Embed Size (px)
Citation preview

Local Circuit for the Computationof Object Approach by an Identified
Visual Neuron in the Locust
F. CLAIRE RIND* AND PETER J. SIMMONS
Neurobiology Department, School of Neurosciences and Psychiatry, University of Newcastleupon Tyne, Newcastle upon Tyne, NE2 4HH, United Kingdom
ABSTRACTThe lobula giant movement detector (LGMD) neuron in the locust visual system is part of
a motion-sensitive pathway that detects objects approaching on a collision course. Here weshow that the retinotopic units presynaptic to the LGMD make synapses directly with eachother and these synapses are immediately adjacent to their outputs onto the LGMD. Synapsesoccur along the fine dendrites of the LGMD in the distal lobula, often in large numbers andcompletely covering the LGMD processes. Gamma aminobutyric acid (GABA) was eliminatedas a possible neurotransmitter at these synapses when immunogold-tagged monoclonalGABA antibody did not specifically label the afferent processes. We used a histochemicalmethod to demonstrate that acetylcholine esterase, the enzyme that hydrolyses acetylcholineat cholinergic synapses, was present in the synaptic clefts between the retinotopic units andalong the membrane of the LGMD. It is well established that acetylcholine has both excitatoryand inhibitory effects and we propose that these retinotopic units excite the LGMD, butinhibit each other; and that the synapses form the substrate for a critical race betweenexcitation caused by edges moving out over successive photoreceptors, and inhibitionspreading laterally. This results in the selective response to objects approaching on a collisioncourse. J. Comp. Neurol. 395:405–415, 1998. r 1998 Wiley-Liss, Inc.
Indexing terms: motion detection; invertebrate; acetylcholine; presynaptic inhibition
In the locust, two identified neurons, the lobula giantmovement detector (LGMD, O’Shea and Williams, 1974)and the postsynaptic descending contralateral movementdetector (DCMD, Rowell, 1971; Rind, 1984), respond selec-tively to the images of an object approaching towards itseye (Rind and Simmons, 1992). The strongest response isgiven to an object approaching on a collision course (Rindand Simmons, 1992, 1997; Hatsopoulos et al., 1995; Judgeand Rind, 1997). Selectivity for approach is achieved byresponding to the increasing extent and velocity of imageedges moving over the retina of one eye (Simmons andRind, 1992). The retinotopic units presynaptic to theLGMD inhibit each other (Simmons and Rind, 1992;O’Shea and Rowell, 1975; Pinter, 1977), and we proposedthat this results in a critical race between excitationcaused by edges moving over successive photoreceptors,and inhibition spreading laterally (Simmons and Rind,1992). A neural network based on the input organisation ofthe LGMD neuron confirmed the effectiveness of this race,between excitation, passing down the network, and inhibi-tion directed laterally, in determining a response that isselective for approaching objects (Rind and Bramwell,1996). For a strong response in the artificial network,
excitation must overtake the lateral inhibition, whichoccurs when an object approaches on a collision coursewith the artificial eye. In this case, the images of approach-ing edges move with increasing velocity over the eye andexpand rapidly. Inhibition then arrives at each retinotopicinput unit after excitation has already excited the unitsufficiently for it to generate a response in the outputelement of the model, equivalent to the LGMD. We showhere that in the locust, there is an appropriate anatomicalsubstrate for this inhibition; the afferents make synapseswith their neighbours locally right at the sites where theyalso provide synaptic input onto the LGMD. Synapsesalways occur in back-to-back pairs each with two postsyn-aptic elements: the LGMD and the neighbouring afferent.For this anatomical circuit to operate in the same way as
Grant sponsor: Royal Society; Grant sponsor: BBSRC*Correspondence to: F.C. Rind, Neurobiology Department, School of
Neurosciences and Psychiatry, University of Newcastle upon Tyne, New-castle upon Tyne, NE2 4HH, United Kingdom. E-mail: [email protected]
Received 21 October 1997; Revised 4 February 1998; Accepted 6 February1998
THE JOURNAL OF COMPARATIVE NEUROLOGY 395:405–415 (1998)
r 1998 WILEY-LISS, INC.
the model circuit, the afferents should excite the LGMDbut inhibit each other. When we stained for two conven-tional neurotransmitters, we obtained positive resultsonly for acetylcholine, a neurotransmitter that, in insects,is well established to mediate both excitatory and inhibi-tory effects depending on the receptor involved (for areview see Trimmer, 1995).
MATERIALS AND METHODS
Visual stimuli
The visual stimuli were generated by a VSG2 board(Cambridge Research Systems, Rochester, UK), fitted in amicrocomputer and were displayed on a Kikusui monitor.The screen was parallel to the long axis of the locust, withits centre 60 mm from the locust’s left eye. Screen dimen-sions were 120 by 100 mm. The background intensity was4.12 3 1022 mW/cm2 measured at the screen; objects whichmoved against this background had an intensity of 1.64 31022 mW/cm2. The refresh rate for the screen was 200 Hz, arefresh rate unlikely to cause appreciable flicker on alocust’s photoreceptors (Pinter, 1972).
Electrophysiology and neurobiotin staining
Intracellular recordings were made from the dendritesof the LGMD in the left lobula. The LGMD was identifiedby correlating, 1-for-1, spikes recorded extracellularlyfrom the axon of the DCMD in the contralateral nervecord, with those recorded intracellularly in the optic lobe.To reveal the fine morphological detail of the LGMDneuron in the optic lobe, the neuron was injected withneurobiotin and then reacted with a monoclonal anti-
biotin antibody tagged with Cy3 (Jackson Immunore-search Laboratories, West Grove, PA) according to themethod of Mesce et al. (1993). The optic lobe was thenviewed from behind using a confocal microscope and astack of 50 3-µm, optical sections taken through the opticlobe.
LGMD identification by electron microscopy
Initially, the LGMD was identified by injecting it withhexamminecobaltic ions, which selectively damage mito-chondria (Littlewood and Simmons, 1988), but we foundthat the consistent position of the LGMD profiles in thelobula and their lightly staining cytoplasm made thempossible to identify without intracellular marking. Subse-quently, the LGMD profiles were identified using thesecriteria.
GABA immunocytochemistry
To test for GABA immunoreactivity, the optic lobes werefixed in ice-cold 0.1 M phosphate-buffered gluteraldehydesupplemented with 10% sucrose for 4 hours, postfixed inOsmium followed by 2% Uranyl acetate. The tissue wasdehydrated in a graded acetone series and embedded inaraldite 502 (Agar Scientific, Stansted, UK). Sections werecut and mounted on nickel slot grids coated with pioloform.The material was etched with 2% aqueous periodic acid (20minutes) then washed in saturated sodium metaperiodate(30 minutes) before being treated with 5% normal goatserum in Tris buffer (pH 7, 30 minutes). The sections werethen washed briefly in Tris buffer with 1% bovine serumalbumin before being transferred for 2 hours to rabbit
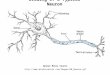
Fig. 1. Morphology of the lobula giant movement detector (LGMD)in the left lobula. The neuron was injected with neurobiotin accordingto the method of Mesce et al. (1993) then viewed from behind using aconfocal microscope. A shows the complete stack of images (50 3 3 mmsections) through the lobula. The dendritic fan of the LGMD in the
distal lobula is indicated by an asterisk. B: The top 20 images havebeen removed to reveal previously obscured detail. The dendritic fan ofthe LGMD in the distal lobula (A) clearly lies in register above theproximal branches (B). An arrow indicates the area shown in Figure 3.Scale bars 5 100 µm.
406 F.C. RIND AND P.J. SIMMONS
anti-GABA serum (Sigma, St. Louis, MO) in Tris buffer(pH 7) at a dilution of 1:800. After washing in Tris bufferwith 1% bovine serum albumin at pH 7 then pH 8.3, thesections were incubated for 1 hour in 15-nm gold-labelled,goat anti-rabbit antibody diluted 1:20 in Tris at pH 8.3.After washing, they were stained with lead citrate andurayl acetate and viewed on the electron microscope.
AChE histochemistry
To reveal the distribution of acetylcholinesterase, amethod modified from Navaratnam and Lewis (1970) wasused. Tissue was fixed at 4oC for 4 hours in 2.5% glutaral-
dehyde, with 0.1 M phosphate buffer (pH 7.2) and 10%sucrose. It was washed in phosphate buffer (pH 5.6) for 5minutes, incubated for 4 hours in 13 mM (0.188 g/50 ml)acetylthiocholine iodide, 2 3 1024 M (0.0035 g/50 ml)ethopropazine hydrochloride, 18 mM (0.225 g/50 ml) cop-per sulphate, 33 mM (0.124 g/50 ml) glycine, and 25 mM(0.338 g/50 ml) succinate buffer (pH 5.6 with 0.1 M HCl).Tissue was washed three times in buffer before beingtransferred to a 1% ammonium ferric sulphide solution (10minutes). The tissue was kept at 4oC to improve tissuepreservation. The tissue was postfixed in 1% osmiumtetroxide at pH 5.6.
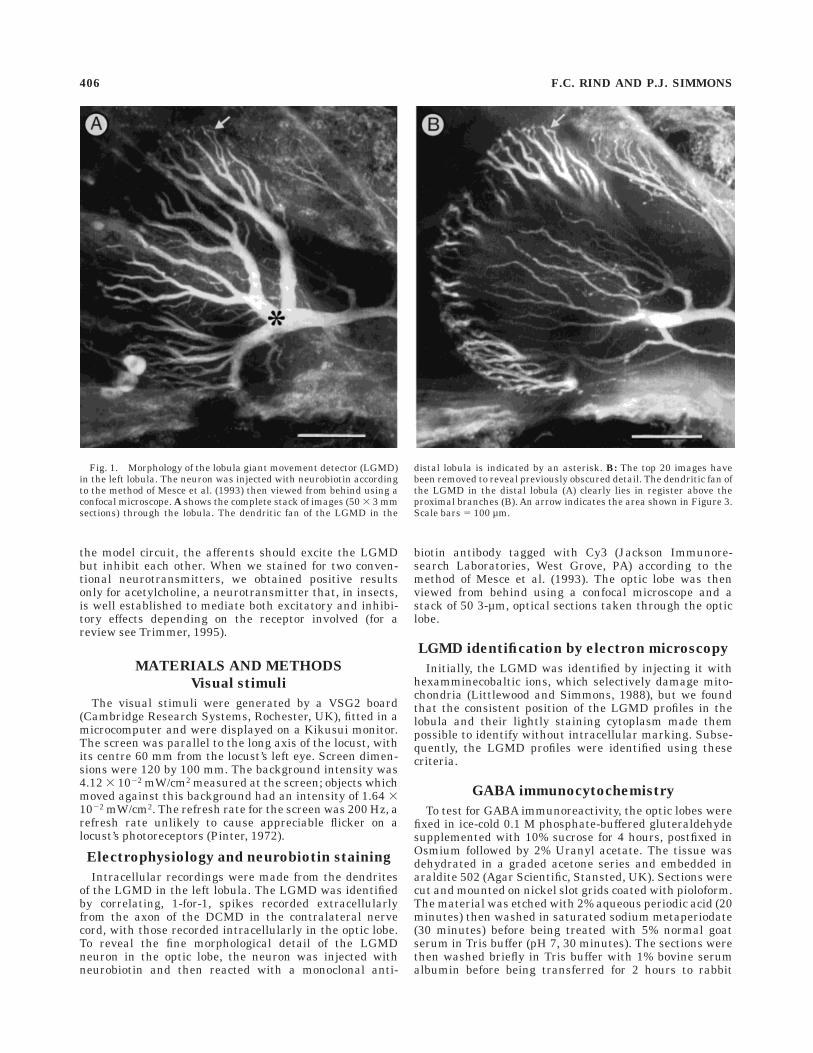
Fig. 2. Intracellular responses to images moving posteriorly overthe eye. Below each set of four recordings, the stimulus configurationis shown. A: One or two bars moved over the eye: (i) left bar (L) alone;(ii) left bar followed by right bar (R); (iii and iv) left bar moving alonewith dc currents injected into lobula giant movement detector (LGMD)as indicated. The left bar subtended 67 by 9.5o at the eye; and the rightbar 9.5 by 9.5o. Bars moved at an angular velocity over the eye of 160o/s
at the times indicated on each recording. B: Each moving stimulus wasa 9.5o square alone (i) or accompanied by two rows of stripes, eachsubtending 3.8o (ii) or 34o (iii, iv) at the eye. In iv, dc current wasinjected into the LGMD. Stimulus movement started at the beginningof each recording and continued at 62o/second for 0.6 seconds. The tworows of stripes were separated by 20o.
A LOCAL CIRCUIT FOR SIGNALLING OBJECT APPROACH 407
RESULTS
The locust LGMD has extensive fan-shaped dendrites inthe third optic neuropile, the lobula (Fig. 1A), whichreceive excitatory input (excitatory postsynaptic poten-tials, epsps) from small-field, retinotopic units projectingfrom the second optic neuropile, the medulla (O’Shea andRowell, 1976). The identity of the medullary neuronsdirectly presynaptic to the LGMD has not yet been con-firmed by paired recordings, although Osorio has character-ised neurons within the medulla which share responsefeatures with the LGMD (Osorio, 1987; James and Osorio,1996).
The initial evidence for lateral inhibition among retino-topic units presynaptic to the LGMD was provided byexperiments which predated the demonstration that theLGMD was selective for movements in depth. In theseexperiments, the number of spikes generated by theLGMD in response to a small, translating visual targetwas reduced by the presence of drifting gratings, and thestrength of the inhibition declined as the distance betweenthe gratings and the target increased (O’Shea and Rowell,1975; Rowell, et al., 1977). Intracellular recordings fromthe LGMD revealed inhibitory postsynaptic potentials(ipsps) if overall luminance changed rapidly, or if a wide-field grating moved rapidly, but not in response to wide-field, drifting gratings with the same contrast frequencyused to demonstrate lateral inhibition (less than 10–20 Hz;Rowell et al., 1977). These ipsps are thought to be medi-ated by synapses onto a more proximal arbor of the LGMD(Fig. 1B) than the epsps.
We extended this earlier work in order to investigatewhether stimulation of a small retinal field activates thepathways responsible for lateral inhibition in a way thatwould be consistent with a critical race between excitationand inhibition. In seven experiments, we demonstrateddirectly that stimulation of one part of the LGMD recep-
tive field decreases the response to stimulation of anotherpart without directly inhibiting the LGMD itself. In theseexperiments, the eye was stimulated by movement of twovertically orientated bars. In the experiment shown inFigure 2A, when only the right bar moved (Fig. 2A, i), theLGMD was excited with a 5-mV depolarising potentialupon which rode 10 spikes (mean of four repetitions, 10.25spikes). If movement of the right bar was preceded bymovement of the left (Fig. 2A, ii), the amplitude of thedepolarising potential and the number of spikes in theLGMD was reduced (mean number of spikes, 4.5). Thistype of inhibition was not dependent on the direction ofmovement of the bars, or on the relative lengths of the twobars. When responses to movement of the left bar alonewere examined, no sign of any synaptic activity caused bythis movement was apparent by the time the right barwould normally have moved, even when depolarising (Fig.2A, iii) or hyperpolarizing (Fig. 2A, iv) d.c. currents wereinjected to enhance any psps. In a second stimulus configu-ration, lateral movement of a small square caused astrong, maintained depolarisation of the LGMD (Fig. 2B,i). When the central square was accompanied in itsmovement by two rows of identical spots (Fig. 2B, ii–iv),the initial excitation of the LGMD was enhanced, butsubsequently reduced to a barrage of excitatory postsynap-tic potentials well before the end of movement. Increasingthe heights of the two rows of spots emphasised thetransient nature of the initial excitation and reduced theexcitatory postsynaptic activity (compare ii and iii in Fig.2B). Injection of d.c. currents again failed to reveal ipsps(Fig. 2B, iv). This second stimulus configuration is similarto that used in extracellular recordings by Rowell et al.(1977), but the extent of the gratings we used was smallerand is less likely to activate wide-field pathways, whichprobably accounts for the initial transitory excitation inour experiments.
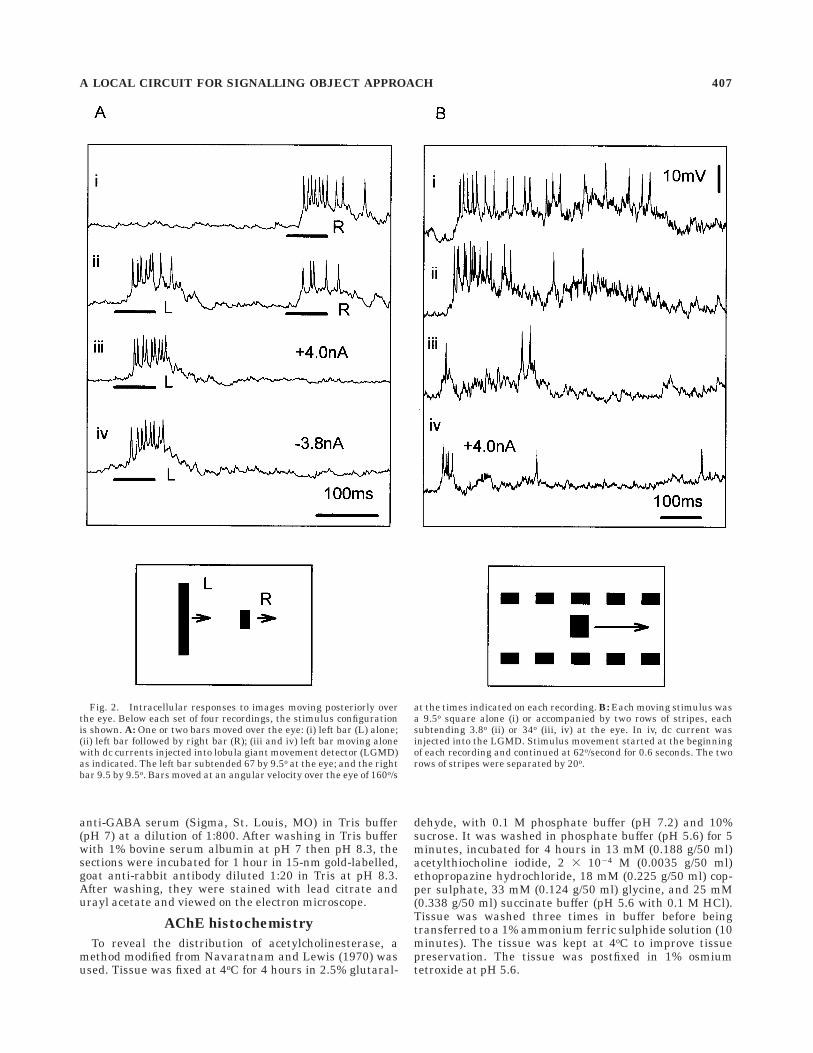
Fig. 3. Electron micrographs showing a section through the den-drites of a lobula giant movement detector (LGMD) neuron (indicatedby an asterisk) which has been injected with hexamminecobalticchloride. A shows a low power view of an affected dendrite with many
enlarged mitochondria with disrupted internal cristae. B showsanother LGMD process at higher magnification. Scale bar 5 1 µm in A,200 nm in B.
408 F.C. RIND AND P.J. SIMMONS
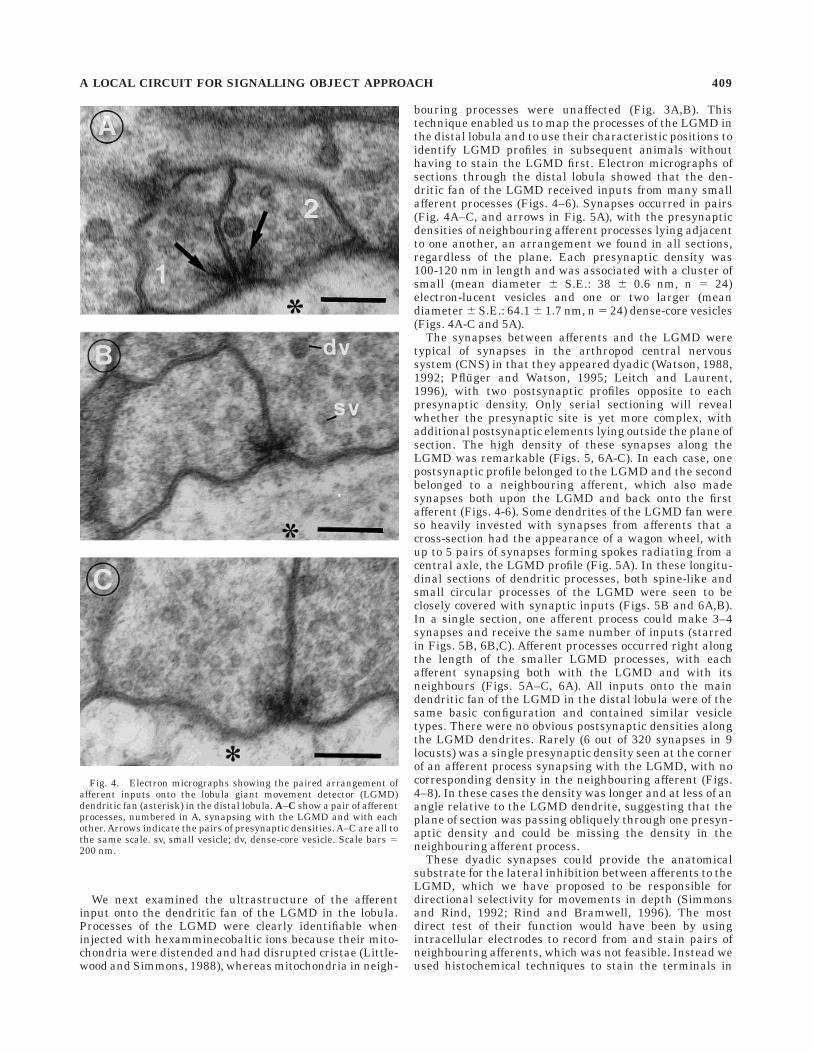
We next examined the ultrastructure of the afferentinput onto the dendritic fan of the LGMD in the lobula.Processes of the LGMD were clearly identifiable wheninjected with hexamminecobaltic ions because their mito-chondria were distended and had disrupted cristae (Little-wood and Simmons, 1988), whereas mitochondria in neigh-
bouring processes were unaffected (Fig. 3A,B). Thistechnique enabled us to map the processes of the LGMD inthe distal lobula and to use their characteristic positions toidentify LGMD profiles in subsequent animals withouthaving to stain the LGMD first. Electron micrographs ofsections through the distal lobula showed that the den-dritic fan of the LGMD received inputs from many smallafferent processes (Figs. 4–6). Synapses occurred in pairs(Fig. 4A–C, and arrows in Fig. 5A), with the presynapticdensities of neighbouring afferent processes lying adjacentto one another, an arrangement we found in all sections,regardless of the plane. Each presynaptic density was100-120 nm in length and was associated with a cluster ofsmall (mean diameter 6 S.E.: 38 6 0.6 nm, n 5 24)electron-lucent vesicles and one or two larger (meandiameter 6 S.E.: 64.1 6 1.7 nm, n 5 24) dense-core vesicles(Figs. 4A-C and 5A).
The synapses between afferents and the LGMD weretypical of synapses in the arthropod central nervoussystem (CNS) in that they appeared dyadic (Watson, 1988,1992; Pfluger and Watson, 1995; Leitch and Laurent,1996), with two postsynaptic profiles opposite to eachpresynaptic density. Only serial sectioning will revealwhether the presynaptic site is yet more complex, withadditional postsynaptic elements lying outside the plane ofsection. The high density of these synapses along theLGMD was remarkable (Figs. 5, 6A-C). In each case, onepostsynaptic profile belonged to the LGMD and the secondbelonged to a neighbouring afferent, which also madesynapses both upon the LGMD and back onto the firstafferent (Figs. 4-6). Some dendrites of the LGMD fan wereso heavily invested with synapses from afferents that across-section had the appearance of a wagon wheel, withup to 5 pairs of synapses forming spokes radiating from acentral axle, the LGMD profile (Fig. 5A). In these longitu-dinal sections of dendritic processes, both spine-like andsmall circular processes of the LGMD were seen to beclosely covered with synaptic inputs (Figs. 5B and 6A,B).In a single section, one afferent process could make 3–4synapses and receive the same number of inputs (starredin Figs. 5B, 6B,C). Afferent processes occurred right alongthe length of the smaller LGMD processes, with eachafferent synapsing both with the LGMD and with itsneighbours (Figs. 5A–C, 6A). All inputs onto the maindendritic fan of the LGMD in the distal lobula were of thesame basic configuration and contained similar vesicletypes. There were no obvious postsynaptic densities alongthe LGMD dendrites. Rarely (6 out of 320 synapses in 9locusts) was a single presynaptic density seen at the cornerof an afferent process synapsing with the LGMD, with nocorresponding density in the neighbouring afferent (Figs.4–8). In these cases the density was longer and at less of anangle relative to the LGMD dendrite, suggesting that theplane of section was passing obliquely through one presyn-aptic density and could be missing the density in theneighbouring afferent process.
These dyadic synapses could provide the anatomicalsubstrate for the lateral inhibition between afferents to theLGMD, which we have proposed to be responsible fordirectional selectivity for movements in depth (Simmonsand Rind, 1992; Rind and Bramwell, 1996). The mostdirect test of their function would have been by usingintracellular electrodes to record from and stain pairs ofneighbouring afferents, which was not feasible. Instead weused histochemical techniques to stain the terminals in
Fig. 4. Electron micrographs showing the paired arrangement ofafferent inputs onto the lobula giant movement detector (LGMD)dendritic fan (asterisk) in the distal lobula. A–C show a pair of afferentprocesses, numbered in A, synapsing with the LGMD and with eachother. Arrows indicate the pairs of presynaptic densities. A–C are all tothe same scale. sv, small vesicle; dv, dense-core vesicle. Scale bars 5200 nm.
A LOCAL CIRCUIT FOR SIGNALLING OBJECT APPROACH 409
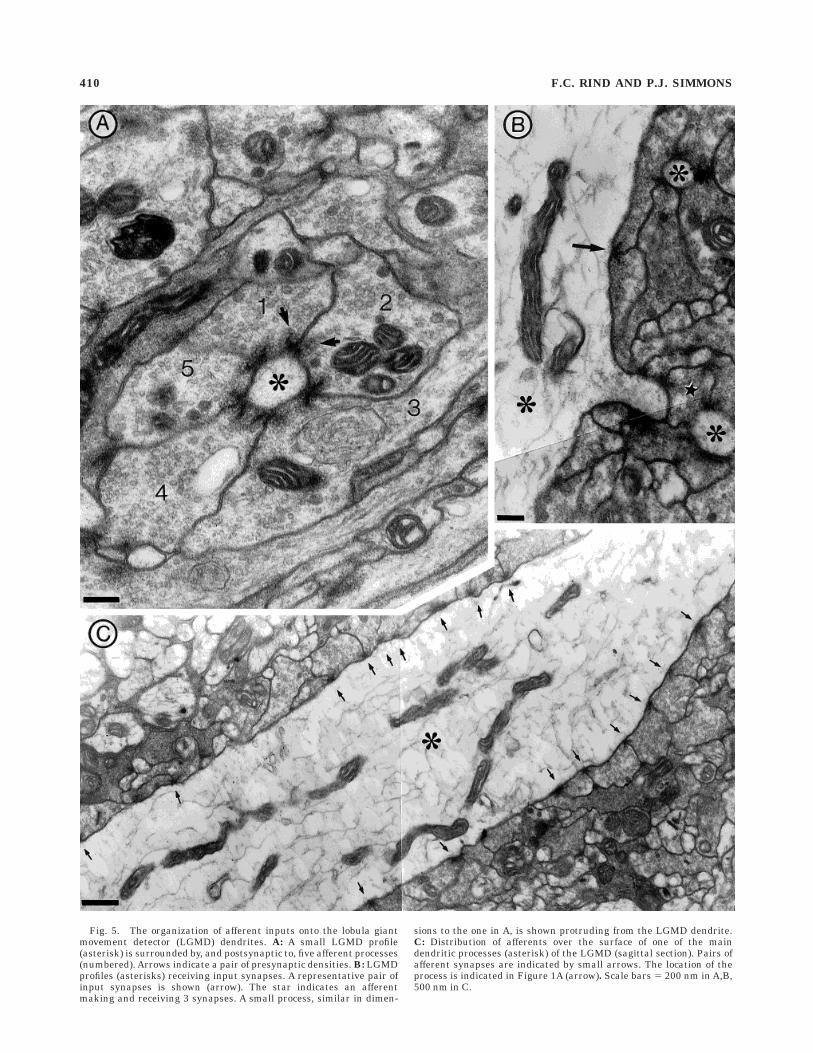
Fig. 5. The organization of afferent inputs onto the lobula giantmovement detector (LGMD) dendrites. A: A small LGMD profile(asterisk) is surrounded by, and postsynaptic to, five afferent processes(numbered). Arrows indicate a pair of presynaptic densities. B: LGMDprofiles (asterisks) receiving input synapses. A representative pair ofinput synapses is shown (arrow). The star indicates an afferentmaking and receiving 3 synapses. A small process, similar in dimen-
sions to the one in A, is shown protruding from the LGMD dendrite.C: Distribution of afferents over the surface of one of the maindendritic processes (asterisk) of the LGMD (sagittal section). Pairs ofafferent synapses are indicated by small arrows. The location of theprocess is indicated in Figure 1A (arrow). Scale bars 5 200 nm in A,B,500 nm in C.
410 F.C. RIND AND P.J. SIMMONS
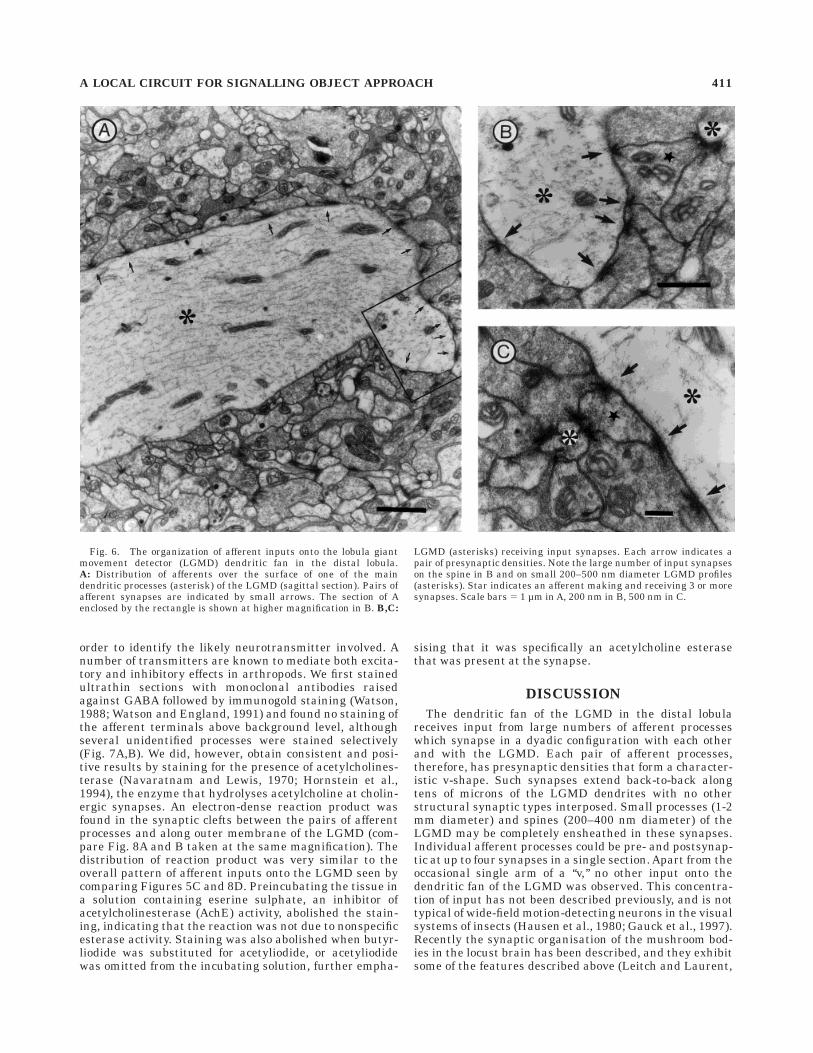
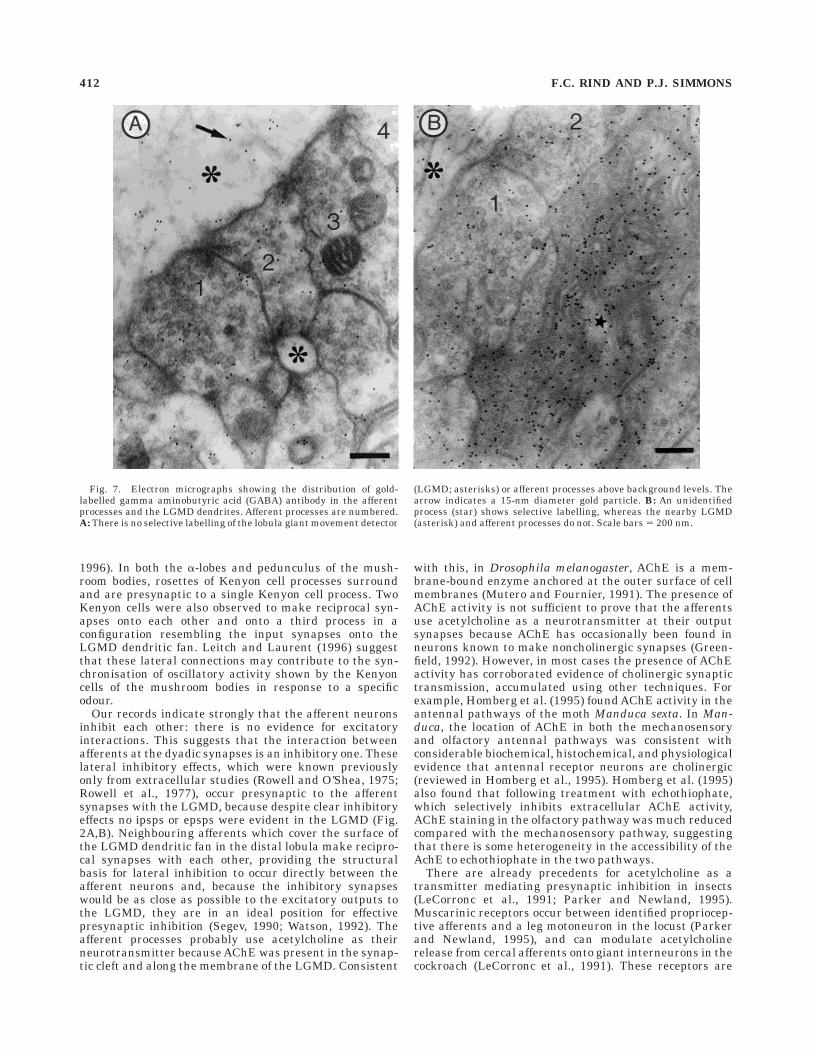
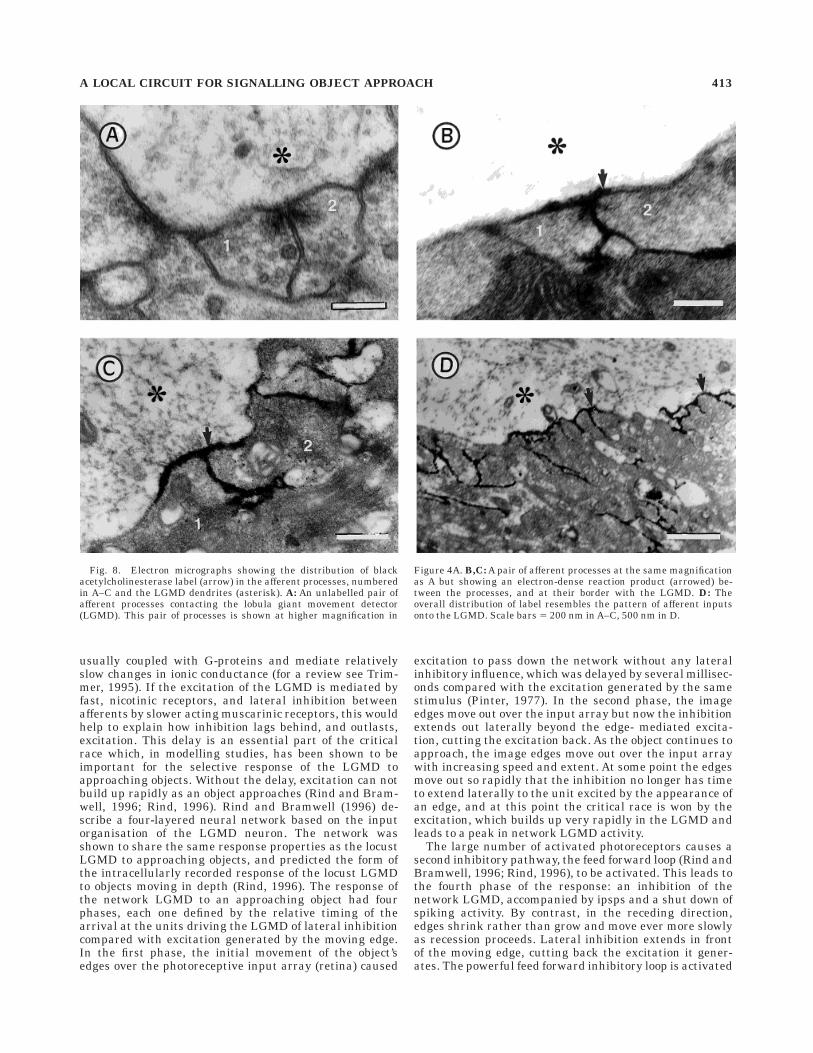
order to identify the likely neurotransmitter involved. Anumber of transmitters are known to mediate both excita-tory and inhibitory effects in arthropods. We first stainedultrathin sections with monoclonal antibodies raisedagainst GABA followed by immunogold staining (Watson,1988; Watson and England, 1991) and found no staining ofthe afferent terminals above background level, althoughseveral unidentified processes were stained selectively(Fig. 7A,B). We did, however, obtain consistent and posi-tive results by staining for the presence of acetylcholines-terase (Navaratnam and Lewis, 1970; Hornstein et al.,1994), the enzyme that hydrolyses acetylcholine at cholin-ergic synapses. An electron-dense reaction product wasfound in the synaptic clefts between the pairs of afferentprocesses and along outer membrane of the LGMD (com-pare Fig. 8A and B taken at the same magnification). Thedistribution of reaction product was very similar to theoverall pattern of afferent inputs onto the LGMD seen bycomparing Figures 5C and 8D. Preincubating the tissue ina solution containing eserine sulphate, an inhibitor ofacetylcholinesterase (AchE) activity, abolished the stain-ing, indicating that the reaction was not due to nonspecificesterase activity. Staining was also abolished when butyr-liodide was substituted for acetyliodide, or acetyliodidewas omitted from the incubating solution, further empha-
sising that it was specifically an acetylcholine esterasethat was present at the synapse.
DISCUSSION
The dendritic fan of the LGMD in the distal lobulareceives input from large numbers of afferent processeswhich synapse in a dyadic configuration with each otherand with the LGMD. Each pair of afferent processes,therefore, has presynaptic densities that form a character-istic v-shape. Such synapses extend back-to-back alongtens of microns of the LGMD dendrites with no otherstructural synaptic types interposed. Small processes (1-2mm diameter) and spines (200–400 nm diameter) of theLGMD may be completely ensheathed in these synapses.Individual afferent processes could be pre- and postsynap-tic at up to four synapses in a single section. Apart from theoccasional single arm of a ‘‘v,’’ no other input onto thedendritic fan of the LGMD was observed. This concentra-tion of input has not been described previously, and is nottypical of wide-field motion-detecting neurons in the visualsystems of insects (Hausen et al., 1980; Gauck et al., 1997).Recently the synaptic organisation of the mushroom bod-ies in the locust brain has been described, and they exhibitsome of the features described above (Leitch and Laurent,
Fig. 6. The organization of afferent inputs onto the lobula giantmovement detector (LGMD) dendritic fan in the distal lobula.A: Distribution of afferents over the surface of one of the maindendritic processes (asterisk) of the LGMD (sagittal section). Pairs ofafferent synapses are indicated by small arrows. The section of Aenclosed by the rectangle is shown at higher magnification in B. B,C:
LGMD (asterisks) receiving input synapses. Each arrow indicates apair of presynaptic densities. Note the large number of input synapseson the spine in B and on small 200–500 nm diameter LGMD profiles(asterisks). Star indicates an afferent making and receiving 3 or moresynapses. Scale bars 5 1 µm in A, 200 nm in B, 500 nm in C.
A LOCAL CIRCUIT FOR SIGNALLING OBJECT APPROACH 411
1996). In both the a-lobes and pedunculus of the mush-room bodies, rosettes of Kenyon cell processes surroundand are presynaptic to a single Kenyon cell process. TwoKenyon cells were also observed to make reciprocal syn-apses onto each other and onto a third process in aconfiguration resembling the input synapses onto theLGMD dendritic fan. Leitch and Laurent (1996) suggestthat these lateral connections may contribute to the syn-chronisation of oscillatory activity shown by the Kenyoncells of the mushroom bodies in response to a specificodour.
Our records indicate strongly that the afferent neuronsinhibit each other: there is no evidence for excitatoryinteractions. This suggests that the interaction betweenafferents at the dyadic synapses is an inhibitory one. Theselateral inhibitory effects, which were known previouslyonly from extracellular studies (Rowell and O’Shea, 1975;Rowell et al., 1977), occur presynaptic to the afferentsynapses with the LGMD, because despite clear inhibitoryeffects no ipsps or epsps were evident in the LGMD (Fig.2A,B). Neighbouring afferents which cover the surface ofthe LGMD dendritic fan in the distal lobula make recipro-cal synapses with each other, providing the structuralbasis for lateral inhibition to occur directly between theafferent neurons and, because the inhibitory synapseswould be as close as possible to the excitatory outputs tothe LGMD, they are in an ideal position for effectivepresynaptic inhibition (Segev, 1990; Watson, 1992). Theafferent processes probably use acetylcholine as theirneurotransmitter because AChE was present in the synap-tic cleft and along the membrane of the LGMD. Consistent
with this, in Drosophila melanogaster, AChE is a mem-brane-bound enzyme anchored at the outer surface of cellmembranes (Mutero and Fournier, 1991). The presence ofAChE activity is not sufficient to prove that the afferentsuse acetylcholine as a neurotransmitter at their outputsynapses because AChE has occasionally been found inneurons known to make noncholinergic synapses (Green-field, 1992). However, in most cases the presence of AChEactivity has corroborated evidence of cholinergic synaptictransmission, accumulated using other techniques. Forexample, Homberg et al. (1995) found AChE activity in theantennal pathways of the moth Manduca sexta. In Man-duca, the location of AChE in both the mechanosensoryand olfactory antennal pathways was consistent withconsiderable biochemical, histochemical, and physiologicalevidence that antennal receptor neurons are cholinergic(reviewed in Homberg et al., 1995). Homberg et al. (1995)also found that following treatment with echothiophate,which selectively inhibits extracellular AChE activity,AChE staining in the olfactory pathway was much reducedcompared with the mechanosensory pathway, suggestingthat there is some heterogeneity in the accessibility of theAchE to echothiophate in the two pathways.
There are already precedents for acetylcholine as atransmitter mediating presynaptic inhibition in insects(LeCorronc et al., 1991; Parker and Newland, 1995).Muscarinic receptors occur between identified propriocep-tive afferents and a leg motoneuron in the locust (Parkerand Newland, 1995), and can modulate acetylcholinerelease from cercal afferents onto giant interneurons in thecockroach (LeCorronc et al., 1991). These receptors are
Fig. 7. Electron micrographs showing the distribution of gold-labelled gamma aminobutyric acid (GABA) antibody in the afferentprocesses and the LGMD dendrites. Afferent processes are numbered.A: There is no selective labelling of the lobula giant movement detector
(LGMD; asterisks) or afferent processes above background levels. Thearrow indicates a 15-nm diameter gold particle. B: An unidentifiedprocess (star) shows selective labelling, whereas the nearby LGMD(asterisk) and afferent processes do not. Scale bars 5 200 nm.
412 F.C. RIND AND P.J. SIMMONS
usually coupled with G-proteins and mediate relativelyslow changes in ionic conductance (for a review see Trim-mer, 1995). If the excitation of the LGMD is mediated byfast, nicotinic receptors, and lateral inhibition betweenafferents by slower acting muscarinic receptors, this wouldhelp to explain how inhibition lags behind, and outlasts,excitation. This delay is an essential part of the criticalrace which, in modelling studies, has been shown to beimportant for the selective response of the LGMD toapproaching objects. Without the delay, excitation can notbuild up rapidly as an object approaches (Rind and Bram-well, 1996; Rind, 1996). Rind and Bramwell (1996) de-scribe a four-layered neural network based on the inputorganisation of the LGMD neuron. The network wasshown to share the same response properties as the locustLGMD to approaching objects, and predicted the form ofthe intracellularly recorded response of the locust LGMDto objects moving in depth (Rind, 1996). The response ofthe network LGMD to an approaching object had fourphases, each one defined by the relative timing of thearrival at the units driving the LGMD of lateral inhibitioncompared with excitation generated by the moving edge.In the first phase, the initial movement of the object’sedges over the photoreceptive input array (retina) caused
excitation to pass down the network without any lateralinhibitory influence, which was delayed by several millisec-onds compared with the excitation generated by the samestimulus (Pinter, 1977). In the second phase, the imageedges move out over the input array but now the inhibitionextends out laterally beyond the edge- mediated excita-tion, cutting the excitation back. As the object continues toapproach, the image edges move out over the input arraywith increasing speed and extent. At some point the edgesmove out so rapidly that the inhibition no longer has timeto extend laterally to the unit excited by the appearance ofan edge, and at this point the critical race is won by theexcitation, which builds up very rapidly in the LGMD andleads to a peak in network LGMD activity.
The large number of activated photoreceptors causes asecond inhibitory pathway, the feed forward loop (Rind andBramwell, 1996; Rind, 1996), to be activated. This leads tothe fourth phase of the response: an inhibition of thenetwork LGMD, accompanied by ipsps and a shut down ofspiking activity. By contrast, in the receding direction,edges shrink rather than grow and move ever more slowlyas recession proceeds. Lateral inhibition extends in frontof the moving edge, cutting back the excitation it gener-ates. The powerful feed forward inhibitory loop is activated
Fig. 8. Electron micrographs showing the distribution of blackacetylcholinesterase label (arrow) in the afferent processes, numberedin A–C and the LGMD dendrites (asterisk). A: An unlabelled pair ofafferent processes contacting the lobula giant movement detector(LGMD). This pair of processes is shown at higher magnification in
Figure 4A. B,C: A pair of afferent processes at the same magnificationas A but showing an electron-dense reaction product (arrowed) be-tween the processes, and at their border with the LGMD. D: Theoverall distribution of label resembles the pattern of afferent inputsonto the LGMD. Scale bars 5 200 nm in A–C, 500 nm in D.
A LOCAL CIRCUIT FOR SIGNALLING OBJECT APPROACH 413

early during recession when large numbers of photorecep-tive units are activated, and the loop cuts back all but thefirst few milliseconds of excitation in the network LGMD.
Our investigation suggests that the critical race betweenexcitation and inhibition occurs over the surface of theLGMD neuron itself, in the lobula. This phenomenon is ofgeneral significance because most of the mechanisms thataccount for directional selectivity involve lateral inhibition/excitation passing between retinotopic units which isdelayed relative to the excitation passing down throughthe network (Barlow and Levick, 1965; Poggio and Rei-chardt, 1976; Emerson and Gerstein, 1977; Torre andPoggio, 1978; Van Santen and Sperling, 1985; Adelson andBergen, 1985; Osorio, 1986; Vaney, 1990; Borg-Grahamand Grzywacs, 1992; Douglas and Martin, 1992; Bouzer-doum, 1993). For these proposed circuits to operate cor-rectly, precision is required both in the location of synapsesthat connect different elements, and in the timing of theirresponses (O’Carroll et al., 1996). These requirementscould be met by the compact microcircuits that provide theinput to the LGMD neuron. This is a very simple andprecise way of achieving a selective response using thecontrasting properties of two receptor types for the sametransmitter activated by a common input. As an exampleof a process creating a selective response to one particularimage feature, this microcircuit may be a common mecha-nism for the selective response to motion direction or toedge orientation.
At present, there is no consensus as to how theseselective processes arise in the visual system (fly: Poggioand Reichardt, 1976; Bouzerdoum, 1993; locust: Osorio,1986; salamander: Borg-Graham and Grzywacs, 1992;rabbit: Barlow and Levick, 1965; Torre and Poggio, 1978;Vaney, 1990; He and Masland, 1997; cat: Reid et al., 1991;Douglas and Martin, 1992; human: Van Santen and Sper-ling, 1985; Adelson and Bergen, 1985). Whole cell record-ings from cortical orientation selective cells in the cat haverevealed that in any particular cell, selectivity can becreated by excitatory, inhibitory, or by a combination ofboth mechanisms (Volgushev et al., 1993). In insects, too,both facilitatory and inhibitory mechanisms have beenproposed to underlie directional selectivity in the sameanimal (Schmid and Bulthoff, 1988; Franceschini et al.,1989; Bouzerdoum, 1993). In part, this may be becausedirectionally selective motion detection is not a one stageprocess, but emerges gradually through successive synap-tic interaction as has been suggested previously for thelocust (Rind, 1990) and the fly (Borst and Egelhaaf, 1990).Direction selectivity of the LGMD for motion in depth is anexample of a selective response and demonstrates howmuch can be done at a single synaptic level within thevisual system.
LITERATURE CITED
Adelson, E.H. and J.R. Bergen (1985) Spatio-temporal energy models forthe perception of motion. J. Opt. Soc. Am. A 2:284–299.
Barlow, H.B. and W.R. Levick (1965) The mechanism of directionallyselective units in the rabbit’s retina. J. Physiol. 178:477–504.
Borg-Graham, L.J. and N.M. Grzywacz (1992) A model of the directionselectivity circuit in the retina. In T. McKenna, J. Davis, and S.F.Zornetzer (eds): Single Neuron Computation. Orlando, Florida: Aca-demic Press, pp. 347–375.
Borst, A. and M. Egelhaaf (1990) Direction selectivity of blowfly motion-sensitive neurons is computed in a two stage process. Proc. Natl. Acad.Sci. USA 87:9363–9367.
Bouzerdoum, A. (1993) The elementary movement detection mechanism ininsect vision. Phil. Trans. R. Soc. Lond. B 339:375–384.
Douglas, R.J. and K.A.C. Martin (1992) Exploring cortical microcircuits: Acombined anatomical, physiological and computational approach. In T.McKenna, J. Davis, and SF. Zornetzer (eds): Single Neuron Computa-tion. Orlando, Florida: Academic Press, pp. 381–412.
Emerson, R.C. and G.L. Gerstein (1977) Simple striate neurons in the cat.II. Mechanisms underlying directional asymmetry and directionalselectivity. J. Neurophysiol. 40:136–155.
Franceschini, N., A. Riehle, and A. Le Nestour (1989) Directionally selectivemotion detection by insect neurons. In D. G. Stavenga and R.C. Hardie(eds): Facets of Vision. Berlin: Springer, pp. 360–390.
Gauck, V., M. Egelhaaf, and A. Borst (1997) Synapse distribution on VCH,an inhibitory, motion-sensitive interneuron in the fly visual system. J.Comp. Neurol. 381:489–499.
Greenfield, S.A. (1992) Acetylcholinesterase as a modulatory neuroproteinand its influence on motor control. In A. Shafferman and B. Velan (eds):Multidisciplinary Approaches to Cholinesterase Functions. New York:Plenun Press, pp. 233–242.
Hatsopoulos, N., F. Gabbiani, and G. Laurent (1995) Elementary computa-tion of object approach by a wide-field visual neuron. Science 270:1000–1003.
Hausen, K., K. Wolburg-Buchholz, and W.A. Ribi (1980) The synapticorganization of visual interneurons in the lobula complex of flies. CellTissue Res. 208:371–387.
He, S. and R.H. Masland (1997) Retinal direction selectivity after targetedlaser ablation of starburst amacrine cells. Nature 389:378–382.
Homberg, U., S.G. Hoskins, and J.G. Hildebrand (1995) Distribution ofacetylcholinesterase activity in the deutocerebrum of the sphinx mothManduca sexta. Cell Tissue Res. 279:249–259.
Hornstein, E.P., D.L. Sambursky, and S.C. Chamberlin (1994) Histochemi-cal localization of acetylcholinesterase in the lateral eye and brain ofLimulus polyphemus: Might acetylcholine be a neurotransmitter forlateral inhibition in the lateral eye? Vis. Neurosci. 11:989–1001.
James, A.C. and D. Osorio (1996) Characterisation of columnar neuronsand visual signal processing in the medulla of the locust optic lobe bysystem identification techniques. systems approach.J. Comp. Physiol.178:183–199.
Judge, S.J. and F.C. Rind (1997) The locust DCMD, A movement detectingneurone tightly tuned to collision trajectories. J. Exp. Biol. 200:2209–2216.
LeCorronc, H., B. Lapied, and B. Hue (1991) M2 like presynaptic receptorsmodulate acetylcholine release in the cockroach (Periplaneta ameri-cana) central nervous system. J. Insect Physiol. 37:647–652.
Leitch, B. and G. Laurent (1996) GABAergic synapses in the antennal lobeand mushroom body of the locust olfactory system. J. Comp. Neurol.372:487–514.
Littlewood, P.M.H. and P.J. Simmons (1988) Hexamminecobaltic chlorideprovides a simple method for marking neurones for electron microscopy.Brain Res. 445:165–170.
Mesce, K.A., K.A. Klukas, and T.C. Brelje (1993) Improvements for theanatomical characterization of insect neurones in whole mount: The useof cyanine-derived fluorophores and laser scanning confocal microscopy.Cell Tissue Res. 271:381–397.
Mutero, A. and D. Fournier (1991) Drosophila acetylcholinesterase struc-ture. In J. Massoulie, F. Bacou, E. Barnard, A. Chatonnet, B. P. Doctor,and D.M. Quin (eds): Cholinesterases. Structure, Function, Geneticsand Cell Biology. Washington: American Chemical Society, pp. 141–145.
Navaratnam, V. and P.R. Lewis (1970) Cholinesterase-containing neuronesin the spinal cord of the rat. Brain Res. 18:411–425.
O’Carroll, D.C., N.J. Bidwell, S.B. Laughlin, and E.J. Warrant (1996) Insectmotion detectors matched to visual ecology. Nature 382:63–66.
O’Shea, M. and C.H.F. Rowell (1975) Protection from habituation by lateralinhibition. Nature 254:53–55.
O’Shea, M. and C.H.F. Rowell (1976) Neuronal basis of a sensory analyzer,the acridid movement detector system. II. Response decrement, conver-gence, and the nature of the excitatory afferents to the fan-likedendrites of the LGMD. J. Exp. Biol. 65:289–308.
O’Shea, M. and J.L.D. Williams (1974) The anatomy and output connec-tions of a locust visual interneurone: The lobula giant movementdetector (LGMD) neurone.J. Comp. Physiol. 91:257–266.
Osorio, D. (1986) Directionally selective cells in the locust medulla. J. CompPhysiol. 159:841–847.
Osorio, D. (1987) The temporal properties of non-linear, transient cells inthe locust medulla. J. Comp. Physiol. 161:431–440.
414 F.C. RIND AND P.J. SIMMONS

Parker, D. and P.I. Newland (1995) Cholinergic synaptic transmissionbetween proprioceptive afferents and a hind leg motor neuron in thelocust. J. Neurophysiol. 73:586–594.
Pfluger, H.-J. and A.H.D Watson (1995) GABA and glutamate-like immuno-reactivity at synapses received by dorsal unpaired median neurones inthe abdominal nerve cord of the locust. Cell Tissue Res. 280:325–333.
Pinter, R.B. (1972) Frequency and time domain properties of retinular cellsof the desert locust (Shistocerca gregaria) and the house cricket (Achetadomesticus). J. Comp. Physiol. 77: 383–397.
Pinter, R.B. (1977) Visual discrimination between small objects and largetextured backgrounds. Nature 270:429–431.
Poggio, T. and W. Reichardt (1976) Visual control of orientation behavior inthe fly. Part ii. towards the underlying neural interactions. Quart. Rev.Biophys. 9:377–438.
Reid, R.C., R.E. Soodak, and R.M. Shapley (1991) Directional selectivityand spatiotemporal structure of simple cells in cat striate cortex. J.Neurophysiol. 66:505–529.
Rind, F.C. (1984) A chemical synapse between two motion detectingneurones in the locust brain. J. Exp. Biol. 110:143–167.
Rind, F.C. (1990) Identification of directionally selective motion-detectingneurones in the locust lobula and their synaptic connections with anidentified descending neurone. J. Exp. Biol. 149:21–43.
Rind, F.C. (1996) Intracellular characterization of neurons in the locustbrain signalling impending collision. J. Neurophysiol. 75: 986–995.
Rind, F.C. and D.I. Bramwell (1996) A neural network based on the inputorganisation of an identified neuron signalling impending collision. J.Neurophysiol. 75:967–985.
Rind, F.C. and P. J. Simmons (1992) Orthopteran DCMD neuron: Are-evaluation of responses to moving objects. I. Selective responses toapproaching objects. J. Neurophysiol. 68:1654–1666.
Rind, F.C. and P.J. Simmons (1997) Signalling of object approach by theDCMD neuron of the locust. J. Neurophysiol. 77:1029–1033.
Rowell, C.H.F. (1971) The orthopteran descending movement detector(DMD) neurones: A characterisation and review. Z. Vgl. Physiol.73:167–194.
Rowell, C.H.F., M. O’Shea, and J.L.D. Williams (1977) Neuronal basis of asensory analyzer, the acridid movement detector system. IV. Thepreference for small field stimuli. J. Exp. Biol. 68: 157–185.
Schmid, A. and H. Bulthoff (1988) Using neuropharmacology to distinguishbetween excitatory and inhibitory movement detection mechanisms inthe fly Calliphora erythrocephala. Biol. Cyber. 59: 71–80.
Segev, I. (1990) Computer study of presynaptic inhibition controlling thespread of action potentials into axon terminals. J. Neurophysiol.63:987–998.
Simmons, P.J. and F.C. Rind (1992) Orthopteran DCMD neuron: A re-evaluation of responses to moving objects. II. Critical cues for detectingapproaching objects. J. Neurophysiol. 68:1667–1682.
Torre, V. and T. Poggio (1978) A synaptic mechanism possibly underlyingdirectional selectivity to motion. Proc. Roy. Soc. Lond. B. 202:409–416.
Trimmer, B.A. (1995) Current excitement from insect muscarinic receptors.Trends Neurosci. 18:104–111.
Vaney, D.I. (1990) The mosaic of amacrine cells in the mammalian retina. InN. Osborne and J. Chader (eds): Progress in Retinal Research, Vol. 9.Oxford: Pergamon Press, pp. 49–100.
Van Santen, J.P.H. and G. Sperling (1985) Elaborated Reichardt detectors.J. Opt. Soc. Am. 2:300–320.
Volgushev, M., X. Pei, T.R. Vidyasagar, and T. Creutzfeldt (1993) Excitationand inhibition in orientation selectivity of cat visual cortical neuronsrevealed by whole-cell recordings in vivo. Vis. Neurosci. 10:1151–1155.
Watson, A.H.D. (1988) Antibodies against GABA and glutamate labelneurones with morphologically distinct synaptic vesicles in locustcentral nervous system. Neuroscience 26:33–44.
Watson, A.H.D. (1992) Presynaptic modulation of sensory afferents in theinvertebrate and vertebrate nervous system. Comp. Biochem. Physiol.103:227–239.
Watson, A.D.H. and R.C.D. England (1991) The distribution of and interac-tions between GABA-immunoreactive and non-immunoreactive pro-cesses presynaptic to campaniform sensilla on the trochanter of thelocust leg. Cell. Tissue Res. 226:331–341.
A LOCAL CIRCUIT FOR SIGNALLING OBJECT APPROACH 415