Embed Size (px)
Citation preview

Live-Cell Fluorescent Imaging of Membrane or Mitochondrion Transfer
between Connected Cells in Culture
A. Cselenyák, E. Pankotai, A. Csordás, L. Kiss, and Z. Lacza
Department of Human Physiology and Clinical Experimental Research, Semmelweis University, Tűzoltó utca 37-47., H1094, Budapest, Hungary
Exchange of cellular elements has become the focus of intense research which revealed that cells actively interact with each other and various parts can be exchanged through direct cell-to-cell contacts. Live cell confocal microscopy is a state-of-the-art tool for experiments designed to elucidate these communications, pushing the capabilities of fluorescent microscopy to its limits. It is often required to use multiple staining without any trace of overlap to label the various cell populations or organelles which must remain unmixed for long periods of time in co-culture. Another important feature is the spatial and temporal resolution of live cell imaging as structures like tunneling nanotubes are barely visible with the highest optical magnification of light microscopy. Since cells interact in 3-dimensions, one must consider 3D imaging in order to differentiate between cell fusion or cellular overlap. Furthermore, time-lapse imaging is required to observe cell-to-cell interactions for hours or even days which bring a new dimension of problems into live cell microscopy such as focus shift and environmental control on the stage of the microscope. In most cases all of the above specifications must be met to gain convincing evidence for a certain question resulting in complex experimental setups where the exclusion of technical artifacts is of paramount importance but cannot be easily achieved.
Keywords mitochondria; cell fusion; tunneling nanotubes; FRAP; confocal; time lapse; video microscopy
1. Introduction
Cell-to-cell communication is a crucial requirement for the development and maintenance of multicellular organisms. This communication may occur via different mechanisms, such as synapses, transport of small molecules through gap junctions or releasing different messengers like hormones or growth factors [1-3]. In 2004 a new route of cell-to-cell communication was discovered between rat pheochromocytoma (PC12) cells [4]. The structures of this new route were named ‘tunneling nanotubes’ (TNTs) and have a diameter of 50-200 nm and a length of up to several cell diameters. This is a dinamically forming and constantly changing nanotubular network, in which TNTs connect cells at their closest distance and they do not attach to the surface but hover in the medium. Based on the morphological criteria defined for TNTs between PC12 cells, the presence of these membrane bridges has been observed in many different cell types, such as bone marrow derived mesenchymal stem cells, endothelial progenitor cells, cardiomyocytes [5-7], macrophages, dendritic or Jurkat T cells [8, 9].
The importance of direct cell-to-cell interactions was also proved in tissue regeneration processes. Wang et al isolated mesenchymal stem cells (MSCs) from rat bone marrow, amplified them in vitro and cultured together with rat cardiomyocytes in direct co-culture – when cells were grown together –, or in indirect co-culture – when cells were separated by an insert in the culture dish – or in conditioned culture – when MSCs were fed for one week with the media collected from the direct co-culture. A week later immunofluorescence staining was positive against alpha-actin, desmin and cardiac troponin T only in the direct co-culture, but not in the indirect co-culture, nor in the conditioned culture group. They concluded that direct cell-to-cell contact between MSCs and adult cardiomyocytes, but not the soluble signaling molecules are obligatory in the differentiation of MSCs into cardiomyocytes or smooth muscle cells [10].
During short term intercellular cell-to-cell interactions neighboring cells can exchange membrane, mitochondria and other cytoplasmic components [11]. The advantage of developing direct cell-to-cell membrane bridges is the selective communication even over long distances. In agreement with this assumption, the common feature of all TNT-like structures is the transfer of cargo between connected cells.
1.1Mitochondria and organelle transfer
Mitochondria are essential organelles in plant and animal cells and play a key role in processes such as oxidative phosphorylation, aerobic metabolism of glucose and fat, calcium signaling, and apoptosis [12, 13]. These organelles play essential roles in aging, in the determination of lifespan and a variety of heritable and acquired diseases are linked to mitochondrial dysfunction. Spees and his colleagues reported in 2005 that mitochondria were more dynamic than had been thought earlier as mitochondria or mtDNA were shown to move between cells [14]. The active transfer from adult stem cells and somatic cells could rescue aerobic respiration in mammalian cells with nonfunctional mitochondria. In the experiments they used cells pretreated with ethidium bromide to mutate and deplete mtDNA. These cells became
Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.)
764 ©FORMATEX 2010
______________________________________________

incapable of aerobic respiration and growth (A549 ρ° cells), unless kept in a permissive medium containing uridine and pyruvate to supplement anaerobic glycolysis [15, 16]. The A549 ρ° cells were co-cultured with either adult non-hemopoietic stem/progenitor cells from human bone marrow or with skin fibroblasts. The co-cultures produced clones of rescued A549 ρ° cells with functional mitochondria. In the same year Koyanagi et al. showed the formation of nanotubes between endothelial progenitor cells and cardiomyocytes [5]. They investigated the formation of intercellular connections, which may allow the transport of macromolecular structures between labeled adult human endothelial progenitor cells (EPCs) and neonatal rat cardiomyocytes (CM) expressing green fluorescent protein (GFP) in a co-culture system. To determine whether the nanotubular structures allowed the transport of organelles, they labeled CM with a vital mitochondrial dye (MitoTracker). Using time lapse video microscopy, they observed the transport of stained complexes between CM and EPC resulting in the uptake of MitoTracker-positive structures in EPC. Mesenchymal stem cells and cardiomyocytes also develop nanotubes and mesenchymal stem cells (MSCs) can donate mitochondria to cardiomyocytes restoring their energetic state [17, 18]. Light and fluorescence microscopy and scanning electron microscopy as well showed that cells formed intercellular contacts and the exchange of cytosolic elements could be observed. The characterization of TNTs revealed that these filaments contain actin, myosin and in some cases, microsomes or mitochondria [14]. TNTs facilitate the intercellular transfer of vesicles of endocytic origin as well as of membrane components and cytoplasmic components. Direct evidence for the intercellular exchange of cargo via TNT-like structures was obtained by video-microscopic studies. Employing fluorescent dyes, organelles of the endosomal/lysosomal system [4], and mitochondria [5] were shown to travel uni-directionally along TNT-like structures between cells over long distances. The transferred endocytic vesicles were shown to fuse with their counterparts in the target cells [4], and to provide signaling information from one single cell to a larger community. In addition to the transfer of organelles, Ca2+ signals, plasma membrane components can transfer along TNT-like bridges into the connected cells.
1.2 Cell fusion
Cell fusion was observed in several in vitro co-culture studies and in some cases also in in vivo experiments following tissue injury [19-21]. Cell fusion occurs between healthy cells [17] and also between transplanted stem cells and host cells after tissue injury [22-24]. Stem cells in vitro are able to fuse with each other and also with other differentiated cells, which was proved by co-culturing GFP-labeled, puromycin resistant neuronal stem cells and hygromycin-resistant embryonic stem cells; resulting in GFP-positive, puromycin and hygromycin-resistant tetraploid cells [25]. MSCs can also fuse with cardiomyoblasts as shown in a flow cytometry analysis; in which co-culture of fluorescent Vybrant DiO-labeled cardiomyoblasts and Vybrant DiD-labeled MSCs provided double labeled cells, although some of these cells had a size of a single cell [17]. Cselenyák et al. also found double labeled cells without double nuclei in the co-culture that may result from direct cell-to-cell interactions, when cells are able to exchange membrane parts and Vybrant dye molecules. The Vybrant fluorescence dyes are high molecular weight, lipophilic stains that are not able to transfer between cells through gap junctions [11]. Interpretation of cell fusion is difficult, because of the variations between the used cell cultures and different detection techniques [26-28]. On the contrary, several studies have shown that cell fusion may result in transdifferentiation, which is a favorable explanation of increased tissue regeneration after stem cell transplantation [29], and wide range of evidence supports the physiological importance of stem cell fusion in general [30-32].
2. Fluorescent Imaging Techniques for the Investigation of Cell-to-Cell Interactions
The investigations of the abovementioned phenomena necessitate accurate and stable staining techniques for the temporal and spatial differentiation of cellular membranes and mitochondria. Unfortunately these criteria cannot be met easily with the available staining methods.
2.1 Labeling of mitochondria
2.1.1. Staining with MitoTracker
MitoTracker dyes are available in several fluorescent colors and were described as selective markers for mitochondria. MitoTracker Green (excitation/emission: 490/516nm), a cell-permeant dye, is non fluorescence in aqueous solutions and becomes fluorescent only on sequestration and association with lipids within the mitochondria without requiring oxidation or reduction. The dye contains a mildly thiol-reactive chloromethyl moiety that keeps it inside the mitochondrion and essentially measures cellular mitochondria content and distribution. MitoTracker Red CM-H2XRos is a red fluorescence X-rosamine derivative (excitation/emission: 579/599nm) that is also cell-permeable and sequesters in the mitochondria, but does not emit fluorescence unless it is oxidized, implying that actively respiring mitochondria are required to obtain fluorescence [33].
Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.)
©FORMATEX 2010 765
______________________________________________
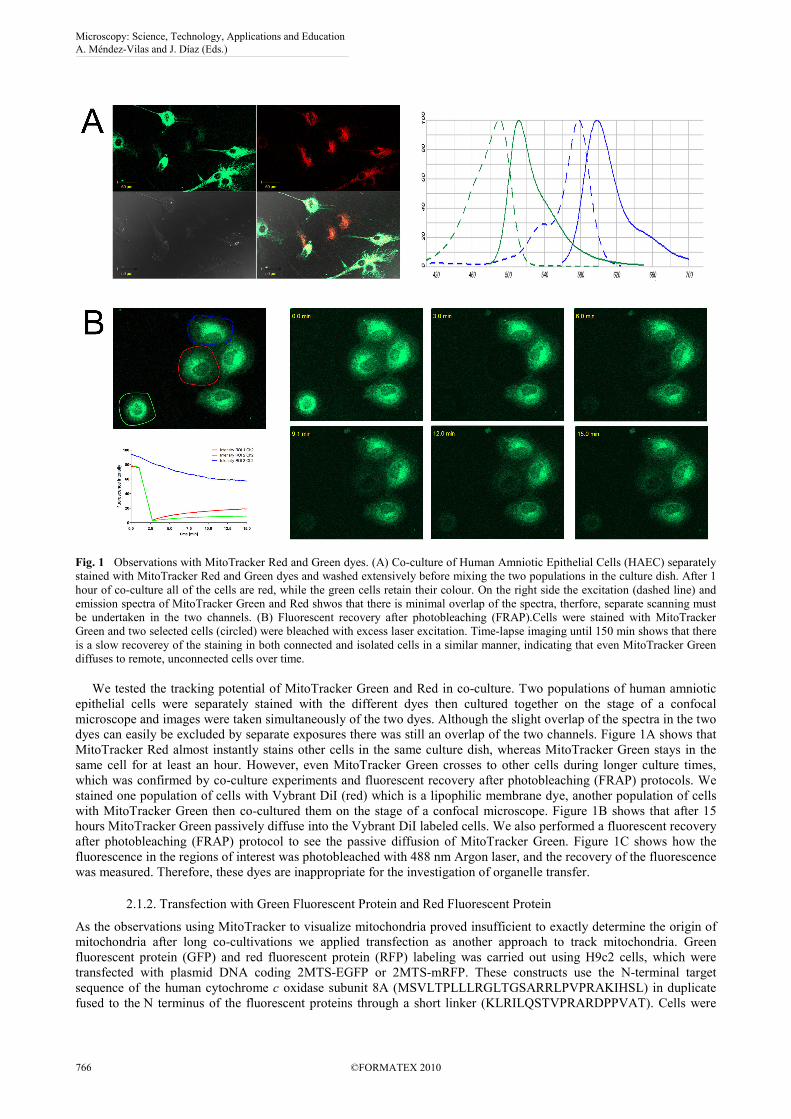
Fig. 1 Observations with MitoTracker Red and Green dyes. (A) Co-culture of Human Amniotic Epithelial Cells (HAEC) separately stained with MitoTracker Red and Green dyes and washed extensively before mixing the two populations in the culture dish. After 1 hour of co-culture all of the cells are red, while the green cells retain their colour. On the right side the excitation (dashed line) and emission spectra of MitoTracker Green and Red shwos that there is minimal overlap of the spectra, therfore, separate scanning must be undertaken in the two channels. (B) Fluorescent recovery after photobleaching (FRAP).Cells were stained with MitoTracker Green and two selected cells (circled) were bleached with excess laser excitation. Time-lapse imaging until 150 min shows that there is a slow recoverey of the staining in both connected and isolated cells in a similar manner, indicating that even MitoTracker Green diffuses to remote, unconnected cells over time. We tested the tracking potential of MitoTracker Green and Red in co-culture. Two populations of human amniotic epithelial cells were separately stained with the different dyes then cultured together on the stage of a confocal microscope and images were taken simultaneously of the two dyes. Although the slight overlap of the spectra in the two dyes can easily be excluded by separate exposures there was still an overlap of the two channels. Figure 1A shows that MitoTracker Red almost instantly stains other cells in the same culture dish, whereas MitoTracker Green stays in the same cell for at least an hour. However, even MitoTracker Green crosses to other cells during longer culture times, which was confirmed by co-culture experiments and fluorescent recovery after photobleaching (FRAP) protocols. We stained one population of cells with Vybrant DiI (red) which is a lipophilic membrane dye, another population of cells with MitoTracker Green then co-cultured them on the stage of a confocal microscope. Figure 1B shows that after 15 hours MitoTracker Green passively diffuse into the Vybrant DiI labeled cells. We also performed a fluorescent recovery after photobleaching (FRAP) protocol to see the passive diffusion of MitoTracker Green. Figure 1C shows how the fluorescence in the regions of interest was photobleached with 488 nm Argon laser, and the recovery of the fluorescence was measured. Therefore, these dyes are inappropriate for the investigation of organelle transfer.
2.1.2. Transfection with Green Fluorescent Protein and Red Fluorescent Protein
As the observations using MitoTracker to visualize mitochondria proved insufficient to exactly determine the origin of mitochondria after long co-cultivations we applied transfection as another approach to track mitochondria. Green fluorescent protein (GFP) and red fluorescent protein (RFP) labeling was carried out using H9c2 cells, which were transfected with plasmid DNA coding 2MTS-EGFP or 2MTS-mRFP. These constructs use the N-terminal target sequence of the human cytochrome c oxidase subunit 8A (MSVLTPLLLRGLTGSARRLPVPRAKIHSL) in duplicate fused to the N terminus of the fluorescent proteins through a short linker (KLRILQSTVPRARDPPVAT). Cells were
Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.)
766 ©FORMATEX 2010
______________________________________________
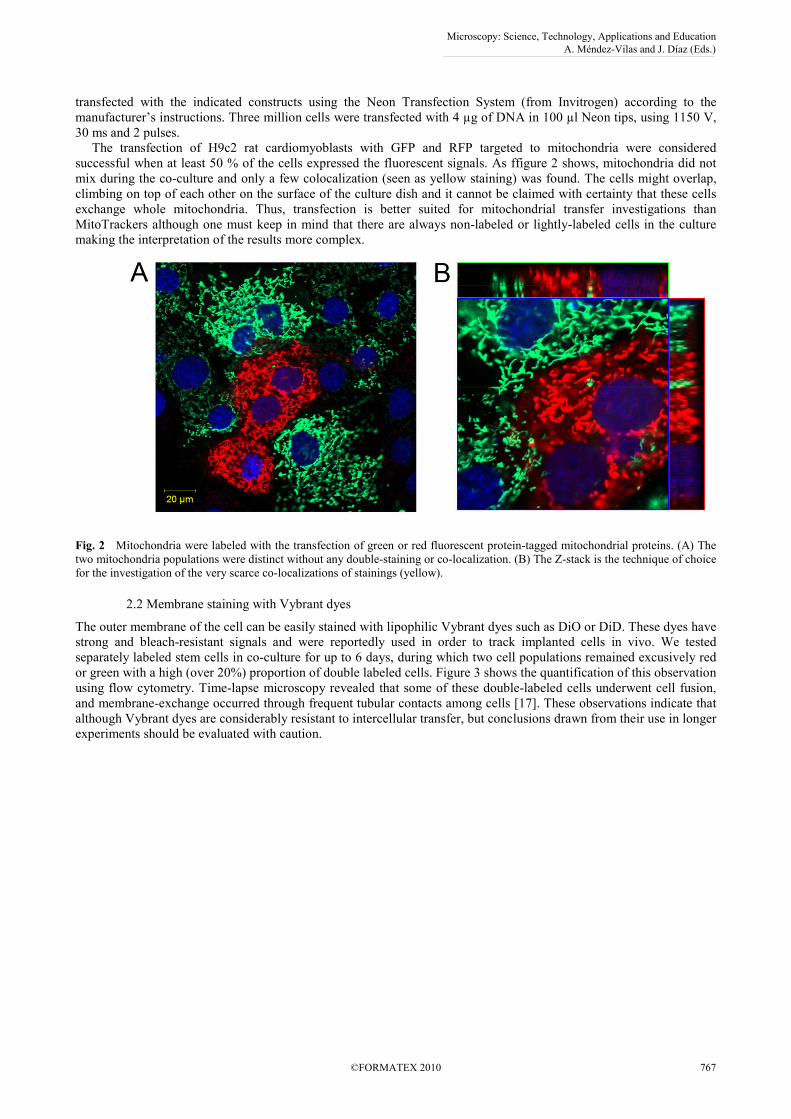
transfected with the indicated constructs using the Neon Transfection System (from Invitrogen) according to the manufacturer’s instructions. Three million cells were transfected with 4 µg of DNA in 100 µl Neon tips, using 1150 V, 30 ms and 2 pulses. The transfection of H9c2 rat cardiomyoblasts with GFP and RFP targeted to mitochondria were considered successful when at least 50 % of the cells expressed the fluorescent signals. As ffigure 2 shows, mitochondria did not mix during the co-culture and only a few colocalization (seen as yellow staining) was found. The cells might overlap, climbing on top of each other on the surface of the culture dish and it cannot be claimed with certainty that these cells exchange whole mitochondria. Thus, transfection is better suited for mitochondrial transfer investigations than MitoTrackers although one must keep in mind that there are always non-labeled or lightly-labeled cells in the culture making the interpretation of the results more complex.
Fig. 2 Mitochondria were labeled with the transfection of green or red fluorescent protein-tagged mitochondrial proteins. (A) The two mitochondria populations were distinct without any double-staining or co-localization. (B) The Z-stack is the technique of choice for the investigation of the very scarce co-localizations of stainings (yellow).
2.2 Membrane staining with Vybrant dyes
The outer membrane of the cell can be easily stained with lipophilic Vybrant dyes such as DiO or DiD. These dyes have strong and bleach-resistant signals and were reportedly used in order to track implanted cells in vivo. We tested separately labeled stem cells in co-culture for up to 6 days, during which two cell populations remained excusively red or green with a high (over 20%) proportion of double labeled cells. Figure 3 shows the quantification of this observation using flow cytometry. Time-lapse microscopy revealed that some of these double-labeled cells underwent cell fusion, and membrane-exchange occurred through frequent tubular contacts among cells [17]. These observations indicate that although Vybrant dyes are considerably resistant to intercellular transfer, but conclusions drawn from their use in longer experiments should be evaluated with caution.
Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.)
©FORMATEX 2010 767
______________________________________________
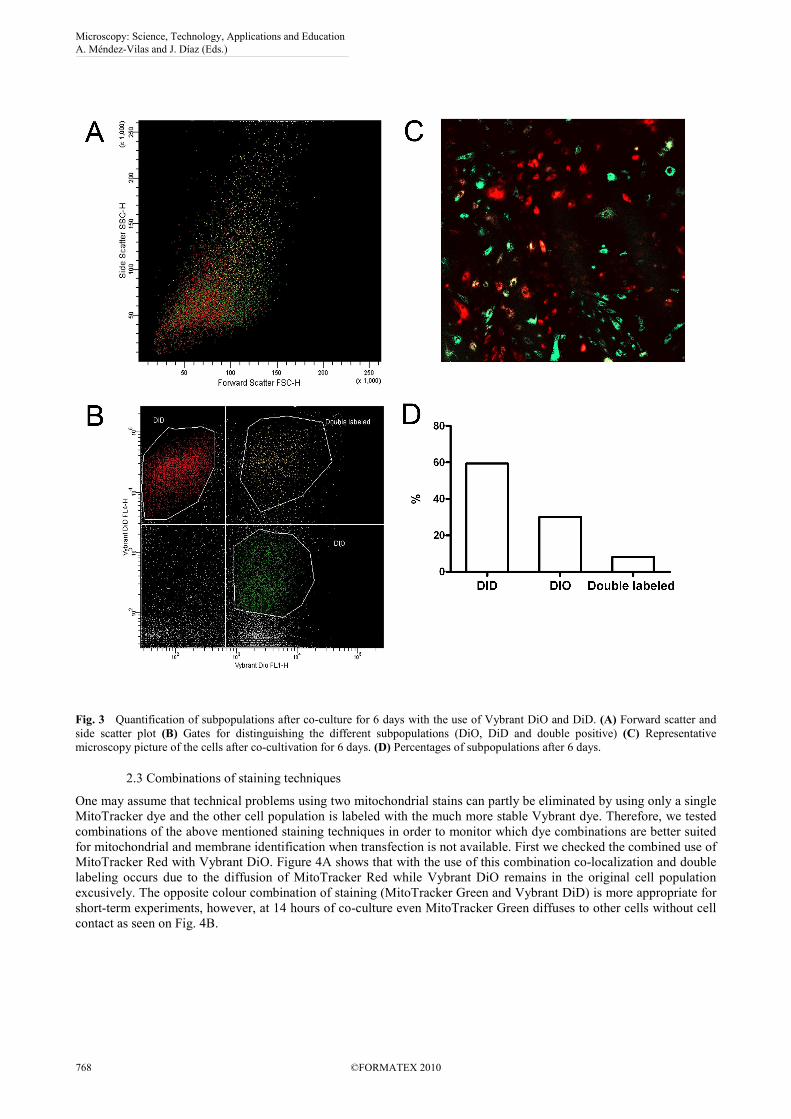
Fig. 3 Quantification of subpopulations after co-culture for 6 days with the use of Vybrant DiO and DiD. (A) Forward scatter and side scatter plot (B) Gates for distinguishing the different subpopulations (DiO, DiD and double positive) (C) Representative microscopy picture of the cells after co-cultivation for 6 days. (D) Percentages of subpopulations after 6 days.
2.3 Combinations of staining techniques
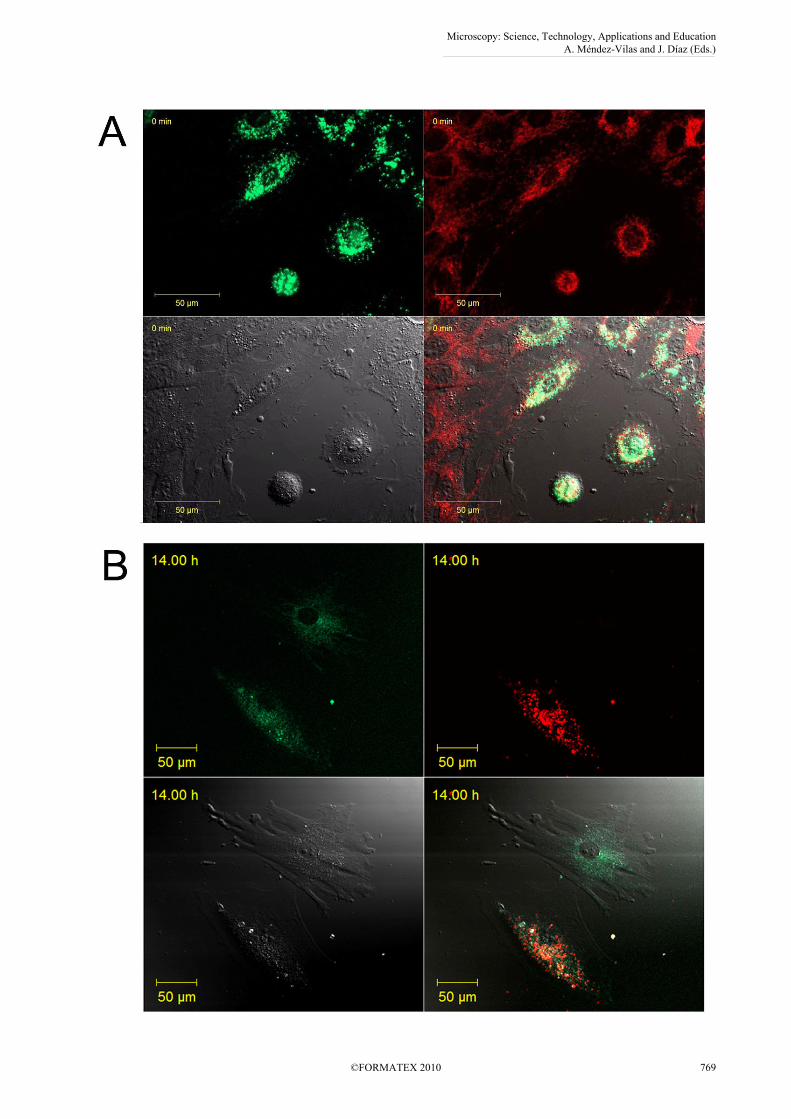
One may assume that technical problems using two mitochondrial stains can partly be eliminated by using only a single MitoTracker dye and the other cell population is labeled with the much more stable Vybrant dye. Therefore, we tested combinations of the above mentioned staining techniques in order to monitor which dye combinations are better suited for mitochondrial and membrane identification when transfection is not available. First we checked the combined use of MitoTracker Red with Vybrant DiO. Figure 4A shows that with the use of this combination co-localization and double labeling occurs due to the diffusion of MitoTracker Red while Vybrant DiO remains in the original cell population excusively. The opposite colour combination of staining (MitoTracker Green and Vybrant DiD) is more appropriate for short-term experiments, however, at 14 hours of co-culture even MitoTracker Green diffuses to other cells without cell contact as seen on Fig. 4B.
Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.)
768 ©FORMATEX 2010
______________________________________________
Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.)
©FORMATEX 2010 769
______________________________________________

Fig 4. The use of combined staining for the differentiation of cellular components. (A) MitoTracker Red and the cell membrane stain Vybrant DiO. Immediately after mixing the two cell populations MitoTracker Red diffuses to the other cells, while the green cell membrane dye remains separated. (B) MitoTracker Green and Vybrant DiI. MitoTracker Green diffuses to nearby cells over longer periods of time time as shown in this picture taken at 14 hours after co-culture. Thus, neither MitoTracker dyes are suitable for determining mitochondrial transfer in co-culture even in a setting where only one population is labeled.
3. Conclusions
We tested the potential of MitoTracker green and red to differentiate mitochondria in cellular co-cultures. Two cell populations were separately stained with the different dyes then cultured together on the stage of a confocal microscope and images were taken simultaneously of the two dyes. Although the slight overlap of the spectra in the two dyes can easily be excluded by separate exposure there was still an overlap of the two channels. We observed that MitoTracker red almost instantly stains other cells in the same culture dish, whereas MitoTracker green stays in the same cell for at least an hour. However, even MitoTracker green crosses to other cells during longer culture times, which was confirmed by fluorescent recovery after photobleaching (FRAP) protocols. Therefore, these dyes are inappropriate for the investigation of organelle transfer. When mitochondria were labeled with the transfection of green or red fluorescent protein-tagged mitochondrial proteins, the two mitochondria populations were distinct without any double-staining or co-localization. Thus, transfection is better suited for mitochondrial transfer investigations than MitoTracker although one must keep in mind that there are always non-labeled or lightly labeled cells in the culture making the interpretation of the results more complex. The outer membrane of the cell can be easily stained with lipophilic Vybrant dyes such as DiI, DiO or DiD. These dyes have strong and bleach-resistant signals and were reportedly used in order to track implanted cells in vivo. We tested separately labeled stem cells in co-culture for up to 6 days, during which two cell populations remained exclusively red or green, with a high proportion of double labeled cells. This observation was quantified with flow cytometry. Time-lapse microscopy revealed that some of these double labeled cells underwent cell fusion, and membrane-exchange occurred through frequent tubular contacts among cells. Taken together, our observations may lead to better and more relevant experimental protocols for the investigation of the transfer of cellular elements with confocal fluorescent microscopy.
Acknowledgements The supports by OTKA (Hungarian Scientific Research Fund) D45933, T049621, TÉT (Hungarian Science and Technology Foundation) A4/04, 08-SG-STAR-CELLTHER and Arg-17/2006, Bolyai, Öveges Fellowships and TÁMOP 4.2.2 08/1/KMR-2008-0004 and 4.2.1. are gratefully acknowledged.
References
[1] Kumar NM, Gilula NB. The gap junction communication channel. Cell. 1996;84(3):381-388. [2] Yeager M, Harris AL. Gap junction channel structure in the early 21st century: facts and fantasies. Curr Opin Cell Biol.
2007;19(5):521-528. [3] Kresh JY. Cell replacement therapy: the functional importance of myocardial architecture and intercellular gap-junction
distribution. J Thorac Cardiovasc Surg. 2006;131(6):1310-1313. [4] Rustom A, Saffrich R, Markovic I, Walther P, Gerdes HH. Nanotubular highways for intercellular organelle transport. Science.
2004;303(5660):1007-1010. [5] Koyanagi M, Brandes RP, Haendeler J, Zeiher AM, Dimmeler S. Cell-to-cell connection of endothelial progenitor cells with
cardiac myocytes by nanotubes: a novel mechanism for cell fate changes? Circ Res. 2005;96(10):1039-1041. [6] Onfelt B, Nedvetzki S, Yanagi K, Davis DM. Cutting edge: Membrane nanotubes connect immune cells. J Immunol.
2004;173(3):1511-1513. [7] Onfelt B, Purbhoo MA, Nedvetzki S, Sowinski S, Davis DM. Long-distance calls between cells connected by tunneling
nanotubules. Sci STKE. 2005;2005(313):pe55. [8] Gerdes HH, Bukoreshtliev NV, Barroso JF. Tunneling nanotubes: a new route for the exchange of components between animal
cells. FEBS Lett. 2007;581(11):2194-2201. [9] Gurke S, Barroso JF, Gerdes HH. The art of cellular communication: tunneling nanotubes bridge the divide. Histochem Cell
Biol. 2008;129(5):539-550. [10] Wang T, Xu Z, Jiang W, Ma A. Cell-to-cell contact induces mesenchymal stem cell to differentiate into cardiomyocyte and
smooth muscle cell. Int J Cardiol. 2006;109(1):74-81. [11] Driesen RB, Dispersyn GD, Verheyen FK, van den Eijnde SM, Hofstra L, Thone F, Dijkstra P, Debie W, Borgers M,
Ramaekers FC. Partial cell fusion: a newly recognized type of communication between dedifferentiating cardiomyocytes and fibroblasts. Cardiovasc Res. 2005;68(1):37-46.
[12] Wallace DC. Mitochondrial DNA sequence variation in human evolution and disease. Proc Natl Acad Sci U S A. 1994;91(19):8739-8746.
[13] Dyall SD, Brown MT, Johnson PJ. Ancient invasions: from endosymbionts to organelles. Science. 2004;304(5668):253-257. [14] Spees JL, Olson SD, Whitney MJ, Prockop DJ. Mitochondrial transfer between cells can rescue aerobic respiration. Proc Natl
Acad Sci U S A. 2006;103(5):1283-1288. [15] King MP, Attardi G. Human cells lacking mtDNA: repopulation with exogenous mitochondria by complementation. Science.
1989;246(4929):500-503.
Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.)
770 ©FORMATEX 2010
______________________________________________

[16] Bodnar AG, Cooper JM, Holt IJ, Leonard JV, Schapira AH. Nuclear complementation restores mtDNA levels in cultured cells from a patient with mtDNA depletion. Am J Hum Genet. 1993;53(3):663-669.
[17] Cselenyak A, Pankotai E, Horvath EM, Kiss L, Lacza Z. Mesenchymal stem cells rescue cardiomyoblasts from cell death in an in vitro ischemia model via direct cell-to-cell connections. BMC Cell Biol. 11(29.
[18] Plotnikov EY, Khryapenkova TG, Vasileva AK, Marey MV, Galkina SI, Isaev NK, Sheval EV, Polyakov VY, Sukhikh GT, Zorov DB. Cell-to-cell cross-talk between mesenchymal stem cells and cardiomyocytes in co-culture. J Cell Mol Med. 2008;12(5A):1622-1631.
[19] Nygren JM, Jovinge S, Breitbach M, Sawen P, Roll W, Hescheler J, Taneera J, Fleischmann BK, Jacobsen SE. Bone marrow-derived hematopoietic cells generate cardiomyocytes at a low frequency through cell fusion, but not transdifferentiation. Nat Med. 2004;10(5):494-501.
[20] Ishikawa F, Shimazu H, Shultz LD, Fukata M, Nakamura R, Lyons B, Shimoda K, Shimoda S, Kanemaru T, Nakamura K, Ito H, Kaji Y, Perry AC, and Harada M. Purified human hematopoietic stem cells contribute to the generation of cardiomyocytes through cell fusion. Faseb J. 2006;20(7):950-952.
[21] Lacza Z, Horvath E, Busija DW. Neural stem cell transplantation in cold lesion: a novel approach for the investigation of brain trauma and repair. Brain Res Brain Res Protoc. 2003;11(3):145-154.
[22] Horvath EM, Lacza Z, Csordas A, Szabo C, Kollai M, Busija DW. Graft derived cells with double nuclei in the penumbral region of experimental brain trauma. Neurosci Lett. 2006;396(3):182-186.
[23] Alvarez-Dolado M, Pardal R, Garcia-Verdugo JM, Fike JR, Lee HO, Pfeffer K, Lois C, Morrison SJ, Alvarez-Buylla A. Fusion of bone-marrow-derived cells with Purkinje neurons, cardiomyocytes and hepatocytes. Nature. 2003;425(6961):968-973.
[24] Weimann JM, Johansson CB, Trejo A, Blau HM. Stable reprogrammed heterokaryons form spontaneously in Purkinje neurons after bone marrow transplant. Nat Cell Biol. 2003;5(11):959-966.
[25] Ying QL, Nichols J, Evans EP, Smith AG. Changing potency by spontaneous fusion. Nature. 2002;416(6880):545-548. [26] Menasche P. You can't judge a book by its cover. Circulation. 2006;113(10):1275-1277. [27] Garbade J, Schubert A, Rastan AJ, Lenz D, Walther T, Gummert JF, Dhein S, Mohr FW. Fusion of bone marrow-derived stem
cells with cardiomyocytes in a heterologous in vitro model. Eur J Cardiothorac Surg. 2005;28(5):685-691. [28] Kajstura J, Rota M, Whang B, Cascapera S, Hosoda T, Bearzi C, Nurzynska D, Kasahara H, Zias E, Bonafe M, Nadal-Ginard
B, Torella D, Nascimbene A, Quaini F, Urbanek K, Leri A, and Anversa P. Bone marrow cells differentiate in cardiac cell lineages after infarction independently of cell fusion. Circ Res. 2005;96(1):127-137.
[29] Terada N, Hamazaki T, Oka M, Hoki M, Mastalerz DM, Nakano Y, Meyer EM, Morel L, Petersen BE, Scott EW. Bone marrow cells adopt the phenotype of other cells by spontaneous cell fusion. Nature. 2002;416(6880):542-545.
[30] Rodic N, Rutenberg MS, Terada N. Cell fusion and reprogramming: resolving our transdifferences. Trends Mol Med. 2004;10(3):93-96.
[31] Wurmser AE, Gage FH. Stem cells: cell fusion causes confusion. Nature. 2002;416(6880):485-487. [32] Wurmser AE, Nakashima K, Summers RG, Toni N, D'Amour KA, Lie DC, Gage FH. Cell fusion-independent differentiation
of neural stem cells to the endothelial lineage. Nature. 2004;430(6997):350-356. [33] Paxinou E, Weisse M, Chen Q, Souza JM, Hertkorn C, Selak M, Daikhin E, Yudkoff M, Sowa G, Sessa WC, and
Ischiropoulos H. Dynamic regulation of metabolism and respiration by endogenously produced nitric oxide protects against oxidative stress. Proc Natl Acad Sci U S A. 2001;98(20):11575-11580.
Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.)
©FORMATEX 2010 771
______________________________________________





























