Embed Size (px)
Citation preview

Making a vertebrate limb: newplayers enter from the wingsGail Martin
SummaryWhat initiates vertebrate limb development and induceslimbs to form where they do? For several years theanswer to this intriguing question has been framed interms of a working model that limb induction depends ona dialogue between two members of the FibroblastGrowth Factor (FGF) family of intercellular signalingmolecules, FGF8 and FGF10. Now, a recent paper haswritten roles for signals encoded by WNT genes, thevertebrate relatives of the Drosophila wingless gene, intothe script.(1) BioEssays 23:865±868, 2001.ß 2001 John Wiley & Sons, Inc.
Observing limb developmentÐin front of the
curtain and behind the scenes
Much of what we know about vertebrate limb development has
been learned by watching the chicken embryo develop in ovo
through a window in the egg's shell. The first sign that limbs are
developing is the appearance of a small protrusion on each
side of the body, near the anterior end of the torso. Several
hours after these forelimb buds become visible, the hindlimb
buds begin to protrude near the posterior end. Each limb bud is
composed of lateral plate mesoderm (LPM) and the overlying
surface ectoderm. The mesoderm contains the progenitors of
skeletal elements, tendons, and other connective tissue of the
mature limb. The surface ectoderm gives rise to skin and
cutaneous appendages such as feathers on chick wings or
hairs on mouse legs.
A few hours after the limb buds first become visible,
changes in cell shape and position within the surface ectoderm
result in the appearance of a ridge that runs along the distal
margin of each limb bud. Once this apical ectodermal
ridge (AER) has formed, the limb bud elongates along its
proximodistal axis (e.g., shoulder to fingers), flattens along
its dorsoventral axis (e.g., back of hand to palm), and becomes
asymmetric along its anteroposterior axis (e.g., thumb to
little finger). AER formation is a critical event, because the
ridge produces signals that are essential for limb development.
If the AER fails to form, the nascent limb bud regresses and no
limb develops.(2) If the AER is removed, the limb that develops
is truncated, i.e., it contains proximal skeletal elements but
lacks distal ones.(3)
To gain insight into what causes limb buds to develop at
specific positions along the body axis, tissue grafting and other
experimental manipulations have been performed in ovo.(4) A
key finding was that the LPM along most of the length of the
torso is competent to respond to a limb-inducing signal. This
implies that under normal circumstances, the induction of limb
development at specific positions involves some mechanism
for restricting the availability of the limb inducer. A second
important finding was that the signal(s) for limb induction is
produced by tissue medial to the LPM (Fig. 1). But what is the
limb-inducing signal? Studies by Cohn et al.(5) turned the
spotlight on FGFs.
Outlining the scriptÐFGFs have a starring role
FGFs are secreted proteins that signal by binding to high-
affinity tyrosine kinase receptors on the cell surface (FGF
receptors; FGFRs), thereby activating intracellular signaling
pathways, including the RAS-MAP kinase pathway.(6,7) FGFs
are known to play major roles in multiple aspects of embryonic
development,(8) including outgrowth of the limb bud.(4) But
their function as signals that initiate limb development was not
appreciated until a bead soaked in FGF protein was inserted
into the prospective interlimb region.(5) This induced cells that
would not normally participate in limb development to form an
ectopic limb (Fig. 1A). The type of limb that forms (wing or leg)
depends on whether the FGF source was placed closer to the
prospective forelimb or hindlimb territory, respectively.
These results suggested that the endogenous limb inducer
might be an FGF. However, several different members of the
FGF family (of which there are now 22 in mouse and human
Ref. 9) proved capable of inducing ectopic limbs. Studies of
FGF gene expression patterns at early stages of normal and
ectopic chick limb development identified FGF8 and FGF10 as
potential regulators of limb development.(10±12) The model
that was proposed to explain their function (Fig. 1B) posits that
FGF10 produced in the LPM is the signal that initiates limb
development. At early stages of embryogenesis, however,
Fgf10 is expressed throughout the LPM, so the key to inducing
limbs at specific locations is to restrict Fgf10 expression to the
appropriate territories at the stage when limb induction occurs.
FGF8 produced in medial tissues (intermediate mesoderm for
the forelimb and primitive streak for the hindlimb) was
proposed to be the signal that maintains Fgf10 expression in
BioEssays 23:865±868, ß 2001 John Wiley & Sons, Inc. BioEssays 23.10 865
Department of Anatomy, Box 0452, University of California, Rm.
S-1457, 513 Parnassus Ave, San Francisco, CA 94143-0452.
E-mail: [email protected]
What the papers say
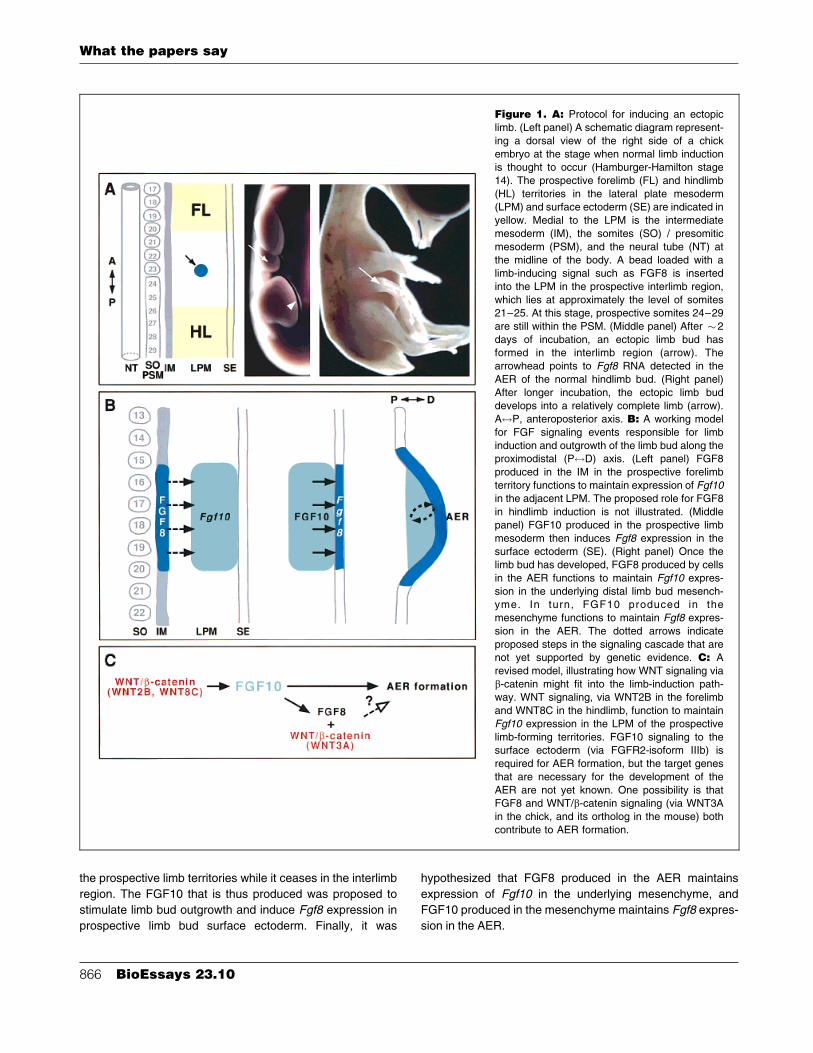
the prospective limb territories while it ceases in the interlimb
region. The FGF10 that is thus produced was proposed to
stimulate limb bud outgrowth and induce Fgf8 expression in
prospective limb bud surface ectoderm. Finally, it was
hypothesized that FGF8 produced in the AER maintains
expression of Fgf10 in the underlying mesenchyme, and
FGF10 produced in the mesenchyme maintains Fgf8 expres-
sion in the AER.
Figure 1. A: Protocol for inducing an ectopic
limb. (Left panel) A schematic diagram represent-
ing a dorsal view of the right side of a chick
embryo at the stage when normal limb inductionis thought to occur (Hamburger-Hamilton stage
14). The prospective forelimb (FL) and hindlimb
(HL) territories in the lateral plate mesoderm(LPM) and surface ectoderm (SE) are indicated in
yellow. Medial to the LPM is the intermediate
mesoderm (IM), the somites (SO) / presomitic
mesoderm (PSM), and the neural tube (NT) atthe midline of the body. A bead loaded with a
limb-inducing signal such as FGF8 is inserted
into the LPM in the prospective interlimb region,
which lies at approximately the level of somites21±25. At this stage, prospective somites 24±29
are still within the PSM. (Middle panel) After �2
days of incubation, an ectopic limb bud hasformed in the interlimb region (arrow). The
arrowhead points to Fgf8 RNA detected in the
AER of the normal hindlimb bud. (Right panel)
After longer incubation, the ectopic limb buddevelops into a relatively complete limb (arrow).
A$P, anteroposterior axis. B: A working model
for FGF signaling events responsible for limb
induction and outgrowth of the limb bud along theproximodistal (P$D) axis. (Left panel) FGF8
produced in the IM in the prospective forelimb
territory functions to maintain expression of Fgf10in the adjacent LPM. The proposed role for FGF8
in hindlimb induction is not illustrated. (Middle
panel) FGF10 produced in the prospective limb
mesoderm then induces Fgf8 expression in thesurface ectoderm (SE). (Right panel) Once the
limb bud has developed, FGF8 produced by cells
in the AER functions to maintain Fgf10 expres-
sion in the underlying distal limb bud mesench-yme. In turn, FGF10 produced in the
mesenchyme functions to maintain Fgf8 expres-
sion in the AER. The dotted arrows indicate
proposed steps in the signaling cascade that arenot yet supported by genetic evidence. C: A
revised model, illustrating how WNT signaling via
b-catenin might fit into the limb-induction path-way. WNT signaling, via WNT2B in the forelimb
and WNT8C in the hindlimb, function to maintain
Fgf10 expression in the LPM of the prospective
limb-forming territories. FGF10 signaling to thesurface ectoderm (via FGFR2-isoform IIIb) is
required for AER formation, but the target genes
that are necessary for the development of the
AER are not yet known. One possibility is thatFGF8 and WNT/b-catenin signaling (via WNT3A
in the chick, and its ortholog in the mouse) both
contribute to AER formation.
What the papers say
866 BioEssays 23.10

To validate this model, which is based primarily on data
from ``gain-of-function'' experiments (i.e. inserting a source of
a specific FGF in the interlimb region and determining what
genes are subsequently expressed), some form of genetic test
needs to be performed. If one assumes that the signals for limb
induction have been evolutionarily conserved, then an
analysis of the limb phenotypes in mutant mice can provide a
critical test of the model, as well as offer new insights into the
specific functions of individual molecules in the proposed
signaling cascade. Importantly, the central tenet of the working
model, namely the concept that an FGF10 signal is essential
for limb development, has been confirmed by loss-of-function
studies in the mouse. The data show that, although limb bud
outgrowth is initiated, the AER does not form and limb
development therefore fails in mutant mouse embryos that
cannot produce either FGF10(13,14) or its receptor, an isoform
of FGFR2 (FGFR2-IIIb).(15,16) Furthermore, as predicted by
the working model, Fgf8 is not expressed in the surface
ectoderm of Fgf10ÿ /ÿor Fgfr2ÿ /ÿ limb buds.(13,14,16) There
are, however, some conflicting data on this point, since Fgf8
RNA has been detected in the limb bud rudiments of embryos
that cannot produce FGFR2-IIIb, the specific isoform of
FGFR2 that transduces the FGF10 signal in the limb bud.(15)
On the other hand, the genetic data clearly contradict the
assumption implicit in the working model that induction of Fgf8
expression by FGF10 is vital for limb development. When Fgf8
is inactivated specifically in prospective limb ectoderm, the
AER forms and a limb develops, although it is not completely
normal.(17,18) Therefore, the failure of AER formation in
Fgf10ÿ /ÿ or Fgfr2ÿ /ÿ limb buds cannot be attributed solely
to a lack of FGF8, and other targets of FGF10 signaling must
be required for the formation of an AER.
Questions about other features of the working model
remain unresolved. For example, does FGF8 produced in
medial tissue act upstream of Fgf10? This hypothesis has
not yet been tested genetically because complete loss of
Fgf8 function in the embryo causes early lethality,(19) and
the mice necessary for conditionally inactivating Fgf8 in the
intermediate mesoderm or primitive streak at the appropriate
stages of development are not yet available. Furthermore, it
has been difficult to test the proposal that there is an FGF8/
FGF10-positive regulatory loop in the established limb bud.
Embryos that lack Fgf8 function in the AER continue to
express Fgf10, suggesting that signaling via FGF8 is not
essential to maintain Fgf10 expression.(17,18) This result is not
so easily interpreted, however, because, as the AER devel-
ops, other FGF family members in addition to FGF8 begin to be
produced by AER cells, and these other FGFs might maintain
Fgf10 expression in the absence of FGF8. The question of
whether FGF10 signaling is required to maintain Fgf8
expression in the AER cannot be answered by analyzing
mutant mouse embryos that cannot signal via FGF10,
because they never develop an AER. In principle, however,
this question could be addressed by inactivating Fgf10 in the
limb bud mesenchyme or its receptor in the AER, after the AER
has been established.
In summary, we know that FGF10 produced in the
mesoderm is necessary for AER formation. But we do not
yet know for certain what signals are responsible for ensuring
that Fgf10 is expressed appropriately in prospective limb
territories or what targets of FGF10 signaling are responsible
for forming the AER (Fig. 1B). The recent paper by Kawakami
et al.(1) turns the spotlight on members of another family of
signaling molecules, the WNTs.
Introducing new playersÐWNTs take center
stage
Like FGFs, members of the WNT superfamily are secreted
proteins that play major roles in multiple aspects of embryonic
development.(20) WNTs act as ligands for transmembrane
receptors belonging to the Frizzled family. Some WNTs signal
via an intracellular pathway that involves the stabilization of
cytoplasmic b-catenin, which then translocates to the nucleus
and becomes a transcriptional co-activator of WNT target
genes.(21) One member of the WNT gene family, Wnt7a, which
is expressed in the dorsal ectoderm of the limb bud, is essential
for normal dorsoventral patterning of the mouse limb.(4,22) But
until now it had not been shown that WNT genes play a role at
the earliest stages of limb development (Fig. 1C).
Kawakami et al.(1) performed both gain- and loss-of-
function experiments to show that signaling via b-catenin
functions upstream of FGF10 in the limb-induction pathway.
First, they used a retroviral vector to focally express an
activated form of b-catenin in the chick interlimb region. This
resulted in ectopic Fgf10 expression and the formation of an
ectopic limb. Second, they used an adenoviral vector to
express Axin, a negative regulator of b-catenin, in the normal
limb-forming territory several hours before limb induction is
thought to occur. This completely blocked limb formation.
Together these data provide strong support for the hypothesis
that signaling via b-catenin functions to ensure that Fgf10
expression is maintained in the limb-forming territories,
which in turn is required for AER formation in the nascent
limb bud.
The question of which specific WNT family members might
be responsible for stabilizing b-catenin and thus affecting
Fgf10 expression was addressed by studying the normal
patterns of WNT family gene expression in the early chick
embryo. Interestingly, two different WNT genes were identified
as candidate limb inducers. WNT2B was proposed to be
involved in forelimb and WNT8C in hindlimb induction. In
support of the hypothesis that these genes function upstream
of Fgf10 during limb development, the authors showed that
ectopic expression of either Wnt2b or Wnt8c in the chick
interlimb region can lead to Fgf10 expression in cells
near the source of the WNT signal and, subsequently, to the
What the papers say
BioEssays 23.10 867

induction of an ectopic limb bud. Furthermore, they provided
evidence that both of these WNT signals act by stabilizing
b-catenin.
In addition to its role upstream of FGF10 signaling,
Kawakami et al. propose that WNT signaling via b-catenin
also functions downstream of FGF10. This hypothesis is
based on the finding that Fgf8 expression is severely reduced
and the AER is disrupted in the normal limb when Axin is
expressed throughout the limb territory after Fgf10 expression
has become normally restricted to the LPM of the limb-forming
region. They suggest that the WNT family member that effects
this b-catenin-dependent function is Wnt3a, which is normally
expressed in prospective chick limb bud ectoderm, and
subsequently in the AER.(23) In support of the concept that
activation of Fgf8 expression and AER formation by FGF10 is
mediated by WNT3A (Fig. 1C), they showed that implanta-
tion of a pellet of FGF10-producing cells in the interlimb region
induces first Wnt3a expression and then Fgf8 expression in
the overlying surface ectoderm.
An open casting call
From its inception, the model that FGF signaling is responsible
for inducing limb development was regarded as a framework
upon which to build a more complete description of the
molecular mechanism of limb induction. The evidence that
WNT family genes play a role in this process adds a new
dimension to the model. If the hypothesis that a medial FGF8
signal maintains Fgf10 expression in the limb-forming regions
is incorrect, then WNT/b-catenin signaling (via WNT2B in the
forelimb and WNT8C in the hindlimb) may prove to be the
earliest known regulator of Fgf10 expression in the prospec-
tive limb territories.
Perhaps the most interesting question raised by the data
accumulated to date is what are the target genes of FGF10
signaling to the surface ectoderm that are required for AER
formation? As noted above, genetic studies have shown that
Fgf8, which is the only FGF family member known to be
expressed in prospective limb ectoderm prior to AER forma-
tion, is not necessary for AER formation.(17,18) Kawakami et al.
propose that WNT3A plays a critical role in AER formation.
However, when b-catenin signaling is blocked by Axin
expression after Fgf10 expression has become restricted to
the limb-forming territory, a truncated limb develops, in which
proximal elements are present and distal elements are
missing. This phenotype is more similar to what is observed
when the AER is removed at a relatively early stage in the
outgrowth of an established limb bud than to what is seen when
the AER fails to form. One intriguing possibility is that FGF and
WNT signals contribute to AER formation, and that the failure
to form an AER in mouse embryos that cannot signal via
FGF10 is due to a lack of both FGF8 and the WNT3A ortholog
in mice. As research into the question of what induces limb
development continues, it will be exciting to learn how the
known players interact and what other molecules have a place
at the center of the stage.
References1. Kawakami Y, Capdevila J, Buscher D, Itoh T, Rodriguez Esteban
C, IzpisuÂa Belmonte JC. WNT signals control FGF-dependent limb
initiation and AER induction in the chick embryo. Cell 2001;104:891±900.
2. Fallon JF, Frederick JM, Carrington JL, Lanser ME, Simandl BK. Studies
on a limbless mutant in the chick embryo. In Fallon JF, Caplan AI ed;
Limb Development and Regeneration, Part A. New York: Alan R. Riss.
1983. 33±43.
3. Summerbell D. A quantitative analysis of the effect of excision of the AER
from the chick limb bud. J Embryol Exp Morph 1974;32:651±660.
4. Martin GR. The roles of FGFs in the early development of vertebrate
limbs. Genes Dev 1998;12:1571±1586.
5. Cohn MJ, IzpisuÂa-Belmonte J-C, Abud H, Heath JK, Tickle C. Fibroblast
growth factors induce additional limb development from the flank of
chick embryos. Cell 1995;80:739±746.
6. Pawson T. Protein modules and signalling networks. Nature 1995;373:
573±580.
7. Klint P, Claesson-Welsh L. Signal transduction by fibroblast growth factor
receptors. Front Biosci 1999;4:D165±177.
8. Szebenyi G, Fallon JF. Fibroblast growth factors as multifunctional
signaling factors. Int Rev Cytol 1999;185:45±106.
9. Ornitz DM, Itoh N. Fibroblast growth factors. Genome Biol 2001;2:in
press.
10. Crossley PH, Minowada G, MacArthur CA, Martin GR. Roles for FGF8 in
the induction, initiation, and maintenance of chick limb development. Cell
1996;84:127±136.
11. Vogel A, Rodriguez C, IzpisuÂa-Belmonte JC. Involvement of FGF-8 in
initiation, outgrowth and patterning of the vertebrate limb. Development
1996;122:1737±1750.
12. Ohuchi H et al. The mesenchymal factor, FGF10, initiates and maintains
the outgrowth of the chick limb bud through interaction with FGF8, an
apical ectodermal factor. Development 1997;124:2235±2244.
13. Min H, Danilenko DM, Scully SA, Bolon B, Ring BD, Tarpley JE,
DeRose M, Simonet WS. Fgf-10 is required for both limb and lung
development and exhibits striking functional similarity to Drosophila
branchless. Genes Dev 1998;12:3156±3161.
14. Sekine K et al. Fgf10 is essential for limb and lung formation. Nat Genet
1999;21:138±141.
15. Revest JM, Spencer-Dene B, Kerr K, De Moerlooze L, Rosewell I,
Dickson C. Fibroblast growth factor receptor 2-IIIb acts upstream of Shh
and Fgf4 and is required for limb bud maintenance but not for the
induction of Fgf8, Fgf10, Msx1, or Bmp4. Dev Biol 2001;231:47±62.
16. Xu X, Weinstein M, Li C, Naski M, Cohen RI, Ornitz DM, Leder P, Deng C.
Fibroblast growth factor receptor 2 (FGFR2)-mediated reciprocal
regulation loop between FGF8 and FGF10 is essential for limb induction.
Development 1998;125:753±765.
17. Lewandoski M, Sun X, Martin GR. Fgf8 signalling from the AER is
essential for normal limb development. Nat Genet 2000;26:460±463.
18. Moon AM, Capecchi MR. Fgf8 is required for outgrowth and patterning of
the limbs. Nat Genet 2000;26:455±459.
19. Sun X, Meyers EN, Lewandoski M, Martin GR. Targeted disruption of
Fgf8 causes failure of cell migration in the gastrulating mouse embryo.
Genes Dev 1999;13:1834±1846.
20. Wodarz A, Nusse R. Mechanisms of Wnt signaling in development. Annu
Rev Cell Dev Biol 1998;14:59±88.
21. Willert K, Nusse R. Beta-catenin: a key mediator of Wnt signaling. Curr
Opin Genet Dev 1998;8:95±102.
22. Parr BA, McMahon AP. Dorsalizing signal Wnt-7a required for normal
polarity of D-V and A-P axes of mouse limb. Nature 1995;374:350±353.
23. Kengaku M, Capdevila J, Rodriguez-Esteban C, De La Pena J, Johnson
RL, Belmonte JC, Tabin CJ. Distinct WNT pathways regulating AER
formation and dorsoventral polarity in the chick limb bud. Science
1998;280:1274±1277.
What the papers say
868 BioEssays 23.10





























