Embed Size (px)
Citation preview

Vol. 171, No. 2JOURNAL OF BACTERIOLOGY, Feb. 1989, p. 1118-11250021-9193/89/021118-08$02.00/0Copyright © 1989, American Society for Microbiology
Mechanism of L-Glutarmate Transport in Membrane Vesicles fromBacillus stearothermophilus
W. DE VRIJ,* R. A. BULTHUIS, P. R. VAN IWAARDEN, AND W. N. KONINGS
Department of Microbiology, University of Groningen, Kerklaan 30, 9751 NN Haren, The Netherlands
Received 27 June 1988/Accepted 14 November 1988
In the presence of electrochemhical energy, several branched-chain neutral and acidic amino acids were foundto accumulate in membrane vesicles of Bacillus stearothermophilus. The membrane vesicles contained astereo-specific transport system for the acidic amino acids L-glutamate and L-aspartate, which could nottranslocate their respective amines, L-glutamihe and L-asparagine. The transport system was thermostable (T1= 700C) and showed highest activities at elevated temperatures (60 to 65°C). The membrane potential or pHgradient could act as the driving force for L-glutamate uptake, which indicated that the transport process ofL-glutamate is electrogenic and that protons are involved in the translocation process. The electrogeniccharacter implies that the anionic L-glutamate is cotransported with at least two monovalent cations. Todetermine the mechanistic stoichiometry of L-glutamate transport and the nature of the cotranslocated cations,the relationship between the components of the proton motive force and .the chemical gradient of L-glutamatewas investigated at different external pH values in the absence and presence of ionophores. In the presence ofeither a membrane potential or a pH gradient, the chemical gradient of L-glutamate was equivalent to thatspecific gradient at different pH values. These results cannot be explained by cotransport of L-glutamate withtwo protons, assuming thermodynamic equilibrium between the driving force for uptake and the chemicalgradient of the substrate. To determine the character, of the cotranslocated cations, L-glutamate uptake wasmonitored with artificial gradients. It was established that either the membrane potential, pH gradient, orchemical gradient of sodium ions could act as the driving force for L-glutamate uptake, which indicated thatL-glutamate most likely is cotranslocated in symport with one proton and one sodium ion.
Recently, several studies have been performed on thermo-philic bacteria, mainly because the metabolic activity ofthese microorganisms at elevated temperatures and the(thermo)stability of their proteins can be applicable forbiotechnological purposes. These studies involve extracellu-lar hydrolytic enzymes, which are frequently used in indus-trial processes, as well as intracellular and membrane-associated proteins (1). An important feature of membraneproteins from thermophilic bacteria is their greater stabilityin the presence of various dissociating agents and detergents(16, 26, 27). This has been demonstrated for energy-trans-ducing enzymes such as H+-ATPase (16), cytochrome coxidase (26, 27), and the alanine transport system (12, 14)isolated from the thermophilic bacterium PS-3. Thermophilicbacteria therefore offer distinct advantages for the purifica-tion and biochemical characterization of membrane-associ-ated proteins. This aspect has been given limited attentionwith respect to solute transport proteins in thermophilicmicroorganisms (11, 12, 14, 22, 28). An advanced stage offunctional characterization has been reached only with theL-alanine transport system from the thermophile PS-3, abacterium that strongly resembles Bacillus stearothermophi-lus. The alanine transport system has been purified tohomogeneity and reconstituted in proteoliposomes. It wasdemonstrated that transport of alanine occurred via an Na'symport mechanism (15). Goto et al. (9) noted Na+/H'exchange activity in PS-3 which may be involved in thegeneration of an Na' gradient required for Na' symportsystems in this organism. Sodium solute symport is not anunusual feature. Sodium is the predominant cotransportedcation for solute transport in marine and halophilic bacteriaand obligate alkalophiles (4). High-affinity Na' solute trans-
* Corresponding author.
port has been reported frequently, even in the enterobacte-rium Escherichia coli, for serine, proline, and glutamate (4,7, 8). Na+ solute symporters with low affinity for sodiumhave been identified just recently in the thermophilic fermen-tative bacterium Clostridium fervidus (G. Speelmans, un-published results) and the ruminal bacterium Streptococcusbovis (23).
In this study, membrane vesicles from the thermophilicbacterium B. stearothermophilus were used to study aminoacid transport. Isolated bacterial membrane vesicles havebeen shown to be excellent model systems for studies ofsolute transport (5, 6, 10, 17, 19). The properties of theenzymes involved in energy transduction of this organismhave already been extensively studied (3). It was found thatthe efficiency of energy transduction declined substantiallyat elevated temperatures due to high proton permeability.Nevertheless, B. stearothermophilus membrane vesicleswere able to maintain a high proton motive force at elevatedtemperatures as a result of extremely high respiratory activ-ity (3). In our study, the functional properties of the L-glutamate transport system were characterized, with specialemphasis on the mechanistic stoichiometry, temperaturedependence, and thermostability of this transport protein.
MATERIALS AND METHODSCell growth and preparation of membrane vesicles. B.
stearothermophilus ATCC 7954 was grown at 55°C withvigorous aeration in a medium containing 2% (wt/vol) tryp-tone, 1% (wt/vol) yeast extract, and 1% (wt/vol) NaCI.Logarithmically growing cells were harvested at an A6. of1.5 to 2.0.Membrane vesicles were prepared by a slight modification
of the procedure of Konings et al. (17). The incubationtemperature of the cells in the presence of lysozyme was
1118
on Novem
ber 8, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
L-GLUTAMATE TRANSPORT IN B. STEAROTHERMOPHILUS 1119
increased to 500C to facilitate cell wall breakdown. Themembrane preparations (10 to 20 mg of membrane proteinper ml) were suspended in 50 mM potassium phosphate (pH6.6), rapidly frozen, and stored in liquid nitrogen.Uptake of amino acid in membrane vesicles. Amino acid
transport was measured as described previously (19). Theassay mixture contained 0.1 ml of oxygen-saturated 50 mMpotassium phosphate adjusted to the appropriate pH and 10mM MgSO4. Ascorbate (20 mM) and N,N,N',N'-tetra-methyl-p-phenylenediamine (TMPD) (100 FiM) were used asthe electron donor system. Uptake experiments were carriedout at 40'C unless stated otherwise. Membrane vesicles wereincubated at this temperature for 1 min before addition of theelectron donor.
L-Glutamate counterflow activity of membrane vesicleswas measured by preequilibration of membrane vesicles (25mg of protein per ml) with 10 mM L-[12C]glutamate in thepresence of 1 nmol of valinomycin per mg of membraneprotein. After 1 h of incubation at 30'C, the membranevesicles were diluted 200-fold into 50 mM potassium phos-phate (pH 6.5-10 mM MgSO4 supplemented with L-[I4C]glutamate (final concentration, 53.6 piM). Uptake ofL-[14C]glutamate was followed at various temperatures.Transport of L-glutamate driven by artificial gradients was
measured by using membrane vesicles (20 mg of membraneprotein per ml) suspended in 20 mM morpholineethanesulfo-nic acid (MES)-KOH (pH 6.0) and 100 mM potassiumacetate. The transmembrane chemical gradient of sodiumruns (APNa) was generated upon 100-fold dilution of mem-brane vesicles into 20 mM MES-NaOH (pH 6.0-100 mMsodium acetate-10 mM MgSO4. AiNa and transmembraneelectrical potential (Aqi) were generated by dilution of vali-nomycin-preincubated membrane vesicles (2 nmol/mg ofprotein) into 20 mM MES-NaOH (pH 6.0-100 mM sodiumacetate-10 mM MgS04. AjiNa and transmembrane protongradient (ApH) were generated by 100-fold dilution of mem-brane vesicles into 120 mM MES-NaOH (pH 6.0)-10 mMMgSO4. ApH was generated by 100-fold dilution of mem-brane vesicles into 20 mM choline-MES (pH 6.0)-100 mMcholine chloride-10 mM MgSO4. Aq, was generated by100-fold dilution into 20 mM choline-MES (pH 6.0)-100 mMcholine chloride-10 mM MgSO4 of membranes that had beenloaded with 120 mM MES-KOH (pH 6.0) and preincubatedwith valinomycin. Proton motive force (Ap) was generatedby 100-fold dilution of valinomycin-preincubated vesiclesinto 20 mM choline-MES (pH 6.0)-100 mM choline chloride-100 mM MgSO4. Ap and Ai!Na were generated by dilution ofvalinomycin-incubated vesicles into 120 mM MES-NaOH(pH 6.0)-10 mM MgSO4.
Control experiments were performed by 100-fold dilutionin the same buffer in which the membrane vesicles weresuspended. Uptake of L-glutamate was measured at 45°C bysupplementing the dilution medium with L-[U-14C]glutamate(final concentration, 1.75 ,uM). At given time intervals, 2 mlof ice-cold 0.1 M KCI was added, and the samples werefiltered immediately over 0.45-jim-pore-size cellulose-nitratefilters (Millipore Corp., Bedford, Mass.). Filters werewashed with 2 ml of 0.1 M KCl, dried, and transferred toscintillation vials; 4 ml of scintillation fluid was then added.Radioactivity was measured with a liquid scintillationcounter (Tris-Carb 460 CD; Packard Instruments Co., Inc.,Rockville, Md.).
Determination of transmembrane gradients. The mem-brane potential (interior negative) was determined from thedistribution of the tetraphenylphosphonium ion across themembrane by using an electrode selective for tetraphenyl-
phosphonium ion (25). The reaction mixture (1 ml) contained50 mM potassium phosphate of appropriate pH, 10 mMMgSO4, 4 FLM tetraphenylphosphonium ion, and membranevesicles (0.1 mg of membrane protein per ml). Ascorbate (20mM) and TMPD (100 puM) were added as the electron donorsystem. Nigericin and valinomycin were used at final con-centrations of 100 and 300 nM, respectively. The experi-ments were performed at 40'C. The membrane potential wasdetermined by applying a correction for concentration-de-pendent probe binding according to the model described byLolkema et al. (20). For calculation of the membrane poten-tial, an internal volume of 3 pA per mg of membrane proteinwas used. Qualitative measurements of membrane potentialwere performed by an alternative method based on use of themembrane potential indicator 3,3'-dipropylthiocarbocyanineiodide by measuring membrane potential-dependent absorb-ance changes (AA683-660) as described previously (3).The transmembrane pH gradient (interior alkaline) was
determined from the fluorescence changes of pyranine en-trapped within the membrane vesicles (2). Measurementswere performed at 40'C in an MPF-4 spectrophotofluorim-eter (The Perkin-Elmer Corp., Norwalk, Conn.), using ex-citation and emission wavelengths of 460 and 508 nm.Pyranine was entrapped in membrane vesicles by addingpyranine (final concentration, 200 VLM) to the membranevesicles (10 mg of membrane protein per ml) before freeze-thaw sonication. External pyranine was removed by chro-matography of the membrane suspension over SephadexG-25 (course grade, 1 by 20 cm) preequilibrated with 50 mMpotassium phosphate of appropriate pH and 10 mM MgSO4.The reaction mixture contained 50 mM potassium phosphateof desired pH, 10 mM MgSO4, and membranes (0.1 to 0.2 mg
160
120
5
'j)w
0Ia
80
40
5.5 6.0 6.5 7.0 7.5 8.0
pH external
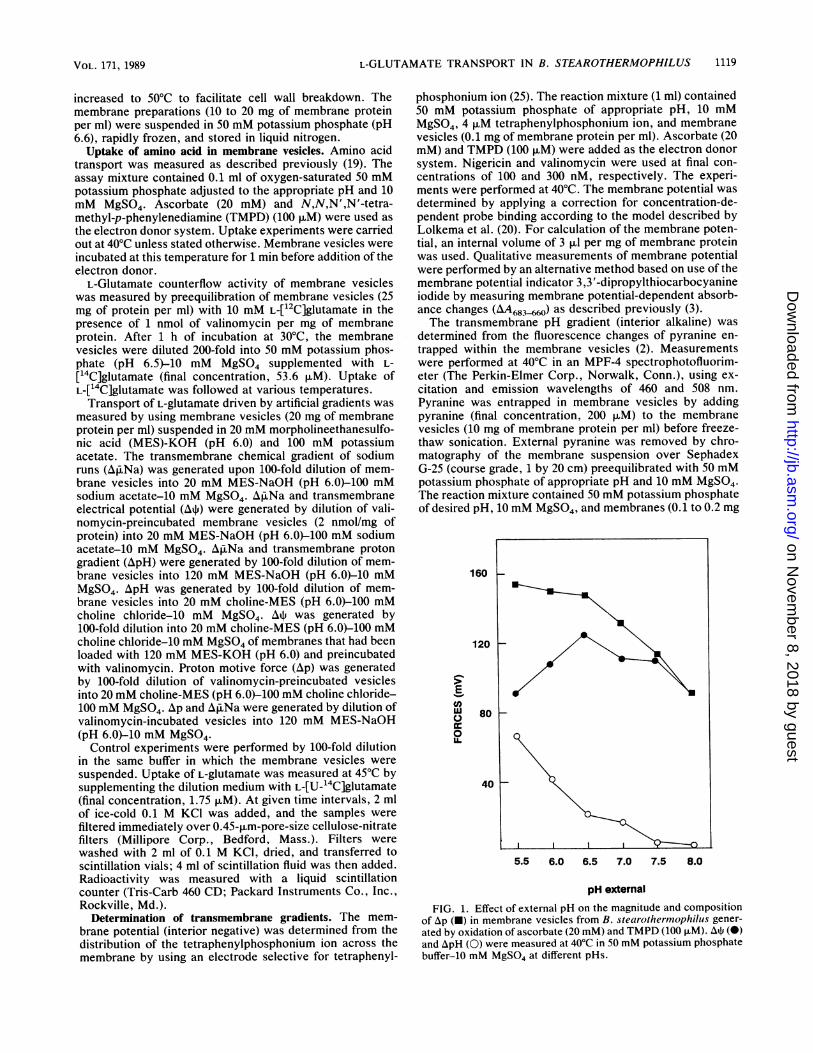
FIG. 1. Effect of external pH on the magnitude and compositionof Ap (U) in membrane vesicles from B. stearothermophilus gener-ated by oxidation of ascorbate (20 mM) and TMPD (100 [iM). AtP (0)and ApH (0) were measured at 40'C in 50 mM potassium phosphatebuffer-10 mM MgSO4 at different pHs.
VOL. 171, 1989
on Novem
ber 8, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
1120 DE VRIJ ET AL.
00Ch
E0
0JwEo£5
6
4
2
1 2 3 4 5
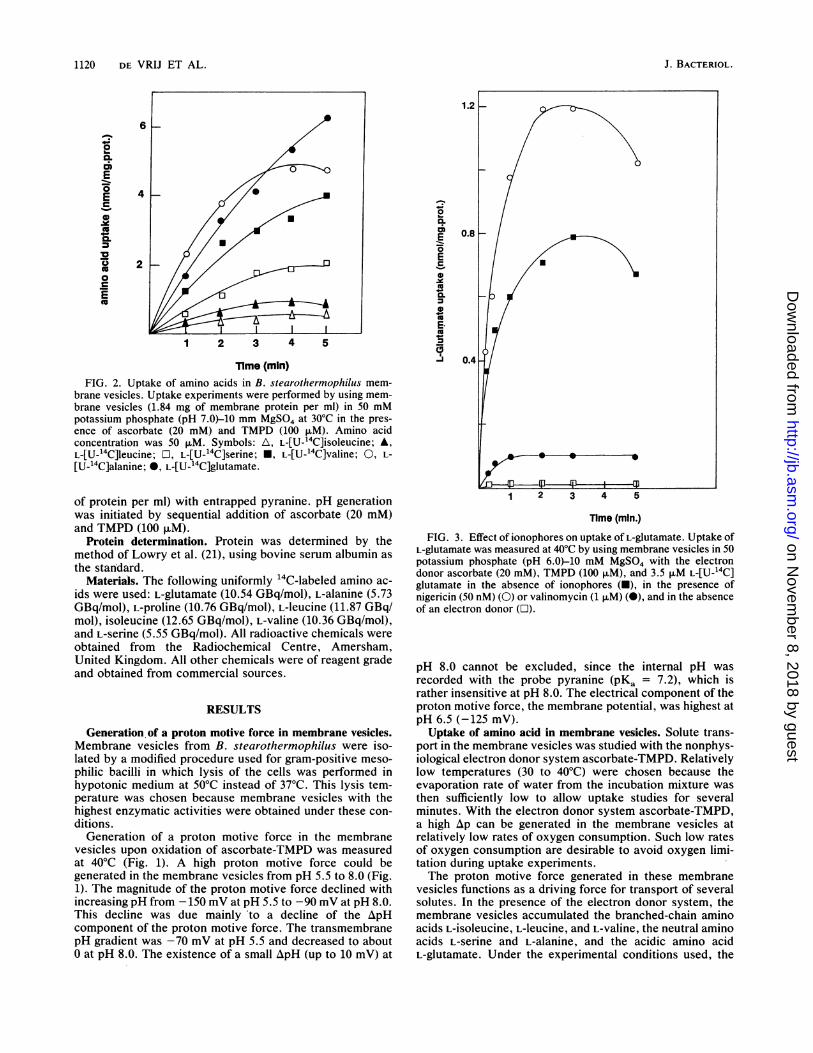
Time (min)FIG. 2. Uptake of amino acids in B. stearothermophilus mem-
brane vesicles. Uptake experiments were performed by using mem-brane vesicles (1.84 mg of membrane protein per ml) in 50 mMpotassium phosphate (pH 7.0)-10 mm MgSO4 at 30°C in the pres-ence of ascorbate (20 mM) and TMPD (100 p.M). Amino acidconcentration was 50 p.M. Symbols: A, L-[U-'4C]isoleucine; A,L-[U-14C]jeucine; Li, L-[U-14C]serine; *, L-[U-14C]valine; 0, L-
[U-_4C]alanine; 0, L-[U-'4C]glutamate.
of protein per ml) with entrapped pyranine. pH generationwas initiated by sequential addition of ascorbate (20 mM)and TMPD (100 IxM).
Protein determination. Protein was determined by themethod of Lowry et al. (21), using bovine serum albumin asthe standard.
Materials. The following uniformly 14C-labeled amino ac-ids were used: L-glutamate (10.54 GBq/mol), L-alanine (5.73GBq/mol), L-proline (10.76 GBq/mol), L-leucine (11.87 GBq/mol), isoleucine (12.65 GBq/mol), L-valine (10.36 GBq/mol),and L-serine (5.55 GBq/mol). All radioactive chemicals were
obtained from the Radiochemical Centre, Amersham,United Kingdom. All other chemicals were of reagent gradeand obtained from commercial sources.
RESULTS
Generation of a proton motive force in membrane vesicles.Membrane vesicles from B. stearothermophilus were iso-lated by a modified procedure used for gram-positive meso-
philic bacilli in which lysis of the cells was performed inhypotonic medium at 50°C instead of 37°C. This lysis tem-perature was chosen because membrane vesicles with thehighest enzymatic activities were obtained under these con-
ditions.Generation of a proton motive force in the membrane
vesicles upon oxidation of ascorbate-TMPD was measuredat 40°C (Fig. 1). A high proton motive force could begenerated in the membrane vesicles from pH 5.5 to 8.0 (Fig.1). The magnitude of the proton motive force declined withincreasing pH from -150 mV at pH 5.5 to -90 mV at pH 8.0.This decline was due mainly to a decline of the ApHcomponent of the proton motive force. The transmembranepH gradient was -70 mV at pH 5.5 and decreased to about0 at pH 8.0. The existence of a small ApH (up to 10 mV) at
0)0.8
Ch0 .E0CLC
0.4
1 2 3 4 5
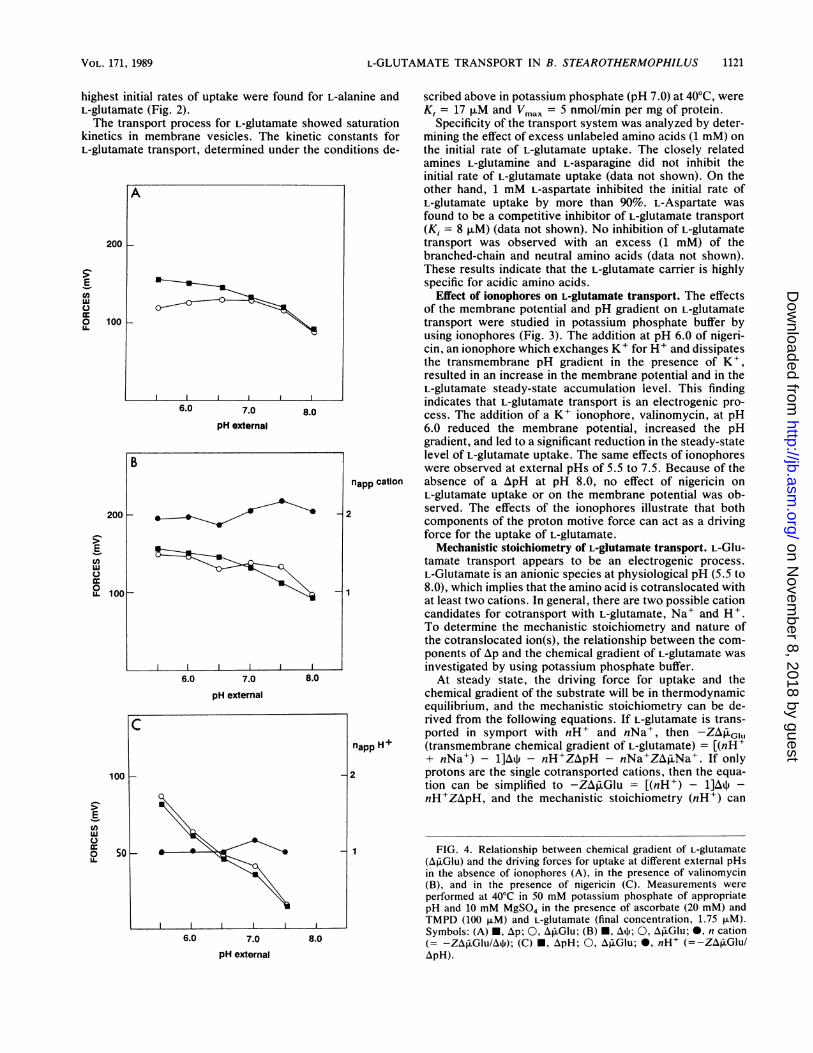
Time (min.)FIG. 3. Effect of ionophores on uptake of L-glutamate. Uptake of
L-glutamate was measured at 40'C by using membrane vesicles in 50potassium phosphate (pH 6.0)-10 mM MgSO4 with the electrondonor ascorbate (20 mM), TMPD (100 ,uM), and 3.5 ,uwM L-[U-14C]glutamate in the absence of ionophores (L), in the presence ofnigericin (50 nM) (0) or valinomycin (1 ,uM) (L), and in the absenceof an electron donor (O).
pH 8.0 cannot be excluded, since the internal pH wasrecorded with the probe pyranine (pKa = 7.2), which israther insensitive at pH 8.0. The electrical component of theproton motive force, the membrane potential, was highest atpH 6.5 (-125 mV).Uptake of amino acid in membrane vesicles. Solute trans-
port in the membrane vesicles was studied with the nonphys-iological electron donor system ascorbate-TMPD. Relativelylow temperatures (30 to 40°C) were chosen because theevaporation rate of water from the incubation mixture wasthen sufficiently low to allow uptake studies for severalminutes. With the electron donor system ascorbate-TMPD,a high Ap can be generated in the membrane vesicles atrelatively low rates of oxygen consumption. Such low ratesof oxygen consumption are desirable to avoid oxygen limi-tation during uptake experiments.The proton motive force generated in these membrane
vesicles functions as a driving force for transport of severalsolutes. In the presence of the electron donor system, themembrane vesicles accumulated the branched-chain aminoacids L-isoleucine, L-leucine, and L-valine, the neutral aminoacids L-serine and L-alanine, and the acidic amino acidL-glutamate. Under the experimental conditions used, the
J. BACTERIOL.
on Novem
ber 8, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
L-GLUTAMATE TRANSPORT IN B. STEAROTHERMOPHILUS 1121
highest initial rates of uptake were found for L-alanine andL-glutamate (Fig. 2).The transport process for L-glutamate showed saturation
kinetics in membrane vesicles. The kinetic constants forL-glutamate transport, determined under the conditions de-
200 k
E
0U-
IL 100 k
200
E
wcn.
0U.
6.0 7.0 8.0
pH external
1001
100
Eu,w
0
0U-
SC
6.0 7.0 8.0
pH external
C
I
2
1
6.0 7.0 8.0
pH external
scribed above in potassium phosphate (pH 7.0) at 40'C, wereK, = 17 ILM and Vmax = 5 nmol/min per mg of protein.
Specificity of the transport system was analyzed by deter-mining the effect of excess unlabeled amino acids (1 mM) onthe initial rate of L-glutamate uptake. The closely relatedamines L-glutamine and L-asparagine did not inhibit theinitial rate of L-glutamate uptake (data not shown). On theother hand, 1 mM L-aspartate inhibited the initial rate ofL-glutamate uptake by more than 90%. L-Aspartate wasfound to be a competitive inhibitor of L-glutamate transport(Ki = 8 1iM) (data not shown). No inhibition of L-glutamatetransport was observed with an excess (1 mM) of thebranched-chain and neutral amino acids (data not shown).These results indicate that the L-glutamate carrier is highlyspecific for acidic amino acids.
Effect of ionophores on L-glutamate transport. The effectsof the membrane potential and pH gradient on L-glutamatetransport were studied in potassium phosphate buffer byusing ionophores (Fig. 3). The addition at pH 6.0 of nigeri-cin, an ionophore which exchanges K+ for H' and dissipatesthe transmembrane pH gradient in the presence of K+,resulted in an increase in the membrane potential and in theL-glutamate steady-state accumulation level. This findingindicates that L-glutamate transport is an electrogenic pro-cess. The addition of a K+ ionophore, valinomycin, at pH6.0 reduced the membrane potential, increased the pHgradient, and led to a significant reduction in the steady-statelevel of L-glutamate uptake. The same effects of ionophoreswere observed at external pHs of 5.5 to 7.5. Because of theabsence of a ApH at pH 8.0, no effect of nigericin onL-glutamate uptake or on the membrane potential was ob-served. The effects of the ionophores illustrate that bothcomponents of the proton motive force can act as a drivingforce for the uptake of L-glutamate.
Mechanistic stoichiometry of L-glutamate transport. L-Glu-tamate transport appears to be an electrogenic process.L-Glutamate is an anionic species at physiological pH (5.5 to8.0), which implies that the amino acid is cotranslocated withat least two cations. In general, there are two possible cationcandidates for cotransport with L-glutamate, Na' and H'.To determine the mechanistic stoichiometry and nature ofthe cotranslocated ion(s), the relationship between the com-ponents of Ap and the chemical gradient of L-glutamate wasinvestigated by using potassium phosphate buffer.At steady state, the driving force for uptake and the
chemical gradient of the substrate will be in thermodynamicequilibrium, and the mechanistic stoichiometry can be de-rived from the following equations. If L-glutamate is trans-ported in symport with nH+ and nNa+, then -Z4iiGI.(transmembrane chemical gradient of L-glutamate) = [(nH++ nNa+) - 1]Aqi - nH+ZApH - nNa+ZAfrNa+. If onlyprotons are the single cotransported cations, then the equa-tion can be simplified to -ZAPGlu = [(nH+) - 1]>i\ -nH+ZApH, and the mechanistic stoichiometry (nH+) can
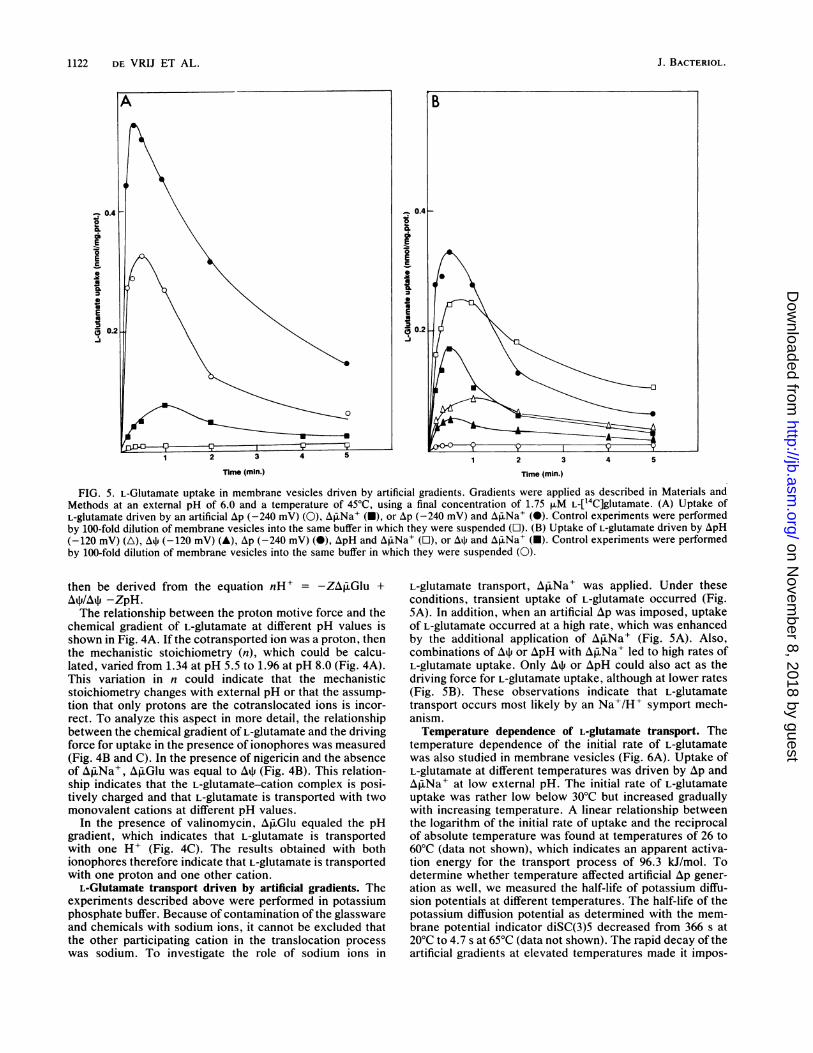
FIG. 4. Relationship between chemical gradient of L-glutamate(A Glu) and the driving forces for uptake at different external pHsin the absence of ionophores (A), in the presence of valinomycin(B), and in the presence of nigericin (C). Measurements were
performed at 40°C in 50 mM potassium phosphate of appropriatepH and 10 mM MgSO4 in the presence of ascorbate (20 mM) andTMPD (100 p.M) and L-glutamate (final concentration, 1.75 ,uM).Symbols: (A) E, Ap; 0, AjiGlu; (B) *, Ati; 0, A Glu; *, n cation(= -ZA~iGluI/At); (C) *, ApH; 0, Ap.Glu; *, nH+ (=-ZAfrGlu/ApH).
A
B
_,,II
VOL. 171, 1989
1
on Novem
ber 8, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
1122 DE VRIJ ET AL.
E
0.
Ei-I
a.
0E-S
CI
aID-i
Time (min.)2 3 4 5
Time (min.)
FIG. 5. L-Glutamate uptake in membrane vesicles driven by artificial gradients. Gradients were applied as described in Materials andMethods at an external pH of 6.0 and a temperature of 450C, using a final concentration of 1.75 p.M L-[14C]glutamate. (A) Uptake ofL-glutamate driven by an artificial Ap (-240 mV) (0), AjINa' (U), or Ap (-240 mV) and A Na' (0). Control experiments were performedby 100-fold dilution of membrane vesicles into the same buffer in which they were suspended (L). (B) Uptake of L-glutamate driven by ApH(-120 mV) (A), Aip (-120 mV) (A), Ap (-240 mV) (0), ApH and APNa' (E), or Aip and AjiNa' (U). Control experiments were performedby 100-fold dilution of membrane vesicles into the same buffer in which they were suspended (0).
then be derived from the equation nH' = -ZAfiGlu +Aq4/AP -ZpH.The relationship between the proton motive force and the
chemical gradient of L-glutamate at different pH values isshown in Fig. 4A. If the cotransported ion was a proton, thenthe mechanistic stoichiometry (n), which could be calcu-lated, varied from 1.34 at pH 5.5 to 1.96 at pH 8.0 (Fig. 4A).This variation in n could indicate that the mechanisticstoichiometry changes with external pH or that the assump-tion that only protons are the cotranslocated ions is incor-rect. To analyze this aspect in more detail, the relationshipbetween the chemical gradient of L-glutamate and the drivingforce for uptake in the presence of ionophores was measured(Fig. 4B and C). In the presence of nigericin and the absenceof AfrNa+, ApiGlu was equal to Ali (Fig. 4B). This relation-ship indicates that the L-glutamate-cation complex is posi-tively charged and that L-glutamate is transported with twomonovalent cations at different pH values.
In the presence of valinomycin, AiJGlu equaled the pHgradient, which indicates that L-glutamate is transportedwith one H' (Fig. 4C). The results obtained with bothionophores therefore indicate that L-glutamate is transportedwith one proton and one other cation.
L-Glutamate transport driven by artificial gradients. Theexperiments described above were performed in potassiumphosphate buffer. Because of contamination of the glasswareand chemicals with sodium ions, it cannot be excluded thatthe other participating cation in the translocation processwas sodium. To investigate the role of sodium ions in
L-glutamate transport, AjiNa' was applied. Under theseconditions, transient uptake of L-glutamate occurred (Fig.5A). In addition, when an artificial Ap was imposed, uptakeof L-glutamate occurred at a high rate, which was enhancedby the additional application of AjiNa' (Fig. 5A). Also,combinations of Aqj or ApH with AfrNa' led to high rates ofL-glutamate uptake. Only Al\ or ApH could also act as thedriving force for L-glutamate uptake, although at lower rates(Fig. SB). These observations indicate that L-glutamatetransport occurs most likely by an Na+/H' symport mech-anism.Temperature dependence of L-glutamate transport. The
temperature dependence of the initial rate of L-glutamatewas also studied in membrane vesicles (Fig. 6A). Uptake ofL-glutamate at different temperatures was driven by Ap andApNa' at low external pH. The initial rate of L-glutamateuptake was rather low below 30'C but increased graduallywith increasing temperature. A linear relationship betweenthe logarithm of the initial rate of uptake and the reciprocalof absolute temperature was found at temperatures of 26 to60'C (data not shown), which indicates an apparent activa-tion energy for the transport process of 96.3 kJ/mol. Todetermine whether temperature affected artificial Ap gener-ation as well, we measured the half-life of potassium diffu-sion potentials at different temperatures. The half-life of thepotassium diffusion potential as determined with the mem-brane potential indicator diSC(3)5 decreased from 366 s at20'C to 4.7 s at 650C (data not shown). The rapid decay of theartificial gradients at elevated temperatures made it impos-
J. BACTERIOL.
on Novem
ber 8, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
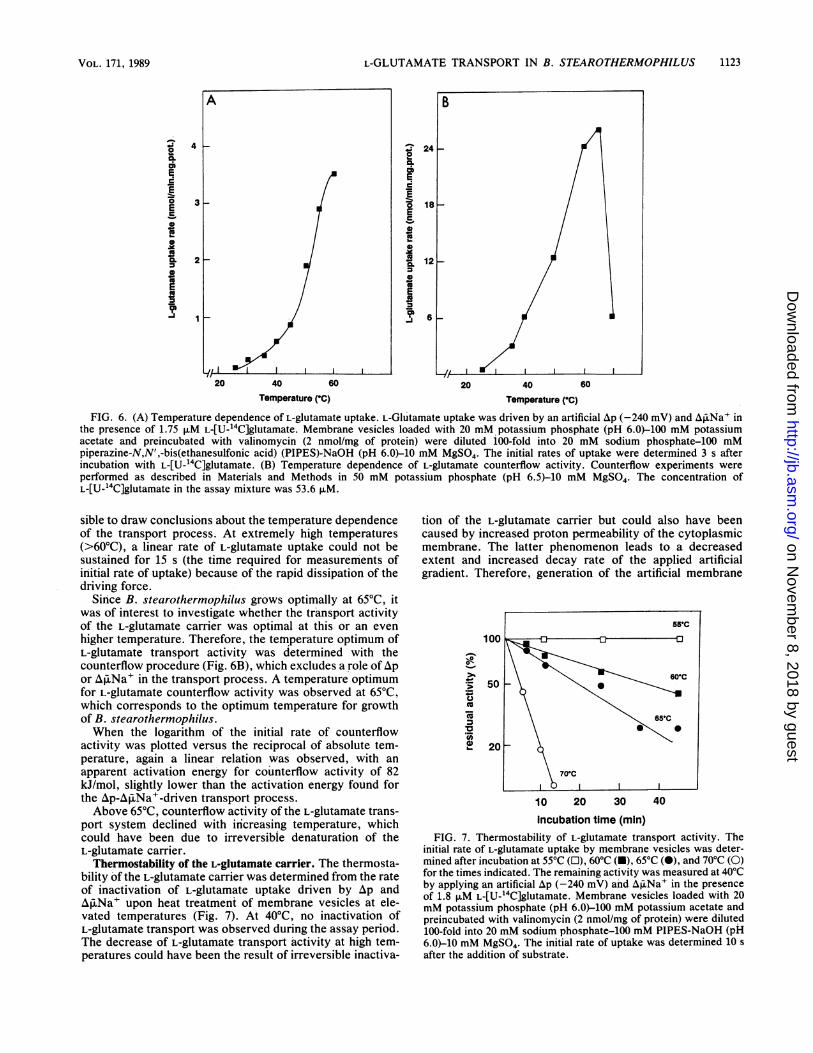
L-GLUTAMATE TRANSPORT IN B. STEAROTHERMOPHILUS
20 40 60
Temperature (,C)
0.
C.
Ed
-
0.
0
I
s
20 40 60
Temperature (,C)
FIG. 6. (A) Temperature dependence of L-glutamate uptake. L-Glutamate uptake was driven by an artificial Ap (-240 mV) and AliNa' inthe presence of 1.75 jiM L-[U-14C]glutamate. Membrane vesicles loaded with 20 mM, potassium phosphate (pH 6.0)-100 mM potassiumacetate and preincubated with valinomycin (2 nmol/mg of protein) were diluted 100-fold into 20 mM sodium phosphate-100 mMpiperazine-N,N',-bis(ethanesulfonic acid) (PIPES)-NaOH (pH 6.0)-10 mM MgSO4. The initial rates of uptake were determined 3 s afterincubation with L-[U-14C]glutamate. (B) Temperature dependence of L-glutamate counterflow activity. Counterflow experiments were
performed as described in Materials and Methods in 50 mM potassium phosphate (pH 6.5-10 mM MgSO4. The concentration ofL-[U-14C]glutamate in the assay mixture was 53.6 p.M.
sible to draw conclusions about the temperature dependenceof the transport process. At extremely high temperatures(>60'C), a linear rate of L-glutamate uptake could not besustained for 15 s (the time required for measurements ofinitial rate of uptake) because of the rapid dissipation of thedriving force.
Since B. stearothermophilus grows optimally at 650C, itwas of interest to investigate whether the transport activityof the L-glutamate carrier was optimal at this or an evenhigher temperature. Therefore, the temperature optimum ofL-glutamate transport activity was determined with thecounterflow procedure (Fig. 6B), which excludes a role ofApor APiNa' in the transport process. A temperature optimumfor L-glutamate counterflow activity was observed at 650C,which corresponds to the optimum temperature for growthof B. stearothermophilus.When the logarithm of the initial rate of counterflow
activity was plotted versus the reciprocal of absolute tem-perature, again a linear relation was observed, with anapparent activation energy for counterflow activity of 82kJ/mol, slightly lower than the activation energy found forthe Ap-Aj3pNa'-driven transport process.Above 650C, counterflow activity of the L-glutamate trans-
port system declined with increasing temperature, whichcould have been due to irreversible denaturation of theL-glutamate carrier.
Thermostability of the L-glutamate carrier. The thermosta-bility of the L-glutamate carrier was determined from the rateof inactivation of L-glutamate uptake driven by Ap andAPNa' upon heat treatment of membrane vesicles at ele-vated temperatures (Fig. 7). At 40'C, no inactivation ofL-glutamate transport was observed during the assay period.The decrease of L-glutamate transport activity at high tem-peratures could have been the result of irreversible inactiva-
tion of the L-glutamate carrier but could also have beencaused by increased proton permeability of the cytoplasmicmembrane. The latter phenomenon leads to a decreasedextent and increased decay rate of the applied artificialgradient. Therefore, generation of the artificial membrane
A
(N
0-'aa)
L-
100
50
20
10 20 30 40
Incubation time (min)
FIG. 7. Thermostability of L-glutamate transport activity. Theinitial rate of L-glutamate uptake by membrane vesicles was deter-mined after incubation at 550C (1), 60'C (U), 650C (0), and 70'C (0)for the times indicated. The remaining activity was measured at 400Cby applying an artificial Ap (-240 mV) and APiNa' in the presenceof 1.8 p.M L-[U-_4C]glutamate. Membrane vesicles loaded with 20mM potassium phosphate (pH 6.0)-100 mM potassium acetate andpreincubated with valinomycin (2 nmol/mg of protein) were diluted100-fold into 20 mM sodium phosphate-100 mM PIPES-NaOH (pH6.0)-10 mM MgSO4. The initial rate of uptake was determined 10 s
after the addition of substrate.
9C.E0E
-6
S
Eu0.
0'UaILz1
4
3
2
I
1123VOL. 171, 1989
on Novem
ber 8, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

1124 DE VRIJ ET AL.
potential was monitored in these membrane vesicles bymeasuring absorbance changes of the membrane potentialindicator diSC(3)5 (data not shown). Incubation of mem-brane vesicles at a temperature of 70'C or higher led to asignificant decrease in the generated membrane potential anda higher decay tate of the gradient, This finding indicates anirreversible increase in the passive ion permeability of thecytoplasmic membrane at these extremely high tempera-tures. However, incubation of the membrane vesicles atlower temperatures (<70'C) did not significantly affect thegeneration and extent of the applied membrane potential.Therefore, inactivation observed at temperatures lower than70'C reflects the thermostability of the L-glutamate transportprotein in isolated membranes. An inactivation temperature(Ti), defined as the temperature at which enzyme activitywithin 10 min of incubation is reduced to 50% of approxi-mately 70'C, can be derived.
DISCUSSION
This study demonstrates that in membrane vesicles iso-lated from B. stearotherrnophilus a high proton motive forcecan be generated, which can drive the uptake of severalamino acids in a manner similar to that observed in otherthermophilic bacteria (12, 13, 22, 28, 29).The L-glutamate transport system has been studied in
detail. Competition experiments in membrane vesicles of B.stearothermophilus indicate that L-glutamate is transportedvia an uptake-system specific for acidic amino acids. Theaffinities and specificities for the acidic amino acids in thisorganism are similar to those found in membrane vesicles ofBacillus subtilis and E. coli (18, 24).
Several lines of evidence can be presented which indicatea symport system for Na', H', and L-glutamate in B.stearothermophilus membranes: (i) a membrane potential,ApH, or AfrNa+ can act separately as a driving force foruptake of L-glutamrate; (ii) the relationship between theindividual components of Ap (Aiv or ApH) and AjiGlu indi-cates that L-glutamate is cotranslocated with two cations;and (iii) ApH appears to be equivalent to AQfGlu in theabsence of other gradients, which indicates that L-glutamateis cotransported with one H'. Measurements in which therelationship between the components of Ap and A iGlu wasdetermined were performed in potassium phosphate buffer.Despite the apparent lack of Na' ions, uptake of L-glutamatewas observed. These observations indicate that contaminat-ing sodium is sufficient to enable transport and that theaffinity of the transport system for sodium is high (micro-molar range). Since AiJ and ApH are equivalent drivingforces for L-glutamate uptake at different external pHs, weconclude that no AjJNa' existed under the experimentalconditions used. Goto et al. (9) noted pH-dependent Na+/H'exchange activity in the related thermophilic bacteriumPS-3. The exchanger plays a role in internal pH regulationand conversion of ApH into AfrNa', which can be used forNa' solute symport systems. The exchanger is inactivebelow external pH 7.0, but above pH 7.0 activity increaseswith increase in external pH. This phenomenon is caused bya pH-dependent increase in the affinity for sodium. Even atthe optimum pH (8.5), the affinity for sodium is low (Kl =1.4 mM). If a similar Na+/H' exchange characteristic existsin B. stearothermophilus membranes, no significant A/INa'would have been formed under the conditions used. At lowexternal pH, Ap deviates from AfrGlu. An explanation forthis finding could be thermodynamic disequilibrium between
the driving force for uptake and the chemical gradient of thesubstrate. pH-dependent coupling (in)efficiency, caused byintrinsic (carrier-mediated) or extrinsic (non-carrier-medi-ated) leaks could explain thermodynamic disequilibrium (5,6). However, in the presence of only A* or ApH as thedriving force for uptake, no significant pH-dependent alter-ations in napp were observed, which would argue againstpH-dependent thermodynamic disequilibrium. An explana-tion for this phenomenon remains to be established. Symportof Na', H', and L-glutamate has also been found in E. coliB (7, 8). This carrier has been intensively investigated, and itwas shown that L-glutamate can bind only to an Na'-H'carrier complex that is formed by random-order binding ofNa' and H' to the unloaded carrier. Translocation ofL-glutamate occurs in 1:1 stoichiometry with the two cat-ions, identical to findings for Na'-H+-L-glutamate in B.stearothermophilus (7). Furthermore, the kinetic constantsof L-glutamate transport in E. coli B were greatly affected byalterations in concentrations of either one of the symportedcations (8). It.therefore seems of interest to. study the effectsof pH and of various Na+ concentrations on L-glutamatetransport in B. stearothermophilus. L-Alanine transport inthe thermophilic bacterium PS-3, which strongly resemblesB. stearothermophilus, is mediated by an Na+ symportsystem (11-14)..Possibly other solute transport systems in B.stearothermophilus are Na+ symport systems as well.
Interesting features of the L-glutamate carrier in B. stear-othermophilus membrane vesicles are the temperature de-pendence and thermostability of the protein. The tempera-ture optimum of the L-glutamate carrier has been determinedby measuring coUnterflow activity and corresponds ratherwell to the optimum temperature for growth of the organism.The linearity of the Arrhenius plot of L-glutamate counter-flow activity indicates that the activity of the transportprotein is affected mainly by the temperature and not by themicroviscosity of the hydrophobic environment. Inflectionsin the Arrhenius plot at 20°C have been reported andascribed to a phase transition of the membrane lipid bilayer,as has been observed for alanine and sodium uptake in PS-3membrane vesicles (9, 13). An inflection was not observedfor L-glutamate transport in B. stearothermophilus mem-brane vesicles in the temperature range tested (26 to 60°C).The L-glutamate carrier in B. stearothermophilus was
found to be rather thermostable (T, = 70°C). The heatstability of this transport protein was less than the stability ofthe proteins involved in energy transduction in B. stearo-thermophilus (3). In a previous study, higher thermostabili-ties were found for cytochrome c oxidase (T, = 86°C) andNADH oxidase (T, = 77°C) in membrane vesicles of B.stearothermophilus. Whether the thermostability is deter-mined by intrinsic properties of the enzyme or influenced bythe hydrophobic environment must be elucidated. The hy-drophobic environment can have a pronounced effect on thethermostability of the carrier, as has been reported for theleucine carrier in Thermus thermophilus HB-8 (28). Thisfinding does not account for all membrane-associated pro-teins, as has been demonstrated for cytochrome c oxidasefrom B. stearothermophilus (W. de Vrij, R. A. Bulthuisj andW. N. Konings, Eur. J. Biochem., in press). This enzymehas been found not to be protected against thermoinactiva-tion by its hydrophobic lipid environment (de Vrij et al., inpress).Membranes of B. stearothermophilus irreversibly lose the
capacity to maintain a potassium diffusion potential afterincubation at temperatures higher than 70°C. This irrevers-ible change in membrane integrity occurs at a temperature
J. BACtERIOL.
on Novem
ber 8, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

L-GLUTAMATE TRANSPORT IN B. STEAROTHERMOPHILUS 1125
which corresponds to the maximal growth temperature (T,,,= 70°C). It seems likely, therefore, that the maximumgrowth temperature is determined by the combined effects ofthermoinactivation of intracellular and membrane-associ-ated proteins and loss of integrity of the cytoplasmic mem-
brane.
LITERATURE CITED1. Brock, T. D. (ed.). 1986. Thermophiles, general, molecular and
applied microbiology. John Wiley & Sons, Inc., New York.2. Clement, N. R., and M. J. Gould. 1981. Pyranine(8-hydroxy-
1,3,6-pyrenotrisulfonate) as a probe of internal aqueous hydro-gen ion concentration in phospholipid vesicles. Biochemistry20:1534-1538.
3. de Vrij, W., R. A. Bulthuis, and W. N. Konings. 1988. Compar-ative study of the energy-transducing properties of cytoplasmicmembranes from mesophilic and thermophilic Bacillus species.J. Bacteriol. 170:2359-2366.
4. Dimroth, P. 1987. Sodium ion transport decarboxylases andother aspects of sodium ion cycling in bacteria. Microbiol. Rev.51:320-340.
5. Driessen, A. J. M., K. J. Hellingwerf, and W.N. Konings. 1987.Mechanism of energy coupling to entry and exit of neutral andbranched amino acids in membrane vesicles of Streptococcuscreemoris. J. Biol. Chem. 262:12438-12443.
6. Driessen, A. J. M., J. Kodde, S. De Jong, and W. N. Konings.1987. Neutral amino acid uptake by membrane vesicles ofStreptococcus cremoris is subjected to regulation by the internalpH. J. Bacteriol. 196:2748-2754.
7. Fujimura, T., I. Yamato, and Y. Anraku. 1983. Mechanism ofglutamate transport in Escherichia coli B. 1. Proton dependentand sodium-ion dependent binding of glutamate to a a glutamatecarrier in the cytoplasmic membrane. Biochemistry 22:1954-1959.
8. Fujimura, T., I. Yamato, and Y. Anraku. 1983. Mechanism ofglutamate transport in Escherichia coli B. 2. Kinetics of gluta-mate transport driven by artificially imposed proton and sodiumion gradients across the cytoplasmic membrane. Biochemistry22:1959-1965.
9. Goto, K., Y. Kagawa, and H. Hirata. 1980. A stable Na+/H+antiporter of thermophilic bacterium PS3. J. Bioenerg. Bi-omembr. 12:297-308.
10. Hellingwerf, K. J., and W. N. Konings. 1985. The energy flow inbacteria: the main free-energy intermediates and their regula-tory role. Adv. Microb. Physiol. 26:125-154.
11. Hirata, H., T. Kambe, and Y. Kagawa. 1984. A purified alaninecarrier composed of a single polypeptide from thermophilicbacterium PS3 driven by either proton or sodium gradient. J.Biol. Chem. 259:10653-10656.
12. Hirata, H., N. Sone, H. Yoshida, and Y. Kagawa. 1976. Solubi-lization and partial purification of alanine carrier from mem-
branes of a thermophilic bacterium and its reconstitution infunctional vesicles. Biochem. Biophys. Res. Commun. 69:655-671.
13. Hirata, H., N. Sone, M. Yoshida, and Y. Kagawa. 1976. Activetransport of alanine by thermostable membrane vesicles isolatedfrom a thermophilic bacterium. J. Biol. Chem. 79:1157-1166.
14. Hirata, H., N. Sone, H. Yoshida, and Y. Kagawa. 1977. Isolationof the alanine carrier from the membranes of a thermophilicbacterium and its reconstitution into vesicles capable of trans-port. J. Supramol. Struct. 6:77-84.
15. Kaback, H. R. 1974. Transport in isolated bacterial membranevesicles. Methods Enzymol. 31:698-709.
16. Kagawa, Y. 1978. Reconstitution of the energy transformer, gateand channel, subunit reassembly, crystalline ATPase and ATPsynthesis. Biochim. Biophys. Acta 505:45-93.
17. Konings, W. N., A. Bisschop, and M. C. C. Daatselaar. 1972.Transport of L-glutamate and L-aspartate by membrane vesiclesof Bacillus subtilis W23. FEBS Lett. 24:260-264.
18. Konings, W. N., A. Bisschop, M. Veenhuis, and C. A. Ver-meulen. 1973. New procedure for the isolation of membranevesicles of Bacillus subtilis and an electron microscopy study oftheir ultrastructure. J. Bacteriol. 116:1456-1465.
19. Konings, W. N., and E. Freese. 1972. Aminoacid transport inmembrane vesicles of B. subtilis. J. Biol. Chem. 247:2408-2418.
20. Lolkema, J. S., K. J. Hellingwerf, and W. N. Konings. 1982. Theeffect of "probe binding" on the quantitative determination ofthe proton motive force in bacteria. Biochim. Biophys. Acta681:85-94.
21. Lowry, 0. H., N. J. Rosebrough, A. L. Farr, and R. J. Randall.1951. Protein measurement with the Folin phenol reagent. J.Biol. Chem. 193:265-275.
22. Reizer, J., and N. Grosswicz. 1974. Properties of a isoaminobu-tyric acid transport in a thermophilic microorganism. J. Bacte-riol. 118:414-424.
23. Russell, J. B., H. J. Strobel, A. J. M. Driessen, and W. N.Konings. 1988. Sodium-dependent transport of neutral aminoacids by whole cells and membrane vesicles of Streptococcusboli's, a ruminal bacterium. J. Bacteriol. 170:3531-3536.
24. Schellenberg, G. D., and C. E. Furlong. 1977. Resolution of themultiplicity of the glutamate and aspartate transport systems ofEscherichia coli. J. Biol. Chem. 252:9055-9064.
25. Shinbo, T., N. Kamo, K. Kurihara, and Y. Kobataka. 1978. APVC-based electrode sensitive to DDA+ as a device for moni-toring the membrane potential in biological systems. Arch.Biochem. Biophys. 187:414-422.
26. Sone, N. 1986. Cytochrome oxidase from thermophilic bacte-rium PS-3. Methods Enzymol. 126:145-153.
27. Sone, N., and Y. Yanagita. 1982. A cytochrome aa3-type termi-nal oxidase of a thermophilic bacterium purification, propertiesand proton pumping. Biochim. Biophys. Acta 682:216-226.
28. Wakayama, N. 1978. Membrane properties of an extreme ther-mophile. J. Biochem. 83:1693-1698.
VOL. 171, 1989
on Novem
ber 8, 2018 by guesthttp://jb.asm
.org/D
ownloaded from





























