and J.M. Essigmann (1985) Possible role for thymine glycol in the selective inhibition of DNA synthesis on oxidized DNA templates, Cancer Res., 45, 6113-6118.
Scholes, G. (1983) Radiation effects on DNA, Br. J. Radiat., 56, 221-231.
Shibutani, S., M. Takeshita and A.P. Grollman (1991) Inser- tion of specific bases during DNA synthesis past the oxidation-damaged base 8-oxoG, Nature (London), 349, 431-434.
Shirnam~-Mor& L., T.G. Rossman, W. Troll, G.W. Teebor and K. Frenkel (1987) Genetic effects of 5-hydroxymethyl- 2'-deoxyuridine, a product of ionizing radiation, Mutation Res., 178, 177-186.
Simic, M.G., and S.V. Jovanovic (1986) Free radical mecha- nism of DNA base damage, in: M.G. Simic, L. Grossman and A.C. Upton (Eds.), Mechanisms of DNA Damage and Repair, Plenum, New York, pp. 39-69.
T~oule, R. (1987) Radiation-induced DNA damage and its repair, Int. J. Radiation Biol., 51,573-589.
Thomas, H.F., R.M. Herriott, B.S. Hahn and S.Y. Wang (1976) Thymine hydroperoxide as a mediator in ionizing radiation mutagenesis, Nature (London), 259, 341-342.
Tofigh, S., and K. Frenkel (1989) Effect of metals on nucleo- side hydroperoxide, a product of ionizing radiation in DNA, Free Rad. Biol. Med., 7, 131-143.
Troll, W., K. Frenkel and G.W. Teebor (1984) Free oxygen radicals: Necessary contributors to tumor promotion and cocarcinogenesis, in: H. Fujiki, E. Hecker, R.E. Moore, T. Sugimura and I.B. Weinstein (Eds.), Cellular Interactions by Environmental Tumor Promoters, Japan Scientific So- cieties Press, Tokyo, pp. 207-218.
Vuillaume, M. (1987) Reduced oxygen species, mutation, in- duction and cancer initiation, Mutation Res., 186, 43-47.

156
Wang, S.Y., B.S. Hahn, R.P. Batzinger and E. Bueding (1979) Mutagenic activities of hydroperoxythymine derivatives, products of radiation and oxidation reactions, Biochem. Biophys. Res. Commun., 89, 259-263.
Waschke, S., J. Reefschlager, D. Barwolff and P. Langen (1975) 5-Hydroxymethyl-2'-deoxyuridine, a normal DNA constituent in certain Bacillus subtilis phages is cytostatic for mammalian cells, Nature (London), 255, 629-630.
Wataya, Y., D.V. Santi and C. Hansch (19771 Inhibition of Lactobacillus casei thymidylate synthetase by 5-substituted 2'-deoxyuridylates, Preliminary quantitative structure-ac- tivity relationship, J. Med. Chem., 20, 1469-1473.
Wei, H., and K. Frenkel (1991) In vivo formation of oxidized DNA bases in tumor promoter-treated mouse skin, Cancer Res., 5l, 4443-4449.
Weitzman, S.A., and L.I. Gordon (19901 Inflammation and cancer: role of phagocyte-generated oxidants in carcino- genesis, Blood, 76, 655-663.
Wood, M.L., M. Dizdaroglu, E. Gajewski and J.M. Essigmann (1990) Mechanistic studies of ionizing radiation and oxida- tive mutagenesis: Genetic effects of a single 8-hydroxy- guanine (7-hydro-8-oxoguanine) residue inserted at a unique site in a viral genome, Biochemistry, 29, 7(124-7032.
Zhong, Z., W. Troll, K.L. Koenig and K. Frenkel (199/)) Carcinogenic sulfide salts of nickel and cadmium induce H20 2 formation by human polymorphonuclear leukocytes, Cancer Res., 50, 7564-7570.
Communicated by H. Hayatsu





























