Embed Size (px)
Citation preview

Photochemistry andphorobiology. 1965 Vol. 4, pp. 613-619. Pergamon Press Ltd. Printed in Great Britain.
LIGHT REACTION SYSTEMS IN CYANOPHYCEAE"
W. NULTSCH
Department of Botany, University of Tuebingen, Germany.
(Received 15 February 1965)
Abstract-In the blue green algae, Phormidium uncinafum and Ph. autumnale, movement in the dark is arrested by 2 x lo-' M DCPIPt, whereas photokinesis, i.e. the acceleration of the movement by light, is not markedly impaired. Thus at this concentration movement occurs in the light, but not in the dark. Obviously, the dark movement is coupled with oxidative phosphoiylation and the inhibitory effect of DCPLP is probably due to trapping of electrons from the cytochrome chain and uncoupling of the oxidative phosphorylation whereas photophosphorylation in vivo is not uncoupled and accordingly photokinesis is not inhibited. On the other hand, the dark movement is not influenced by M DCMU, a high concentration, which otherwise stops both cyclic and non-cyclic photophosphorylation and hence photokinesis. The inhibition by DCMU of that part of the light induced movement which is due to non-cyclic phosphorylation is reversible by the addition of ferrocyanide (lo-* to
The photo-phobotactic response is less sensitive to DCMU and is not completely sup- pressed even at M. The inhibition is not significantly reversible by the addition of ferrocy- anide. However, M DCPIP, that does not considerably impair photokinesis, suppresses the photo-phobotactic responx completely. Thus, contrary to Links' hypothesis, photo- phobotaxis cannot be linked with photophosphorylation. But there is some evidence that photo-phobotaxis is coupled with the non-cyclic electron transport of photosynthesis at another point, which seems to lie between electron supply from oxygen and the inhibition point of DCMU, probably in the region of the acmssory pigments.
M) as external electron donor.
INTRODUCTION IN BLUE green algae of the Phormidium-Oscillatoria-type, movement is influenced by light in three different ways :
(1) photo-topotaxis, i.e. the orientation of the movement by the direction of incident
(2) photo-phobotaxis, i.e. the reversal of movement caused by sudden changes in light
(3) photokinesis, i.e. the influence of light on the speed of the movement, in most
In earlier investigations it was found that these three reaction types are controlled by three different photoreactive systems, which have different energy requirements and differ- ent action spectra.(l) The present report is restricted to the discussion of the mechanisms of the photo-phobotactic and the photokinetic response.
*Manuscript of a lecture, delivered at the Fourth International Photobiology Congress, 26-30 July 1964,
tAbbreviations: DCPIP = dichlorophenol indophenol, DCMU = 3-(3,4-dichloropheny1)-l,l-dimethylurea. 613
light,
intensity, and
cases, an acceleration of the speed.
Oxford. The Investigation was supported by Deutsche Forschungsgemeinschaft.

614 W. NULTSCH
The action spectra of photo-phobotaxis and photokinesis show(1,2,3) that the photo- receptors of these two reactions in the visible region are photosynthetically active pigments. The photoreceptor of photokinesis is chlorophyll a, without considerable participation of biliproteins and carotenoids. On the other hand, the main maxima in the action spectrum of photo-phobotaxis coincide with the absorption maxima of the biliproteins, phycocyanin and phycoerythrin, and with the longwave maximum of chlorophyll a, blue light being less active. Thus the effect of visible light on photo-phobotaxis corresponds to the absorp- tion by the accessory pigments and chlorophyll a. This leads to the supposition that both reactions, photo-botaxis and photokinesis, are linked with the electron transport chain of photosynthesis, but at different points and in different ways. Therefore, it should be possible to influence both reactions by means of photosynthetic inhibitors or cofactors.
MATERIALS A N D METHODS In our experiments Ph. uncinatum and Ph. autumnale, both containing large amounts of
phycoerythrins but only small amounts of phycocyanins,(4) were used. The culture tech- nique, the experimental conditions and the methods of evaluation of the photokinetic and photo-phobotactic reactions have been described in earlier papers. (233) The photokinetic experiments were carried out in white light (1000 Ix), the photo-phobotactic experiments in red light (RG 2 filter, Schott and Gen., Mainz) to avoid possible interactions between photo-phobotaxis and photo-topotaxis.(3,5) The substances were added to a 0.5 % agar.
RESULTS It is generally accepted that movement needs ATP or any other energy-rich compound
as an energy source. This being the case, the speed of the movement must be a function of the intensity of the phosphorylation processes. Since movement occurs to a certain extent in both the species of Phormidium even in the dark, we may suppose that this part of the movement is due to oxidative phosphorylation and anaerobic phosphorylation whereas photokinesis is caused by photophosphorylation.
Effect of DCPIP It is possible, in fact, to separate the photoinduced and the dark movement by chemical
treatment with DCPIP. DCPIP and other indophenols have been reported to uncouple the electron transport from the phosphorylation processes in chloroplasts in vi~ro.(6,7) As seen in Fig. 1 , in Ph. autumnale movement in the dark is increasingly inhibited with in- creasing concentrations of DCPIP in the range of 10-5 to 10-4 M and becomes completely arrested at 2 x 10-4 M. The total movement in light, which is the sum of the light-induced movement and that due to oxidative and anaerobic phosphorylation, is decreased to the same degree as the movement in the dark, whereas, the photoinduced part of the movement is not markedly inhibited. Thus, at 2 x 1W M DCPIP, movement occurs in the light but not in the dark. The same is valid in the case of Ph. uncinatum. We may conclude that 2 x 10-4 M DCPIP completely inhibits oxidative phosphorylation, by trapping of electrons from the cytochrome chain but that electrons from the electron transport chain in photo- synthesis are not trapped in the living cell. Accordingly, DCPIP is reduced by the two Phormidium species only in the dark. Analogous experiments were carried out to compare the effect of DCPIP on the photo-phobotactic reaction of both species. As shown in Fig. 2, DCPIP markedly inhibits the phobotactic response at concentrations between 5 x 106

Light reaction systems in cyanophyceae 615
O h
10 0 -, -jp7 - - - - - - - - -1 Ighl Con t LoL
5 1;-5 2 i I;-' 5 M
FIG. 1 . Effect of DCPIP on the speed of movement in Ph. autumnale. The speed values are expressed in % of the light control. Solid line and solid circles: movement in the light (1000 Ix).
dotted line and open circles: movement in the dark.
FIG. 2. Effect of DCPIP on the photo-phobotactic reaction of Ph. uncinaturn in red light (Schott-filter RG 2). Abscisse: light intensity in erg/cmE sec, ordinate: value of photo-
phobotactic reaction. At M DCPIP there was no reaction.
and 10-5 M, which do not influence movement either in the light or in the dark (Fig. 1). The inhibitory effect of DCPIP becomes stronger with increasing concentrations and at 10-4 M the photo-phobotactic reaction is completely suppressed, whereas movement, although partially inhibited, occurs both in light and in dark. Thus photo-phobotaxis is much more sensitive to DCPTP than photokinesis.
Effect of DCM I / DCMU is a strong inhibitor of the non-cyclic electron transport in photosynthesis. It
has been reported to inhibit the evolution of oxygen by interrupting the electron supply from oxygen to system 2.(8) According to Duysens(9~10) and Amez,"" however, DCMU inhibits the non-cyclic electron transport between Q and quinone. In order to investigate whether photokinesis and photo-phobotaxis are coupled with the photosynthetic electron transport chain, we carried out some experiments with a medium, containing DCMU.
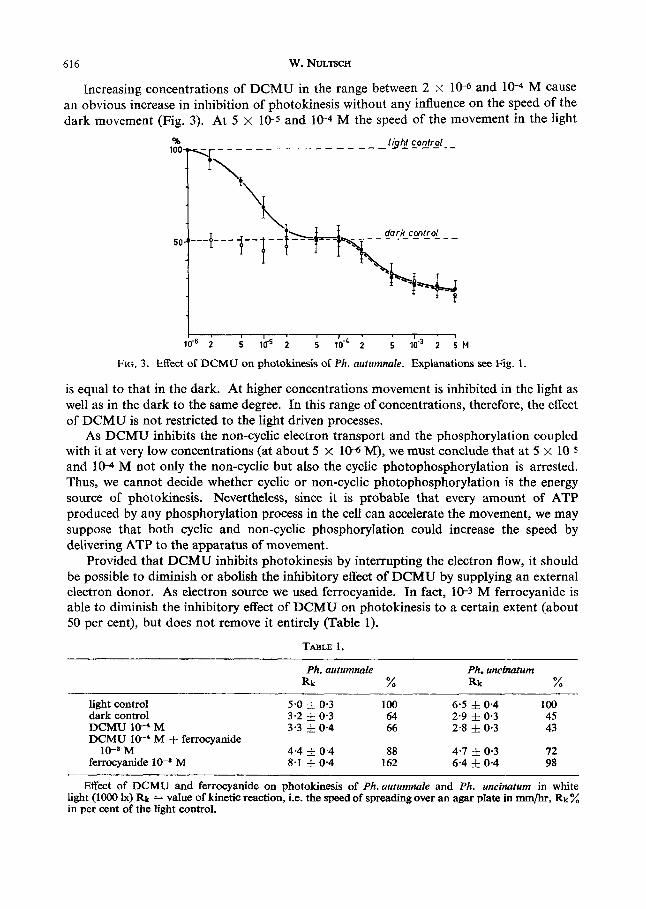
SI6 W. NULTSCH
Increasing concentrations of DCMU in the range between 2 x 106 and 10-4 M cause an obvious increase in inhibition of photokinesis without any influence on the speed of the dark movement (Fig. 3). At 5 x 10-5 and 10-4 M the speed of the movement in the light
1;-6 2 5 l k S 2 ; ;o-& 2 5 I k 3 2 ; M
FIG. 3. Effect of DCMU on photokinesis of Ph. autumnak. Explanations see Fig. 1 .
is equal to that in the dark. At higher concentrations movement is inhibited in the light as well as in the dark to the same degree. In this range of concentrations, therefore, the effect of DCMU is not restricted to the light driven processes.
As DCMU inhibits the non-cyclic electron transport and the phosphorylation coupled with it at very low concentrations (at about 5 x 10-6 M), we must conclude that at 5 x 10-5 and 10-4 M not only the non-cyclic but also the cyclic photophosphorylation is arrested. Thus, we cannot decide whether cyclic or non-cyclic photophosphorylation is the energy source of photokinesis. Nevertheless, since it is probable that every amount of ATP produced by any phosphorylation process in the cell can accelerate the movement, we may suppose that both cyclic and non-cyclic phosphorylation could increase the speed by delivering ATP to the apparatus of movement.
Provided that DCMU inhibits photokinesis by interrupting the electron flow, it should be possible to diminish or abolish the inhibitory effect of DCMU by supplying an external electron donor. As electron source we used ferrocyanide. In fact, 10-3 M ferrocyanide is able to diminish the inhibitory effect of DCMU on photokinesis to a certain extent (about 50 per cent), but does not remove it entirely (Table 1).
TABLE 1.
Ph. autumnale Rk %
~ ~~ ~
light control 5.0 f 0.3 100 dark control 3.2 f 0.3 64 DCMU 10-4 M 3.3 f 0-4 66 DCMU lW4 M + ferrocyanide
M 4.4 f 0.4 88 ferrocyanide lo-* M 8.1 f 0.4 162
Ph. uncinatum R k %
6.5 f 0.4 100 2.9 f 0.3 45 2.8 f 0.3 43
4.7 f 0.3 72 6.4 f 0.4 98
~~ ~~ ~~ ______
Effect of DCMU and ferrocyanide on photokinesis of Ph.autumnale and Ph. uncinatum in white light (loo0 Ix) Rk = value of kinetic reaction, i.e. the speed of spreading over an agar plate in m/hr, Rk% in per cent of the light control.

Light reaction systems in cyanophyceae 611
Further increase in the concentration of ferrocyanide does not abolish the residual inhibition. Obviously, only that part of photokinesis which i s due to non-cyclic phos- phorylation is activated again by the external electron supply, whereas, the inhibition of the cyclic phosphorylation is not reversible in this way. In fact, ferrocyanide entirely cancels the inhibitory effect of DCMU at lower concentrations (2 x 10-6 and 5 x 106 M) which are known to block non-cyclic phosphorylation without any effect on cyclic phosphory- lation in the living celL(12.13)
As observed in the same table, ferrocyanide, applied alone, causes a considerable increase in the speed of the movement in light in Ph. autumnale, but not in Ph. uncinatum. A more detailed investigation of this phenomenon has shown that in Ph. autumnale, ferro- cyanide accelerates movement in light in a wide range of concentrations (10-4 - 10-2 M), without any effect in the dark (Fig. 4). Obviously, in Ph. autumnale, the electron supply
FIG. 4. Effect of ferrocyanide on the speed of the movement both in light and in dark in Ph. autumnale. Explanations see Fig. 1.
by system 2, i.e. the system of the accessory pigments, under our experimental conditions, is not sufficient to cover the requirements of the phosphorylation apparatus, so that an additional electron source increases the rate of photophosphorylation and hence photo- kinesis. On the other hand, in Ph. uncinaturn the electron requirement of the photophos- phorylation apparatus seems to be covered so that no stimulation of movement in the light occurs after the addition of ferrocyanide.
Photo-phobotaxis is also inhibited by DCMU, but the concentration of 10-4 M, which totally inhibits photokinesis, has only a slight effect on photo-phobotaxis (Fig. 5). Although at higher concentrations the inhibitory effect of DCMU on phobotaxis increases, the phobo- tactic response is not completely suppressed even at 10-3 M. Thus, both photokinesis and photo-phobotaxis are inhibited by DCMU, but photokinesis is much more sensitive to it than phobotaxis. This leads to the conclusion that both reactions are coupled with photo- synthesis, even though in a different manner and at different points. This is further confirmed by the fact that the inhibition of photo-phobotaxis by DCMU is not significantly reversible when ferrocyanide is added.

618 w. NULTSCH
i C o n l r
J O - ~ M
5 l o - 4 ~ l O 4 M
10 100 1000 10000 100 000 e r g / c m ’ s c c
Fiti . 5 . Effect of DCMU on pholo-phobotactic reaction of fh. mri/iuti/m in red light. Explana- tions see Fig. 2.
DISCUSSION According to the hypothesis of Links,(l4,15) phobotactic responses are caused by sudden
changes in ATP production. This would mean that photo-phobotaxis is coupled with photo- phosphorylation. But in the present paper it has been shown that 5 x 10-5 M DCMU, which stops the photophosphorylation processes in vivo, does not impair the photo-phobotactic response, whereas, at the same concentration, photokinesis is totally arrested. Thus, contrary to the hypothesis of Links, photokinesis, but not photo-phobotaxis is coupled with photophosphorylation. Probably, both cyclic and non-cyclic phosphorylations are the ATP sources for the light-induced movement, because, the inhibition by DCMU is reversible only up to about 50 per cent by the addition of ferrocyanide as an external elec- tron donor. The reversible and the non-reversible part of photokinesis are obviously due to the non-cyclic and cyclic phosphorylations respectively.
DCPIP, at 2 x 10-4 M, completely inhibits movement in the dark but not in the light. The fact that DCPIP is reduced by the species of Phormidium investigated in the dark and oxidized in the light, permits us to conclude that DCPIP traps the electrons from the cyto- chrome chain and uncouples oxidative phosphorylation, but does not uncouple photo- phosphorylation in vivo, as it has been reported to do in vitro.(6,7)
From the results, we conclude that movement requires ATP and that every supply of ATP, produced by any phosphorylation process, increases the speed of the movement. The extent of participation of each phosphorylation process in the supply of energy for movement depends obviously on the experimental conditions. That the amount of ATP produced by oxidative phosphorylation varies with the oxygen tension and the available amount of oxidizable substrate within the cell, deserves special mention here.
The results of our previous studies(l6) show that photo-phobotaxis is also coupled with photosynthesis. In fact, it is also inhibited by DCMU, but as it is less sensitive to this substance than photokinesis, the linkage should lie before the DCMU block, probably in the region of the accessory pigments. This supports the findings of the Duysens group [ ~ J O A ) that DCMU inhibits the non-cyclic electron transport between the two pigment systems. Accordingly, the inhibition of photo-phobotaxis by DCMU at high concentrations is not reversible by ferrocyanide.

Light reaction sytems in cyanophyceae 619
On the other hand, photo-phobotactic response is completely suppressed by 10-4 M DCPIP (oxidized), even though the movement continues with decreased speed. Obviously, the linkage between the reaction centre of photo-phobotaxis and the electron transport chain of photosynthesis is either broken or bypassed by DCPIP. However, further investi- gations are necessary to locate the position of linkage in the electron transport chain of photosynthesis.
REFERENCES 1 . W. NULTSCH, Ber. deutsch. botan. Ges. 75, 443 (1963). 2. W. NULTSCH, Planta 57, 613 (1962). 3. W. NULTSCH, Planta 58, 647 (1962). 4. W. NULTSCH, Biochim. et Biophys. Acta 59, 213 (1962). 5. W. NULTSCH, Planta 56, 632 (1961). 6. D. W. KROGMANN and A. T. JAGENDORF, Plant Physiol. 34, 277 (1959). 7. M. LOSADA, F. R. WHATLEY and D. I. ARNON, Natrrre 190, 606 (1961). 8. A. MULLER, B. RUMBERG and H. T. W i n , Proc. Roy. SOC. (London) 157, 313 (1963). 9. L. N . M. DUYSENS, Progr. Biophys. 14, 1 (1964).
10. L. N. M. DUYSENS, Photosynthesis Mechanisms in Green Plants. Publ. 1145, Nai. Acad. Sci.-Nut.
11. J. AMEZ, Biochim, et Biophys. Acta 79, 257 (1964). 12. W. URBACH and W. SIMONIS, Biochem. Biophys. Res. Comm. 13, 241 (1963). 13. G. FORTI and B. PARISI, Biochim. et Biophys. Acta 71, 1 (l963). 14. J. LINKS, A hypothesis for the mechanism of (phobo) chemotaxis. Diss. Leiden 1955. 15. R. K. CLAYTON, Handb. Pflanzenphysiol. 17/1, 371 (1959). 16. W. NULTSCH and G. RICHTER, Arch. Mikrobiol. 47, 207 (1963).
Research Council (1963).