Embed Size (px)
Citation preview

Journalof Soil Science, 1990,41,341-358
Mechanical impedance to root growth: a review of experimental techniques and root growth responses
A. G. B E N G O U G H & C. E. MULLINS* Cellular and Environmental Physiology Department, Scottish Crop Research Institute, Dundee O D 2 5DA and *Department of Plant and Soil Science, University of Aberdeen.
Aberdeen AB9 ?UE, UK
SUMMARY
Mechanicalimpedancetorootgrowthisoneofthemostimportant factorsdeterminingroot elongation and proliferation within a soil profile. Penetrometers overestimate resistance to root growth in soil by a factor of between two and eight and, although they remain the most convenient method for predicting root resistance, careful interpretation of results and choice of penetrometer design are essential if improved estimates of soil resistance to root elongation are to be obtained. Resistance to root growth through pressurized cells contain- ing ballotini considerably exceeds the confining pressure applied externally to these cells. Results from this work are reappraised. Existing models of soil penetration by roots and penetrometers are reviewed together with the factors influencing penetration resistance. The interpretation of results from mechanical impedance experiments is examined in some detail and root responses, including possible mechanisms of response, are discussed.
I N T R O D U C T I O N The type of soil strength characteristic (i.e. the variation of soil strength with soil water content) favourable to crop growth depends on both the amount and the distribution of the annual rainfall, and on the nature of the crop. The soil must have sufficient mechanical strength to provide adequate anchorage for the plant throughout its development, and to prevent the collapse of soil water and air pathways by soil overburden pressure and the weight of vehicle and animal traffic. Dense regions of high strength may limit root growth and crop yield (Jamieson et al., 1988; Oussible, 1988) by creating a large mechanical resistance to root growth and/or restricting the rate of oxygen supply to roots. These dense regions occur in naturally compact soil horizons and also arise from compaction by heavy farm machinery and by the formation of plough pans.
Mechanical impedance is experienced to varying degrees by virtually all roots growing through soil. If continuous pores of sufficiently large diameter do not already exist, a root tip must exert a force to deform the soil. This process may considerably decrease root elongation rates, increase the root diameter and change the pattern of lateral root initiation (Russell, 1977).
In this paper, the effects of mechanical impedance on root morphology are reviewed and some direct comparisons between soil resistance to root growth and resistance to a penetrometer are discussed. The physical process of root growth through soil and artificial media is considered, with emphasis on the interpretation of results from different experimental techniques. Changes which occur in root elongation rate under both constant, and temporally and spatially varying levels of mechanical impedance are considered together with the complicating effects of soil aeration and water status. Finally, possible physiological mechanisms for the root responses are discussed.
Terminology Penetrometers provide the best estimates of resistance to root growth in soil, short of direct measure- ment of root force. Most penetrometers consist of a metal probe with a conical tip fixed onto a
34 1

342 A . G . Bengough & C. E. Mullins
cylindrical shaft (Fig. 1) that is generally of smaller diameter than the cone (normally 80% of the cone diameter; Gill, 1968; Barley & Greacen, 1967; Bengough, 1990). Penetrometer resistance, Q,, is defined in Equation (l), where F, is the force required to push the penetrometer probe through the soil, and A , is the cross-sectional area of the penetrometer cone:
Qp.r = Fp,rlAp,r (1)
The pressure that is exerted on the soil by a growing root cannot at present be measured at every point on the root surface. In this review, root penetration resistance, Q,, is defined similarly to penetrometer resistance, but where F, is the component of force directed along the root-axis that the section of root that moves through the soil must exert on the soil in order to extend, and A , is the root cross-sectional area, measured behind the elongation region. In common with the literature, the terms mechanical impedance and root penetration resistance have been used interchangeably.
- Shaft
7
Root hairs 41 ifl Elongating region
Meristemat i( region
Phloem
Endodermis
Cortex
Epidermis
Xylem
‘‘j ,,’/ f Muclgel sheoth
‘l;
I I
Fig. 1. (a) A penetrometer, where F,, A,, oN. and a are as defined in Equations (1 ) and (2). and (b) a root tip.
E F F E C T S OF M E C H A N I C A L I M P E D A N C E O N R O O T M O R P H O L O G Y
When a root tip encounters an obstacle that resists penetration, the root cap becomes less pointed and the surface cells may slough off (Souty, 1987). Mechanical impedance decreases the rate of root elongation because of both a decrease in the rate of cell division in the meristem, and a decrease in cell length (rather than volume). Eavis (1967) found a decrease of 40% in the cell division rate at a root penetration resistance (0.34 MPa) sufficient to decrease the root elongation rate by 70%. Cell length is decreased and the volume of the inner cortical cells may decrease, but the diameter and volume of the outer cortical and epidermal cells can be considerably greater (Barley, 1965; Wilson et al., 1977). The increase in root diameter in mechanically impeded roots results mainly from an increased thickness of the cortex; this is a consequence of both the increase in the diameter of the outer cells, and an increase in the number of cells per unit length of root.

Mechanical impedance to root growth 343
The apical meristem and zone of cell extension of impeded roots is shorter (Barley, 1962; Souty, 1987), and root hairs develop closer to the tip of impeded roots (Goss & Russell, 1980). Lateral initiation occurs nearer the tip and laterals occur together along the impeded axis (Goss & Russell, 1980; Barley, 1962). Where mechanical deflection causes roots to curve around an obstacle, the initiation of laterals generally occurs on the convex side of the root (Goss & Russell, 1980). Root hair development is greater on the opposite (concave) side and, in highly impeding media, the growing zone of the root is much distorted. The growth of impeded lateral roots is affected by impedance similarly to the main axis (Goss, 1977). However, if the pore size in the growing medium is such that only the main root axes are impeded, the freely penetrating laterals attain much greater length than in completely unimpeded root systems.
C O M P A R I S O N OF ROOT RESISTANCE WITH PENETROMETER RESISTANCE
There have been relatively few studies involving the measurement of root force (F, in Equation (1)) because of the experimental difficulties. Root force must be measured after the root has penetrated the surface of the soil to a depth of several times its diameter (since root penetration resistance is initially lower because the surface of the surrounding soil is displaced upwards; Gill, 1968), but before root hairs anchor the tip (Stolzy & Barley, 1968; Ennos, 1989). To calculate the root penetration resistance requires measurement of the root cross-sectional area. Root tip diameter increases in impeded roots and, since simultaneous measurements of root diameter and force can not normally be obtained, it is not obvious whether the initial or the final root diameter should be measured. Ideally, root diameter should be recorded just behind the elongating zone and level with the soil surface at the time of force measurement.
The results of experiments involving direct comparisons of root and penetrometer resistance indicate that penetrometers experience two to eight times greater resistance than plant roots penetrating soil (Table 1). Dexter (1987) suggests that this ratio of penetrometer resistance to root resistance is positively correlated with soil strength, being greater in ‘stronger’ soils. At present there is neither theoretical basis for this suggestion nor sufficient published data to justify such a conclusion, although the need for accurate prediction of this ratio is clear.
Indirect evidence for the difference between root and penetrometer resistance arises from com- paring the maximum pressures exerted by roots with penetrometer resistance in soil of sufficient strength to virtually halt root elongation. The maximum axial pressure that a root can exert is between about 0.9 MPa and 1.3 MPa (Misra et al., 1986b), whereas root elongation stops in soil with a penetrometer resistance of 0.8 to 5.0 MPa (Greacen et al., 1969). The results are variable because of differences between plant species and soil types, and possibly the temperatures at which the exper- iments were performed (Greacen, 1986). Thus, roots cease elongating in soil with a penetrometer resistance up to six or more times greater than the maximum axial pressure that they can exert. The reason for this difference must be physical differences in the way in which plant roots and metal probes penetrate soil.
M O D E L L I N G M E C H A N I C A L I M P E D A N C E T O P L A N T ROOTS A N D TO P E N E T R O M E T E R S I N SOIL
Barley & Greacen (1967) comprehensively reviewed the mechanics of soil deformation and failure which occur around penetrometer probes, roots and underground shoots. There have since been several attempts to predict penetration resistances in soil and in ballotini beads from bulk mechan- ical properties. All but one of these models estimate root resistance by predicting the theoretical pressure required to expand a cavity in the soil or ballotini. Penetrometer resistance, Qp, is then given by
Qp=a, ( l + p c o t a )
(Greacen et al., 1968) where uN is the pressure required to expand a cavity in the soil (and is equal to the normal stress on the surface of the penetrometer cone), p is the coefficient of soil-metal friction,
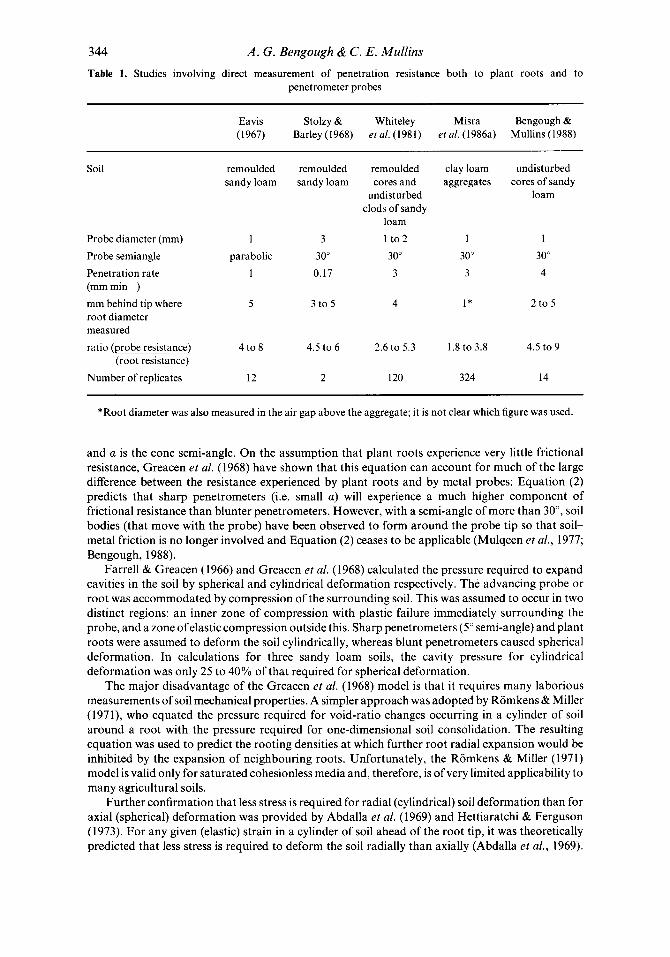
344 A . G . Bengough & C . E. Mullins
penetrometer probes Table 1. Studies involving direct measurement of penetration resistance both to plant roots and to
Eavis Stolzy & Whiteley Misra Bengough & (1967) Barley(1968) etal. (1981) etal . (1986a) Mullins(1988)
Soil
Probe diameter (mm) Probe semiangle Penetration rate (mm min- I)
mm behind tip where root diameter measured ratio (orobe resistance)
remoulded remoulded remoulded sandy loam sandy loam cores and
undisturbed clods of sandy
loam 1 3 1 t o2
parabolic 30" 30" 1 0.17 3
5 3 t 0 5 4
4 t 0 8 4.5 to 6 2.6 to 5.3
clay loam undisturbed aggregates cores of sandy
loam
1 I 30" 30" 3 4
I * 2 to 5
1.8 to 3.8 4.5 to 9 (root resistance)
Number of replicates 12 2 120 324 14
*Root diameter was also measured in the air gap above the aggregate; it is not clear which figure was used.
and a is the cone semi-angle. On the assumption that plant roots experience very little frictional resistance, Greacen et al. (1968) have shown that this equation can account for much of the large difference between the resistance experienced by plant roots and by metal probes: Equation ( 2 ) predicts that sharp penetrometers (i.e. small a) will experience a much higher component of frictional resistance than blunter penetrometers. However, with a semi-angle of more than 30", soil bodies (that move with the probe) have been observed to form around the probe tip so that soil- metal friction is no longer involved and Equation (2 ) ceases to be applicable (Mulqeen et al., 1977; Bengough, 1988).
Farrell & Greacen (1966) and Greacen et al. (1968) calculated the pressure required to expand cavities in the soil by spherical and cylindrical deformation respectively. The advancing probe or root was accommodated by compression of the surrounding soil. This was assumed to occur in two distinct regions: an inner zone of compression with plastic failure immediately surrounding the probe, and a zone of elastic compression outside this. Sharp penetrometers (5" semi-angle) and plant roots were assumed to deform the soil cylindrically, whereas blunt penetrometers caused spherical deformation. In calculations for three sandy loam soils, the cavity pressure for cylindrical deformation was only 25 to 40% of that required for spherical deformation.
The major disadvantage of the Greacen et al. (1968) model is that it requires many laborious measurements of soil mechanical properties. A simpler approach was adopted by Romkens & Miller (1971), who equated the pressure required for void-ratio changes occurring in a cylinder of soil around a root with the pressure required for one-dimensional soil consolidation. The resulting equation was used to predict the rooting densities at which further root radial expansion would be inhibited by the expansion of neighbouring roots. Unfortunately, the Romkens & Miller (1971) model is valid only for saturated cohesionless media and, therefore, is of very limited applicability to many agricultural soils.
Further confirmation that less stress is required for radial (cylindrical) soil deformation than for axial (spherical) deformation was provided by Abdalla et al. (1969) and Hettiaratchi & Ferguson (1973). For any given (elastic) strain in a cylinder of soil ahead of the root tip, it was theoretically predicted that less stress is required to deform the soil radially than axially (Abdalla et al., 1969).

Mechanical impedance to root growth 345 This theory was complemented by experiments using a large modified penetrometer to demonstrate that radial expansion behind a penetrometer (or root) tip can reduce axial resistance to soil pen- etration. Hettiaratchi & Ferguson (1973) predicted theoretically that the pressure required for cylindrical soil deformation in a frictionless cohesive medium was always less than for spherical deformation, the difference increasing with cohesion.
Collis-George & Yoganathan (1985) used the spherical cavity expansion model of VesiC (1972) to define limiting mechanical conditions for seed germination and root growth. Although this model may be suitable to describe germination conditions, use of spherical expansion theory will have resulted in overestimates of the resistance to root growth. The VesiC model requires fewer inputs than the Greacen model, and includes a volumetric strain term. Collis-George & Yoganathan assumed the volumetric strain to be zero, so that fewer soil mechanical measurements were needed to perform their calculation. However, their zero-strain assumption is questionable because even a tiny volumetric strain may, under certain conditions, alter the cavity pressure considerably (VesiC, 1972).
PHYSICAL D I F F E R E N C E S BETWEEN SOIL P E N E T R A T I O N BY P L A N T ROOTS A N D P E N E T R O M E T E R PROBES
Roots are flexible organs that follow tortuous paths through the soil, apparently seeking out the path of least resistance. They extract water from the soil, excrete mucilage from around their tips, and swell when physically impeded. In contrast, penetrometers are rigid metal probes constrained to a linear path through the soil. Penetrometers vary from about 0.1 mm in diameter for a small (needle) penetrometer (e.g. Groenevelt et al., 1984) to over lOmm for a large (field) penetrometer (the standard ASAE penetrometer cone has a diameter of 20.27mm; ASAE, 1969), and often penetrate the soil at rates up to two or more orders of magnitude greater than roots (Whiteley et al., 1981).
The differences between penetrometers and roots have resulted in the expression of much doubt as to the usefulness of penetrometers (e.g. Russell, 1977, p. 188), but despite their limitations they remain the best available method of estimating resistance to root growth in soil. It is important, therefore, to determine what are the most important physical differences between the action of roots and penetrometers.
Rootflexibility and spatial variation of soil strength Because roots often grow through cracks and holes in the soil, or follow planes of weakness between soil peds (Russell, 1977), penetrometers are of limited use in some structured soils. Detailed work has been done on the behaviour of roots growing along cracks and through pores (Whiteley & Dexter, 1983; Dexter, 1986; Scholefield & Hall, 1985), but is beyond the scope of this review. However, in coarsely structured soil, individual soil peds may be considered as continuous even though the soil is structured on a larger scale (Greacen et al., 1969) and root penetration into these peds may be important for nutrient uptake and plant growth. The forces required to buckle root tips growing across air gaps were measured by Whiteley & Dexter (198 1 ). The buckling stress decreased as the size of the air gap increased, but attempts to predict the buckling stress from the elastic modulus of the root tip were only partly successful. Dexter (1978) has modelled root growth through a bed of aggregates by relating root growth rates to penetrometer resistance within individual aggregates, and combining this with information on the probability of roots penetrating the aggre- gates. To date, this model has not been tested against independent experimental data over a range of realistic conditions.
Penetrometers average soil resistance in a zone surrounding the probe tip; thus, they cannot detect changes in soil strength that are on a scale much smaller than the tip dimensions. Groenevelt et al. (1984) investigated small-scale variations in strength by using a 0.15mm diameter pen- etrometer to determine the proportion of linear depth in a soil core with penetrometer resistance less than 1 MPa, and inferred that this fraction of the soil has a relatively low resistance to root growth. This ‘percentage linear penetrability’ of the soil decreased at higher soil bulk density, and good correlations between penetrability and rooting density have been obtained using a larger (1 3 mm
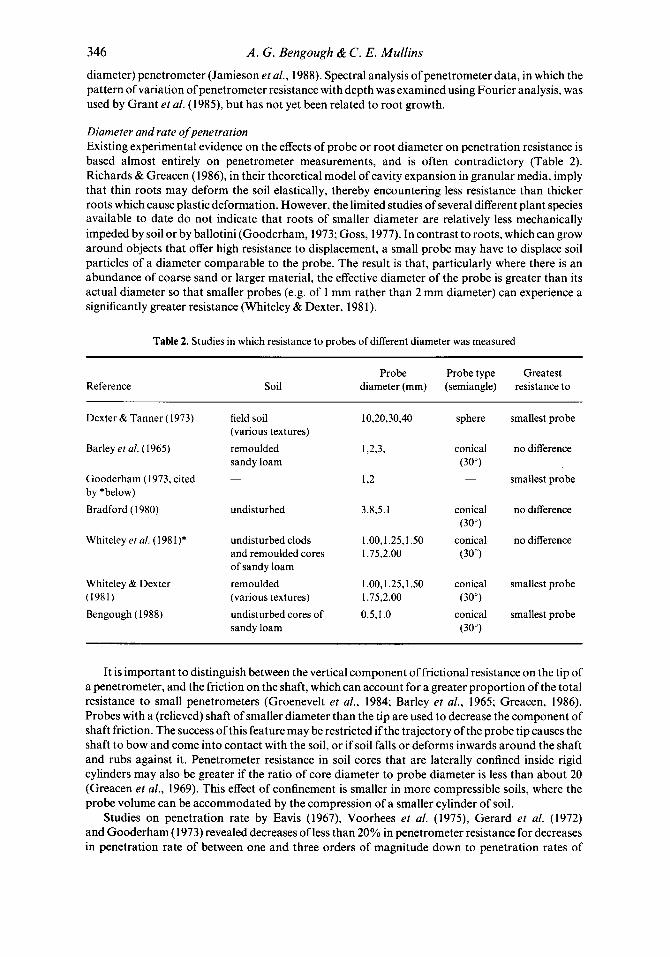
346 A . G . Bengough & C . E. Mullins
diameter) penetrometer (Jamieson et al., 1988). Spectral analysis of penetrometer data, in which the pattern of variation of penetrometer resistance with depth was examined using Fourier analysis, was used by Grant et al. (1989, but has not yet been related to root growth.
Diameter and rate ofpenetration Existing experimental evidence on the effects of probe or root diameter on penetration resistance is based almost entirely on penetrometer measurements, and is often contradictory (Table 2). Richards & Greacen (1986), in their theoretical model of cavity expansion in granular media, imply that thin roots may deform the soil elastically, thereby encountering less resistance than thicker roots which cause plastic deformation. However, the limited studies of several different plant species available to date do not indicate that roots of smaller diameter are relatively less mechanically impeded by soil or by ballotini (Gooderham, 1973; Goss, 1977). In contrast to roots, which can grow around objects that offer high resistance to displacement, a small probe may have to displace soil particles of a diameter comparable to the probe. The result is that, particularly where there is an abundance of coarse sand or larger material, the effective diameter of the probe is greater than its actual diameter so that smaller probes (e.g. of 1 mm rather than 2 mm diameter) can experience a significantly greater resistance (Whiteley & Dexter, 198 1).
Table 2. Studies in which resistance to probes of different diameter was measured
Reference Probe Probe type Greatest
Soil diameter (mm) (semiangle) resistance to
Dexter & Tanner (1973)
Barley ef al. (1965)
Gooderham (1973, cited by *below) Bradford (1980)
Whiteley ef al. (1981)*
Whiteley & Dexter (1981) Bengough (1988)
field soil (various textures) remoulded sandy loam
undisturbed
undisturbed clods and remoulded cores of sandy loam remoulded (various textures) undisturbed cores of sandy loam
10,20,30,40
3.8,5.1
1.00,1.25,1.50 1.75.2.00
1.00,1.25,1.50 1.75,2.00 0.5,l .O
sphere smallest probe
conical no difference
- smallest probe (30")
conical no difference
conical no difference (307
(30")
conical smallest probe
conical smallest probe (307
(307
It is important to distinguish between the vertical component of frictional resistance on the tip of a penetrometer, and the friction on the shaft, which can account for a greater proportion of the total resistance to small penetrometers (Groenevelt et al., 1984; Barley et al., 1965; Greacen, 1986). Probes with a (relieved) shaft of smaller diameter than the tip are used to decrease the component of shaft friction. The success of this feature may be restricted if the trajectory of the probe tip causes the shaft to bow and come into contact with the soil, or if soil falls or deforms inwards around the shaft and rubs against it. Penetrometer resistance in soil cores that are laterally confined inside rigid cylinders may also be greater if the ratio of core diameter to probe diameter is less than about 20 (Greacen et al., 1969). This effect of confinement is smaller in more compressible soils, where the probe volume can be accommodated by the compression of a smaller cylinder of soil.
Studies on penetration rate by Eavis (1967), Voorhees et al. (1975), Gerard et al. (1972) and Gooderham (1973) revealed decreases of less than 20% in penetrometer resistance for decreases in penetration rate of between one and three orders of magnitude down to penetration rates of

Mechanical impedance to root growth 347
1 mm h-’ or slower. Although Waldron & Constantin (1970) found a large effect of penetration rate, an intermittently rotated penetrometer was used, which would have resulted in larger decreases in soil frictional resistance at slower rates of penetration. In very wet soil, penetrometer resistance is more clearly linked to penetration rate because of its interaction with pore water pressure (Cockroft et al., 1969). This effect will be greater in less permeable soils (especially remoulded soil) containing a higher propertion of silt and clay, than in sands. Penetrometer resistance doubled in a sandy loam soil remoulded at approximately field capacity for a 100-fold increase in penetration rate, whereas a 250-fold increase in penetration rate resulted in only a 25% increase in penetrometer resistance in air-dry sand (Bengough, 1988). Similarly, Cockroft et al. (1969) found a doubling of penetration resistance for a 350-fold increase in penetration rate in saturated remoulded clay. Thus, excluding very wet and remoulded soils, penetrometer resistance is only weakly dependent on penetration rate for speeds between those normally used in needle penetrometer measurements and typical rates of root elongation.
Shape and friction Root or probe shape determines both the mode of soil deformation and the amount of frictional resistance on the tip. Observations of soil movement and density patterns surrounding probes and roots suggest that both narrowly tapered probes and plant roots deform the soil cylindrically compared with the spherical deformation caused by blunt probes (Cockroft et al., 1969; Greacen et al., 1968). By using both lubricated and rotated penetrometer probes it has been demonstrated that a large component of penetrometer resistance is frictional (Tollner & Verma, 1984; Bengough & Mullins, 1988). Greacen et al. (1968) suggested that root tips experience virtually no frictional resistance because of the lubricating action of mucilage secretion and the sloughing off of root cap cells. If this is so, the best estimate of root resistance may be obtained by measuring the resistance to a narrowly-tapered probe, and then subtracting the component of frictional resistance using Equation ( 2 ) (Greacen &Oh, 1972; Voorhees et al., 1975).
Interactions between roots, water extraction by roots, root swelIing and root nutation Because roots seldom grow through soil in complete isolation, it is important to consider inter- actions between neighbouring roots. Greacen et al. (1969) measured resistance to penetration of a narrowly-tapered probe, surrounded by six identical probes. Penetration resistance for the central probe of a group was considerably lower than when the probe was used on its own. Tensile cracking occurred between the probes, and similar cracks were also observed in a separate experiment between neighbouring pea radicles growing into a loam. The drying action of roots is very important in the formation of such cracks, which must facilitate the subsequent growth of lateral roots in a soil of high resistance (Gerard et al., 1972). Although this cracking is generally likely to be advan- tageous, there are soils of high tensile strength (e.g. Mullins et al., 1987, 1990) which may not crack readily under the drying action of roots. In such soils, the increase in penetration resistance caused by the soil drying may further impede root growth.
It has been suggested by Abdalla et al. (1969) that roots penetrate soil by an alternating series of radial and axial enlargements. Graf & Cooke (1980) used a finite-element model and, assuming a low coefficient of root-soil friction and that the soil behaved as a homogenous linear elastic medium, predicted that the radial expansion of impeded root tips could reduce the axial stress on the root cap. A zone of stress relief caused by radial enlargement was also predicted by Richards & Greacen (1986), and by Hettiaratchi & O’Callaghan (1974, 1978) who also analysed theoretically the mech- anics of the changes in cell size and shape that can occur in mechanically impeded root tips. It is clear that rigid penetrometers cannot mimic this radial expansion of roots, but the potential importance of this as a mechanism for soil penetration by roots has not yet been fully investigated.
Finally, oae factor rarely commented on with respect to soil penetration is the tendency for roots to nutate (Greacen et al., 1969; Ney & Pilet, 1981). The magnitude of this motion in highly resistant soil is probably very small, but it could be a process by which roots locate low-resistance pathways through heterogeneous soils, and may also reduce soil frictional resistance to root tip penetration. Nutation has also been suggested as a mechanism that aids soil penetration by rhizomes (Fisher, 1964).

348 A . G . Bengough & C . E. Mullins
E F F E C T S O F M E C H A N I C A L I M P E D A N C E O N ROOT G R O W T H
Experimental techniques Existing experimental techniques can be divided into several different catagories.
Soil. Experiments in soil are more realistic, but it is difficult to ensure that mechanical impedance is the only soil factor limiting root growth. If a soil is compacted to increase resistance to root growth, the resulting decrease in porosity may result in poor aeration. Similarly, increasing soil- water tension to increase soil strength may result in water stress. However, the greatest difficulty is in determining the penetration resistance experienced by roots in soil. This is ideally determined by direct measurement (e.g. Stolzy & Barley, 1968; Eavis & Payne, 1969; Bengough & Mullins, 1988), but practical difficulties have led most researchers to use penetrometer resistance measurements to estimate root resistance (e.g. Greacen &Oh, 1972).
Pressurizedcelis. Root growth has been studied in artificial systems where it was intended that a constant, uniform and known value of mechanical impedance could be imposed, while simul- taneously maintaining a carefully controlled supply of aerated nutrient solution to the roots. Goss (1977), Abdalla et al. (1969) and Barley (1963) grew roots in ballotini contained in flexible-sided cells which could be subjected to an external confining pressure (Fig. 2).
( a )
Pressure inlet
Root inlet ( b )
Root inlet
Gas inlet
Gas under pressure, ud Polythene diaphragm
Nylon cloth
u Lu 0 2 4 01 2
cm cm Fig. 2. (a) A triaxial cell (after Barley, 1963), and (b) a pressurized diaphragm apparatus (after Barley, 1962). Fig. 2(a) is reproduced from K.P. Barley, Influence of soil strength on growth of roots, Soil Science 1963,96(3), 175-180 (0 Williams & Wilkins, 1963).
Resistance to root growth in ballotini has been taken as either equal to (Russell & Goss, 1974; Goss, 1977; Goss & Russell, 1980; Veen, 1982), or an order of magnitude greater than (Richards & Greacen, 1986; Bengough & Mullins, 1990) the pressure applied externally to the boundary of the growth medium (u3 in Fig. 2). Goss (1977), Veen (1982), Richards & Greacen (1986) and Bengough & Mullins (1990) all estimated resistance to root growth by measuring the pressure required to inflate a small rubber tube within the pressurized ballotini cell. The tube inflation pressure depends entirely on the detailed procedure followed (Bengough & Mullins, 1990).
Goss (1977), Veen (1982) and Bengough & Mullins (1990) inserted a tube inflated under atmospheric pressure only and unsealed at one end into an unpressurized cell of ballotini. An external confining pressure was then applied to the cell, causing the tube partly to deflate. The
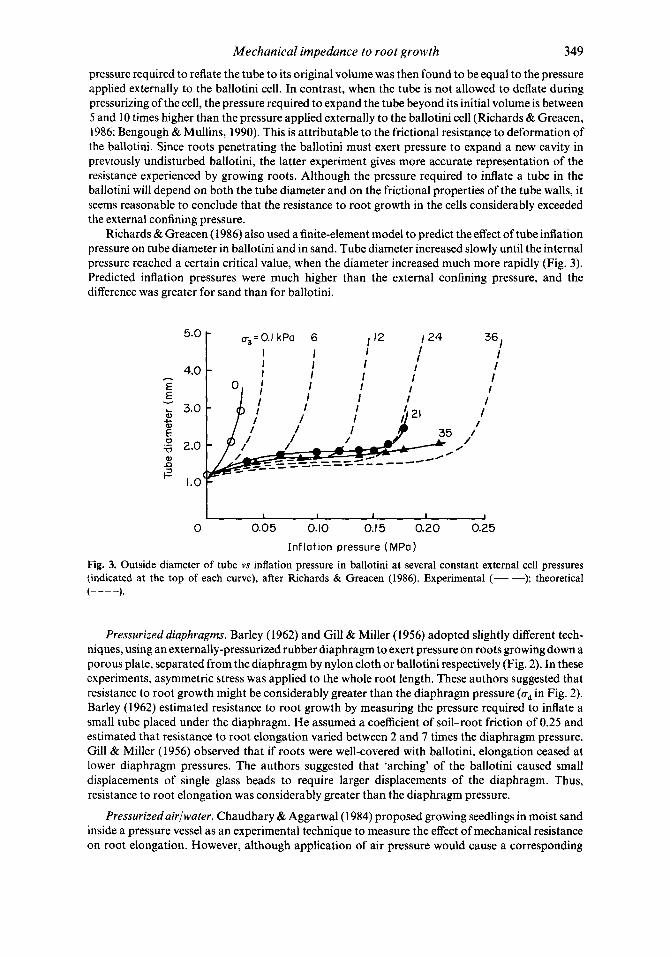
Mechanical impedance to root growth 349
pressure required to reflate the tube to its original volume was then found to be equal to the pressure applied externally to the ballotini cell. In contrast, when the tube is not allowed to deflate during pressurizing of the cell, the pressure required to expand the tube beyond its initial volume is between 5 and 10 times higher than the pressure applied externally to the ballotini cell (Richards & Greacen, 1986; Bengough & Mullins, 1990). This is attributable to the frictional resistance to deformation of the ballotini. Since roots penetrating the ballotini must exert pressure to expand a new cavity in previously undisturbed ballotini, the latter experiment gives more accurate representation of the resistance experienced by growing roots. Although the pressure required to inflate a tube in the ballotini will depend on both the tube diameter and on the frictional properties of the tube walls, it seems reasonable to conclude that the resistance to root growth in the cells considerably exceeded the external confining pressure.
Richards & Greacen (1986) also used a finite-element model to predict the effect of tube inflation pressure on tube diameter in ballotini and in sand. Tube diameter increased slowly until the internal pressure reached a certain critical value, when the diameter increased much more rapidly (Fig. 3). Predicted inflation pressures were much higher than the external confining pressure, and the difference was greater for sand than for ballotini.
E E v
I I
I I I I I I i I
I I i I I
3.0} I / I
3 6 ~ I I
I I
I I
I /
/ /
I 1 I 1 I I
0 0.05 0.10 0.15 0.20 0.25
Inflation pressure (MPa)
Fig. 3. Outside diameter of tube vs inflation pressure in ballotini at several constant external cell pressures (indicated at the top of each curve), after Richards & Greacen (1986). Experimental (-); theoretical (----)
Pressurized diaphragms. Barley (1962) and Gill & Miller (1956) adopted slightly different tech- niques, using an externally-pressurized rubber diaphragm to exert pressure on roots growing down a porous plate, separated from the diaphragm by nylon cloth or ballotini respectively (Fig. 2). In these experiments, asymmetric stress was applied to the whole root length. These authors suggested that resistance to root growth might be considerably greater than the diaphragm pressure (o,, in Fig. 2). Barley (1962) estimated resistance to root growth by measuring the pressure required to inflate a small tube placed under the diaphragm. He assumed a coefficient of soil-root friction of 0.25 and estimated that resistance to root elongation varied between 2 and 7 times the diaphragm pressure. Gill & Miller (1956) observed that if roots were well-covered with ballotini, elongation ceased at lower diaphragm pressures. The authors suggested that ‘arching’ of the ballotini caused small displacements of single glass beads to require larger displacements of the diaphragm. Thus, resistance to root elongation was considerably greater than the diaphragm pressure.
Pressurized airlwater. Chaudhary & Aggarwal(l984) proposed growing seedlings in moist sand inside a pressure vessel as an experimental technique to measure the effect of mechanical resistance on root elongation. However, although application of air pressure would cause a corresponding
350 A . G . Bengough & C . E. Mullins
increase in the absolute value of root cell turgor pressure, the decrease in osmotic potential or cell wall tension required for root cell extension would be independent of the externally applied pressure (assuming the cell permeability to water remained unchanged). Thus, the experiment does not truly represent the situation of roots elongating against an external mechanical resistance. The reason for the large decrease in root growth rates observed by the authors could be the 10-fold increase in dissolved gas concentration in the water inside the pressure chamber, and ultimately in the plant, resulting from the 10-fold increase in air pressure that they applied (Henry's law).
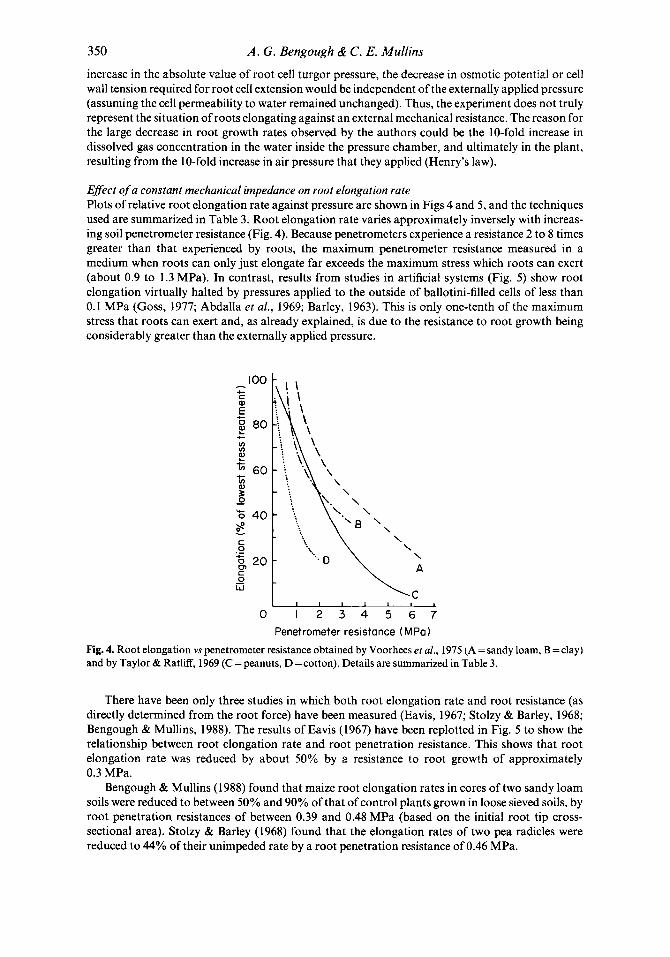
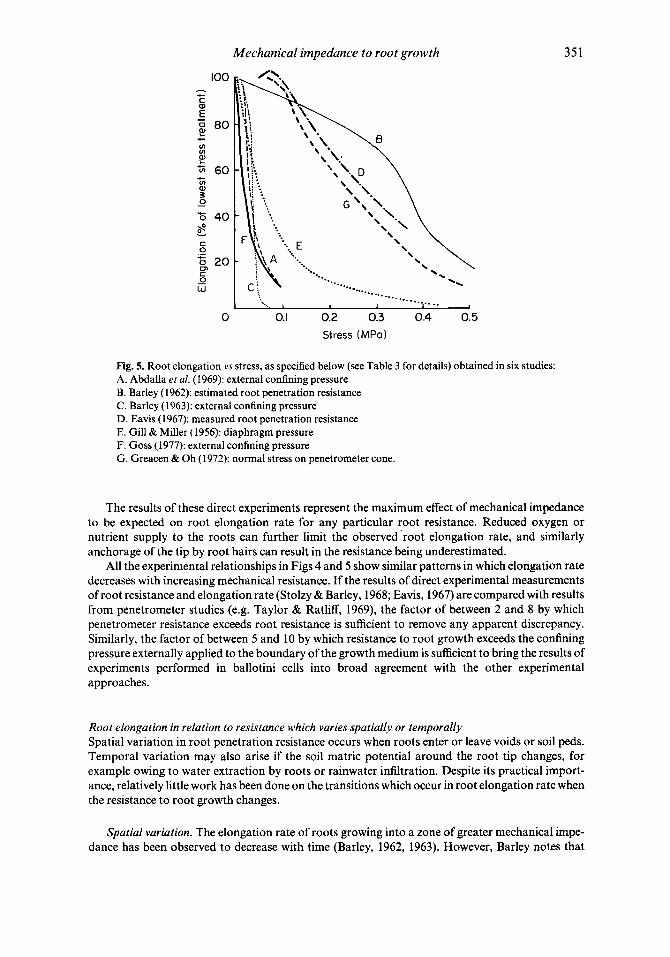
Effect of a constant mechanical impedance on root elongation rate Plots of relative root elongation rate against pressure are shown in Figs 4 and 5 , and the techniques used are summarized in Table 3. Root elongation rate varies approximately inversely with increas- ing soil penetrometer resistance (Fig. 4). Because penetrometers experience a resistance 2 to 8 times greater than that experienced by roots, the maximum penetrometer resistance measured in a medium when roots can only just elongate far exceeds the maximum stress which roots can exert (about 0.9 to 1.3 MPa). In contrast, results from studies in artificial systems (Fig. 5 ) show root elongation virtually halted by pressures applied to the outside of ballotini-filled cells of less than 0.1 MPa (Goss, 1977; Abdalla et al., 1969; Barley, 1963). This is only one-tenth of the maximum stress that roots can exert and, as already explained, is due to the resistance to root growth being considerably greater than the externally applied pressure.
m m ? 5; 60 t m W 3 0 -
c 0 .- 5 20 c 0 W -
Penetrometer resistance (MPa) Fig. 4. Root elongation vs penetrometer resistance obtained by Voorhees et al., 1975 (A = sandy loam, B =clay) and by Taylor & Ratliff, 1969 (C = peanuts, D =cotton). Details are summarized in Table 3.
There have been only three studies in which both root elongation rate and root resistance (as directly determined from the root force) have been measured (Eavis, 1967; Stolzy & Barley, 1968; Bengough & Mullins, 1988). The results of Eavis (1967) have been replotted in Fig. 5 to show the relationship between root elongation rate and root penetration resistance. This shows that root elongation rate was reduced by about 50% by a resistance to root growth of approximately 0.3 MPa.
Bengough & Mullins (1988) found that maize root elongation rates in cores of two sandy loam soils were reduced to between 50% and 90% of that of control plants grown in loose sieved soils, by root penetration resistances of between 0.39 and 0.48 MPa (based on the initial root tip cross- sectional area). Stolzy & Barley (1968) found that the elongation rates of two pea radicles were reduced to 44% of their unimpeded rate by a root penetration resistance of 0.46 MPa.
Mechanical impedance to root growth 35 1
100 - + c E + 0 80 P + ul ul
P 60
c 0 ._ 20
c 0 - W
0 0. I 0.2 0.3 0.4 0.5 Stress (MPa)
Fig. 5. Root elongation vs stress, as specified below (see Table 3 for details) obtained in six studies: A. Abdalla et al. (1969): external confining pressure 9. Barley (1962): estimated root penetration resistance C. Barley (1963): external confining pressure D. Eavis (1967): measured root penetration resistance E. Gill & Miller (1956): diaphragm pressure F. Goss (1977): external confining pressure G. Greacen & Oh (1972): normal stress on penetrometer cone.
The results of these direct experiments represent the maximum effect of mechanical impedance to be expected on root elongation rate for any particular root resistance. Reduced oxygen or nutrient supply to the roots can further limit the observed 'root elongation rate, and similarly anchorage of the tip by root hairs can result in the resistance being underestimated.
All the experimental relationships in Figs 4 and 5 show similar patterns in which elongation rate decreases with increasing mechanical resistance. If the results of direct experimental measurements of root resistance and elongation rate (Stolzy & Barley, 1968; Eavis, 1967) are compared with results from penetrometer studies (e.g. Taylor & Ratliff, 1969), the factor of between 2 and 8 by which penetrometer resistance exceeds root resistance is sufficient to remove any apparent discrepancy. Similarly, the factor of between 5 and 10 by which resistance to root growth exceeds the confining pressure externally applied to the boundary of the growth medium is sufficient to bring the results of experiments performed in ballotini cells into broad agreement with the other experimental approaches.
Root elongation in relation to resistance which varies spatially or temporally Spatial variation in root penetration resistance occurs when roots enter or leave voids or soil peds. Temporal variation may also arise if the soil matric potential around the root tip changes, for example owing to water extraction by roots or rainwater infiltration. Despite its practical import- ance, relatively little work has been done on the transitions which occur in root elongation rate when the resistance to root growth changes.
Spatial variation. The elongation rate of roots growing into a zone of greater mechanical impe- dance has been observed to decrease with time (Barley, 1962, 1963). However, Barley notes that
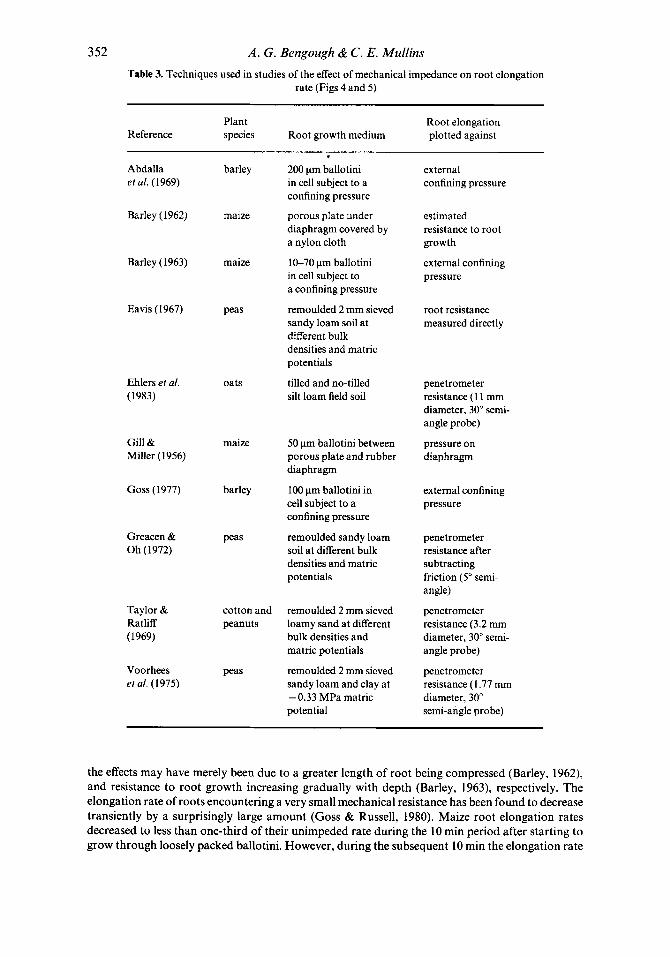
352 A . G . Bengough & C . E. Mullins Table 3. Techniques used in studies of the effect of mechanical impedance on root elongation
rate (Figs 4 and 5 )
Plant Root elongation Reference species Root growth medium plotted against
Abdalla e? a/ . (1969)
Barley (1962)
Barley (1963)
Eavis (1967)
Ehlers et a/ . (1983)
Gill & Miller (1956)
Goss (1977)
Greacen & Oh (1972)
Taylor & Ratliff (1 969)
Voorhees et a/ . (1 975)
barley
maize
maize
peas
oats
maize
barley
cotton and peanuts
peas
200 pm ballotini in cell subject to a confining pressure
porous plate under diaphragm covered by a nylon cloth
1C70 pm ballotini in cell subject to a confining pressure
remoulded 2 mm sieved sandy loam soil at different bulk densities and matric potentials
tilled and no-tilled silt loam field soil
50 pm ballotini between porous plate and rubber diaphragm
100 pm ballotini in cell subject to a confining pressure
remoulded sandy loam soil at different bulk densities and matric potentials
remoulded 2 mm sieved loamy sand at different bulk densities and matric potentials
remoulded 2 mm sieved sandy loam and clay at -0.33 MPa matric potential
external confining pressure
estimated resistance to root growth
external confining pressure
root resistance measured directly
penetrometer resistance ( 1 1 mm diameter, 30" semi- angle probe)
pressure on diaphragm
external confining pressure
penetrometer resistance after subtracting friction (5" semi- angle)
penetrometer resistance (3.2 mm diameter, 30" semi- angle probe)
penetrometer resistance (1.77 mm diameter, 30" semi-angle probe)
the effects may have merely been due to a greater length of root being compressed (Barley, 1962), and resistance to root growth increasing gradually with depth (Barley, 1963), respectively. The elongation rate of roots encountering a very small mechanical resistance has been found to decrease transiently by a surprisingly large amount (Goss & Russell, 1980). Maize root elongation rates decreased to less than one-third of their unimpeded rate during the 10 min period after starting to grow through loosely packed ballotini. However, during the subsequent 10 min the elongation rate

Mechanical impedance to root growth 353
returned to its original unimpeded value. Identical experiments performed using roots that first had their root caps removed showed no significant changes in root growth rate. Thus, the root cap must have a role in regulating root response to very low resistances, although it is uncertain whether the same mechanism operates at the greater resistances more often found in soil.
Temporal variation. Mechanical pressure applied to the apical 15 mm of previously unimpeded roots can halt root elongation which resumes only after a lag time (13 to 50 h), which itself increases with increasing stress (Barley, 1962). Localized moderate pressure (0.1 MPa) applied to the root apex results in a bigger reduction in elongation rate than when fully enlarged tissue is compressed (Barley, 1965). Higher pressures applied to the older tissue caused browning, loss of turgor and stopped elongation, although no tissue damage resulted if the root cells were allowed to enlarge and mature under the mechanical pressure (Barley, 1965). When the external confining pressure is reduced to zero, mechanically impeded roots growing in pressurized cells of ballotini only gradually return to the unimpeded elongation rate (Goss & Russell, 1980). The delay, of 2 to 7 d, was longer for the more severely impeded roots. Thus, roots which develop under mechanical impedance probably undergo physiological changes to adapt to the stress.
Interaction of mechanical impedance with matric potential and aeration Roots grow more slowly in poorly aerated soil (Blackwell & Wells, 1983; Greenwood, 1969) and in soil of low (i.e. more negative) matric potential (Eavis & Payne, 1969; Eavis, 1972; Yappa et al., 1988): but do these two stresses interact with mechanical impedance to produce an effect that is greater than would be expected from each stress acting independently?
Gill & Miller (1956) and Barley (1962) grew maize roots in ballotini compressed by a diaphragm and circulated with nutrient solution containing different concentrations of oxygen. Gill & Miller (1956) found that root elongation ceased when the oxygen concentration was reduced to 1% in mechanically impeded roots at an externally applied diaphragm pressure of 0.15 MPa or greater, whereas growth continued in the better-aerated treatments. Barley (1962) found that increasing the externally applied diaphragm pressure from zero to 0.05 MPa resulted in a 31% decrease in root elongation rate for roots grown in solution containing a 20% oxygen concentration, whereas a 50% reduction in root elongation rate resulted in roots grown in solution containing a 5% oxygen concentration. Goss et al. (1989) found a small interaction of mechanical impedance with aeration in maize roots, but the interaction was not statistically significant in wheat. The explanation for the interaction between oxygen deficiency and mechanical impedance is still uncertain, but may be due to mechanical impedance changing the morphology of the cortical root cells and the spaces between these cells. This might result in a shorter and a more tortuous path for oxygen transport along the root to the tip, and for ethylene transport away from the tip.
It is more difficult to ascertain whether low matric potentials interact with mechanical impedance, because soil strength and root penetration resistance increase as the matric potential decreases. Taylor & Ratliff (1969) measured the root elongation rates of cotton and peanuts in remoulded soil at several different bulk densities and matric potentials in the range -17 to -700 kPa for cotton, and - 19 to - 1250 kPa for peanuts. Root elongation rate clearly depended on the penetrometer resistance of the soil and not on the matric potential per se. Similar results were obtained by Greacen & Oh (1972) for peas at matric potentials below -0.5 MPa, and by Taylor & Gardner (1963) for cotton root penetration at matric potentials between - 20 kPa and - 67 kPa. These results imply that there is no interaction between matric potential and mechanical impedance.
In contrast to these findings, Mirreh & Ketcheson (1972) found that, in soil with penetrometer resistance in the range 0 to 2.5 MPa, the root elongation rate of maize was progressively reduced for any given penetrometer resistance as matric potential was decreased from -0.1 to -0.8 MPa. The reduction in elongation rate was greatest in soil with high penetrometer resistance, suggesting an interaction between matric potential and mechanical impedance.
A source of uncertainty in these experiments is that the matric potential occurring close to the root surface may be considerably lower than that in the bulk of the soil, and will decrease as the

354 A . G . Bengough & C . E. Mullins
transpirational demand increases (Cowan, 1965; Hainsworth & Aylmore, 1986). Thus, roots grow- ing in soils of differing bulk matric potential may experience similar matric potentials at the root surface (Mirreh & Ketcheson, 1972).
Possible mechanisms for the response of roots to mechanical impedance The growth rate of root cells is controlled by the balance of the internal and external pressures together with the rheological properties of the cell wall. Lockhart’s (1965) equation for cell growth was modified by Greacen (1986) to allow for root penetration resistance gr (Pa) giving,
dlldt = ZKrlm(n - W - 0,)/(2K + r’m) ( 3 )
where dl/dt is the rate of increase in cell length (m s-’), K(m Pa-ls-’) is the water permeability of the cell membrane per unit surface area, m (m-’Pa-Is-’) is the cell wall extensibility, n (Pa) is the osmotic pressure, r (m) is the cell radius and W(Pa) is the yield stress of the cell wall. Greacen (1986) used a simplified version of the above equation together with Greacen & Oh’s (1972) model of osmotic adjustment to generate a relationship between soil resistance and root elongation rate. The calculations produced a curve similar to that found by Eavis (1967) and Greacen & Oh (1972) by assuming a progressive decrease in the ratio of cell length to diameter, and also a linear reduction in cell production with increase in root penetration resistance.
Greacen & Oh (1972) proposed that root cells adjust their osmotic potential (the term ‘osmo- regulation’ has recently been criticized as misleading; Munns, 1988; Reed, 1984) to compensate for increased soil resistance and for low soil-water potential. The efficiency of this osmotic adjustment was measured to be 70% for mechanical impedance and 100% for water potentials down to -0.8 MPa. The agreement of their results with such an ‘efficiency’ hypothesis was questioned by Russell & Goss (1974) although Greacen (1986) argued that any discrepancy was caused by the high degree of variability associated with this aspect of root behaviour. Atwell (1988) recently suggested that the decrease in osmotic potential observed in mechanically impeded root tips occurred because of a simple build-up of solutes because of the slower root growth rate. Such an excess of solute supply over demand has also been suggested as the source of osmotic adjustment in plants growing in dry or saline soils (Munns, 1988).
However, a model of root response to mechanical impedance based entirely on osmotic adjust- ment does not explain the many morphological changes that occur in mechanically impeded roots. Barley (1976) and Feldman (1984) suggested that ethylene may be involved in root response to mechanical impedance. Kays et al. (1974) noted that the rate of ethylene evolution increased by up to six times when bean roots were impeded by a barrier. Exogenous applications of ethylene can cause root radial thickening, a reduction in the root elongation rate, and a profusion of root hairs within 1 to 2 mm of the swollen root apices (Kays et al., 1974; Smith & Robertson, 197 I); all of these have been observed in mechanically impeded roots.
Cell shape can be determined by the orientation of the cellulose microfibrils as they are laid down in the cell wall (Osborne, 1976). Initially these are aligned transversely to the root axis, but as cellular elongation begins to slow, the orientation of the microfibrils becomes more oblique (Hogetsu, 1986; Barlow, 1989). Veen (1982) observed longitudinal deposition of microfibrils in mechanically impeded roots, which resulted in shorter fatter cells. Such longitudinal deposition also occurs in roots exposed to ethylene (Osborne, 1976). The smaller diameter of the inner compared with the outer cortical cells, observed in impeded roots by Wilson et al. (1977), may have resulted from the innermost cell layers experiencing considerable wall pressure from the outer cells (Veen, 1982). Thus, despite the presence of longitudinal microfibrils, lateral expansion of the innermost cells could not occur.
Dexter (1987) presented a physical model of root growth in terms of the balance of pressures acting on the root, based largely on reworking published results using previously derived empirical formulae. An implicit assumption of this model is that the cell wall ‘extensibility’ is independent of the mechanical impedance experienced by the root. Surprisingly he does not comment on this assumption, although the evidence on microfibril deposition already presented suggests that cell wall extensibility may be restricted in the longitudinal direction. 3,5-diiodo-4-hydroxy benzoic acid

Mechanical impedance to root growth 355 (DIHB) is thought to be antagonistic to endogenous ethylene production/action, and increases the elongation rate of mechanically impeded roots, possibly by influencing the action of ethylene on cell wall extensibility (Goss et al., 1987).
Recently the involvement of ethylene in root response to mechanical impedance has been seriously questioned (Moss et al., 1988). Moss et al. measured a two- to 2.5-fold increase in the rate of ethylene evolution by mechanically impeded maize roots, and found that supplying ethylene in the air-flow to unimpeded roots simulated closely the effects of impedence. However, two inhibitors of ethylene production, aminoethoxyvinylglycine, and action, 2,5-norbornadiene, which overcame the effects of ethylene in unimpeded roots, did not modify the growth of mechanically impeded roots, although the increased rate of ethylene evolution was entirely suppressed.
Moss et al. (1988) suggest that the increased rate of ethylene evolution by impeded roots may not be a result of a direct effect of mechanical impedance, but of the physical wounding of the radially expanding root by the ballotini (such self-inflicted damage has been observed during time-lapse studies of bean root growth in ballotini; personal observation, AGB). However, the wounding hypothesis does not entirely explain why an increase in the rate of ethylene evolution occurred within 1 h ofwhen bean roots grown inside perforated tubes were mechanically impeded by a plastic barrier (Kays et al., 1974). The likelihood of physical wounding of the root tissue in this system should be considerably smaller than in the ballotini, although it is possible that some damage to the root tip occurred as it buckled upon reaching the barrier. Clearly the role of ethylene in the response of roots to mechanical impedance must be re-examined, as much of the supporting evidence for its involvement is largely correlative.
CONCLUSIONS
Except where direct measurements of root force are available, penetrometer resistance is currently the best method for estimating resistance to root growth in soil, although subject to many limi- tations. The relationship between penetrometer resistance and root resistance needs to be more clearly established by direct measurements of both root force and penetrometer resistance in a variety of differing soils over a range of bulk densities and matric potentials. It is probable that improved estimates of root resistance may be obtained by subtracting the frictional component of penetrometer resistance (determined by independent measurement of the coefficient of soil-metal friction) from the total resistance to a narrowly tapered probe. The effects on penetration resistance of penetrometer or root diameter still require a fuller investigation.
Root elongation rate is progressively decreased by increasing mechanical resistance to growth, and ceases at root penetration resistances of about 1 MPa. Resistance to root elongation within a ballotini cell confined by an externally applied pressure may be up to one order of magnitude greater than the externally applied pressure. The mistaken interpretation of results from such experiments has caused the apparent disagreement with results from penetrometer studies in soil. Pressurized ballotini cells still provide a valuable technique for studying the effects of mechanical impedence on root morphology and nutrient uptake (Goss, 1977; Lindberg & Pettersson, 1985), and are poten- tially of considerable use for investigating the largely unresearched ability of different species and varieties to grow against mechanical impedance (Goss, 1974). The changes in root growth rate which occur as a root enters or leaves a zone of high mechanical impedance are of considerable relevance to real soils, but have received surprisingly little attention.
The mechanism of root response to mechanical impedance may involve osmotic adjustment, although insufficient evidence has been presented to test such a model. The role of ethylene must be re-assessed in the light of some recent experiments, and its type of involvement in the response of roots to mechanical impedance must be more clearly established.
A C K N O W L E D G E M E N T S
We thank Drs D. Linehan, B. Marshall, I. Young, D. Robinson and other colleagues at SCRI for their helpful discussions and constructive criticism of this manuscript.

356 A . G. Bengough & C. E. Mullins
R E F E R E N C E S
ABDALLA, A.M., HETTIARATCHI, D.R.P. & REECE, A.R. 1969. The mechanics of root growth in granu- lar media. Journal of Agricultural Engineering Research 14,263-248.
ASAE 1969. ASAE Recommendation: ASAE R313: Soil Cone Penetrometer. American Society of Agricultural Engineers, St. Joseph, MI.
ATWELL, B.J. 1988. Physiological response of lupin roots to soil compaction. Plant and Soil 111,
BARLEY, K.P. 1962. Theeffects ofmechanical stresson the growth of roots. Journal of Experimental Botany 13,95-110.
BARLEY, K.P. 1963. Influence of soil strength on growth of roots. Soil Science 96,175-180.
BARLEY, K.P. 1965. The effect oflocalized pressure on the growth of the maize radicle. Australian Journal of Biological Science 18,499-503.
BARLEY, K.P. 1976. Mechanical resistance of the soil in relation to the growth of roots and emerging shoots. Agrochimica 20,173-182.
BARLEY, K.P. & GREACEN, E.L. 1967. Mechanical resistance as a soil factor influencing the growth of roots and underground shoots. Advances in Agronomy 19.1-40.
BARLEY, K.P., GREACEN, E.L. & FARRELL, D.A. 1965. The influence of soil strength on the penetration of a loam by plant roots. Australian Journal of Soil Research 3,69-79.
BARLOW, P.W. 1989. Anatomical controls of root growth. Aspects of Applied Biology 22,57-66.
BENGOUGH, A.G. 1988. The use of penetrometers in estimating mechanical impedance to root growth. Ph.D. thesis, University of Aberdeen.
BENGOUGH, A.G. 1990. The penetrometer in relation to mechanical resistance to root growth. In Soil Analysis: Physical Methods (eds K.A. Smith & C.E. Mullins), pp. 431435. Marcel Dekker, New York.
BENGOUGH, A.G. & MULLINS, C.E. 1988. Use of a low- friction penetrometer to estimate soil resistance to root growth. Proceedings of the 11th Conference of the International Soil Tillage Research Organisation
BENCOUGH, A.G. & MULLINS, C.E. 1990. The resist- ance experienced by roots growing in a pressurised cell-a reappraisal. Plant and Soil 123.73-82.
BLACKWELL, P.S. & WELLS, E.A. 1983. Limiting oxy- gen flux densities for oat root extension. Plant and Soil73,129-139.
BRADFORD, J.M. 1980. The penetration resistance of a soil with well defined structural units. Soil Science Society of American Journal44,601-606.
CHAUDHARY, M.R. & AGGARWAL, G.C. 1984. A simple technique to evaluate the effect of mechan- ical stress on root growth. Journal of Agricultural Science 102,79-80.
COCKROFT, B., BARLEY, K.P. & GREACEN, E.L. 1969. The penetration of clays by fine probes and root tips. Australian Journalof Soil Research 7,333-348.
COLLIS-GEORGE, N. & YOGANATHAN, P. 1985. The effect of soil strength on germination and emerg- ence of wheat (Triticum aestivum L.). 11. High shear strength conditions. Australian Journal of Soil Research 23,589-601.
277-28 1.
1, 1-6.
COWAN, I.R. 1965. Transport of water in the soil- plant-atmosphere system. Journal of Applied
DEXTER, A.R. 1978. A stochasticmodel for the growth of roots in tilled soils. Journal of Soil Science 29,
DEXTER, A.R. 1986. Model experiments on the behav- iour of roots at the interface between a tilled seed- bed and a compacted sub-soil. 11. Entry of pea and wheat roots into subsoil cracks. Plant and Soil 95,
DEXTER, A.R. 1987. Mechanics of root growth. Hunt andSoil98,303-312.
DEXTER, A.R. & TANNER, D.W. 1973. The force on spheres penetrating soil. Journal of Terramechanics
EAVIS, B.W. 1967. Mechanical impedance to root growth. Agricultural Engineering Symposium, Silsoe. Paper 4/F/39, pp. 1-1 1.
EAVIS, B.W. 1972. Soil physical conditions affecting seedling root growth. I. Mechanical impedance. aeration and moisture availability as influenced by bulk density and moisture levels in a sandy loam soil. Plant and Soil36,613-622.
EAVIS, B.W. & PAYNE, D. 1969. Soil physical con- ditions and root growth. In Root Growth (ed. W.J. Whittington), pp. 315-336. Butterworths, London.
EHLERS, W., KOPKE, U., HESSE, F. and BOHM, W. 1983. Penetration resistance and root growth of oats in tilled and untilled Loess soil. Soil and Tillage Research 3,261-275.
ENNOS, R. 1989. The mechanics of anchorage in seed- lings of sunflower, Helianthus annuus L. New Phyrologist 113, 185-192.
FARRELL, D.A. & GREACEN, E.L. 1966. Resistance to penetration of fine probes in compressible soil. Australian Journal of Soil Research 4, 1-1 7.
FELDMAN, L.J. 1984. Regulation of root development. Annual Review of Plant Physiology 35,223-242.
FISHER, J.E. 1984. Evidence of circumnutational growth movements of rhizomes of Poapratensis L. that aid in soil penetration. Canadian Journal of Botany 42,293-299.
GERARD, C.J., MEHTA, H.C. & HINOJOSA, E. 1972. Root growth in a clay soil. Soil Science 114,37-49.
GILL, W.R. 1968. Influence of compaction hardening of soil on penetration resistance. Transactions of the American Society of Agricultural Engineers 11,
GILL, W.R. &MILLER, R.D. 1956. A method for study of the influence of mechanical impedance and aeration on the growth ofseedling roots. SoilScience Society of America Proceedings 20, 154-1 57.
GOODERHAM, P.T. 1973. Soil physical conditions and plant growth. Ph.D. Thesis, University of Reading.
GOB, M.J 1974. Effects of mechanical impedance on root growth. Ph.D. Thesis, University of Reading.
Goss, M.J. 1977. Effects of mechanical impedance on root growth in barley (Hordeum vulgare L.). I. Effects on elongation and branching of seminal roots. Journal of Experimental Botany 28,96-11 I .
Goss, M.J. & RUSSELL, R.S. 1980. Effects of mechan- ical impedance on root growth in barley (Hordeum vulgare L.). 111. Observations on the mechanism
Ecology 2,221-239.
102-1 16.
135-147.
9,31-39.
74 1-753.

Mechanical impedance to root growth 357 of response. Journal of Experimental Botany 31, 577-588.
Goss, M.J., BARRACLOUGH, P.B. & POWELL, B.A. 1989. The extent to which physical factors in the rooting zone limit crop growth. Aspects of Applied Biology22,173-183.
Go% M.J., DEXTER, A.R. & EVANS, M. 1987. Mechanics of root elongation and the effects of3,5- diiodo-4-hydroxybenzoic acid (DIHB). Plant and S0il99,211-218.
GRAF, G.L. &COOKE, J.R. 1980. Soil stressesaround a growing root tip. Proceedings of the American Society of Agricultural Engineers: 80-1024.
GRANT, C.D., KAY, B.D., GROENEVELT, P.H.9 KIDD, G.E. & THURTELL, G.W. 1985. Spectral analysis of micropenetrometer data to characterize soil struc- ture. Canadian Journal ofsoil Science 65,789-804.
GREACEN, E.L. 1986. Root response to soilmechanical properties. Transactions of the 13th Congress of the International Society of Soil Science 5,20-47.
GREACEN, E.L. OH, J.s. 1972. Physics Of root growth. Mature, New Biology 235,24-25.
The mechanics of root growth in soils with particu- lar reference to the implications for root distri-
pp. 256-268. Butterworths, London.
LOCKHART, J.A. 1965. An analysis of irreversible cell elongation. Journal of Theoretical Bioiogy 8, 264-275.
MIRREH, H.F. & KETCHESON, J.W. 1972. Influence of soil water matric potential on corn root elongation. Canadian Journal of Soil Science 53,383-388.
MISRA, R.K., DEXTER, A.R. &ALSTON, A.M. l986a. Penetration ofsoil aggregates of finite size. 11. Plant roots. Plant and Soil 94,59-85.
MISRA, R.K., DEXTER, A.R. & ALSTON, A.M. 1986b. Maximum axial and radial growth pressures of plant roots, Plant andSoil95,315-326.
Moss, G.I., HALL, K.C. and JACKSON, M.B. 1988. Ethylene and the responses of roots of maize (Zea mays L.) to physical impedance. New Phytologist 109,303-31 1,
M ~ ~ ~ ~ ~ ~ , c,E,, yOUNG, I,M,, B E N ~ U G H , A,G, & L ~ ~ , G.J, 1987, Hard setting soils, soit use and M~~~~~~~~~ 3, 79-83,
MULLINS, C.E., MACLEOD, D.A., NORTHCOTE, K.H., TISDALL, J.M. & YOUNG, I.M. 1990. Hardsetting soils: Behaviour, occurrence and managment.
M ~ ~ ~ ~ ~ ~ ~ , J,, sTAFFORD, J.V. & T ~ ~ ~ ~ ~ , D,w. 1977, Evaluation of penetrometers for measuring soil
MUNNS, R. 1988. Why measure osmotic adjustment? Australian Journal of Plant Physiology 15,717-726.
georeacting roots, and Environment 4, 339-343.
OSBORNE, D.J. 1976. Control ofcell shape and cell size
Perspectives in Experimental Biology, Vol. 2 (ed. N'
ousslBLE, M. 1988. Effect Of subsoiling a compacted clay loam soil on the nitrogen uptake ofwheat. Pro- ceedings Of the ' I t h Conference Of lhe International
GREACEN, E.L., BARLEY, K.P. & FARRELL, D.A. 1969. Advances in ~ ~ i l ~ ~ i ~ ~ ~ ~ 11,37-107.
bution. In Root Growth (ed. w.J. Whittington), strength, Journal of Terramechanics 14, 137-1 51,
GREACEN3 E'L', FARRELL3 D'A' ' CoCKRoFT, B' 1968.
Transactions of the 9th Congress of the International Society of Soil Science 1,769-779.
GREENWOOD, D.J. 1969. Effect of oxygen distribution in the soil on plant growth, In Root Growth (ed,
London.
'Oil resistance to probes and plant roots' N E ~ , D, & PILET, P.E, 1981. Nutation ofgrowing and
W,J, Whittington), pp, 202-221. Butterworths, by the regulation Of auxin and ethylene' In
GROENEVELT, P.H., KAY, B.D. & GRANT, C.D. 1984. pp' 89-102' 'ergamon' Oxford' physical assessment ofa with respect to rooting
HAINSWORTH, J.M. & AYLMOE, L.A,G, 1986, Water extraction by single plant roots, soil Science society
potential. Geoderma 34, 101-1 14.
Research Organisation 2, 797-802. ofAmerica Journal 50,841-848.
HETTIARATCHI, D.R.P. & FERGUSON, C.A. 1973. Stress deformation behaviour of soil in root growth
Research IS, 309-320. in granular media. Australian Journal of Soil HETTIARATCHI, D.R.P. & ~'CALLAGHAN, J.R. 1974. Research 249 393-404.
A membrane model of plant cell extension. Journal ROMKENS, M.J.M. ~4 MILLER, R.D. 1971. Predicting of Theoretical Biology 45,459-465. root size and frequency from one-dimensional con-
HETT~ARATCH~, D,R,P, & OcALLAGHAN, J,R, 1978, solidation data-a mathematical model. Plant and Structural mechanics of plant cells. Journal of Theoretical Biology 14,235-257. RUSSELL, R.S., 1977. Plant Root Systems: Their
HOGETSU, T. 1986. Orientation of wall microfibril Function and Interaction wifh rhe Soil. McGraw- deposition in root cells of Pisum sativum L. var. Hill, London. Alaska. Plant Cell Physiology 27,947-951, RUSSELL, R.S. & Goss, M.J. 1974. Physical aspects of
JAMIESON, J.E., MORRIS, R.J. & MULLINS, C.E. 1988. soil fertility-the response of roots to mechanical Effect of subsoiling on physical properties and crop impedance. Netherlands Journal of Agricultural growth on a sandy soil with a naturally compact Science 2% 305-318. subsoil. Proceedings of the Zlth Conference of the SCHOLEFIELD, D. & HALL, D.M. 1985. Constricted International Soil Tillage Research Organisation 2, growth of grass roots through rigid pores. Plant and 499-503. Soil85,153-162.
KAYS, S.J., NICKLOW, C.W. & SIMONS, D.H. 1974. SMITH, K.A. & ROBERTSON P.D. 1971. Effect of Ethylene in relation to the response of roots to ethylene on root extension in cereals. Nature 234,
LINDBERG, S. & PETERSON, S. 1985. Effects ofmechan- SOUTY, N. 1987. Aspect mkhanique de la croissance ical stress on uptake and distribution of nutrient in des racines. I. Mesure de la force de ptnetration. barley. Plant and Soil 83,295-309. Agronomie 7,623-630.
REED, R.H. 1984. Useandabuseofosmo-terminology. Celland Enviromenr7~ 165-170.
B.G. 8~ GREACENy E.L. 1986. mechanics, Journal of Agricultural Engineering stresses on an expanding root analogue
237-248.
physical impedance. Plant and Soil40,565-571. 148-149.

358 A . G . Bengough & C . E. Mullins
(Received24 May 1989; accepted29 March 1990)
STOLZY, L.H. & BARLEY, K.P. 1968. Mechanical resistance encountered by roots entering compact soils. Soil Science 105,297-301.
TAYLOR, H.M. & GARDNER, H.R. 1963. Penetration of cotton seedling tap roots as influenced by bulk density, moisture content and strength of soil. Soil Science 96, 153-156.
TAYLOR, H.M. & RATLIFF, L.F. 1969. Root elongation rates of cotton and peanuts as a function of soil strength and water content. Soil Science 108, 113-119.
TOLLNER, E.W. & VERMA, B.P. 1984. Modified cone penetrometer for measuring soil mechanical impe- dance. Transactions of the American Society of Agricultural Engineers 27,33 1-336.
VEEN, B.W. 1982. The influence of mechanical impe- dance on the growth of maize roots. Rant and Soil 66,101-109.
VESIC, A.S. 1972. Expansion of cavities in infinite soil mass. Journal of Soil Mechanics and Foundations Division, American Society of Civil Engineers
VOORHEES, W.B., FARRELL, D.A. & LARSON, W.E. 1975. Soil strength and aeration effects of root
98(SM3), 265-290.
elongation. Soil Science Society of America Pro- ceedings 39,948-953.
WALDRON, L.J. & CONSTANTIN, G.K. 1970. Soil resist- ance to a slowly moving penetrometer. Soil Science
WHITELEY, G.M. & DEXTER, A.R. 1981. The depen- dence ofsoil penetrometer pressure on penetrometer size. Journal of Agricultural Engineering Research
WHITELEY, G.M., UTOMO, W.H. & DEXTER, A.R. 198 I . A comparison of penetrometer pressures and the pressures exerted by roots. Plant and Soil 61, 351-364.
WHITELEY, G.M. &DEXTER, A.R. 1983. Behaviour of roots in cracks between soil peds. Plant and Soil74,
WILSON, A.J., ROBARDS, A.W. & Goss, M.J. 1977. Effects of mechanical impedance on root growth in barley, Hordeum vulgare L. 11. Effects on cell devel- opment in seminal roots. Journal of Experimental Botany 28,1216-1227.
YAPPA, L.G.G., FRITTON, D.D. & WILLATT, S.T. 1988. Effect of soil strength on root growth under different water conditions. Plant and Soil 109,9-16.
109,221-226.
26,467-476.
153-162.





























