Embed Size (px)
Citation preview

Biophysics. In the article “Membrane permeabilization byListeria monocytogenes phosphatidylinositol-specific phospho-lipase C is independent of phospholipid hydrolysis and coop-erative with listeriolysin O” by Howard Goldfine, ChristopherKnob, Dennis Alford, and Joe Bentz, which appeared innumber 7, March 28, 1995, of Proc. Natl. Acad. Sci. USA (92,2979–2983), the authors wish to retract the data showing thatthe phosphatidylinositol-specific phospholipase C (PI-PLC) iscapable of permeabilizing membranes consisting of phosphati-dylcholine and cholesterol, and that it cooperates with list-eriolysin O in this activity. With similar preparations ofPI-PLC, the authors have separated the PI-PLC activity fromthe permeabilizing activity by hydrophobic interaction chro-matography. The latter activity in these preparations comi-grated with listeriolysin O.
Howard GoldfineChristopher KnobDennis AlfordJoe Bentz
Copyright q 1997 by THE NATIONAL ACADEMY OF SCIENCES OF THE USA0027-8424y97y942772-1$2.00y0PNAS is available online at http:yywww.pnas.org.
2772 Retraction Proc. Natl. Acad. Sci. USA 94 (1997)

Proc. Natl. Acad. Sci. USAVol. 92, pp. 2979-2983, March 1995Biophysics
Membrane permeabilization by Listeria monocytogenesphosphatidylinositol-specific phospholipase C isindependent of phospholipid hydrolysis andcooperative with listeriolysin 0HOWARD GOLDFINE*t, CHRISTOPHER KNOB*t, DENNIS ALFORD§I, AND JOE BENTZ§*Department of Microbiology, University of Pennsylvania School of Medicine, Philadelphia, PA 19104-6076; and §Department of Bioscience and Biotechnology,Drexel University, Philadelphia, PA 19104-2875
Communicated by Mildred Cohn, University of Pennsylvania School of Medicine, Philadelphia, PA, December 5, 1994
ABSTRACT We have examined potential cooperative in-teractions of Listeria monocytogenes phosphatidylinositol-specific phospholipase C (PI-PLC) and listeriolysin 0 (LLO),a pore-forming hemolysin, in a liposome lysis assay. Largeunilamellar vesicles, -0.1 ,um in diameter, encapsulating thefluorescent probe calcein, were treated with PI-PLC or LLOat pH 6.0, and each was capable of causing dye release. Withphosphatidylcholine/phosphatidylinositol/cholesterol lipo-somes at 0.1 IuM lipid, minimal release of dye was observed onaddition of 80 pM LLO or 7 nM PI-PLC. Addition of the twoproteins together produced rapid dye release. Unexpectedly,essentially identical results were obtained with phosphatidyl-choline/cholesterol liposomes. Thus, the effect of PI-PLC didnot depend on lipid hydrolysis. Both proteins also releasedinulin (Mr 5200) from liposomes. Membrane permeabiliza-tion was not accompanied by membrane fusion. Very littledye releasefrom phosphatidylcholine/phosphatidylinositol/cholesterol liposomes was seen with PI-PLC from Bacillusthuringiensis, and addition of this enzyme to LLO produced noadditional dye release; however PI-PLC from L. monocytogenescooperated with perfringolysin 0 from Clostridium perfrin-gens. PI-PLC from L. monocytogenes and LLO bind to phos-phatidylcholine/cholesterol liposomes, and the rate of bind-ing of each protein was not influenced by the presence of theother. These data support a postulated accessory role forPI-PLC with LLO in lysing the primary phagosome of amacrophage.
The facultative intracellular human pathogen Listeria mono-cytogenes is a Gram-positive non-spore-forming rod, whichgrows rapidly in laboratory culture and in the cytoplasm ofinfected cells (1-3). Infection in humans can cause severemorbidity and mortality in immunocompromised adults andpregnant women (2). Recently, several genes encoding deter-minants of pathogenesis have been identified and found to beclustered on the chromosome. Among these are hly, whichencodes listeriolysin 0 (LLO)-a member of a family ofpore-forming, sulfhydryl-activated hemolysins secreted by bac-teria; picA, which encodes a phosphatidylinositol (Ptdlns)-specific phospholipase C (PI-PLC); andplcB, which encodes abroad-range, phosphatidylcholine (PtdCho)-preferring phos-pholipase C (PC-PLC). The genetics and regulation of thesegenes have been reviewed (3).The search for the role of L. monocytogenes PI-PLC in
pathogenesis has recently focused on the phagosome resultingfrom the initial infection of cells by bacteria-induced phago-cytosis (4). Studies on infections in murine macrophagesshowed that only LLO is required for growth (3); however, anin-frameplcA deletion mutant was delayed in escape from the
primary phagosome compared with wild type (5). Growth ofthis mutant in the livers of infected mice was also slower thanthat of wild type (5). The homogeneous protein is specific forPtdlns, with no activity on PtdIns 4-phosphate or Ptdlns4,5-bisphosphate (6). In contrast to other bacterial PI-PLCs, ithas very weak or no activity on several PtdIns-anchoredmembrane proteins (6, 7). We have studied the ability ofPI-PLC to release solutes from large unilamellar lipid vesicles(LUV). These studies have shown that PI-PLC is capable ofinducing solute release from liposomes alone or in cooperationwith LLO. Remarkably, this activity is independent of itsability to hydrolyze Ptdlns.
MATERIALS AND METHODS
Materials. L. monocytogenes strains were obtained fromDaniel Portnoy (University of Pennsylvania). PI-PLC waspurified as described from the culture supernatants of L.monocytogenes DP-L1470 (6). It did not contain LLO detect-able by immunoblotting (Western blotting) with antibody toLLO. Furthermore, it was active in LUV permeabilization atpH 7.0 and did not require prior treatment with a reducingagent. PI-PLC from Bacillus thuringiensis was a gift fromMartin G. Low (Columbia University). Both PI-PLC prepa-rations had specific activities of 1000-1500 ,umol/min per mgof protein on detergent-dispersed Ptdlns (6, 8), and bothhydrolyzed Ptdlns in liposomes (data not shown). PC-PLC wasisolated as described (9, 10) from L. monocytogenes DP-L1553AplcA (5). LLO was isolated by using essentially the sameprocedure (9) from the culture supernatant of L. monocyto-genes DP-L1545 mpl-. LLO was more stable in the absence ofcontaminating L. monocytogenes metalloprotease. Bio-GelP-100 fractions containing homogeneous LLO were detectedby SDS/PAGE, followed by staining with Coomassie blue.Perfringolysin 0 was a gift from Rodney K. Tweten (Univer-sity of Oklahoma Health Science Center). All proteins wereassayed for activity at the time of these experiments. LLO andperfringolysin 0 were assayed on sheep erythrocytes (11).PI-PLC (6) and PC-PLC (10) were assayed as described.Rabbit polyclonal antiserum against PI-PLC from this labo-ratory and anti-LLO, a gift from Pascale Cossart (Pasteur
Abbreviations: LLO, listeriolysin 0; LUV, large unilamellar vesicles;PtdCho, phosphatidylcholine; Ptdlns, phosphatidylinositol; PLC,phospholipase C; PI-PLC, Ptdlns-specific PLC; PC-PLC, PtdCho-specific PLC; CPT, [4-(7-diethylamino-4-methylcoumarin-3-yl)phenyl]-N-methylthiourea; DABS, [4-dimethylaminophenylazo-benzene-4'-(N'-methyl)sulfonamide].tTo whom reprint requests should be addressed.lPresent address: Virginia Institute of Marine Science, GloucesterPoint, VA 23062.TPresent address: Center for Blood Research Laboratories, Boston,MA 02115.
2979
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement" inaccordance with 18 U.S.C. §1734 solely to indicate this fact.
Proc. Natl. Acad. Sci. USA 92 (1995)
Institute, Paris), were used to detect liposome-bound proteinsby Western blotting.
Preparation of LUV. Dioleoyl PtdCho, plant Ptdlns (AvantiPolar Lipids), and cholesterol (Sigma) were used in theproportions listed to generate LUV encapsulating 50 mMcalcein (Molecular Probes)/10 mM 2-{[tris(hydroxymethyl)methyl]amino}ethane sulfonic acid (Tes)/0.1 mM EDTA [ti-trated to pH 6.0, followed by adjustment of osmolarity to thatof the incubation buffer (see below)] by reversed-phase evap-oration and 10 extrusions through 0.1-,um polycarbonate filters(12). Unencapsulated calcein was removed by chromatographyon Sephadex G-75. LUV consisting of PtdCho/cholesterol,70:30 molar ratio, encapsulating [methoxy-3H]inulin [275,uCi/mg (1 Ci = 37 GBq); New England Nuclear] dissolved inthe buffer used for dye release experiments, were prepared byfreeze/thaw cycling followed sequentially by 10 extrusionsthrough 0.4-,um pores in polycarbonate filters and 8 extrusionsthrough a 0.2-,um filter. The liposomes were separated fromunencapsulated [3H]inulin on a 60 x 1 cm Sephadex G-75column. Liposome concentrations were measured by phos-phate analysis after (20%) perchloric acid digestion (12).
Assays. Dye release. LUV (0.1 ,uM total lipid) were dilutedinto pH 6.0 incubation buffer containing 150mM NaCl, 10mMTes, and 0.1 mM EDTA. PI-PLC was added after dilution inincubation buffer containing 0.1% bovine serum albumin,which did not permeabilize LUV by itself. LLO was prere-duced in incubation buffer at pH 6.0 containing 20 mMcysteine for 20 min at room temperature. Most experimentswere done at pH 6.0. In addition, we repeated all criticalexperiments in 10mM Tes/Mes, which was well buffered at pH6.0, and measured the pH of representative incubations withboth buffers and found them to give the stated pH ± 0.1 unit.While it is known that calcein fluorescence is sensitive to pH,it remains membrane impermeant down to pH 4.0 (13), and thepH was the same inside and outside the vesicles. Initially,calcein at 50 mM is self-quenched inside the LUV, and thisfluorescence level was set to 0% leakage. Upon release,fluorescence (Aex = 494 nm and Aem = 518 nm, where ex =excitation and em = emission) increases due to dequenching;100% leakage was set to the fluorescence level achievedfollowing detergent lysis (C12E8, Calbiochem). Measurementswere made on a PTI Alphascan fluorometer (South Bruns-wick, NJ) in a thermostated, stirred cuvette. Encapsulatedvolume for LUV was 2.5 ± 0.5 p,l/,umol of total lipid. In thecuvette, the lipid concentration was always 0.1 ,uM, whichimplies 12.5 nM calcein after lysis.Liposome fusion. To examine whether either LLO or
PI-PLC could induce membrane fusion, we employed the[4-(7-diethylamino-4-methylcoumarin-3-yl)phenyl]-N-methyl-thiourea (CPT)/[4-dimethylaminophenylazobenzene-4'-(N-methyl)sulfonamide] (DABS)-conjugated lipid mixing assay(14). Briefly, 1-palmitoyl-2-{ 12-N'-[4-(7-diethylamino-4-methylcoumarin-3-yl)phenyl]-N-methylthioureidyloctadeca-noyl}-sn-glycerol-3-phosphocholine (12-CPT-18-PtdCho) is afluorescent phospholipid and 1-palmitoyl-2-{ 12-[4-dimethyl-aminophenylazobenzene-4'-(N'-methyl)sulfonamidooctadeca-noyl]}sn-glycerol-3-phosphocholine (12-DABS-18-PtdCho) isa nonfluorescent phospholipid collisional quencher of CPT.Labeled Ptdlns/PtdCho/cholesterol LUV were made with 1.0mol % each of 12-CPT-18-PC and 12-DABS-18-PC. LabeledLUV (0.1 ,uM) were mixed with 0.9 ,uM unlabeled LUV,followed by addition of protein. The total lipid concentrationswere 10-fold higher than in the calcein leakage experiments tocompensate for the lower sensitivity of the assay. Fusion wouldresult in increased CPT fluorescence (Aex = 395 nm and Aem= 477 nm) due to dilution of the probes.Inulin release. LUV encapsulating [methoxy-3H]inulin were
treated with LLO or PI-PLC in Tes (pH 6.0) at 37°C. Afterincubation the reaction mixture was placed on a 60 x 1 cmSephadex G-75 column equilibrated at pH 7.0 to stop LLO
action; elution with 10 mM Tes, pH 7.0/150 mM NaCl bufferseparated released inulin from intact liposomes.Liposome binding. The ability of PI-PLC and LLO to bind to
liposomes was determined as described by Long et at (15).Proteins were incubated with liposomes at 37°C for the timesindicated, layered under a sucrose gradient, and centrifuged at200,000 x g in a swinging-bucket rotor for 3 hr. Liposomeswere removed from the top of the gradient, precipitatedwith trichloroacetic acid, and subjected to SDS/PAGE. Pro-teins were detected with appropriate antibodies by Westernblotting.
RESULTS
Induction of Calcein Leakage by LLO, PI-PLC, or PC-PLCfrom L. monocytogenes. LLO 2 230 pM (12.5 ng/ml), PI-PLC2 41 nM (1.35 ,Lg/ml), or PC-PLC 2 6.6 nM (185 ng/ml) wasindependently capable of causing rapid release of calcein at pH6.0 from LUV (0.1 ,M total lipid) consisting of PtdCho/PtdIns/cholesterol, 60:10:30 molar ratio. LLO has very lowactivity at neutral pH (16, 17), which we have confirmed in thecalcein release assay. In contrast, PI-PLC and PC-PLC per-meabilized LUV at pH 7.0.
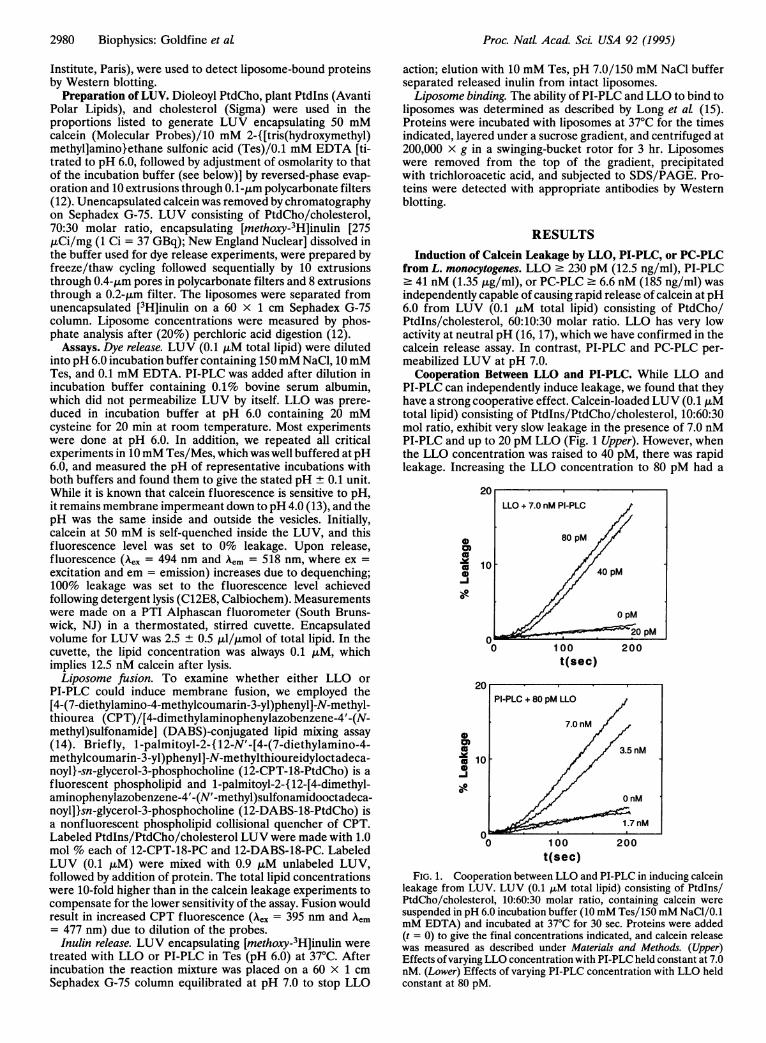
Cooperation Between LLO and PI-PLC. While LLO andPI-PLC can independently induce leakage, we found that theyhave a strong cooperative effect. Calcein-loaded LUV (0.1 ,uMtotal lipid) consisting of Ptdlns/PtdCho/cholesterol, 10:60:30mol ratio, exhibit very slow leakage in the presence of 7.0 nMPI-PLC and up to 20 pM LLO (Fig. 1 Upper). However, whenthe LLO concentration was raised to 40 pM, there was rapidleakage. Increasing the LLO concentration to 80 pM had a
0
0)
co0
0-
1
0
0
c oco-
0)0i
1 00
t(sec)200
100
t(sec)
FIG. 1. Cooperation between LLO and PI-PLC in inducing calceinleakage from LUV. LUV (0.1 ALM total lipid) consisting of Ptdlns/PtdCho/cholesterol, 10:60:30 molar ratio, containing calcein were
suspended in pH 6.0 incubation buffer (10mM Tes/150mM NaCl/0.1mM EDTA) and incubated at 37°C for 30 sec. Proteins were added(t = 0) to give the final concentrations indicated, and calcein releasewas measured as described under Materials and Methods. (Upper)Effects ofvarying LLO concentration with PI-PLC held constant at 7.0nM. (Lower) Effects of varying PI-PLC concentration with LLO heldconstant at 80 pM.
LLO + 7.0 nM PI-PLC
80 pM
0 , 40 pM
0 pM
a) _ _o20 pM
2980 Biophysics: Goldfine et at
Proc. Natl. Acad Sci. USA 92 (1995) 2981
small additional effect. In Fig. 1 Lower we see that these LUVexhibit slow leakage in the presence of 80 pM LLO with up to1.7 nM PI-PLC. Increasing the PI-PLC concentration to 3.5nM induced rapid leakage, and 7.0 nM PI-PLC caused a
further modest increase in the leakage kinetics.We next examined whether this cooperativity depended
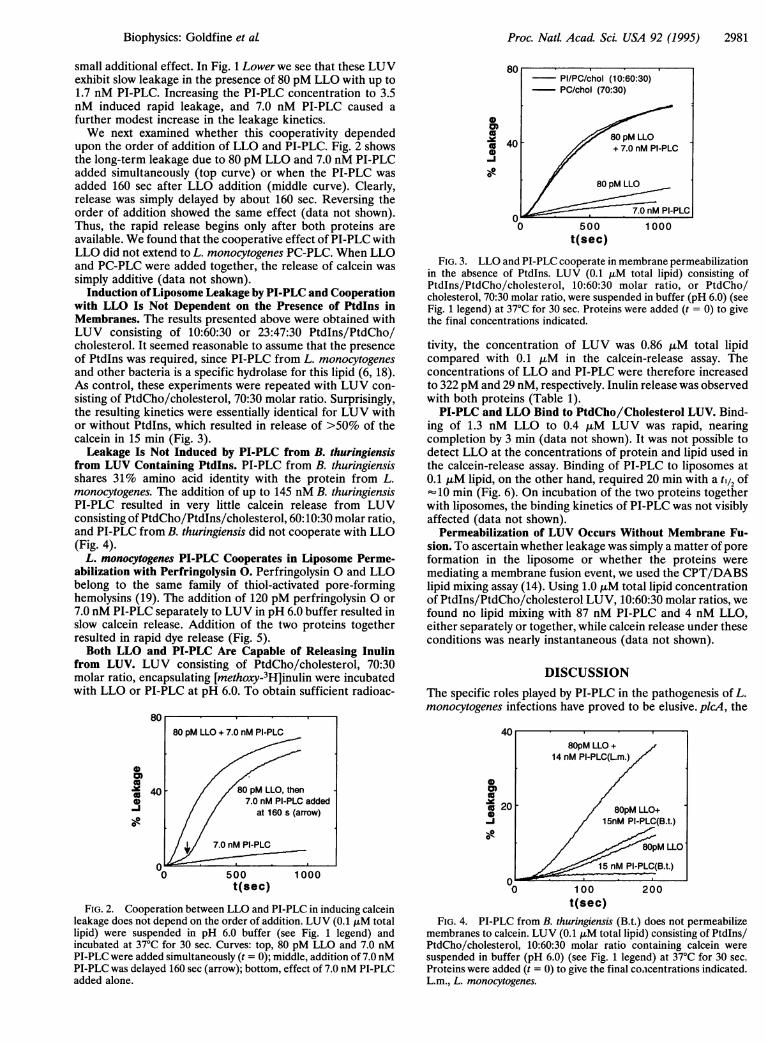
upon the order of addition of LLO and PI-PLC. Fig. 2 showsthe long-term leakage due to 80 pM LLO and 7.0 nM PI-PLCadded simultaneously (top curve) or when the PI-PLC wasadded 160 sec after LLO addition (middle curve). Clearly,release was simply delayed by about 160 sec. Reversing theorder of addition showed the same effect (data not shown).Thus, the rapid release begins only after both proteins areavailable. We found that the cooperative effect of PI-PLC withLLO did not extend to L. monocytogenes PC-PLC. When LLOand PC-PLC were added together, the release of calcein wassimply additive (data not shown).
Induction ofLiposome Leakage by PI-PLC and Cooperationwith LLO Is Not Dependent on the Presence of PtdIns inMembranes. The results presented above were obtained withLUV consisting of 10:60:30 or 23:47:30 Ptdlns/PtdCho/cholesterol. It seemed reasonable to assume that the presenceof Ptdlns was required, since PI-PLC from L. monocytogenesand other bacteria is a specific hydrolase for this lipid (6, 18).As control, these experiments were repeated with LUV con-sisting of PtdCho/cholesterol, 70:30 molar ratio. Surprisingly,the resulting kinetics were essentially identical for LUV withor without Ptdlns, which resulted in release of >50% of thecalcein in 15 min (Fig. 3).Leakage Is Not Induced by PI-PLC from B. thuringiensis
from LUV Containing Ptdlns. PI-PLC from B. thuringiensisshares 31% amino acid identity with the protein from L.monocytogenes. The addition of up to 145 nM B. thuringiensisPI-PLC resulted in very little calcein release from LUVconsisting of PtdCho/Ptdlns/cholesterol, 60:10:30 molar ratio,and PI-PLC from B. thuringiensis did not cooperate with LLO(Fig. 4).
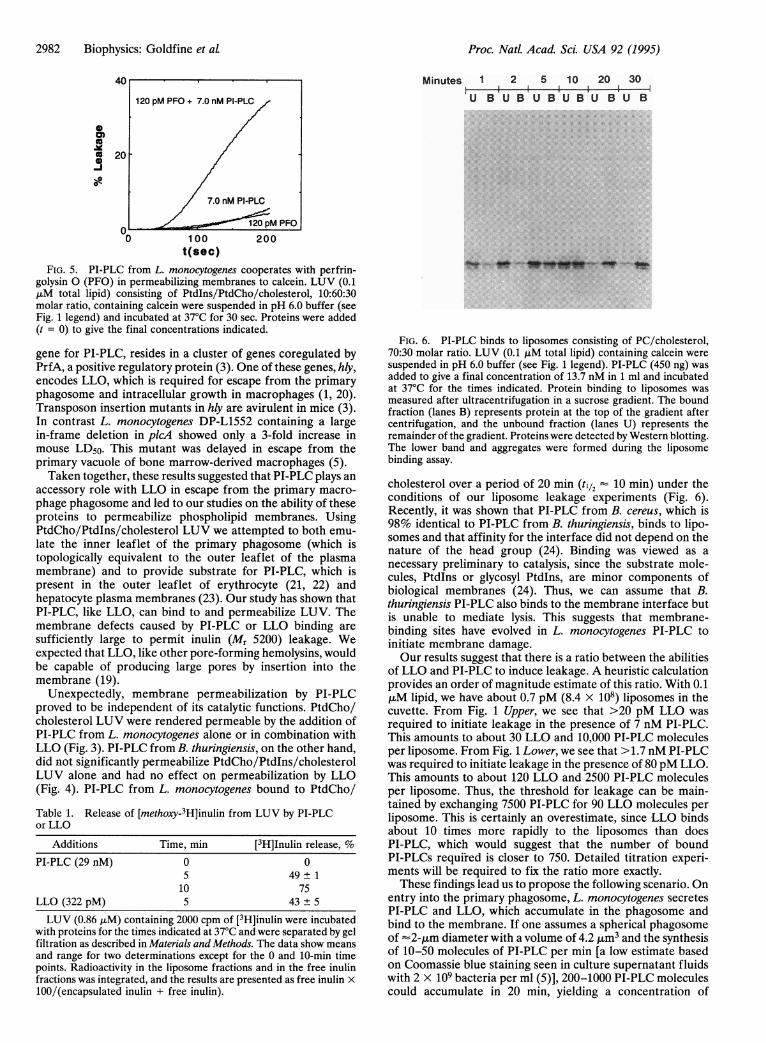
L. monocytogenes PI-PLC Cooperates in Liposome Perme-abilization with Perfringolysin 0. Perfringolysin 0 and LLObelong to the same family of thiol-activated pore-forminghemolysins (19). The addition of 120 pM perfringolysin 0 or7.0 nM PI-PLC separately to LUV in pH 6.0 buffer resulted inslow calcein release. Addition of the two proteins togetherresulted in rapid dye release (Fig. 5).Both LLO and PI-PLC Are Capable of Releasing Inulin
from LUV. LUV consisting of PtdCho/cholesterol, 70:30molar ratio, encapsulating [methoxy-3H]inulin were incubatedwith LLO or PI-PLC at pH 6.0. To obtain sufficient radioac-
8080 pM LLO + 7.0 nM PI-PLC
1 40 - / /80 pM LLO, then
/ / 7.0 nM PI-PLC added
_J / / at 160 s (arrow)
7.0 nM PI-PLC
00 500 1000
t(sec)
FIG. 2. Cooperation between LLO and PI-PLC in inducing calceinleakage does not depend on the order of addition. LUV (0.1 ,AM totallipid) were suspended in pH 6.0 buffer (see Fig. 1 legend) andincubated at 37°C for 30 sec. Curves: top, 80 pM LLO and 7.0 nMPI-PLC were added simultaneously (t = 0); middle, addition of 7.0 nMPI-PLC was delayed 160 sec (arrow); bottom, effect of 7.0 nM PI-PLCadded alone.
0
as
-i0y
0 500t(sec)
1 000
FIG. 3. LLO and PI-PLC cooperate in membrane permeabilizationin the absence of PtdIns. LUV (0.1 ,uM total lipid) consisting ofPtdlns/PtdCho/cholesterol, 10:60:30 molar ratio, or PtdCho/cholesterol, 70:30 molar ratio, were suspended in buffer (pH 6.0) (seeFig. 1 legend) at 37°C for 30 sec. Proteins were added (t = 0) to givethe final concentrations indicated.
tivity, the concentration of LUV was 0.86 ,uM total lipidcompared with 0.1 ,uM in the calcein-release assay. Theconcentrations of LLO and PI-PLC were therefore increasedto 322pM and 29 nM, respectively. Inulin release was observedwith both proteins (Table 1).PI-PLC and LLO Bind to PtdCho/Cholesterol LUV. Bind-
ing of 1.3 nM LLO to 0.4 ,sM LUV was rapid, nearingcompletion by 3 min (data not shown). It was not possible todetect LLO at the concentrations of protein and lipid used inthe calcein-release assay. Binding of PI-PLC to liposomes at0.1 ,uM lipid, on the other hand, required 20 min with a til2 ofo10 min (Fig. 6). On incubation of the two proteins together
with liposomes, the binding kinetics of PI-PLC was not visiblyaffected (data not shown).
Permeabilization of LUV Occurs Without Membrane Fu-sion. To ascertain whether leakage was simply a matter of poreformation in the liposome or whether the proteins weremediating a membrane fusion event, we used the CPT/DABSlipid mixing assay (14). Using 1.0 ,uM total lipid concentrationof PtdIns/PtdCho/cholesterol LUV, 10:60:30 molar ratios, wefound no lipid mixing with 87 nM PI-PLC and 4 nM LLO,either separately or together, while calcein release under theseconditions was nearly instantaneous (data not shown).
DISCUSSIONThe specific roles played by PI-PLC in the pathogenesis of L.monocytogenes infections have proved to be elusive. picA, the
0
0
v0 100
t(sec)200
FIG. 4. PI-PLC from B. thuringiensis (B.t.) does not permeabilizemembranes to calcein. LUV (0.1 ,uM total lipid) consisting of PtdIns/PtdCho/cholesterol, 10:60:30 molar ratio containing calcein weresuspended in buffer (pH 6.0) (see Fig. 1 legend) at 37°C for 30 sec.Proteins were added (t = 0) to give the final concentrations indicated.L.m., L. monocytogenes.
PI/PC/chol (10:60:30)PC/chol (70:30)
40 80 pM LLO+ 7.0 nM PI-PLC
80 pM LLO
7.0 nM PI-PLCO
4080pM LLO +
14 nM PI-PLC(L.m.)
20 / 80pM LLO+15nM PI-PLC(B.t.)
8OpM LLO
15 nM PI-PLC(B.t.)r}_
Biophysics: Goldfine et aL
Proc. Natl. Acad. Sci. USA 92 (1995)
0)(U
9 20
7.0 nM PI-PLC
O z , 120 pM PFO0 100 200
t(sec)
FIG. 5. PI-PLC from L. monocytogenes cooperates with perfrin-golysin 0 (PFO) in permeabilizing membranes to calcein. LUV (0.1,uM total lipid) consisting of Ptdlns/PtdCho/cholesterol, 10:60:30molar ratio, containing calcein were suspended in pH 6.0 buffer (seeFig. 1 legend) and incubated at 37°C for 30 sec. Proteins were added(t = 0) to give the final concentrations indicated.
gene for PI-PLC, resides in a cluster of genes coregulated byPrfA, a positive regulatory protein (3). One of these genes, hly,encodes LLO, which is required for escape from the primaryphagosome and intracellular growth in macrophages (1, 20).Transposon insertion mutants in hly are avirulent in mice (3).In contrast L. monocytogenes DP-L1552 containing a largein-frame deletion in plcA showed only a 3-fold increase inmouse LD50. This mutant was delayed in escape from theprimary vacuole of bone marrow-derived macrophages (5).Taken together, these results suggested that PI-PLC plays an
accessory role with LLO in escape from the primary macro-phage phagosome and led to our studies on the ability of theseproteins to permeabilize phospholipid membranes. UsingPtdCho/Ptdlns/cholesterol LUV we attempted to both emu-late the inner leaflet of the primary phagosome (which istopologically equivalent to the outer leaflet of the plasmamembrane) and to provide substrate for PI-PLC, which ispresent in the outer leaflet of erythrocyte (21, 22) andhepatocyte plasma membranes (23). Our study has shown thatPI-PLC, like LLO, can bind to and permeabilize LUV. Themembrane defects caused by PI-PLC or LLO binding aresufficiently large to permit inulin (Mr 5200) leakage. Weexpected that LLO, like other pore-forming hemolysins, wouldbe capable of producing large pores by insertion into themembrane (19).
Unexpectedly, membrane permeabilization by PI-PLCproved to be independent of its catalytic functions. PtdCho/cholesterol LUV were rendered permeable by the addition ofPI-PLC from L. monocytogenes alone or in combination withLLO (Fig. 3). PI-PLC from B. thuringiensis, on the other hand,did not significantly permeabilize PtdCho/Ptdlns/cholesterolLUV alone and had no effect on permeabilization by LLO(Fig. 4). PI-PLC from L. monocytogenes bound to PtdCho/
Table 1. Release of [methoxy-3H]inulin from LUV by PI-PLCor LLO
Additions Time, min [3H]Inulin release, %PI-PLC (29 nM) 0 0
5 49± 110 75
LLO (322 pM) 5 43 ± 5
LUV (0.86 ,LM) containing 2000 cpm of [3H]inulin were incubatedwith proteins for the times indicated at 37°C and were separated by gelfiltration as described in Materials and Methods. The data show meansand range for two determinations except for the 0 and 10-min timepoints. Radioactivity in the liposome fractions and in the free inulinfractions was integrated, and the results are presented as free inulin x100/(encapsulated inulin + free inulin).
Minutes 1 2 5 10 20 30I I I I I
U B U B U B U B U B U B
FIG. 6. P1-PLC binds to liposomes consisting of PC/cholesterol,70:30 molar ratio. LUV (0.1 liM total lipid) containing calcein weresuspended in pH 6.0 buffer (see Fig. 1 legend). P1-PLC (450 ng) wasadded to give a final concentration of 13.7 nM in 1 ml and incubatedat 370C for the times indicated. Protein binding to liposomes wasmeasured after ultracentrifugation in a sucrose gradient. The boundfraction (lanes B) represents protein at the top of the gradient aftercentrifugation, and the unbound fraction (lanes U) represents theremainder of the gradient. Proteins were detected by Western blotting.The lower band and aggregates were formed during the liposomebinding assay.
cholesterol over a period of 20 mmn (t112 10 mmn) under theconditions of our liposome leakage experiments (Fig. 6).Recently, it was shown that PT-PLC from B. cereus, which is98% identical to P1-PLC from B. thuringiensis, binds to lipo-somes and that affinity for the interface did not depend on thenature of the head group (24). Binding was viewed as anecessary preliminary to catalysis, since the substrate mole-cules, PtdIns or glycosyl Ptdlns, are minor components ofbiological membranes (24). Thus, we can assume that B.thuringiensis PT-PLC also binds to the membrane interface butis unable to mediate lysis. This suggests that membrane-binding sites have evolved in L. monocytogenes P1-PLC toinitiate membrane damage.Our results suggest that there is a ratio between the abilities
of LLO and PI-PLC to induce leakage. A heuristic calculationprovides an order of magnitude estimate of this ratio. With 0.1gM lipid, we have about 0.7 pM (8.4 X 108) liposomes in thecuvette. From Fig. 1 Upper, we see that >20 pM LLO wasrequired to initiate leakage in the presence of 7 nM PT-PLC.This amounts to about 30 LLO and 10,000 P1-PLC moleculesper liposome. From Fig. 1 Lower, we see that >1.7 nM PI-PLCwas required to initiate leakage in the presence of 80 pM LLO.This amounts to about 120 LLO and 2500 P1-PLC moleculesper liposome. Thus, the threshold for leakage can be main-tained by exchanging 7500 P1-PLC for 90 LLO molecules perliposome. This is certainly an overestimate, since LLO bindsabout 10 times more rapidly to the liposomes than doesPT-PLC, which would suggest that the number of boundPI-PLCs requi-red is closer to 750. Detailed titration experi-ments will be required to fix the ratio more exactly.These findings lead us to propose the following scenario. On
entry into the primary phagosome, L. monocytogenes secretesPT-PLC and LLO, which accumulate in the phagosome andbind to the membrane. If one assumes a spherical phagosomeof -~2-/im diameter with a volume of 4.2 ltm3and the synthesisof 10-50 molecules of Pi-PLC per m in[a low estimate basedon Coomassie blue staining seen in culture supernatant fluidswith 2 x 109 bacteria per ml (5)], 200-1000 PI-PLC moleculescould accumulate in 20 min, yielding a concentration of
2982 Biophysics: Goldfine et al

Proc. NatL Acad ScL USA 92 (1995) 2983
80-400 nM in the aqueous compartment of the phagosome.This is at least 25- to 100-fold higher than the concentrationsof PI-PLC required for cooperation with LLO. Thus, theamounts of PI-PLC available should be sufficient to initiaterapid membrane damage in concert with LLO. Without Pl-PLC, LLO can disrupt the macrophage phagosome, albeitmore slowly (5).
In addition to sulfhydryl-activated pore-forming hemolysinssuch as LLO and perfringolysin 0, other bacterial pore-forming proteins or peptides are known to bind to andpermeabilize membranes. These include Staphylococcus au-reus 8 toxin, "CAMP"-factor (25), and Escherichia coli hemo-lysin (26). These molecules are believed to associate withmembranes through one or more amphiphilic helices, and inthe case of E. coli hemolysin, several pore-forming domainshave been identified by deletion analysis (26).
We thank Martin Low for PI-PLC from B. thuringiensis, PascaleCossart for antibody to LLO, Rodney Tweten for perfringolysin 0, andJohn Silvius for reagents used in the fusion assay. We also appreciatethe donation of strains by Daniel Portnoy and his advice and continuedinterest in this research. This work was supported by National Insti-tutes of Health Grants AI-31537 to H.G. and GM-31506 to J.B.
1. Tilney, L. G. & Portnoy, D. A. (1989)J. Cell Biol. 109,1597-1608.2. Farber, J. M. & Peterkin, P. I. (1991) Microbiol. Rev. 55,476-511.3. Portnoy, D. A., Chakraborty, T., Goebel, W. & Cossart, P. (1992)
Infect. Immun. 60, 1263-1267.4. Gaillard, J.-L., Berche, P., Frehel, C., Gouin, E. & Cossart, P.
(1991) Cell 65, 1127-1141.5. Camilli, A., Tilney, L. G. & Portnoy, D. A. (1993) Mol. Microbiol.
8, 143-157.6. Goldfine, H. & Knob, C. (1992) Infect. Immun. 60, 4059-4067.7. Gandhi, A. J., Perussia, B. & Goldfine, H. (1993) J. Bacteriol. 175,
8014-8017.8. Camilli, A., Goldfine, H. & Portnoy, D. A. (1991) J. Exp. Med.
173, 751-754.
9. Geoffroy, C., Raveneau, J., Beretti, J.-L., Lecroisey, A., Vazquez-Boland, J.-A., Alouf, J. E. & Berche, P. (1991) Infect. Immun. 59,2382-2388.
10. Goldfine, H., Johnston, N. C. & Knob, C. (1993)J. Bactenol. 175,4298-4306.
11. Portnoy, D. A., Jacks, P. S. & Hinrichs, D. J. (1988)J. Exp. Med.167, 1459-1471.
12. Alford, D., Ellens, H. & Bentz, J. (1994) Biochemistry 33,1977-1987.
13. Staubinger, R. M., Hong, K., Friend, D. S. & Papahadjopoulous,D. (1983) Cell 32, 1069-1079.
14. Silvius, J. R., Leventis, R., Brown, P. M. & Zuckermann, M.(1987) Biochemistry 26, 4279-4287.
15. Long, D., Berson, J. F., Cook, D. G. & Doms, R. W. (1994) J.Virol. 68, 5890-5898.
16. Geoffroy, C., Gaillard, J.-L., Alouf, J. E. & Berche, P. (1987)Infect. Immun. 55, 1641-1646.
17. Portnoy, D. A., Tweten, R. K., Kehoe, M. & Bielecki, J. (1992)Infect. Immun. 60, 2710-2717.
18. Ikezawa, H. & Taguchi, R. (1981) Methods Enzymol. 71,731-741.19. Alouf, J. E. & Geoffroy, C. (1991) in Sourcebook of Bacterial
Protein Toxins, eds. Alouf, J. E. & Freer, J. H. (Academic,London), pp. 147-186.
20. Gaillard, J. L., Berche, P., Mounier, J., Richard, S. & Sansonetti,P. (1987) Infect. Immun. 55, 2822-2829.
21. Gascard, P., Tran, D., Sauvage, M., Sulpice, J.-C., Fukami, K,Takenawa, T., Claret, M. & Giraud, F. (1991) Biochim. Biophys.Acta 1069, 27-36.
22. Butikofer, P., Lin, Z. W., Chiu, D. T.-Y., Lubin, B. & Kuypers,F. A. (1991) J. Biol. Chem. 265, 16035-16038.
23. Higgins, J. A., Hitchin, B. W. & Low, M. G. (1989) Biochem. J.259, 913-916.
24. Volwerk, J. J., Filthuth, E., Griffith, 0. H. & Jain, M. K. (1994)Biochemistry 33, 3464-3474.
25. Fehrenbach, F. J. & Jurgens, D. (1991) in Sourcebook ofBacterialProtein Toxins, eds. Alouf, J. E. & Freer, J. H. (Academic,London), pp. 187-213.
26. Ludwig, A., Schmid, A., Benz, R. & Goebel, W. (1991) Mol. Gen.Genet. 226, 198-208.
Biophysics: Goldfine et at















![I i pase-cat al y zed hydrolysis phosphatidylcholine of ... · PDF filephosphatidylcholine of guinea pig very low density ... [ ''C]palm~toyl phosphatidylcholine and purified bovine](https://img.pdfslide.us/doc/110x75/5a83b0f17f8b9a682c8ef22c/i-i-pase-cat-al-y-zed-hydrolysis-phosphatidylcholine-of-of-guinea-pig-very-low.jpg)













