Embed Size (px)
Citation preview

International Dairy Journal 14 (2004) 467–494
ARTICLE IN PRESS
Contents
1. Intr
2. Ha
2.1
2.2
2.3
3. Hea
3.1
3.2
3.3
4. Cha
4.1
4.2
4.3
5. Gro
5.1
5.2
5.3
*Correspondin
E-mail addres
0958-6946/$ - see
doi:10.1016/j.ida
Review
Leuconostoc, characteristics, use in dairy technology andprospects in functional foods
Denis Hemme, Catherine Foucaud-Scheunemann*
Institut National de la Recherche Agronomique, Unit!e de Recherches Laiti"eres et G !en!etique Appliqu!ee, 78352 Jouy en Josas Cedex, France
Received 14 March 2003; accepted 2 October 2003
This review is dedicated to the memory of Charles Divies, professor at ENSBANA, Universite de Bourgogne, Dijon, France who passed away
January 8th, 2003
Abstract
This review presents up to date comprehensive data about Leuconostoc, relative to their habitat, taxonomy, metabolism, and
genetics, their implications in health and safety, and their present and potential use in dairy technology and functional foods.
r 2003 Elsevier Ltd. All rights reserved.
Keywords: Leuconostoc; Review; Cheese; Dairy products; Habitat; Classification; Taxonomy; Bacteriophages; Technology; Bacteriocins;
Metabolism; Plasmids; Genetics; Polysaccharides; Clinical occurrence; Infection
oduction . . . . . . . . . . . . . . . . . . . . . .
bitat of Leuconostoc . . . . . . . . . . . . . . . . .
. Presence in various niches . . . . . . . . . . . .
. Presence in fermented products, general considera
. Presence in dairy products . . . . . . . . . . . .
lth, disease and safety . . . . . . . . . . . . . . .
. Infection and opportunism . . . . . . . . . . . .
. Antibiotic resistance . . . . . . . . . . . . . . .
. Health-related metabolic activities . . . . . . . .
racterization and taxonomy . . . . . . . . . . . .
. Identification, characterisation and biodiversity .
. Recent evolution and up to date taxonomy . . .
. Comments about methods of comparison . . . .
wth of Leuconostoc . . . . . . . . . . . . . . . . .
. Cultivation, maintenance and preservation of cult
5.1.1. Media for isolation and enumeration . .
5.1.2. Preservation of cultures . . . . . . . . .
. Industrial production . . . . . . . . . . . . . . .
. Involvement of Leuconostoc in microbial interacti
5.3.1. Co-aggregation reactions . . . . . . . .
5.3.2. Growth and metabolism of Leuconostoc i
g author. Tel.: +33-1-3465-2091; fax: + 33-1-3465-2065.
s: [email protected] (C. Foucaud-Scheunema
front matter r 2003 Elsevier Ltd. All rights reserved.
iryj.2003.10.005
. . . . . . . . . . . . . . . . . . . . . . . . . . 468
. . . . . . . . . . . . . . . . . . . . . . . . . . 469
. . . . . . . . . . . . . . . . . . . . . . . . . . 469
tions . . . . . . . . . . . . . . . . . . . . . . . 469
. . . . . . . . . . . . . . . . . . . . . . . . . . 469
. . . . . . . . . . . . . . . . . . . . . . . . . . 471
. . . . . . . . . . . . . . . . . . . . . . . . . . 471
. . . . . . . . . . . . . . . . . . . . . . . . . . 471
. . . . . . . . . . . . . . . . . . . . . . . . . . 471
. . . . . . . . . . . . . . . . . . . . . . . . . . 472
. . . . . . . . . . . . . . . . . . . . . . . . . . 472
. . . . . . . . . . . . . . . . . . . . . . . . . . 473
. . . . . . . . . . . . . . . . . . . . . . . . . . 473
. . . . . . . . . . . . . . . . . . . . . . . . . . 474
ures . . . . . . . . . . . . . . . . . . . . . . . . 474
. . . . . . . . . . . . . . . . . . . . . . . . . . 474
. . . . . . . . . . . . . . . . . . . . . . . . . . 475
. . . . . . . . . . . . . . . . . . . . . . . . . . 475
on . . . . . . . . . . . . . . . . . . . . . . . . . 475
. . . . . . . . . . . . . . . . . . . . . . . . . . 475
n mixed strain cultures . . . . . . . . . . . . . . 475
nn).

ARTICLE IN PRESS
5.3.3. Metabolic end products and interference with microbial growth . . . . . . . . . . . . . . 476
5.3.4. Bacteriophages of dairy Leuconostoc . . . . . . . . . . . . . . . . . . . . . . . . . . . . 476
5.3.5. Bacteriocins produced by dairy Leuconostoc . . . . . . . . . . . . . . . . . . . . . . . . . 476
6. Survival of Leuconostoc to stress conditions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 477
7. Metabolism of Leuconostoc . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 478
7.1. Carbohydrate utilisation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 478
7.1.1. Sugar transport . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 478
7.1.2. Sugar catabolism . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 478
7.2. Organic acid metabolism . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 479
7.3. Nitrogenous compounds metabolism . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 480
7.3.1. Amino acid requirements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 480
7.3.2. The proteolytic system . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 480
7.4. Metabolism in the presence of oxygen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 481
7.5. Other metabolic considerations . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 481
8. Genetics of Leuconostoc . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 481
8.1. Plasmid biology . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 482
8.2. Cloning vectors and gene transfer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 482
8.3. Gene cloning and expression . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 482
8.4. Genomics . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 482
9. Roles of Leuconostoc in technology . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 482
9.1. Roles in dairy technology . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 483
9.1.1. Openness . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 483
9.1.2. Aroma production . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 483
9.1.3. Deliberate addition of Leuconostoc as NSLAB . . . . . . . . . . . . . . . . . . . . . . . 483
9.2. Roles in functional foods . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 484
9.2.1. Leuconostoc as potential probiotics . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 484
9.2.2. Production of polysaccharides . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 484
9.2.3. Production of mannitol . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 485
9.2.4. Hydrolysis of a-galactosides . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 485
9.2.5. Production of vitamins . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 485
10. Conclusion and prospects . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 485
10.1. Identification and ecology . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 485
10.2. Health, safety and benefits . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 485
10.3. Technological implications . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 486
Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 486
References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 486
D. Hemme, C. Foucaud-Scheunemann / International Dairy Journal 14 (2004) 467–494468
1. Introduction
Strains belonging to the genus Leuconostoc are Gram-positive lactic acid bacteria (LAB) of economic im-portance, related to numerous positive aspects:
* Fermentation of foodstuffs (sauerkraut, pickles, meatproducts, etc.).
* Production of gas (CO2) in cheeses presenting open-ness ( in particular Blue-veined cheeses).
* Production of flavour compounds in multiple dairyproducts.
* In situ production of dextran in saccharose contain-ing (dairy) products, or as high value polymers forindustrial or clinical use, or as biological auxiliary inthe oil industry.
* Potential roles in functional foods.
Leuconostoc strains are also linked to a few negativeaspects including spoilage in the sugar cane industry(Day, 1992) and food products (Ennahar, Cai, &Fujita, 2003; Lyhs, 2002; Susiluoto, Korkeala, &Bjorkroth, 2003) by formation of slime, and safety asthey have been identified in human clinical isolates(Section 3). However, the history of human exposureand consumption has led to the conclusion thatLeuconostoc is a microorganism generally regardedas safe.
In dairy technology, the importance of Leuconostoc
strains is widely recognized, although their physiologyand genetics are less developed than those of Lactococ-
cus. They are often present in dairy starter cultures andalso in the dairy environment and thus could beconsidered as non-starter lactic acid bacteria (NSLAB)in the same way as mesophilic lactobacilli (Cogan,

ARTICLE IN PRESSD. Hemme, C. Foucaud-Scheunemann / International Dairy Journal 14 (2004) 467–494 469
2002). Their role in the formation of aroma and textureof certain dairy products is essential.
This review will synthesize data concerning Leuco-
nostoc species, which play a role in the dairy field,excluding in particular information relative to Oeno-
coccus oeni and Weissella paramesenteroides. Maincomprehensive reviews dealing with Leuconostoc wereeither ancient (Devoyod & Poullain, 1988) or partialwith emphasis on taxonomy (Thunell, 1995), metabo-lism (Cogan & Jordan, 1994), biotechnology (Dessart &Steenson, 1995), bacteriocins (Stiles, 1994) or dairytechnology (Vedamuthu, 1994) or are included inextensive review on LAB (Stiles & Holzapfel, 1997;McSweeney & Sousa, 2000; Carr, Chill, & Maida, 2002)or in a book chapter (Bjorkroth & Holzapfel, 2003). Toavoid extensive list of references, those cited in thesereviews or in original articles are omitted and the readersmight refer to references given herein.
2. Habitat of Leuconostoc
2.1. Presence in various niches
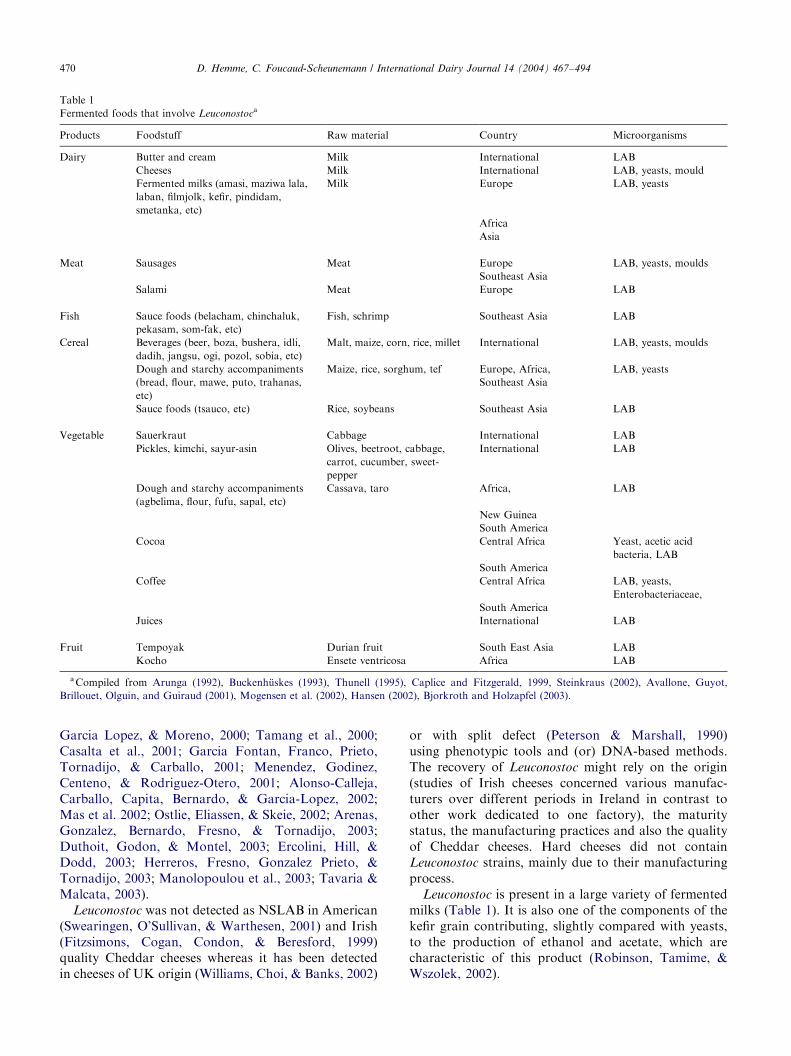
Leuconostoc strains present in many environmentsoriginate from green vegetation and roots that consti-tute their natural ecological niche (Mundt, 1970),although their population is often small (less than 1%)compared with that of aerobic bacteria and yeasts(Buckenh .uskes, 1993). From this natural habitat, theycan easily propagate in various niches including plantmaterials such as vegetables and silage (Ennahar et al.,2003) and fermentation food products from various rawmaterials (Table 1). Their presence in milk andconsequently in dairy products, is due to contaminationduring milking and (or) manufacturing, enhanced bytheir peculiarity of surviving for a long time on materialsurfaces such as wooden ‘‘gerle’’ and other moulds(Devoyod & Poullain, 1988) or through the pasteuriser(Martley & Crow, 1993).
Leuconostoc is not usually considered as part of thehuman flora although strains have been isolated fromhuman faeces, vaginal samples and breast milk samples(Auge, Donnio, Le D!eaut, & Avril, 1987; Green,Wadowsky, & Barbadora, 1990; Heikkil.a, & Saris,2003; dal Bello, Walter, Hammes, & Hertel, 2003) andalso in internal fluids (blood, urine and cerebrospinalfluid) of disabled patients (Farrow, Facklam, & Collins,1989). They have also been isolated from the microfloraof cattle (Brashears, Jaroni, & Trimble, 2003), fish(Ringo & Gatesoupe, 1998), insects (Ohkuma & Kudo,1998; Reeson, Jankovic, Kasper, Rogers, & Austin,2003) and from other environments such as treatedbauxite residues (Hamdy & Williams, 2001).
Leuconostoc was one of the first bacterial groupsstudied for their causative role in commercial losses in
sugar industry (Day, 1992). They have also beenassociated with spoilage of fish (Lyhs, 2002) and meatproducts (Bjorkroth et al., 2000; Anifantaki, Metax-opoulos, Kammenou, Drosinos, & Vlassi, 2002; Hama-saki, Ayaki, Fuchu, Sugiyama, & Morita, 2003).
2.2. Presence in fermented products, general
considerations
LAB including Leuconostoc, play an important role inthe fermentation of various products including foods(Table 1). The source of the microbial flora can be eitherthe raw material as for the production of raw milkcheeses, sauerkraut and some fermented sausages or acommercial starter cultures. The first procedure is oftenin use for propagation of sourdough for private use, andfor the production of some artisanal cheeses where theinoculum is maintained by keeping a sample from theprevious production, the so-called ‘‘old-young techni-que’’ (Limsowtin, Powell, & Parente, 1996).
During fermentation, the end products of carbohy-drate metabolism contribute not only to the preserva-tion upon acidification but also to the flavour andtexture of the product. Fermentation may also increasethe nutritional quality of food by increasing itsdigestibility, as in the fermentation of milk to cheese,or by reducing its toxicity (Gueguen, Chemardin,Labrot, Arnaud, & Galzy, 1997).
2.3. Presence in dairy products
Ln. mesenteroides subsp. cremoris or Ln. lactis strainsare of classical use in butter and cream production andsome fermented fresh dairy products (Vedamuthu, 1994;Section 9.1).
The presence of Leuconostoc in numerous cheesevarieties made without addition of Leuconostoc starter isregular, in particular in raw milk cheeses. In an extensivestudy, among 4379 isolates from 35 different Europeandairy products including 24 artisanal cheeses, 10% ofthe strains were Leuconostoc (Cogan et al., 1997). Thislevel has to be compared to the levels of lactobacilli(mesophile, 12% and thermophile, 14%) and entero-cocci (17%). These results confirm previous dataindicating the presence of Leuconostoc in the majorityof raw milk French cheeses, and other European cheeses(Devoyod & Poullain, 1988).
Numerous studies are still devoted to the ecology ofartisanal cheeses, i.e. nature and evolution of the mainmicrobial groups during manufacture. The studiespublished in the last 5 years concerned mostly raw milkcheeses from South-European countries, and alsoFrance, Great-Britain, Sweden, and India and reliedprogressively on new DNA-based methods (Estepar, delMar Sanchez, Alonso, & Mayo, 1999; Morea, Baruzzi,& Cocconcelli, 1999; Lopez-Diaz, Alonso, Roman,
ARTICLE IN PRESS
Table 1
Fermented foods that involve Leuconostoca
Products Foodstuff Raw material Country Microorganisms
Dairy Butter and cream Milk International LAB
Cheeses Milk International LAB, yeasts, mould
Fermented milks (amasi, maziwa lala,
laban, filmjolk, kefir, pindidam,
smetanka, etc)
Milk Europe LAB, yeasts
Africa
Asia
Meat Sausages Meat Europe LAB, yeasts, moulds
Southeast Asia
Salami Meat Europe LAB
Fish Sauce foods (belacham, chinchaluk,
pekasam, som-fak, etc)
Fish, schrimp Southeast Asia LAB
Cereal Beverages (beer, boza, bushera, idli,
dadih, jangsu, ogi, pozol, sobia, etc)
Malt, maize, corn, rice, millet International LAB, yeasts, moulds
Dough and starchy accompaniments
(bread, flour, mawe, puto, trahanas,
etc)
Maize, rice, sorghum, tef Europe, Africa,
Southeast Asia
LAB, yeasts
Sauce foods (tsauco, etc) Rice, soybeans Southeast Asia LAB
Vegetable Sauerkraut Cabbage International LAB
Pickles, kimchi, sayur-asin Olives, beetroot, cabbage,
carrot, cucumber, sweet-
pepper
International LAB
Dough and starchy accompaniments
(agbelima, flour, fufu, sapal, etc)
Cassava, taro Africa, LAB
New Guinea
South America
Cocoa Central Africa Yeast, acetic acid
bacteria, LAB
South America
Coffee Central Africa LAB, yeasts,
Enterobacteriaceae,
South America
Juices International LAB
Fruit Tempoyak Durian fruit South East Asia LAB
Kocho Ensete ventricosa Africa LAB
aCompiled from Arunga (1992), Buckenh .uskes (1993), Thunell (1995), Caplice and Fitzgerald, 1999, Steinkraus (2002), Avallone, Guyot,
Brillouet, Olguin, and Guiraud (2001), Mogensen et al. (2002), Hansen (2002), Bjorkroth and Holzapfel (2003).
D. Hemme, C. Foucaud-Scheunemann / International Dairy Journal 14 (2004) 467–494470
Garcia Lopez, & Moreno, 2000; Tamang et al., 2000;Casalta et al., 2001; Garcia Fontan, Franco, Prieto,Tornadijo, & Carballo, 2001; Menendez, Godinez,Centeno, & Rodriguez-Otero, 2001; Alonso-Calleja,Carballo, Capita, Bernardo, & Garcia-Lopez, 2002;Mas et al. 2002; Ostlie, Eliassen, & Skeie, 2002; Arenas,Gonzalez, Bernardo, Fresno, & Tornadijo, 2003;Duthoit, Godon, & Montel, 2003; Ercolini, Hill, &Dodd, 2003; Herreros, Fresno, Gonzalez Prieto, &Tornadijo, 2003; Manolopoulou et al., 2003; Tavaria &Malcata, 2003).
Leuconostoc was not detected as NSLAB in American(Swearingen, O’Sullivan, & Warthesen, 2001) and Irish(Fitzsimons, Cogan, Condon, & Beresford, 1999)quality Cheddar cheeses whereas it has been detectedin cheeses of UK origin (Williams, Choi, & Banks, 2002)
or with split defect (Peterson & Marshall, 1990)using phenotypic tools and (or) DNA-based methods.The recovery of Leuconostoc might rely on the origin(studies of Irish cheeses concerned various manufac-turers over different periods in Ireland in contrast toother work dedicated to one factory), the maturitystatus, the manufacturing practices and also the qualityof Cheddar cheeses. Hard cheeses did not containLeuconostoc strains, mainly due to their manufacturingprocess.
Leuconostoc is present in a large variety of fermentedmilks (Table 1). It is also one of the components of thekefir grain contributing, slightly compared with yeasts,to the production of ethanol and acetate, which arecharacteristic of this product (Robinson, Tamime, &Wszolek, 2002).

ARTICLE IN PRESSD. Hemme, C. Foucaud-Scheunemann / International Dairy Journal 14 (2004) 467–494 471
3. Health, disease and safety
A number of reports associate LAB, includingLeuconostoc, with clinical infections. Attention was alsodevoted to the enzymatic activities implicated in theproduction of potential deleterious substances.
3.1. Infection and opportunism
Originally considered to be clinically insignificant,probably as it was not specifically detected, Leuconostoc
is now slightly emerging as opportunistic pathogen untilthe first report of bacteraemia caused by Leuconostoc
came out in 1985 (Dhodapkar & Henry, 1996). Lessthan 100 cases of Leuconostoc infections have beenreported since, mostly involving adults with compro-mised immunity or a wide spectrum of underlyingdiseases although infections have also been documentedin few healthy patients. Predisposing factors also includethe use of enteral or central venous catheter and otherinvasive access devices (tracheotomy, gastrostomy orendotracheal tubes) and previous antimicrobial treat-ment (Dhodapkar & Henry, 1996; Zinner, 1999;Casanova-Roman, Rios, Sanchez-Porto, Gomar, &Casanova-Bellido, 2003).
Clinically, Leuconostoc has been described to causeprimary bacteraemia, pulmonary infections andempyema, peritonitis and infection of the peritonealfluid, endocarditis, meningitis, osteomyelitis, ventriculi-tis, abscess and septicaemia. Most strains belongto the species mesenteroides, the isolation of Ln. lactis
being rare although most species except Ln. mesenter-
oides subsp. cremoris are able to grow at 37�C (Barreau& Wagener, 1990; Aguirre & Collins, 1993; Giacometti,Ranaldi, Siquini, & Scalise, 1993; Kikuchi et al.,1994; Facklam & Elliott, 1995; Dhodapkar & Henry,1996; del Nozal-Nalda, Jimenez-Mejias, Suarez, &Viciana, 1997; Jimenez-Mejias, Becerril, Gomez-Cia,del Nozal, & Palomino-Nicas, 1997; Nelson, 1998;Rodriguez, Saavedra, Fernandez-Jurado, & Prados,1999; Schabereiter-Gurtner et al., 2001; Golan, Poutsia-ka, Tozzi, Hadley, & Snydman, 2001; Gillespie, Symons,& Mc Donald, 2002; Vagiakou-Voudris et al., 2002;Deye, Lewis, Patterson, & Jorgensen, 2003; Fauchaiset al., 2003).
The source of these infections remain unknown, mostof them being hospitally acquired. Skin represents apossible portal in cases with co-isolation from the skinflora and access to the bloodstream through thegastrointestinal tract has been also suggested. In twocases, the organism was isolated from the infant formulaor the enteral feeding, indicating that these are possiblesources (Dhodapkar & Henry, 1996). It is worth notingthat vancomycin treatment might offer Leuconostoc aselective advantage in an ecological niche devoid ofbacteria that would have normally inhibited their
growth, which allows them to multiply and causedisease (Auge et al., 1987).
3.2. Antibiotic resistance
Resistance of Leuconostoc to vancomycin is a generalintrinsic feature and is linked to the presence of apentadepsipeptide with a C terminal d-lactate instead ofa d-alanine in the peptidoglycan (Delcour, Ferain,Deghorain, Palumbo, & Hols, 1999). The vancomycinresistance d-Ala-d-Ala ligase gene from Ln. mesenter-
oides has been cloned (Park & Walsh, 1997). In contrast,few reports are available on other antibiotics (Swenson,Facklam, & Thornsberry, 1990; Kikuchi et al., 1994;Herrero, Mayo, Gonzales, & Suarez, 1996; Cappelli,Barros, Camello, Teixeira, & Merquior, 1999; Zarazagaet al., 1999; Zinner, 1999; Katla, Kruse, Johnsen, &Herikstad, 2000; Holt, Al-Sheikh, & Shin, 2001; Deyeet al., 2003). The studies were either partial or theantimicrobial agents tested and susceptibility methodsused (broth dilution, disk diffusion or Etest) differed, sofew concordant results are available. Converging studieson antibiotic susceptibility of dairy or human clinicalisolates of Leuconostoc strains showed that they areresistant to fosfomycin, ‘‘old’’ quinolones, and glyco-peptides. They are susceptible or give intermediatesensitivity to macrolides and tetracyclines.
Nevertheless, the use of commercial starter culturebacteria in the production of dairy products couldrepresent a potential for spread of genes encodingresistance to antimicrobial agents; the utilization ofantibiotics in animal husbandry, horticulture or indus-trial processes might contribute to the emergence ofantibiotic resistance among human isolates (Patel, 1999;Luh et al., 2000; Teale, 2002); Leuconostoc is recognizedas an opportunistic pathogen and antibiotic-resistantstrains of Leuconostoc were isolated from infection sites(Kikuchi et al., 1994; Cappelli et al., 1999).
3.3. Health-related metabolic activities
Bile salt hydrolysis is dependent on intestinal bacterialactions on body secretions. This reaction has a facilitat-ing effect for bile salt excretion but can also be involvedin various illnesses. Bile salt hydrolase activity is absentin Ln. mesenteroides strains (Tanaka, Doesburg, Iwasa-ki, & Mierau, 1999).
Biogenic amines are organic bases, which have beenimplicated in food poisoning incidents usually from theconsumption of different kind of foods, includingcheese, meat and fish products, and red wine. They aremainly generated by microbial decarboxylation ofamino acids. Biogenic amine production in Leuconostoc
has never been reported (Gonzalez de Llano, Cuesta, &Rodriguez, 1998; Bover-Cid & Holzapfel, 1999) so that

ARTICLE IN PRESSD. Hemme, C. Foucaud-Scheunemann / International Dairy Journal 14 (2004) 467–494472
Leuconostoc strains are first–grade components ofstarter cultures used in the dairy industry.
Leuconostoc produces d-lactate from carbohydratemetabolism as do Lactobacillus delbrueckii (Carr et al.,2002). Human adults metabolise d-lactate slowly so thatthe recommended daily intake should not exceed100mg kg�1 body weight (FAO, 1966). Neither d nordl-lactate should be used in infant foods, except fortherapeutic purposes. The normal consumption of dairyproducts could not lead to an excess of d-lactate.
4. Characterization and taxonomy
The reader may refer to the reviews of Thunell (1995),Stiles and Holzapfel (1997) and Bjorkroth and Holzap-fel (2003), which give a comprehensive view regardingthe history of the genus Leuconostoc and the currenttaxonomy of LAB present in foods, respectively.
Leuconostoc strains are non-motile and non spore-forming cocci, usually present as pairs or short chainsthat belong to the low G+C branch of the Gram-positive bacteria. They do not possess arginine dehy-drolase and catalase, are heterofermentative thusproducing CO2 from glucose metabolism besided-lactate and ethanol or acetate. They are facultativeanaerobic bacteria, which harbour mesophilic charac-teristics and grow at 10�C.
4.1. Identification, characterisation and biodiversity
Phenotypic characters have been used for a long timeto isolate and characterise Leuconostoc and, sometimes,
Table 2
Presumptive identification of Leuconostoc by phenotypical tests
General characters
Gram-positive
Cocci (ovoid-shaped), non motile, non-sporeforming
Facultative anaerobic, catalase negative
Vancomycin resistant
Production of gas from glucose
No arginine hydrolysis
Production of d-lactate from glucose
Additional characters
Growth at 8�C, no growth at 45�C
No growth at pH 4.8
Growth with NaCl 7%
No H2S formation
Acid production from: glucose (all strains); arabinose, arbutin, cellulose,
melibiose, raffinose, ribose, salicin, sucrose, trehalose, and xylose (variabl
Features of some species or subspecies
No production of slime from sucrose by Ln. mesenteroides subsp. cremori
No acid production from fructose by Ln. mesenteroides subsp. cremoris (a
subsp. cremoris (and some Ln. gelidum strains)
No malate decarboxylation by Ln. fallax
to differentiate between species or sub-species (Table 2).As it is difficult to identify species or sub-speciesunequivocally using these classical methods, new mole-cular techniques permit a reliable and consistentidentification, including that of Leuconostoc. They areused alone or combined to estimate the moleculardiversity and (or) for identification of species or sub-species (G .urtler & Mayall, 2001). They were taken intoconsideration to claim recommendations regardingspecies definition (Stackebrandt et al., 2002).
The molecular diversity of 221 dairy strains has beenestimated by RAPD (-Cibik, Lepage, & Tailliez, 2000).Strains were classified as Ln. mesenteroides or Ln.
citreum and these data were confirmed using 16S rDNAsequence and 16S rDNA fragment amplification, exceptfor one strain. Strains formerly described as Ln.
paramesenteroides using phenotypic characters areindeed variants of Ln. mesenteroides, which did notproduce slime, rather than W. paramesenteroides strains.Thus, it will be interesting to test strains described asW. paramesenteroides in other collections.
DNA-based methods are becoming widely used fordifferentiation of Leuconostoc (Lee, Park, & Kim, 2000;Sch .onhuber, Le Bourhis, Tremblay, Amann, & Kula-kauskas, 2001; Matte-Tailliez et al., 2001; P!erez,Cardell, & Zarate, 2002; Randazzo, Torriani, Akker-mans, de Vos, & Vaughan, 2002; dal Bello et al., 2003;Ennahar et al., 2003; Jang, Kim, Lee, & Han, 2003;Reeson et al., 2003). Thus, they permitted to describethe microbial community in Stilton cheese, Leuconostoc
colonies being homogenously distributed in all partsobserved (Ercolini et al., 2003) and in Salers cheese,Leuconostoc being predominant during manufacturing
cellobiose, fructose, galactose, lactose, maltose, mannitol, mannose,
e within the species or subspecies)
s, Ln. lactis, Ln. argentinum
nd some Ln. argentinum strains), from maltose by Ln. mesenteroides

ARTICLE IN PRESSD. Hemme, C. Foucaud-Scheunemann / International Dairy Journal 14 (2004) 467–494 473
and ripening (Duthoit et al., 2003). Separation of 16Sribosomal DNA by Temporal Temperature GradientGel Electrophoresis (TTGE) is of peculiar interestbecause it permits a rapid identification of mostbacterial species present in dairy products, includingLeuconostoc (Ogier, Son, Gruss, Tailliez, & Delacroix-Buchet, 2002). The distinction between the subspeciesmesenteroides, dextranicum and cremoris of Ln. mesen-
teroides was confirmed using proteins patterns orribotyping (Villani, Moschetti, Blaiotta, & Coppola,1997; P!erez et al., 2002) whereas it was questioned by
-Cibik et al. (2000) who proposed that these subspeciesmight be biovars, using other molecular methods.RAPD permits the identification of strains of the newspecies Ln. fallax (Barrangou, Sung-Sik, Breidt, Flem-ing, & Klaenhammer, 2002) although the 16S rPCR-RFLP analysis, which allows the identification of nineLeuconostoc species, did not include the species fallax,
filcuneum, and fructosus (Jang et al., 2003).
4.2. Recent evolution and up to date taxonomy
Only four Leuconostoc species are included in theBergey’s Manual of Systematic Bacteriology (Garvie,1986), the species Ln. mesenteroides comprising the threesubspecies mesenteroides, dextranicum and cremoris
(Table 3). The two main facts concerning Leuconostoc
are the creation of the genus Weissella that comprisesW. paramesenteroides (previously Ln. paramesenter-
oides) and some heterofermentative species of the genusLactobacillus (Collins, Samelis, Metaxopoulos, & Wall-banks, 1993), and the assignment of Ln. oenos as a newgenus, Oenococcus oeni (Dicks, Dellaglio, & Collins,1995). Additionally, eleven new species have beendescribed (Table 3): Ln. gelidum and Ln. carnosum
Table 3
Species included in the genus Leuconostoc
Leuconostoc species Previous nomenclature
Ln. mesenteroides
subsp. cremoris
subsp. dextranicum
subsp. mesenteroides
Ln. lactis
Ln. pseudomesenteroides
Ln. carnosum
Ln. gelidum
Ln. fallax
Ln. citreum Ln. amelibiosum
Ln. argentinum
Ln. gasicomitatum
Ln. kimchi
Ln. ficulneum
Ln. fructosum Lactobacillus fructosus
Ln. inhae
isolated from meat products, Ln. citreum and Ln.
pseudomesenteroides from clinical isolates; Ln. fallax
from sauerkraut, Ln. argentinum from Argentine rawmilk, Ln. gasicomitatum associated with meat spoilage,Ln. kimchii and Ln. inhae from kimchi, Ln. ficulneum
from a ripe fig and Ln. fructosum (previously Lb.
fructosus).Strains found in the microflora of the wasp Vespula
germanica present 90% homology with known Leuco-
nostoc species and may constitute a new taxon (Reesonet al., 2003).
4.3. Comments about methods of comparison
Although molecular methods are useful for taxonomyand phylogeny of strains, phenotypic characters remainrealistic and play a predominant role in food science(Section 9). Interest for strains used as tools in humanactivities is often linked to at least one importantproperty and understanding the relationship betweengenotype and phenotype is a great challenge (O’Donnell,Embley, & Goodfellow, 1993; Morris et al, 2002).
Thus, Ln. fallax strains were characterized usingDNA-based methods (Barrangou et al., 2002) but couldalso be distinguished from Ln. mesenteroides by themalolactic reaction, which is absent in the former butpresent in the latter. This underlines the interest and thefacility of phenotypic discriminatory features that couldbe used routinely and prior (or accompanying) tomolecular based techniques, e.g. arabinose, raffinoseand fructose utilization (-Cibik & Chapot-Chartier,2000), vancomycin resistance, CO2 and dextran produc-tion, or citrate utilization. In the same way, the ability ofconcentrated non-proliferating Leuconostoc cells toacidify milk varied from pH 6.5 to 4.1 and permitted
References
Garvie (1986)
Garvie (1986)
Farrow et al. (1989)
Shaw and Harding (1989)
Shaw and Harding (1989)
Martinez-Murcia and Collins (1991)
Takahashi, Okada, Uchimura, and Kozaki (1992)
Dicks, Fantuzzi, Gonzalez, Du Toit, and Dellaglio (1993)
Bjorkroth et al. (2000)
Kim, Chun, and Han (2000)
Antunes et al. (2002)
Antunes et al. (2002)
Kim, Lee, Jang, Kim, and Han (2003)

ARTICLE IN PRESSD. Hemme, C. Foucaud-Scheunemann / International Dairy Journal 14 (2004) 467–494474
to discriminate between the 107 strains tested forapplications in dairy technology (Demirci & Hemme,1995). Similarly, a recent study proposed a characteriza-tion of the LAB strains on the basis of the neutralcompounds produced in the whey (Mauriello et al.,2001). Some tools that are not suitable for characteriza-tion of species could nevertheless be used to describe areduced group of strains, e.g. plasmid profile andcarbohydrate fermentation to distinguish between phy-siologically related dextran-producing strains (Holtet al., 2001) or citrate metabolism, carbohydratefermentation pattern and peptidase activities to selectdairy strains for technological applications (Server-Busson, Foucaud, & Leveau, 1999). Clinical Leuconos-
toc strains were identified as vancomycin resistant(VAN), and lacking Leu-aminopeptidase (LAP) andpyrrolidonylarylamidase (PYR) activities using a diskmethod (Facklam, Pigott, Franklin, & Elliot, 1995).However, strains of dairy origin utilized Leu-containingpeptides for growth and showed LAP activity on leucyl-paranitroanilide derivatives (Foucaud, Hemme, &Desmazeaud, 2001; Herreros et al., 2003). This dis-crepancy may result from the absence of externalhydrolysis activity or the absence of transport of thesubstrate and further experiments are required toconclude about peptidase specificity.
5. Growth of Leuconostoc
5.1. Cultivation, maintenance and preservation of
cultures
5.1.1. Media for isolation and enumeration
When processing samples of LAB, chilled 0.1%peptone water or 0.9% sodium chloride should be usedfor dilutions, and should not be refrozen prior to plating(Bellengier, Richard, & Foucaud, 1997a, b; Foucaudet al., 2001). Phosphate buffer diluent results in lowerrecovery rates. Cell chains should be mechanicallydisrupted before bacterial enumeration (Lange, Plihon,Taillandier, & Riba, 1995; Bellengier et al., 1997a, b).
The cultivation of Leuconostoc may be accomplishedusing enrichment broths and selective or non-selectivemedia, depending on a need to isolate either a particulargenus from a mixture of microorganisms or to maintainisolates in culture (Bjorkroth & Holzapfel, 2003).Various media satisfying the general nutritional require-ments (also called elective) of Leuconostoc normallyallow high recovery rates, without completely inhibitingother groups. The most common are APT, Briggs,MRS, La and BHIYE. Because of their low selectivity,their application is limited to associations dominated byone type or group of organisms. Selective media, whichcontain one or more restrictive factors, which still allowgrowth of Leuconostoc but inhibit unwanted bacterial
groups in a mixed population, have been proposed.However, no completely effective medium is yet avail-able. Most selective media based on the organism’sability to metabolise citrate have proven to be unsa-tisfactory because many organisms in the same habitat(e.g. Lactobacillus, Lactococcus lactis subsp. lactis
biovar diacetylactis) also metabolise citrate and not allLeuconostoc strains utilize citrate (Section 7.2). Theaciduric nature or their preference for microaerophilicto anaerobic conditions was taken into consideration.Inhibitory factors such as potassium sorbate (MRSS pH5.7), thallous acetate (MRST pH 6.5), sodium azide(MSE), antibiotics such as vancomycin or tetracyclinehave been used, alone or in combination (Benkerroum,Misbah, Sandine, & Elaraki, 1993; Mathot, Kihal,Pr!evost, & Divi"es, 1994).
Usual medium for cultivation of pure strains ofLeuconostoc is the classical MRS medium modified ornot by omission of citrate and meat extract (C. Divi"es,personal communication). Media for physiologicalstudies of sugar fermentation pattern, gas production,dextran formation, citrate degradation and others havebeen developed (Bjorkroth & Holzapfel, 2003). Achemically defined medium was also elaborated, whichfulfils the nutritional requirements of a representativenumber of Leuconostoc strains and allows rapid andheavy growth comparable to that in complex medium(Foucaud, Fran@ois, & Richard, 1997).
When co-cultures of Leuconostoc with Lactococcus
are involved, MRS containing 30 mgmL�1 vancomycinremains the more simple media to detect Leuconostoc,
Lactococcus being inhibited by lower concentrations(minimum inhibitory concentration o 2.5 mgmL�1;Mathot et al., 1994). M17 containing 5% glucose couldalso be used since Lactococcus gives good-size coloniesafter 24 h while Leuconostoc requires 48 h (Liu, As-mundson, Holland, & Crow, 1997). In contrast, when5% sucrose is added to M17, the formation of slimepermits detection of Leuconostoc colonies before Lacto-
coccus colonies (D. Hemme, unpublished). Moreover,this medium supplemented with sucrose allows morerapid detection of Leuconostoc than MRS supplementedwith vancomycin or MSE medium, that also containssucrose but delays growth, probably because of thepresence of sodium azide (0.075%). b-galactosidase(b-gal) positive strains of Leuconostoc developing asblue colonies can be distinguished from Lactococcus
strains that are b-gal negative on MRS agar platesspread with 5-bromo-4-chloro-3-indolyl-b-d-galactopyr-anoside (X-Gal; Mathot et al., 1994; Bellengier et al.,1997a, b). This is also convenient for discriminationbetween b-gal-positive and b-gal-negative Leuconostoc
isolates. In the presence of vancomycin-resistantLactobacillus strains, LUSM medium containing vanco-mycin, tetracyclin and tomato juice should be utilized(Benkerroum et al., 1993).

ARTICLE IN PRESSD. Hemme, C. Foucaud-Scheunemann / International Dairy Journal 14 (2004) 467–494 475
Strains of Leuconostoc grow at 30�C like mesophilicLactococcus starters but are favoured by a lowtemperature (Section 5.3.2). Most strains grow well at10�C and even at 4�C (Hemme, unpublished; Hamasakiet al., 2003). In contrast, some strains were describedas thermoduric as they resist pasteurization, theirpresence on the equipment favouring contamination ofthe milk, which is further pasteurized (Martley & Crow,1993).
5.1.2. Preservation of cultures
For short-term maintenance of dairy Leuconostoc,stab cultures (MRS agar containing 1% lactose) may bestored at 4�C for 1–2 weeks. Viability may be retained inlitmus milk supplemented with 5% yeast extract and 5%glucose or in MRS broth containing 1% lactose and10% glycerol as protecting agents for 6 months to 1 yearat –20�C or over a longer period at –80�C. The use ofexponential phase cultures gives maximum viabilityupon freezing (Bellengier et al., 1997a, b; Bjorkroth &Holzapfel, 2003). Lyophilization of dairy Leuconostoc
in milk supplemented with 4% lactose resulted ingood long-term survival and retention of metaboliccharacteristics.
5.2. Industrial production
As complex laboratory media are often too expensive,other media are used by the industry and theircomposition is often not available (proprietary proce-dures). Papa.ın-hydrolysed casein or milk or whey basedmedium or the same medium as these utilized forLactococcus (1.4% yeast extract, 4% tryptone, 9%lactose) may be used in batch cultures under regulatedpH conditions (often pH 6.5) and controlled parameters(C. Bouillanne, personal communication; Accolas &Auclair, 1967; Vedamuthu, 1994; Champagne & Gard-ner, 2002). The use of lactose instead of glucose avoidsthe development of lactose-negative microorganisms.However, low yields (max. 5� 109 cellsmL�1) wereobtained compared with those obtained with Lactococ-
cus or Lactobacillus in the same conditions (Vedamuthu,1994).
The type of centrifuge used to concentrate the cellsdid not seem to affect the viable counts recovery, but acontinuous freeze-drying process had a negative effect(�33%) compared to a batch laboratory process(Champagne & Gardner, 2002).
The production of mixed cultures containing strainsof Lc. lactis, Ln. mesenteroides subsp. cremoris and Lb.
rhamnosus, on commercial starter media has beenassayed to replace blending mixture; population ratios,acidifying activities and aroma production being influ-enced by the incubation temperature (Barrette, Cham-pagne, Roy, & Rodrigue, 2000).
5.3. Involvement of Leuconostoc in microbial interaction
Mesophilic LAB starters used in the dairy industryare mixtures of genera, species, strains and evendifferent variants. Their composition is thus not alwaysknown, particularly in the case of natural starters. Inaddition, microbial interactions, either beneficial (co-operation) or deleterious (inhibition) may lead touncontrollable changes in the composition of the starter(Juillard et al., 1998). Positive interactions can result in abetter growth or a better metabolite production. Theyare either direct, involving physical contacts of indivi-duals or indirect, due to the modification of substratesor of medium parameters. Negative interactions can bedirect and involved phages or they might also be indirectand concerned toxic metabolites (e.g. bacteriocin),competition for substrate consumption, and cataboliteaccumulation.
5.3.1. Co-aggregation reactions
Extracellular material isolated from Ln. mesenteroides
subsp. dextranicum 663 was shown to physically interactwith lactococcal cells causing co-aggregation (Gopalet al., 1996). This may facilitate intergeneric interactionsand (or) co-culture during cheese manufacture.
5.3.2. Growth and metabolism of Leuconostoc in mixed
strain cultures
Leuconostoc strains grow associatively with acid-producing lactococci and their associative growth hasbeen studied with respect to citrate metabolism andaroma formation. It was described as a synergisticfunctional relationship (Walsh & Cogan, 1973; Boquien,Corrieu, & Desmazeaud, 1988; Vedamuthu, 1994;Monnet, Schmitt, & Divi"es, 1994; Jordan & Cogan,1995). Leuconostoc, metabolized citrate over a pH rangefrom 6.3 to 4.5, and diacetyl and acetoin are onlyproduced at acidic pH (Section 7.2). Thus, to initiatecitrate metabolism and to produce aroma compounds,sufficient acid production by Lc. lactis is required todepress the pH in milk (Cogan & Jordan, 1994).
Less attention has been devoted to the growth rate,acid production and final biomass of these bacteria,which also reflect the interaction occurring in mixed-strain cultures of Leuconostoc and Lactococcus. Leuco-
nostoc growth was either unchanged or was inhibitedduring the exponential phase and was systematicallyinhibited in the stationary phase (Vedamuthu, 1994;Bellengier et al., 1997a). Inhibition was usually sup-pressed upon addition of peptides or amino acids,showing that the competition for nitrogenous nutrientsis a general feature of mixed-strain culture of Leuconos-
toc and Lactococcus in milk (Bellengier et al., 1997a). Itis worth noting that, in limiting or excessive nitrogenousnutrient concentrations, Ln. mesenteroides was alwaysdisadvantaged over Lc. lactis (Foucaud et al., 2001;

ARTICLE IN PRESSD. Hemme, C. Foucaud-Scheunemann / International Dairy Journal 14 (2004) 467–494476
Gendrot, Foucaud-Scheunemann, Ferchichi, & Hemme,2002). In contrast, the improved maximum growth ofLn. lactis CNRZ 1091 in mixed-strain cultures with Lc.
lactis subsp. cremoris AM2 in milk illustrated microbialcooperation between the two species (Boquien et al.,1988). Various types of interaction may occur dependingon the strains used in the associative growth in milk butalso on the growth temperature. A balanced growth ofspecies occurs during incubation between 21�C and25�C. At temperature above 25�C, the ratio is skewedtoward the metabolically more active Lc. lactis, whichgrows at a faster rate at higher temperature (between25�C and 32�C) relative to the Leuconostoc (Veda-muthu, 1994; Liu et al., 1997; Baron, Roy, &Vuillemard, 2000; Barrette et al., 2000).
Differences in the growth pattern of the variousstrains constituting a mixed-strain starter can lead tounavoidable and important changes in its compositionin a relatively short cultivation period. Moreover, incomposite commercial culture concentrates containingacid-producers and aroma producing Leuconostoc, theproportion of Leuconostoc varied from 5% to 10% ofthe culture. Thus, the use of Leuconostoc strainconcentrates may avoid the imbalance of populationand the rapid dominance of Lc. lactis over Leuconostoc
(Vedamuthu, 1994; Bellengier et al., 1997a).Although acetaldehyde (100mgL�1) might enhance
the production of acetoin and diacetyl as well as thegrowth of Leuconostoc, normal concentrations ofacetaldehyde produced by mesophilic cultures (3 to8mgL�1) would be unlikely to stimulate the growth ofLeuconostoc in mixed starter cultures (Vedamuthu,1994; Cogan & Jordan, 1994).
NSLAB may also influence the behaviour of Leuco-
nostoc in dairy technology. Devoyod and Poullain(1988) reported the stimulatory action of enterococciand yeasts on Leuconostoc growth and gas productionthrough the production of amino acids and peptidesfrom the degradation of caseins.
5.3.3. Metabolic end products and interference with
microbial growth
The mechanism by which Leuconostoc inhibits micro-bial growth of LAB as well as spoilage or pathogenicstrains has been also attributed to degradation productsof carbohydrate and citrate metabolism (Dessart &Steenson, 1995; Caplice & Fitzgerald, 1999). Theantimicrobial effects of organic acids, including lacticand acetic acids, affect the membrane properties such asthe membrane potential and the integrity of the cell(Mans de Marion, Raffellini, Fantuzzi, & Gonzalez,1995; Davidson, 1997; Alakomi et al., 2000; Cabo,Braber, & Koenraad, 2002).
The antibacterial properties of diacetyl or ethanol arealso described, although their effects are slight inusual lactic fermentations. In the presence of oxygen,
Leuconostoc accumulates hydrogen peroxide thatcould be inhibitory to some microorganisms (Condon,1987).
5.3.4. Bacteriophages of dairy Leuconostoc
The absence of reports indicating problems in dairymanufacture associated with Leuconostoc phages mayexplain why studies on such phages are rare. Addition-ally, the slow growth of Leuconostoc strains in mixedstarter cultures might also account for the difficulty torecognize those phages in fermentations in which acidproduction is normal. However, phages active againstLeuconostoc were isolated from dairy products (wheysamples, cheese, etc.). They were characterized as to hostrange, morphology, structural proteins, and genomefingerprint and belonged to the Siphoviridae family(Boizet, Mata, Mignot, Ritzenthaler, & Sozzi, 1992;Davey, Ward, & Brown, 1995; Dessart & Steenson,1995; Ackermann, 2001).
5.3.5. Bacteriocins produced by dairy Leuconostoc
There are few reports on antimicrobial action ofLeuconostoc against pathogenic and spoilage micro-organisms in which the inhibitory activity is attributedto bacteriocin-like substances (Stiles, 1994). Studies areeither old or preliminary, limited to the isolation ofbacteriocin producers, and the incomplete characteriza-tion of antibacterial substances (Devoyod & Poullain,1988; Serot, Dousset, Zucca, & Torcatis, 1990; Martley& Crow, 1993; Malik, Naveen, & Mathur, 1995; Sarkar& Sharmistha, 1996; Fantuzzi, Raffellini, & Gonzalez,1999; Rodriguez, Gonzalez, Gaya, Nunez, & Medina,2000; Brashears et al., 2003; Heikkil.a & Saris, 2003;Pepe, Blaiotta, Moschetti, Greco, & Villani, 2003).
Bacteriocins produced by some Leuconostoc strainsbelong to the subclass IIa bacteriocins that are pediocin-like, small heat-stable non-modified peptides, and activeagainst Listeria (Ennahar, Sashihara, Sonomoto, &Ishizaki, 2000). Mesentericin Y105 and B105 andmesenterocin 52A and 52B from dairy Leuconostoc
strains have been extensively described and theirbiological activity, structure and properties havebeen analysed (Mathieu, Sudirman, Rekhif, Milli"ere, &Lef"ebvre, 1993; Sudirman, Mathieu, Benoit, & Lef"ebvre,1994; Krier, Revol-Junelles, & Germain, 1998; H!echard,Berjeaud, & Cenatiempo, 1999; Ennahar et al., 2000;Corbier, Krier, Mulliert, Vitoux, & Revol-Junelles,2001; Morisset & Fr"ere, 2002). The mesentericin Y105and B105 structural genes, mesY and mesB, respectivelywere cloned (Fr!emaux, H!echard, & Cenatiempo, 1995;H!echard et al., 1999). Additionally, leucocin H (Blomet al., 1999) and dextranicin 24 (Revol-Junelles &Lef"ebvre, 1996) have been partly depicted.
Different mechanisms, including the membrane fattyacid content or a putative immunity protein, induceresistance phenomena to bacteriocins in Leuconostoc

ARTICLE IN PRESSD. Hemme, C. Foucaud-Scheunemann / International Dairy Journal 14 (2004) 467–494 477
(Dayem et al., 1996; Limonet, Revol-Junelles, & Milli-"ere, 2002a, b).Although bacteriocins produced by Leuconostoc of
dairy origin (mesentericin Y105) are being studied fortheir possible use in food preservation, the addition ofbacteriocin from Leuconostoc or that of bacteriocinproducers to food has never been reported. Actually, foroptimal effectiveness against foodborne pathogenic andspoilage bacteria, bacteriocins may be used as part of ageneral multihurdle food preservation system, whichinvolves a set of antimicrobial factors (Ennahar et al.,2000; Cleveland, Montville, Nes, & Chikindas, 2001).
Fig. 1. Adhesion of Leuconostoc on glazed sandstone moulds,
examination by scanning electron microscopy. Reproduced from
Devoyod and Poullain (1988) with authorization of Le Lait.
6. Survival of Leuconostoc to stress conditions
Little is known about the effect of stress onLeuconostoc cells (-Cibik & Chapot-Chartier, 2000).They are able to survive for a long time in unfavourableenvironments as diverse as the sugar, oil or dairyindustries. They remained viable for many years on thesurface of wooden ‘‘gerle’’, glazed sandstone, iron orplastic moulds used and other wooden or glazed toolsused in traditional cheese making (Fig. 1; Devoyod &Poullain, 1988). Hostile environmental conditions fa-vour interaction phenomena through the formation ofslime or glycocalix in the presence of saccharose andtrace minerals, resulting in a biofilm, which protects cellsagainst detrimental agents (Kim, Thomas, & Scott-Fogler, 2000; Section 2). Curd of ripened cheeseprobably permits the survival of Leuconostoc that isdetected at high cellular levels (Devoyod & Poullain,1988; Mor-Mur, Carretero, Pla, & Guamis, 1994).
In potassium buffer, the ability of non-proliferatingcells to lyse was lower for cells grown in lactose orgalactose containing MRS than in glucose containingMRS and varied with the strain from 7% to 44% after24 h at 30�C and pH 6.5, remaining stable thereafter(-Cibik & Hemme, 1997; -Cibik & Chapot-Chartier,2000). In cheese technology, the strain dependency wasalso evidenced and the low lysis observed corroboratedthat observed in buffer, pH above 6.5 in the presence ofCa2+ and Na+, underlying the importance of thosefactors (Hemme, Bienvenu, & Ogier, unpublished;Turner, 1988; Martley & Crow, 1993).
Two major peptidoglycan hydrolases (PGH), aglycosidase and an N-acetylmuramyl-l-Ala amidase(or endopeptidase) were described. Additional PGHswere only detected in Ln. lactis with Ln. mesenteroides
cell wall as a substrate, suggesting that Ln. lactis strainscan be separated from other species by their PGHprofile. An atypical PGH, called LnMur, has beendescribed in Ln. citreum 22R and the gene cloned. Itlacks the amino acid repeats implicated in the cell wallbinding. The encoded protein of 209 amino acid residuesincluded a 31 amino acids putative signal peptide. A
chimeric fusion protein between LnMur and Lc. lactis
AcmA C-terminal domain is able to complement AcmAdeficiency in Lc. lactis AcmA (-Cibik & Chapot-Chartier,2000; -Cibik, Tailliez, Langella, & Chapot-Chartier,2001).
Homeostasis of the internal pH is essential for growthand survival of all biological cells, including Leuconostoc
but bacterial growth is a self-limiting process throughthe acidification of the external medium and acidaccumulation (Cogan & Jordan, 1994; Konings, 2002;Section 7). As pHout decreased, the pHin of Ln.
mesenteroides decreased in contrast to that Lc. lactis.Growth ceased when internal pH values of 5.4 to 5.7were reached, independent of the composition of themedium. In contrast, the limiting external cellular pHwas greatly influenced by the growth medium, i.e. natureand concentration of organic acids.
The response of Ln. mesenteroides cells to heat shockconsisted in the overexpression of stress proteins(Salotra et al., 1995), the N-terminal sequence of whichis homologous to the highly conserved chaperoneproteins DnaK and GroEL of Escherichia coli. Ctsrhomologues were identified in Oenococcus oeni andother Gram-positive bacteria, suggesting that heat-shock regulation by Ctsr might also exist in Leuconostoc
(Derre, Rapoport, & Msadek, 1999).Preservation technologies, such as high pressure
technology, which combine efficient germ reductionwith a maximal retention of the chemical and physico-chemical product properties, are currently under in-vestigation. High pressure homogenisation treatments at25�C for 15min and pressure ranging between 100 and300MPa showed that Gram-positive bacteria, includingLn. mesenteroides were more resistant than Gram-negative bacteria, the structure of the peptidoglycanprobably contributing to the resistance whereas both

ARTICLE IN PRESSD. Hemme, C. Foucaud-Scheunemann / International Dairy Journal 14 (2004) 467–494478
groups overlapped at high hydrostatic pressure (Wuy-tack, Diels, & Michiels, 2002). The destruction ofLeuconostoc depended both of the pressure level andthe pressure treatment time, the pressure resistance ofLn. mesenteroides being lower than that of Sacchar-
omyces cerevisiae (Basak, Ramaswamy, & Piette, 2002).After 5min at 25�C and a pressure of at 345MPa thecell viability loss was over 6 log cycles in 1 h. In thepresence of pediocin AcH, cell wall degradation wasfaster and more extensive (Kalchayanand, Frethem,Dunne, Sikes, & Ray, 2002).
7. Metabolism of Leuconostoc
The general metabolism of Leuconostoc is presentedFig. 2.
7.1. Carbohydrate utilisation
Leuconostoc does not possess functional cytochromesand lacks some enzymes of Krebs cycle. They only
Fig. 2. General metabolism of Leuconostoc. Major products formed
are indicated in bold. Numbers refer to enzymes involved or steps: (1)
dextransucrase; (2) mannitol-dehydrogenase; (3) b-galactosidase; (4)esterase; (5) NADH oxydase; (6) alcohol dehydrogenase; (7) phos-
phoketolase; (8) phosphotransacetylase, (9) a-acetolactate decarbox-
ylase; (10) acetate kinase; (11) a-acetolactate synthase; (12) non-
enzymatic formation; (13) diacetyl reductase; (14) oxaloacetate
decarboxylase; (15) lactate dehydrogenase; (16) citrate lyase; (17)
malate dehydrogenase; (18) formation of aspartate; (19) malolactic
enzyme; (20) ATPase.
obtained energy by fermentation, always producinglactic acid as all other LAB and CO2 and ethanol oracetate as reviewed by Cogan and Jordan (1994).
A large variety of mono- and disaccharides supportgrowth of Leuconostoc strains (Vaughan, David, & deVos, 1996; Server-Busson et al., 1999; Holt et al., 2001;Section 4). The sugar fermentation pattern (e.g.arabinose, xylose, ribose, and fructose) of the strainsconstitutes a phenotypical criterion for species, sub-species and strain characterization and is of greatinterest to choose strain for potential application(Section 4).
7.1.1. Sugar transport
Leuconostoc takes up carbohydrates by permeasesthat permit the entry of sugars into the cell withoutmodification of the substrate (e.g. phosphorylationwhen the PEP-PTS system is involved). The lactosetransport gene, lacS from Ln. lactis has been character-ized, using functional complementation of lactosepermease-deficient E. coli mutants (Vaughan, David, &de Vos, 1996). It encodes a 24 kDa hydrophobic proteinand presents significant homology with the Gln and Histransport protein genes of enteric bacteria (Cogan,1995).
7.1.2. Sugar catabolism
The heterofermentative utilization of glucose followsthe phosphoketolase (or pentose-P) pathway and leadsto CO2, a C2 compound (ethanol or acetate with anoxidative mechanism) and pyruvate. Pyruvate is furtherreduced by a d-lactate dehydrogenase in d-lactate that ispartly used for the peptidoglycan synthesis, thusreducing the quantity to be rejected by the cell andavoiding d-Ala synthesis (Section 3.2). The d-lactatedehydrogenase gene, d-ldh from Ln. mesenteroides hasbeen cloned (Phalip, Dartois, Schmitt, & Divi"es, 1994).The final reaction might be affected by exogenousacetaldehyde, which is taken from the medium whenpresent and utilized as an electron acceptor (Section9.1.2). The proportion of the metabolites depends of theconversion of the acetyl-P formed from xylulose-5-P.The acetate kinase directs the flux to the formation ofacetate with production of ATP, the additional energybeing used for growth, whereas the phosphotransacety-lase (PTA) produces acetyl-CoA that is used forbiosynthesis or for reoxydation of NADH with produc-tion of ethanol by alcohol dehydrogenase. The pta genehas been cloned and appears not to be associated withacetate kinase gene, both genes being as a monocistronictranscript (Bourel et al., 2001).
Leuconostoc ferments fructose to fructose-6-P, whichthen enters the pentose-P pathway and is converted tomannitol without the co-formation of sorbitol (vonWeymarn, Hujanen, & Leisola, 2002). The oxidation ofNADH to NAD+ is achieved by reducing fructose to

ARTICLE IN PRESSD. Hemme, C. Foucaud-Scheunemann / International Dairy Journal 14 (2004) 467–494 479
mannitol by an NADH-dependent mannitol dehydro-genase. This affects the cellular redox balance andfavours the production of acetate instead of ethanol sothat an ATP is gained. The enzyme is a tetramer of 4identical sub-unit of about 38 kDa (Aarnikunnas,Ronnholm, & Palva, 2002; Hahn, Kaup, Bringer-Meyer,& Sahm, 2003). The mannitol dehydrogenase gene (mdh)was cloned (Aarnikunnas et al., 2002; Hahn et al., 2003).The sequences of the gene and of the enzyme are distinctfrom those of other bacteria, hybridization occurredonly weakly with some heterofermentative Lactobacillus
strains. The question remained to know if this peculiar-ity is related to the use of fructose as an electronacceptor. The fermentation balance and pathwayregulation have been explored (Grobben et al., 2001).
Galactose and mannose are probably used via theLeloir pathway (Cogan & Jordan, 1994). The galactosegenes are chromosomally encoded and the galK genefrom Ln. lactis has been cloned (Grossiord, Vaughan,Luesink, & de Vos, 1998). Sequence analysis of theDNA flanking galK revealed an open reading frame(ORF) upstream with homology to a a-galactosidasegene and a downstream ORF with homology to galT
from other LAB. The utilization of d-tagatose has beenrecently studied (Bertelsen, Andersen, & Tvede, 2001).
Pentoses are converted to xylulose-5-P that is furthercatabolized in glyceraldehyde-3-P and acetyl-P. Neitherethanol nor acetaldehyde is produced and 1 mole ofATP and acetate are formed from acetyl-P (Cogan &Jordan, 1994).
Lactose is cleaved to galactose and glucose by a b-galencoded by two overlapping genes (lacL and lacM),which are highly homologous (99%) to those of Lb.
casei (Cogan, 1995). The two monosaccharides are thenused via the Leloir and pentose-P pathway, respectively.Most strains grow in the presence of a low lactoseconcentration (20 gL�1), whereas few required a highconcentration, equivalent to that in milk (50 gL�1) ormore. In this case, the induction of the b-gal may limitgrowth (Hemme, unpublished).
Raffinose utilization by Leuconostoc is a variablecharacter and the enzymes involved (a-galactosidase andb-fructosidase) have been studied in term of fermenta-tion balance and induction (Milli"ere, Mathot, Schmitt,& Divi"es, 1989; Pr!evost, Phalip, Huang, & Divi"es, 1993;Server-Busson et al., 1999; Carr et al., 2002).
During sucrose-grown cultures, a large fraction ofsucrose is converted outside the cell into dextran(Section 9) and fructose and does not support growth.The other fraction enters the cell, where it is phos-phorylated by a sucrose phosphorylase and converted toglucose-6-P. The sucrose phosphorylase gene from Ln.
mesenteroides has been cloned (Kitao & Nakano, 1992;Kawasaki, Nakamura, Ohmori, & Sakai, 1996).
Some polysaccharides (e.g. cellulose) are catabolized(Carr et al., 2002) and dextrans are susceptible to
degradation (Eggleston & Legendre, 2003). Althoughthe immunological detection of CcpA-like protein andthe partial characterization of the gene has beenreported in Ln. lactis (Luesink, Kuipers, & de Vos,1998), few are known relative to the excretion ofmetabolic end products and to the regulation of sugarcatabolism.
7.2. Organic acid metabolism
Citrate and malate are the two major organic acidsmetabolized by Leuconostoc. Metabolism of the formeris important for aroma formation and gas production infermented dairy products whereas the latter is unim-portant because milk lacks malate (Cogan & Jordan,1994; McSweeney & Sousa, 2000; Konings, 2002).
Citrate present in the milk (8mmol L�1) and furtherin the cheese curd is not utilized as a sole energy sourcein Leuconostoc but is degraded in the presence of afermentable sugar. As Leuconostoc grows aroundneutral pH, the dianionic form (Hcit2�) of citrate istaken up in symport with a proton. The Arg 425 of thepermease is implicated in the transport process of citrateand S-enantiomers of other substrates with a 2-hydro-xycarboxylate motif (Bandell & Lolkema, 2000; Bourelet al., 2001). Internally, Hcit2� is further degraded inacetate and oxaloacetate, which is decarboxylated topyruvate, thus increasing the pyruvate pool. Theaddition of citrate to glucose both increases the specificgrowth rate and molar growth yield of Ln. mesenter-
oides. Under these conditions, lactate is produced bothfrom citrate and glucose metabolism and exchangebetween citrate and lactate favoured citrate metabolism.Metabolic energy conservation and pH homeostasis bycitrate metabolism in Ln. mesenteroides has beenreviewed by Konings (2002).
Utilization of citrate leads to the formation ofdiacetyl, which is an important component of theflavour of dairy products, and other compounds suchas acetate, acetoin and 2,3-butanediol (McSweeney &Sousa, 2000). The part of the putative reactions involvedin the formation of diacetyl remains unclear. Diacetylcould be produced via the diacetyl synthase althoughthis enzyme has not been clearly detected in all LABtested. Another way to obtain diacetyl is the oxydativedecarboxylation of a-acetolactate formed from pyruvatethat could occur spontaneously or may be protein-mediated. Recently, the acetolactate decarboxylase fromLn. lactis has been purified and sequenced (O’Sullivan,Condon, Cogan, & Sheehan, 2001).
The co-metabolism of citrate plus xylose in Ln.
mesenteroides results in a growth stimulation, anincrease in d-lactate and acetate production andrepression of ethanol formation (Schmitt et al., 1997).This correlates with the formation of acetyl-P frompentose (Section 7).

ARTICLE IN PRESSD. Hemme, C. Foucaud-Scheunemann / International Dairy Journal 14 (2004) 467–494480
Genetic aspects of citrate transport and metabolismhave been progressively studied. The gene encoding thecitrate permease is localized on a 23 kb plasmid in Ln.
mesenteroides, whereas it is chromosomal in Ln. lactis.
Genes involved in the degradation of citrate arechromosomal and arranged in a clyR-mae-citDEF locusin Ln. mesenteroides (Bourel et al., 2001). The citratepermease (citP) genes from Ln. lactis (Vaughan et al.,1995) or Ln. mesenteroides (Bourel et al., 2001), thecitrate utilization gene cluster (cit CDEFG) from Ln.
mesenteroides or (cit MCDEFGRP) from Ln. parame-
senteroides (Martin, Corrales, de Mendoza, Lopez, &Magni, 1999; Bourel et al., 2001) and the gene of aputative regulatory protein, clyR (Bourel et al., 2001)were cloned. The diacetyl reductase has been purifiedand the gene cloned and sequenced (Rattray, Myling-Petersen, Larsen, & Nilsson, 2003).
Most LAB, including Leuconostoc but not Ln. fallax,
carry out malo-lactic fermentation with the formation ofl-lactate and CO2 from malate by malo-lactic enzyme(Cogan & Jordan, 1994; Barrangou et al., 2002;Konings, 2002). Recently, two open reading framesencoding for an NAD-dependent malic enzyme werefound in the upstream region of the citrate utilizationlocus in Ln. mesenteroides (Bourel et al., 2001).
7.3. Nitrogenous compounds metabolism
7.3.1. Amino acid requirements
Leuconostoc strains are fastidious organisms thatrequire amino acids and vitamins in addition to afermentable carbohydrate. Nutritional requirements foramino acids were determined by assessing growth in achemically defined medium lacking one of the compo-nents. Although this varied considerably betweenspecies and strains, the branch-chained amino acidsLeu, Ile and Val, as well as Gln were found to beessential (Cogan & Jordan, 1994; Bellengier et al.,1997b; Foucaud et al., 2001). Ser stimulated the growthof few strains, while Ala was not required by any.
Branched chain amino acid biosynthesis genes sharedhigh similarity with the genes of the BCCA operon ofLactococcus and have been cloned from Ln. mesenter-
oides subsp. cremoris. Additionally, a catabolic a-acetolactate synthase gene is also present but is locatedoutside the putative BCCA operon (Cavin, Dartois,Labarre, & Divi"es, 1999).
Ln. mesenteroides can synthesise Asp from oxaloace-tate via transamination, which can further be convertedto Asn and participates in the biosynthesis of pyrimi-dines and purines (Konings, 2002). The ability ofLeuconostoc to utilize amino acids independently or asa mixture in the presence or not of an added aminoacceptor varied with the strains and the species,suggesting that Leuconostoc offer either a great potentialfor flavour generation for amino acid or would impact
slightly on both physiology and cheese ripening (Crow,Liu, & Holland, 1998; Tavaria, Dahl, Carballo, &Malcata, 2002; Liu, Holland, & Crow, 2003a).
7.3.2. The proteolytic system
Leuconostoc grows poorly in milk since no strainsexceed 5� 108 cfumL�1 (Cogan & Jordan, 1994; De-mirci & Hemme, 1994; Bellengier et al., 1997b). Growthof Leuconostoc can be stimulated up to 109 cfumL�1
when the non-protein nitrogen content (NPN) of milk isartificially increased by the addition of amino acids orpeptides (e.g. amino acid mixture, yeast extract, etc).This indicates that they lack adequate proteolyticactivities that might provide them with assimilableNPN (Vedamuthu, 1994; Bellengier et al., 1997a, b;Server-Busson et al., 1999).
However, preculturing milk with proteolytic strains ofLc. lactis is not recommended for promoting thesubsequent growth of Leuconostoc in cheese manufac-turing because the NPN accumulated in preculturedmilk has been shown to be generally insufficient tostimulate high Leuconostoc growth (Foucaud, Furlan,Bellengier, Juillard, & Richard, 1998).
7.3.2.1. Protein utilization. In contrast with lactococci,little attention has been devoted to the proteolyticsystem of Leuconostoc. Though few strains displayedsome caseinolytic activities, this did not contribute muchto growth (Vafopoulou-Mastrojiannaki & Litopoulou-Tzanetaki, 1996; Bellengier et al., 1997b; Vafopoulou-Mastrojiannaki, 1999; Herreros et al., 2003). Thehydrolysis of caseins may result from the activity ofintracellular housekeeping proteases detected in intra-cellular extracts (Vafopoulou-Mastrojiannaki, 1999) orreleased into the external medium upon cell lysis (Ezzat,El Soda, El Shafei, & Olson, 1993), which probably haverestricted practical importance (if any indeed exist).
7.3.2.2. Peptide utilisation and transport. Ln. mesenter-
oides utilized a large set of di- and tripeptides andoligopeptides up to seven amino acids residues forgrowth (Foucaud et al., 2001), or in non-proliferatingstate (Hemme, Giraudon & Vassal, 1996; Lepeuple &Hemme, unpublished), the ability varying with thestrains. No extracellular breakdown of peptides occursduring peptide transport as peptides are transported inthe peptide form and peptidases are located intracellu-larly (Foucaud et al., 2001).
Separate transport systems were shown to mediate theuptake of amino acids and their dipeptides in Ln.
mesenteroides (Foucaud et al., 2001). Recently, anoligopeptide transport system specific for peptidescontaining at least four amino acid residues wascharacterized in Ln. mesenteroides. It was enhanced inthe presence of Mg2+ or Ca2+ ions, and driven by an

ARTICLE IN PRESSD. Hemme, C. Foucaud-Scheunemann / International Dairy Journal 14 (2004) 467–494 481
energy-rich phosphorylated intermediate (Germain-Alpettaz & Foucaud-Scheunemann, 2002).
Although there were large differences between strains,the overall peptidase profiles of most Leuconostoc
strains are comparable to those of Lc. lactis (Vafopou-lou-Mastrojiannaki & Litopoulou-Tzanetaki, 1996; Ser-ver-Busson et al., 1999; Foucaud et al., 2001; Herreroset al., 2003). However, in cell-free extracts of Leuconos-
toc, carboxypeptidase activity was either significant orlow or absent (Vafopoulou-Mastrojiannaki & Litopou-lou-Tzanetaki, 1996; Macedo, Vieira, Po@as, & Malcata,2000; Herreros et al., 2003), while such activity wasdefinitively not observed using whole cells (Foucaudet al., 2001), suggesting the absence of a carboxypepti-dase or that of appropriate transport system or thespecificity of the existing peptidase. Aminopeptidase anddipeptidylaminopeptidase activities have seldom beencharacterized (Foucaud et al., 2001).
7.3.2.3. Amino acid transport and utilization. The trans-port of branched-chain amino acids in cytoplasmicmembranes of Ln. mesenteroides has been characterizedas a proton-motive-force driven system (Winters, Pool-man, Hemme, & Konings, 1991). Recently, multipleamino acid transport systems were characterized usingwhole cells. Some of these systems were shared byseveral amino acids and their kinetic characteristicscould be an additional tool to estimate the biodiversityof Leuconostoc (Gendrot, Ferchichi, Winters, &Hemme, 2000; Gendrot et al., 2002) although theiractivity does not limit growth.
7.4. Metabolism in the presence of oxygen
In aerobic conditions, oxygen consumption is linkedto an increase of growth yield and specific growth rate(Cogan & Jordan, 1994; Plihon, Taillandier, & Stre-haiano, 1995, 1996). Oxygen is used as an alternativeelectron acceptor and is reduced to hydrogen peroxideand water, and acetate is largely substituted for ethanol(Condon, 1987; Nuraida, Grogolava, Owens, & Camp-bell-Platt, 1992; Cogan & Jordan, 1994). This is due toincreased synthesis of NADH oxidase, which effectivelyregenerates reducing equivalents for continued fermen-tation and allows the acetyl-P to be used for ATP andacetate formation rather than reduction to ethanol as inco-metabolism of sugar and citrate (Section 7). TheNADH oxidase from Ln. mesenteroides was described asa dimer or a tetramer with a subunit molecular mass of53 or 55 kDa, respectively, depending probably of thestrain and FAD as a cofactor (Koike, Kobayashi, Ito,&Saitoh, 1985; Sakamoto, Uchimura, & Komagata,1996). Immobilized Ln. mesenteroides cells possessinghigh NADH oxidase activity have been successfully usedfor the regeneration of the coenzyme (Ergan, Thomas, &Chang, 1984).
The high intracellular levels of Mn2+ (ca. 6–10mmol L�1) might provide Leuconostoc as some otherLAB, with an important defence mechanism againstendogenous O2
� and other reactive oxygen species. Thiscould be related to their original habitat where highMn2+ concentrations are present and explained theabsence of superoxydismutase or manganicatalase(Boyaval, 1989; Horsburgh, Wharton, Karavolos, &Foster, 2002).
7.5. Other metabolic considerations
The esterase activities have been detected withenzymatic test strips (Biom!erieux, France) using b-naphtyl substrates (Milli"ere et al, 1989; Hemme &Bellengier, unpublished). Leuconostoc showed a pro-gressively lower hydrolysis rates as ester chain lengthincreased from C4 to C12 substrates with C14 and C16not being cleaved at all, and C18 being the bestsubstrate.
The hydrolysis of tributyrin, triolein, tributyrin orbutter was not detected on Elliker-rhodamine b agarplates (Meyers, Cuppett, & Hutkins, 1996) whereas thehydrolysis of tributyrin was recently described in Ln.
mesenteroides using naphthyl derivatives and postelec-trophoretic detection (Katz, Medina, Gonzalez, &Oliver, 2002). Leuconostoc lactis synthesized mainlyethyl and butyl esters from tributyrin and ethanolduring incubation in a broth via a transferase reaction(Liu, Holland, & Crow, 2003b). S-methylthioacetate wasthe only thioester formed by Leuconostoc cells incubatedwith methanethiol alone or in conjunction with variousshort-chain fatty acids (Lamberet, Auberger, & Berg"ere,1997).
A d-2-hydroxycaproate dehydrogenase with broadsubstrate specificity, which converts 2-ketocarboxylicacids (including d-lactate i.e. 2-hydroxypropionate) in2-hydroxycarboxylic acids while reducing NAD, hasbeen found in Leuconostoc and in Lactobacillus strains(Hummel, Sch .utte, & Kula, 1985). The question remainsto know if this enzyme differs from the d-lactatedehydrogenase and could be involved in different partsof the metabolism.
8. Genetics of Leuconostoc
The genus Leuconostoc has not been subjected toextensive genetic research yet. Most genetic interestin Leuconostoc has focused on the instability ofphenotypes essential in milk fermentations and onthe transfer of genetic material, especially plasmidDNA, into Leuconostoc strains to improve theirfermentation capacities as reviewed by Dessart andSteenson (1995).

ARTICLE IN PRESSD. Hemme, C. Foucaud-Scheunemann / International Dairy Journal 14 (2004) 467–494482
8.1. Plasmid biology
Leuconostoc harbours one or more native plasmids ofvarious sizes. Lactose utilization, citrate permeaseactivity (Dessart & Steenson, 1995), bacteriocin produc-tion (Fr!emaux et al., 1995), diacetyl reductase (Rattrayet al., 2003) are plasmid-linked but the majority ofplasmids have no known function. The genetic organi-zation and mode of replication of small cryptic plasmidshave been recently described (Coffey, Harrington,Kearney, Daly, & Fitzgerald, 1994; Biet, Cenatiempo,& Fr!emaux, 1999, 2002).
The nucleotide sequence and structural organizationof the plasmid pCI411 (2.9 kb) from Ln. lactis 533(Coffey et al., 1994) and pFR18 (1.8 kb) from Ln.
mesenteroides FR52 (Biet et al., 1999) suggested thattheir replication is done by the rolling circle mechanism.The plasmid pTLX1 (2.7 kb) isolated from Ln. mesen-
teroides Y110 most likely replicates via a theta mechan-ism (Biet et al., 2002). All these plasmids may be usefulfor the construction of food grade vectors.
8.2. Cloning vectors and gene transfer
There are no reports of cloning vectors developedspecifically for Leuconostoc. They either representedexisting antibiotic resistance plasmids or were developedfrom cryptic lactococcal plasmids (Platteeuw, Simons, &de Vos, 1994; Dessart & Steenson, 1995; Kleerebezem,Beerthuyzen, Vaughan, de Vos, & Kuipers, 1997; Bourelet al., 2001). Most of these vectors were efficientlyintroduced in Leuconostoc by conjugation or transfor-mation, thus indicating the feasibility of geneticmanipulations.
The conjugative transfer of the transposon Tn919
from Enterococccus faecalis, nisin production, andlactose fermenting ability were performed and broad-host-range self-transmissible plasmids were used toestablish and develop conjugal transfer protocols inLeuconostoc (Dessart & Steenson, 1995). Plasmid-mediated DNA transformation by electroporation wasoptimized and used in molecular cloning experimentsdespite the frequencies of transfer remained sometimeslow, suggesting the existence of a restriction modifica-tion system (Otts & Day, 1987; Dessart & Steenson,1995, Bourel et al., 2001; Hemme & Vogel, unpub-lished).
Natural (horizontal) gene transfer has been seldomexamined in Leuconostoc. Insertion sequence elementswere identified and analysed in Leuconostoc strains,suggesting the possibility of genetic exchange betweenthese genera (Johansen & Kibenich, 1992; Vaughan &de Vos, 1995; Ward, Brown, & Davey, 1996). Aminoacid sequences of bacteriocins produced by Leuconostoc
are quite similar (e.g. mesentericin Y105 and leucocinA-UAL 187 from Ln. gellidum, Fleury et al., 1996) so
that Leuconostoc strains may constitute a good modelfor studying bacteriocin gene transfer and evolution.The vancomycin resistance genes in Leuconostoc are notthe source of the high-level vancomycin resistance-associated genes in enterococci as determinants differ(Delcour et al., 1999; Patel, 1999). The complete genomesequence of Lb. plantarum WCSF1 revealed that thegene cluster citR-mae-citCDEF is closely related to Ln.
mesenteroides whereas the lactose permease LacS andb-gal are highly related to Ln. lactis (Kleerebezem et al.,2003).
8.3. Gene cloning and expression
The first cloning studies concerned the chromosomalglucose-6-phosphodehydrogenase and phosphoglucoseisomerase genes and a 16-kb restriction fragment of thelactose plasmid pNZ63 from Ln. lactis in E. coli. The Lc.
lactis lacG gene and E. coli lacZ gene were expressed inW. paramesenteroides under the control of Lc. lactis,
demonstrating the feasibility of recombinant DNAtechnology (Dessart & Steenson, 1995). The recA genefrom Ln. mesenteroides has also been cloned (Duwat,Ehrlich, & Gruss, 1992).
Since then, the cloned Leuconostoc genes reported sofar increased and dealt with various aspects oftechnological interest. In the dairy field, this concernsbacteriocin production (Section 5.3.5), peptidoglycanhydrolase (Section 6), carbohydrate utilisation (Section7.1), organic acid metabolism (Section 7.2), andnitrogenous compounds metabolism (Section 7.3.1).
8.4. Genomics
The neotype strain Ln. mesenteroides ATCC 8293 thatwas isolated from fermenting olives and produceddextrans is currently sequenced by the Joint GenomeInstitute/Lactic acid Bacteria Genome Consortium(Walnut Creek, CA, USA; http://www.jgi.doe.gov/JGI microbial/html/index.html). Basic facts and se-quence annotation are in progress (draft sequencecurrently in pre-finishing phase): the genome size is2.0Mb in 116 contigs of 20 reads or greater with anaverage G+C content of 37.4%.
9. Roles of Leuconostoc in technology
Leuconostoc plays important roles in the technologyof dairy products, in particular through the productionof gas and aroma compounds. Despite often beingpresent at high levels in the product with a stablepopulation, all their roles are not clearly defined(Devoyod & Poullain, 1988; Alonso-Calleja et al.,2002) and other functions could be assumed in shelf-life extension through the inhibition of undesirable

ARTICLE IN PRESSD. Hemme, C. Foucaud-Scheunemann / International Dairy Journal 14 (2004) 467–494 483
bacteria (Section 5) and in the new domain of functionalfoods.
9.1. Roles in dairy technology
The role of Ln. mesenteroides subsp. cremoris inaroma production has been well described (Vedamuthu,1994), but these of other Ln. mesenteroides is lessrecognized. A major reason is that for a long time Ln.
mesenteroides subsp. cremoris, known for its aromapotential, was the unique adjunct available. In the sameway, the literature reflects more the use of Ln.
mesenteroides subsp. cremoris than that other speciesor subspecies mostly due to a lack of knowledge. Indeed,strains isolated from raw milk cheeses belong to thesubspecies mesenteroides and dextranicum of Ln. me-
senteroides and to Leuconostoc citreum but not to thesubspecies cremoris of Ln. mesenteroides (Section 4.1;Devoyod & Poullain, 1988; Cogan et al., 1997; -Cibiket al., 2001).
9.1.1. Openness
The demand for opening is the rule for Blue-veinedcheeses and in particular Roquefort in which this allowsthe Penicillium roqueforti colonization. In soft ripenedcheeses in which enough rigid curd cubes avoid collapseand thus permit mechanical opening, Leuconostoc
creates an intracurd opening resulting from CO2
production. In pressed ripened Dutch cheeses such asEdam, Gouda and other brine salted cheese varieties,small and shiny openings are due to CO2 produced byLeuconostoc present in the starter and not to mechanicalopenness related to technological steps, i.e. pressingunder the whey to avoid air entrapment. Strains are hereselected for their moderate production of CO2 to avoidexcessive openness (Vedamuthu, 1994; Martley & Crow,1996).
In Roquefort cheese production, Leuconostoc repre-sents about 5% to 10% of the Lactococcus concentra-tion, i.e. 5� 107 cellsmL�1 of milk, the minimalconcentration being 106 cellsmL�1. Optimal results wereobtained with selected strains of the subspecies mesen-
teroides of Ln. mesenteroides used as concentrated cellssuspensions (Devoyod & Poullain, 1988; Pradel, perso-nal communication). In a few cases, the population ofLeuconostoc originally present in milk and coming fromthe environment creates the desired openness (Reiteret al., 1967; Devoyod & Poullain, 1988; Martley &Crow, 1993, 1996; Hemme, unpublished). In contrast,the addition of Leuconostoc is required with pasteurizedand raw milk of increasing microbial quality, thuscontaining a low bacterial flora.
The maximum concentration of CO2 produced is16mmol kg�1 of cheese, corresponding to the utilizationof the 8mmol L�1 citrate present in milk, but the activityvaries both with the species and the strains (Bellengier,
Foucaud, & Hemme, 1993; Bellengier, Hemme, &Foucaud, 1994). Strains degrading citrate must growduring the early cheese making stages (pressing)although classical conditions (30�C) are unfavourableto Leuconostoc (Section 5.3). They must survive tocontinue this metabolism during ripening in the absenceof sugar. However, some strains, active in low pH curdscould be inactive in washed-curd cheeses having a higherpH (Turner, 1988; Martley & Crow, 1993).
Leuconostoc has been involved in early and lateblowing of some cheeses (Devoyod & Poullain, 1988). InNorwegian cheese of Gouda type, Leuconostoc strainswere present in 7 out of 9 cheeses with atypical eyeformation (Narvhus, Hulbaekdal, & Abrahamsen,1992).
9.1.2. Aroma production
The major compound related to the utilization ofLeuconostoc in the dairy field is diacetyl, acetate andethanol contributing also to aroma formation (Veda-muthu, 1994). The level of diacetyl that is able to givethe desired aroma is low (1.5 to 5 ppm) due to its lowflavour threshold. Sufficient number of cells andpeculiar physico-chemical conditions are required foroptimal citrate utilization and aroma production (Sec-tions 5.3 and 7.2). When allowed by legislation, additionof citrate to the milk (i.e. 0.15% in the USA) or in thewash water (i.e. when lactose removal technology isused, replacing part of the whey with water) permits amore intense aroma production.
Leuconostoc may further transform diacetyl to acetoinand 2,3-butanediol, which do not give aroma. Thisunfavourable transformation could be lowered whenproducts such as fermented milks are cooled after thearoma production. The storage of ripened cheeses isgenerally done at higher temperature (10–13�C), abovethose that are required to stop the process. Theincorporation of oxygen in products such as fermentedmilks favours diacetyl production (Section 7).
Excess acetaldehyde, which may be produced bystarter cultures in butter and fermented milks leads tothe development of a ‘‘green’’ flavour defect (Section 5).Under refrigeration, Leuconostoc is able to reduceacetaldehyde to ethanol, the activity being maximal forLn. mesenteroides subsp. cremoris (Vedamuthu, 1994;Cogan & Jordan, 1994). In cheese, low pH, increasedlevel of salt and low water activity reduce theacetaldehyde metabolism of Ln. mesenteroides subsp.cremoris (Liu et al., 1997).
9.1.3. Deliberate addition of Leuconostoc as NSLAB
Adjuncts of Leuconostoc comparable with those ofother NSLAB (e.g. Lactobacillus) have been tested indifferent cheeses where they are not normally presentsince a real interest exists, in particular for raw milkflora (Crow et al., 2002).

ARTICLE IN PRESSD. Hemme, C. Foucaud-Scheunemann / International Dairy Journal 14 (2004) 467–494484
Ln. mesenteroides subsp. cremoris strains usuallyused for butter, cream or fresh cheeses are not re-covered when used in other cheese and have only aminor effect, if any. Ln. mesenteroides subsp. mesenter-
oides or dextranicum strains are preferred and the cellnumbers in the products need to be at least107 cellsmL�1 of milk. Most strains gave low or nomultiplication in the milk and in the curd, and this is notfavoured by the presence of Lactococcus (Section 5).Milk can also be matured with about 106 cellsmL�1, for15 h at 13�C, to reach this level (Hemme & Boulanger,unpublished).
In Gouda-type cheese manufactured in aseptic in-dustrial conditions, the effect was not clear despite adecrease in citrate content was observed (Martley,personal communication). On a pilot scale, pressed-cheeses containing 108 cells g�1 present a higher pH (5.3vs. 5.1), a higher amino acids content and an acceleratedripening than the control (Hemme et al., 1996; Hemme,Boulanger M .unchner, & Ogier, unpublished). Thishigher pH results from a slight inhibition of l-lactateproduction by Lactococcus and could partly explain themore rapid ripening observed. Leuconostoc could thusbe a good candidate to counteract acidifying starters.These results were confirmed by trials in asepticconditions using the micro-model developed for pressedtype washed-curd cheeses (Hynes, Ogier, & Delacroix-Buchet, 2000). Aroma was modified when cells concen-trations were greater than 5� 106 cellsmL�1 of milk,whereas openness appeared only between 5� 106 and5� 107 cellsmL�1, depending on the strain and relyingpossibly on its autolytic properties (Section 6). Thetexture of the product after two weeks was thenequivalent to that in absence of Leuconostoc after 1month so that the ripening process was accelerated(Hemme, Bienvenu, & Ogier, unpublished).
Leuconostoc strain 5201 was also shown to inhibitNSLAB, which are involved in off-flavour production inCheddar (Martley & Crow, 1993; Section 5). Theinhibition by metabolic end products, combined withthe reduced oxido-reduction potential that exists incheese, has been described to inhibit Lb. plantarum
(Ouvry, Wach!e, Tourdot-Mar!echal, Divi"es, & Cachon,2002).
Ras cheese made in a pilot plant with addition offreeze- or heat shocked cells or cell free extracts ofLeuconostoc strains isolated from the natural floraof French cheese, showed a decreasing bitterness(El Shafei, 1994). Manchego cheeses made frompasteurized milk using defined starters comprising Ln.
mesenteroides obtained higher scores for flavour qualityand intensity and overall impression that cheeses madewith commercial starters (Poveda, Sousa, Cabezas, &Mc Sweeney, 2003).
Very high numbers of cells in cheese trials withdeliberately added cells (108 cellsmL�1 of milk) give an
excess of opening and concomitantly a typical silage off-flavour (Hemme, unpublished).
9.2. Roles in functional foods
A functional food comprises a component that servesto promote health or helps to prevent disease, and ingeneral, the term is used to indicate a food that containssome health-promoting component(s) beyond tradi-tional nutrients. This includes the addition of probioticsand the production of metabolites as well as activitiesthat may contribute to the health benefit of foodmaterials will also be considered hereafter (Hugenholtzet al., 2002; Ouwehand et al., 2003).
9.2.1. Leuconostoc as potential probiotics
As some microorganisms currently proposed to theconsumers, Leuconostoc does not colonize the intestinaltract, and their effect on the host through microbialactions is thus expected to be small except when ingestedat high cell concentrations. Recently, studies of thefeasibility of diarrhoea control in children by feedingfermented milk revealed that Indian Dahi containing108 g�1 Lc. lactis and Ln. mesenteroides reduced themean duration of diarrhoea by 0.3 day (Agarwal &Bhasin, 2002).
9.2.2. Production of polysaccharides
Ln. mesenteroides produces exopolysaccharides(EPS), which are homopolysaccharide consisting ofa-d-glucans such as dextrans mainly composed ofa-1,6-linked residues with variable (strain specific)degrees of branching and alternans composed of a-1,3and a-1,6 linkages. The biosynthesis process is externaland requires sucrose. Specific glycosyltransferase anddextran or levan sucrase enzymes are involved in thebiosynthesis process (de Vuyst & Degeest, 1999;Monchois, Willemot, & Monsan, 1999). Eight glucan-sucrase-encoding genes from Ln. mesenteroides werecloned (Bozonnet et al., 2002). The gene encoding thedextransucrase DsrD can be efficiently expressed andsecreted in a heterologous host (i.e. Lc. lactis MG1363)and is able to drive dextran synthesis (Neubauer,Bauche, & Mollet, 2003). Recently, the gene codingfor an inulosucrase in Ln. citreum was cloned, sequencedand expressed in E. coli. Bacterial inulosucrases,encoding for the production of inulin, have only beenreported in Streptococcus mutans and Lb. reuteri, yet(Olivares-Illana, Lopez-Munguia, & Olvera, 2003).Industrial dextrans are currently used in the manufac-ture of gel filtration products and as blood volumeextenders and blood flow improvers.
In dairy technology, dextrans, as with other EPS, areused as food additives and act as texturizers byincreasing viscosity and as stabilizers through strength-ening the rigidity of the casein network by binding

ARTICLE IN PRESSD. Hemme, C. Foucaud-Scheunemann / International Dairy Journal 14 (2004) 467–494 485
hydration water and interacting with milk constituents.As a consequence, EPS decreases syneresis and im-proves product stability. They play a recognized rolein the manufacturing of fermented milk, culturedcream, milk based dessert and flavoured milk (Pucci& Kunka, 1995; de Vuyst & Degeest, 1999; Duboc &Mollet, 2001; Cooke, Foster, Galema, Ledeboer, &Sanders, 2002).
In addition to technological benefits, certain EPS areclaimed to have beneficial physiological effects on theconsumer. It is assumed that the increased viscosity ofEPS containing foods may increase the residence time ofingested fermented milk in the gastrointestinal tract andtherefore be beneficial to a transient colonization byprobiotic bacteria (German et al., 1999).
a-gluco-oligosaccharides produced by Ln. mesenter-
oides NRRL-B-18242, are highly resistant to attack bydigestive enzymes. They have been proposed to have apotential prebiotic effect in human neonates as theystimulated the growth of beneficial bacteria of theintestinal flora (Djouzi et al., 1995). Such oligosacchar-ides were catabolized by bifidobacteria and lactobacillibut not by Salmonella or E. coli, pointing toward theireffect on intestinal microflora modification (Chung &Day, 2002).
9.2.3. Production of mannitol
Mannitol is a low-calorie sugar that could replacesucrose, lactose, glucose or fructose in food products.It is metabolized independently of insulin and isalso applicable in diabetic food products. Ln. pseudo-
mesenteroides and Ln. mesenteroides are known fortheir ability to produce mannitol in the fermentationof fructose (Section 7). Recently, high yields ofmannitol have been reported, using a membrane cell-recycle bioreactor (von Weymarn, Kiviharju, & Leisola,2002).
9.2.4. Hydrolysis of a-galactosides
a-galactosides such as stachyose and raffinose that arecommonly present in plants (including soya, maize andpulse derived food products) are not metabolized byhumans nor animals owing to the lack of a-galactosidasein intestinal mucosa, hence causing flatulence. Toovercome these drawbacks and to boost the consump-tion of otherwise highly nutritional food products,attempts have been made to eliminate a-galactosidesusing physical methods or a-galactosidase (Hugenholtzet al., 2002). Despite the ability of Leuconostoc toferment a-galactosides (Boquien, Desmazeaud, & Cor-rieu, 1989; Pr!evost et al., 1993; Huang, Pr!evost, &Divi"es, 1994) and the proposal to use a-galactosidasefrom Ln. mesenteroides as a biotechnological approachto remove a-galactosides, attempts did not come out(Huang et al., 1994).
9.2.5. Production of vitamins
Ln. mesenteroides strains producing significantamounts of menaquinones have been characterizedand would be useful as starter cultures for dairy (andother) food fermentations or dietetary supplements toprevent vitamin K deficiency diseases (Morishita,Tamura, Makino, & Kudo, 1999). Recently, theproduction of folate (vitamin B9) has been reported inLn. lactis and Ln. paramesenteroides (Sybesma, Starren-burg, Tijsseling, Hoefnagel & Hugenholtz, 2003).
10. Conclusion and prospects
10.1. Identification and ecology
In the last few decades, new molecular tools, inparticular DNA-based techniques, have contributedwidely to clarify the phylogeny of Leuconostoc and toidentify new species. Knowledge of microbial ecology invarious niches and fermentations might also improvethrough the emergence of molecular identification andtyping methods even though biochemical charactersremain of major interest for technological applications.In addition, the undergoing genomic sequencing andanalysis of LAB, including Leuconostoc will also allow amore comprehensive approach of both the potential andthe relationships of Leuconostoc through genomics(Klaenhammer et al., 2002).
10.2. Health, safety and benefits
Considering the wide distribution of Leuconostoc inthe environment, the large numbers that areingested daily in the vast range of fermented foodssuch as dairy products and the relatively few infectionsthey cause, these bacteria have very little, if any,virulence for healthy human beings. No cases ofinfection have been directly linked to the consumptionof fermented foods or to high occupational exposureto LAB. Even if the potential expansion in the areaof functional foods would have to take safetyaspects into detailed consideration (Feord, 2002), thishistory of human exposure and consumption hasled to the reasonable conclusion that Leuconostoc isgenerally safe. The proper detection and characteriza-tion of clinical isolates of Leuconostoc as well ascareful applications of infection control principles,judicious prophylaxis, and prompt effective therapyshould maximize benefits for patients at risk ofinfection.
Products containing Leuconostoc (as Lactobacillus)may also be a non-neglectible source of Mn2+ as cellscontain millimolar concentrations of Mn2+and variantswith increased concentration could be of potentialinterest (Boyaval, 1989).

ARTICLE IN PRESSD. Hemme, C. Foucaud-Scheunemann / International Dairy Journal 14 (2004) 467–494486
Moreover, three aspects of Leuconostoc metabolismdeserve considerations for the design of functional foodsor the exploitation of metabolic properties in the contextof industrial use. These are the ability of Leuconostoc (i)to hydrolyse a-galactosides such as raffinose fromvegetable products, (ii) to synthesize acarbose analo-gues, which may improve the glycemic profile andinsulin sensitivity in patients with type 2 diabetes (Yoon& Robyt, 2002) or (iii) to produce inulin-like polymers,which may serve as a prebiotic (Olivares-Illana, Wacher-Rodarte, Le Borgne, & Lopez-Munguia, 2002).
10.3. Technological implications
When considering their overall capabilities, Leuco-
nostoc strains could be used to obtain a sufficientproduction of ethanol for ester production (Crow et al.,2002) or acetate for thioester, in cheeses in whichmethane thiol is present (Hemme, 1992), or for theconversion of acetaldehyde to ethanol and acetate (Liuet al., 1997). The fermentation of xylose or otherpentoses will decrease the ethanol content of the productand enhance that of diacetyl, thus answering somedemand for alcohol-free products. This metabolism,which occurs in vegetable fermentation, could also takeplace in mixed dairy-vegetable fermented food. Thesurvival of most Leuconostoc strains in the cheese curd israther high so that they can act as reactors for theproduction and (or) the consumption of metabolitesduring ripening.
When considering their overall potential, Leuconos-
toc, as with other heterofermentative LAB, has a higherpyruvate dissipating potential than homofermentativeLAB so that mutants could be obtained that reroute thepyruvate to the desired pathway. As with Lc. lactis
subsp. lactis biovar diacetylactis, it could be possible tobuild up strains that would not transform the diacetylproduced. The butA mutant reported by Rattray et al.(2003) might be useful in improving the flavour stabilityof certain fermented dairy products, such as culturedbuttermilk, sour cream, and ripened cream butter, wherethe characteristic butter flavour note is due to thepresence of diacetyl. In the same way, if the productionof d-lactate by Leuconostoc appears to limit its use,it could also be possible to obtain variants as withLb. plantarum, that produce a low concentration ofd-lactate, satisfying at least the synthesis of thepeptidoglycan.
Strains belonging to the subspecies mesenteroides ofLn. mesenteroides are more adapted to cheese manu-facturing than Ln. mesenteroides subsp. cremoris strainscommonly found in most commercial preparations.Efforts should be continued for an optimal selectionbased on technological potentials (aptitude to lyse, CO2
production, bacteriocin production, phage sensitivity,compatibilities with other starters, absence of biogenic
amines production, etc). In this context, manufacturersmay increase the development of their proprietarystarter cultures to fulfil the required quality linked totechnological properties (e.g. flavour and texture) and tohygiene, as proposed for dairy fermentations (Povedaet al., 2003) and sauerkraut fermentation (Harris,Fleming, & Klaenhammer, 1992). Additionally, asadjunct cultures, Leuconostoc strains may also limitthe growth and acid production of Lactococcus, thusavoiding negative impacts such as bitterness. They arealso candidates as mixed-starter or pure cultures tomature the milk through the degradation of the NPNfraction and a slight acidification of the medium forsubsequent fermentation and removal of the indigenousNPN fraction that may favour growth of potentialcontaminants. This concept would emerge if the use ofmilk treated by microfiltration expands in dairy plants.
Acknowledgements
The authors acknowledge the contribution of F.Martley (Fonterra Research Centre, Palmerston North,New Zealand), and J. Gall!e (INRA, Jouy en Josas,France).
References
Aarnikunnas, J., Ronnholm, K., & Palva, A. (2002). The mannitol
dehydrogenase gene (mdh) from Leuconostoc mesenteroides is
distinct from other known bacterial mdh genes. Applied Micro-
biology and Biotechnology, 59, 665–671.
Accolas, J.-P., & Auclair, J. E. (1967). Conservation "a l’!etat congel!e de
suspensions de bact!eries lactiques concentr!ees sous faible volume.
I. Bact!eries lactiques m!esophiles. Le Lait, 47, 253–260.
Ackermann, H. W. (2001). Frequency of morphological phage
descriptions in the year 2000. Brief review. Archives of Virology,
146, 843–857.
Agarwal, K. N., & Bhasin, S. K. (2002). Feasibility studies to control
acute diarrhoea in children by feeding fermented milk preparations
Actimel and Dahi. European Journal of Clinical Nutrition, 56,
S56–S59.
Aguirre, M., & Collins, M. D. (1993). Lactic acid bacteria and human
clinical infection. Journal of Applied Bacteriology, 75, 95–107.
Alakomi, H. L., Skytta, E., Saarela, M., Mattila-Sandholm, T., Latva-
Kala, K., & Helander, I. M. (2000). Lactic acid permeabilizes
gram-negative bacteria by disrupting the outer membrane. Applied
and Environmental Microbiology, 66, 2001–2005.
Alonso-Calleja, C., Carballo, J., Capita, R., Bernardo, A., & Garcia-
Lopez, M. L. (2002). Changes in the microflora of Valdeteja raw
goat’s milk cheese throughout manufacturing and ripening.
Lebensmittel-Wissenschaft und- Technologie, 35, 222–232.
Anifantaki, K., Metaxopoulos, J., Kammenou, M., Drosinos, E. H., &
Vlassi, M. (2002). The effect of smoking, packaging and storage
temperature on the bacterial greening of frankfurters caused by
Leuconostoc mesenteroides subsp. mesenteroides. Italian Journal of
Food Science, 14, 135–144.
Antunes, A., Rainey, F. A., Nobre, M. F., Schumann, P., Ferreira, A.
M., Ramos, A., Santos, H., & de Costa, M. S. (2002). Leuconostoc
ficulneum sp. nov., a novel lactic acid bacterium isolated from a ripe

ARTICLE IN PRESSD. Hemme, C. Foucaud-Scheunemann / International Dairy Journal 14 (2004) 467–494 487
fig, and reclassification of Lactobacillus fructosus as Leuconostoc
fructosum comb. nov. International Journal of Systematic and
Evolutionary Microbiology, 52, 647–655.
Arenas, R., Gonzalez, L., Bernardo, A., Fresno, J. M., & Tornadijo,
M. E. (2003). Microbiological and physico-chemical changes in
Genestoso cheese, a Spanish acid curd variety, throughout
ripening. Food Control, Papers in Press, published on line 23
May 2003, doi:10.1016/S095667135(03)0067-7.
Arunga, R. O. (1992). Lactic acid bacteria in coffee and cocoa
fermentation. In B. J. B. Wood (Ed.), The lactic acid bacteria
(pp. 409–430). London & New York: Elsevier Applied Science.
Auge, B., Donnio, P. Y., Le D!eaut, P., & Avril, J. L. (1987). Influence
de la vancomycine intraveineuse sur les flores a!erobies salivaires et
f!ecales. Pathologie Biologie, 35, 673–675.
Avallone, S., Guyot, B., Brillouet, J. M., Olguin, E., & Guiraud, J. P.
(2001). Microbiological and biochemical study of coffee fermenta-
tion. Current Microbiology, 4, 2252–2256.
Bandell, M., & Lolkema, J. S. (2000). Arg-425 of the citrate
transporter CitP is responsible for high affinity binding of
di- and tricarboxylates. Journal of Biological Chemistry, 275,
39130–39136.
Baron, M., Roy, D., & Vuillemard, J. C. (2000). Biochemical
characteristics of fermented milk produced by mixed-cultures of
lactic starters and bifidobacteria. Le Lait, 80, 465–478.
Barrangou, R., Sung-Sik, Y., Breidt Jr., F., Fleming, H. P., &
Klaenhammer, T. R. (2002). Identification and characterization of
Leuconostoc fallax strains isolated from an industrial Sauerkraut
fermentation. Applied and Environmental Microbiology, 68,
2877–2884.
Barreau, C., & Wagener, G. (1990). Characterization of Leuconostoc
lactis strains from human sources. Journal of Clinical Microbiology,
28, 1728–1733.
Barrette, J., Champagne, C. P., Roy, D., & Rodrigue, N. (2000). The
production of mixed cultures containing strains of Lactococcus
lactis, Leuconostoc cremoris and Lactobacillus rhamnosus, on
commercial starter media. Journal of Industrial Microbiology and
Biotechnology, 25, 288–297.
Basak, S., Ramaswamy, H. S., & Piette, J. P. G. (2002). High pressure
destruction kinetics of Leuconostoc mesenteroides and Sacchar-
omyces cerevisiae in single strength and concentrated orange juice.
Innovative Food Science and Emerging Technologies, 3, 223–231.
Bellengier, P., Foucaud, C., & Hemme, D. (1993). Carbon dioxide
production from citrate and glucose in Leuconostoc species
determined by an adapted enzymatic method. Milchwissenschaft,
48, 548–551.
Bellengier, P., Hemme, D., & Foucaud, C. (1994). Citrate metabolism in
sixteen Leuconostoc mesenteroides subsp. mesenteroides and subsp.
dextranicum strains. Journal of Applied Bacteriology, 77, 54–60.
Bellengier, P., Richard, J., & Foucaud, C. (1997a). Associative growth
of Lactococcus lactis and Leuconostoc mesenteroides strains in milk.
Journal of Dairy Science, 80, 1520–1527.
Bellengier, P., Richard, J., & Foucaud, C. (1997b). Nutritional
requirements of Leuconostoc mesenteroides subsp. mesenteroides
and subsp. dextranicum for growth in milk. Journal of Dairy
Research, 64, 95–103.
Benkerroum, N., Misbah, M., Sandine, W. E., & Elaraki, A. T. (1993).
Development and use of a selective medium for isolation of
Leuconostoc spp. from vegetables and dairy products. Applied and
Environmental Microbiology, 59, 607–609.
Bertelsen, H., Andersen, H., & Tvede, M. (2001). Fermentation of
d-tagatose by human intestinal bacteria and lactic acid bacteria.
Microbial Ecology in Health and Disease, 13, 87–95.
Biet, F., Cenatiempo, Y., & Fr!emaux, C. (1999). Characterization of
pFR18, a small cryptic plasmid from Leuconostoc mesenteroides
ssp. mesenteroides FR52, and its use as a foodgrade vector. FEMS
Microbiology Letters, 179, 375–383.
Biet, F., Cenatiempo, Y., & Fr!emaux, C. (2002). Identification of a
replicon from pTXL1, a small cryptic plasmid from Leuconostoc
mesenteroides subsp. mesenteroides Y110, and development of a
food-grade vector. Applied and Environmental Microbiology, 68,
6451–6456.
Bjorkroth, K. J., Geisen, R., Schillinger, U., Weiss, N., de Vos, P.,
Holzapfel, W. H., Korkeala, H. J., & Vandamme, P. (2000).
Characterization of Leuconostoc gasicomitatum sp. nov., associated
with spoiled raw tomato-marinated broiler meat strips packaged
under modified-atmosphere conditions. Applied and Environmental
Microbiology, 66, 3764–3772.
Bjorkroth & Holzapfel, 2003. Genera Leuconostoc, Oenococcus and
Weissella. In: M. Dworkin, et al. (Eds.), The prokaryotes: An
evolving electronic resource for the microbiological community (3rd
ed), release 3.12, March 28, 2003. Berlin: Springer.
Blom, H., Katla, T., Holck, A., Sletten, K., Axelsson, L., & Holo, H.
(1999). Characterization, production, and purification of leucocin
H, a two-peptide bacteriocin from Leuconostoc MF215B. Current
Microbiology, 39, 43–48.
Boizet, B., Mata, M., Mignot, O., Ritzenthaler, P., & Sozzi, T. (1992).
Taxonomic characterization of Leuconostoc mesenteroides and
Leuconostoc oenos bacteriophage. FEMS Microbiology Letters,
90, 211–216.
Boquien, C. Y., Corrieu, G., & Desmazeaud, M. J. (1988). Effect
of fermentation conditions on growth of Streptococcus cremoris
AM2 and Leuconostoc lactis CNRZ 1091 in pure and
mixed culture. Applied and Environmental Microbiology, 54,
2527–2531.
Boquien, C. Y., Desmazeaud, M. J., & Corrieu, G. (1989). Caract-
!erisation d’une aminopeptidase chez Streptococcus cremoris AM2
et d’une a-galactosidase chez Leuconostoc lactis CNRZ 1091. Le
Lait, 69, 71–81.
Bourel, G., Henini, S., Krantar, K., Oraby, M., Divi"es, C., & Garmyn,
D. (2001). M!etabolisme sucre-citrate chez Leuconostoc mesenter-
oides. Le Lait, 81, 75–82.
Bover-Cid, S., & Holzapfel, W. H. (1999). Improved screening
procedure for biogenic amine production by lactic acid bacteria.
International Journal of Food Microbiology, 53, 33–41.
Boyaval, P. (1989). Lactic acid bacteria and metal ions. Le Lait, 69,
87–113.
Bozonnet, S., Dols-Laffargue, M., Fabre, E., Pizzut, S., Remaud-
Simeon, M., Monsan, P., & Willemot, R. M. (2002). Molecular
characterization of DSR-E, an alpha-1,2 linkage-synthesizing
dextransucrase with two catalytic domains. Journal of Bacteriology,
184, 5753–5761.
Brashears, M. M., Jaroni, D., & Trimble, J. (2003). Isolation, selection,
and characterization of lactic acid bacteria for a competitive
exclusion product to reduce shedding of E. coli O157:H7 in cattle.
Journal of Food Protection, 66, 355–363.
Buckenh .uskes, H. J. (1993). Selection criteria for lactic acid bacteria to
be used as starter cultures for various food commodities. FEMS
Microbiology Reviews, 12, 253–272.
Cabo, M. L., Braber, A. F., & Koenraad, P. M. (2002). Apparent
antifungal activity of several lactic acid bacteria against Penicillium
discolor is due to acetic acid in the medium. Journal of Food
Protection, 65, 1309–1316.
Caplice, E., & Fitzgerald, G. F. (1999). Food fermentations: Role of
microorganisms in food production and preservation. International
Journal of Food Microbiology, 50, 131–149.
Cappelli, E. A., Barros, R. R., Camello, T. C., Teixeira, L. M., &
Merquior, V. L. (1999). Leuconostoc pseudomesenteroides as a
cause of nosocomial urinary tract infections. Journal of Clinical
Microbiology, 37, 4124–4126.
Carr, F. J., Chill, D., & Maida, N. (2002). The lactic acid bacteria:
A literature survey. Critical Reviews in Microbiology, 28,
281–370.

ARTICLE IN PRESSD. Hemme, C. Foucaud-Scheunemann / International Dairy Journal 14 (2004) 467–494488
Casalta, E., No.el, Y., Le Bars, D., Carr!e, C., Achilleos, C., &
Maroselli, M. X. (2001). Characterization of Bastelicaccia cheese.
Le Lait, 81, 529–546.
Casanova-Roman, M., Rios, J., Sanchez-Porto, A., Gomar, J. L., &
Casanova-Bellido, M. (2003). Leuconostoc bacteremia in a healthy
infant. Minerva Pediatrica, 55, 83–86.
Cavin, J. F., Dartois, V., Labarre, C., & Divi"es, C. (1999). Cloning of
branched chain amino acid biosynthesis genes and assays of a-acetolactate synthase activities in Leuconostoc mesenteroides subsp.
cremoris. Research in Microbiology, 150, 189–198.
Champagne, C. P., & Gardner, N. J. (2002). Effect of process
parameters on the production of Leuconostoc mesenteroides
cultures. Journal of Industrial Microbiology and Biotechnology, 28,
291–296.
Chung, C. H., & Day, D. F. (2002). Glucooligosaccharides from
Leuconostoc mesenteroides B-742 (ATCC 13146): A potential
prebiotic. Journal of Industrial and Microbiological Biotechnology,
29, 196–199.
-Cibik, R., & Chapot-Chartier, M.-P. (2000). Autolysis of dairy
leuconostocs and detection of peptidoglycan hydrolases by
renaturing SDS-PAGE. Journal of Applied Microbiology, 89,
862–869.
-Cibik, R., & Hemme, D. (1997). Variabilit!e du caract"ere autolytique
chez le genre Leuconostoc. 8th club des bact!eries lactiques, Dijon,
France, May 28–30, 1997.
-Cibik, R., Lepage, E., & Tailliez, P. (2000). Molecular diversity of
Leuconostoc mesenteroides and Leuconostoc citreum isolated from
traditional French cheeses as revealed by RAPD fingerprinting,
16S rDNA sequencing and 16S rDNA fragment amplification.
Systematic and Applied Microbiology, 23, 267–278.
-Cibik, R., Tailliez, P., Langella, P., & Chapot-Chartier, M. P. (2001).
Identification of Mur, an atypical peptidoglycan hydrolase derived
from Leuconostoc citreum. Applied and Environmental Microbiol-
ogy, 67, 858–864.
Cleveland, J., Montville, T. J., Nes, I. F., & Chikindas, M. L. (2001).
Bacteriocins: Safe, natural antimicrobials for food preservation.
International Journal of Food Microbiology, 71, 1–20.
Coffey, A., Harrington, A., Kearney, K., Daly, C., & Fitzgerald, G.
(1994). Nucleotide sequence and structural organization of the
small, broad-host-range plasmid pCI411 from Leuconostoc lactis
533. Microbiology, 140, 2263–2269.
Cogan, T. M. (1995). Flavour production by dairy starter cultures.
Journal of Applied Bacteriology Symposium Supplement, 79,
49S–64S.
Cogan, T. M. (2002). Improving Cheddar cheese flavour by micro-
biological way. 26th International Dairy Congress, Paris, France,
September 24–27, 2002.
Cogan, T. M., Barbosa, M., Beuvier, E., Bianchi-Salvadori, B.,
Cocconcelli, P. S., Fernandes, I., Gomez, J., Gomez, R.,
Kalantzopoulos, G., Ledda, A., Medina, M., Rea, M. C., &
Rodriguez, E. (1997). Characterization of the lactic acid bacteria in
artisanal dairy products. Journal of Dairy Research, 64, 409–421.
Cogan, T. M., & Jordan, K. N. (1994). Metabolism of Leuconostoc
bacteria. Journal of Dairy Science, 77, 2704–2717.
Collins, M. D., Samelis, J., Metaxopoulos, J., & Wallbanks, S. (1993).
Taxonomic studies on some leuconostoc-like organisms from
fermented sausages: Description of a new genus Weissella for the
Leuconostoc paramesenteroides group of species. Journal of Applied
Bacteriology, 75, 595–603.
Condon, S. (1987). Responses of lactic acid bacteria to oxygen. FEMS
Microbiology Reviews, 46, 269–280.
Cooke, D., Foster, T. J., Galema, S. A., Ledeboer, A. M., & Sanders,
J. W. (2002). Method of preparing a fermented milk-based food
product. Unilever, European Patent No. 1 180 329.
Corbier, C., Krier, F., Mulliert, G., Vitoux, B., & Revol-Junelles, A.
M. (2001). Biological activities and structural properties of the
atypical bacteriocins mesenterocin 52B and leucocin B-TA33a.
Applied and Environmental Microbiology, 67, 1418–1422.
Crow, V. L., Curry, B., Christison, M., Hellier, K., Holland, R., & Liu,
S. Q. (2002). Raw milk flora and NSLAB as adjuncts. Australian
Journal of Dairy Technology, 57, 99–105.
Crow, V. L., Liu, S. Q., & Holland, R. (1998). The diversity of amino
acid utilisation and ethyl butyrate formation by dairy bacteria.
Australian Journal of Dairy Technology, 53, 118.
dal Bello, F., Walter, J., Hammes, W. P., & Hertel, C. (2003).
Increased complexity of the species composition of lactic acid
bacteria in human feces revealed by alternative incubation
condition. Microbial Ecology, 45, 455–463.
Davey, G. P., Ward, L. J. H., & Brown, J. C. S. (1995).
Characterisation of four bacteriophages isolated from dairy
fermentations. FEMS Microbiology Letters, 128, 21–26.
Davidson, P. M. (1997). Chemical preservatives and natural anti-
microbial compounds. In M. P. Doyle, R. P. Beuchat, & T. J.
Montville (Eds.), Food Microbiology: Fundamentals and frontiers
(pp. 520–526). Washington: ASM Press.
Day, D. F. (1992). Spoilage in the sugar industry. In B. J. B. Wood
(Ed.), The lactic acid bacteria (pp. 343–362). London & New York:
Elsevier Applied Science.
Dayem, M. A., Fleury, Y., Devilliers, G., Chaboisseau, E., Girard, R.,
Nicolas, P., & Delfour, A. (1996). The putative immunity protein
of the Gram-positive bacteria Leuconostoc mesenteroides is
preferentially located in the cytoplasm compartment. FEMS
Microbiology Letters, 138, 251–259.
Delcour, J., Ferain, T., Deghorain, M., Palumbo, E., & Hols, P.
(1999). The biosynthesis and functionality of the cell-wall of lactic
acid bacteria. Antonie van Leeuwenhoek, 76, 159–184.
del Nozal-Nalda, M., Jimenez-Mejias, M. E., Suarez, A. I., & Viciana,
P. (1997). Sepsis caused by Leuconostoc cremoris, neutropenia and
human immunodeficiency virus infection. Medicina Clinica (Bar-
celona), 108, 799.
Demirci, Y., & Hemme, D. (1994). Growth of Leuconostoc mesenter-
oides strains isolated from French raw milk cheeses in a reference
milk. Milchwissenschaft, 43, 483–485.
Demirci, Y., & Hemme, D. (1995). Acidification test based on the use
of biomass for screening of Leuconostoc. Application to Ln.
mesenteroides strains isolated from French raw milk cheeses.
Journal of Dairy Research, 62, 521–527.
Derre, I., Rapoport, G., & Msadek, T. (1999). CtsR, a novel regulator
of stress and heat shock response, controls clp and molecular
chaperone gene expression in Gram-positive bacteria. Molecular
Microbiology, 31, 117–131.
Dessart, S. R., & Steenson, L. R. (1995). Biotechnology of dairy
Leuconostoc. In Y. H. Hui, & G. G. Khatchatourians (Eds.), Food
Biotechnology: Microorganisms (pp. 665–702). New York: VCH
Publishers.
Devoyod, J.-J., & Poullain, F. (1988). Les leuconostocs. Propri!et!es:
Leur r #ole en technologie laiti"ere. Le Lait, 68, 249–280.
de Vuyst, L., & Degeest, B. (1999). Heteropolysaccharides from lactic
acid bacteria. FEMS Microbiology Reviews, 23, 153–177.
Deye, G., Lewis, J., Patterson, J., & Jorgensen, J. (2003). A case of
Leuconostoc ventriculitis with resistance to carbapenem antibiotics.
Clinical Infectious Diseases, 37, 869–870.
Dhodapkar, K. M., & Henry, N. K. (1996). Leuconostoc bacteremia in
an infant with short-gut syndrome: Case report and literature
review. Mayo Clinical Proceedings, 71, 1171–1174.
Dicks, L. M., Dellaglio, F., & Collins, M. D. (1995). Proposal to
reclassify Leuconostoc oenos as Oenococcus oeni [corrig.] gen. nov.,
comb. nov. International Journal of Systematic Bacteriology, 45,
395–397.
Dicks, L. M. T., Fantuzzi, L., Gonzalez, F. C., Du Toit, M., &
Dellaglio, F. (1993). Leuconostoc argentinum sp. nov. isolated from

ARTICLE IN PRESSD. Hemme, C. Foucaud-Scheunemann / International Dairy Journal 14 (2004) 467–494 489
Argentine raw milk. International Journal of Systematic Bacteriol-
ogy, 43, 347–351.
Djouzi, Z., Andrieux, C., Pelenec, V., Somarriba, S., Popot, F., Paul,
F., Monsan, P., & Szylit, O. (1995). Degradation and fermentation
of a-gluco-oligosaccharides by bacterial strains from human colon:
In vitro and in vivo studies in gnotobiotic rats. Journal of Applied
Bacteriology, 79, 117–127.
Duboc, P., & Mollet, B. (2001). Applications of exopolysaccharides in
the dairy industry. International Dairy Journal, 11, 759–768.
Duthoit, F., Godon, J. J., & Montel, M. C. (2003). Bacterial
community dynamics during production of registered designation
of origin Salers cheese as evaluated by 16S rRNA gene single-
strand conformation polymorphism analysis. Applied and Environ-
mental Microbiology, 69, 3840–3848.
Duwat, P., Ehrlich, S. D., & Gruss, A. (1992). A general method for
cloning recA genes of gram-positive bacteria by polymerase chain
reaction. Journal of Bacteriology, 174, 5171–5175.
Eggleston, G., & Legendre, B. (2003). Mannitol and oligosaccharides
as new criteria for determining cold tolerance in sugarcane
varieties. Food Chemistry, 80, 451–461.
El Shafei, H. (1994). Accelerated ripening of Ras cheese using cell free
extract, freeze- and heat-shocked Leuconostoc spp. cultures. Die
Nahrung, 6, 599–605.
Ennahar, S., Cai, Y., & Fujita, Y. (2003). Phylogenetic diversity of
lactic acid bacteria associated with paddy rice silage as determined
by 16S ribosomal DNA analysis. Applied and Environmental
Microbiology, 69, 444–451.
Ennahar, S., Sashihara, T., Sonomoto, K., & Ishizaki, A. (2000). Class
IIa bacteriocins: Biosynthesis, structure, and activity. FEMS
Microbiology Reviews, 24, 85–106.
Ercolini, D., Hill, P. J., & Dodd, C. E. R. (2003). Bacterial community
structure and location in Stilton cheese. Applied and Environmental
Microbiology, 69, 3540–3548.
Ergan, F., Thomas, D., & Chang, T. M. (1984). Selection and
microencapsulation of an ‘‘NADH-oxidizing’’ bacterium and its
use for NAD regeneration. Applied Biochemistry and Biotechnol-
ogy, 10, 61–71.
Estepar, J., del Mar Sanchez, M., Alonso, L., & Mayo, B. H.
(1999). Biochemical and microbiological characterization of
artisanal ‘‘Penamellera’’ cheese. International Dairy Journal, 9,
737–746.
Ezzat, N., El Soda, M., El Shafei, H., & Olson, N. F. (1993). Cell-wall
associated peptide hydrolase and esterase activities in several
cheese-related bacteria. Food Chemistry, 48, 19–23.
Facklam, R. R., & Elliot, J. A. (1995). Identification, classification,
and clinical relevance of catalase-negative, Gram-positive cocci,
excluding the streptococci and enterococci. Clinical Microbiology
Reviews, 8, 479–495.
Facklam, R. R., Pigott, N., Franklin, R., & Elliott, J. (1995).
Evaluation of three disk tests for identification of enterococci,
leuconostocs, and pediococci. Journal of Clinical Microbiology, 33,
885–887.
Fantuzzi, L., Raffellini, S., & Gonzalez, F. C. (1999). Inhibition of
psychotrophic bacteria growth in raw milk stored at low
temperature by Leuconostoc mesenteroides subsp. dextranicum
LL152. Microbiologie Aliments Nutrition, 17, 101–106.
FAO. (1966). Toxicological evaluation of some antimicrobials,
antioxydants, emulsifiers, stabilizers, flour-treatment agents, acids
and bases. FAO Nutrition Meetings Report Series, 40, 339.
Farrow, J. A. E., Facklam, R. R., & Collins, M. D. (1989). Nucleic
acid homologies of some vancomycin-resistant leuconostocs and
description of Leuconostoc citreum sp. nov. and Leuconostoc
pseudomesenteroides sp. nov. International Journal of Systematic
Bacteriology, 39, 279–283.
Fauchais, A. L., Roques, S., Druesne, L., Verdonck, A., Chassagne,
P., Bercoff, E., Doucet, J., & Fr!ebourg, N. (2003). Rave
opportunistic infection due to Leuconostoc species. La Revue de
M!edecine Interne, 24, 268–275.
Feord, J. (2002). Lactic acid bacteria in a changing legislative
environment. Antonie van Leeuwenhoek, 82, 353–360.
Fitzsimons, N. A., Cogan, T. M., Condon, S., & Beresford, T. (1999).
Phenotypic and genotypic characterization of non-starter lactic
acid bacteria in mature Cheddar cheese. Applied and Environmental
Microbiology, 65, 3418–3426.
Fleury, Y., Dayem, M. A., Montagne, J. J., Chaboisseau, E.,
Le Caer, J. P., Nicolas, P., & Delfour, A. (1996). Covalent
structure, synthesis, and structure-function studies of mesen-
tericin Y 105, a defensive peptide from Gram-positive bacteria
Leuconostoc mesenteroides. Journal of Biological Chemistry, 271,
14421–14429.
Foucaud, C., Fran@ois, A., & Richard, J. (1997). Development of a
chemically defined medium for the growth of Leuconostoc
mesenteroides. Applied and Environmental Microbiology, 63,
301–304.
Foucaud, C., Furlan, S., Bellengier, P., Juillard, V., & Richard, J.
(1998). Nutritional value of the non-protein N that accumulates
during growth of Lactococcus lactis in milk for dairy lactococcal
and leuconostoc isolates. Journal of Dairy Research, 65, 491–501.
Foucaud, C., Hemme, D., & Desmazeaud, M. (2001). Peptide
utilization by Lactococcus lactis and Leuconostoc mesenteroides.
Letters in Applied Microbiology, 31, 20–25.
Fr!emaux, C., H!echard, Y., & Cenatiempo, Y. (1995). Mesentericin
Y105 gene clusters in Leuconostoc mesenteroides Y105. Microbiol-
ogy, 141, 1637–1645.
Garcia Fontan, M. C., Franco, I., Prieto, B., Tornadijo, M. E., &
Carballo, J. (2001). Microbiological changes in San Simon cheese
throughout ripening and its relationship with physico-chemical
parameters. Food Microbiology, 18, 25–33.
Garvie, E. I. (1986). Genus Leuconostoc van Tieghem 1878. In P. H. A.
Sneath, N. S. Mair, M. E. Sharpe, & J. G. Holt (Eds.), Bergey’s
manual of systematic bacteriology (pp. 1071–1075). Baltimore:
Williams and Wilkins.
Gendrot, F., Ferchichi, M., Winters, D., & Hemme, D. (2000). Valine
transport and biodiversity of Leuconostoc wild strains from French
raw milk cheeses. Systematic and Applied Microbiology, 23,
461–468.
Gendrot, F., Foucaud-Scheunemann, C., Ferchichi, M., & Hemme, D.
(2002). Characterization of amino acid transport in the dairy strain
Leuconostoc mesenteroides subsp. mesenteroides CNRZ 1273.
Letters in Applied Microbiology, 35, 291–295.
Germain-Alpettaz, V., & Foucaud-Scheunemann, C. (2002). Identifi-
cation and characterization of an oligopeptide transport system in
Leuconostoc mesenteroides subsp. mesenteroides CNRZ 1463.
Letters in Applied Microbiology, 35, 68–73.
German, B., Schiffrin, E. J., Renerio, R., Mollet, B., Pfeifer, A., &
Neese, J. R. (1999). The development of functional foods: Lessons
from the gut. Trends in Biotechnology, 17, 492–499.
Giacometti, A., Ranaldi, R., Siquini, F. M., & Scalise, G. (1993).
Leuconostoc citreum isolated from lung in AIDS patient. Lancet,
342, 622.
Gillespie, R. S., Symons, J. M., & Mc Donald, R. A. (2002). Peritonitis
due to Leuconostoc species in a child receiving peritoneal dialysis.
Pediatric Nephrology, 17, 966–968.
Golan, Y., Poutsiaka, G. G., Tozzi, S., Hadley, S., & Snydman, D. R.
(2001). Daptomycin for line-related Leuconostoc bacteraemia.
Journal of Antimicrobial Chemotherapy, 47, 357–368.
Gonzalez de Llano, D., Cuesta, P., & Rodriguez, A. (1998). Biogenic
amine production by wild lactococcal and leuconostoc strains.
Letters in Applied Microbiology, 26, 270–274.
Gopal, P. K., Davey, G. P., Crow, V. L., Pillidge, C. J., Coolbear, T.,
& Holland, R. (1996). Extracellular material from Leuconostoc
mesenteroides subsp. dextranicum mediates the aggregation of

ARTICLE IN PRESSD. Hemme, C. Foucaud-Scheunemann / International Dairy Journal 14 (2004) 467–494490
lactococcal cells: Implications for the isolation of single strains.
Journal of Applied Bacteriology, 81, 48–56.
Green, M., Wadowsky, R. M., & Barbadora, K. (1990). Recovery of
vancomycin-resistant Gram-positive cocci from children. Journal of
Clinical Microbiology, 28, 484–488.
Grobben, G. J., Peters, S. W., Wisselink, H. W., Weusthuis, R. A.,
Hoefnagel, M. H., Hugenholtz, J., & Eggink, G. (2001).
Spontaneous formation of a mannitol-producing variant of
Leuconostoc pseudomesenteroides grown in the presence of fructose.
Applied and Environmental Microbiology, 67, 2867–2870.
Grossiord, B., Vaughan, E. E., Luesink, E., & de Vos, W. M. (1998).
Genetics of galactose utilisation via the Leloir pathway in lactic
acid bacteria. Le Lait, 78, 77–84.
Gueguen, Y., Chemardin, P., Labrot, P., Arnaud, A., & Galzy, P.
(1997). Purification and characterization of an intracellular b-glucosidase from a new strain of Leuconostoc mesenteroides isolated
from cassava. Journal of Applied Microbiology, 82, 469–476.
G .urtler, V., & Mayall, B. C. (2001). Genomic approaches to typing
taxonomy and evolution of bacterial isolates. International Journal
of Systematic and Evolutionary Microbiology, 51, 3–16.
Hahn, G., Kaup, B., Bringer-Meyer, S., & Sahm, H. (2003). A zinc-
containing mannitol-2-dehydrogenase from Leuconostoc pseudo-
mesenteroides ATCC 12291: Purification of the enzyme and cloning
of the gene. Archives of Microbiology, 179, 101–107.
Hamasaki, Y., Ayaki, M., Fuchu, H., Sugiyama, M., & Morita, H.
(2003). Behavior of psychrotrophic lactic acid bacteria isolated
from spoiling cooked meat products. Applied and Environmental
Microbiology, 69, 3668–3671.
Hamdy, M. K., & Williams, F. S. (2001). Bacterial amelioration of
bauxite residue water of industrial alumina plants. Journal of
Industrial Microbiology and Biotechnology, 27, 228–233.
Hansen, E. B. (2002). Commercial bacterial starter cultures for
fermented foods of the future. International Journal of Food
Microbiology, 78, 119–131.
Harris, L. J., Fleming, H. P., & Klaenhammer, T. R. (1992). Novel
paired starter culture system for sauerkraut, consisting of a nisin-
resistant Leuconostoc mesenteroides strain and a nisin-producing
Lactococcus lactis strain. Applied and Environmental Microbiology,
58, 1484–1489.
H!echard, Y., Berjeaud, J. M., & Cenatiempo, Y. (1999). Characteriza-
tion of the mesB gene and expression of bacteriocins by
Leuconostoc mesenteroides Y105. Current Microbiology, 39,
265–269.
Heikkil.a, M. P., & Saris, P. E. J. (2003). Inhibition of Staphylococcus
aureus by the commensal bacteria of human milk. Journal of
Applied Microbiology, 95, 471–478.
Hemme, D. (1992). Pathways to the formation of flavour volatiles in
cheese. Symposium FIL/IDF on Cheese ripening, April 7–9, 1992.
Lund, Sweden.
Hemme, D., Giraudon, C., & Vassal, L. (1996). Addition of
Leuconostoc to the milk in cheesemaking. 5th symposium on lactic
acid bacteria, September, 8–12, 1996. Veldhoven, The Netherlands.
Herrero, M., Mayo, B., Gonzales, B., & Suarez, J. E. (1996).
Evaluation of technologically important traits in lactic acid
bacteria isolated from spontaneous fermentations. Journal of
Applied Bacteriology, 81, 565–570.
Herreros, M. A., Fresno, J. M., Gonzalez Prieto, M. E., & Tornadijo,
M. E. (2003). Technological characterization of lactic acid bacteria
isolated from Armada cheese (a Spanish goats’milk cheese).
International Dairy Journal, 13, 469–479.
Holt, S. M., Al-Sheikh, H., & Shin, K. J. (2001). Characterization of
dextran-producing Leuconostoc strains. Letters in Applied Micro-
biology, 32, 185–189.
Horsburgh, M. J., Wharton, S. J., Karavolos, M., & Foster, S. J.
(2002). Manganese: Elemental defence for a life with oxygen.
Trends in Microbiology, 10, 496–501.
Huang, D. Q., Pr!evost, H., & Divi"es, C. (1994). Principal character-
istics of a-galactosidase from Leuconostoc mesenteroides subsp.
mesenteroides. Journal of Basic Microbiology, 34, 87–95.
Hugenholtz, J., Sybesma, W., Groot, M. N., Wisselink, W., Ladero,
V., Burgess, K., van Sinderen, D., Piard, J. C., Eggink, G., Smid, E.
J., Savoy, G., Sesma, F., Jansen, T., Hols, P., & Kleerebezem, M.
(2002). Metabolic engineering of lactic acid bacteria for the
production of neutraceuticals. Antonie van Leeuwenhoek, 82,
217–235.
Hummel, W., Sch .utte, H., & Kula, M. R. (1985). d-2-hydroxycaproate
dehydrogenase from Lactobacillus casei. Applied Microbiology and
Biotechnology, 21, 7–15.
Hynes, E., Ogier, J.-C., & Delacroix-Buchet, A. (2000). Protocol for
the manufacture of miniature washed-curd cheeses under con-
trolled microbiological conditions. International Dairy Journal, 10,
733–737.
Jang, J., Kim, B., Lee, J., & Han, H. (2003). A rapid method for
identification of typical Leuconostoc species by 16S rDNA PRC-
RFLP analysis. Journal of Microbiological Methods, 55, 295–302.
Jimenez-Mejias, M. E., Becerril, B., Gomez-Cia, T., del Nozal, M., &
Palomino-Nicas, J. (1997). Bacteremia caused by Leuconostoc
cremoris in a patient with severe burn injuries. European Journal of
Clinical and Microbiological Infectious Diseases, 16, 533–535.
Johansen, E., & Kibenich, A. (1992). Isolation and characterization of
IS1165, an insertion sequence of Leuconostoc mesenteroides subsp.
cremoris and other lactic acid bacteria. Plasmid, 27, 200–206.
Jordan, K. N., & Cogan, T. M. (1995). Growth and metabolite
production by mixed-strain starter cultures. Irish Journal of
Agricultural and Food Research, 34, 39–47.
Juillard, V., Foucaud, C., Flambard, B., Furlan, S., Bellengier, P., &
Richard, J. (1998). Interactions entre bact!eries lactiques m!esophiles
dans le lait: R #ole des facteurs nutritionnels. Le Lait, 78, 91–97.
Kalchayanand, N., Frethem, C., Dunne, P., Sikes, A., & Ray, B.
(2002). Hydrostatic pressure and bacteriocin-triggered cell wall
lysis of Leuconostoc mesenteroides. Innovative Food Science &
Emerging Technologies, 3, 33–40.
Katla, A. K., Kruse, H., Johnsen, G., & Herikstad, H. (2001).
Antimicrobial susceptibility of starter culture bacteria used in
Norwegian dairy products. International Journal of Food Micro-
biology, 67, 147–152.
Katz, M., Medina, R., Gonzalez, S., & Oliver, G. (2002). Esterolytic
and lipolytic activities of lactic acid bacteria isolated from ewe’s
milk and cheese. Journal of Food Protection, 65, 1997–2001.
Kawasaki, H., Nakamura, N., Ohmori, M., & Sakai, T. (1996).
Cloning and expression in E. coli of sucrose phosphorylase gene
from Leuconostoc mesenteroides No. 165. Bioscience, Biotechnology
and Biochemistry, 60, 322–324.
Kikuchi, K., Totsuka, K., Shimizu, K., Yoshida, K., Kobayashi, M.,
Tomonaga, O., Oomori, Y., & Okada, S. (1994). (Microbiological
and clinical studies of vancomycin resistant Leuconostoc spp. and
Pediococcus spp. isolated from septicemia patients). Kansenshogaku
Zasshi, 68, 1084–1092 (in Japanese).
Kim, B., Lee, J., Jang, J., Kim, J., & Han, H. (2003). Leuconostoc inhae
sp. nov, a lactic acid bacterium isolated from kimchi. International
Journal of Systematic and Evolutionary Microbiology, 53,
1123–1126.
Kim, D. S., Thomas, S., & Scott-Fogler, H. (2000). Effect of pH and
trace minerals on log-term starvation of Leuconostoc mesenteroides.
Applied and Environmental Microbiology, 66, 976–981.
Kim, J., Chun, J., & Han, H. U. (2000). Leuconostoc kimchii sp. nov., a
new species from kimchi. International Journal of Systematic and
Evolutionary Microbiology, 50, 1915–1919.
Kitao, S., & Nakano, E. (1992). Cloning of the sucrose phosphorylase
gene from Leuconostoc mesenteroides and its overexpression using a
‘‘sleeper’’ bacteriophage vector. Journal of Fermentation and
Bioengineering, 73, 179–184.

ARTICLE IN PRESSD. Hemme, C. Foucaud-Scheunemann / International Dairy Journal 14 (2004) 467–494 491
Klaenhammer, T., Altermann, E., Arigoni, F., Bolotin, A., Breidt, F.,
Broadbent, J., Cano, R., Chaillou, S., Deutscher, J., Gasson, M.,
van de Guchte, M., Guzzo, J., Hartke, A., Hawkins, T., Hols, P.,
Hutkins, R., Kleerebezem, M., Kok, J., Kuipers, O., Lubbers, M.,
Maguin, E., McKay, L., Mills, D., Nauta, A., Overbeek, R., Pel,
H., Pridmore, D., Saier, M., van Sinderen, D., Sorokin, A., Steele,
J., O’Sullivan, D., de Vos, W. M., Weimer, B., Zagorec, M., &
Siezen, R. (2002). Discovering lactic acid bacteria by genomics.
Antonie Van Leeuwenhoek, 82, 29–58.
Kleerebezem, M., Beerthuyzen, M. M., Vaughan, E. E., de Vos, W.
M., & Kuipers, O. P. (1997). Controlled gene expression systems
for lactic acid bacteria: Transferable nisin-inducible expression
cassettes for Lactococcus, Leuconostoc, and Lactobacillus spp.
Applied and Environmental Microbiology, 63, 4581–4584.
Kleerebezem, M., Boekhorst, J., van Kranenburg, R., Molenaar, D.,
Kuipers, O. P., Leer, R., Tarchini, R., Peters, S. A., Sandbrink, H.
M., Fiers, M. W., Stiekema, W., Lankhorst, R. M., Bron, P. A.,
Hoffer, S. M., Groot, M. N., Kerkhoven, R., de Vries, M., Ursing,
B., de Vos, W. M., & Siezen, R. J. (2003). Complete genome
sequence of Lactobacillus plantarum WCFS1. Proceedings of the
National Academy of Sciences USA, 100, 1990–1995.
Koike, K., Kobayashi, T., Ito, S., & Saitoh, M. (1985). Purification
and characterization of NADH oxidase from a strain of
Leuconostoc mesenteroides. Journal of Biochemistry, 97, 1279–1288.
Konings, W. N. (2002). The cell membrane and the struggle for life of
lactic acid bacteria. Antonie van Leeuwenhoek, 82, 3–27.
Krier, F., Revol-Junelles, A. M., & Germain, P. (1998). Influence
of temperature and pH on production of two bacteriocins
by Leuconostoc mesenteroides subsp. mesenteroides FR52 during
batch fermentation. Applied Microbiology Biotechnology, 50,
359–363.
Lamberet, G., Auberger, B., & Berg"ere, J. L. (1997). Aptitude of
cheese bacteria for volatile S-methyl thioester synthesis. II.
Comparison of coryneform bacteria, Micrococcaceae and some
lactic acid bacteria starters. Applied Microbiology and Biotechnol-
ogy, 48, 393–397.
Lange, H., Plihon, F., Taillandier, P., & Riba, J. P. (1995). A new
method for the determination of Leuconostoc mesenteroides cell
number. Journal of Microbiological Methods, 24, 93–97.
Lee, H. J., Park, S. Y., & Kim, J. (2000). Multiplex PCR-based
detection and identification of Leuconostoc species. FEMS Micro-
biology Letters, 15, 243–247.
Limonet, M., Revol-Junelles, A. M., & Milli"ere, J. B. (2002a).
Variations in the membrane fatty acid composition of resistant or
susceptible Leuconostoc or Weissella strains in the presence or
absence of mesenterocin 52A and mesenterocin 52B produced by
Leuconostoc mesenteroides subsp. mesenteroides FR52. Applied and
Environmental Microbiology, 68, 2910–2916.
Limonet, M., Revol-Junelles, A. M., & Milli"ere, J. B. (2002b). Les
ph!enom"enes de r!esistance induite aux bact!eriocines de bact!eries
lactiques. Science des Aliments, 22, 123–132.
Limsowtin, G. K. Y., Powell, I. B., & Parente, E. (1996). Type of
starters. In T. M. Cogan, & J. P. Accolas (Eds.), Dairy starter
cultures (pp. 101–130). London: VCH Publishers Inc.
Liu, S. Q., Asmundson, R. V., Holland, R., & Crow, V. L. (1997).
Acetaldehyde metabolism by Leuconostoc mesenteroides subsp.
cremoris under stress conditions. International Dairy Journal, 7,
175–183.
Liu, S. Q., Holland, R., & Crow, V. L. (2003a). The potential of dairy
lactic acid bacteria to metabolise amino acids via non-transaminat-
ing reactions and endogenous transamination. International Jour-
nal of Food Microbioliogy, 86, 257–269.
Liu, S. Q., Holland, R., & Crow, V. L. (2003b). Ester synthesis in an
aqueous environment by Streptococcus thermophilus and other
dairy lactic acid bacteria. Applied Microbiology and Biotechnology,
63, 81–88.
Lopez-Diaz, T. M., Alonso, C., Roman, C., Garcia Lopez, M. L., &
Moreno, B. (2000). Lactic acid bacteria isolated from a hand-made
Blue cheese. Food Microbiology, 17, 23–32.
Luesink, E. J., Kuipers, O. P., & de Vos, W. M. (1998). Regulation of
the carbohydrate metabolism in Lactococcus lactis and other lactic
acid bacteria. Le Lait, 78, 69–76.
Luh, K. T., Hsueh, P. R., Teng, L. J., Paan, H. J., Chen, Y. C., Lu, J.
J., Wu, J. J., & Ho, S. W. (2000). Quinupristin-dalfopristin
resistance among Gram-positive bacteria in Taiwan. Antimicrobial
Agents and Chemotherapy, 44, 3374–3380.
Lyhs, U. (2002). Lactic acid bacteria associated with the spoilage of
fish products. Ph.D. Thesis, University of Helsinki, Finland.
Macedo, A. C., Vieira, M., Po@as, R., & Malcata, F. X. (2000). Peptide
hydrolase system of lactic acid bacteria isolated from Serra da
Estrela cheese. International Dairy Journal, 10, 769–774.
Malik, R. K., Naveen, K., & Mathur, D. K. (1995). Bacteriocinogenic
activity of Leuconostoc spp. against food spoilage and pathogenic
bacteria. Cultured Dairy Products Journal, 30, 10–12.
Manolopoulou, E., Sarantinopoulos, P., Zoidou, E., Aktypis, A.,
Moschopoulou, E., Kandarakis, I. G., & Anifantakis, E. M.
(2003). Evolution of microbial populations during traditional Feta
cheese manufacture and ripening. International Journal of Food
Microbiology, 82, 153–161.
Mans de Marion, M. C., Raffellini, S. M., Fantuzzi, L., &
Gonzalez, F. C. (1995). Inhibitory effect of Leuconostoc mesenter-
oides subsp. dextranicum on Pseudomonas fluorescens in milk
stored at low temperature. Australian Journal of Dairy Technology,
50, 1–5.
Martin, M., Corrales, M. A., de Mendoza, D., Lopez, P., & Magni, C.
(1999). Cloning and molecular characterization of the citrate
utilization citMCDEFGRP cluster of Leuconostoc paramesenter-
oides. FEMS Microbiology Letters, 174, 231–238.
Martinez-Murcia, A. J., & Collins, M. D. (1991). A phylogenetic
analysis of an atypical leuconostoc: Description of Leuconostoc
fallax sp. nov. FEMS Microbiology Letters, 82, 55–60.
Martley, F. G., & Crow, V. L. (1993). Interactions between non-starter
microorganisms during cheese manufacture and ripening. Interna-
tional Dairy Journal, 3, 461–483.
Martley, F. G., & Crow, V. L. (1996). Open texture in cheese: The
contributions of gas production by microorganisms and cheese
manufacturing practices. Journal of Dairy Research, 63, 489–507.
Mas, M., Tabla, R., Moriche, J., Roa, I., Gonzalez, J., Rebollo, J. E.,
& Caceres, P. (2002). Ibores goat’s milk cheese: Microbiological
and physiochemical changes throughout ripening. Le Lait, 82,
579–587.
Mathieu, F., Sudirman, I., Rekhif, N., Milli"ere, J. B., & Lef"ebvre, G.
(1993). Mesenterocin 52, a bacteriocin produced by Leuconostoc
mesenteroides subsp. mesenteroides FR 52. Journal of Applied
Bacteriology, 74, 372–379.
Mathot, A. G., Kihal, M., Pr!evost, H., & Divi"es, C. (1994). Selective
enumeration of Leuconostoc on vancomycin agar medium. Inter-
national Dairy Journal, 4, 459–469.
Matte-Tailliez, O., Qu!en!ee, P., -Cibik, R., van Opstal, J., Dessevre, F.,
Firmesse, O., & Tailliez, P. (2001). Detection and identification of
lactic acid bacteria in milk and industrial starter culture with
fluorescently labeled rRNA-targeted peptide nucleic acid probes.
Le Lait, 81, 237–248.
Mauriello, G., Moio, L., Moschetti, G., Piombino, P., Addeo, F., &
Coppola, S. (2001). Characterization of lactic acid bacteria strains
on the basis of neutral volatile compounds produced in whey.
Journal of Applied Microbiology, 90, 928–942.
McSweeney, P. L. H., & Sousa, M. J. (2000). Biochemical pathways
for the production of flavour compounds in cheeses during
ripening. A review. Le Lait, 80, 293–324.
Menendez, S., Godinez, R., Centeno, J. A., & Rodriguez-Otero, J. L.
(2001). Identification and characterisation of lactic acid bacteria

ARTICLE IN PRESSD. Hemme, C. Foucaud-Scheunemann / International Dairy Journal 14 (2004) 467–494492
isolated from Tetilla cheese made from crude milk. Alimentaria, 38,
77–83.
Meyers, S. A., Cuppett, S. L., & Hutkins, R. W. (1996). Lipase
production by lactic acid bacteria and activity on butter oil. Food
Microbiology, 13, 383–389.
Milli"ere, J. B., Mathot, A. G., Schmitt, P., & Divi"es, C. (1989).
Phenotypic characterization of Leuconostoc species. Journal of
Bacteriology, 67, 529–542.
Mogensen, G., Salminen, S., O’Brien, J., Ouwehand, A., Holzapfel,
W., Shortt, C., Fonden, R., Miller, G. D., Donohue, D., Playne,
M., Crittenden, R., Salvadori, B. B., & Zink, R. (2002). Inventory
of microorganisms with a documented history of use in food. FIL/
IDF Bulletin, 377, 10–19.
Monchois, V., Willemot, R. M., & Monsan, P. (1999). Glucansucrases:
Mechanism of action and structure-function relationships. FEMS
Microbiology Reviews, 23, 131–151.
Monnet, C., Schmitt, P., & Divi"es, C. (1994). Method for assaying
volatile compounds by headspace gas chromatography and
application to growing starter cultures. Journal of Dairy Science,
77, 1809–1815.
Morea, M., Baruzzi, F., & Cocconcelli, P. S. (1999). Molecular and
physiological characterization of dominant populations in tradi-
tional Mozzarella cheese processing. Journal of Applied Microbiol-
ogy, 87, 574–582.
Morishita, T., Tamura, N., Makino, T., & Kudo, S. (1999).
Production of menaquinones by lactic acid bacteria. Journal of
Dairy Science, 82, 1897–1903.
Morisset, D., & Fr"ere, J. (2002). Heterologous expression
of bacteriocins using the mesentericin Y105 dedicated
transport system by Leuconostoc mesenteroides. Biochimie, 84,
569–576.
Mor-Mur, M., Carretero, C., Pla, R., & Guamis, B. (1994). Microbial
changes during ripening of Cendrat del Montsec, a goat’s milk
cheese. Food Microbiology, 11, 177–185.
Morris, C. E., Bardin, M., Berge, O., Frey-Klett, P., Fromin, N.,
Girardin, H., Guinebreti"ere, M. H., Lebaron, P., Thi!ery, J. M., &
Troussellier, M. (2002). Microbial diversity: Approaches to
experimental design and hypothesis testing in primary literature
from 1975 to 1999. Microbiology and Molecular Biology Reviews,
66, 592–616.
Mundt, J. O. (1970). Lactic acid bacteria associated with raw
plant food material. Journal of Milk and Food Technology, 33,
550–553.
Narvhus, J. A., Hulbaekdal, A., & Abrahamsen, R. K. (1992).
Occurrence of lactobacilli and leuconostoc in Norwegian cheese of
Gouda type and their possible influence on cheese ripening and
quality. Symposium FIL/IDF on cheese ripening, April 7–9, 1992.
Lund, Sweden.
Nelson, R. R. S. (1998). Intrinsically vancomycin-resistant Gram-
positive organisms: Clinical relevance and implications for infec-
tion control. Journal of Hospital Infection, 42, 275–282.
Neubauer, H., Bauche, A., & Mollet, B. (2003). Molecular character-
ization and expression analysis of the dextransucrase DsrD of
Leuconostoc mesenteroides Lcc4 in homologous and heterologous
Lactococcus lactis cultures. Microbiology, 149, 973–982.
Nuraida, L., Grigolava, I., Owens, J. D., & Campbell-Platt, G. (1992).
Oxygen and pyruvate as external electron acceptors for Leuconos-
toc sp. Journal of Applied Bacteriology, 72, 517–522.
O’Donnell, A. G., Embley, T. M., & Goodfellow, M. (1993). Future of
bacterial systematics. In M. Goodfellow, & A. G. O’Donnell
(Eds.), Handbook of new bacterial systematic (pp. 513–524).
London: Academic Press.
Ogier, J.-C., Son, O., Gruss, A., Tailliez, P., & Delacroix-Buchet, A.
(2002). Identification of the bacterial microflora in dairy products
by temporal temperature gradient gel electrophoresis. Applied and
Environmental Microbiology, 68, 3691–3701.
Ohkuma, M., & Kudo, T. (1998). Phylogenetic analysis of the
symbiotic intestinal microflora of the termite Cryptotermes
domesticus. FEMS Microbiology Letters, 164, 389–395.
Olivares-Illana, V., Lopez-Munguia, A., & Olvera, C. (2003).
Molecular characterization of inullosucrase from Leuconostoc
citreum: A fructosyltransferase within a glucosyltransferase.
Journal of Bacteriology, 185, 3606–3612.
Olivares-Illana, V., Wacher-Rodarte, C., Le Borgne, S., &
L "opez-Munguia, A. (2002). Characterization of a cell-associated
inulosucrase from a novel source: A Leuconostoc citreum
strain isolated from Pozol, a fermented corn beverage of Mayan
origin. Journal of Industrial Microbiology and Biotechnology, 28,
112–117.
Ostlie, H. M., Eliassen, L., & Skeie, S. (2002). Characterization of
NSLAB in Scandinavien cheese during ripening. 7th symposium on
lactic acid bacteria, September 1–5, 2002. Egmond aan Zee, The
Netherlands.
O’Sullivan, S. M., Condon, S., Cogan, T. M., & Sheehan, D. (2001).
Purification and characterisatioon of acetolactate decarboxylase
from Leuconostoc lactis NCW1. FEMS Microbiology Letters, 194,
245–249.
Otts, D. R., & Day, D. F. (1987). Optimization of protoplast
formation and regeneration in Leuconostoc mesenteroides. Applied
and Environmental Microbiology, 53, 1694–1695.
Ouvry, A., Wach!e, Y., Tourdot-Mar!echal, R., Divi"es, C., & Cachon,
R. (2002). Effects of oxydoreduction potential combined with
acetic acid, NaCl and temperature on the growth, acidification, and
membrane properties of Lactobacillus plantarum. FEMS Micro-
biology Letters, 214, 257–261.
Ouwehand, A. C., Bianchi Salvadori, B., Fonden, R., Mogensen, G.,
Salminen, S., & Sellars, R. (2003). Health effects of probiotics and
culture-containing dairy products in humans. FIL/IDF Bulletin,
380, 4–19.
Park, I. S., & Walsh, C. T. (1997). D-Alanyl-d-lactate and d-alanyl-d-
alanine synthesis by d-alanyl-d-alanine ligase from vancomycin-
resistant Leuconostoc mesenteroides. Effects of a phenylalanine 261
to tyrosine mutation. Journal of Biological Chemistry, 272,
9210–9214.
Patel, R. (1999). Enterococcal-type glycopeptide resistance genes in
non-enterococcal organisms. FEMS Microbiology Letters, 185,
1–7.
Pepe, O., Blaiotta, G., Moschetti, G., Greco, T., & Villani, F. (2003).
Rope-producing strains of Bacillus spp. from wheat bread and
strategy for their control by lactic acid bacteria. Applied and
Environmental Microbiology, 69, 2321–2329.
P!erez, G., Cardell, E., & Zarate, V. (2002). Random amplified
polymorphic DNA analysis for differentiation of Leuconostoc
mesenteroides subspecies isolated from Tenerife cheese. Letters in
Applied Microbiology, 34, 82–85.
Peterson, S. D., & Marshall, R. T. (1990). Nonstarter lactobacilli in
Cheddar cheese: A review. Journal of Dairy Science, 73, 1395–1410.
Phalip, V., Dartois, V., Schmitt, P., & Divi"es, C. (1994). Cloning of the
d-lactate dehydrogenase gene from Leuconostoc mesenteroides
subsp. cremoris. Biotechnology Letters, 16, 221–226.
Platteeuw, C., Simons, G., & de Vos, W. M. (1994). Use of the E. coli
beta-glucuronidase (gusA) gene as a reporter gene for analyzing
promoters in lactic acid bacteria. Applied and Environmental
Microbiology, 60, 587–593.
Plihon, F., Taillandier, F., & Strehaiano, P. (1995). Oxygen effect on
batch cultures of Leuconostoc mesenteroides: Relationship between
oxygen uptake, growth and end-products. Applied Microbiology
and Biotechnology, 43, 117–122.
Plihon, F., Taillandier, F., & Strehaiano, P. (1996). Oxygen effect on
lactose catabolism by a Leuconostoc mesenteroides strain: Model-
ling of general O2-dependant stoichiometry. Biotechnology and
Bioengineering, 49, 63–69.

ARTICLE IN PRESSD. Hemme, C. Foucaud-Scheunemann / International Dairy Journal 14 (2004) 467–494 493
Poveda, J. M., Sousa, M. J., Cabezas, L., & McSweeney, P. L. H.
(2003). Preliminary observations on proteolysis in Manchego
cheese made with a defined-strain starter culture and adjunct
starter (Lactobacillus plantarum) or a commercial starter. Interna-
tional Dairy Journal, 13, 169–178.
Pr!evost, H., Phalip, V., Huang, D. Q., & Divi"es, C. (1993). Metabolism
and physiological behaviour of Leuconostoc dextranicum subsp.
mesenteroides growing on raffinose. Journal of Basic Microbiology,
33, 433–441.
Pucci, M. J., & Kunka, B. S. (1995). Novel dextran produced by
Leuconostoc dextranicum. Microlife Technics Incorporation, Uni-
ted States Patent No. 4 933 191.
Randazzo, C. L., Torriani, S., Akkermans, A. D. L., de Vos, W. M., &
Vaughan, E. E. (2002). Diversity, dynamics, and activity of
bacterial communities during production of an artisanal Sicilian
cheese as evaluated by 16S rRNA analysis. Applied and Environ-
mental Microbiology, 68, 1882–1892.
Rattray, F. P., Myling-Petersen, D., Larsen, D., & Nilsson, D. (2003).
Plasmid-encoded diacetyl (acetoin) reductase in Leuconostoc
pseudomesenteroides. Applied and Environmental Microbiology, 69,
304–311.
Reeson, A. F., Jankovic, T., Kasper, M. L., Rogers, S., & Austin, A.
D. (2003). Application of 16S rDNA-DGGE to examine the
microbial ecology associated with a social wasp Vespula germanica.
Insect Molecular Biology, 12, 85–91.
Reiter, B., Fryer, T. F., Pickering, A., Chapman, H. R., Lawrence, R.
C., & Sharpe, M. E. (1967). The effect of microbial flora on the
flavour and free fatty acid composition of Cheddar cheese. Journal
of Dairy Research, 34, 257–272.
Revol-Junelles, A. M., & Lef"ebvre, G. (1996). Purification and N-
terminal amino acid sequence of dextranicin 24, a bacteriocin of
Leuconostoc sp. Current Microbiology, 33, 136–137.
Ringo, E., & Gatesoupe, F. J. (1998). Lactic acid bacteria in fish: A
review. Aquaculture, 160, 177–203.
Robinson, R. K., Tamime, A. Y., & Wszolek, M. (2002). Microbiology
of fermented milks. In R. K. Robinson (Ed.), Dairy microbiology
handbook: the microbiology of milk and milk products (pp. 367–430).
New York: Wiley.
Rodriguez, E., Gonzalez, B., Gaya, P., Nunez, M., & Medina, M.
(2000). Diversity of bacteriocins produced by lactic acid bacteria
isolated from raw milk. International Dairy Journal, 10, 7–15.
Rodriguez, J., Saavedra, J., Fernandez-Jurado, A., & Prados, D.
(1999). Leuconostoc pseudomesenteroides bacteremia. Sangre (Bar-
celona), 44, 82–83.
Sakamoto, M., Uchimura, T., & Komagata, K. (1996). Comparison of
H2O-forming NADH oxidase from Leuconostoc mesenteroides
subsp. mesenteroides NRIC 1541T and H2O2-forming NADH
oxidase from Sporolactobacillus inulinus NRIC 1133T. Journal of
Biofermentation and Bioengineering, 82, 531–537.
Salotra, P., Singh, D. K., Seal, K. P., Krishna, N., Jaffe, H., &
Bhatnagar, R. (1995). Expression of DnaK and GroEL homologs
in Leuconostoc mesenteroides in response to heat shock, cold shock
or chemical stress. FEMS Microbiology Letters, 131, 57–62.
Sarkar, P. K., & Sharmistha, B. (1996). Antibacterial activity of lactic
acid bacterial isolates obtained from natural habitats. Journal of
Food Science and Technology-Mysore, 33, 231–233.
Schabereiter-Gurtner, C., Maca, S., Rolleke, S., Nigl, K., Lukas, J.,
Hirschl, A., Lubitz, W., & Barisani-Asenbauer, T. (2001). 16S
rDNA-based identification of bacteria from conjunctival swabs by
PCR and DGGE fingerprints. Investigative Ophthalmology and
Visual Science, 42, 1164–1171.
Schmitt, P., Vasseur, C., Phalip, V., Huang, D. Q., Divi"es, C., &
Pr!evost, H. (1997). Diacetyl and acetoin production from the co-
metabolism of citrate and xylose by Leuconostoc mesenteroides
subsp. mesenteroides. Applied Microbiology and Biotechnology, 47,
715–718.
Sch .onhuber, W., Le Bourhis, G., Tremblay, J., Amann, R., &
Kulakauskas, S. (2001). Utilization of tmRNA sequences for
bacterial identification. BMC Microbiology, 1, 20.
Serot, T., Dousset, X., Zucca, J., & Torcatis, N. (1990). Isolation and
partial purification of antibacterial substances produced by
Leuconostoc mesenteroides and Lactobacillus plantarum isolated
from kefir. Microbiologie Aliments Nutrition, 8, 71–76.
Server-Busson, C., Foucaud, C., & Leveau, J. Y. (1999). Selection of
dairy Leuconostoc isolates for important technological properties.
Journal of Dairy Research, 66, 245–256.
Shaw, B. G., & Harding, C. D. (1989). Leuconostoc gelidum sp. nov.
and Leuconostoc carnosum sp. nov. from chill-stored meats.
International Journal of Systematic Bacteriology, 39, 217–223.
Stackebrandt, E., Frederiksen, W., Garrity, G. M., Grimont, P. A. D.,
K.ampfer, P., Maiden, M. C. J., Nesme, X., Rossello-Mora,
R., Swings, J., Tr .uper, H. G., Vauterin, L., Ward, A. C., &
Whitman, W. B. (2002). Report of the ad hoc committee for the
re-evaluation of the species definition in bacteriology. International
Journal of Systematic and Evolutionary Microbiology, 52,
1043–1047.
Steinkraus, K. H. (2002). Fermentations in world food processing.
Comprehensive Reviews in Food Science and Food Safety, 1, 23–32.
Stiles, M. E. (1994). Bacteriocins produced by Leuconostoc species.
Journal of Dairy Science, 77, 2718–2724.
Stiles, M. E., & Holzapfel, W. H. (1997). Lactic acid bacteria of foods
and their current taxonomy. International Journal of Food
Microbiology, 36, 1–29.
Sudirman, I., Mathieu, F., Benoit, V., & Lef"ebvre, G. (1994).
Properties of two bacteriocins synthesized by Leuconostoc strains.
Current Microbiology, 28, 155–159.
Susiluoto, T., Korkeala, H., & Bjorkroth, K. J. (2003). Leuconostoc
gasicomitatum is the dominating lactic acid bacterium in
retail modified-atmosphere-packaged marinated broiler meat
strips on sell-by-day. International Journal of Food Microbiology,
80, 89–97.
Swearingen, P. A., O’Sullivan, D. J., & Warthesen, J. J. (2001).
Isolation, characterization, and influence of native, non starter
lactic acid bacteria on Cheddar cheese quality. Journal of Dairy
Science, 84, 50–59.
Swenson, J. M., Facklam, R. R., & Thornsberry, C. (1990).
Antimicrobial susceptibility of vancomycin-resistant Leuconostoc,
Pediococcus, and Lactobacillus species. Antimicrobial Agents and
Chemotherapy, 34, 543–549.
Sybesma, W., Starrenburg, M., Tijsseling, L., Hoefnagel, M. H. N., &
Hugenholtz, J. (2003). Effects of cultivation conditions on folate
production by lactic acid bacteria. Applied and Environmental
Microbiology, 69, 452–458.
Takahashi, M., Okada, S., Uchimura, T., & Kozaki, M. (1992).
Leuconostoc amelibiosum Schillinger, Holzapfel, and Kandler 1989
is a later subjective synonym of Leuconostoc citreum Farrow,
Facklam, and Collins 1989. International Journal of Systematic
Bacteriology, 42, 649–651.
Tamang, J. P., Dewan, S., Thapa, S., Olasupo, N. A., Schillinger, U.,
Wijaya, A., & Holzapfel, W. H. (2000). Identification and
enzymatic profiles of the predominant lactic acid bacteria isolated
from soft-variety chhurpi, a traditional cheese typical of the Sikkim
Himalayas. Food Biotechnology, 14, 99–112.
Tanaka, H., Doesburg, K., Iwasaki, T., & Mierau, I. (1999). Screening
of lactic acid bacteria for bile salt hydrolase activity. Journal of
Dairy Science, 82, 2530–2535.
Tavaria, F. K., Dahl, S., Carballo, F. J., & Malcata, F. X. (2002).
Amino acid catabolism and generation of volatiles by lactic acid
bacteria. Journal of Dairy Science, 85, 2462–2470.
Tavaria, F. K., & Malcata, F. X. (2003). Enzymatic activities of non-
starter lactic acid bacteria isolated from a traditional Portuguese
cheese. Enzyme and Microbial Technology, 33, 236–243.

ARTICLE IN PRESSD. Hemme, C. Foucaud-Scheunemann / International Dairy Journal 14 (2004) 467–494494
Teale, C. J. (2002). Antimicrobial resistance and the food chain.
Symposium Series of the Society of Applied Microbiology, 31,
85S–89S.
Thunell, R. K. (1995). Taxonomy of the leuconostocs. Journal of Dairy
Science, 78, 2514–2522.
Turner, K. W. (1988). Some aspects of the microbiology of cheese
ripening investigated using aseptic manufacturing techniques.
Ph.D. Thesis, Massey University, Palmerston North, New Zealand.
Vafopoulou-Mastrojiannaki, A. (1999). Influence of pH and NaCl on
proteolytic and esterolytic activity of intracellular extract of
Leuconostoc mesenteroides subsp. mesenteroides K1G8 strain.
Milchwissenschaft, 54, 314–316.
Vafopoulou-Mastrojiannaki, A., & Litopoulou-Tzanetaki, E. (1996).
Protease and peptidase activity from whole cells and crude cell-free
extracts of Leuconostoc mesenteroides and Leuconostoc paramesen-
teroides isolates from cheese. Microbiologie Aliments Nutrition, 14,
167–174.
Vagiakou-Voudris, E., Mylona-Petropoulou, D., Kalogeropoulou, E.,
Chantzis, A., Chini, S., Tsiodra, P., & Malamou-Lada, E. (2002).
Multiple liver abscesses associated with bacteremia due to
Leuconostoc lactis. Scandinavian Journal of Infection Disease, 34,
766–767.
Vaughan, E. E., David, S., & de Vos, W. M. (1996). The lactose
transporter in Leuconostoc lactis is a new member of the LacS
subfamily of galactoside-pentose-hexuronide translocators. Applied
and Environmental Microbiology, 62, 1574–1582.
Vaughan, E. E., David, S., Harrington, A., Daly, C., Fitzgerald, G. F.,
& de Vos, W. M. (1995). Characterization of plasmid-encoded
citrate permease (citP) genes from Leuconostoc species reveals high
sequence conservation with the Lactococcus lactis citP gene.
Applied and Environmental Microbiology, 61, 3172–3176.
Vaughan, E. E., & de Vos, W. M. (1995). Identification and
characterization of the insertion element IS1070 from Leuconostoc
lactis NZ6009. Gene, 155, 95–100.
Vedamuthu, E. R. (1994). The dairy Leuconostoc: Use in dairy
products. Journal of Dairy Science, 77, 2725–2737.
Villani, F., Moschetti, G., Blaiotta, G., & Coppola, S. (1997).
Characterization of strains of Leuconostoc mesenteroides by
analysis of soluble whole-cell protein pattern, DNA fingerprinting
and restriction of ribosomal DNA. Journal of Applied Microbiol-
ogy, 82, 578–588.
von Weymarn, N., Hujanen, M., & Leisola, M. (2002). Production of
d-mannitol by heterofermentative lactic acid bacteria. Process
Biochemistry, 37, 1207–1213.
von Weymarn, N., Kiviharju, K., & Leisola, M. (2002). High-level
production of d-mannitol with membrane cell-recycle bioreactor.
Journal of Industrial Microbiology and Biotechnology, 29, 44–49.
Walsh, B., & Cogan, T. M. (1973). Diacetyl, acetoin, and acetaldehyde
production by mixed-species lactic starter cultures. Applied
Microbiology, 26, 820–825.
Ward, L. J. H., Brown, J. C. S., & Davey, G. P. (1996). Identification
and sequence analysis of IS1297, an ISS1-like insertion sequence in
a Leuconostoc strain. Gene, 174, 259–263.
Williams, A. G., Choi, A. G., & Banks, J. M. (2002). Variability of
the species phenotype composition of the non-starter lactic
acid bacterial population of cheddar cheese manufactured
in a commercial creamery. Food Research International, 35,
483–493.
Winters, D. A., Poolman, B., Hemme, D., & Konings, W. N. (1991).
Branched-chain amino acid transport in cytoplasmic membranes of
Leuconostoc mesenteroides subsp. dextranicum CNRZ 1273.
Applied and Environmental Microbiology, 57, 3350–3354.
Wuytack, E. Y., Diels, A. M. J., & Michiels, C. W. (2002). Bacterial
inactivation by high-pressure homogenisation and high hydrostatic
pressure. International Journal of Food Microbiology, 77, 205–212.
Yoon, S. H., & Robyt, J. F. (2002). Synthesis of acarbose analogues by
transglycosylation reactions of Leuconostoc mesenteroides B-
512FMC and B-742CB dextransucrases. Carbohydrate Research,
337, 2427–2435.
Zarazaga, M., Saenz, Y., Portillo, A., Tenorio, C., Ruiz-Larrea, F.,
Del Campo, R., Baquero, F., & Torres, C. (1999). In vitro activities
of ketolide HMR3647, macrolides, and other antibiotics against
Lactobacillus, Leuconostoc, and Pediococcus isolates. Antimicrobial
Agents and Chemotherapy, 43, 3039–3041.
Zinner, S. H. (1999). Changing epidemiology of infections in patients
with neutropenia and cancer: Emphasis on gram-positive and
resistant bacteria. Clinical Infectious Diseases, 29, 490–494.
Further reading
Collins, Y. F., McSweeney, P. L. H., & Wilkinson, M. G. (2003).
Lipolysis and free fatty acid catabolism in cheese: A review of
current knowledge. International Dairy Journal, 13, 841–866.
Kaup, B., Bringer-Meyer, S., & Sahm, H. (2003). Metabolic
engineering of Escherichia coli: Construction of an efficient
biocatalyst for d-mannitol formation in a whole-cell biotransfor-
mation. Applied Microbiology and Biotechnology, Papers in Press,
published on line 28 October 2003, doi:10.1007/S00253-003-1470-9.
Menendez, S., Godinez, R., Hermida, M., Centeno, J. A., &
Rodriguez-Otero, J. L. (2004). Characteristics of ‘‘Tetilla’’ pasteur-
ized milk cheese manufactured with the addition of autochthonous
cultures. Food Microbiology, 21, 97–104.
Seifu, E., Buys, E. M., & Donkin, E. F. (2003). Effect of the
lactoperoxidase system on the activity of mesophilic cheese starter
cultures in goat milk. International Diary Journal, 13, 953–959.
Sievers, M., Uermosi, C., Fehlmann, M., & Kriger, S. (2003). Cloning,
sequence analysis and expression of the F1F0-ATPase beta-subunit
from wine lactic acid bacteria. Systematic and Applied Microbiol-
ogy, 26, 350–356.





























