Embed Size (px)
Citation preview

IntroductionThe mammary gland is an excellent example of the inductiveprinciple maintained in an adult organ. Tissue composition andthe state of differentiation change drastically according tofunctional requirement and are dependent on mesenchymal-parenchymal interactions, extracellular matrix and localparacrine and endocrine interactive stimuli (Schmeichel et al.,1998). Furthermore, members of at least two gene familiesspecifying developmental pathways during embryonicdevelopment (Wntand Hox) are expressed in a developmentalstage-specific manner in the mammary gland (Gavin andMcMahon, 1992; Friedmann et al., 1994).
The mammary gland consists of two main components, theectodermal parenchyma and the mesodermal stroma. Theparenchyma, which is composed of secretory and ductalepithelial cells, contractile myoepithelial cells and pluripotentstem cells, develops and functions within the stroma, whichconsists of fibroblasts and adipose cells (Smith and Cepko,2001). Unlike other organs, the mammary gland developsmainly in the juvenile and adult organism. With the onset of
ovarian function at puberty, the rudimentary epithelial anlagenare induced to proliferate and to invade the surrounding fattytissue, giving rise to a primitive epithelial ductal treecharacteristic of the virgin gland. During pregnancy, themammary epithelium differentiates and expands drasticallyuntil the entire gland is filled with secretory epitheliumproducing milk to nourish the young. After weaning, themammary epithelium regresses by massive apoptotic cell death(Richert et al., 2000). Although the mediators of the complexinterplay involved in mammary gland development andfunction are not fully characterized, protein tyrosine kinases,either as receptors or intracellular signal transducers, have beenimplicated (Fox and Harris, 1997; Hynes et al., 1997).
The murine EphB4 receptor protein tyrosine kinase wasoriginally isolated from the mature mouse mammary gland,and tightly controlled expression was observed duringmammary gland development and experimental carcinogenesis(Andres et al., 1994). The Eph family of receptor proteintyrosine kinases (RPTKs), which has 14 characterizedmembers, represents the largest family of RPTKs to date
25
We have previously documented the cell-type-specific andhormone-dependent expression of the EphB4 receptor inthe mouse mammary gland. To investigate its role in thebiology of the mammary gland, we have establishedtransgenic mice bearing the EphB4 receptor under thecontrol of the MMTV-LTR promoter, which represents thefirst transgenic mouse model to investigate the effect(s) ofunscheduled expression of EphB4 in adult organisms.Transgene expression in the mammary epithelium wasinduced at puberty, increased during pregnancy,culminated at early lactation and persisted until day threeof post-lactational involution. In contrast, expression ofthe endogenous EphB4 gene is downregulated duringpregnancy, is essentially absent during lactation and is re-induced after day three of post-lactational involution. Theunscheduled expression of EphB4 led to a delayeddevelopment of the mammary epithelium at puberty andduring pregnancy. During pregnancy, less lobules wereformed, these however exhibited more numerous but
smaller alveolar units. Transgenic mammary glands werecharacterized by a fragile, irregular morphology atlactation; however, sufficient functionality was maintainedto nourish the young. Transgenic mammary glandsexhibited untimely epithelial apoptotic cell death duringpregnancy and abnormal epithelial DNA synthesis at earlypost-lactational involution, indicating a disturbed responseto proliferative/apoptotic signals. Mammary tumours werenot observed in the EphB4 transgenic animals; however, indouble transgenic animals expressing both EphB4 andthe neuT genes, tumour appearance was significantlyaccelerated and, in contrast to neuT-only animals,metastases were observed in the lung. These resultsimplicate EphB4 in the regulation of tissue architecture,cellular growth response and establishment of the invasivephenotype in the adult mammary gland.
Key words: Mammary gland, Mammary tumour, Apoptosis, Wholemount, Epithelial proliferation
Summary
Altered mammary epithelial development, patternformation and involution in transgenic miceexpressing the EphB4 receptor tyrosine kinaseNadia Munarini 1,*, Richard Jäger 2,*, Susanne Abderhalden 1, Gisela Zuercher 1, Valeria Rohrbach 1,Saemi Loercher 1, Brigitte Pfanner-Meyer 1, Anne-Catherine Andres 1 and Andrew Ziemiecki 1,‡
Department of Clinical Research, University of Berne, Tiefenaustrasse 120, CH-3004 Bern, Switzerland2Institute for Genetics, Karlsruhe Research Center, D-76021 Karlsruhe, Germany*Both authors contributed equally to this work‡Author for correspondence (e-mail: [email protected])
Accepted 2 October 2001Journal of Cell Science 115, 25-37 (2002) © The Company of Biologists Ltd
Research Article

26
(Pasquale, 1997). The ligands of the Eph family RPTKs, theprotein ephrins, are also membrane associated either by aglycosyl-phosphatidylinositol tail (ephrin A family) or arebona fide transmembrane proteins (ephrin B family) (Pandeyet al., 1995). The cytoplasmic domains of both the receptors,as well as the ligands (ephrin-B family), becomephosphorylated on conserved tyrosine residues followinginteraction, suggesting that signalling cascades can ensue notonly from the receptors but also from the ligands. This mayprovoke bi-directional signalling and mutual cell-cellcommunication (Holland et al., 1996; Brückner et al., 1997).This contention is supported by the demonstrations that bothEph receptors and ephrin ligands interact with PDZ-domain-containing proteins (Hock et al., 1998; Torres et al., 1998;Brückner et al., 1999; Lin et al., 1999), proteins implicated inthe formation of submembranous scaffolds for the assembly ofmacromolecular signalling complexes (Garner et al., 2000).Possible mechanism(s) for modulating both receptor andligand activities exist. Both molecule types associate withphosphatases (Dodelet and Pasquale, 2000) and recently anextracellular metalloprotease, Kuzbanian, has been found toassociate with the ephrin-A2 ligand. This protease is activatedafter ligand activation and cleaves the extracellular moiety ofthe ligand molecule, thereby terminating ligand signalling andthe physical association between cells (Hattori et al., 2000).
The observation that Eph receptors and their ligands exhibitreciprocal expression patterns during embryonic developmenthas led to the suggestion that these molecules play a role in thedevelopment and patterning of a variety of tissues duringembryogenesis. Indeed, Eph family members are involved ingastrulation, cell migration from the neural crest, segmentationof the early embryo and formation of the somites (Holder andKlein, 1999). The best-studied system so far is the developmentof the nervous system, where the Eph family and its ligandshave a pivotal role in axon guidance, fasciculation and, togetherwith NMDA receptors, in synaptogenesis (Klein, 2001).Recently, it has been shown that the Eph family, in particularEphB4 and its ligand ephrin-B2, are intimately involved in thedevelopment of the vascular system during embryogenesis(Wang et al., 1998; Gerety et al., 1999). In contrast toembryonic development, little is known about the function ofthe Eph family in post-natal and adult life, although somemembers, including EphB4, are expressed in adult organs suchas the kidney, lung and mammary gland (Andres et al., 1994).
We have previously investigated the expression of theEphB4 receptor and the ephrin-B2 ligand proteins duringmammary gland development. Expression of both wasparenchyma specific, developmentally regulated and estrogendependent, implicating this receptor-ligand pair in thehormone-dependent morphogenesis of the mammary gland(Nikolova et al., 1998). Expression of the ephrin-B2 ligand wasconfined to the epithelial cells, whereas the EphB4 receptorwas expressed in both the myoepithelial and epithelial cells.Interestingly, the epithelial expression of EphB4 was onlyobserved during proliferative phases of mammary glanddevelopment, such as puberty and the follicular phase of thecycle (Nikolova et al., 1998). We have now establishedtransgenic mice exhibiting overexpression of the EphB4receptor in the mammary epithelium to investigate the effectsof untimely epithelial expression of this receptor on glandulargrowth and differentiation. We demonstrate that unscheduled
expression of EphB4 interferes with the architecture of themammary epithelial tree, alters the response of the epithelialcells to proliferative and apoptotic signals and contributes tothe invasive phenotype of mouse mammary tumours.
Materials and MethodsEstablishment of transgenic miceAn expression cassette comprising the 1.2 kb MMTV-LTR promoterand a 0.3 kb fragment encoding the SV40 late polyadenylation sitewas used for construction of the transgene. This expression cassetteconfers dexamethasone-inducible expression on linked oncogenes incell culture (Jaggi et al., 1986). A 0.2 kb fragment encoding the SV40splice donor/acceptor sequence and a 3.5 kb fragment encompassingthe murine EphB4 cDNA depleted of its own polyadenylation sitewere inserted adjacent to the MMTV-LTR promoter (Andres et al.,1994). Transgenic mice were established commercially by pronuclearinjection using fertilized eggs from a F2 C57Bl6×DBA/2 hybrid cross(Animal Facility of the Centre for Molecular Biology, University ofHeidelberg, Germany). Mouse tail DNA was prepared according toAndres et al. (Andres et al., 1991), and transgenic mice were identifiedby PCR using primers recognizing sequences in the LTR promoterand the EphB4 cDNA. The PCR reactions were performed using thePCR Core Kit according to the manufacturers instructions (RocheDiagnostics, Rotkreuz, Switzerland). Two transgenic lines wereobtained with stable integration and high expression of the transgenein the mammary gland. Animals of these lines were bred tohomozygosity. Homozygosity was ascertained by Southern blotting(Andres et al., 1991) and confirmed by crossing suspectedhomozygous animals repeatedly with control animals and determiningthe pattern of transgene inheritance. In order to establish doubletransgenic mice, EphB4 transgenic animals were first crossed over sixgenerations with inbred C57Bl6 mice (Charles River Wiga, Sulzfeld,Germany) in order to reduce the complexity of the geneticbackground. MMTV-LTR-neuT transgenic mice of the FeBV strain(Muller et al., 1988) were obtained commercially (Charles RiverWiga, Sulzfeld, Germany). Tumour development was followed in theF1 generation of EphB4/C57Bl6 and neuT/FeBV crossings. EphB4negative littermates served as controls in all experiments.
Histological analysesThe fourth inguinal mammary glands were used routinely forhistological examination. For whole-mount staining, the mammaryglands were spread on coated slides and fixed for four hours inCarnoy’s solution (ethanol:chloroform:glacial acid, 6:3:1). Tissueswere washed for 15 minutes in 70% ethanol, rehydrated and stainedovernight in carmine alum (2 g/L carmine, 5 g/L potassium sulfate).The next day, tissues were washed in 70% ethanol, dehydrated,cleared with xylene and mounted in Eukitt. For paraffin embedding,the contralateral fourth inguinal mammary glands were fixed for 24hours in 4% formaldehyde, dehydrated and embedded in paraffin. 4µm sections were either stained with hematoxilin and eosin or weresubjected to immunohistochemical detection of the EphB4 protein asdescribed previously (Nikolova et al., 1998). Sections were examinedusing a Leica DMRD microscope and images were recorded digitallyusing a DC200 camera and the Leica LMS programme (Leica,Glattbrug, Switzerland).
Detection of cell proliferation and apoptosisAnimals were injected with 200 µg per g body weight ofbromodeoxyuridine (BdUr, Sigma, Buchs, Switzerland) in PBS threehours before sacrificing. Incorporation of BdUr into DNA wasdetermined immunohistochemically on formaldehyde fixed sectionsusing anti-BdUr-specific antibodies (Roche Diagnostics) and
Journal of Cell Science 115 (1)

27Mammary development in EphB4 transgenic mice
peroxidase-labelled AB Complex (Dako, Glostrup, Denmark).Apoptotic cell death was analysed by the TUNEL assay onformaldehyde fixed sections using the In Situ Cell Death DetectionKit with TMR-red-labelled dUTP, according to the manufacturersinstructions (Roche Diagnostics, Rotkreuz, Switzerland).
RNA and protein analysesFor RNA analyses, the third mammary gland was snap frozen in liquidnitrogen and stored at –70°C until required. RNA preparation andnorthern blot analyses were done as described in Andres et al. (Andreset al., 1994). Equal loading of the gels was verified by ethidiumbromide staining. For RT-PCR, the RNA was treated with 10 U ofDNase for 60 minutes at room temperature and re-isolated by phenolextraction and ethanol precipitation. RT-PCR was performed using theTitan One Tube RT-PCR Kit according to the manufacturersinstructions (Roche Diagnostics, Rotkreuz, Switzerland). The primersused to detect transgene-derived transcripts were directed tosequences corresponding to the 3′ untranslated region of the EphB4cDNA and the SV40 polyadenylation signal of the transgene constructyielding a transgene-specific fragment of 480 bp. The correctness ofthe PCR product was verified by cloning and sequencing. The absenceof contaminating DNA was controlled by conventional PCR. Forprotein analyses, the snap-frozen contra-lateral third mammary glandswere macerated in SDS-PAGE sample buffer, boiled and subjectedto immunoprecipitation and/or western blot analyses. Protein
concentration was determined visually by amido black staining of 5µl aliquots of extracts spotted onto nitrocellulose and approximatelyequal loading confirmed by Coomassie staining of western blot filters.The EphB4 antibodies (Nikolova et al., 1998) and the clone 4G10phospho-tyrosine antibodies were utilised using conventionalimmunoprecipitation and western blotting methodology (Küng et al.,1997).
ResultsEstablishment and characterization of transgenic linesexpressing EphB4In order to investigate the role of the EphB4 receptor in thebiology of the mammary gland, we have established transgenicmice bearing the murine EphB4 receptor cDNA under thecontrol of the MMTV-LTR promoter. This promoter inducesexpression of linked transgenes in the mammary glandpredominantly during late pregnancy and lactation(Hennighausen, 2000). Two transgenic lines, line 3 and 9, wereobtained and bred to homozygosity. Both lines transmit thetransgene to the progeny in a Mendelian manner typical ofsomatic integration and exhibit high transgene expression inthe lactating mammary glands. In the first instance, weanalysed transgene expression throughout mammary gland
Fig. 1. EphB4transgene expression.(A) Northern blot analysis of transgeneexpression throughout mammary glanddevelopment. 10 µg of total RNA wasprepared from mammary glands of control(C) and transgenic mice (T) at thedevelopmental stages indicated and analysedusing an EphB4-specific probe. (B) RT-PCRanalysis of EphB4transgene and ephrin-B2expression during mammary glanddevelopment. RNA was prepared frommammary glands of control (C) andtransgenic mice (T) at the developmentalstages indicated and amplified usingtransgene and ephrin-B2-specific primers.Std, Molecular weight marker; +co, positivecontrol using transgene DNA as template;–co, negative control without RNA.(C) Western blot analysis of EphB4transgene expression. Protein extracts frommammary glands of control (1),heterozygous (2) and homozygoustransgenic mice (3) at the developmental
stages indicated were analysed using a murine EphB4-specific polyclonal antibody. (D) Tyrosine phosphorylation of the transgenic EphB4protein. Protein extracts of lactating control and transgenic mammary glands were immunoprecipitated (IP) with either EphB4 (Eph) orphosphotyrosine (p-tyr) antibodies, the immunoprecipitates resolved by SDS-PAGE and western blotted with EphB4 antibodies.(E and F) Immunohistochemical localization of the EphB4transgene protein. Sections of lactating mammary glands were reacted withaffinity-purified EphB4 antibodies and visualized with peroxidase-coupled anti-rabbit IgG. Sections were counterstained with hematoxilin.Bars represent 25 µm.

28
development. Northern blot analysis revealed strong transgenicEphB4expression, which was induced at puberty and increasedduring pregnancy (Fig. 1A). Transgene expression culminatedat early lactation and declined after day three of post-lactational involution. In contrast, expression of theendogenous EphB4 gene is downregulatedduring pregnancy, is essentially absentduring lactation and re-appears after daythree of post-lactational involution (Andreset al., 1994). As the transcripts of thetransgene and the endogenous gene wereindistinguishable in size, the transgenicorigin of EphB4 overexpression observedat the RNA level was verified by RT-PCRanalysis taking advantage of the SV40polyadenylation signal sequence specificfor the transgene (Fig. 1B). Thediscrepancy between northern blot and RT-PCR results for transgene expression inmature virgins most probably reflectsdifferential transgene expression duringthe estrous cycle (Andres et al., 1995). RT-PCR analysis of the identical RNAs usingan ephrin-B2 specific primer pair showedno striking difference in endogenousephrin-B2 ligand expression betweentransgenic and control animals (Fig. 1B).As previously observed, ephrin-B2expression was less tightly controlledduring the life cycle of the mammary gland(Nikolova et al., 1998).
Western blot analyses revealing that theEphB4 receptor protein levels in mammaryglands of transgenic animals paralleledthose observed at the RNA level (Fig. 1C).Breeding the mice to homozygosity for theEphB4 transgene resulted in a markedincrease in transgene expression (Fig. 1C).Immunoprecipitation of protein extractsfrom lactating mammary glands witha phospho-tyrosine-specific antibodyfollowed by western blot analysis withanti-EphB4 antibody revealed that aportion of the transgenic EphB4 receptormolecules was phosphorylated at tyrosineresidues (Fig. 1D). Immunohistochemicalstaining confirmed, as expected, theabsence of EphB4 protein in controllactating animals (Fig. 1E). In contrast, inlactating transgenic mammary glands, theEphB4 protein was readily detected andlocalized to the parenchymal cells oftenconcentration at the cell membrane. Theseresults confirm that the transgene protein ismade by the expected cell type and exhibitsthe correct subcellular localization.Interestingly, the immunohistochemicalstaining revealed patchy expression of thetransgene in the mammary tissue: theepithelial cells showed large variations inthe amount of EphB4 protein in the lobular
units and even within the same alveolus (Fig. 1F). In summary,animals homozygous for the EphB4 transgene exhibitconsiderable overexpression of the EphB4 receptor,predominantly during pregnancy, lactation and earlyinvolution. All data presented in this paper were derived
Journal of Cell Science 115 (1)
Fig. 2.Whole-mount staining of mammary glands of control and transgenic mice(transgene) at puberty (5.5 weeks) (A-D), in virgins at 10 weeks (E-H) and 13.5 weeks ofage (I-L). Bars represent 1.5 cm (A,B,E,F,I,J) or 100 µm (C,D,G,H,K,L)
29Mammary development in EphB4 transgenic mice
using homozygote animals, as the phenotypic consequencesobserved in heterozygote animals were very similar but hadlower penetrance.
Transgene expression interferes with the developmentand architecture of the mammary epithelial treeWhole-mount staining of the entire mammary epithelial tree atdifferent stages of development served to assess the effect(s)of unscheduled EphB4expression on the gross architecture ofthe mammary gland. In control animals, at the onset of ovarianfunction at 3.5 weeks of age, the rudimentary mammaryepithelial anlagen are induced to proliferate and invade thefatty tissue by twisting and branching, giving rise to theepithelial tree characteristic of the mature mammary gland(Fig. 2A,C). At 10 weeks of age, the entire fat pad waspopulated by epithelium and the pubertal growth phase wasconcluded, as evidenced by the reshaping of terminal end budsto ductal structures (Fig. 2E,G,I,K). In contrast, the mammaryanlagen of EphB4 transgenic animals exhibited severe growthretardation. At 5.5 weeks of age, the epithelial network wasrudimentary, with reduced side-branching activity and anabsence of second and third order branching (Fig. 2B,D). At10 weeks of age, the epithelial network was expanded and theextent of the epithelialization of the gland and the ductalmorphology were similar to that found in 5.5 week-old controlfemales (Fig. 2F,H). In mature transgenic females at 13.5weeks, the epithelial network was clearly developed; however,epithelial-free areas of adipose tissue and a large number ofgrowing terminal end buds were still visible, indicating that thegrowth phase had not yet been concluded (Fig. 2J,L).
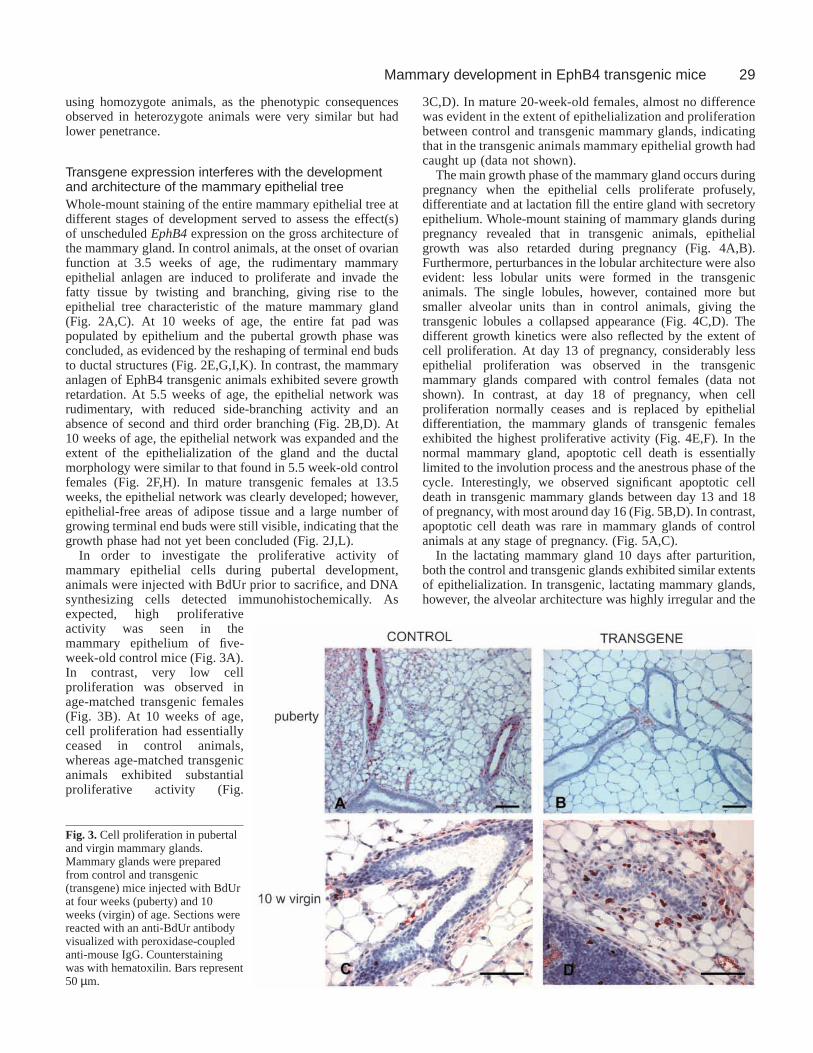
In order to investigate the proliferative activity ofmammary epithelial cells during pubertal development,animals were injected with BdUr prior to sacrifice, and DNAsynthesizing cells detected immunohistochemically. Asexpected, high proliferativeactivity was seen in themammary epithelium of five-week-old control mice (Fig. 3A).In contrast, very low cellproliferation was observed inage-matched transgenic females(Fig. 3B). At 10 weeks of age,cell proliferation had essentiallyceased in control animals,whereas age-matched transgenicanimals exhibited substantialproliferative activity (Fig.
3C,D). In mature 20-week-old females, almost no differencewas evident in the extent of epithelialization and proliferationbetween control and transgenic mammary glands, indicatingthat in the transgenic animals mammary epithelial growth hadcaught up (data not shown).
The main growth phase of the mammary gland occurs duringpregnancy when the epithelial cells proliferate profusely,differentiate and at lactation fill the entire gland with secretoryepithelium. Whole-mount staining of mammary glands duringpregnancy revealed that in transgenic animals, epithelialgrowth was also retarded during pregnancy (Fig. 4A,B).Furthermore, perturbances in the lobular architecture were alsoevident: less lobular units were formed in the transgenicanimals. The single lobules, however, contained more butsmaller alveolar units than in control animals, giving thetransgenic lobules a collapsed appearance (Fig. 4C,D). Thedifferent growth kinetics were also reflected by the extent ofcell proliferation. At day 13 of pregnancy, considerably lessepithelial proliferation was observed in the transgenicmammary glands compared with control females (data notshown). In contrast, at day 18 of pregnancy, when cellproliferation normally ceases and is replaced by epithelialdifferentiation, the mammary glands of transgenic femalesexhibited the highest proliferative activity (Fig. 4E,F). In thenormal mammary gland, apoptotic cell death is essentiallylimited to the involution process and the anestrous phase of thecycle. Interestingly, we observed significant apoptotic celldeath in transgenic mammary glands between day 13 and 18of pregnancy, with most around day 16 (Fig. 5B,D). In contrast,apoptotic cell death was rare in mammary glands of controlanimals at any stage of pregnancy. (Fig. 5A,C).
In the lactating mammary gland 10 days after parturition,both the control and transgenic glands exhibited similar extentsof epithelialization. In transgenic, lactating mammary glands,however, the alveolar architecture was highly irregular and the
Fig. 3.Cell proliferation in pubertaland virgin mammary glands.Mammary glands were preparedfrom control and transgenic(transgene) mice injected with BdUrat four weeks (puberty) and 10weeks (virgin) of age. Sections werereacted with an anti-BdUr antibodyvisualized with peroxidase-coupledanti-mouse IgG. Counterstainingwas with hematoxilin. Bars represent50 µm.

30
epithelial cells appeared to have partly lost cell-cell contactwith their neighbours. Cells characterized by areas of strongbasophilic staining were often observed exfoliating into thelumen. Furthermore, the luminal space was frequently reducedin volume and the secreted material in the alveolar lumen hada much denser appearance than in the lactating mammaryglands of control animals. (Fig. 6A,B). This phenotype was notseen homogenously throughout the gland; areas of near normalmorphology could also be detected. Despite the irregulararchitecture of the mammary glands at lactation, transgenicfemales were able to nourish their young normally, indicatingthat sufficient functionality of the mammary gland wasachieved. Indeed, no striking differences in the amount andquality of milk protein expression could be detected intransgenic and control animals (data not shown).
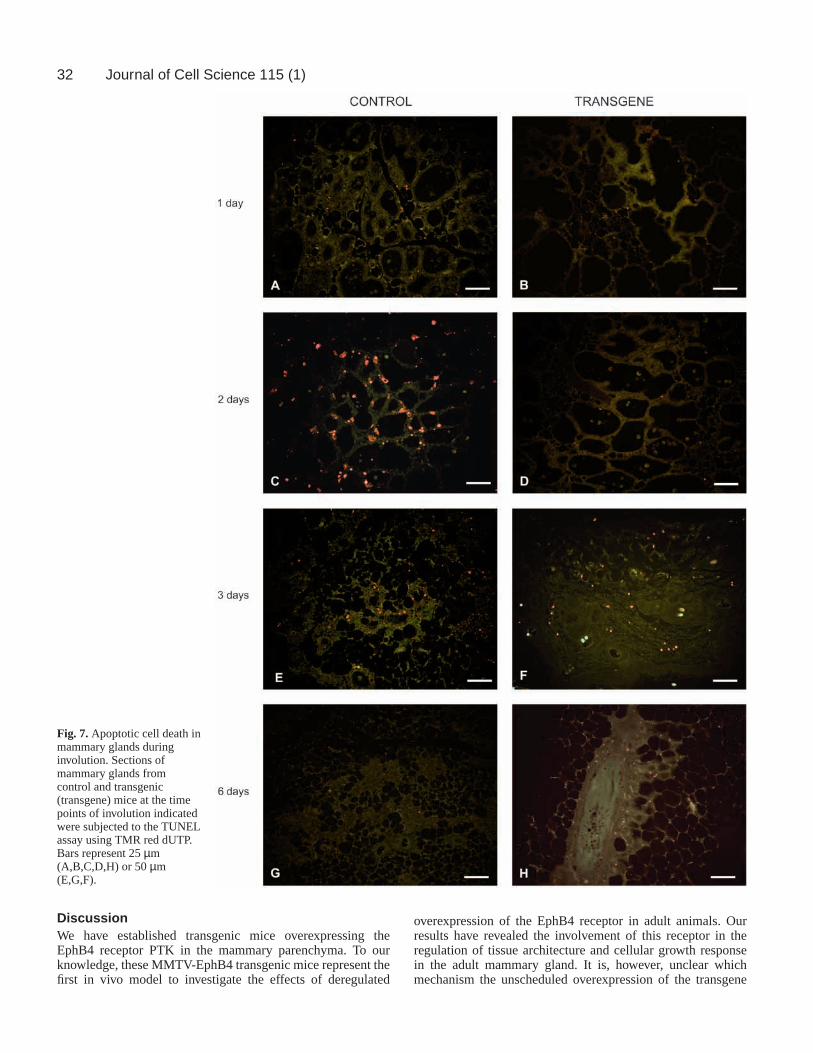
Transgene expression alters thepost-lactational involution processAfter weaning of the young, themammary gland involutes and most ofthe secretory epithelium undergoesapoptotic cell death (Strange et al., 1992).We have compared the involution processin transgenic and control animals in situusing the TUNEL assay. In controlanimals, massive apoptosis was initiatedat day one of involution, culminated atdays two to three and after day six, celldeath was essentially replaced by tissueremodelling (Fig. 7A,C,E,G) (Strange etal., 1992). In contrast, in transgenicanimals, apoptosis was first detected atday three of involution and was stillprominent at day six of involution (Fig.7B,D,F,H).
Interestingly, we frequently detectedBdUr incorporation in nuclei ofparenchymal cells of transgenicmammary glands at day two of weaning(Fig. 8B). This is in contrast to controlanimals, where only occasional BdUrincorporation was seen during earlyinvolution (Fig. 8A). These observationsindicate that in the transgenic animals theinvolution process is delayed and that theepithelial cells respond differently to theapoptotic signals induced by weaning,many replicating their DNA. Clusterin(SGP-2) is a secreted protein originallydescribed as an early marker of cell deathin the prostate and the mammary gland(Montpetit et al., 1986; Strange et al.,1992). Recent observations, however,indicate that clusterin is a cytoprotectiveprotein expressed in cells destined tosurvive apoptosis (Wilson andEasterbrook-Smith, 2000). Northern blotanalyses revealed that transgenic animalsexpressed significantly higher levels ofclusterin RNA during the first two days
of involution and that expression subsided to control levels atday three of involution, paralleling the appearance of TUNELpositive cells in the transgenic animals (Fig. 8C,D).
The delay in the involution process was also evident fromhistological examination of involuting mammary glands.Mammary glands of control animals at day two of involutionwere characterized by dilated alveoli with many dead cellsshed into the lumen (Fig. 9A). Stage-matched transgenicmammary glands also exhibited engorgement of the alveoli;however, dead cells within the lumen were only occasionallyobserved (Fig. 9B). In transgenic animals, histologicalevidence of apoptosis was first observed three days afterweaning (Fig. 9C,D), and after six days, the extent ofinvolution was almost similar to control mice (Fig. 9E,F).Whole-mount staining of regressed mammary glands fiveweeks after the last lactating period revealed that involution
Journal of Cell Science 115 (1)
Fig. 4.Whole-mount staining and cell proliferation of mammary glands during pregnancy.Whole-mount staining (A-D) and BdUr detection (E,F) was performed on control andtransgenic (transgene) mammary glands at day 18 of pregnancy. For BdUr detection, sectionswere reacted with a BdUr antibody visualized with peroxidase-coupled anti-mouse IgG andcounterstained with hematoxilin. Bars represent 200 µm (A,B) or 50 µ (C-F).

31Mammary development in EphB4 transgenic mice
had occurred in both transgenic and control animals. Intransgenic animals, however, the involution process was notas extensive and a larger number of alveolar buds remained inthe tissue (Fig. 9G,H).
Transgene expression favours tumour development andmetastasis formationOverexpression of EphB4 has been described in a subset of humanand mouse mammary tumour cells (Dodelet and Pasquale, 2000).This, and the observation that transgenic animals exhibited lessextensive epithelial regression after weaning, prompted us toinvestigate the influence of EphB4 expression on mammarytumour formation. The results aresummarized in Fig. 10. After anobservation time of up to one year, none ofthe MMTV-EphB4 transgenic femalesdeveloped mammary tumours. In order toinvestigate if EphB4 can influence tumourformation, we have crossed the EphB4transgenic animals with transgenic animalsbearing the neuT oncogene under thecontrol of the MMTV LTR promoter.Females bearing the neuT oncogene in aC57Bl6 genetic background developedmammary gland tumours after a latencytime of 6.5 months. Although tumourswere allowed to grow to a considerablesize, tumour growth was always local andmetastasis formation was never observed(Fig. 10A). Interestingly, the latency time
of mammary tumour appearance in EphB4/neuT doubletransgenic animals was reduced by about 50% to 3.4 months.Moreover, tumour metastasis to the lung was observed in five outof six double transgenic animals (Fig. 10A). Both the primarymammary carcinomas and the lung metastases of double-transgenic animals were histologically very similar to the solidmammary carcinomas developing in the single transgenic NeuTanimals, suggesting that EphB4 expression did not alter thetumour type but its invasive behaviour (Fig. 10B-D). These resultsindicate that, although EphB4 expression itself does not correlatewith an enhanced incidence of tumour development, EphB4 cancooperate with the neuT oncogene and results in earliertumorigenesis and a more aggressive, invasive tumour phenotype.
Fig. 5.Apoptotic cell deathin mammary glands ofcontrol and transgenic(transgene) mice at 16 days(A,B) and 18 days (C,D) ofpregnancy. Sections weresubjected to the TUNELassay using TMR red dUTP.Bars represent 50 µm (A,C)or 25 µm (B,D).
Fig. 6.Histology of lactating mammary glands. Sections of lactating mammary glands fromcontrol and transgenic (transgene) animals at day 10 of lactation were stained with hematoxilinand eosin. Bars represent 25 µm.
32
DiscussionWe have established transgenic mice overexpressing theEphB4 receptor PTK in the mammary parenchyma. To ourknowledge, these MMTV-EphB4 transgenic mice represent thefirst in vivo model to investigate the effects of deregulated
overexpression of the EphB4 receptor in adult animals. Ourresults have revealed the involvement of this receptor in theregulation of tissue architecture and cellular growth responsein the adult mammary gland. It is, however, unclear whichmechanism the unscheduled overexpression of the transgene
Journal of Cell Science 115 (1)
Fig. 7.Apoptotic cell death inmammary glands duringinvolution. Sections ofmammary glands fromcontrol and transgenic(transgene) mice at the timepoints of involution indicatedwere subjected to the TUNELassay using TMR red dUTP.Bars represent 25 µm(A,B,C,D,H) or 50 µm(E,G,F).

33Mammary development in EphB4 transgenic mice
protein uses to exert its effects. We have demonstratedphosphorylation of transgenic EphB4 receptors molecules ontyrosine residues, suggesting that the observed effects may notonly be due to overexpression but also to activation. Theobservation that the greatest effects were seen at developmentalstages when endogenous ephrin-B2 is also expressed suggeststhat transgenic EphB4-ephrinB2 receptor-ligand interactionsmay at least in part be responsible for the observed phenotype.This may be an oversimplification of the situation given theprobable in vivo promiscuity of both Eph receptors and ephrinligands (Dodelet and Pasquale, 2000). However, a ligand-independent mechanism is also conceivable, relying onactivation as a consequence of overexpression. Indeed, Zischet al. (Zisch et al., 1997) have observed Eph receptor activationsolely as a consequence of overexpression.
The EphB4 transgenic mice exhibited developmentallyregulated transgene expression consistent with the propertiesof the MMTV-LTR promoter – the highest expression beingdetected in lactating mammary glands (Hennighausen, 2000;Dickson et al., 2000). The observed patchy expression patternof the transgene appears to be a characteristic of the normallactating mammary gland, as the same phenomenon is also truefor milk proteins. It is thought that this phenomenon may bethe consequence of local tissue regeneration or local adaptationto variable milk consumption by the young (Bchini et al., 1991;Wilde, 1999) and reflects the remarkable plasticity of thisorgan. Alternatively, the patchy EphB4 expression could alsoreflect an attempt of the organ to compensate for possibledetrimental effects by silencing transgene expression (Clark,1998).
Transgenic mammary glands were characterized bydisturbed development of the epithelial tree. Beginning at
puberty, transgenic epithelial ducts exhibited less branchingactivity and developed less alveolar buds. This phenomenonwas even more evident during pregnancy-inducedmorphogenesis of the mammary gland. Although not asextensive, this phenotypic consequence is very reminiscentof the defects observed in the mammary epithelium ofprogesterone receptor (PR) knockout mice (Brisken et al.,1998). These PR knockout animals have confirmed theobservations made in vitro that progesterone is responsible forthe development of ductal side branches during pubertal andpregnancy-induced development. The local regulators of thefrequency of and spacing of the side-branches as well as ofbifurcation of the alveolar buds are still unknown. Thephenotype observed in theEphB4transgenic females suggeststhat this receptor could serve as a negative local controlelement in these processes.
The low branching activity of the epithelial tree in theEphB4 transgenic mice resulted in a smaller number ofindividual lobules during pregnancy. The single lobules of thetransgenic animals, however, contained more, but smaller,alveoli than control animals. Furthermore, the histologicalappearance of the single alveoli at lactation of the transgenicanimals revealed an irregular, fragile morphology, suggestiveof perturbances in cell-cell and cell-matrix interactions. E-andP-cadherin, members of the cell adhesion molecule genefamily, are important factors regulating mammary alveolargrowth and function. Recent data have described a closerelationship between E-cadherin and the Eph receptor familyin terms of expression, localization and activation (Zantek etal., 1999; Orsulic and Kemler, 2000). Moreover, ectopicexpression of EphA4 in early Xenopus embryos has beenshown to disrupt the cadherin-mediated cell adhesion during
Fig. 8.Cell proliferation in mammary glandsat involution. Sections of mammary glandsfrom BdUr-injected control (A) andtransgenic (B; transgene) mice at day two ofinvolution were reacted with an anti-BdUrantibody visualized with peroxidase-coupledanti-mouse IgG. Sections werecounterstained with hematoxilin. Barsrepresent 25 µm. (C) Clusterin expressionduring mammary gland involution. Northernblot analysis of clusterin expression. 10 µgof total RNA prepared from mammaryglands of control (C) and transgenic (T)animals at the developmental stagesindicated were analysed. (D) Western blotanalysis of EphB4 expression. Proteinextracts prepared from mammary glands ofcontrol (C) and transgenic (T) animals atdifferent time points of involution (invol.)

34
gastrulation (Winning et al., 1996; Jones et al., 1998). Inaddition to the cross-talk with the cadherin cell adhesionmolecules, the Eph family has also been shown to mediate cellattachment or detachment by regulating integrin function(Huynh-Do et al., 1999; Davy and Parker, 2000). Thus, it is
likely that the observed phenotypic consequences of EphB4overexpression on the mammary epithelial morphology may,at least in part, be due to a disturbed cell-cell and cell-matrixattachment.
The most striking effect observed in the EphB4 transgenic
Journal of Cell Science 115 (1)
Fig. 9.Histology ofinvoluting mammaryglands. (A-F) Sections ofmammary glands of controland transgenic (transgene)animals taken at theindicated time points afterweaning were stained withhematoxilin and eosin. Barsrepresent 50 µm.(G,H) Whole-mountstaining of regressedmammary glands fromcontrol (G) and transgenic(H; transgene) animalstaken 4.5 weeks afterweaning. Bars represent200 µm.

35Mammary development in EphB4 transgenic mice
mice concerned the altered proliferative and apoptotic responseof the mammary epithelial cells. The analysis of BdUrincorporation revealed that a considerable portion of thetransgenic mammary epithelial cells undergo DNA synthesisat day two of involution, a time point at which in control glandsapoptosis is initiated and little if any BdUr incorporation wasseen. The subsequent, approximately one day delayed,induction of epithelial regression in transgenic glands makes itunlikely that the BdUr-positive cells finished the cell cycle andunderwent mitosis. Conceivably, these cells are arrested at theG2/M checkpoint of the cell cycle and subsequently undergoapoptosis. Normally, cells predetermined to proliferate orundergo cell death enter the cell cycle, traverse early G1 phase,whereupon their exact fate is fulfilled (King and Cidlowski,1998). Recent evidence suggests that, apart from beinginvolved in the control of cell migration, the Eph family is alsoinvolved in the regulation of cell death and survival. EphA4 istransiently expressed in motoneurons, which are predestined toundergo cell death during the development of the spinal cord(Ohta et al., 1996). Moreover, several members of the EphAfamily are responsible for inhibiting cell migration andinduction of cell death in ventral spinal cord neurons (Yue etal., 1999). In contrast, overexpression of the ectodomain of theEphB2 receptor in the subventricular zone of the adult braininhibited cell migration but stimulated proliferation ofneuroblasts (Conover et al., 2000).
Our observation of DNA synthesis in the involutingmammary epithelium may reflect an altered response of EphB4overexpressing cells to death/survival signals in early G1.Furthermore, the high proportion of apoptotic cell deathobserved in transgenic animals during the proliferative phaseat pregnancy may similarly reflect an altered cellular responseto proliferative signals, possibly correlated to a disturbedfunctioning of the E-cadherins. In mammary epithelial cells,E-cadherin can regulate cell survival by activating theretinoblastoma (Rb) gene, and via Rb it can initiate a growthsignal conflict in an epithelial cell population induced to
undergo apoptosis (Day et al., 1999). It is interesting that inthe same time frame that DNA synthesis was observed in theabsence of apoptosis, clusterin, a gene involved in protectingcells from apoptosis by unknown mechanisms (Wilson andEasterbrook, 2000), was highly induced in the transgenicmammary glands. This suggests that epithelial survival is, atleast transiently, favoured in the transgenic mammary glandspredestined for involution.
Although EphB4 overexpression apparently interferes withthe growth response of the mammary epithelial cells duringinvolution, tumour formation was not observed in the EphB4transgenic females. In contrast, in neuT/EphB4 doubletransgenic females, tumour formation was accelerated andtumour growth was more aggressive than in the singletransgenic neuT animals. Similar observations have been madein transgenic animals overexpressing the bcl-2 gene in themammary epithelium where a delay in post-lactationalinvolution was observed. Similar to the results reported here,bcl-2 overexpression alone did not lead to tumorigenesis;instead, it favoured tumour formation in c-myc/bcl2 doubletransgenic mice (Jäger et al., 1997). Interestingly, we haveobserved a considerable increase in the tumour latency time inthe single transgenic neuT females crossed into the C57Bl6genetic background over that in NeuTtransgenic females of thepure FeBV strain. This is in agreement with the observationsmade with other oncogene-bearing mice or with spontaneoustumour formation that the high tumour resistance of theC57Bl6 strain is genetically determined (Macleod and Jacks,1999). In the EphB4/neuT transgenic females, tumours notonly appeared with reduced latency but also metastasised to thelung. Several reports have positively correlated overexpressionof Eph family members with carcinogenesis, although directevidence for the transforming potential is still missing (Dodeletand Pasquale, 2000). Our results indicate that EphB4overexpression by itself is not tumorigenic but instead favoursan invasive phenotype on transformed cells. This notion isfurther supported by our previous observation that in
Fig. 10. Mammary tumour formation in NeuTand NeuT/EphB4double transgenic mice (A) Table summarizing tumour frequency, latency andmetastasis formation. (B-D) Histology of primary mammary tumours from NeuTtransgenic mice (B), of primary mammary tumours (C) andlung metatstasis (D) from EphB4/NeuTdouble transgenic mice. Sections were stained with hematoxilin and eosin. Bars represent 50 µm.

36
experimental mouse mammary tumours induced by the Ha-rasoncogene only the anaplastic, invasive tumour cells expressedhigh levels of EphB4, whereas the majority of the tumour masswas EphB4 negative (Nikolova et al., 1998). Thus, disturbancein the expression of molecules involved in the control of patternformation may not directly induce transformation but may beinstrumental in the acquisition of the malignant, invasivephenotype, possibly by modulating integrin and cadherinfunctions. It remains to be elucidated if epithelialoverexpression of EphB4 can also provoke increasedangiogenic potential of mammary cells and thereby facilitatetumour invasiveness.
The authors wish to thank R. R. Friis and R. Strange for stimulatingdiscussions and Barbara Krieger for photographic artwork. This workhas been supported by the Swiss National Science Foundation (31-54315.98), the Novartis Research Foundation, the Foundation forCinical-Experimental Cancer Research, the Swiss Life InsuranceFoundation, the Stiftung zur Krebsbekämpfung, and the ‘Stiftung zurFörderung der Wissenschaftlichen Forschung an der UniversitätBern’. R.J. was supported by the Bernese Cancer League.
ReferencesAndres, A.-C., Bchini, O., Schubaur, B., Dolder, B., LeMeur, M. and
Gerlinger, P. (1991). Ha-ras induced transformation of mammaryepithelium is favoured by increased oncogene expression or inhibition ofmammary regression. Oncogene6, 771-779.
Andres, A.-C., Reid, H. H., Zuercher, G., Blaschke, R. J., Albrecht, D. andZiemiecki, A. (1994). Expression of two novel eph-related receptor tyrosinekinases in mammary gland development and carcinogenesis. Oncogene9,1461-1467.
Andres, A.-C., Zuercher, G., Djonov, V., Flueck, M. and Ziemiecki, A.(1995). Protein tyrosine kinase expression during the estrous cycle andcarcinogenesis of the mammary gland. Int. J. Cancer63, 288-269.
Bchini, O., Andres, A.-C., Schubaur, B., Mehtali, M., LeMeur, M., Lathe,R. and Gerlinger, P. (1991). Precocious mammary gland development andmilk protein synthesis in transgenic mice ubiquitously expressing humangrowth hormone. Endocrinology128, 539-546
Brisken, C., Park, S., Vass, T., J. P., L., O’Malley, B. W. and Weinberg, R.A. (1998). A paracrine role for the epithelial progesterone receptor inmammary development. Proc. Natl. Acad. Sci. USA95, 5076-5081.
Brueckner, K., Pasquale, E. B. and Klein, R. (1997). Tyrosinephosphorlylation of transmembrane ligands for EphB4 receptors. Science275, 1640-1643.
Brueckner, K., Labrador, J. P., Scheiffele, P., Herb, A., Seeburg, P. H. andKlein, R. (1999). EphrinB ligands recruit GRIP family PDZ adaptorproteins into raft membrane microdomains. Neuron22, 511-524.
Clark, A. J. (1998). Gene expression in the mammary glands of transgenicanimals. Biochem. Soc. Symp. 63, 133-140.
Conover, J. C., Doetsch, F., Garcia-Verdugo, J. M., Gale, N. W.,Yancopoulos, G. D. and Alvarez-Buylla, A.(2000). Disruption of Eph-ephrin signalling affects migration and proliferation in the adultsubventricular zone. Nat. Neurosci. 3, 1091-1097.
Davy, A. and Parker, S. M.(2000). Ephrin-A5 modulates cell adhesion andmorphology in an integrin-dependent manner. EMBO J. 19, 5396-5404.
Day, M. L., Zhao, X., Vallorosi, C. J., Putzi, M., Powell, C. T., Lin, C. andDay, K. C. (1999). E-caherin mediates aggregation-dependent survival ofprostate and mammary epithelial cells through the retinoblastoma cell cyclecontrol pathway. J. Biol. Chem. 274, 9656-9664.
Dickson, C., Creer, A. and Fantl. V.(2000). Mammary gland oncogenes asindicators of pathways important in mammary gland development.Oncogene19, 1097-1101.
Dodelet, V. C. and Pasquale, E. B.(2000). Eph receptors and ephrin ligands:embryogenesis to tumorigenesis. Oncogene19, 5614-5619.
Fox, S. B. and Harris, A. L.(1997). The epidermal growth factor receptor inbreast cancer. J. Mammary Gland Biol. Neopl. 2, 131-142.
Friedmann, Y., Daniel, C. A., Strickland, P. and Daniel, C. W.(1994). Hoxgenes in normal and neoplastic mouse mammary gland. Cancer Res. 54,5981-5985.
Garner, C. C., Nash, J. and Hughanir, R. L. (2000). PDZ domains in synapseassembly and signalling. Trends Cell Biol. 10, 274-280
Gavin, B. J. and McMahon, A. P.(1992). Differential regulation of the Wntgene family during pregnancy and lactation suggests a role in postnataldevelopment of the mammary gland. Mol. Cell Biol. 12, 2418-2423.
Gerety, S. S., Wang, H. U. and Anderson, D. J.(1999). Symmetrical mutantphenotypes of the receptor EphB4 and its specific transmembrane ligandephrin-B2 in cardiovascular development. Mol. Cell 4, 403-414.
Hattori, M., Osterfield, M. and Flanagan, J. G.(2000). Regulated cleavageof a contact-mediated axon repellent. Science289, 1360-1365.
Hennighausen, L.(2000). Mouse models for breast cancer. Oncogene19, 966-967.
Hock, B., Boehme, B., Karn, T., Yamamoto, T., Kaibuchi, K., Holtrich, U.,Holland, S., Pawson, T., Ruebsamen-Waigmann, H. and Streibhardt, K.(1998). PDZ-domain mediated interaction of the Eph-related receptortyrosine kinase EphB3 and the ras-binding protein AF6 depends on thekinase activity of the receptor. Proc. Natl. Acad. Sci. USA95, 9779-9784.
Holder, N. and Klein, R. (1999). Eph receptors and ephrins: effectors ofmorphogenesis. Development126, 2033-2044.
Holland, S. J., Gale, N. W., Mbamalu, G., Yancopoulos, G. D., Henkemeyer,M. and Pawson, T. (1996). Bi-directional signalling through the Eph familyreceptor Nuk and its transmembrane ligands. Nature383, 722-725.
Huynh-Do, U., Stein, E., Lane, A. A., Liu, H., Cerretti, D. P. and Daniel,T. O. (1999). Surface densities of ephrin-B1 determine EphB1-coupledactivation of cell attachment through alphavbeta3 and alpha5beta1 integrins.EMBO J. 18, 2165-2173.
Hynes, N. E., Cella, N. and Wartmann, M.(1997) Prolactin mediatedsignalling in mammary epithelial cells. J. Mammary Gland Biol. Neopl. 2,19-28.
Jäger, R., Hetzer, U., Schenkel, J. and Weiher, H.(1997). Over expressionof bcl-2 inhibits alveolar cell apoptosis during involution and accelerates c-myc induced tumorigenesis of the mammary gland. Oncogene15, 1787-1795.
Jaggi, R., Salmons, B., Muellener, D. and Groner, B.(1986). The v-mos andHa-ras oncogene expression represses glucocorticoid hormone-dependenttrasnscription from the mouse mammary tumor virus LTR. EMBO J. 5,2609-2616.
Jones, T. L., Chong, L. D., Kim, J., Xu, R. H., Kung, H. F. and Daar, I. O.(1998). Loss of cell adhesion in Xenopus laevis embryos mediated by thecytoplasmic domain of Xlerk, an erythropoietin-producing hepatocellularligand. Proc. Natl. Acad. Sci. USA95, 576-581.
King, K. L. and Cidlowski, J. A. (1998). Cell cycle regulation and apoptosis.Annu. Rev. Physiol. 60, 601-617.
Klein, R. (2001). Exitatory Eph receptors and adhesive ephrin ligands. Curr.Opin. Cell Biol. 13, 196-203.
Kueng, P., Nikolova, Z., Djonov, V., Hemphill, A., Rohrbach, V., Boehlen,D., Zuercher, G., Andres, A.-C. and Ziemiecki, A.(1997). A novel familyof serine/threonine kinases participating in spermiogenesis. J. Cell Biol. 139,1851-1859.
Lin, D., Gish, G. D., Songyang, Z. and Pawson, T.(1999). The carboxylterminus of class B ephrins constitutes a PDZ domain binding motif. J. Biol.Chem. 272, 3726-3733.
Macleod, K. F. and Jacks, T.(1999). Insights into cancer from transgenicmouse models. J. Pathol. 187, 43-60.
Monpetit, M. L., Lawless, K. R. and Tenniswood, M. (1986). Androgen-repressed messages in the rat ventral prostrate. Prostate8, 25-36.
Muller, W. J., Sinn, E., Pattengale, P. K., Wallace, R. and Leder, P. (1998).Single-step induction of mammary adenocarcinoma in transgenic micebearing the activated neu oncogene. Cell 54, 105-115.
Nikolova, Z., Djonov, V., Zuercher, G., Andres, A.-C. and Ziemiecki, A.(1998). Cell-type specific and estrogen dependent expression of the receptortyrosine kinase EphB4 and its ligand ephrin-B2 during mammary glandmorphogenesis. J. Cell Sci. 111, 2741-2751.
Ohta, K., Nakamura, M., Hirokawa, K., Tanaka, S., Iwama, A., Suda, T.,Ando, M. and Tanaka, H. (1996). The receptor tyrosine kinase, Cek8, istransiently expressed on subtypes of motoneuron in the spinal cord duringdevelopment. Mech. Dev. 54, 59-69.
Orsulic, S. and Kemler, R.(2000). Expression of Eph receptors and ephrinsis differentially regulated by E-cadherin. J. Cell Sci. 113, 1793-1802.
Pandey, A., Lindberg, R. A. and Dixit, V. M. (1995). Cell signalling.Receptor orphans find a family. Curr. Biol. 5, 986-989.
Pasquale, E. B.(1997). The Eph family of receptors. Curr. Opin. Cell Biol.9, 608-615.
Richert, M. M., Schwertfeger, K. L., Ryder, J. W. and Anderson, S. M.
Journal of Cell Science 115 (1)

37Mammary development in EphB4 transgenic mice
(2000). An atlas of mouse mammary gland development. (2000). J. Mamm.Gland Biol. Neopl. 5, 227-242.
Schmeichel, K. L., Weaver, V. M. and Bissell, M.(1998). Structural cuesfrom tissue microenvironment are essential determinants of the humanmammary epithelial phenotype. J. Mammary Gland Biol. Neopl. 3, 201-214.
Smith, G. H. and Cepko, G.(2001). Mammary epithelial stem cells. Micr.Res. Tech. 52, 190-203.
Strange, R., Li, F., Saurer, S., Burkhardt, A. and Friis, R. R.(1992).Apoptotic cell death and tissue remodelling during mouse mammary glandinvolution. Development115, 49-58.
Torres, R., Firestein, B. L., Dong, H. L., Staudinger, Y., Olson, E. N.,Hughanir, R. L., Bredt, D. S., Gale, N. W. and Yancopoulos, G. D.(1998). PDZ proteins bind, cluster, and synaptically colocalize with Ephreceptors and their ephrin ligands. Neuron21, 1453-1463.
Wang, H. U., Chen, Z. F. and Anderson, D. J.(1998). Molecular distinctionand angiogenic interaction between embryonic arteries and veins revealedby ephrinB2 and its receptor EphB4. Cell 93, 741-753.
Wilde, C. J., Knight, C. H. and Flint, D. J. (1999). Control of milk secretionand apoptosis during mammary involution. J. Mamm. Gland Biol. Neoplasia4, 129-136.
Wilson, M. R. and Easterbrook-Smith, S. B.(2000). Clusterin is a secretedmammalian chaperone. Trends Biochem. Sci. 25, 95-98.
Winning, R. S., Scales, J. B. and Sargent, T. D.(1996). Disruption of celladhesion in Xenopus by Pagliaccio, an Eph-class receptor tyrosine kinase.Dev. Biol. 179, 309-319.
Yue, Y., Su, J., Cerretti, D. P., Fox, G. M., Jing, S. and Zhou, R.(1999).Selective inhibition of spinal cord neurite outgrowth and cell survival by theEph family ligand ephrin-A5. J. Neurosci.15, 10026-10035.
Zantek, N. D., Azimi, M., Fedor-Chaiken, M., Wang, B., Brackenbury, R.and Kinch, M. S. (1999). E-cadherin regulates the function of EphA2receptor tyrosine kinase. Cell Growth Differ. 10, 629-638.
Zisch, A. H., Stallcup, W. B., Chong, L. D., Dahlin-Huppe, K., Voshol, J.,Schachner, M. and Pasquale, E. B.(1997). Tyrosine phosphorylation ofL1 family adhesion molecules: implication of the Eph kinase Cek5. J.Neurosci. Res. 47, 655-665.





























