Embed Size (px)
Citation preview

Mechanism of Cardiovascular Action ofTetrodotoxin in the Cat
BLOCK OF CONDUCTION IN PERIPHERAL SYMPATHETIC FIBERS
By Maurice B. Feinstein, Ph.D., and Marve Paimre, B.S.
ABSTRACTTetrodotoxin given to cats in doses of 1 to 10 fi,g/kg iv caused profound
cardiovascular depression characterized by decreases in systolic and diastolicblood pressures, pulse pressure, heart rate, force of myocardial contraction,cardiac output and peripheral resistance. These effects were associated withmarked inhibition of the responses to stimulation of nerves to the adrenalmedulla and the sympathetic nerves to heart and blood vessels. Sinus brady-cardia due to toxin did not occur after acute cardiac sympathetic denervation.In isolated sympathetic nerve-right atrium preparations, tetrodotoxin at 1 to2 x 10~8 g/ml abolished the positive inotropic and chronotropic responses tonerve stimulation without effect on spontaneous rate or force, or transmitterrelease by tyramine. Toxin-produced vasodilation in perfused hindlimbs wasassociated with marked inhibition of the normal vasoconstrictor responsesto sympathetic vasomotor nerve stimulation. Vasodilation was not due toa direct action on vascular smooth muscle, since it was not seen after a-receptorblockade with phenoxybenzamine, after sympathetic denervation, or afterblockade of adrenergic transmitter release by guanethidine or 8̂-TM 10 ac-companied by adrenalectomy. Vasodilators such as acetylcholine, histamineand isoproterenol consistently produced additional vasodilation under theseconditions. The hypotensive effect of the toxin was also observed in spinalcats. It is concluded that the potent hypotensive action of tetrodotoxin, inthe cat, is primarily due to blockade of conduction in peripheral sympatheticnerves and nerve fibers innervating the adrenal medulla.
ADDITIONAL KEY WORDSmyocardial contractionspinal cat
adrenal medullavasomotor center
cardiac outputlimb blood flow
• Tetrodotoxin is the purified active prin-ciple responsible for tetrodon poisoning, alsoknown as puffer-fish or fugu poisoning. It hasbecome a very important tool in the study ofexcitation phenomena in cells because of itspotent and highly selective inhibition of theinitial changes in membrane permeability re-sponsible for the spike potential in nerve (1).The toxin's poisonous properties in man and
From the Department of Pharmacology, State Uni-versity of New York, Downstate Medical Center,Brooklyn, New York 11203.
This investigation was supported by U. S. PublicHealth Service Grant GM14299.
This paper was presented in part to the AmericanSociety for Pharmacology and Experimental Thera-peutics, August 1967.
Accepted for publication August 20, 1968.
other animals stem from its depressant ac-tions on the cardiovascular and respiratorysystems, thus leading to severe hypotensionand respiratory paralysis. The mechanism ofthe hypotensive action of tetrodotoxin re-mains a subject of controversy, although re-ports on its circulatory effects have appearedsince 1890. The hypotension has been as-cribed variously to depression of central vaso-motor areas (2-5), a histamine-like vasodila-tor action (4), a direct vasodilator action onvascular smooth muscle (6), ganglionic orpreganglionic blockade (7), and blockade ofvasomotor nerves (8). Evidence for andagainst brain vasomotor centers as the prin-cipal site of toxin action has recently beenpublished (5,9).
Circulation Reiearcb, Vol. XXIII, October 1968 553
by guest on May 10, 2018
http://circres.ahajournals.org/D
ownloaded from

554 FEINSTEIN, PAIMRE
It has been rather generally concluded thatthe cardiovascular effects of tetrodotoxin re-sult from a loss of vasomotor tone withoutany significant direct action on the heart (1,5). However, the evidence concerning theeffects of the toxin on the mammalian heartis meager and often contradictory. AlthoughKao and Fuhrman (7) reported little if anyalteration of the heart rate in cats, except atvery high doses of about 20 /xg/kg, Murthaand associates (3) observed some decrease inheart rate in dogs receiving doses of only 3to 6 /ng/kg and a substantial decrease in myo-cardial contractility. The method by whichthe latter effect was studied was not de-scribed, and little attention has been paidsince then to the actions of the toxin on themammalian heart. Tsukada (10) found thattetrodotoxin produced a slight sinus brady-cardia in rodents which was not preventedby vagotomy or atropine. Conduction dis-turbances between atrium and ventricles havealso been described (2, 8, 10).
In this paper we report the effects of tetro-dotoxin on the responses of the cat to stimu-lation of sympathetic fibers that innervatethe heart, blood vessels, and adrenal medulla.We found that peripheral vasodilation anddepression of myocardial function result fromthe potent blockade of conduction in theseperipheral sympathetic nerves by tetrodotox-in, thus confirming the early conclusion ofIshihara (8), which was based solely onvisual observations of blood vessels.
MethodsCats of both sexes, ranging in weight from
1.8 to 2.5 kg, were anesthetized either by intra-peritoneal injection of sodium pentobarbital, 35mg/kg, or by ether followed by intravenous in-jection of a-chloralose, 40 mg/kg, after whichthe ether was discontinued. All animals wereartificially ventilated because of the toxin's abil-ity to produce respiratory paralysis. In five cats,no pentobarbital was given; the spinal cord wassectioned at C1 under ether anesthesia. The etherwas then discontinued, and the animals wereartificially ventilated at a minute volume ofabout 0.3 liters with a respirator (E & M Instru-ment Co. model V5KG) equipped with a non-rebreathing valve. The body temperature wasmaintained by warming the animals with lamps.
VASCULAR PRESSURES, VENTRICULAR FORCEAND CARDIAC OUTPUT
Blood pressures were recorded on a Grassmodel 5D polygraph from either the carotid orfemoral arteries with Statham P23D pressuretransducers. Pulse rates and electrocardiogramswere monitored with a Grass 5P4 preamplifier.Intraventricular pressures were measured by di-rect needle puncture of the left ventricle aftersternal thoracotomy. A Walton-Brodie straingauge arch, modified in size for the catheart (11), was used to assess the force of ven-tricular contraction from a segment of the myo-cardium 5 to 6 mm long. The strip of ven-tricle between the feet of the arch was stretchedby approximately 50% as recommended by Cottenand Bay (12) to minimize errors due to changesin the size of the ventricles.
Cardiac output was determined by the dye di-lution method, with indocyanine green1 as the in-dicator. The dye was injected into the jugularvein, and arterial blood was withdrawn from thecarotid artery at a constant rate by an infusion-withdrawal pump (Harvard Apparatus Co.)through a flow cell2 placed in a Beckman DUspectrophotometer. The dye concentration curvesin the blood were recorded with a Gilford model2000 recorder at 800 mfi, after which the with-drawn blood (about 10 to 15 ml) was rein-fused. The response time of the recorder was 1second for full scale (10-inch chart) deflec-tion. Cardiac output was calculated from thecurves after correction for recirculation of thedye by the method of Kinsman and associ-ates (13). Duplicate determinations of the con-trol cardiac outputs in each cat agreed within10%. The control cardiac output in seven experi-ments was 0.346 liters/min (± .030 SE), whichagrees favorably with a mean value of 0.33 liters/min previously reported for cats (14, 15).
HINDLIMB PERFUSION
Blood flow in the abdominal aorta, just belowthe inferior mesenteric artery, was divertedthrough a polyethylene cannula to a 5-ml reser-voir heated to 37°C, and then into a Holter per-fusion pump. The blood was returned by thepump at a controlled constant rate back into theaorta through a second cannula inserted justrostral to the bifurcation of the external iliacarteries. The perfusion pressure was monitoredby a Statham P23A transducer through the side-arm of a T-tube, inserted into the outflow can-nula from the pump. Under constant flow condi-
1Cardiogreen, donated by Hynson, Westcott &Dunning, Baltimore, Md.
2Hellma Cells Inc., Jamaica, N. Y., cell No. 137(0.3 ml).
Circulation Research, Vol. XXII1, October 1968
by guest on May 10, 2018
http://circres.ahajournals.org/D
ownloaded from

CARDIOVASCULAR ACTIONS OF TETRODOTOXIN 555
tions the changes in perfusion pressure reflectchanges in the vascular resistance of the per-fused area (16).
AUTONOMIC NERVE STIMULATION
The effects of tetrodotoxin on the responsesto stimulation of various pre- and postganglionicautonomic nerves were investigated after surgicalisolation of the respective nerve trunks andganglia. The nerves were placed over platinumwire electrodes and kept moist and electricallyinsulated from the surrounding tissues by warmmineral oil. Stimulation at supramaximal voltage,2-msec duration, and frequencies varying from 5to 35 impulses/sec was provided by a Grass stimu-lator. Among the nerves stimulated and the cor-responding physiologic responses elicited therebywere: (1) pre- and postganglionic fibers of thesuperior cervical ganglion (contraction of thenictitating membrane); (2) pre- and postgan-glionic fibers of the right and left stellate ganglia(increased arterial blood pressure, intraventricu-lar pressure, heart rate, and ventricular force);(3) splanchnic nerve fibers or fibers from theceliac ganglia that innervate the adrenal medulla(increased blood pressure and heart rate); (4)pre- and postganglionic fibers of the lumbar sym-pathetic chain and postganglionic fibers runningin the sciatic nerve trunk (increased vascular re-sistance in the perfused hind limbs and tail);(5) right cervical vagal trunk (decreased bloodpressure and heart rate). In two experiments, avasopressor area located in the sigmoid gyms ofthe cerebral cortex was electrically stimulatedafter trephining the skull.
ISOLATED SYMPATHETIC NERVE-ATRIUM PREPARATIONS
Under artificial respiration, the chest was
opened by splitting the sternum. The right tho-racic sympathetic chain and the stellate ganglionwere isolated. These were dissected out alongwith the entire heart and the tissues throughwhich the postganglionic cardiac nerves course tothe heart. The dissection was continued in a cul-ture dish containing Krebs3 solution perfusedwith 95% O3-5& CO2 so as to remove as much ex-traneous tissue as possible, including the left atri-um and both ventricles. The pre- and postgangli-onic fibers were placed over two pairs of platinumelectrodes held by micromanipulators so thatthey could be lifted out of the bath momentarilyduring the period of stimulation. The right atriumwas fixed in a horizontal position and its force ofcontraction measured with a Grass FT 03 trans-ducer. In some experiments, electrical activity ofthe atria was recorded with a suctionelectrode (17). This method measures the de-marcation or injury potential, which is a variablefraction of the actual membrane potential. Themagnitude of the action potentials measured inour experiments ranged from 5 to 30 mv. Theconfiguration of the action potential is almostexactly the same as that obtained with intracel-lular microelectrodes (18), and a good implan-tation of the suction electrode enables one torecord from a contracting atrium of a cat forconsiderable periods without the misfortune ofelectrode breakage or displacement, which is socommon in this tissue when using glass microelec-trodes.
Crystalline tetrodotoxin was obtained from theSankyo Co., Tokyo, Japan.
•HThe composition of the solution (in ITIM) was:NaCl 120, KC1 4.6, CaCl, 2.6, KH,,PO4 1.2, MgSO41.2, NaHCO3 25, glucose" 11.
TABLE 1Cardiovascular Effects of Tetrodotoxin
Tetrodotoxin (flg/kg iv)Controls 2.5 5.0 10
No. of experiments 36Heart rate (beats/min) 183 ± 5Blood pressure (mm Hg)
Systolic 132 ± 4Diastolic 94 ± 3
ECG intervals (msec)Q-T 249 ± 7Q-S 68 ± 2P-R 72 ± 2P-P 341 ± 1 1
9186 ± 7*
99 ± 663 ± 5
12128 ± 6
63 ± 432 ± 2
322 ± 1576 ±4*85 ± 2
479 ± 29
12130 ± 10
43 ± 522 ± 4
322 ± 1766 ± 5 *80 ± 4
509 ± 40
3132 ± 5
47 ±102.3 ± 7
311 ±1575 ±0*83 ± 0
451 ± 22
Values are mean ± SEM.*P>.05; for all other values P<.01. Calculations by Student's t-test.
Circulation Research, Vol. XXIII, October 1968
by guest on May 10, 2018
http://circres.ahajournals.org/D
ownloaded from

556 FEINSTEIN, PAIMRE
HR /9S
toxin
HR /6S
FIGURE 1
(a-d) Femoral arterial pressure in a 2.3-kg cat. Inhibition of pressor response to electricalstimulation (5 v, 30/sec), at S, of left thoracic sympathetic chain and induction of mechanicalpulsus alternans by tetrodotoxin, 5 ng, injected through the jugular vein. Note abortivepulses at arrows, (d) Partial recovery after 15 minutes, (e, f) 2.6-kg cat. Note fall in heartrate (e) and abortive pulses indicated at arrows, despite normal ECG, after injection of toxin.
ResultsEFFECTS OF TETROOOTOXIN ON BLOOD PRESSURE,HEART RATE, ECG, AND PULSE WAVES
The principal cardiovascular actions oftetrodotoxin are summarized by the data com-piled in Table 1.
In nine experiments, a dose of 1 /^g/kg re-duced blood pressure to an average level of99 ±6/63 ± 5 mm Hg, although in two ofthese this dose was completely without effect.The heart rate fell in three of the seven ex-periments in which the toxin had a hypoten-sive effect, and the greatest slowing amount-ed to 20%. At doses of 2.5 to 10 ^ug/kg, thetoxin always produced a substantial fall insystolic and diastolic blood pressures, pulsepressure, and heart rate. The maximum re-duction of blood pressure occurred at a doseof 5 /u,g/kg; bradycardia was maximal at adose of 2.5 //.g/kg. When the heart rate wasrecorded continuously with a Grass tacho-graph, we observed that the bradycardia wasconcurrent with the fall in arterial pressure(Fig. l,e, and 5, A).
In two cats, acute sympathetic denerva-tion of the heart by removal of the thoracic
sympathetic chains and the stellate gangliaon both sides resulted in a fall in heart ratefrom 144 to 102 in one animal, and from 240to 100 in the other. The administration of asmuch toxin as 15 //-g/kg did not further de-crease the heart rate but, in fact, increased itto 120 and 111, respectively. We believe thatthis effect was due to inhibition of conductionin vagal fibers, since in five other animalswe were able to demonstrate that 2.5 to 5/itg/kg of toxin would effectively preventslowing of the heart due to electrical stimula-tion of the vagus nerve.
The fall in heart rate was a sinus brady-cardia, as mainly the P-P interval was in-creased. There was little or no effect on eitheratrioventricular conduction time (P-R) or in-traventricular conduction (Q-S) (Table 1).However, the lack of effect of toxin on sym-pathetically denervated hearts, as well as ex-periments on spontaneously beating isolatedright atria, to be described below, indicatethat bradycardia is due to blockade of con-duction in sympathetic fibers carrying im-pulses to the pacemaker region and not to adirect effect on cells of the sinoatrial node.
Circulation Research, Vol. XXIII, October 1968
by guest on May 10, 2018
http://circres.ahajournals.org/D
ownloaded from
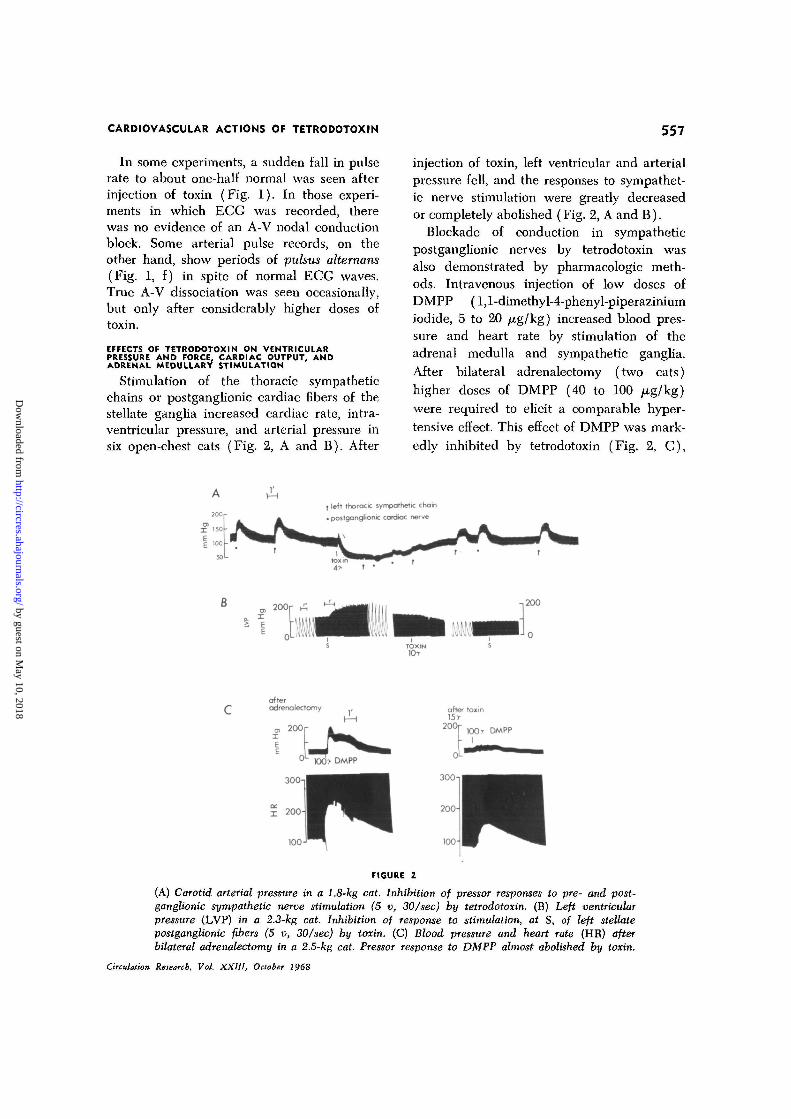
CARDIOVASCULAR ACTIONS OF TETRODOTOXIN 557
In some experiments, a sudden fall in pulserate to about one-half normal was seen afterinjection of toxin (Fig. 1). In those experi-ments in which ECG was recorded, therewas no evidence of an A-V nodal conductionblock. Some arterial pulse records, on theother hand, show periods of pulsus alternans(Fig. 1, f) in spite of normal ECG waves.True A-V dissociation was seen occasionally,but only after considerably higher doses oftoxin.
EFFECTS OF TETRODOTOXIN ON VENTRICULARPRESSURE AND FORCE, CARDIAC OUTPUT, ANDADRENAL MEDULLARY STIMULATION
Stimulation of the thoracic sympatheticchains or postganglionic cardiac fibers of thestellate ganglia increased cardiac rate, intra-ventricular pressure, and arterial pressure insix open-chest cats (Fig. 2, A and B). After
injection of toxin, left ventricular and arterialpressure fell, and the responses to sympathet-ic nerve stimulation were greatly decreasedor completely abolished (Fig. 2, A and B).
Blockade of conduction in sympatheticpostganglionic nerves by tetrodotoxin wasalso demonstrated by pharmacologic meth-ods. Intravenous injection of low doses ofDMPP (l,l-dimethyl-4-phenyl-piperaziniumiodide, 5 to 20 /xg/kg) increased blood pres-sure and heart rate by stimulation of theadrenal medulla and sympathetic ganglia.After bilateral adrenalectomy (two cats)higher doses of DMPP (40 to 100 ju.g/kg)were required to elicit a comparable hyper-tensive effect. This effect of DMPP was mark-edly inhibited by tetrodotoxin (Fig. 2, C),
I 150
1 ,00
50
I left thoracic sympathetic chain• postganglionic cardiac nerve
FIGURE 2
(A) Carotid arterial pressure in a 1.8-kg cat. Inhibition of pressor responses to pre- and post-ganglionic sympathetic nerve stimulation (5 v, 30/sec) by tetrodotoxin. (B) Left ventricularpressure (LVP) in a 2.3-kg cat. Inhibition of response to stimulation, at S, of left stellatepostganglionic fibers (5 v, 30/sec) by toxin. (C) Blood pressure and heart rate (HR) afterbilateral adrenalectomy in a 2.5-kg cat. Pressor response to DMPP almost abolished by toxin.
Circulation Research, Vol. XXIII, October 1968
by guest on May 10, 2018
http://circres.ahajournals.org/D
ownloaded from

558 FEINSTEIN, PAIMRE
FIGURE 3
Effect of tetrodotoxin on arterial blood pressure (ART) and left ventricular force (LVF). (A)2-kg cat. Effect of stimulation of left thoracic sympathetic chain (5 v, 10/secj, at S. Afterintravenous injection of toxin, ventricular force was reduced and response to nerve stimulationwas almost abolished (B). Response to norepinephrine (NE) unaffected (C).
probably by blockade of conduction of im-pulses in the sympathetic postganglionic fi-bers. The persistence of some increase inblood pressure and heart rate following highdoses of DMPP after tetrodotoxin, in adre-nalectomized animals, is most likely the resultof direct release of endogenous tissue cate-cholamines from nerve terminals by DMPP,especially within the heart (19-21).
After intravenous injection of 2.5 /Ag/kgtetrodotoxin, the force of ventricular contrac-tion decreased by 54 ± 4.5% in eight experi-ments (6 left ventricles, 2 right ventricles);the velocity of contraction was also decreased(Fig. 3). Prior to administration of the toxin,stimulation of the pre- or postganglionic fi-bers of the stellate ganglia in these animalsincreased ventricular force by 50% to 76%;however, after the administration of the toxinthis positive inotropic effect was abolished(Fig. 3, A and B). On the other hand, theresponse to norepinephrine was not im-
paired (Fig. 3, C). If the arterial pressurewas maintained in these same animals at alow level, similar to that after toxin, by theinfusion of histamine or acetylcholine, ven-tricular force actually increased during theinfusion of histamine and fell by an averageof only 10% with acetylcholine.
The effect of tetrodotoxin on the cardiacoutput is shown in Table 2. In seven experi-ments, 2.5 //.g/kg toxin administered intra-venously reduced the cardiac output by anaverage of 50%, and the stroke volume by 35%.Thus the decrease in cardiac output is duenot only to a decreased stroke volume, butalso to the attendant bradycardia. In fouropen-chest animals, a Henderson cardiom-eter (22) placed around the ventricles wasused to measure beat-to-beat variations inventricular volume. These experiments con-firmed that stroke volume was decreasedafter administration of tetrodotoxin.
TABLE 2
Effect of Tetrodotoxin on Cardiac Output
No.experiments
Arterial bloodpressure (mm Hg)
Heart rate(beats/min)
Cardiac output(liters/min)
Calculatedstroke volume
(ml)
77
5
ControlsAfter toxin
(2.5*cg/kg)After recovery
from toxin
163 ± 11/113 :74 ± 6/42 ±
166 ± 14/114 :
± 1 04
± 14
176 ± 15145 ± 14
162 ± 19
.346 ± .030
.176 ± .028
.304 ± .043
2.1 ±0.21.3 ± 0.3
2.0 ± 0.3
All values are mean ± SEM.
Circulation Research, Vol. XX111, October 1968
by guest on May 10, 2018
http://circres.ahajournals.org/D
ownloaded from
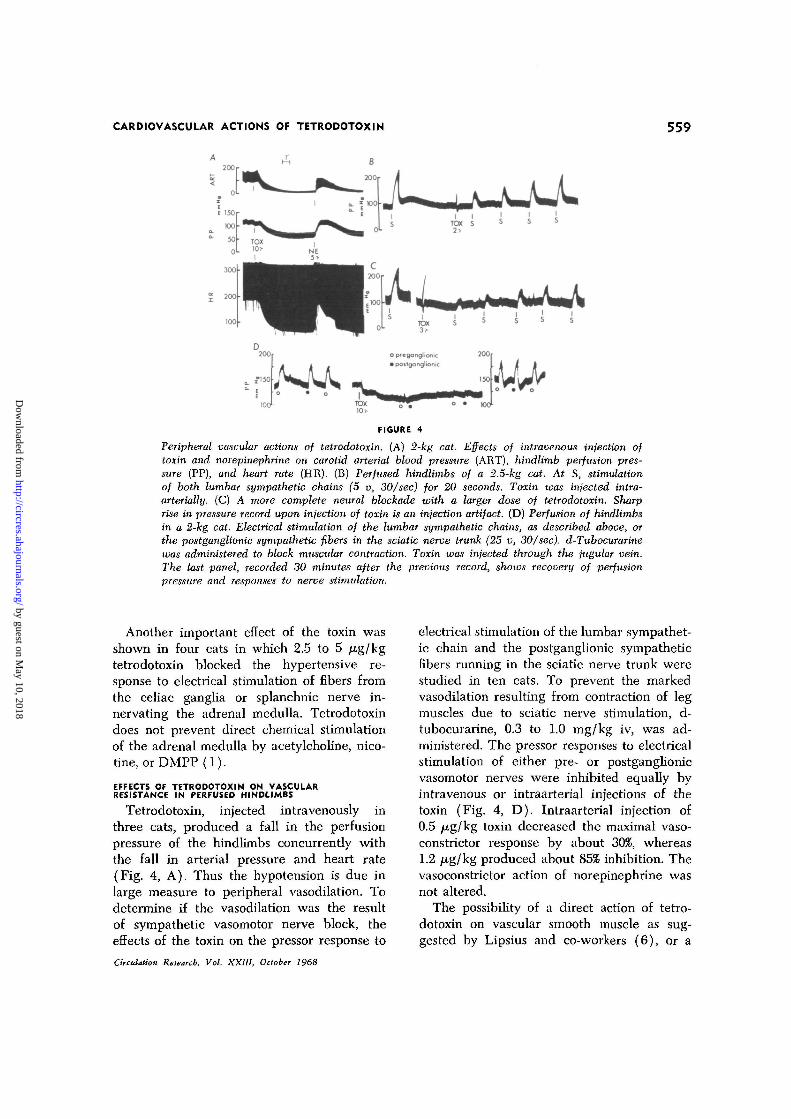
CARDIOVASCULAR ACTIONS OF TETRODOTOXIN 559
FIGURE 4
Peripheral vascular actions of tetrodotoxin. (A) 2-fcg cat. Effects of intravenous injection oftoxin and norepinephrine on carotid arterial blood pressure (ART), hindlimb perfusion pres-sure (PP), and heart rate (HR). (B) Perfused hindlimbs of a 2.5-kg cat. At S, stimulationof both lumbar sympathetic chains (5 v, 30/sec) for 20 seconds. Toxin was injected intra-arterially. (C) A more complete neural blockade with a larger dose of tetrodotoxin. Sharprise in pressure record upon injection of toxin is an injection artifact. (D) Perfusion of hindlimbsin a 2-kg cat. Electrical stimulation of the lumbar sympathetic chains, as described above, orthe postganglionic sympathetic fibers in the sciatic nerve trunk (25 v, 30/sec). d-Tubocurarinewas administered to block muscular contraction. Toxin was injected through the jugular vein.The last panel, recorded 30 minutes after the previous record, shows recovery of perfusionpressure and responses to nerve stimulation.
Another important effect of the toxin wasshown in four cats in which 2.5 to 5 j^g/kgtetrodotoxin blocked the hypertensive re-sponse to electrical stimulation of fibers fromthe celiac ganglia or splanchnic nerve in-nervating the adrenal medulla. Tetrodotoxindoes not prevent direct chemical stimulationof the adrenal medulla by acetylcholine, nico-tine, orDMPP (1).
EFFECTS OF TETRODOTOXIN ON VASCULARRESISTANCE IN PERFUSED HINDLIMBS
Tetrodotoxin, injected intravenously inthree cats, produced a fall in the perfusionpressure of the hindlimbs concurrently withthe fall in arterial pressure and heart rate(Fig. 4, A). Thus the hypotension is due inlarge measure to peripheral vasodilation. Todetermine if the vasodilation was the resultof sympathetic vasomotor nerve block, theeffects of the toxin on the pressor response to
Circulation Research, Vol. XXIII, October 1968
electrical stimulation of the lumbar sympathet-ic chain and the postganglionic sympatheticfibers running in the sciatic nerve trunk werestudied in ten cats. To prevent the markedvasodilation resulting from contraction of legmuscles due to sciatic nerve stimulation, d-tubocurarine, 0.3 to 1.0 mg/kg iv, was ad-ministered. The pressor responses to electricalstimulation of either pre- or postganglionicvasomotor nerves were inhibited equally byintravenous or intraarterial injections of thetoxin (Fig. 4, D). Intraarterial injection of0.5 /Ag/kg toxin decreased the maximal vaso-constrictor response by about 30%, whereas1.2 /Ag/kg produced about 85% inhibition. Thevasoconstrictor action of norepinephrine wasnot altered.
The possibility of a direct action of tetro-dotoxin on vascular smooth muscle as sug-gested by Lipsius and co-workers (6), or a
by guest on May 10, 2018
http://circres.ahajournals.org/D
ownloaded from

560 FEINSTEIN, PAIMRE
50
after sympatheticdenervation andodrenalectomy
FIGURE 5
Peripheral vascular actions of tetrodotoxin. (A) Responses of perfused hindlimbs in 2.3-kgcat to intraartericd injection, of norepinephrine (NE), epinephrine (E), acetylcholine (Ach),histamine (H), and tetrodotoxin (TOX). After maximal vasodilation with toxin, histamineproduced a further fall in perfusion pressure (PP). (B) After administration of 10 mg ofphenoxybenzamine to the same cat. No additional vasodilation produced by twice theoriginal dose of toxin. Histamine and acetylcholine remained effective, and the original pressoreffect of epinephrine was reversed. Initial sharp rise and slight fall in pressure records afterinjections are injection artifacts. (C) 2-hg cat. Effect of toxin and guanethidine on responseto lumbar sympathetic chain stimulation at S (5 v, 10/sec). (D) Fall in perfusion pressure ofhindlimbs after tying ligatures placed around both adrenals. Initial brief fall in perfusionpressure after toxin is an injection artifact, but note marked vasodilation due to isoproterenol(I). (E) 2-kg cat. Response to stimulation of lumbar sympathetic chain at S. After recoveryfrom toxin effects, bilateral adrenalectomy and sympathetic denervation of the hindlimbsresulted in a fall in perfusion pressure. Large dose of toxin had no greater effect thaninjection of same volume of saline (Sal), but other vasodilators remained effective.
histamine-like action, as suggested by Li (4)was investigated. We assumed that any directvasodilator action on vascular smooth muscleshould be apparent even after completeblockade of transmitter release from vasocon-strictor nerves or a-receptor blockade. Theeffect of tetrodotoxin on the circulation in theperfused hindlimbs was therefore studied af-
ter treatment with guanethidine or /3-TM 104
to abolish the release of norepinephrine whenadrenergic nerves were stimulated, or sufficientphenoxybenzamine to abolish the vasocon-strictor response to norepinephrine. In 23
4(2(2,6-dimethylphenoxy) propyl) trimethylammo-nium bromide, the /S-methyl analogue of xylocholine,Smith, Kline & French.
Circulation Research, Vol. XXIII, October 1968
by guest on May 10, 2018
http://circres.ahajournals.org/D
ownloaded from
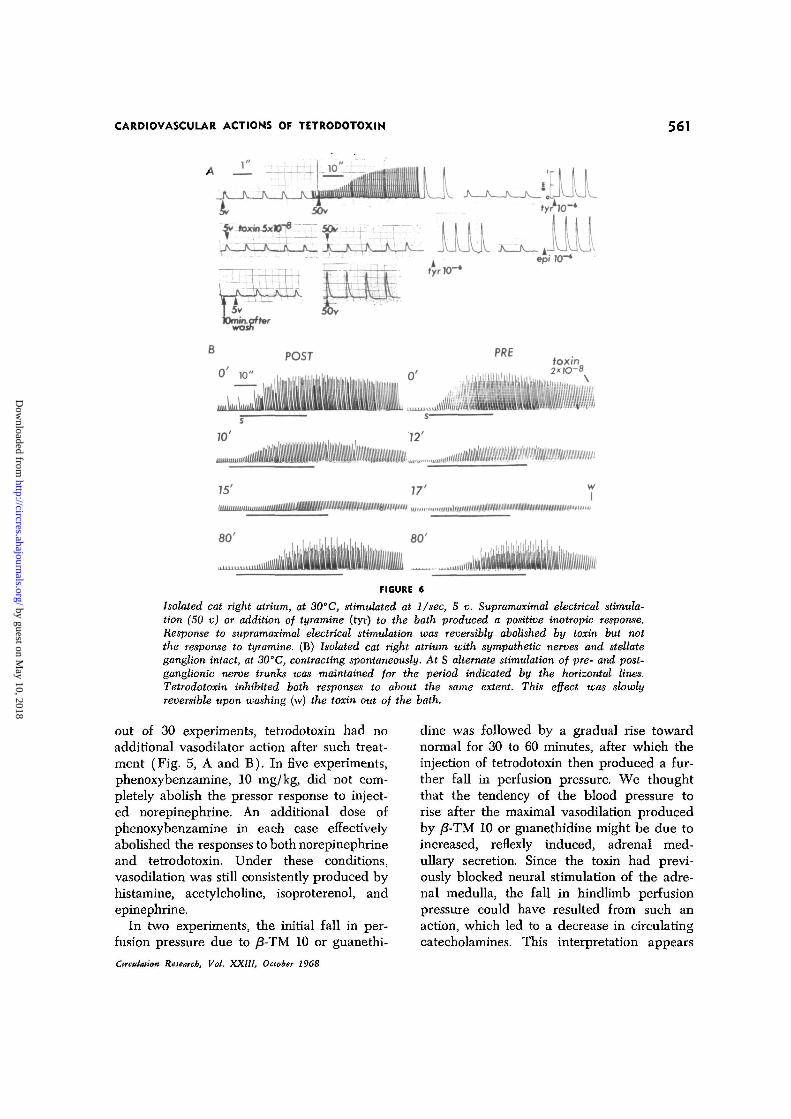
CARDIOVASCULAR ACTIONS OF TETRODOTOXIN 561
A - -K. .
POST PREfoxin2»)O-8
15' 17' wI
FIGURE 6
Isolated cat right atrium, at 30°C, stimulated at 1/sec, 5 v. Supramaximal electrical stimula-tion (50 v) or addition of tyramine (tyr) to the bath produced a positive inotropic response.Response to supramaximal electrical stimulation was reversibly abolished by toxin but notthe response to tyramine. (B) Isolated cat right atrium with sympathetic nerves and stellateganglion intact, at 30°C, contracting spontaneously. At S alternate stimulation of pre- and post-ganglionic nerve trunks was maintained for the period indicated by the horizontal lines.Tetrodotoxin inhibited both responses to about the same extent. This effect was slowlyreversible upon washing (w) the toxin out of the bath.
out of 30 experiments, tetrodotoxin had noadditional vasodilator action after such treat-ment (Fig. 5, A and B). In five experiments,phenoxybenzamine, 10 ing/kg, did not com-pletely abolish the pressor response to inject-ed norepinephrine. An additional dose ofphenoxybenzamine in each case effectivelyabolished the responses to both norepinephrineand tetrodotoxin. Under these conditions,vasodilation was still consistently produced byhistamine, acetylcholine, isoproterenol, andepinephrine.
In two experiments, the initial fall in per-fusion pressure due to /3-TM 10 or guanethi-
Circulation Research, Vol. XXlll, October 1968
dine was followed by a gradual rise towardnormal for 30 to 60 minutes, after which theinjection of tetrodotoxin then produced a fur-ther fall in perfusion pressure. We thoughtthat the tendency of the blood pressure torise after the maximal vasodilation producedby /3-TM 10 or guanethidine might be due toincreased, reflexly induced, adrenal med-ullary secretion. Since the toxin had previ-ously blocked neural stimulation of the adre-nal medulla, the fall in hindlimb perfusionpressure could have resulted from such anaction, which led to a decrease in circulatingcatecholamines. This interpretation appears
by guest on May 10, 2018
http://circres.ahajournals.org/D
ownloaded from

562 FEINSTEIN, PAIMRE
to be correct, since acute bilateral adrenalec-tomy performed in four cats 30 minutes afteradministration of /3-TM 10 or guanethidineresulted in a prompt fall in hindlimb perfusionpressure, ranging from 18 to 35 mm Hg (Fig.5, D). Thereafter, injections of tetrodotoxin indoses as high as 10 /Ag/kg into the hindlimbcirculation were without effect. Histamine,isoproterenol, and acetylcholine, however,always produced additional vasodilationwith falls in perfusion pressure of 30 to45 mm Hg. In two cats, acute sympatheticdenervation of the hindlimbs (lumbar sym-pathetic chains removed and sciatic nervetrunks sectioned) accompanied by bilateraladrenalectomy also resulted in a fall in per-fusion pressure, after which tetrodotoxin waswithout further effect (Fig. 5, E). Underthese conditions, the direct vasodilators re-ferred to above still produced additional vaso-dilation.
In three intact cats, after several doses of2.5 to 5 /ug/kg of toxin, or a single dose of 10/Mg/kg, a state was obtained in which therewas prolonged, severe hypotension and aboli-tion of sympathetic vasoconstrictor nerve re-sponses. Under these conditions, repeated ad-ditional doses of toxin no longer affectedthe arterial blood pressure. Yet histamine,isoproterenol, and acetylcholine consistentlyand repeatedly lowered the blood pressureeven further. A similar observation was madeby Koizumi and her co-workers (5) usingacetylcholine. They also were unable to pre-vent the hypotensive action of the toxin withan antihistaminic compound.
EFFECTS OF TETRODOTOXIN ON ISOLATED RIGHTATRIUM AND SYMPATHETIC NERVE-ATRIUMPREPARATIONS
Stimulation of the intrinsic sympatheticnerve endings in the isolated right atrium bysupramaximal voltage markedly increases theforce of contraction of electrically driven mus-cles (23). This effect was completely abol-ished in five experiments by adding as littleas 1 to 2 X 10~s g/ml toxin to the tissue bathwithout affecting the response to the directaction of catecholamines or the catechola-mine-releasing action of tyramine (Fig. 6, A).Thus the toxin probably acts by inhibiting
the conduction of impulses in nerve fiberswithout affecting the mechanism for releaseof the neurotransmitter. This interpretation isconsistent with the recent experiments ofKatz and Miledi (24), who showed that tet-rodotoxin, while blocking action potentialconduction in motor nerves, did not hinderthe release of acetylcholine due to local de-polarization of the nerve terminals.
In five atria isolated with their sympatheticnerves intact, the stimulation of either pre- orpostganglionic fibers increased the force andspontaneoas frequency of contractions (Fig.6, B). As little as 5 X 10"9 g/ml toxin partiallyinhibited these responses to nerve stimulation,and complete block was attained at 1 to 2 X10~8 g/ml. Responses to pre- and postgangli-onic stimulation were reduced equally.
Assuming that after a single intravenous in-jection into an intact cat, tetrodotoxin is dis-tributed mainly in the extracellular fluid dur-ing the first few minutes required to attainmaximal pharmacologic effect, one can cal-culate an approximation of the possible toxincontent of the extracellular water. These cal-culations give figures of 5 X 10"9 to 2.5 X 10"8
g/ml after doses of 1 to 5 /ug/kg, which isin the range found to block sympathetic neu-rons in vitro.
FIGURE 7
Effect of high doses of tetrodotoxin on spontaneouscontractions (upper trace) and action potentials (suc-tion electrode) (lower trace) in isolated cat rightatrium. Temperature 35°C. (A) Control. (B) Afteraddition of 2 X 1O~7 g/ml norepinephrine. (C) Afterwashout of norepinephine and 5 minutes after addi-tion of 2 X 1O-" g/ml tetrodotoxin. (D) 10 minutesafter addition of toxin. (E) Addition of 2 X 10~7 g/mlnorepinephrine in the presence of toxin. (F) 10 minutesafter washout of both norepinephrine and toxin.
Circulation Research, Vol. XXIII, October 1968
by guest on May 10, 2018
http://circres.ahajournals.org/D
ownloaded from

CARDIOVASCULAR ACTIONS OF TETRODOTOXIN 563
A 100'
X 50
E 0
fi 200r
E 100-
FIGURE S(A) Hypotensive effect of tetrodotoxin, injected intra-venously in a spinal cat. Femoral arterial pressure ina 1.8-kg cat. (B) Femoral arterial pressure in a 2-fcgcat. At S the sigmoid gyrus of the cerebral cortex onthe left side was stimulated electricalltj with a bipolarelectrode (10 v, 20/sec for 15 seconds). The pressorresponse is markedly inhibited after the intravenousinjection of tetrodotoxin. After 20 minutes the bloodpressure returned to normal and stimulation of thecortex evoked a pressor response equal to that of thecontrol.
At the time that the responses to sympa-thetic nerve stimulation were abolished, neitherthe spontaneous rate nor force of contrac-tion of the atria was altered. In one experi-ment, a slight reduction in force was notedat a toxin concentration of 5 X 10~7 g/ml.Higher concentrations caused a slowing ofrate, arrhythmias, and further reduction offorce, until at 1 to 2 X l(h6 g/ml, contrac-tions were abolished in the five atria tested.The atrial action potential was also abolishedat these concentrations, but spontaneous elec-trical and mechanical activity could be re-stored by applying norepinephrine (Fig. 7).
EFFECTS OF TETRODOTOXIN IN SPINAL CATS AND ONA CEREBRAL VASOMOTOR CENTER
Two observations tend to support the con-tention of Kao and his co-workers (9) thatthe hypotensive action of the toxin does notresult from a selective action on the brain.First, we found that 2.5 /^g/kg toxin loweredblood pressure in five spinal cats (Fig. 8, A)from 94/48 mm Hg (range: 105/55 to 75/40)to 65/26 mm Hg (range: 75/35 to 50/20).This is almost precisely the same level toCirculation Research, Vol. XXIII, October 1968
which the arterial pressure was lowered innormal animals at this dose of toxin. Ofcourse, the total magnitude of the pressuredrop is less in spinal animals because of thepreexistent low pressure resulting from theremoval of vasomotor influence of the brain.Second, in two animals, electrical stimulationof the vasomotor area of the sigmoid gyrusof the cerebral cortex increased arterial pres-sure (Fig. 8, B). This effect has been shownto be accompanied by increased frequencyof action potential discharge in sympatheticnerves to the heart (25). The dose of toxinrequired to abolish this response to cerebralstimulation was the same as that whichblocked the effects of peripheral sympatheticneuron stimulation, namely 2.5 to 5 jag/kg.Responses to brain stimulation were not affect-ed by doses which were too low to reducethe effects of peripheral sympathetic nervestimulation.
Discussion
We conclude from the results describedabove that the hypotensive action of tetro-dotoxin in the cat is the result of a combina-tion of peripheral vasodilation and decreasedcardiac output. The latter is a consequenceof a decrease in both heart rate and strokevolume. These effects of tetrodotoxin on theperipheral circulation and the heart coincidein time and intensity with an inhibition ofresponses to stimulation of postganglionicsympathetic nerve fibers innervating bloodvessels and the heart, as well as splanchnicnerve fibers innervating the adrenal medulla.The recovery of the blood pressure to normalis associated with a return of these nervesto their normal responsiveness.
It should be pointed out that in the doserange producing hypotension, tetrodotoxinhas also been found to inhibit motor nervesto skeletal muscle, some afferent nerves, andcertain areas in the central nervous system (1,5). Thus it is found that muscular weakness,respiratory depression, and hypotension arealways concurrent (1).
The toxin did not block responses to stimu-lation of preganglionic sympathetic nerves
by guest on May 10, 2018
http://circres.ahajournals.org/D
ownloaded from

564 FEINSTEIN, PAIMRE
without a comparable effect on responses topostganglionic nerve stimulation. Nor did itinterfere with adrenergic transmitter releaseby such agents as tyramine, or the direct ac-tion of the catecholamines on their target tis-sues. Thus the action of tetrodotoxin, whichis due to the blockade of conduction in axons,is clearly distinguishable pharmacologicallyfrom adrenergic receptor-blocking agentssuch as phenoxybenzamine or agents whichblock transmitter release from adrenergicnerve terminals, such as /3-TM 10 and guan-ethidine. Peripheral vasodilation in the cat isthe result of abolition of sympathetic vaso-motor tone and not to a direct action on vas-cular smooth muscle, because it did not occurafter effective a-receptor blockade, or afteradrenalectomy accompanied by blockade ofadrenergic transmitter release or denervation.The inability to demonstrate a direct vaso-dilating action of tetrodotoxin on vascularsmooth muscle in the cat was not entirely un-expected. The toxin has been shown to haveno effect on the tone of isolated arterial strips,although it abolished responses to vasocon-strictor nerve stimulation (26) and it alsofailed to block action potentials in arterial(27) and venous (28) smooth muscle. Anumber of other isolated smooth muscle prep-arations have also been shown to be unaf-fected by tetrodotoxin (29, 30). Whether thereports of direct vasodilating action in dog(6) and rat (4) can be attributed to differ-ences in the vascular muscle of these speciesis not known at present.
Our observations indicate that the hypo-tensive action of the toxin in the cat cannotbe ascribed to a selective action on the brainvasomotor centers. The cardiovascular depres-sion induced by the toxin was consistentlyelicited in spinal animals, and the inhibitionof responses to stimulation of a brain vaso-motor center was apparent only at doses ofthe drug which produced peripheral sympa-thetic neuron blockade. Recent experimentsin other laboratories concerning the centralactions of tetrodotoxin are contradictory.Cross-perfusion experiments in cats anddogs (9) failed to show any hypotensive ac-
tion of the toxin when it circulated throughthe recipient animal's head only. However,the doses may not have been sufficient, sincethey were calculated on the basis of brainweight rather than the proportion of the totalcardiac output which goes to the brain. Onthe other hand, Koizumi and co-workers (5)clearly demonstrated that doses of toxin inthe range of 1 to 4 fig/kg produced partialto complete block of mono- and polysynapticspinal reflexes, as well as cortically evokedresponses in cats. The significance of thesefindings vis-a-vis the circulatory effects ofthe toxin is difficult to assess for two rea-sons. First, neither the spinal reflexes nor thecortical areas studied were shown to be in-volved in vasomotor regulation; and second,no temporal correspondence between thespinal and cortical effects and the hypoten-sion was found. The latter investigators at-tribute the hypotension primarily to actionof the toxin on the medullary vasomotor cen-ters, because it presumably did not occur inspinal animals. Both we and Murtha and co-workers (3) have found that the toxin acts onspinal animals, although quantitatively theabsolute magnitude of the fall in pressuremust of necessity be less than in intact animalsbecause of the preexistent loss of vasomotorinfluences of the brain. One experimentdescribed by Koizumi and her co-workers(5) does, in fact, show this. Tetrodotoxin,2 jug/kg, did reduce blood pressure in aspinal cat to precisely the same level towhich it had previously reduced it in thesame animal prior to spinal section. Wewould conclude that, although the toxin mayin fact actually depress central nervous systemvasomotor centers, no selective action—thatis, without concurrent blockade of peripheralcardiac and vasomotor nerves—on such areashas yet been convincingly demonstrated. Theperipheral nerve-blocking effects of tetrodo-toxin which have been demonstrated corre-spond in time and intensity to the observeddepression of the circulation and are there-fore entirely sufficient to account for theprincipal pharmacologic actions of the toxinon the cardiovascular system.
Circulation Research, Vol. XXIII, October 1968
by guest on May 10, 2018
http://circres.ahajournals.org/D
ownloaded from

CARDIOVASCULAR ACTIONS OF TETRODOTOXIN 565
References1. KAO, C. Y.: Tetrodotoxin, saxitoxin and their
significance in the study of excitation phenom-ena. Pharmacol. Rev. 18: 997, 1966.
2. TAKAHASHI, D., AND INOKO, Y.: ExperimentelUntersuchungen iiber das Fugugift. Arch. Ex-ptl. Pathol. Pharmakol. 26: 401, 1890.
3. MUBTHA, E. F., STABILE, D. E., AND WILLS,J. H.: Some pharmacological effects of pufferpoison. J. Pharmacol. 122: 247, 1957.
4. Li, K. M.: Action of puffer fish poison. Nature200: 791, 1963.
5. KOIZUMI, K., LEVINE, D. G., AND BROOKS, C.McC: Effect of tetrodotoxin (puffer fish poi-son) on the central nervous system. Neurology17: 395, 1967.
6. LlPSIUS, M., SlECMAN, M. J., AND KAO, C. Y.:Direct action of procaine and tetrodotoxin onvascular muscle (abstr.). Federation Proc.26: 736, 1967.
7. KAO, C. Y., AND FUHRMAN, F. A.: Pharmacologi-cal studies on tarichatoxin, a potent neurotoxin.J. Pharmacol. 140: 31, 1963.
8. ISHIHARA, F.: Uber die physiologischen Wir-kungen des Fugutoxins. Mitt. Med. Fak. TokioUniv. 20: 375, 1918.
9. KAO, C. Y., SUZUKI, T., KLEINHAUS, A. L.,AND SIECMAN, M. J.: Vasomotor and respiratorydepressant actions of tetrodotoxin and saxitox-in. Arch. Intern. Pharmacodyn. 165: 438, 1967.
10. TSUKADA, O.: Sur le mecanisme de la brady-cardie provequee par la tetrodotoxine crystal-isee. Ann. Rep. Inst. Food Microbiol. ChibaUniv. 10: 78, 1957.
11. ROSEN, A.: Augmented cardiac contraction, heartacceleration and skeletal muscle vasodilationproduced by hypothalamic stimulation in cats.Acta Physiol. Scand. 52: 291, 1961.
12. COTTEN, M. DE V., AND BAY, E.: Direct meas-urement of changes in cardiac contractile force.Am. J. Physiol. 187: 122, 1956.
13. KINSMAN, J. M., MOORE, J. W., AND HAMILTON,W. F.: Studies on the circulation: I. Injectionmethod: physical and mathematical considera-tions. Am. J. Physiol. 89: 322, 1929.
14. GUYTON, A. C : Circulatory Physiology: cardiacoutput and its regulation. W. B. Saunders Co.,1963.
15. CROSS, K. W., GROOM, A. C , MOTTHAM, R. F.,AND ROWLANDS, S.: Cardiac output in the cat:Comparison between the Fick method anda radioactive indicator dilution method (ab-str.). J. Physiol. (London) 136: 24P, 1957.
16. BECK, L.: Active reflex dilatation in the in-nervated perfused hind limb of the dog. Am.J. Physiol. 201: 123, 1961.
17. FLOREY, E., AND MAHLON, E. K.: A new suc-
Circulation Research, Vol. XXIII, October 1968
tion electrode system. Comp. Biochem. Phys-iol. 18: 175, 1966.
18. HOFFMAN, B. F., CRANEFIELD, P. F., LEPESCHKIN,E., SURAWICZ, B., AND HERSHICH, H.: Com-parison of cardiac monophasic action potentialsrecorded by intracellular and suction electrodes.Am. J. Physiol. 196: 1297, 1959.
19. CHIANG, T. S., AND LEADERS, F. E.: Mechanismfor nicotine and DMPP on the isolated ratatria-vagus nerve preparation. J. Pharmacol.Exptl. Therap. 149: 225, 1965.
20. LINDMAB, R., AND MUSCHOLL, E.: Die Wirkungvon Pharmaka auf die Elimination von Nor-adrenalin aus die Perfusionsflussigkeit und dieNoradrenalinaufnahme in das isolierte Herz.Arch. Exptl. Pathol. Pharmakol. 247: 469,1964.
21. BHAGAT, B., BHATTACHARYA, I. C, AND DHAL-LA, N. S.: Retention of exogenous norepineph-rine and recovery of the responses to tyramineand l,l-dimethyl-4-phenylpiperazinium iodideafter reserpine. J. Pharmacol. Exptl. Therap.153: 434, 1966.
22. HENDERSON, Y.: Volume curve of the ventriclesof the mammalian heart, and the significanceof this curve in respect to the mechanismof the heart beat and the filling of the ven-tricles. Am. J. Physiol. 16: 325, 1906.
23. FURCHGOTT, R. F., DE GUBAREFF, T., ANDGROSSMAN, A.: Release of autonomic mediatorsin cardiac tissue by suprathreshold stimulation.Science 129: 328, 1959.
24. KATZ, B., AND MILEDI, R.: Tetrodotoxin andneuromuscular transmission. Proc. Roy. Soc.(London), Ser. B: 167: 8, 1967.
25. ALANIS, J., MASCHER, D., AND MIYAMOTO, J.:Relationship between the self-sustained ac-tivity of the cerebral cortex and the efferentdischarge of cardiac sympathetic and para-sympathetic fibers. Jap. J. Physiol. 16: 316,1966.
26. Su, C, AND BEVAN, J. A.: Electrical and me-chanical responses of pulmonary artery mus-cle to neural and chemical stimulation. Biblio-theca. Anat. 8: 30, 1966.
27. KEATINCE, W. R.: Ionic requirements for ar-terial action potential. J. Physiol. (London)194: 169, 1968.
28. HOLMAN, M. E., KASBY, C. B., SUTHERS, M. B.,AND WILSON, J. A. F.: Some properties orthe smooth muscle of rabbit portal vein. J.Physiol. (London) 196: 111, 1968.
29. KURIYAMA, H., OSA, T., AND TOIDA, N.: Effectof tetrodotoxin on smooth muscle cells of theguinea-pig taenia coli. Brit. J. Pharmacol.Chemother. 27: 366, 1966.
30. GERSHON, M. D.: Effects of tetrodotoxin oninnervated smooth muscle preparations. Brit.J. Pharmacol. Chemother. 29: 259, 1967.
by guest on May 10, 2018
http://circres.ahajournals.org/D
ownloaded from

MAURICE B. FEINSTEIN and MARVE PAIMRECONDUCTION IN PERIPHERAL SYMPATHETIC FIBERS
Mechanism of Cardiovascular Action of Tetrodotoxin in the Cat: BLOCK OF
Print ISSN: 0009-7330. Online ISSN: 1524-4571 Copyright © 1968 American Heart Association, Inc. All rights reserved.is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation Research
doi: 10.1161/01.RES.23.4.5531968;23:553-565Circ Res.
http://circres.ahajournals.org/content/23/4/553World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circres.ahajournals.org//subscriptions/
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer about this process is available in the
located, click Request Permissions in the middle column of the Web page under Services. Further informationEditorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center, not theCirculation Research Requests for permissions to reproduce figures, tables, or portions of articles originally published inPermissions:
by guest on May 10, 2018
http://circres.ahajournals.org/D
ownloaded from





























