Embed Size (px)
Citation preview

of May 14, 2018.This information is current as
SAP-Deficient MiceCharacteristics Associated with SLAM- andDefects yet Do Not Share Several Phenotypic Ly9 (CD229)-Deficient Mice Exhibit T Cell
Shane Crotty and David J. McKeanFaubion,Catherine J. Huntoon, Jan van Deursen, William A.
Daniel B. Graham, Michael P. Bell, Megan M. McCausland,
http://www.jimmunol.org/content/176/1/291doi: 10.4049/jimmunol.176.1.291
2006; 176:291-300; ;J Immunol
Referenceshttp://www.jimmunol.org/content/176/1/291.full#ref-list-1
, 17 of which you can access for free at: cites 28 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2006 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on May 14, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on May 14, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Ly9 (CD229)-Deficient Mice Exhibit T Cell Defects yet Do NotShare Several Phenotypic Characteristics Associated withSLAM- and SAP-Deficient Mice1
Daniel B. Graham,2* Michael P. Bell,* Megan M. McCausland,§ Catherine J. Huntoon,*Jan van Deursen,† William A. Faubion,‡ Shane Crotty,§ and David J. McKean3*
Signaling lymphocyte activation molecule (SLAM) family receptors are critically involved in modulating innate and adaptiveimmune responses. Several SLAM family receptors have been shown to interact with the adaptor molecule SAP; however,subsequent intracellular signaling is poorly defined. Notably, mutations in SLAM-associated protein (SAP) lead to X-linkedlymphoproliferative disease, a rare but fatal immunodeficiency. Although the SLAM family member Ly9 (CD229) is known tointeract with SAP, the functions of this receptor have remained elusive. Therefore, we have generated Ly9�/� mice and comparedtheir phenotype with that of SLAM�/� and SAP�/� mice. We report that Ly9�/� T cells exhibit a mild Th2 defect associated withreduced IL-4 production after stimulation with anti-TCR and anti-CD28 in vitro. This defect is similar in magnitude to thepreviously reported Th2 defect in SLAM�/� mice but is more subtle than that observed in SAP�/� mice. In contrast to SLAM�/�
and SAP�/� mice, T cells from Ly9�/� mice proliferate poorly and produce little IL-2 after suboptimal stimulation with anti-CD3in vitro. We have also found that Ly9�/� macrophages exhibit no defects in cytokine production or bacterial killing as wasobserved in SLAM�/� macrophages. Additionally, Ly9�/� mice differ from SAP�/� mice in that they foster normal developmentof NKT cells and mount appropriate T and B cell responses to lymphocytic choriomeningitis virus. We have identified significantphenotypic differences between Ly-9�/� mice as compared with both SLAM�/� and SAP�/� mice. Although Ly9, SLAM, and SAPplay a common role in promoting Th2 polarization, Ly-9 is uniquely involved in enhancing T cell activation. The Journal ofImmunology, 2006, 176: 291–300.
T he signaling lymphocyte activation molecule (SLAM)4
family of immunomodulatory receptors is comprised ofSLAM (CD150, IPO-3, Slamf1), Ly9 (CD229, Slamf3),
2B4 (CD244, NAIL, C9.1, Slamf4), Ly108 (SF2000, NTB-A,Slamf6), CS-1 (CRACC, 19A, nLy9, Slamf7), and CD2F10(SF2001, CD84-H1, Slamf9). Like SLAM, Ly9 is reported to be ahomophilic receptor with broad expression throughout the hemopoi-etic system (Refs. 1 and 2 and D. B. Graham, M. P. Bell, C. J.Huntoon, and D. J. McKean, submitted for publication). Ly9 expres-sion has been confirmed in T cells, B cells, macrophages, dendriticcells, and granulocytes (our unpublished observations). All of theSLAM family receptors contain tandem IgV and IgC2 sets in theirextracellular domains and several contain immune receptor tyrosinebased switch motifs (ITSMs) in their cytoplasmic tails. These ITSMs
contain the amino acid sequence TxYxxV/I, which comprises a con-sensus sequence for the Src homology 2 domains of the adaptor mol-ecules SLAM-associated protein (SAP) and Eat-2. Ly9 contains twopairs of IgV IgC2 sets in its extracellular domain and two ITSMs inits cytoplasmic tail. We and others have previously shown that Ly9can interact with both SAP and Eat-2, although the functional rele-vance of Ly9 signaling has remained unclear (Refs. 3 and 4 and D. B.Graham, M. P. Bell, C. J. Huntoon, and D. J. McKean, submitted forpublication).
SAP signaling downstream of SLAM has been extensively stud-ied in the context of T cell activation. As an adaptor molecule,SAP bridges SLAM to the Src family kinase Fyn, which regulatesprotein kinase C�-dependent NF-�B activation (5, 6). The impor-tance of this SAP signaling cascade was illustrated when mutationsin SAP were found to cause X-linked lymphoproliferative (XLP)disease in humans (7–9). This fatal immunodeficiency is charac-terized by fulminant infectious mononucleosis, dysgammaglobu-linemia, and/or lymphoproliferative disorders (6). Several aspectsof human XLP disease are mimicked in SAP�/� mice, particularlysusceptibility to viral infections. Lymphocytic choriomeningitisvirus (LCMV) infection in SAP�/� mice results in an exaggeratedacute response associated with lymphoproliferation and increasednumbers of LCMV-specific CD4 and CD8 T cells secreting largequantities of IFN-� (10–12). Interestingly, B cell responses toLCMV are severely compromised in SAP�/� mice, and this defectis dependent on lack of help from CD4 T cells (10). Specifically,SAP�/� mice produce few germinal center B cells, long-livedplasma cells, and LCMV-specific memory B cells (10). A severedefect in B cell memory has also been observed in human XLP
*Department of Immunology, †Department of Pediatric and Adolescent Medicine,and ‡Department of Gastroenterology, Mayo Clinic College of Medicine, Rochester,MN 55905; and §Division of Vaccine Discovery, La Jolla Institute for Allergy andImmunology, San Diego, CA 92121
Received for publication July 1, 2005. Accepted for publication October 20, 2005.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by grants from the National Institutes of Health (AI44959to D.J.M.; T32 AI07425 to D.B.G.) and by a Cancer Research Institute InvestigatorAward (to S.C.).2 Current address: Department of Pathology and Immunology, Washington Univer-sity, St Louis, MO.3 Address correspondence and reprint requests to Dr. David J. McKean, Departmentof Immunology, First Street S.W., Mayo Clinic College of Medicine, Rochester, MN55905. E-mail address: [email protected] Abbreviations used in this paper: SLAM, signaling lymphocyte activation molecule;SAP, SLAM-associated protein; ITSM, immune receptor tyrosine based switch motif;
�GalCer, �-galactosylceramide; LCMV, lymphocytic choriomeningitis virus; LCM-Varm, Armstrong strain of LCMV; XLP, X-linked lymphoproliferative.
The Journal of Immunology
Copyright © 2005 by The American Association of Immunologists, Inc. 0022-1767/05/$02.00
by guest on May 14, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
patients (13, 14) and is consistent with the hypogammaglobuline-mia observed in these patients (6, 15).
As was observed in LCMV-infected mice, CD4 T cells fromuninfected SAP�/� mice produce abnormally large quantities ofIFN-� after stimulation in vitro with anti-TCR and anti-CD28 orAg-pulsed APCs (11, 12). Upon further investigation, SAP�/�
CD4 T cells show a strong skewing toward Th1 responses anddefects in the production of Th2 cytokines such as IL-4, IL-10, andIL-13 (11, 12). In addition to CD4 T cells, NKT cells contribute toTh1/Th2 polarization by producing large quantities of polarizingcytokines (IFN-� or IL-4) early in immune responses. Interest-ingly, SAP�/� mice have virtually no NKT cells, which may ex-acerbate the observed Th2 defect or impart distinct immunologicdefects (16–18). Also consistent with a Th2 defect is the obser-vation that class switching to IgE is defective in SAP�/� mice,which emphasizes that this T cell defect directly impacts B cellresponses (12). Put into context, Th2 responses are very relevant tohost defense. Accordingly, SAP�/� mice are more effective atmounting protective Th1 responses to leishmania than their wild-type counterparts (11).
Consistent with the notion that some SLAM signaling is trans-duced through SAP, the SLAM knockout phenotype resemblesseveral aspects of SAP deficiency in humans. Purified CD4 T cellsfrom SLAM�/� mice produce very little IL-4 and slightly elevatedlevels of IFN-� after stimulation with anti-TCR and anti-CD28 orAg-pulsed APCs in vitro (19). This Th2 defect is much less severein SLAM�/� mice compared with SAP�/� mice, presumably be-cause additional SLAM family members that signal through SAPare still functional. It was expected that the Th1 skewing observedin SLAM�/� mice would protect them from infection with leish-mania, as was observed in SAP�/� mice. Surprisingly, SLAM�/�
mice were more susceptible to leishmaniasis than control mice(19). Closer examination revealed that SLAM�/� mice have aSAP-independent macrophage defect that accounts for their inabil-ity to control leishmania. Macrophages from SLAM�/� mice pro-duce unusually large quantities of IL-6 and significantly lessTNF-�, IL-12, and NO in response to LPS (19).
Taken together, the SLAM knockout phenotype cannot be ex-plained entirely by disrupted SAP signaling. Furthermore, the SAP
knockout phenotype cannot be explained entirely by disrupted sig-naling downstream of SLAM. Therefore, additional SLAM familyreceptors must regulate SAP signaling. Because Ly9 resemblesSLAM in structure and interacts with SAP, we have generatedLy9�/� mice to address these issues.
Materials and MethodsAbs and reagents
Fluorescently conjugated Abs specific for CD8 (53-6.7), CD4 (H129.19),CD19 (1D3), CD28 (37.51), CD11c (HL3), IL-4 (11B11), IFN-�(XMG1.2), Fas (Jo2), B220 (RA3-6B2), CD138, TNF-�, IFN-�, andTCR� (H57-597) were purchased from BD Pharmingen or eBioscience.Goat anti-rat IgG, goat anti-rabbit IgG, and streptavidin-HRP were alsoobtained from BD Pharmingen. Biotinylated anti-IgD was purchased fromeBioscience. Polyclonal antisera specific for Ly9 was collected from rab-bits that had been immunized with His-tagged Ly9 extracellular domain orLy9 cytoplasmic tail produced in Escherichia coli. Anti-CD3 (145-2C11)and anti TCR� (H57-597) were purified from hybridoma culture superna-tant. CD1d tetramers were generously provided by M. Kronenberg (LaJolla Institute for Allergy and Immunology, San Diego, CA). MHC class Itetramer of Db loaded with LCMV gp33–41 was provided by Dr. H. Cheroutre(La Jolla Institute for Allergy and Immunology).
Generation of Ly9�/� mice
A murine genomic library packaged in � phage was screened for clonescontaining the full-length Ly9 gene. A positive clone was used as a tem-plate to amplify portions of the Ly9 gene by PCR. The 5� arm was flankedby HpaI restriction sites and contained exon 1 and part of exon 2. The 3�arm was flanked by SmaI restriction sites and contained exons 3 and 4. The5� and 3� arms were then cloned into the pKO Scrambler targeting vector(Stratagene) flanking a neomycin resistance gene. The targeting vector wasthen introduced into 129/SvEES cells by homologous recombination, andclones containing the mutant Ly9 allele were screened by Southern blotusing a probe complementary to exon 5. A positive embryonic stem cellclone was injected into blastocysts derived from C57BL/6 mice and im-planted into pseudopregnant surrogate mothers to generate chimeric mice.These chimeras were subsequently bred with C57BL/6 mice to generatemice heterozygous for the mutated Ly9 allele. F1 heterozygotes were usedas breeding stock to generate homozygous Ly9 mutants. All mice weregenotyped by PCR. All procedures involving mice were approved by theInstitutional Animal Care and use Committees at the Mayo Clinic Collegeof Medicine and the La Jolla Institute for Allergy and Immunology.
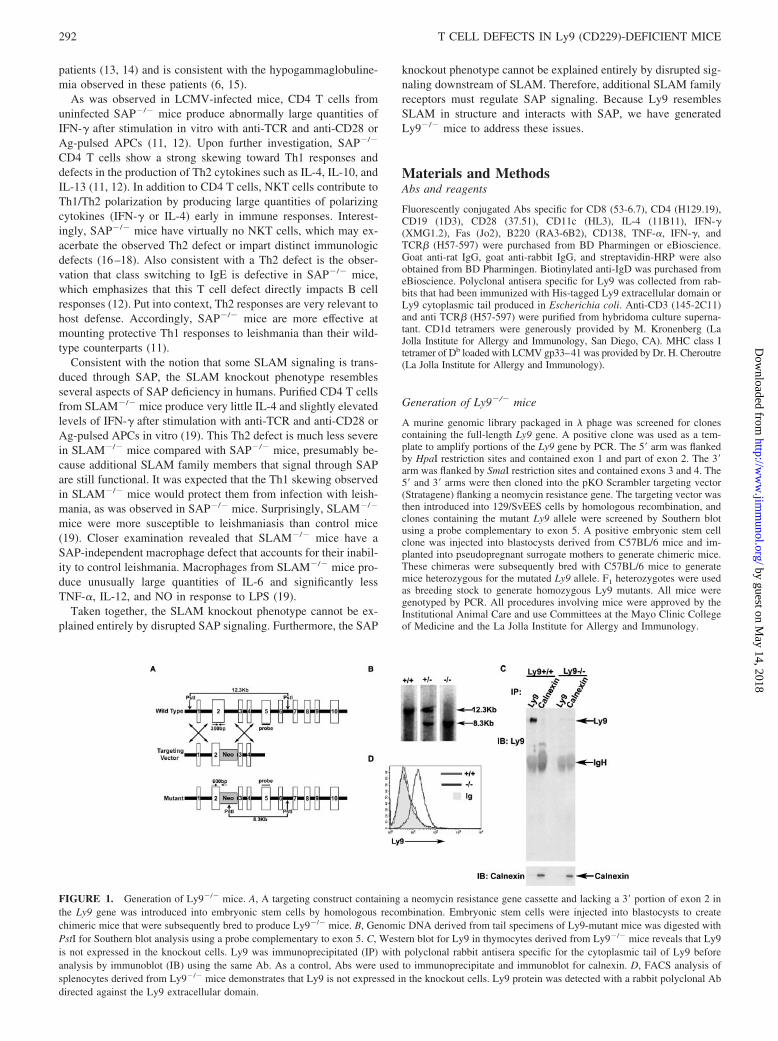
FIGURE 1. Generation of Ly9�/� mice. A, A targeting construct containing a neomycin resistance gene cassette and lacking a 3� portion of exon 2 inthe Ly9 gene was introduced into embryonic stem cells by homologous recombination. Embryonic stem cells were injected into blastocysts to createchimeric mice that were subsequently bred to produce Ly9�/� mice. B, Genomic DNA derived from tail specimens of Ly9-mutant mice was digested withPstI for Southern blot analysis using a probe complementary to exon 5. C, Western blot for Ly9 in thymocytes derived from Ly9�/� mice reveals that Ly9is not expressed in the knockout cells. Ly9 was immunoprecipitated (IP) with polyclonal rabbit antisera specific for the cytoplasmic tail of Ly9 beforeanalysis by immunoblot (IB) using the same Ab. As a control, Abs were used to immunoprecipitate and immunoblot for calnexin. D, FACS analysis ofsplenocytes derived from Ly9�/� mice demonstrates that Ly9 is not expressed in the knockout cells. Ly9 protein was detected with a rabbit polyclonal Abdirected against the Ly9 extracellular domain.
292 T CELL DEFECTS IN Ly9 (CD229)-DEFICIENT MICE
by guest on May 14, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

FACS analysis
Cell surface staining was performed according to standard procedures. Allsamples were subsequently analyzed on a FACScan flow cytometer withCellQuest software (BD Biosciences) (see Figs. 2–5) or acquired on aFACSCalibur and analyzed with FlowJo (see Figs. 6 and 7).
T cell activation
For proliferation assays, splenocytes were stimulated with anti-CD3(2C11) for 3 days in round-bottom 96-well plates at a density of 2 � 105
cells per 200 �l per well. During the last 20 h of culture, [3H]thymidine(6.7 Ci/mmol; MB Biochemicals) was added to the cells to achieve a finalconcentration of 1 �Ci/well. Cells were then harvested and [3H]thymidineincorporation was measured on a microtiter plate counter. Alternatively,culture supernatant was harvested after 24 or 48 h in culture, and IL-2production was measured by ELISA kit (BD Pharmingen).
T cell stimulation and polarization
CD4 T cells were purified from spleen and lymph nodes by positive se-lection on MACS beads (Miltenyi Biotec) and cultured under neutral con-ditions or Th1 or Th2 polarizing conditions. T cells were cultured in 24-well plates at a density of 5 � 105 cells per well. Under neutral conditions,T cells were stimulated with the indicated concentrations of plate-boundanti-TCR and anti-CD28 for 3 days. Preliminary experiments identified Abconcentrations that maximized CD28 costimulatory effects. Under Th1 po-larizing conditions, cells were stimulated with anti-TCR (0.5 �g/ml) andanti-CD28 (5 �g/ml) with the addition of IL-12 (1 ng/ml; R&D Systems)and anti-IL-4 (10 �g/ml, clone 11B11; BD Pharmingen) for 3 days. Th2polarizing conditions consisted of anti-TCR (0.5 �g/ml) and anti-CD28 (5
�g/ml) with the addition of IL-4 (100 ng/ml; R&D Systems), anti-IL-12(10 �g/ml, clone C17.8; BD Pharmingen), and anti-IFN-� (10 �g/ml, cloneR4-6A2; BD Pharmingen) for 3 days. After 3 days in culture, cells werewashed and restimulated overnight in 24-well plates at a density of 5 � 105
cells/well. Overnight restimulation consisted of anti-TCR and anti-CD28 atthe same concentrations as the initial stimulation. The following day, cul-ture supernatants were harvested for ELISAs, and Golgi Stop (BD Pharm-ingen) was added to the cells for 8 h before intracellular cytokine staining.ELISAs for IL-4 and IFN-� were performed as per the manufacturer’srecommendations (R&D Systems). Intracellular cytokine staining with theCytofix/Cytoperm kit (BD Pharmingen) was performed according to themanufacturer’s recommendations. Cells were then analyzed by FACS.
Quantitative PCR
CD4 T cells were purified from spleen and lymph nodes by positive se-lection on MACS beads (Miltenyi Biotec) and stimulated for 2 days withanti-TCR (0.5 �g/ml plate bound) and anti-CD28 (5 �g/ml plate bound).RNA was then isolated with TRIzol reagent (Invitrogen Life Technologies)and amplified by quantitative PCR. Reactions contained TaqMan One StepRT-PCR master mix (PE Applied Biosystems), 290 ng template RNA, 25pmol of primers, and 0.2 pmol of TaqMan probe in a total volume of 50 �l.The following primers and probes were used to detect GATA-binding pro-tein 3: forward CTACCGGGTTCGGATGTAAGTC, reverse GTTCACACACTCCCTGCCTTCT, probe 6FAM-AGGCCCAAGGCACGATCCAGC-TAMRA. The following primers and probes were used to detectT-bet: forward ACCAGAACGCAGAGATCACTCA, reverse CAAAGTTCTCCCGGAATCCTT, probe 6FAM-CTGAAAATCGACAACAACCCCTTTGCC-TAMRA. Primers and probes specific for murine GAPDHwere purchased from PE Applied Biosystems. Thermal cycling conditions
FIGURE 2. T cell activation in Ly9�/� mice. A,Thymus and spleen tissues were harvested from miceand stained with the indicated Abs before FACS anal-ysis. B, Splenocytes were stimulated with varying con-centrations of anti-CD3 (2C11) for 48 h and labeledwith [3H]thymidine for an additional 20 h. Splenocytesfrom three wild-type and three Ly9�/� mice were ana-lyzed in triplicate (n � 2). C, Splenocytes from threeLy9�/� mice and three littermates were stimulated withanti-CD3 (3 �g/ml) as described in A. After 24 or 48 h,IL-2 was measured by ELISA (n � 3).
293The Journal of Immunology
by guest on May 14, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

consisted of a 30 min incubation at 48°C, and an initial denaturation at95°C for 10 min, followed by 40 cycles of 95°C for 15 s and 60°C for 1min. Reactions were performed in a spectrofluorometric thermal cycler(ABI PRISM 7700; PE Applied Biosystems). For each run, standard curveswere generated with dilutions of total RNA to calculate relative copies ofT-bet and GATA3 vs GAPDH.
Quantification of serum IgE
Serum was collected from unimmunized mice (6–12 wk old) and analyzedfor IgE by ELISA. Procedures were performed according to the manufac-turer’s recommendations (BD Biosciences).
Macrophage functions
Mice were injected i.p. with thioglycolate 5 days before sacrifice. Macro-phages were then harvested by peritoneal lavage and cultured overnight ata density of 1 � 106 cells per well in 24-well plates (Corning). A total of1 � 107 F18 E. coli was added to each well of macrophages for 1 h and
then washed thoroughly. Macrophages were cultured for an additional hourin RPMI 1640 containing 100 �g/ml gentamicin. Cells were then lysed in1% Triton X-100 in water or cultured for an additional 4 h in RPMI 1640containing 10 �g/ml gentamicin before lysis. Dilutions of the lysate werethen plated onto Laurie broth agar and incubated overnight. The next day,colonies were counted to determine CFUs per 1 � 106 macrophages. Forcytokine analysis, peritoneal macrophages were harvested from mice thathad not received thioglycolate treatment. Cells were cultured in 96-wellplates (Corning) at a density of 6.5 � 105 cells per 200 �l per well for 3days. Stimulation conditions consisted of LPS (200 ng/ml), CpG (10 �g/ml), or medium alone. After 3 days, culture supernatants were harvestedand analyzed by ELISA for IL-6 or IL-12 according to manufacturer’srecommendations (R&D Systems).
NKT cells
Spleen and thymus tissue was harvested and minced into a single cellsuspension. Cells were then stained with anti-TCR� and CD1d tetramers
FIGURE 3. Mild Th2 defect in Ly9�/� mice. A, CD4T splenic cells were cultured in control, Th1, or Th2polarizing conditions and then stimulated for 3 dayswith plate-bound anti-TCR and anti-CD28. After theinitial 3-day culture, cells were washed and restimulatedovernight with anti-TCR and anti-CD28. The followingday, cells were analyzed for intracellular IL-4 and IFN-�expression. Each sample consisted of T cells pooledfrom four Ly9�/� mice or four wild-type littermates(n � 4). B, T cells were treated as in A. Cytokinespresent in supernatant from overnight restimulation cul-tures were quantified by ELISA. Four pairs of Ly9�/�
and wild-type littermates were analyzed in this experi-ment (n � 4). C, CD4 T cells from four Ly9�/� or fourwild-type littermates were stimulated with plate-boundanti-TCR (0.5 �g/ml) and anti-CD28 (5 �g/ml) for 2days. Subsequently, RNA was isolated from the cells foranalysis of GATA3 and T-bet expression by quantitativeRT-PCR. The values plotted are arbitrary units relativeto GAPDH expression (n � 4). D, Serum was collectedfrom unimmunized mice (6–12 wk old), and IgE wasquantified by ELISA (n � 4).
294 T CELL DEFECTS IN Ly9 (CD229)-DEFICIENT MICE
by guest on May 14, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
(1/100 dilution) loaded with �-galactosylceramide (�GalCer) or unloadedCD1d tetramers (20). Cells were then analyzed by FACS.
LCMV infections
LCMV experiments were performed similarly to those previously de-scribed (10, 12). Plaque-purified clones of the Armstrong strain of LCMV(LCMVarm) were propagated in BHK-21 cells (American Type CultureCollection), and tested for biological activity in vitro and in vivo. A secondpassage stock of subclone SC3 (LCMVarm-sc3) was used for all LCMVarm
experiments. For acute infections, mice received 1 � 105 PFU LCMVarm
in a volume of 0.5 ml (suspended in RPMI 1640) by bilateral i.p.inoculation.
Plasma cell ELISPOT
Plasma cells were quantitated by a modification of the ELISPOT methodpreviously reported (10, 21). Sonicated lysate from LCMV-infectedBHK-21 cells was used as capture Ag for LCMV-specific ELISPOT. Nine-ty-six-well MAHA N4510 filter plates were used (Millipore). Goat anti-mouse IgG�M�A (Caltag Laboratories; 62.5 �l/10 ml) was used as cap-ture Ab for total Ig ELISPOTs. LCMV Ag was UV inactivated (300 mJ inStratalinker 1800; Stratagene) after overnight coating onto ELISPOT plate.Plates were blocked with DMEM plus 10% FCS. Cells of interest wereadded to the plate in 3� serial dilutions in DMEM plus 10% FCS andincubated at 37°C, 5–8% CO2, for 5–6 h. Biotinylated goat anti-mouseIgG� (Caltag Laboratories) followed by streptavidin-HRP (Vector Labo-ratories) was used for detection. 3-Amino-9-ethylcarbazole was used forspot development. Incubation buffers used PBS plus 0.05% Tween 20 plus1% FCS, and wash buffers used PBS plus 0.05% Tween 20. ELISPOTplates were scanned by an ImmunoSpot Analyzer and machine countedusing a standardized set of digital counting parameters we established inImmunoSpot 3.2 (Cell Technology).
Intracellular cytokine staining, LCMV experiments
A total of 1 � 106 cells was cultured in the absence or presence of theindicated peptide and brefeldin A for 5–6 h at 37°C. H-2Db- or H-2Kb-restricted epitopes were used at 0.2 �g/ml, and I-Ab-restricted epitopes at2 �g/ml. After staining for surface Ags, cells were fixed and permeabilizedwith 2% (w/v) paraformaldehyde and 0.1% saponin for 15 min, thenstained for the intracellular cytokine of interest in the presence of 0.1%saponin and 2% NCS (GE Healthcare Life Sciences) for 30 min. Cells werewashed and then fixed in 2% ultrapure formaldehyde (Polysciences).
Statistical analysis
Tests were performed using Prism 4.0 (GraphPad). Statistics were doneusing two-tailed, unpaired t test with 95% confidence bounds. Error barsare �1 SEM. Arithmetic means were used for all analyses.
ResultsGeneration of Ly9�/� mice
To identify immunologic functions of Ly9, mice containing a tar-geted disruption of Ly9 exon 2 were generated by homologousrecombination in embryonic stem cells (Fig. 1A). Western blottingand FACS analysis of thymocytes and splenocytes, respectively,confirmed that mice homozygous for the mutated allele did notexpress Ly-9 protein (Fig. 1, B–D) yet were viable, fertile, andmorphologically indistinguishable from wild-type littermates.
Ly9�/� T cells exhibit activation defects
Both SLAM�/� and SAP�/� mice exhibit normal lymphoid de-velopment (11, 12). Similarly, Ly9�/� mice contained normalnumbers and percentages of T, B, and myeloid cells in the spleenas well as developing T cells in the thymus (Fig. 2A and our un-published observations). Strikingly, Ly9�/� splenocytes stimu-lated for 72 h with anti-CD3 (0.1–10 �g/ml soluble) proliferatedsignificantly less ( p �� 0.05) than wild-type splenocytes (Fig. 2B).Furthermore, Ly9�/� splenocytes produced significantly less IL-2after in vitro stimulation with anti-CD3 for 24 or 48 h ( p � 0.03)(Fig. 2C) This early T cell activation defect in Ly9�/� mice dis-tinguishes Ly9 from the other SLAM family members and sug-gests a potential role for Ly9 in T cell activation or costimulation.
Mild Th2 defect in Ly9�/� T cells
Having shown that Ly9�/� mice exhibit an early T cell activationdefect, we sought to determine whether later-developing Th1 orTh2 responses were skewed. SLAM�/� and SAP�/� T cells aredefective at producing Th2 cytokines and produce excessive quan-tities of Th1 cytokines (5, 11, 12). Similarly, we found that Ly9�/�
CD4 T cells produced less IL-4 compared with wild-type T cells inresponse to stimulation in vitro (Fig. 3). This defect was mild yetsignificant. In these experiments, purified CD4 T cells were stim-ulated in vitro with anti-TCR and anti-CD28 for 3 days. T cells
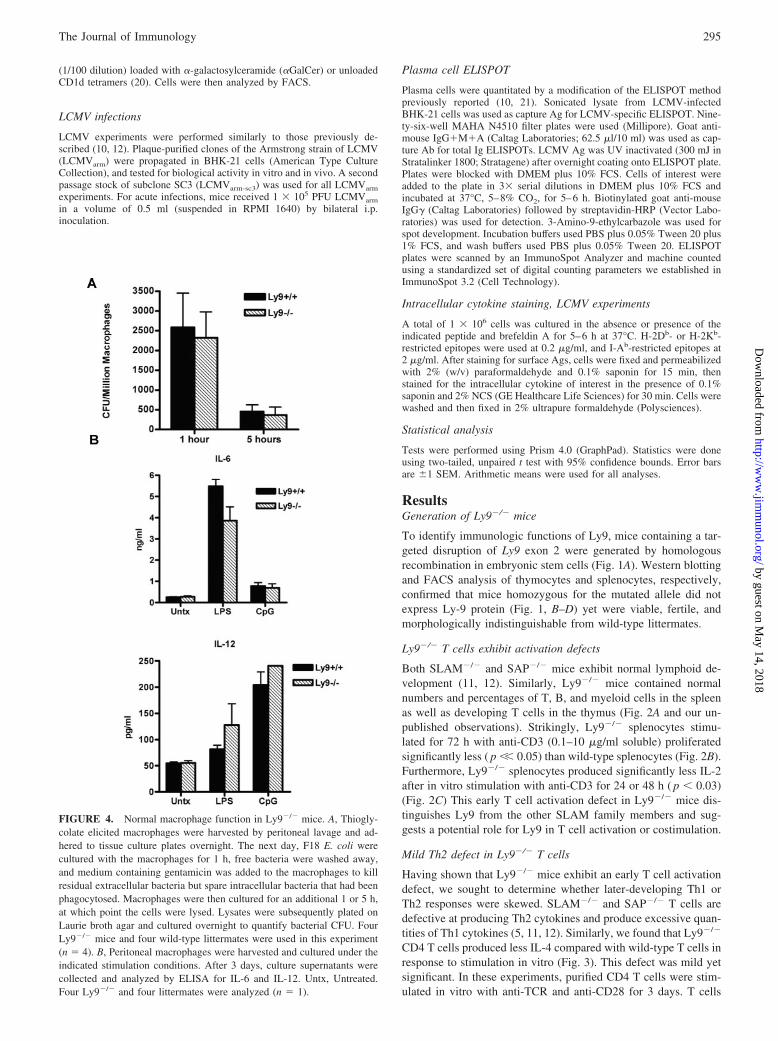
FIGURE 4. Normal macrophage function in Ly9�/� mice. A, Thiogly-colate elicited macrophages were harvested by peritoneal lavage and ad-hered to tissue culture plates overnight. The next day, F18 E. coli werecultured with the macrophages for 1 h, free bacteria were washed away,and medium containing gentamicin was added to the macrophages to killresidual extracellular bacteria but spare intracellular bacteria that had beenphagocytosed. Macrophages were then cultured for an additional 1 or 5 h,at which point the cells were lysed. Lysates were subsequently plated onLaurie broth agar and cultured overnight to quantify bacterial CFU. FourLy9�/� mice and four wild-type littermates were used in this experiment(n � 4). B, Peritoneal macrophages were harvested and cultured under theindicated stimulation conditions. After 3 days, culture supernatants werecollected and analyzed by ELISA for IL-6 and IL-12. Untx, Untreated.Four Ly9�/� and four littermates were analyzed (n � 1).
295The Journal of Immunology
by guest on May 14, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

were then restimulated overnight under identical conditions andevaluated for cytokine production by FACS or ELISA. Intracellu-lar cytokine staining demonstrated that fewer Ly9�/� CD4 T cellsexpressed IL-4 compared with wild-type cells (Fig. 3A). Furtheranalysis showed that Ly9�/� CD4 T cells displayed Th1 skewingas determined by intracellular staining for IFN-� (Fig. 3A). Inter-estingly, culturing Ly9�/� CD4 T cells under Th1 or Th2 polar-izing conditions induced normal production of IL-4 and IFN-�,respectively (Fig. 3, A and B). The implication is that Ly9�/� Tcells are mildly defective in producing Th2 cytokines, but they canrespond appropriately to cytokine stimulation. Notably, IL-2 pro-duction was normal in Ly9�/� T cells after 3 days of stimulationin vitro with anti-TCR and anti-CD28 (data not shown). Thus,CD28-mediated costimulation appeared to rescue the T cell acti-vation defect described in Fig. 2.
ELISA-based quantification of cytokine production revealedthat stimulation with low doses of anti-TCR (0.25 �g/ml platebound) and anti-CD28 elicited reduced IL-4 production in Ly9�/�
T cells compared with wild-type cells ( p � 0.015; Fig. 3B). Athigher doses of anti-TCR (0.5 and 1 �g/ml plate bound) and anti-CD28, Ly9�/� T cells exhibited a modest reduction in IL-4 pro-duction that did not reach statistical significance (Fig. 3B). In con-trast, analysis of cytokine production by ELISA demonstratedcomparable production of IFN-� in Ly9�/� T cells and wild-typeT cells (Fig. 3B).
In addition, quantitative RT-PCR was used to compare levels ofthe Th1-inducing transcription factor T-bet and the Th2-inducingtranscription factor GATA3 in Ly9�/� vs wild-type mice (Fig.3C). Activated CD4 T cells from Ly9�/� mice consistently pro-duced fewer transcripts for GATA3 and more transcripts for T-bet,but the observed differences did not reach statistical significance(Fig. 3C).
The Th2 cytokine defect reported in SAP�/� mice was alsoassociated with reduced serum IgE (12). Based on the observationthat Ly9�/� T cells exhibit defects in IL-4 production, and IL-4plays an important role in class switching to IgE, we measuredlevels of serum IgE in Ly9�/� mice. We observed a slight reduc-tion in serum IgE in unimmunized Ly9�/� mice compared withwild-type mice, but this difference did not reach statistical signif-icance (Fig. 3D). Taken together, Ly9�/� mice appear to have verymild Th2 defects. Similarly, the magnitude of the Th2 defects pre-viously reported in SLAM�/� T cells and Ly108�/� T cells arealso very mild (19, 22). It is likely that several SLAM family
receptors contribute to Th2 polarization and may compensate forLy9 in its absence.
Normal macrophage functions in Ly9�/� mice
In addition to the Th2 defect, the most notable phenotype inSLAM�/� mice was a macrophage defect. Because macrophagesdo not express SAP, this finding identified a SAP-independentfunction for SLAM (19). SLAM and Ly9 share significant homol-ogy within their cytoplasmic tails, and we have shown that bothreceptors interact with several signaling intermediates in commonbesides SAP (D. B. Graham, M. P. Bell, C. J. Huntoon, and D. J.McKean, submitted for publication). Therefore, we pursued afunction for Ly9 in macrophages. Ly9�/� macrophages were ca-pable of killing Gram-negative E. coli as efficiently as wild-typemacrophages (Fig. 4A). In these experiments, thioglycolate-elic-ited peritoneal macrophages were cultured with live E. coli for 1 h,and free bacteria that had not been phagocytosed were subse-quently washed away. Macrophages were then cultured for an ad-ditional 1 or 5 h, at which point, viable bacteria that had beeninternalized by the macrophages were cultured overnight on agarto quantify CFUs. At the 5-h time point, Ly9�/� and wild-typemacrophages contained 3.5-fold fewer CFUs than at the 1-h timepoint (Fig. 4A). During this 4 h culture, Ly9�/� macrophageskilled two-thirds of the bacteria they had phagocytosed during thefirst hour (Fig. 4A). Notably, Ly9�/� macrophages phagocytosedbacteria as efficiently as wild-type macrophages, as indicated bylarge numbers of CFUs measured at the 1-h time point (Fig. 4A).
SLAM�/� macrophages were also previously reported to pro-duce excessive quantities of IL-6 and deficient levels of IL-12 afterstimulation with LPS (19). In contrast, peritoneal macrophagesfrom Ly9�/� and wild-type mice produced comparable quantitiesof IL-6 and IL-12 after stimulation with LPS or CpG, respectively(Fig. 4B). Despite the fact that Ly9 is expressed on wild-type mac-rophages, its absence does not result in a phenotype resemblingthat of SLAM�/� macrophages.
Normal development of NKT cells in Ly9�/� mice
The most recent phenotype documented in SAP�/� mice and inXLP patients is a lack of NKT cells (16–18). Evidence suggeststhat SAP and Fyn are required for the development of NKT cellsbut not traditional T cells or NK cells (16–18, 23). However, it isnot known which receptors may regulate this SAP-dependent Fynactivation. Ly9 and several other SLAM family receptors are
FIGURE 5. Normal development of NKT cells inLy9�/� mice. Thymocytes (A) and splenocytes (B) werestained with anti-TCR� and CD1d �GalCer tetramers todetect NKT cells by FACS. Unloaded (empty) CD1dtetramers were used as a staining control. Data shownare representative of one of four total Ly9�/� miceanalyzed.
296 T CELL DEFECTS IN Ly9 (CD229)-DEFICIENT MICE
by guest on May 14, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
prime candidates, as they become up-regulated on NKT cells dur-ing development. NKT cells from the spleen and thymus ofLy9�/� and wild-type mice were stained with anti-TCR� andCD1d tetramers loaded with �GalCer. As a negative control, cellswere stained with unloaded (empty) CD1d tetramers. Examinationof Ly9�/� mice revealed normal percentages of NKT cells in thethymus and spleen, suggesting that Ly9 alone is not required forNKT cell development (Fig. 5).
Normal responses to LCMV in Ly9�/� mice
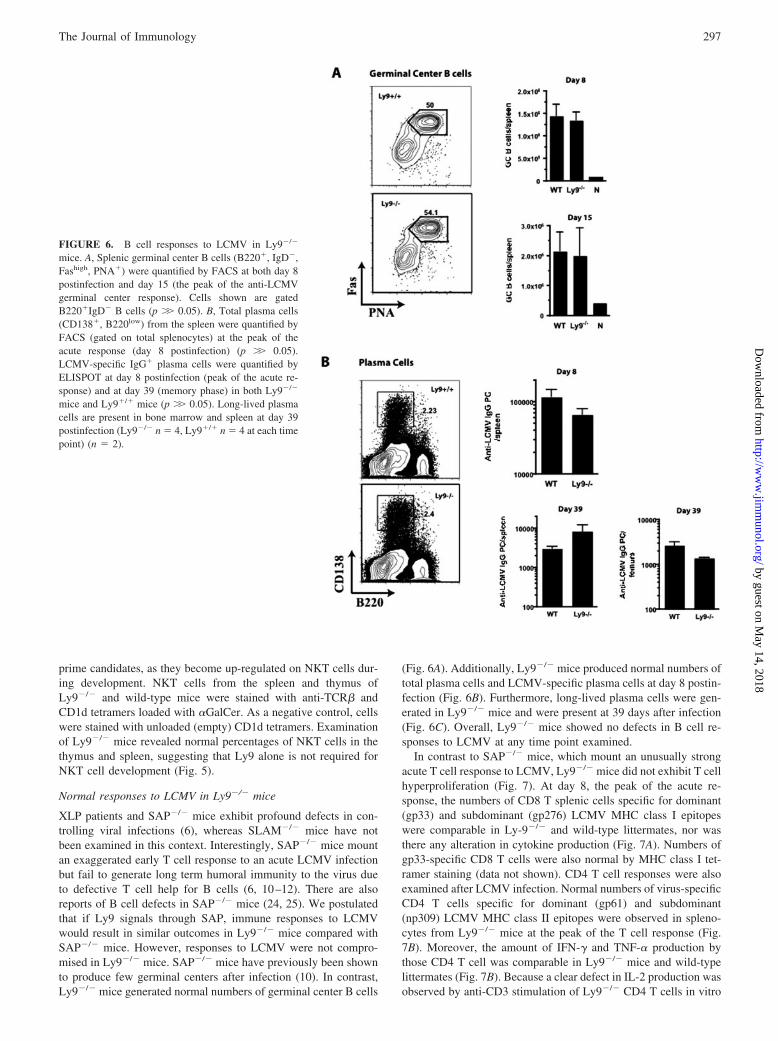
XLP patients and SAP�/� mice exhibit profound defects in con-trolling viral infections (6), whereas SLAM�/� mice have notbeen examined in this context. Interestingly, SAP�/� mice mountan exaggerated early T cell response to an acute LCMV infectionbut fail to generate long term humoral immunity to the virus dueto defective T cell help for B cells (6, 10–12). There are alsoreports of B cell defects in SAP�/� mice (24, 25). We postulatedthat if Ly9 signals through SAP, immune responses to LCMVwould result in similar outcomes in Ly9�/� mice compared withSAP�/� mice. However, responses to LCMV were not compro-mised in Ly9�/� mice. SAP�/� mice have previously been shownto produce few germinal centers after infection (10). In contrast,Ly9�/� mice generated normal numbers of germinal center B cells
(Fig. 6A). Additionally, Ly9�/� mice produced normal numbers oftotal plasma cells and LCMV-specific plasma cells at day 8 postin-fection (Fig. 6B). Furthermore, long-lived plasma cells were gen-erated in Ly9�/� mice and were present at 39 days after infection(Fig. 6C). Overall, Ly9�/� mice showed no defects in B cell re-sponses to LCMV at any time point examined.
In contrast to SAP�/� mice, which mount an unusually strongacute T cell response to LCMV, Ly9�/� mice did not exhibit T cellhyperproliferation (Fig. 7). At day 8, the peak of the acute re-sponse, the numbers of CD8 T splenic cells specific for dominant(gp33) and subdominant (gp276) LCMV MHC class I epitopeswere comparable in Ly-9�/� and wild-type littermates, nor wasthere any alteration in cytokine production (Fig. 7A). Numbers ofgp33-specific CD8 T cells were also normal by MHC class I tet-ramer staining (data not shown). CD4 T cell responses were alsoexamined after LCMV infection. Normal numbers of virus-specificCD4 T cells specific for dominant (gp61) and subdominant(np309) LCMV MHC class II epitopes were observed in spleno-cytes from Ly9�/� mice at the peak of the T cell response (Fig.7B). Moreover, the amount of IFN-� and TNF-� production bythose CD4 T cell was comparable in Ly9�/� mice and wild-typelittermates (Fig. 7B). Because a clear defect in IL-2 production wasobserved by anti-CD3 stimulation of Ly9�/� CD4 T cells in vitro
FIGURE 6. B cell responses to LCMV in Ly9�/�
mice. A, Splenic germinal center B cells (B220�, IgD�,Fashigh, PNA�) were quantified by FACS at both day 8postinfection and day 15 (the peak of the anti-LCMVgerminal center response). Cells shown are gatedB220�IgD� B cells (p �� 0.05). B, Total plasma cells(CD138�, B220low) from the spleen were quantified byFACS (gated on total splenocytes) at the peak of theacute response (day 8 postinfection) (p �� 0.05).LCMV-specific IgG� plasma cells were quantified byELISPOT at day 8 postinfection (peak of the acute re-sponse) and at day 39 (memory phase) in both Ly9�/�
mice and Ly9�/� mice (p �� 0.05). Long-lived plasmacells are present in bone marrow and spleen at day 39postinfection (Ly9�/� n � 4, Ly9�/� n � 4 at each timepoint) (n � 2).
297The Journal of Immunology
by guest on May 14, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

(Fig. 2), we examined the ability of Ly9�/� LCMV-specific CD4T cells to produce IL-2 after stimulation with viral peptide (gp61).IL-2 production by Ly9�/� LCMV-specific CD4 T cells was nor-mal (Fig. 7C). However, LCMV infection is strongly immuno-genic, and therefore, the Ly9-dependent defect in IL-2 productionobserved earlier (Fig. 2) is apparently overcome by other costimu-latory pathways in the context of an LCMV infection (Fig. 7C).Finally, we examined the ability of Ly9�/� mice to generate mem-ory T cells, as memory is a key aspect of T cell biology (26).Numbers of LCMV-specific memory CD8 and CD4 T cells were
normal in Ly9�/� mice (Fig. 7, C and D). The fact that Ly9�/�
mice showed no defects in responding to LCMV indicates that Ly9is not responsible, or not solely responsible, for initiating SAPsignaling during viral infections.
DiscussionWe sought to explore the in vivo functions and mechanisms ofLy9, a receptor with poorly characterized biological functions.Therefore, we generated Ly9�/� mice to compare their phenotypeto that of SLAM�/� and SAP�/� mice. The objectives were to
FIGURE 7. T cell responses to LCMV in Ly9�/� mice. A, LCMV-specific effector CD8 T cells in spleen were quantified at day 8 postinfection by 5 hof stimulation with gp33–41 MHC class I peptide (immunodominant peptide) or gp276–286 and then analyzed by intracellular cytokine staining for IFN-�and TNF-� production. (p �� 0.05) B, LCMV-specific CD4 T cells were quantified at day 8 postinfection by 5 h of stimulation with the immunodominantgp61–80 I-Ab MHC class II peptide or subdominant np309–328 I-Ab peptide and then analyzed by intracellular cytokine staining for IFN-� and TNF-�production. (p �� 0.05) C, LCMV-specific CD4 T cells were examined for IL-2 and IFN-� production by intracellular cytokine staining after in vitrorestimulation for 5 h with gp61 peptide, done either at day 15 postinfection or day 39 postinfection. D, Memory CD8 T cell IFN-� and TNF-� responseswere measured at day 39 postinfection by intracellular cytokine staining after in vitro restimulation for 5 h with gp33 peptide (Ly9�/� n � 4, Ly9�/� n �4 at each time point.) (n � 2).
298 T CELL DEFECTS IN Ly9 (CD229)-DEFICIENT MICE
by guest on May 14, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

compare and contrast the functions of SLAM and Ly9 and to de-termine which, if any, of the defects observed in SAP�/� micemay be attributable to defective signaling downstream of Ly9. Thesingle common feature among Ly9�/�, SLAM�/�, and SAP�/�
mice is a Th2 defect, although the magnitude of the defect variesin the three mice (11, 12, 19). The reduction in IL-4 productionobserved in Ly9�/� T cells is modest and could be masked bycompensation of additional SLAM family members that performredundant functions. Our observations support the idea thatbecause SLAM and Ly9 signal through SAP, both receptors maycontribute to Th2 polarization additively. Accordingly, SAP�/�
mice exhibit a nearly complete Th2 defect, whereas SLAM�/� andLy9�/� mice show a partial defect (11, 12, 19).
Although Ly9 and SLAM share structural features and interactwith SAP, we found important differences in the phenotypes ofSLAM�/�, SAP�/�, and Ly9�/� mice. Ly9�/� mice do not ex-hibit macrophage defects resembling those found in SLAM�/�
mice (19) but not in SAP�/� mice. Furthermore, Ly9�/� mice donot display any overt defects in NKT cell development and do notexhibit defective responses to LCMV, as was observed in SAP�/�
mice (10–12, 16, 17). During the acute phase of LCMV infection,SAP�/� mice mount abnormally strong CD8 and CD4 T cell re-sponses, whereas Ly9�/� mice generate CD8 and CD4 T cellresponses to LCMV that are comparable to wild-type mice (10 –12). Ly9�/� mice also differ from SAP�/� mice in that theyexhibit no defects in T cell-dependent B cell responses toLCMV, including germinal center formation and the productionof long-lived plasma cells (10). These observations suggest thatadditional SLAM family receptors regulate diverse SAP func-tions. However, we cannot rule out the possibility that otherSLAM family receptors compensate for Ly9 deficiency and ob-scure any obvious defects.
Unexpectedly, the phenotype of Ly9�/� mice is distinct fromthat of SLAM�/� and SAP�/� mice. Our finding that Ly9�/� Tcells proliferate poorly and produce little IL-2 after in vitro stim-ulation with anti-CD3 suggests that Ly9 may play an importantrole in T cell activation. In contrast, a recent study proposed thatLy9 partially down-regulates human T cell responses (27). Thesestudies examined TCR signaling events after stimulation of T cellswith anti-CD3 and anti-Ly9. However, there is no evidence thatthe anti-Ly9 Abs used in that study are receptor agonists. Ouranalyses of murine Ly9�/� T cells demonstrate that Ly9 may en-hance T cell activation.
Presently, the relevant ligand for Ly9 in the context of T cellactivation is unknown. It is possible that homotypic interactionsbetween Ly9 on T cells and APCs promote T cell activation (Ref.28 and D. B. Graham, M. P. Bell, C. J. Huntoon, and D. J. McKean,submitted for publication). However, artificial APCs expressing Ly9do not enhance IL-2 production in CD4 T cells, which argues againsthomotypic Ly9 interactions being involved in T cell activation (ourunpublished observations). Alternatively, the ligand for Ly9 may notbe itself but an uncharacterized molecule on APCs or other leuko-cytes. Ultimately, it is not clear whether Ly9 is a ligand or a receptor.Further investigation will be required to elucidate the mechanisms ofLy9 signaling in the context of T cell activation.
Although several mechanistic questions remain, in vitro assayswith Ly9�/� T cells indicate a novel role for Ly9 in regulating Tcell proliferation, IL-2 production, and Th2 polarization. The Th2defect observed in Ly9�/� mice is common to SLAM�/� andSAP�/� mice; however, the T cell activation defect is unique toLy9�/� mice. Additionally, we found that Ly9 does not controlseveral of the biological responses previously shown to be regu-lated by SLAM and SAP. Unlike SLAM�/� mice, Ly9-deficiencydoes not lead to macrophage defects. Furthermore, Ly9�/� mice
differ from SAP�/� mice in that they generate NKT cells andmount appropriate T and B cell responses to LCMV. Our findingsreveal novel functions for Ly9 that distinguish it from the otherSLAM family receptors.
AcknowledgmentsWe thank Drs. Pam Schwartzberg and Jennifer Cannons (National Insti-tutes of Health, Bethesda, MD) for technical assistance and valuable dis-cussions, and we are grateful to Rafi Ahmed (Emory University, Atlanta,GA) for providing viral seed stocks. Mike Thompson (Mayo Clinic,Rochester, MN), Chuck Prickett and Fernando Vasquez (La Jolla Institutefor Allergy and Immunology) provided excellent animal care assistance.
DisclosuresThe authors have no financial conflict of interest.
References1. Romero, X., N. Zapater, M. Calvo, S. G. Kalko, M. A. de la Fuente, V. Tovar,
C. Ockeloen, P. Pizcueta, and P. Engel. 2005. CD229 (Ly9) lymphocyte cellsurface receptor interacts homophilically through its N-terminal domain and re-localizes to the immunological synapse. J. Immunol. 174: 7033–7042.
2. Mavaddat, N., D. W. Mason, P. D. Atkinson, E. J. Evans, R. J. C. Gilbert,D. I. Stuart, J. A. Fennelly, A. N. Barclay, S. J. Davis, and M. H. Brown.2000. Signaling lymphocytic activation molecule (CDw150) is homophilicbut self-associates with very low affinity. J. Biol. Chem. 275: 28100 –28109.
3. Sayos, J., M. Martin, A. Chen, M. Simarro, D. Howie, M. Morra, P. Engel, andC. Terhorst. 2001. Cell surface receptors Ly-9 and CD84 recruit the X-linkedlymphoproliferative disease gene product SAP. Blood 97: 3867–3874.
4. Li, C., C. Iosef, C. Y. Jia, V. K. M. Han, and S. S. C. Li. 2003. Dual functionalroles for the X-linked lymphoproliferative syndrome gene product SAP/SH2D1Ain signaling through the signaling lymphocyte activation molecule (SLAM) fam-ily of immune receptors. J. Biol. Chem. 278: 3852–3859.
5. Cannons, J. L., L. J. Yu, B. Hill, L. A. Mijares, D. Dombroski, K. E. Nichols,A. Antonellis, G. A. Koretzky, K. Gardner, and P. L. Schwartzberg. 2004. SAPregulates TH2 differentiation and PKC-�-mediated activation of NF-�B1. Immu-nity 21: 693–706.
6. Nichols, K. E., C. S. Ma, J. L. Cannons, P. L. Schwartzberg, and S. G. Tangye.2005. Molecular and cellular pathogenesis of X-linked lymphoproliferative dis-ease. Immunol. Rev. 203: 180–199.
7. Coffey, A. J., R. A. Brooksbank, O. Brandau, T. Oohashi, G. R. Howell,J. M. Bye, A. P. Cahn, J. Durham, P. Heath, P. Wray, et al. 1998. Host responseto EBV infection in X-linked lymphoproliferative disease results from mutationsin an SH2-domain encoding gene. Nat. Genet. 20: 129–135.
8. Sayos, J., C. Wu, M. Morra, N. Wang, X. Zhang, D. Allen, S. van Schaik,L. Notarangelo, R. Geha, M. G. Roncaralo, et al. 1998. The X-linked lympho-proliferative disease gene product SAP regulates signals induced through theco-receptor SLAM. Nature 395: 462–469.
9. Nichols, K. E., D. P. Harkin, S. Levitz, M. Krainer, K. A. Koloquist,C. Genovese, A. Bernard, M. Ferguson, L. Zuo, E. Snyder, et al. 1998. Inacti-vating mutations in an SH2 domain-encoding gene in X-linked lymphoprolifera-tive syndrome. Proc. Natl. Acad. Sci. USA 95: 13765–13770.
10. Crotty, S., E. N. Kersh, J. Cannons, P. L. Schwartzberg, and R. Ahmed. 2003.SAP is required for generating long-term humoral immunity. Nature 421:282–287.
11. Wu, C., K. B. Nguyen, G. C. Pien, N. Wang, C. Gullo, D. Howie,M. Rodriguez Sosa, M. J. Edwards, P. Borrow, A. R. Satoskar, et al. 2001. SAPcontrols T cell responses to virus and terminal differentiation of TH2 cells. Nat.Immunol. 2: 410–414.
12. Czar, M. J., E. N. Kersh, L. A. Mijares, G. Lanier, J. Lewis, G. Yap, A. Chen,A. Sher, C. S. Duckett, R. Ahmed, and P. L. Schwartzberg. 2001. Altered lym-phocyte responses and cytokine production in mice deficient in the X-linkedlymphoproliferative disease gene SH2D1A/DSHP/SAP. Proc. Natl. Acad. Sci.USA 98: 7449–7454.
13. Ma, C. S., N. J. Hare, K. E. Nichols, L. Dupre, G. Adolfi, M. G. Roncarolo,S. Adelstein, P. D. Hodgkin, and S. G. Tangye. 2005. Impaired humoral immu-nity in X-linked lymphoproliferative disease with defective IL-10 production byCD4� T cells. J. Clin. Invest. 115: 1049–1059.
14. Malbran, A., L. Belmonte, B. Ruibal-Ares, P. Bare, I. Massud, C. Parodi,M. Felippo, R. Hodinka, K. Haines, K. E. Nichols, and M. M. de Bracco. 2004.Loss of circulating CD27� memory B cells and CCR4� T cells occurring inassociation with elevated EBV loads in XLP patients surviving primary EBVinfection. Blood 103: 1625–1631.
15. Morra, M., D. Howie, M. S. Grande, J. Sayos, N. Wang, C. Wu, P. Engel, andC. Terhorst. 2001. X-linked lymphoproliferative disease: a progressive immuno-deficiency. Annu. Rev. Immunol. 19: 657–682.
16. Nichols, K. E., J. Hom, S. Y. Gong, A. Ganguly, C. S. Ma, J. L. Cannons,S. G. Tangye, P. L. Schwartzberg, G. A. Koretzky, and P. L. Stein. 2005. Reg-ulation of NKT cell development by SAP, the protein defective in XLP. Nat.Med. 11: 340–345.
17. Paquier, B., L. Yin, M. C. Fondaneche, F. Relouzat, C. Bloch-Queyrat,N. Lambert, A. Fischer, G. de Saint-Basile, and S. Latour. 2005. DefectiveNKT cell development in mice and humans lacking the adapter SAP, the
299The Journal of Immunology
by guest on May 14, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

X-linked lymphoproliferative syndrome gene product. J. Exp. Med. 201:695–701.
18. Chung, B., A. Aoukaty, J. Dutz, C. Terhorst, and R. Tan. 2005. Signaling lym-phocytic activation molecule-associated protein controls NKT cell functions.J. Immunol. 174: 3153–3157.
19. Wang, N., A. Satoskar, W. Faubion, D. Howie, S. Okamoto, S. Feske, C. Gullo,K. Clarke, M. Rodriquez Sosa, A. H. Sharpe, and C. Terhorst. 2004. The cellsurface receptor SLAM controls T cell and macrophage functions. J. Exp. Med.199: 1255–1264.
20. Matsuda, J. L., O. V. Naidenko, L. Gapin, T. Nakayama, M. Taniguchi,C. R. Wang, Y. Koezuka, and M. Kronenberg. 2000. Tracking the response ofnatural killer T cells to a glycolipid antigen using CD1d tetramers. J. Exp. Med.192: 741–754.
21. Slifka, M. K., R. Antia, J. K. Whitmire, and R. Ahmed. 1998. Humoral immunitydue to long-lived plasma cells. Immunity 8: 363–372.
22. Howie, D., F. S. Laroux, M. Morra, A. R. Satoskar, L. E. Rosas, W. A. Faubion,A. Julien, S. Rietdijk, A. J. Coyle, C. Fraser, and C. Terhorst. 2005. The SLAMfamily receptor Ly108 controls T cell and neutrophil functions. J. Immunol. 174:5931–5935.
23. Gadue, P., L. Yin, S. Jain, and P. L. Stein. 2004. Restoration of NK T celldevelopment in fyn-mutant mice by a TCR reveals a requirement for Fyn duringearly NK T cell ontogeny. J. Immunol. 172: 6093–6100.
24. Morra, M., R. A. Barrington, A. C. Abadia-Molina, S. Okamoto, A. Julien,C. Gullo, A. Kalsy, M. J. Edwards, G. Chen, R. Spolski, et al. 2005. Defective Bcell responses in the absence of SH2D1APNAS. Proc. Natl. Acad. Sci. USA 102:4819–484.
25. Al-Alem, U., C. Li, N. Forey, F. Relouzat, M. C. Fondaneche, S. V. Tavtigian,Z. Wang, S. Latour, and L. Yin. 2005. Impaired Ig class switch in mice deficientfor the X-linked lymphoproliferative disease gene Sap. Blood 106: 2069–2075.
26. Crotty, S., and R. Ahmed. 2004. Immunological memory in humans. Semin.Immunol. 16: 197–203.
27. Martin, M., J. M. Del Valle, I. N. Saborit, and P. Engel. 2005. Identificationof Grb2 as a novel binding partner of the signaling lymphocytic activationmolecule-associated protein binding receptor CD229. J. Immunol. 174:5977–5986.
28. Romero, X., N. Zapater, M. Calvo, S. G. Kalko, M. A. de la Fuente, V. Tovar,C. Ockeloen, P. Pizcueta, and P. Engel. 2005. CD229 (Ly9) lymphocyte cellsurface receptor interacts homophilically through its N-terminal domain and re-localizes to the immunological synapse. J. Immunol. 174: 7033–7042.
300 T CELL DEFECTS IN Ly9 (CD229)-DEFICIENT MICE
by guest on May 14, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from











![Predominant role of FcγRIII in the induction of ... · Recently, FcRI-deficient mice were produced [12, 15], and showed some func-tional impairments in IgG2a-immune complex–induced](https://img.pdfslide.us/doc/110x75/5f9ab31333807d26255f7f68/predominant-role-of-fcriii-in-the-induction-of-recently-fcri-deicient-mice.jpg)





![FcgRIIB Inhibits the Development of Atherosclerosis … · FcgRIIB Inhibits the Development of Atherosclerosis in Low-Density Lipoprotein Receptor-Deficient Mice ... [WT], 59-gCg](https://img.pdfslide.us/doc/110x75/5b9a5f3609d3f2c3468d1eea/fcgriib-inhibits-the-development-of-atherosclerosis-fcgriib-inhibits-the-development.jpg)











