Embed Size (px)
Citation preview
8/9/2019 L13Biol261ERegulation2013
http://slidepdf.com/reader/full/l13biol261eregulation2013 1/30
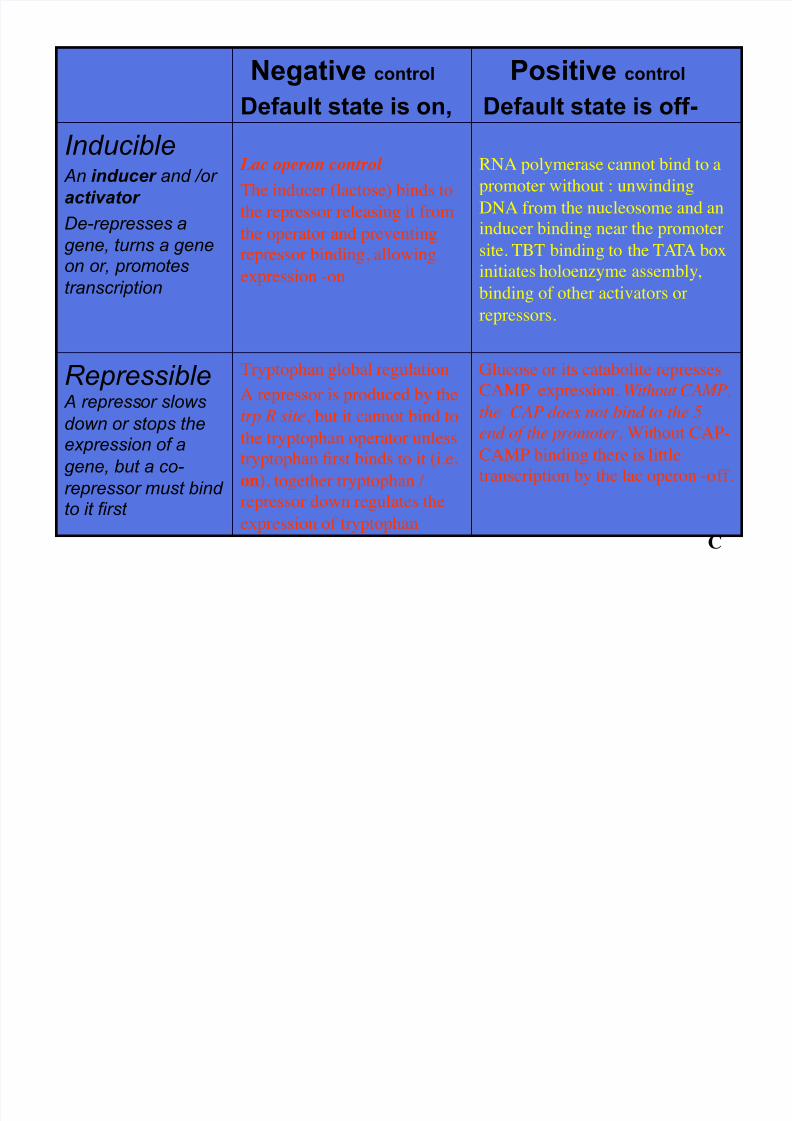
Negative control
Default state is on,
Positive control
Default state is off-
Inducible An inducer and /or
activator
De-represses a
gene, turns a geneon or, promotes
transcription
Lac operon control
The inducer (lactose) binds to
the repressor releasing it from
the operator and preventing
repressor binding, allowing
expression -on
RNA polymerase cannot bind to a
promoter without : unwinding
DNA from the nucleosome and an
inducer binding near the promoter
site. TBT binding to the TATA box
initiates holoenzyme assembly,binding of other activators or
repressors.
Repressible A repressor slows
down or stops theexpression of a
gene, but a co-
repressor must bindto it first
Tryptophan global regulation
A repressor is produced by the
trp R site, but it cannot bind tothe tryptophan operator unless
tryptophan first binds to it (i.e.
on), together tryptophan /
repressor down regulates the
expression of tryptophan
Glucose or its catabolite represses
CAMP expression. Without CAMP,
the CAP does not bind to the 5’
end of the promoter. Without CAP-
CAMP binding there is little
transcription by the lac operon -off.
C
8/9/2019 L13Biol261ERegulation2013
http://slidepdf.com/reader/full/l13biol261eregulation2013 2/30
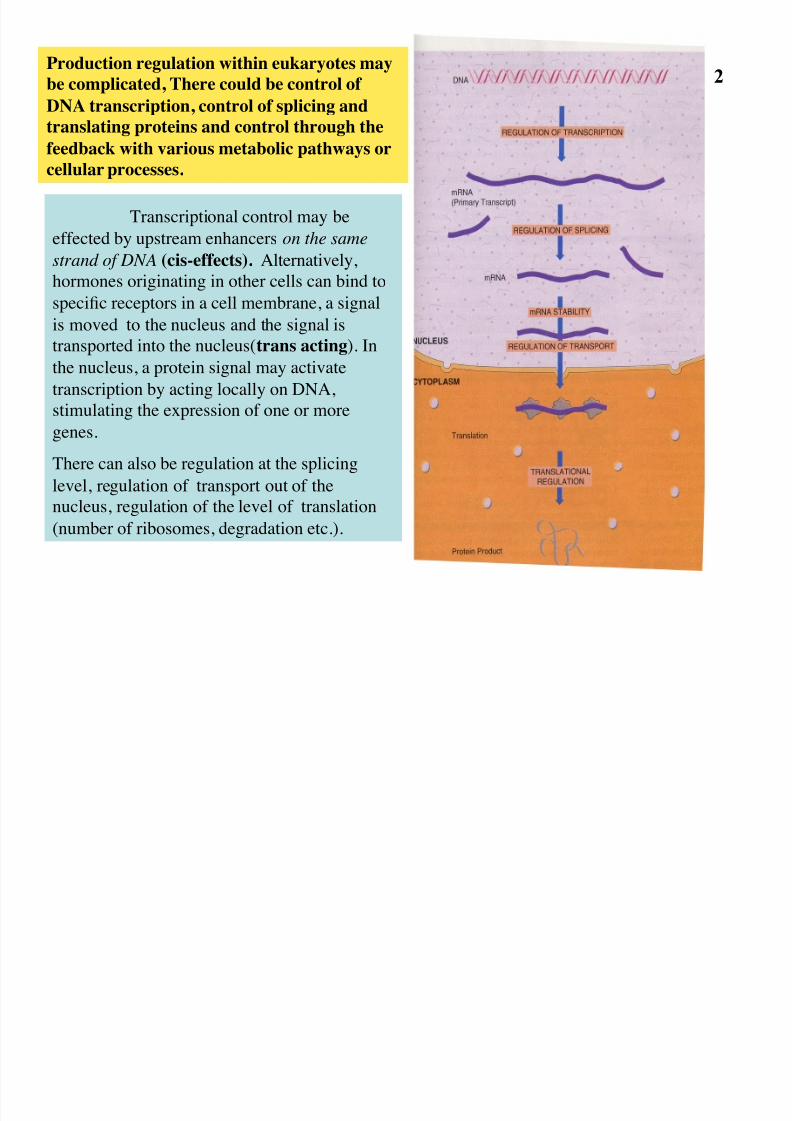
Transcriptional control may be
effected by upstream enhancers on the same
strand of DNA (cis-effects). Alternatively,
hormones originating in other cells can bind to
specific receptors in a cell membrane, a signal
is moved to the nucleus and the signal is
transported into the nucleus(trans acting). In
the nucleus, a protein signal may activate
transcription by acting locally on DNA,
stimulating the expression of one or moregenes.
There can also be regulation at the splicing
level, regulation of transport out of the
nucleus, regulation of the level of translation
(number of ribosomes, degradation etc.).
Production regulation within eukaryotes may
be complicated, There could be control of
DNA transcription, control of splicing and
translating proteins and control through the
feedback with various metabolic pathways or
cellular processes.
2
8/9/2019 L13Biol261ERegulation2013
http://slidepdf.com/reader/full/l13biol261eregulation2013 3/30
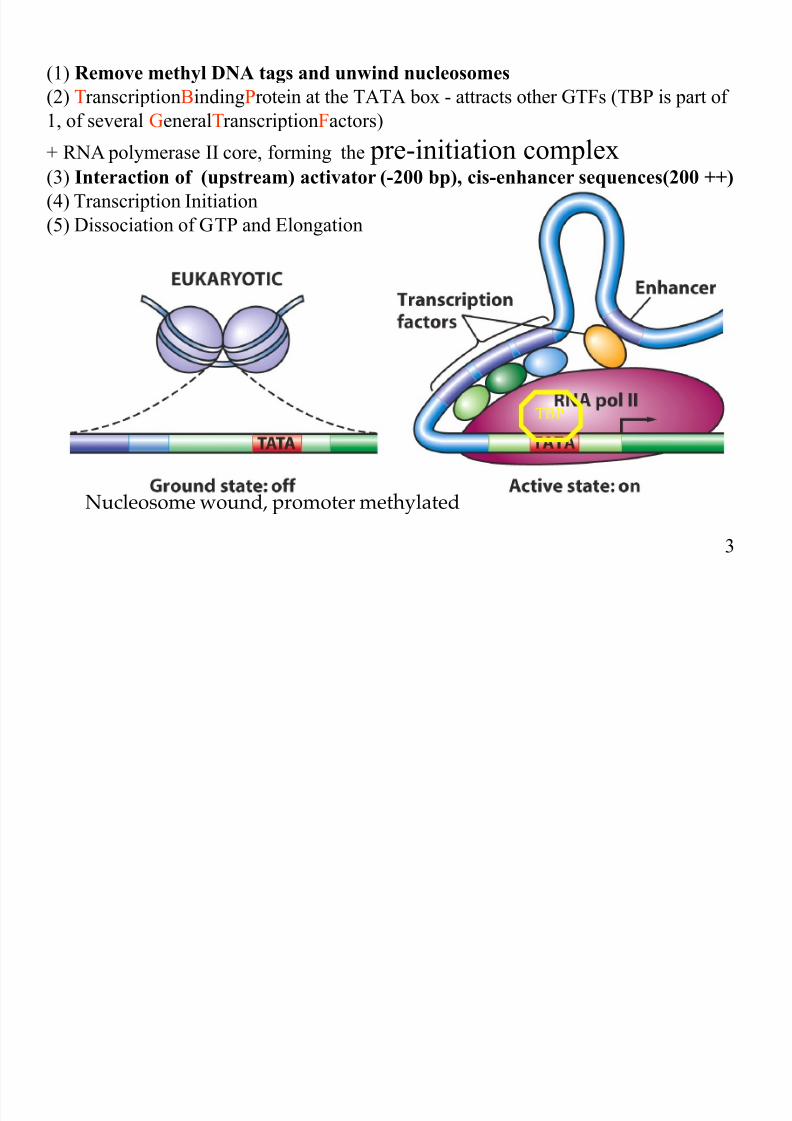
(1) Remove methyl DNA tags and unwind nucleosomes
(2) TranscriptionBindingProtein at the TATA box - attracts other GTFs (TBP is part of
1, of several GeneralTranscriptionFactors)
+ RNA polymerase II core, forming the pre-initiation complex
(3) Interaction of (upstream) activator (-200 bp), cis-enhancer sequences(200 ++)
(4) Transcription Initiation
(5) Dissociation of GTP and Elongation
TBP
3
Nucleosome wound, promoter methylated
8/9/2019 L13Biol261ERegulation2013
http://slidepdf.com/reader/full/l13biol261eregulation2013 4/30
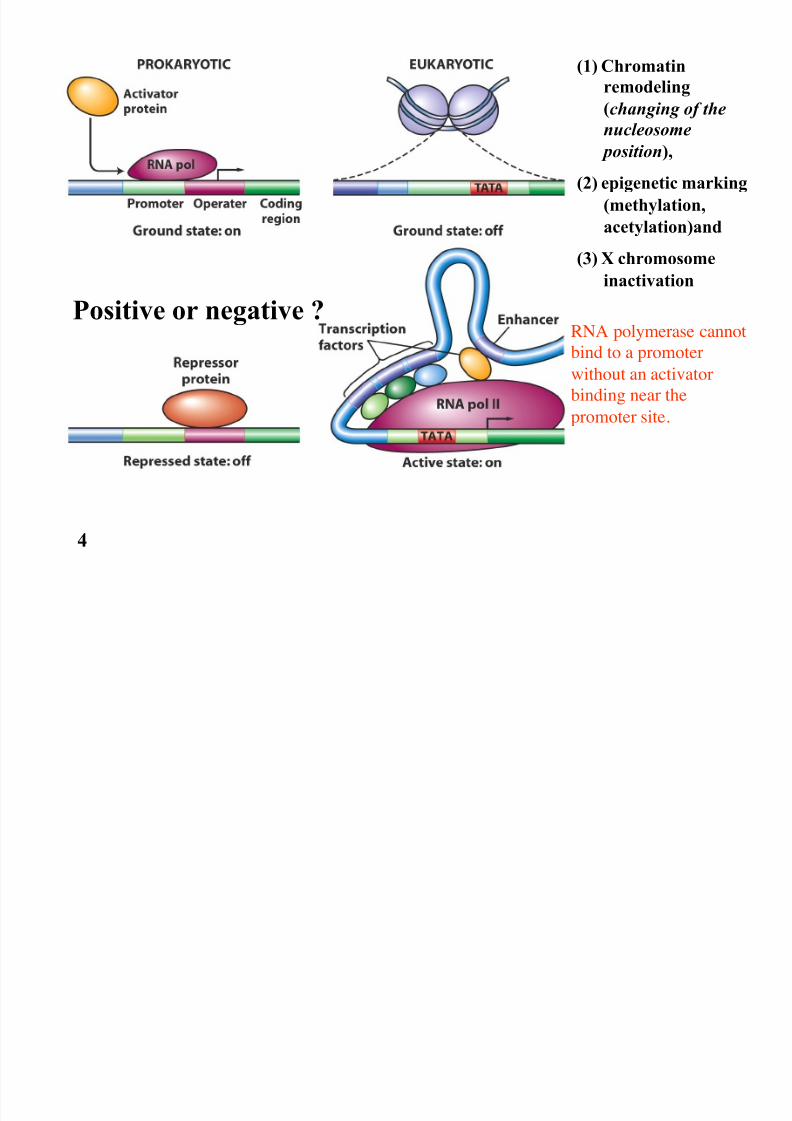
(1) Chromatin
remodeling
(changing of the
nucleosome position),
(2) epigenetic marking
(methylation,
acetylation)and
(3) X chromosome
inactivation
4
Positive or negative ? RNA polymerase cannot
bind to a promoter
without an activator
binding near the
promoter site.
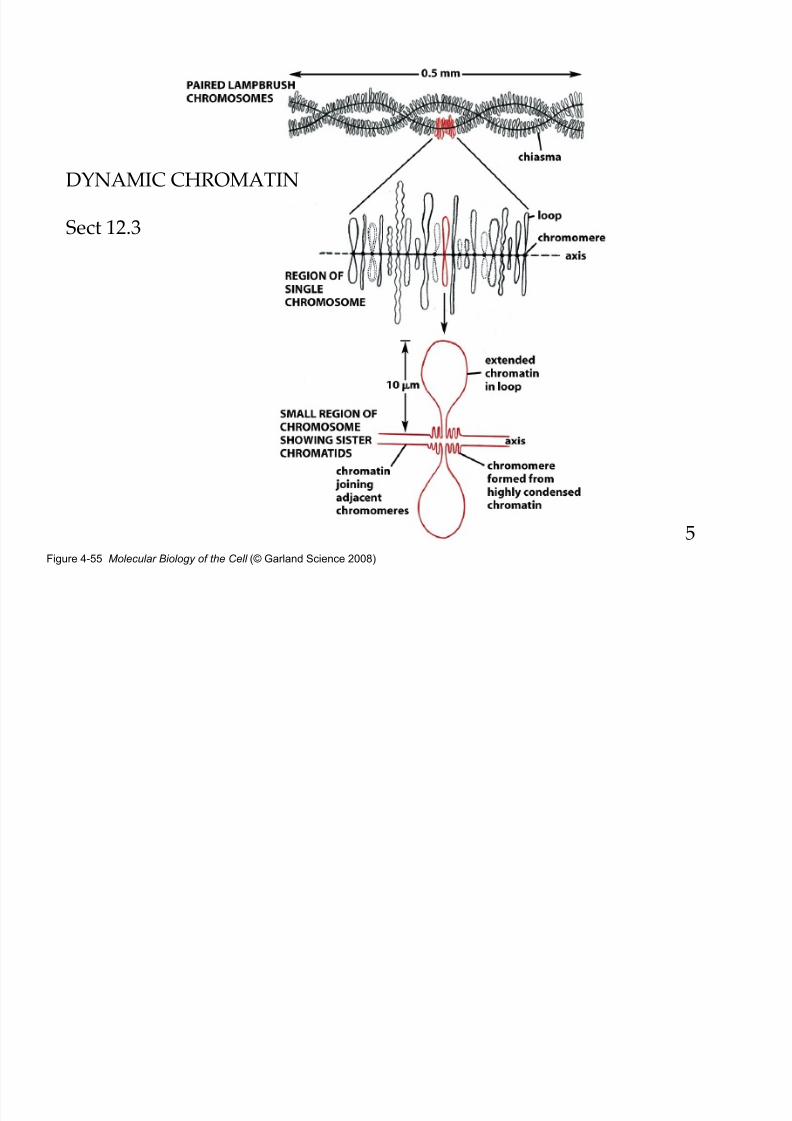
8/9/2019 L13Biol261ERegulation2013
http://slidepdf.com/reader/full/l13biol261eregulation2013 5/30
Figure 4-55 Molecular Biology of the Cell (© Garland Science 2008)
DYNAMIC CHROMATIN
Sect 12.3
5
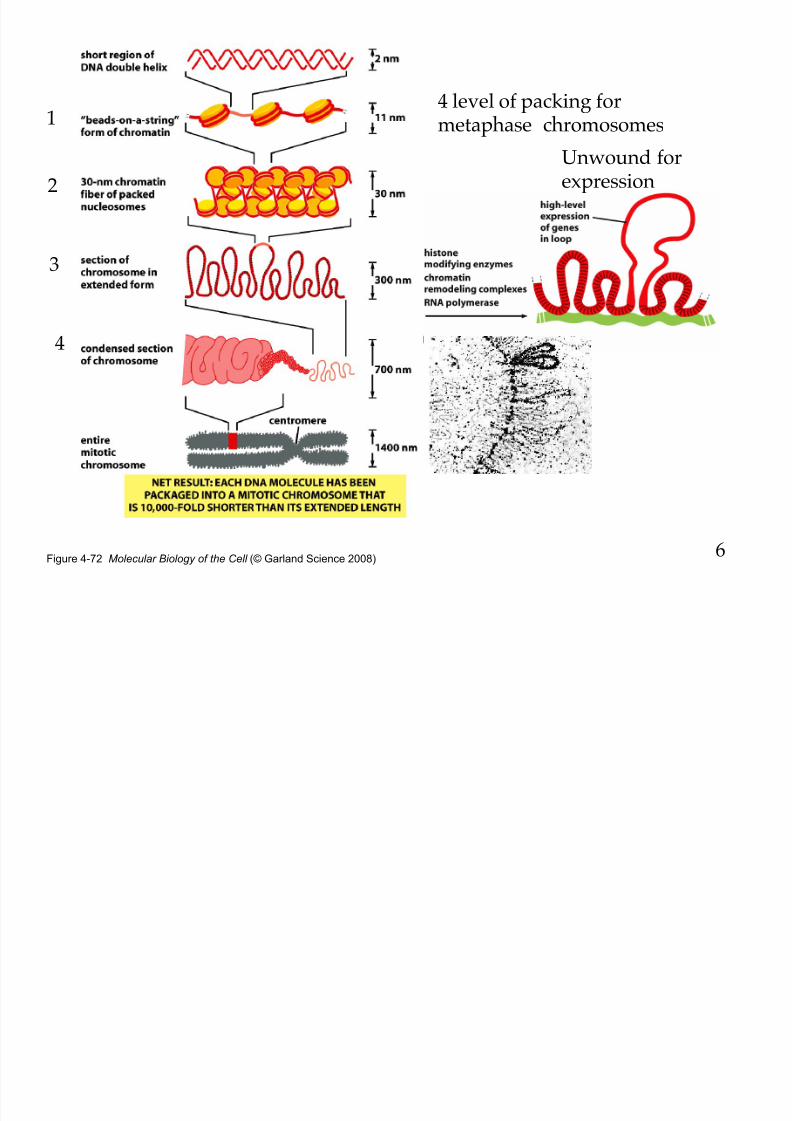
8/9/2019 L13Biol261ERegulation2013
http://slidepdf.com/reader/full/l13biol261eregulation2013 6/30
Figure 4-72 Molecular Biology of the Cell (© Garland Science 2008)
1
2
3
4
2
3
4
4 level of packing formetaphase chromosomes
Unwound forexpression
6
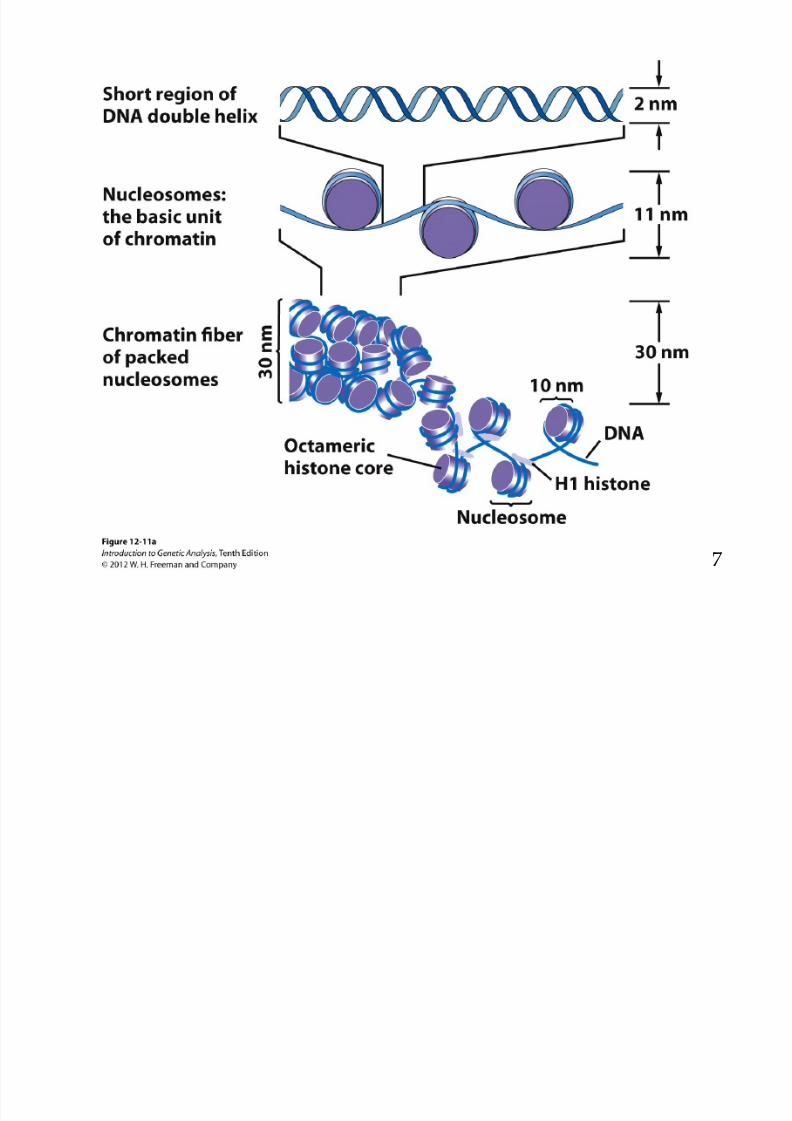
8/9/2019 L13Biol261ERegulation2013
http://slidepdf.com/reader/full/l13biol261eregulation2013 7/30
7
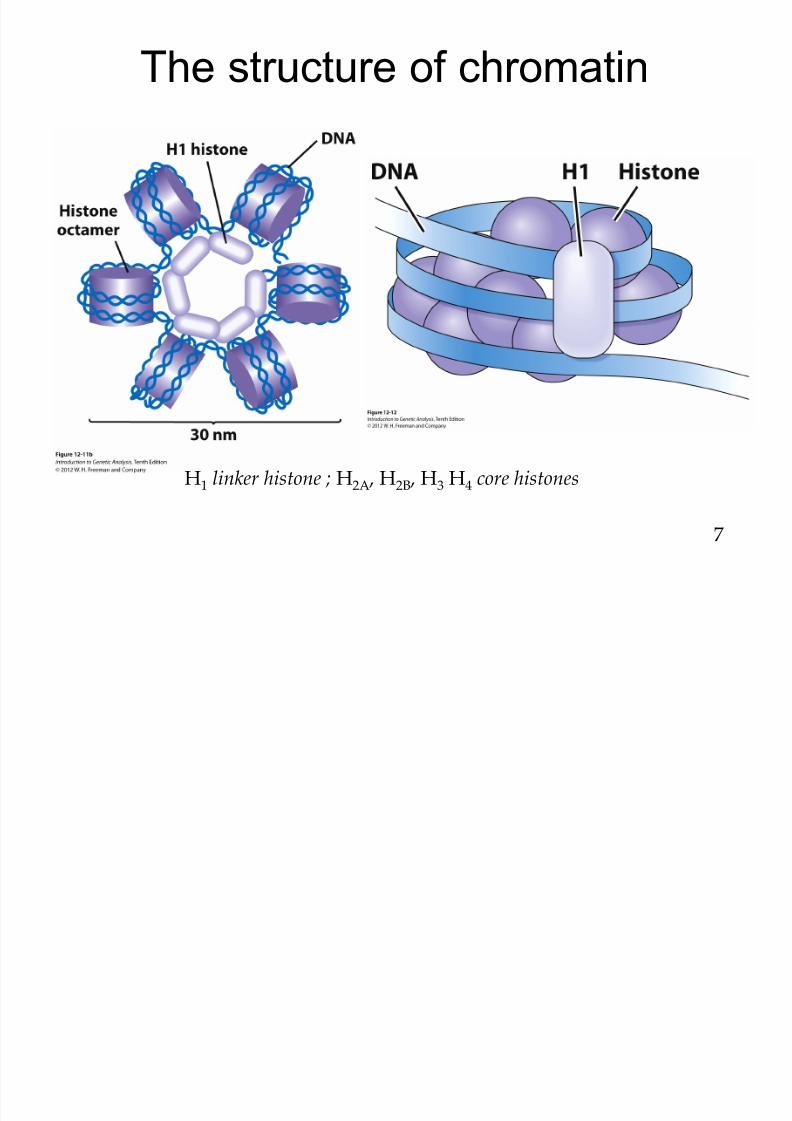
8/9/2019 L13Biol261ERegulation2013
http://slidepdf.com/reader/full/l13biol261eregulation2013 8/30
The structure of chromatin
H1 linker histone ; H2A , H2B , H3 H4 core histones
7
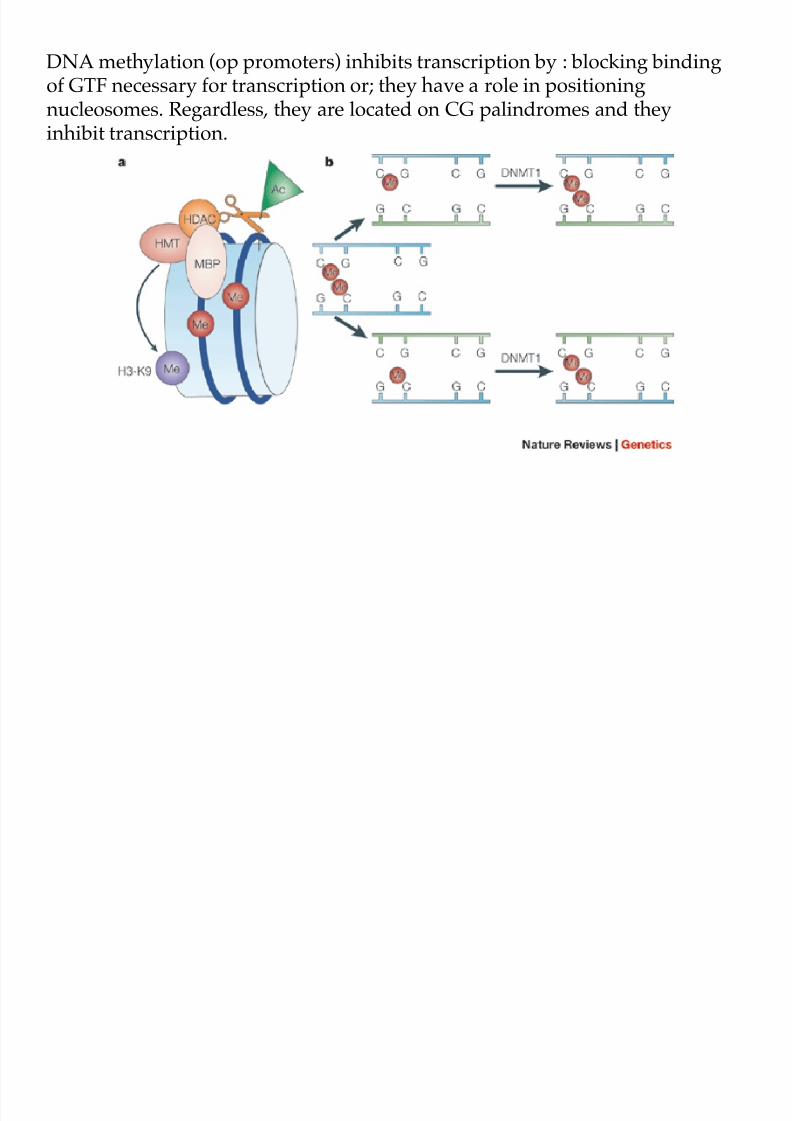
8/9/2019 L13Biol261ERegulation2013
http://slidepdf.com/reader/full/l13biol261eregulation2013 9/30
DNA methylation (op promoters) inhibits transcription by : blocking bindingof GTF necessary for transcription or; they have a role in positioningnucleosomes. Regardless, they are located on CG palindromes and they
inhibit transcription.
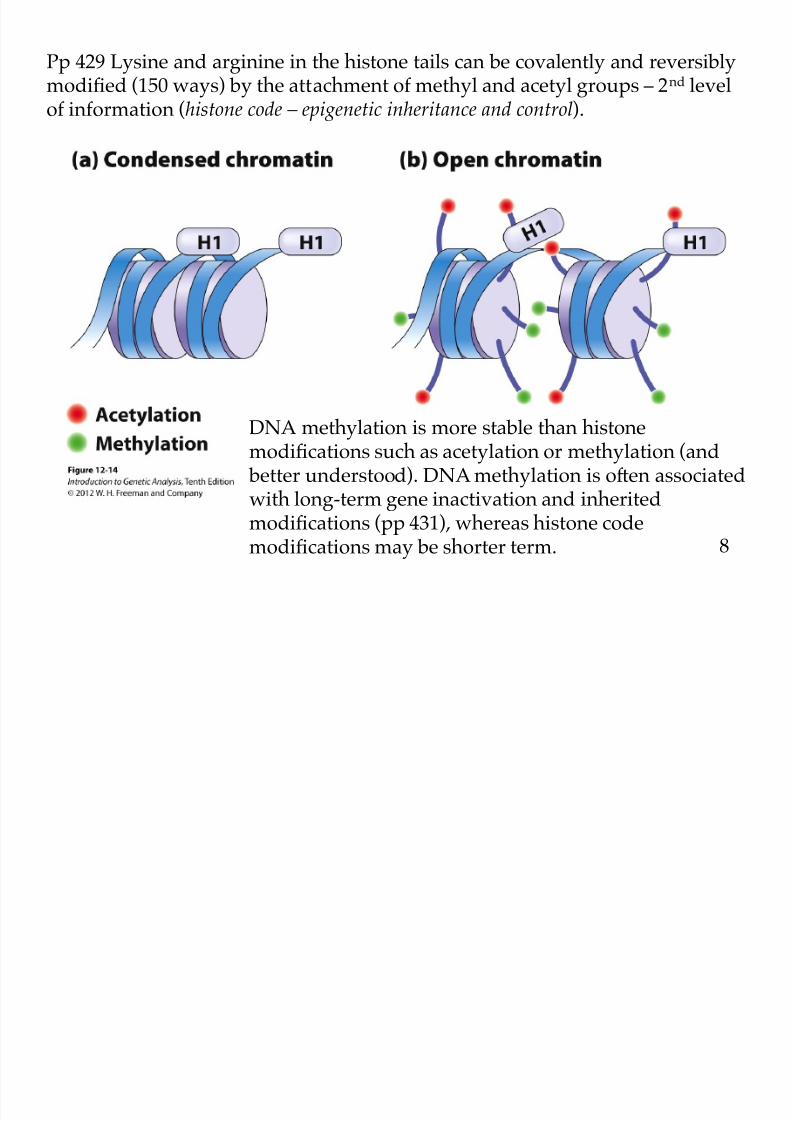
8/9/2019 L13Biol261ERegulation2013
http://slidepdf.com/reader/full/l13biol261eregulation2013 10/30
DNA methylation is more stable than histonemodifications such as acetylation or methylation (and
better understood). DNA methylation is often associatedwith long-term gene inactivation and inheritedmodifications (pp 431), whereas histone code
modifications may be shorter term.
Pp 429 Lysine and arginine in the histone tails can be covalently and reversiblymodified (150 ways) by the attachment of methyl and acetyl groups – 2nd levelof information (histone code – epigenetic inheritance and control).
8

8/9/2019 L13Biol261ERegulation2013
http://slidepdf.com/reader/full/l13biol261eregulation2013 11/30
Histones in active genes are hyperacetylated , +
Inactive genes are underacetylated (hypoacelated -).
How might acetylation affect chromatin remodeling & gene expression ?
a) Acetylation + : predispose a nucleosome to move. remodel
b) Acetylation + : alter the packing interaction between adjacentnucleosomes opening up a strand for transcription. remodel
c) Acetylation+ : with other histone modifications (methylation) influences
the binding of other regulatory proteins to DNA. expression
Replication- old disassembled andmixed with new nucleosomes( epigenetic marks inherited ?), parental strand is methylated
9
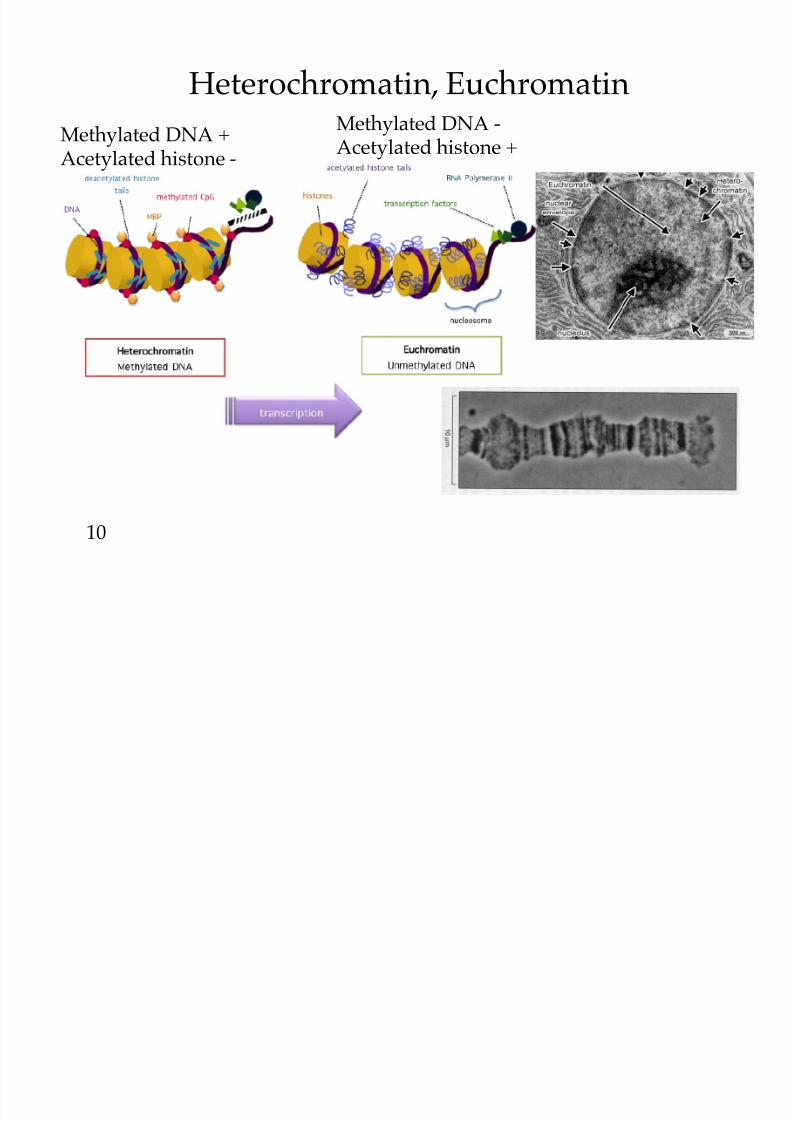
8/9/2019 L13Biol261ERegulation2013
http://slidepdf.com/reader/full/l13biol261eregulation2013 12/30
Heterochromatin, Euchromatin
10
Methylated DNA + Acetylated histone -
Methylated DNA - Acetylated histone +
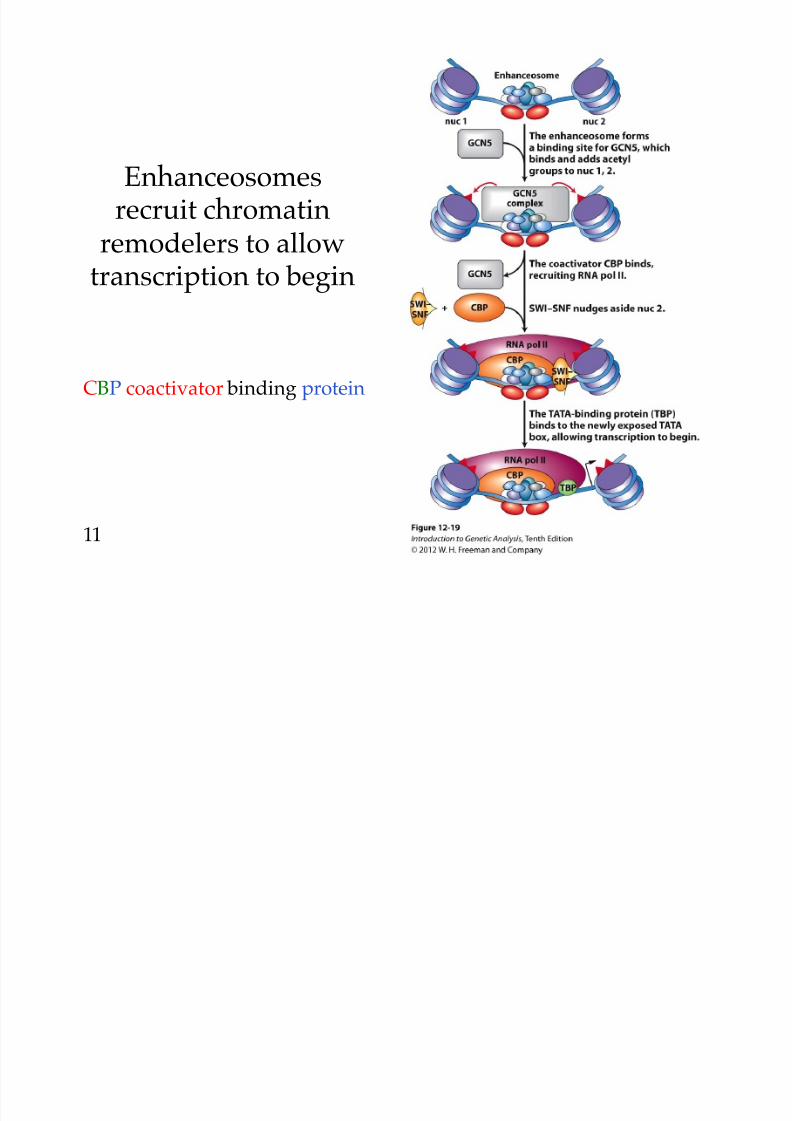
8/9/2019 L13Biol261ERegulation2013
http://slidepdf.com/reader/full/l13biol261eregulation2013 13/30
Enhanceosomesrecruit chromatin
remodelers to allow
transcription to begin
CBP coactivator binding protein
11
8/9/2019 L13Biol261ERegulation2013
http://slidepdf.com/reader/full/l13biol261eregulation2013 14/30
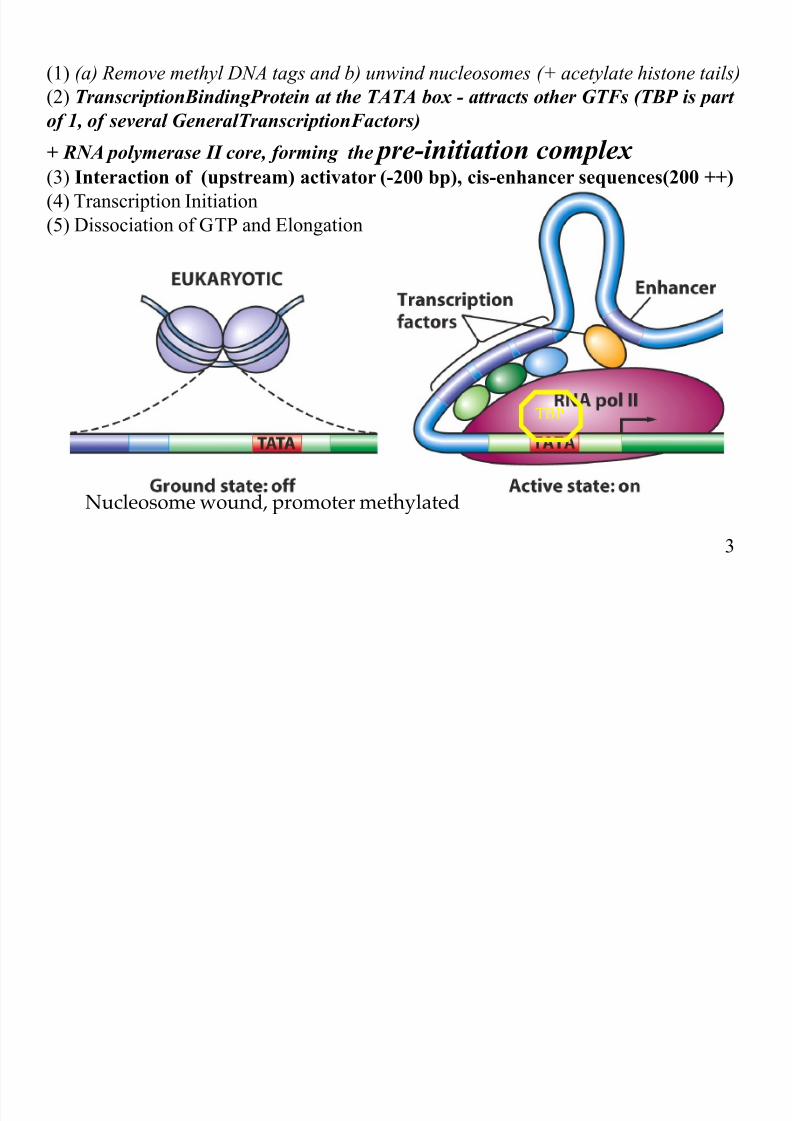
(1) (a) Remove methyl DNA tags and b) unwind nucleosomes (+ acetylate histone tails)
(2) TranscriptionBindingProtein at the TATA box - attracts other GTFs (TBP is part
of 1, of several GeneralTranscriptionFactors)
+ RNA polymerase II core, forming the pre-initiation complex (3) Interaction of (upstream) activator (-200 bp), cis-enhancer sequences(200 ++)
(4) Transcription Initiation
(5) Dissociation of GTP and Elongation
TBP
3
Nucleosome wound, promoter methylated

8/9/2019 L13Biol261ERegulation2013
http://slidepdf.com/reader/full/l13biol261eregulation2013 15/30
Separable, Dual Bindingand Activation Roles inUpstream ActivationSequences
Activators: Gal 4 dimer interacts directlywith DNA and recruits TFIID and
indirectly via a mediator protein torecruit RNA polymeraseII
12

8/9/2019 L13Biol261ERegulation2013
http://slidepdf.com/reader/full/l13biol261eregulation2013 16/30
Model for eukaryotic promoters
13
In eukaryoticgene regulationof expression
there are bothactivators,enhancers,silencers andrepressors that
each affect therate oftranscription

8/9/2019 L13Biol261ERegulation2013
http://slidepdf.com/reader/full/l13biol261eregulation2013 17/30
Eukaryotic Expression control evolved differently in different lineages (TPhillips and L. Hoopes Nature Education 1:1)
“Complexity of transcriptional control can be illustrated by comparingthe number and locations of cis-control elements in higher and lowereukaryotes.
For instance, Drosophila typically has several enhancers for a single geneof 2 to 3 kilobases, scattered over a large (10 kilobase) region of DNA,while,
yeast have no enhancers but instead use one UAS sequence per gene,
located upstream (activators).”
14
8/9/2019 L13Biol261ERegulation2013
http://slidepdf.com/reader/full/l13biol261eregulation2013 18/30
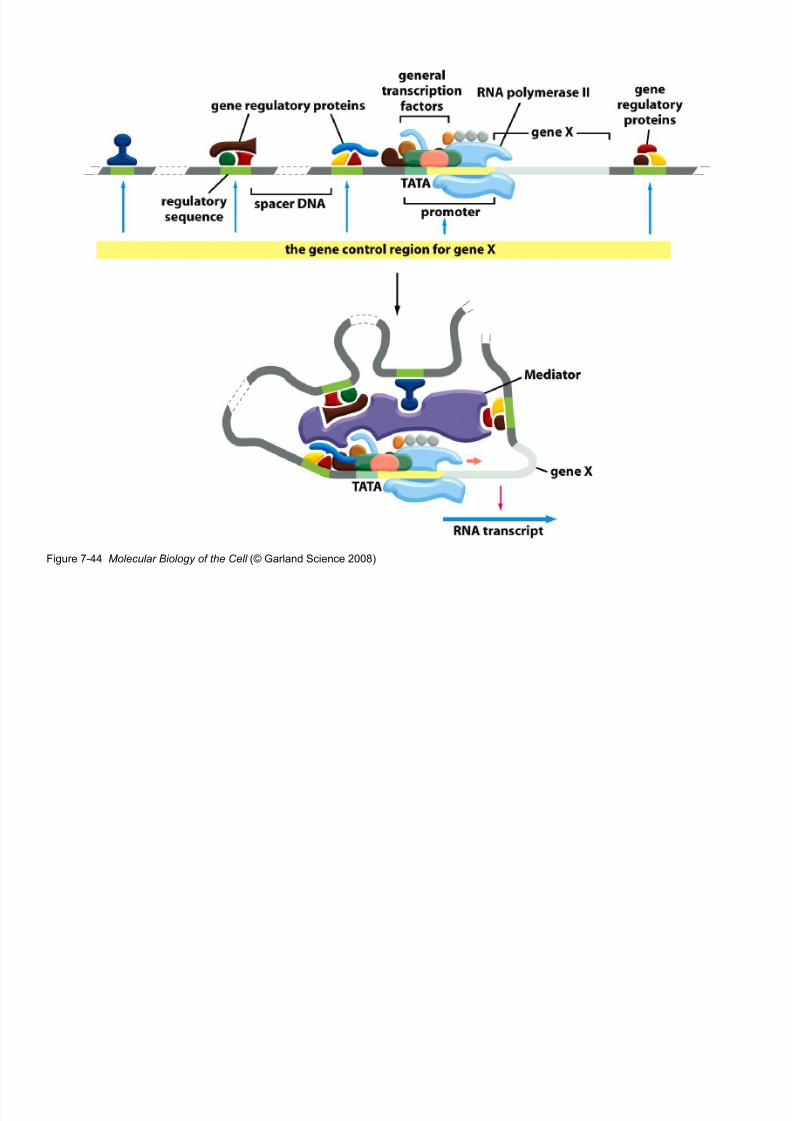
Figure 7-44 Molecular Biology of the Cell (© Garland Science 2008)
8/9/2019 L13Biol261ERegulation2013
http://slidepdf.com/reader/full/l13biol261eregulation2013 19/30
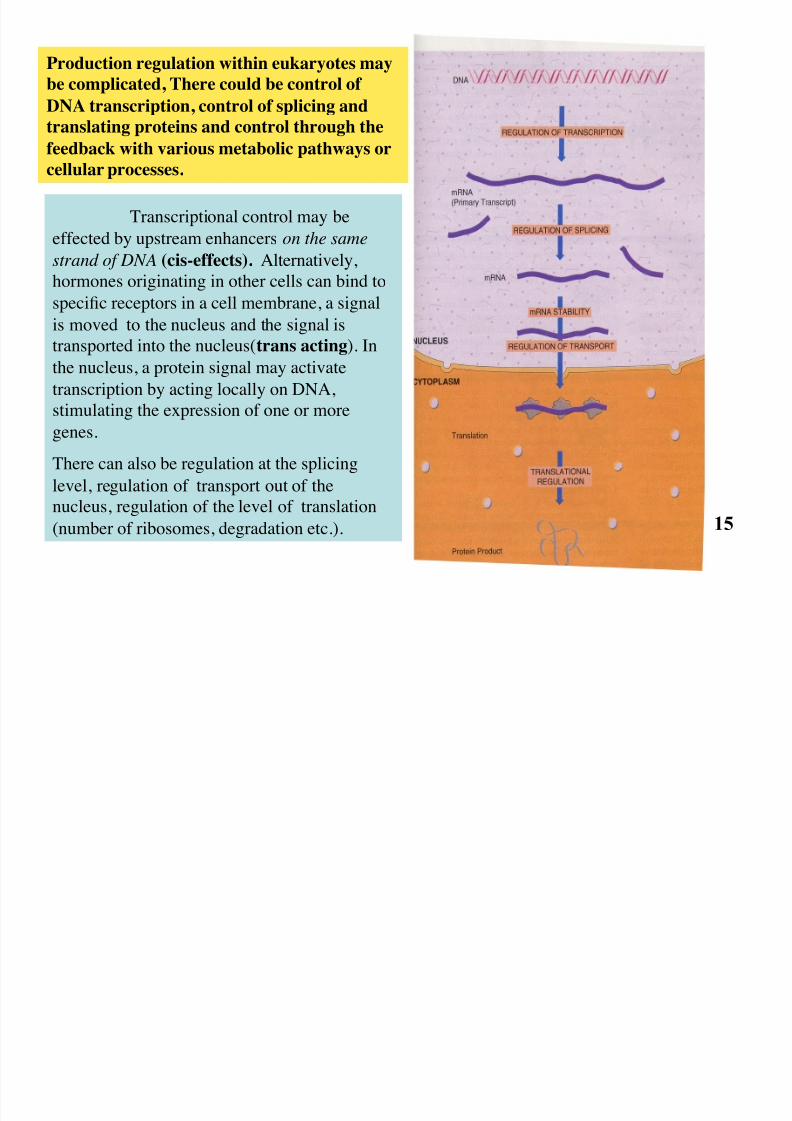
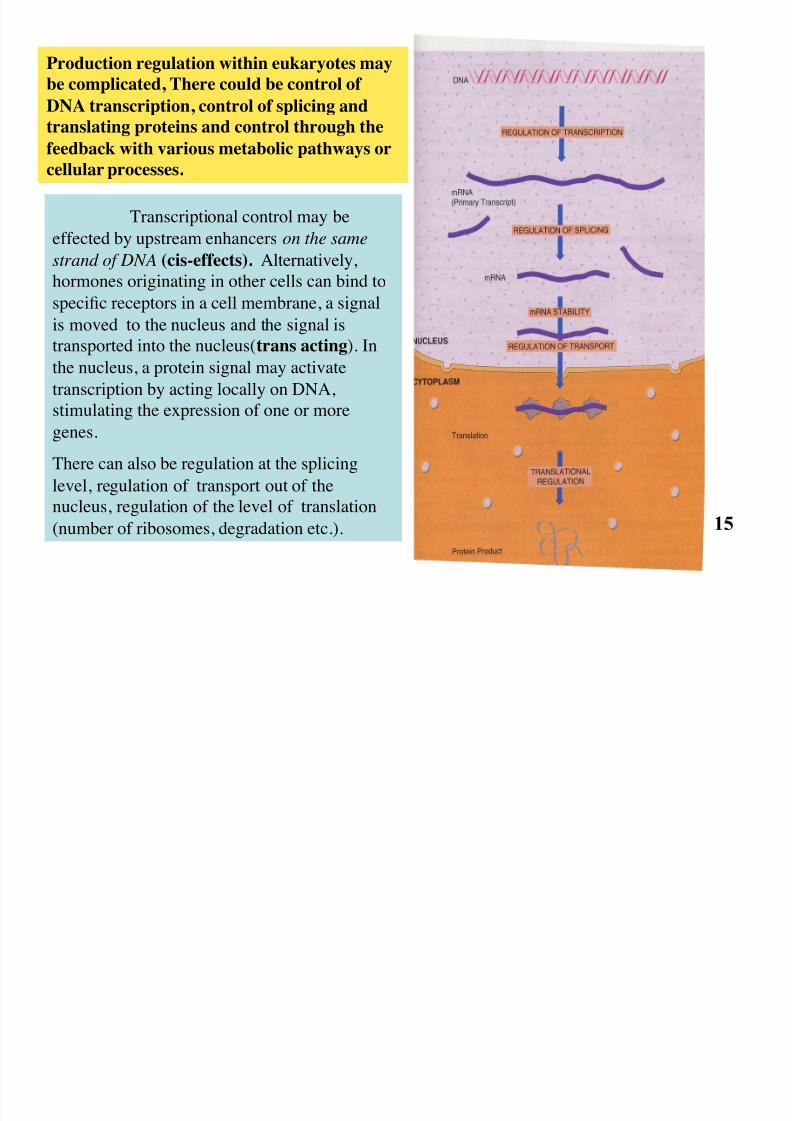
Transcriptional control may be
effected by upstream enhancers on the same
strand of DNA (cis-effects). Alternatively,
hormones originating in other cells can bind tospecific receptors in a cell membrane, a signal
is moved to the nucleus and the signal is
transported into the nucleus(trans acting). In
the nucleus, a protein signal may activate
transcription by acting locally on DNA,
stimulating the expression of one or moregenes.
There can also be regulation at the splicing
level, regulation of transport out of the
nucleus, regulation of the level of translation
(number of ribosomes, degradation etc.).
Production regulation within eukaryotes may
be complicated, There could be control of
DNA transcription, control of splicing and
translating proteins and control through the
feedback with various metabolic pathways or
cellular processes.
15

8/9/2019 L13Biol261ERegulation2013
http://slidepdf.com/reader/full/l13biol261eregulation2013 20/30
Castel and Martensen Nature Reviews Genetics 14
•
Small RNA (<200 bp) that can regulate genes:
• 1) micro RNA (miRNA) – hairpin derived RNA with
imperfect complementarity to targets that interfere withtranslation.
•
2)small interfering RNA (siRNA) perfect complementarity totheir target, and they degrade transcripts.
• 3)PIWI interacting RNA (piRNA) which target transposontranscripts in animals
•
The mechanistic details of 1-3 are converging and are morecommonly all referred to as RNAi
• RNAi - temporary or reversible gene expression knockouts.
See 8.5 fig 8-21to 8-24,
8/9/2019 L13Biol261ERegulation2013
http://slidepdf.com/reader/full/l13biol261eregulation2013 21/30
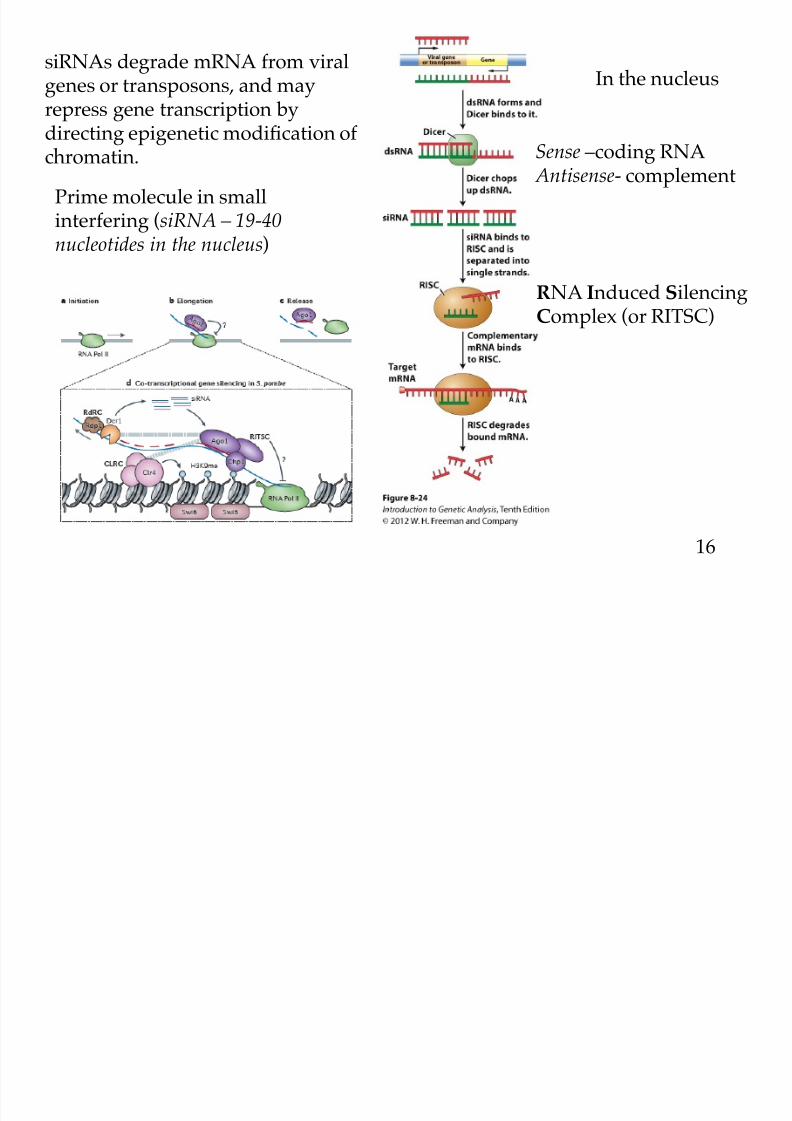
Prime molecule in smallinterfering (siRNA – 19-40nucleotides in the nucleus)
Sense –coding RNA
Antisense- complement
In the nucleus
RNA Induced SilencingComplex (or RITSC)
16
siRNAs degrade mRNA from viralgenes or transposons, and mayrepress gene transcription by
directing epigenetic modification ofchromatin.
8/9/2019 L13Biol261ERegulation2013
http://slidepdf.com/reader/full/l13biol261eregulation2013 22/30
Transcriptional control may be
effected by upstream enhancers on the same
strand of DNA (cis-effects). Alternatively,
hormones originating in other cells can bind tospecific receptors in a cell membrane, a signal
is moved to the nucleus and the signal is
transported into the nucleus(trans acting). In
the nucleus, a protein signal may activate
transcription by acting locally on DNA,
stimulating the expression of one or moregenes.
There can also be regulation at the splicing
level, regulation of transport out of the
nucleus, regulation of the level of translation
(number of ribosomes, degradation etc.).
Production regulation within eukaryotes may
be complicated, There could be control of
DNA transcription, control of splicing and
translating proteins and control through the
feedback with various metabolic pathways or
cellular processes.
15
8/9/2019 L13Biol261ERegulation2013
http://slidepdf.com/reader/full/l13biol261eregulation2013 23/30
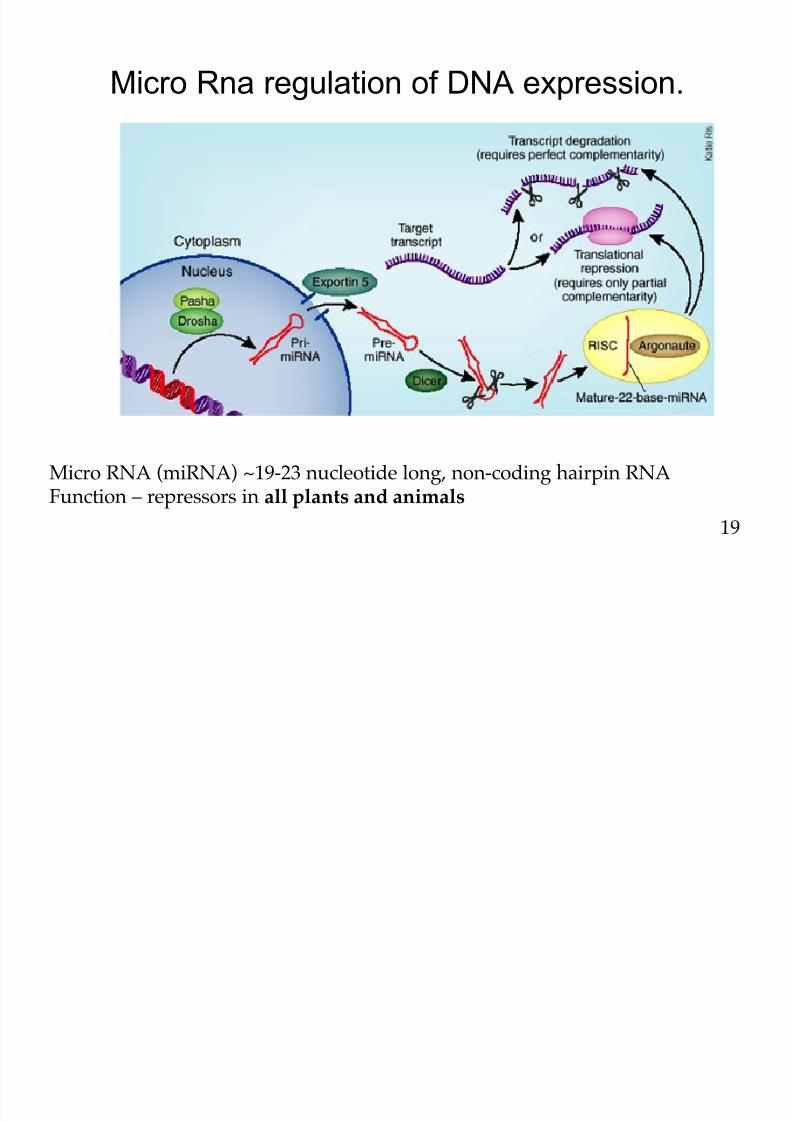
Micro Rna regulation of DNA expression.
19
Micro RNA (miRNA) ~19-23 nucleotide long, non-coding hairpin RNAFunction – repressors in all plants and animals
8/9/2019 L13Biol261ERegulation2013
http://slidepdf.com/reader/full/l13biol261eregulation2013 24/30
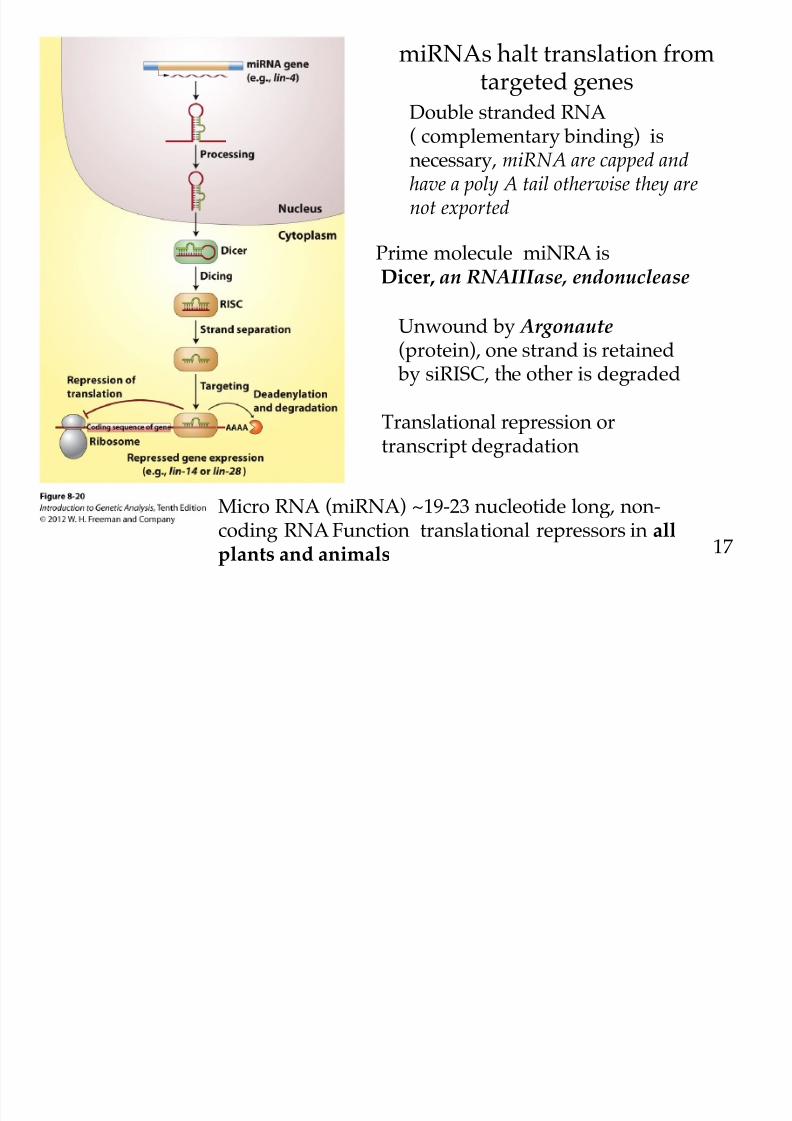
miRNAs halt translation fromtargeted genes
Double stranded RNA
( complementary binding) isnecessary, miRNA are capped andhave a poly A tail otherwise they arenot exported
Prime molecule miNRA is
Dicer, an RNAIIIase, endonuclease
Unwound by Argonaute(protein), one strand is retained
by siRISC, the other is degraded
Translational repression ortranscript degradation
17
Micro RNA (miRNA) ~19-23 nucleotide long, non-coding RNA Function translational repressors in all
plants and animals
8/9/2019 L13Biol261ERegulation2013
http://slidepdf.com/reader/full/l13biol261eregulation2013 25/30
18
8/9/2019 L13Biol261ERegulation2013
http://slidepdf.com/reader/full/l13biol261eregulation2013 26/30
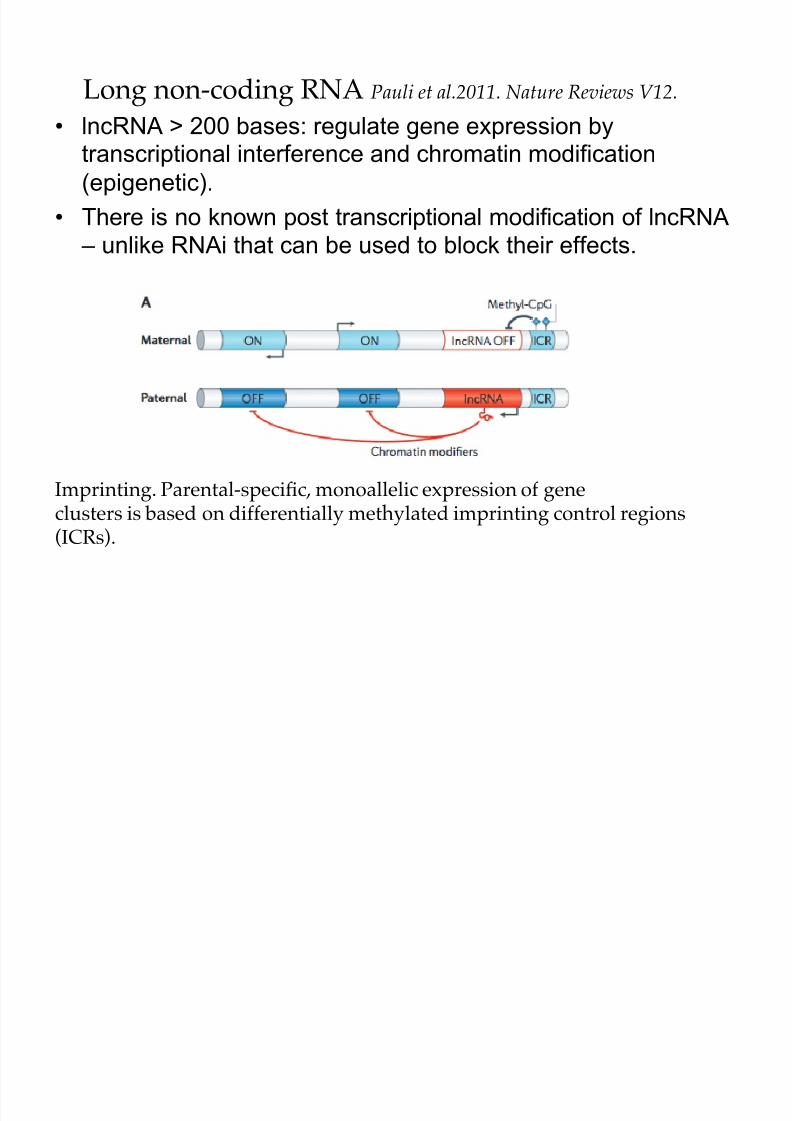
• lncRNA > 200 bases: regulate gene expression bytranscriptional interference and chromatin modification
(epigenetic).
• There is no known post transcriptional modification of lncRNA – unlike RNAi that can be used to block their effects.
Long non-coding RNA Pauli et al.2011. Nature Reviews V12.
Imprinting. Parental-specific, monoallelic expression of geneclusters is based on differentially methylated imprinting control regions(ICRs).
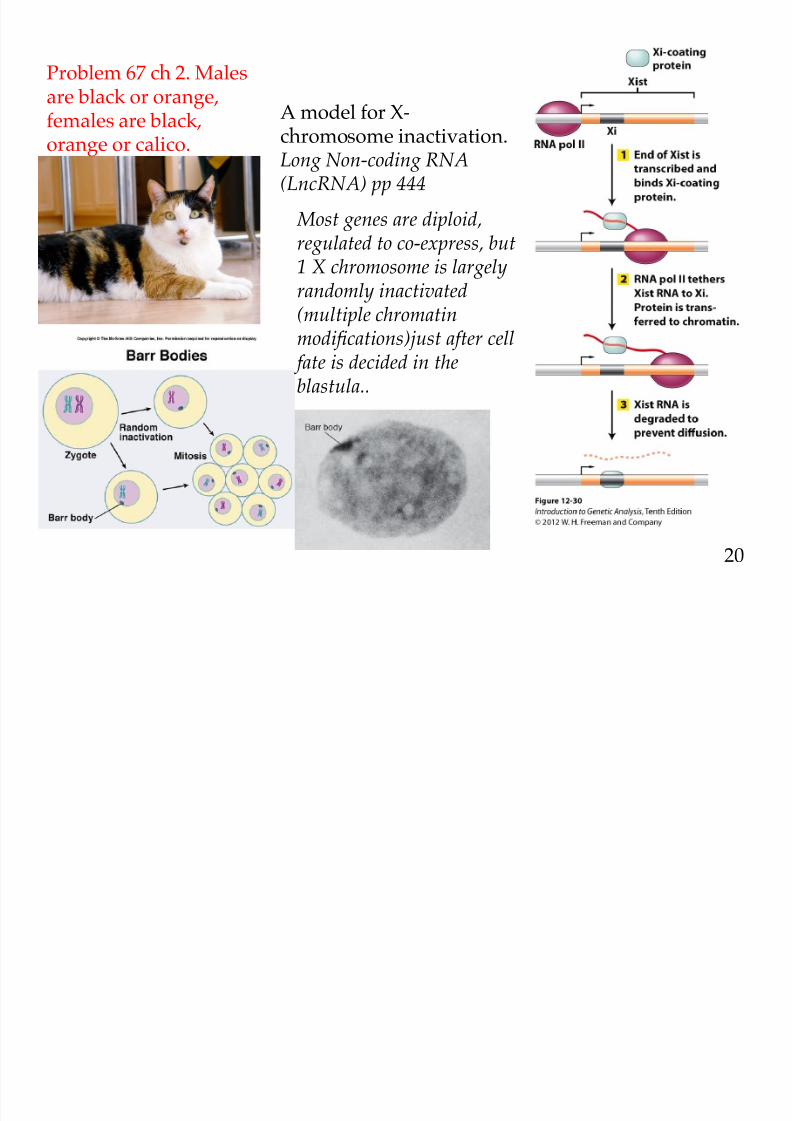
8/9/2019 L13Biol261ERegulation2013
http://slidepdf.com/reader/full/l13biol261eregulation2013 27/30
A model for X-
chromosome inactivation.Long Non-coding RNA(LncRNA) pp 444
Most genes are diploid,regulated to co-express, but1 X chromosome is largelyrandomly inactivated(multiple chromatinmodifications)just after cell fate is decided in theblastula..
Problem 67 ch 2. Malesare black or orange,females are black,
orange or calico.
20
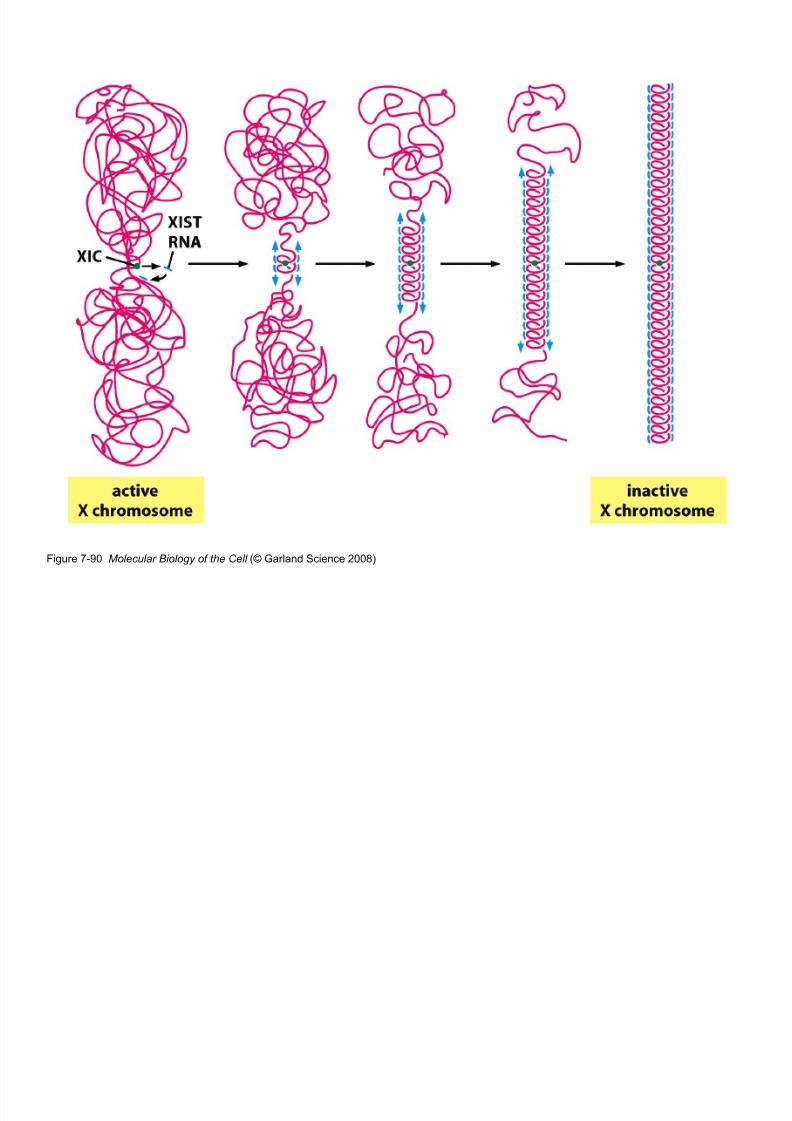
8/9/2019 L13Biol261ERegulation2013
http://slidepdf.com/reader/full/l13biol261eregulation2013 28/30
Figure 7-90 Molecular Biology of the Cell (© Garland Science 2008)
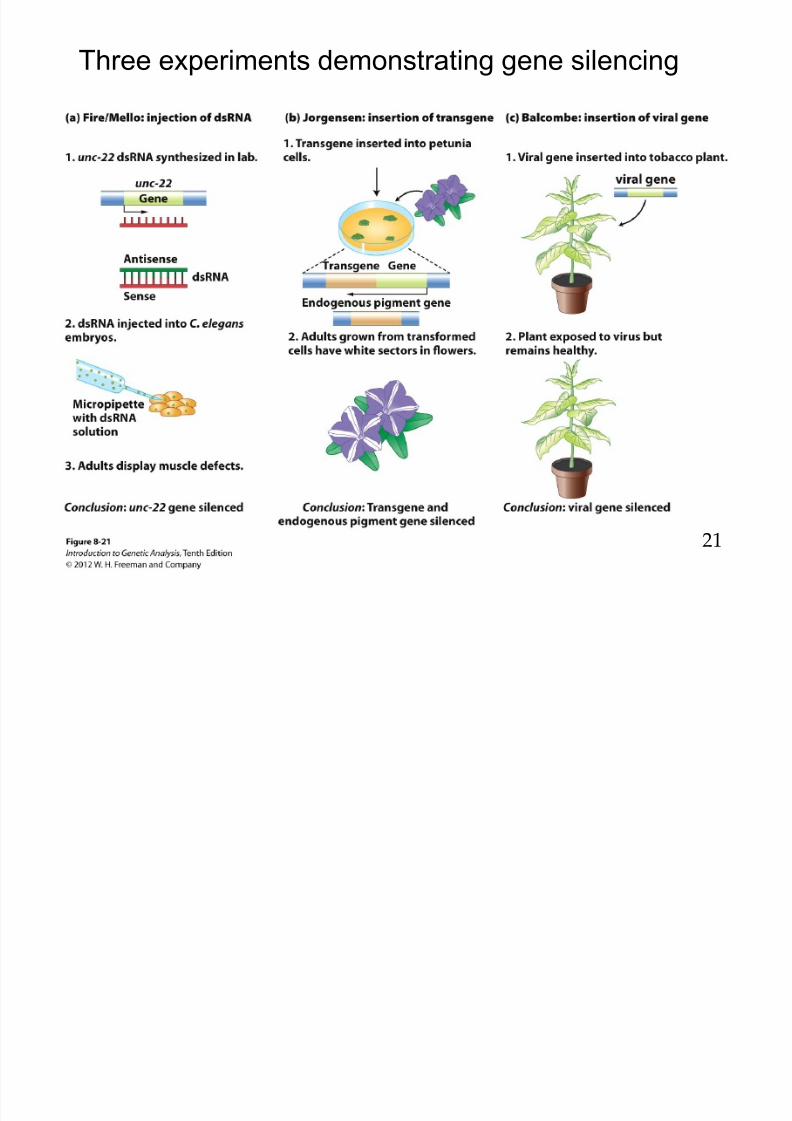
8/9/2019 L13Biol261ERegulation2013
http://slidepdf.com/reader/full/l13biol261eregulation2013 29/30
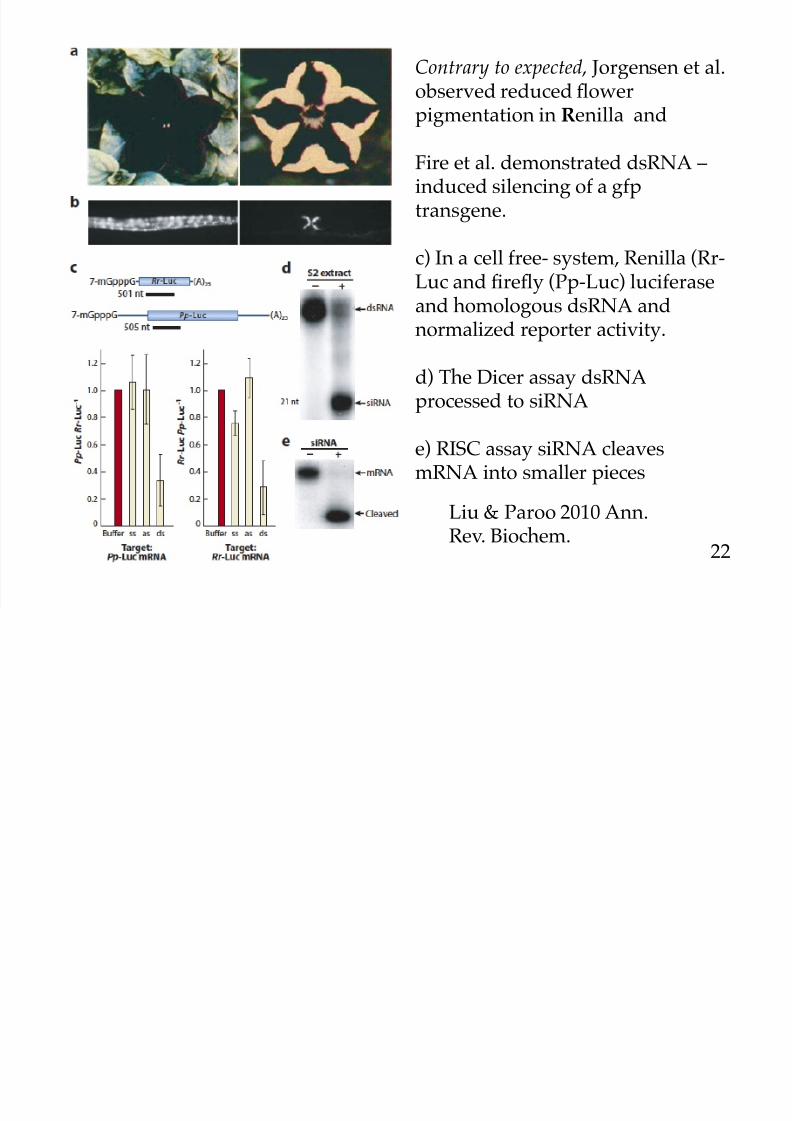
Three experiments demonstrating gene silencing
21
8/9/2019 L13Biol261ERegulation2013
http://slidepdf.com/reader/full/l13biol261eregulation2013 30/30
Contrary to expected , Jorgensen et al.observed reduced flowerpigmentation in Renilla and
Fire et al. demonstrated dsRNA –induced silencing of a gfptransgene.
c) In a cell free- system, Renilla (Rr-
Luc and firefly (Pp-Luc) luciferaseand homologous dsRNA andnormalized reporter activity.
d) The Dicer assay dsRNAprocessed to siRNA
e) RISC assay siRNA cleavesmRNA into smaller pieces
Liu & Paroo 2010 Ann.Rev. Biochem.
22





























