Embed Size (px)
Citation preview

Proc. NatL Acad. Sci. USAVol. 79, pp. 670-674, January 1982Neurobiology
Mapping of odor-related neuronal activity in the olfactory bulb byhigh-resolution 2-deoxyglucose autoradiography
(chemical senses/olfactory processing/glomeruli/cellular localization)
DORON LANCET*, CHARLES A. GREER, JOHN S. KAUER, AND GORDON M. SHEPHERD
Section of Neuroanatomy, Yale University School of Medicine, New Haven, Connecticut 06510
Communicated by Louis Sokoloff, September 22, 1981
ABSTRACT The spatial distribution of odor-induced neu-ronal activity in the olfactory bulb, the first relay station of theolfactory pathway, is believed to reflect important aspects of che-mosensory coding. We report here the application of-high-reso-lution 2-deoxyglucose autoradiography to the mapping of spatialpatterns of metabolic activity at the level of single neurons in theolfactory bulb. It was found that glomeruli, which.are synapticcomplexes containing the first synaptic relay, tend to be uniformlyactive or inactive during odor exposure. Differential 2-deoxyglu-cose uptake-was also observed-in the somata of projection neurons(mitral cells) and interneurons (periglomerular and granule cells).This confirms and extends our previous studies in which odor-spe-cific laminar and focal uptake patterns were revealed by the con-ventional x-ray film 2-deoxyglucose method due to Sokoloff andcolleagues [Sokoloff, L., Reivich, M., Kennedy, C., DesRosiers,M. H., Patlak, C. S., Pettigrew, K. D., Sakurada, 0. & Shinohara,M. (1977)J. Neurochem. 28, 897-916]. Based on results obtainedby the two methods, it is suggested that the glomerulus as a wholeserves as a functional unit of activity. The high-resolution resultsare interpreted in terms of the well.characterized synaptic orga-nization of the olfactory bulb and also serve to illustrate the ca-pability of the 2-deoxyglucose autoradiographic technique to mapmetabolic activity in single neurons of the vertebrate central ner-vous system.
An outstanding problem in vertebrate sensory physiology is thenature of processing of olfactory information (1, 2). It is knownthat olfactory receptors and the cells to which they project inthe olfactory bulb have different, although often broad andoverlapping, odor specificities. These characteristics are shown,in response to single odorants, as spatial patterns ofactivity, theelucidation of which could help to understand the processesunderlying olfactory discrimination (cf. ref. 3). Electrophysio-logical analyses of such patterns have been difficult because ofthe necessity to record from a large number of neuronal units.The introduction, by Sokoloff et al. (4), of 2-deoxyglucose au-toradiography, a method that allows one to map activity in wholebrains of awake and behaving animals, has therefore been par-ticularly useful for studies of olfactory mechanisms. 2-Deoxy-glucose competes with glucose-at the uptake stage but does notundergo glycolysis beyond 6-phosphorylation;)'thus accumulat-ing in metabolically active cells. Applying this method to themammalian olfactory bulb, it has been possible to study laminarpatterns of activity in response to odor exposure (5-8). Foci ofhigh uptake in the glomerular layer, where the olfactory re-ceptor axons terminate, exhibit a topographical distribution re-lated to the nature of the odorant and its concentration (6, 8).Due to inherent resolution limits, however, it was difficult toextend these analyses to the cellular level.
We have used a high-resolution 2-deoxyglucose techniquedeveloped by Sejnowski et al (9) to analyze the fine-structuralbasis of odor-induced activity in the vertebrate olfactory bulb.Through correlation with previous 2-deoxyglucose, as well asanatomical and physiological, studies, new information has beenobtained, about mechanisms of olfactory processing.
METHODSSprague-Dawley albino rats, 19-21 days old (Charles RiverBreeding Laboratories, Wilmington, MA or Camm ResearchLaboratories, Wayne, NJ), were maintained with food andwater ad lib. Tiger salamanders (Ambystoma tigrinum) wereobtained from Amphybians of America (Nashville, TN). Threeanimals ofeach species were used for the high-resolution study.Rats were injected intraperitoneally and salamanders were in-jected in the dorsal lymph sac with 2 uCi (0.2 ,uCi for the x-raymethod; 1 Ci = 3.7 X 1010 becquerels) per gram body weightof2-deoxy['4C]glucose in 0. 15 M NaCl (New England Nuclear,30-60 mCi/mmol). Immediately after injection, animals wereexposed to 3-methylbutyl acetate (isoamyl acetate, Fisher) ina flow dilution olfactometer as described (6). Fractional satu-rations ofthe odorant were 10-1 (1.6 mM) for rats and 10-2 (0.16mM) for salamanders. After 60-90 min, animals were decapi-tated and the olfactory bulbs were removed. For the x-ray filmmethod, we proceeded as described (6). Fir high-resolutionautoradiography, the tissue was processed according to Sejnowskiet al. (9). Briefly, tissue was frozen in Freon 12 or 23 (Matheson),freeze substituted in anhydrous acetone (Baker) at -70°C for48-72 hr, and embedded in Araldite (Ciba Geigy)/1% siliconefluid (Dow Corning). Twelve 2- to 3-,m sections were cut forevery 100 um of tissue. Sections were adhered anhydrously tomicroscope slides, dipped in NTB-2 nuclear emulsion (EastmanKodak), exposed for 3-20 weeks at 4°C, and stained with to-luidine blue at pH 11.0.
RESULTSMapping of Activity at High Resolution in the Rat Olfactory
Bulb. Typical patterns of2-deoxyglucose uptake in rats exposedto isoamyl acetate obtained'by the standard x-ray film methodand the high-resolution emulsion-technique are shown in Fig.1 (A and B vs. C and D). Note the correspondence ofthe resultsgenerated by 'the two techniques: both show regions of highuptake in the glomerular and mitral cell body layers, with loweruptake in the nerve, external plexiform, and granule cell layers,'The x-ray film data are fully consistent with the odor-specificuptake patterns reported previously (5, 6), while the high-res-olution data present a considerably more detailed picture oftheodor-elicited metabolic activity.
*'Present address: Department ofMembrane Research, The WeizmannInstitute of Science, Rehovot, Israel.
670
The publication costs ofthis article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertise-ment" in accordance with 18 U. S. C. §1734 solely to indicate this fact.
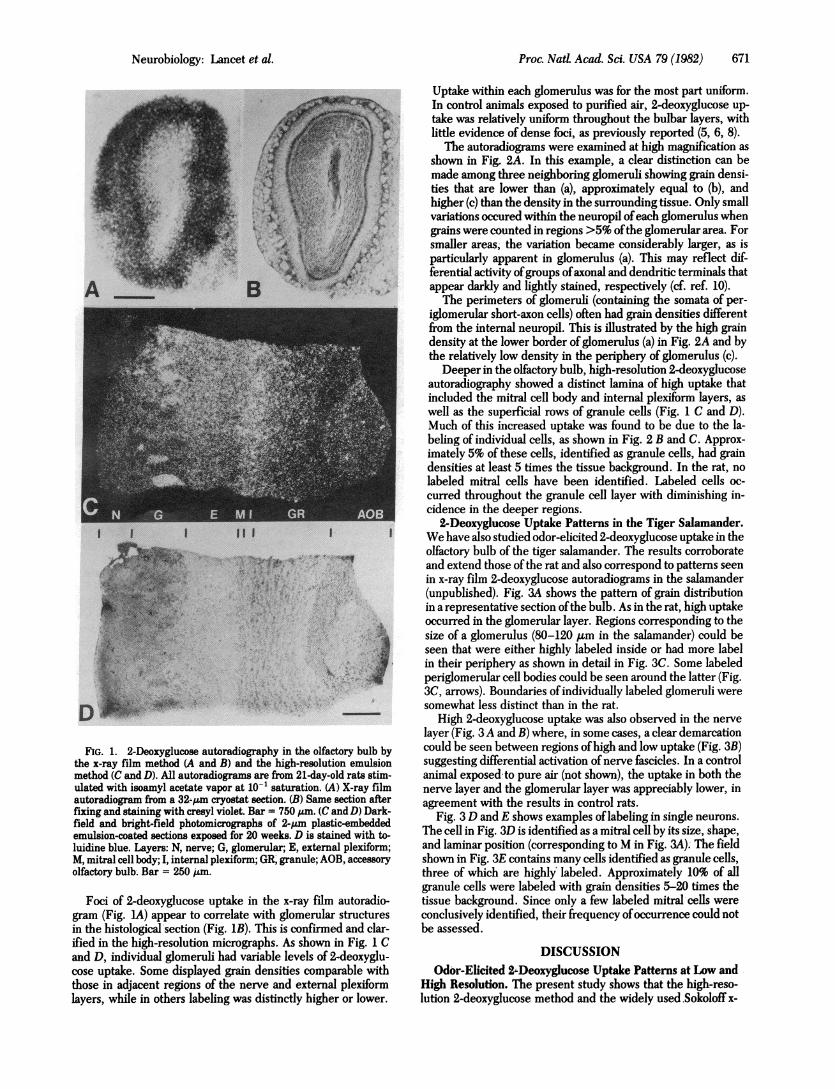
Neurobiology:Lancetetal.~~~~Proc.NatL Acad. Sci. USA 79 (1982) 671
A B
I I 9I
D
FIG. 1. 2-Deoxyglucose autoradiography in the olfactory bulb by
the x-ray film method (A and B) and the high-resolution emulsion
method (C and D). All1 autoradiograms are from 21-day-old rats stim-
ulated with isoamyl. acetate vapor at 10-1 saturation. (A) X-ray film
autoradiogram from a 32-,um cryostat section. (B) Same section after
fixing and staining with cresyl violet. Bar = 750 ILm. (C andD) Dark-
field and bright-field photomicrographs of 2-pam plastic-embeddedemulsion-coated sections exposed -for 20 weeks. D is stained with to-
luidine blue. Layers: N, nerve; G, glomerular; E, external plexiform;
M, mitral cell body; I, internal plexiform; GR granule; AOB, accessory
olfactory bulb. Bar = 250,4m.
Foci of 2-deoxyglucose uptake in the x-ray film autoradio-
gram (Fig. 1A) appear to correlate with glomerular structures
in the histological'section (Fig. iB). This is confirmed and clar-
ified'in the high-resolution micrographs. As shown in Fig. C
and D, individual glomeruli had variable levels of 2-deoxyglu-cose uptake. Some displayed grain densities comparable with
those in adjacent regions of the nerve and external plexiform
layers, while in others labeling was distinctly higher or lower.
Uptake within each glomerulus was for the most part uniform.-In control animals exposed to purified air, 2-deoxyglucose up-take was relatively uniform throughout the bulbar layers, withlittle evidence of dense foci, as previously reported (5, 6, 8).
TFhe autoradiograms were examined at high magnification asshown in Fig. 2A. In this example, a clear distinction can bemade among three neighboring glomeruli showing grain densi-ties that are lower than (a), approximately equal to (b), andhigher (c) than the density in the surrounding tissue. Only smallvariations occured within the neuropil ofeach glomerulus whengrains were counted in regions >5% ofthe glomerular area. Forsmaller areas, the variation became considerably larger, as isparticularly apparent in glomerulus (a). This may reflect dif-ferential activity ofgroups ofaxonal and dendritic terminals thatappear darkly and lightly stained, respectively (cf. ref. 10).The perimeters of glomeruli (containing the somata of per-
iglomerular short-axon cells) often had grain densities differentfrom the internal neuropil. This is illustrated by the high graindensity at the lower border of glomerulus (a) in -Fig. 2A and bythe relatively low density in the periphery of glomerulus (c).
Deeper in the olfactory bulb, high-resolution 2-deoxyglucoseautoradiography showed a distinct lamina of high uptake thatincluded the mitral cell body and internal plexiform layers, aswell as the superficial rows of granule cells (Fig. 1 C and D).Much of this increased uptake was found to be due to the la-beling of individual cells, as shown in Fig. 2 B and C. Approx-imately 5% of these cells, identified as granule cells, had graindensities at least 5 times the tissue background. In the rat, nolabeled mitral cells have been identified. Labeled cells oc-curred throughout the granule cell layer with diminishing in-cidence in the deeper regions.
2-Deoxyglucose Uptake Patterns in the Tiger Salamander.We have also studied odor-elicited 2-deoxyglucose uptake in theolfactory bulb of the tiger salamander. The results corroborateand extend those of the rat and also correspond to patterns seenin x-ray film 2-deoxyglucose autoradiograms in the salamander(unpublished). Fig. 3A shows the pattern of grain distributionin a representative section ofthe bulb. As in the rat, high uptakeoccurred in the glomerular layer. Regions corresponding to thesize of a glomerulus (80-120 A.m in the salamander) could beseen that were either highly labeled inside or had more labelin their periphery as shown in detail in Fig. 3C. Some labeledperiglomerular cell bodies could be seen around the latter (Fig.3C, arrows). Boundaries of individually labeled glomeruli weresomewhat less distinct than in the rat.
High 2-deoxyglucose uptake was also observed in the nervelayer (Fig. 3 A and B) where, in some cases, a clear demarcationcould be seen between regions ofhigh and low uptake (Fig. 3B)suggesting differential activation ofnerve fascicles. In a controlanimal exposed -to pure air (not shown), the uptake in both thenerve layer and the glomerular layer was appreciably lower, inagreement with the results in control rats.
Fig. 3 D and E shows examples oflabeling in single neurons.The cell in Fig. 3D is identified as a mitral cell by its size, shape,and laminar position (corresponding to M in Fig. 3A).- The fieldshown in Fig. 3E contains many cells identified as granule cells,three of which are highly'labeled. Approximately 10% of allgranule cells were labeled with grain densities 5-20 times thetissue background. Since only a few labeled mitral Icells wereconclusively identified, their frequency of'occurrence could notbe assessed.
DISCUSSIONOdor-Eicite 2'.Dexygluose Utake-m~ Pattn.erns at Lws andA
High Resolution. The present study shows that the high-reso-lution 2-deoxyglucose method and the widely used .Sokoloffx-
Neurobiology: Lancet et al.
I I
0j.
i.
.1i in'. -," "., ,.:E
-46. ..t.
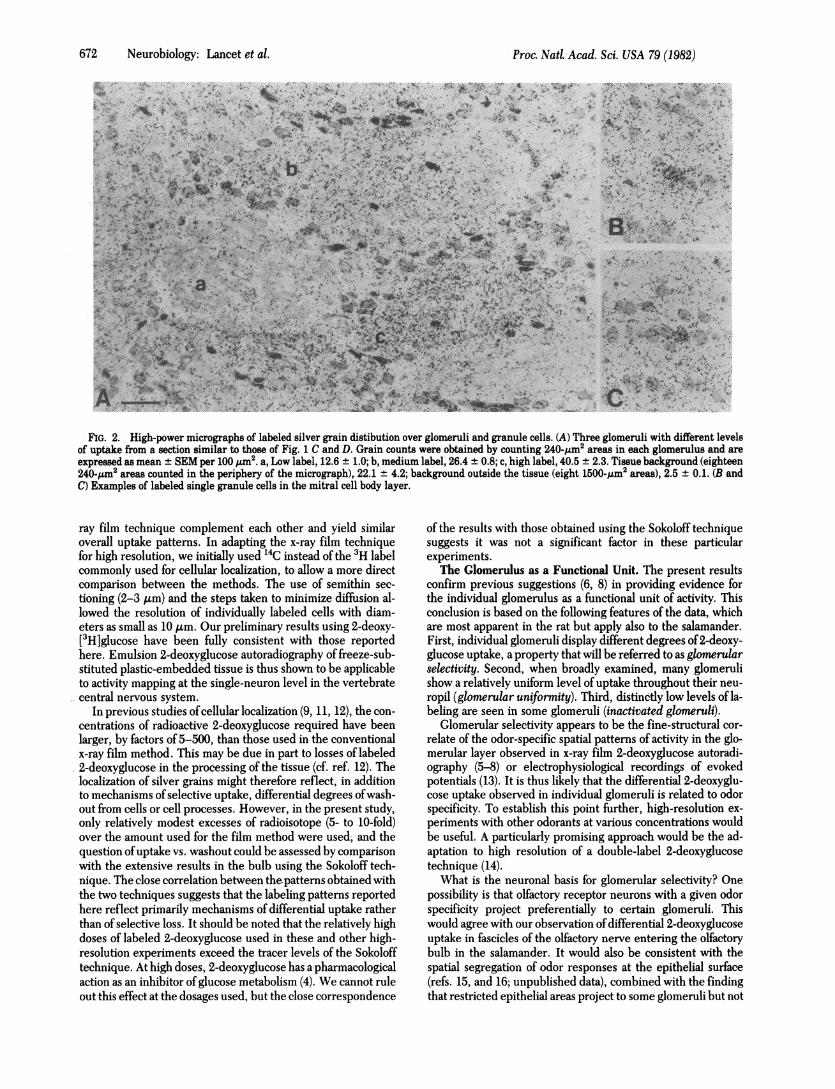
672 Neurobiology: Lancet et al.
A..~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~F
A ,I %f pA..~~~~~~~~~~~~~-
AA
.~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~A
a- - '-~ )m~,.& ¶ I'*~"~Mr(~
W.~~~~~~~~~~~~~~~~~~~~~~..TBy; i:l f of.,,?; . . - .,
,s' e ~-
'- u-,
tt';~-4;.
FIG. 2. High-power micrographs of labeled silver grain distibution over glomeruli and granule cells. (A) Three glomeruli with different levelsof uptake from a section similar to those of Fig. 1 C and D. Grain counts were obtained by counting 240-/Am2 areas in each glomerulus and areexpressed as mean ± SEM per 100 Pjm2. a, Low label, 12.6 ± 1.0; b, medium label, 26.4 ± 0.8; c, high label, 40.5 ± 2.3. Tissue background (eighteen240-,pm2 areas counted in the periphery of the micrograph),. 22.1 ± 4.2; background outside the tissue (eight 1500-,Im2 areas), 2.5 ± 0.1. (B andC) Examples of labeled single granule cells in the mitral cell body layer.
ray film technique complement each other and yield similaroverall uptake patterns. In adapting the x-ray film techniquefor high resolution, we initially used `4C instead of the 3H labelcommonly used for cellular localization, to allow a more directcomparison between the methods. The use of semithin sec-tioning (2-3 ,um) and the steps taken to minimize diffusion al-lowed the resolution of individually labeled cells with diam-eters as small as 10 /im. Our preliminary results using 2-deoxy-[3H]glucose have been fully consistent with those reportedhere. Emulsion 2-deoxyglucose autoradiography of freeze-sub-stituted plastic-embedded tissue is thus shown to be applicableto activity mapping at the single-neuron level in the vertebratecentral nervous system.
In previous studies ofcellular localization (9, 11, 12), the con-centrations of radioactive 2-deoxyglucose required have beenlarger, by factors of 5-500, than those used in the conventionalx-ray film method. This may be due in part to losses of labeled2-deoxyglucose in the processing of the tissue (cf. ref. 12). Thelocalization of silver grains might therefore reflect, in additionto mechanisms of selective uptake, differential degrees ofwash-out from cells or cell processes. However, in the present study,only relatively modest excesses of radioisotope (5- to 10-fold)over the amount used for the film method were used, and thequestion ofuptake vs. washout could be assessed by comparisonwith the extensive results in the bulb using the Sokoloff tech-nique. The close correlation between the patterns obtained withthe two techniques suggests that the labeling patterns reportedhere reflect primarily mechanisms of differential uptake ratherthan of selective loss. It should be noted that the relatively highdoses of labeled 2-deoxyglucose used in these and other high-resolution experiments exceed the tracer levels of the Sokolofftechnique. At high doses, 2-deoxyglucose has a pharmacologicalaction as an inhibitor of glucose metabolism (4). We cannot ruleout this effect at the dosages used, but the close correspondence
of the results with those obtained using the Sokoloff techniquesuggests it was not a significant factor in these particularexperiments.The Glomerulus as a Functional Unit. The present results
confirm previous suggestions (6, 8) in providing evidence forthe individual glomerulus as a functional unit of activity. Thisconclusion is based on the following features of the data, whichare most apparent in the rat but apply also to the salamander.First, individual glomeruli display different degrees of2-deoxy-glucose uptake, a property that will be referred to as glomerularselectivity. Second, when broadly examined, many glomerulishow a relatively uniform level of uptake throughout their neu-ropil (glomerular uniformity). Third, distinctly low levels of la-beling are seen in some glomeruli (inactivated glomeruli).
Glomerular selectivity appears to be the fine-structural cor-relate of the odor-specific spatial patterns of activity in the glo-merular layer observed in x-ray film 2-deoxyglucose autoradi-ography (5-8) or electrophysiological recordings of evokedpotentials (13). It is thus likely that the differential 2-deoxyglu-cose uptake observed in individual glomeruli is related to odorspecificity. To establish this point further, high-resolution ex-periments with other odorants at various concentrations wouldbe useful. A particularly promising approach would be the ad-aptation to high resolution of a double-label 2-deoxyglucosetechnique (14).What is the neuronal basis for glomerular selectivity? One
possibility is that olfactory receptor neurons with a given odorspecificity project preferentially to certain glomeruli. Thiswould agree with our observation ofdifferential 2-deoxyglucoseuptake in fascicles of the olfactory nerve entering the olfactorybulb in the salamander. It would also be consistent with thespatial segregation of odor responses at the epithelial surface(refs. 15, and 16; unpublished data), combined with the findingthat restricted epithelial areas project to some glomeruli but not
Proc. NadAcad. Sci. USA 79 (1982)

Proc. NatL Acad. Sci. USA 79 (1982) 673
M GR
a
H ..i S.0..~~~~~V
A..
X:"9v'-
~2W4:
FIG. 3. High-resolution uptake of 2-deoxyglucose in the olfactorybulb of the tiger salamander exposed to isoamyl acetate at 10-2 sat-uration. (A) Overall view (bright field) with lamina labeled as in Fig.2. Bar = 125 ,um. (B and C) Regions of high and low uptake in the nerveand glomerular layers (C is from the section shown in A; B is from adifferent section). Arrows, labeled periglomerular cells. Bar = 50 ,um.(D andE) Single cell labeling [E is from the section shown inA (marked*), D is from a different section]. The cell inD is identified as a mitralcell, those in E are granule cells. Bar = 25 Am. Scans across such la-beled structures (cf. ref 9) showed a transition from maximum graindensity to tissue background levels over distances of 1-3 pm. Auto-radiographic exposures were 7 (A, C, and E) or 14 (B and D) weeks.
to others (17, 18). In addition to preferential projections, lateralinhibitory mechanisms could contribute to an accentuation ofglomerular selectivity.
Each glomerulus is a highly complex structure containingsynapses of104-105 receptor axons and dendrites of 100 mitraland tufted cells and several hundred periglomerular cells (1, 2).Uniformity of activity within glomeruli was, therefore, an un-
expected finding and would appear to be an important aspectof glomerular function. In view of the fact that individual re-
ceptor axons or even whole olfactory nerve fascicles may projectto only parts of individual glomeruli (if. ref. 18), the observed
glomerular uniformity implies that many receptor axons witha given odor specificity converge onto one glomerulus (cf. ref.16) and terminate throughout its volume. An additional contri-bution to glomerular uniformity may arise through interactionsin the extensive network of synapses between dendrites withinthe glomerulus.
Inactivated glomeruli appear to comprise a special exampleof glomerular uniformity. Their occurrence probably reflectslow levels of activity in receptor axon terminals as well as theeffect of glomerular inhibitory mechanisms. The high 2-deox-yglucose uptake at the perimeter ofsome ofthese glomeruli maybe related to the diminished uptake in their interior. Thus, ac-tivity in immediately adjacent periglomerular cells could exertinhibitory effects via dendrodendritic synapses with mitral celldendrites (intraglomerular inhibition; cf. ref. 1). Some of theactivity at the perimeter not found near cell nuclei may correlatewith inhibitory terminals of periglomerular axons of more re-mote origin, which are known to occur at the glomerular pe-riphery (interglomerular inhibition).The evidence that the glomerulus constitutes a unit related
to odor specificity has implications for the question of how ol-factory acuity is preserved despite continuous turnover of ol-factory receptor cells (19). In view ofthe present results, it couldbe that the target recognized by the newly growing receptoraxons is a whole glomerulus, thereby reducing (by a factor ofup to 100) the amount of information involved in specifying thecorrect axo-dendritic synaptic contacts.
Labeling of Granule and Mitral Cells. Another importantresult of the high-resolution method is the ability to observe 2-deoxyglucose uptake in individual cells. The low proportion oflabeled mitral cells may be related to their low numbers relativeto granule cells (1:5 in the salamander and 1:100 in the rat). Inaddition, a larger percentage of granule cells may be active;mitral cells are subjected to extensive inhibition while most ofthe input onto granule cells is excitatory (1, 2). The labeling ofsome mitral cells, however, agrees with the electrophysiologicaldata showing their selective excitation in response to odor stim-ulation (20).
Differential activity of individual granule cells is a novel ob-servation. Granule cells receive excitatory inputs from manymitral cells (at their reciprocal dendrodendritic synapses) as wellas from centrifugal telencephalic axons. Their activity may thusreflect a process of integration of such multiple effects, whichin turn determines the pattern of inhibition of mitral cells. Thepresent results thus indicate the complex relationships thatmust exist between the processing of olfactory input in the glo-meruli and the interactions that take place between mitral andgranule cells at the level of output from the olfactory bulb tohigher brain centers.
We thank Drs. Terrence J. Sejnowsky and Jan J. Dekker for help andadvice in implementing the high-resolution 2-deoxyglucose method.This research was supported in part by Grant BNS-78-16546 from theNational Science Foundation and by Grants F32-NS06150 and NS-10174 from the National Institute for Neurological and CommunicativeDisorders and Stroke.
1. Shepherd, G. M. (1972) Physiol Rev. 52, 864-917.2. Shepherd, G. M. (1977) in Handbook of Physiology: Nervous
System, ed. Kandel, E. R. (Am. Physiol. Soc., Bethesda, MlD),Sect. 1, Vol. 1, pt. 2, pp. 945-967.
3. Moulton, D. G. (1976) Physiot Rev. 56, 578-593.4. Sokoloff, L., Reivich, M., Kennedy, C., DesRosiers, M. H., Pat-
lak, C. S., Pettigrew, K. D., Sakurada, 0. & Shinohara, M.(1977) J. Neurochem. 28, 897-916.
5. Sharp, F. R., Kauer, J. S. & Shepherd, G. M. (1977)J. Neuro-physioL 40, 800-813.
Neurobiology: Lancet et al.

674 Neurobiology: Lancet et al.
6. Stewart, W. B., Kauer, J. S. & Shepherd, G. M. (1979)J. Comp.Neurot 185, 715-734.
7. Skeen, L. C. (1977) Brain Res. 124, 147-153.8. Jourdan, F., Duveau, A., Astic, L. & Holley, A. (1980) Brain Res.
188, 139-154.9. Sejnowski, T. J., Reingold, S. C., Kelley, D. B. & Gelperin, A.
(1980) Nature (London) 287, 449-451.10. Richardson, K. C., Jarrett, L. & Finke, E. (1960) Stain TechnoL
35, 313-323.11. Basinger, S. F., Gordon, W. C. & Lam, D. M. K. (1979) Nature
(London) 280, 682-684.12. Durham, D., Woolsey, T. A. & Kruger, L. (1981)J. Neurosci. 1,
519-526.13. Leveteau, J. & MacLeod, P. (1966) Science 153, 175-176.14. Livingston; M. S. & Hubel, D. H. (1981) Nature (London) 291,
554-561.
Proc. Nati. Acad. Sci. USA 79 (1982)
15. Kubie, J., Mackay-Sim, A. & Moulton, D. G. (1980) in Proceed-ings of the Seventh International Symposium on Olfaction andTaste, ed. van der Starre, H. (Information Retrieval, London),Vol. 7, pp. 163-166.
16. Kauer, J. S. (1980) in Proceedings of the Seventh InternationalSymposium on Olfaction and Taste, ed. van der Starre, H. (In-formation Retrieval, London), Vol. 7, pp. 227-236.
17. Clark, W. E. LeG. (1957) Proc. R. Soc. London Ser. B 146,299-319.
18. Land, L. J. & Shepherd, G. M. (1974) Brain Res. 70, 506-510.19. Graziadei, P. P. C. & Monti Graziadei, G. A. (1978) in Handbook
of Sensory Physiology, ed. Jacobson, M. (Springer, Berlin), Vol.9, pp. 55-83.
20. Kauer, J. S. (1974) J. Physiol (London) 243, 695-715.




























