Embed Size (px)
Citation preview

MOLECULAR AND CELLULAR BIOLOGY,0270-7306/99/$04.0010
Apr. 1999, p. 2585–2593 Vol. 19, No. 4
Copyright © 1999, American Society for Microbiology. All Rights Reserved.
Ku-Dependent Nonhomologous DNA End Joiningin Xenopus Egg Extracts
PAUL LABHART*
Department of Molecular and Experimental Medicine, The Scripps Research Institute,La Jolla, California 92037
Received 9 June 1998/Returned for modification 13 July 1998/Accepted 17 December 1998
An extract from activated Xenopus eggs joins both matching and nonmatching ends of exogenous linear DNAsubstrates with high efficiency and fidelity (P. Pfeiffer and W. Vielmetter, Nucleic Acids Res. 16:907–924, 1988).In mammalian cells, such nonhomologous end joining (NHEJ) is known to require the Ku heterodimer, acomponent of DNA-dependent protein kinase. Here I investigated whether Ku is also required for the in vitroreaction in the egg extract. Immunological assays indicate that Ku is very abundant in the extract. I found thatall NHEJ was inhibited by autoantibodies against Ku and that NHEJ between certain combinations of DNAends was also decreased after immunodepletion of Ku from the extract. The formation of a joint between a DNAend with a 5*-protruding single strand (PSS) and an end with a 3*-PSS, between two ends with 3*-PSS, andbetween two blunt ends was most Ku dependent. On the other hand, NHEJ between two DNA ends bearing5*-PSS was Ku independent. These results show that the Xenopus cell-free system will be useful to biochemicallydissect the role of Ku in eukaryotic NHEJ.
Xenopus laevis has proved to be a useful system for studyingboth homologous recombination and nonhomologous DNAend joining (NHEJ) of exogenous DNA molecules. Both pro-cesses were studied in vivo by microinjection of DNA as well asin vitro in extracts derived from various stages of oogenesis andearly embryogenesis (12, 18, 26). In oocytes, homologous re-combination is the prevalent mechanism for the joining of twolinear DNA molecules and NHEJ is virtually undetectable.Upon oocyte maturation and in early embryos, NHEJ becomesthe prominent mechanism, even though absolute levels of ho-mologous recombination remain little changed. An extractfrom fertilized or activated Xenopus eggs has been an invalu-able tool for the detailed characterization of the NHEJ prod-ucts generated from defined substrates (32). These experi-ments have shown that the egg extract has the capability to joinpairs of DNA ends bearing various combinations of 59-pro-truding single strands (PSS), 39-PSS, and blunt ends, as well aschemically modified ends (15), with high efficiency and preci-sion. Thus, DNA ends are typically joined without nucleotideloss by end-to-end alignment and filling-in of any gaps (“fill-in”mode). Somewhat more heterogeneous and less-predictableproducts are formed with pairs of nonmatching 59- or 39-PSS,in which case the antiparallel PSS align by forming overlapswhose extent is influenced by the sequence in the PSS (“over-lap” mode) (31). This largely error-free NHEJ appears to be acharacteristic of the Xenopus egg extract and sets it apart fromsimilar cell-free systems derived from mammalian cells where,possibly because of higher levels of exonucleases, deletionsduring NHEJ are more frequent (9, 10, 29).
Based on the findings with the Xenopus egg extract it waspostulated that there must be an “alignment factor” that holdsthe two DNA ends in place for the nucleotide fill-in and strandligation reaction. The existence of such a factor was particu-larly suggested by the finding that fill-in of 39-PSS termini canprecede ligation, which implies that fill-in DNA synthesis of
one strand can proceed past a nick in the opposite strand (39).Such a process is difficult to envision without an apparatus thatholds the two DNA ends together.
Independent of this work in X. laevis, genetic studies withmammalian cells established that the three protein compo-nents of DNA-dependent protein kinase (DNA-PK), the 470-kDa catalytic subunit (DNA-PKcs), and the Ku heterodimer(Ku70 and Ku80) are all required for double-strand breakrepair upon damage by ionizing radiation and also for therelated process of V(D)J recombination during lymphoid de-velopment (4, 21, 37, 38; reviewed in references 2, 7, 11, and20). Some of these genetic findings have been confirmed withbiochemical experiments in cell-free systems (8, 40). DNA-PKcs is a member of the phosphatidylinositol kinase-relatedkinases, is activated by double-stranded DNA ends, and pref-erentially phosphorylates proteins that are bound to the sameDNA molecule. Even though the Ku proteins are thought totarget DNA-PK to DNA and thus cause the activation of thecatalytic subunit, there is evidence that DNA-PKcs and Ku donot always act as a complex. Genetic studies show that only theKu heterodimer, but not DNA-PKcs, is required for the for-mation of the signal joints during V(D)J recombination (4).Furthermore, Ku appears to have a separate function in main-taining telomere length and end structure in yeast cells (5, 14).Finally, atomic force microscopy studies provide evidence thatboth the catalytic subunit and Ku by themselves can holdtogether two linear DNA molecules (30, 41). Both componentsof DNA-PK could thus fulfill a structural alignment functionduring double-strand break repair.
Despite immense progress in this field in recent years, theprecise role of DNA-PK during NHEJ remains unknown.However, the data summarized above suggest that NHEJ inthe Xenopus egg extract is also a DNA-PK-dependent reactionand that this system thus might be useful to further elucidatethe role of DNA-PK during NHEJ. In this study I have usedantibody inhibition and immunodepletion experiments to showthat the DNA-PK component Ku is indeed required for NHEJin this cell-free system. I discuss the possibility that Ku is thepostulated alignment factor present in the egg extract.
* Mailing address: Department of Molecular and ExperimentalMedicine, The Scripps Research Institute, Mail drop SBR10, 10550North Torrey Pines Rd., La Jolla, CA 92037. Phone: (619) 784-2406.Fax: (619) 784-2960. E-mail: [email protected].
2585
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 17
Oct
ober
202
1 by
114
.69.
161.
189.

MATERIALS AND METHODS
Reagents. Purified HeLa Ku was generously provided by W. S. Dynan and S.Yoo (Augusta, Ga.). Ku protein was stored in 0.1 M KCl–50 mM Tris-HCl (pH7.9)–1 mM EDTA–0.02% Tween 20–20% glycerol–1 mM dithiothreitol (DTT)(“Ku buffer”). Purified monoclonal antibody (MAb) N3H10 was obtained fromKamiya Biomedical Company (Seattle, Wash.). Human autoimmune sera werereceived from J. A. Hardin (Augusta, Ga.). The identifying initials of sera aKu-3and aKu-4 were HT and TT, respectively, while the origin of sera aKu-1 andaKu-2 could no longer be established. Ascites fluid containing MAbs 18-2 and42-26 was provided by W. S. Dynan. Purified immunoglobulin G2b (IgG2b) werefrom Pharmingen (San Diego, Calif.), and purified human DNA-PK was pur-chased from Promega (Madison, Wis.).
Extract preparation. The extract from Ca21-ionophore-activated Xenopuseggs was prepared as described by Schaal et al. (36), except that in the extractionbuffer HEPES was used instead of Tris (90 mM KCl, 30 mM HEPES [pH 7.9],2 mM EGTA, 10 mM b-glycerophosphate, 1 mM DTT) and only 1/3 volumeinstead of 1 volume of extraction buffer was added to the packed eggs beforecentrifugation. The protein concentration of the extract was about 16 mg/ml.
DNA repair reactions and analysis of repair products. DNA repair substrateswere prepared by digestion of pBluescript SK (pBSK; Stratagene) with a singleor two different restriction enzymes, followed by gel purification of the ;2.9-kblinear DNA. A typical in vitro reaction consisted of 8 ml of undiluted extract, 1ml of DNA (10 ng/ml), and 1 ml of experimental addition. Human serum wasadded in a volume of 1 ml after dilution in 0.1 M KCl–20 mM HEPES (pH7.9)–20% glycerol–0.2 mM EDTA–0.5 mM DTT–0.5 mM Phenylmethylsulfonylfluoride–50 mg of bovine serum albumin (BSA) per ml (DB/BSA buffer). Reac-tions were incubated at 15°C for 2 to 3 h and stopped by the addition of 90 ml of0.3 M sodium acetate, 10 mM EDTA, 0.5% sodium dodecyl sulfate (SDS), 50mM Tris (pH 7.6), 0.5 mg of proteinase K per ml, and 10 mg of carrier Esche-richia coli RNA and incubation at 37°C for 30 min. DNA was recovered byorganic extraction and ethanol precipitation and then electrophoresed in 1%agarose gels in 50 mM Tris (pH 7.8)–20 mM sodium acetate–2 mM EDTA (TAEbuffer) containing 1 mg of ethidium bromide per ml. Southern blot hybridizationwas performed according to standard procedures by using labeled pBSK se-quences as a probe. For the experiment shown in Fig. 3, lanes 14 to 22, DNA waselectrophoresed in 1% agarose in 50 mM Tris–50 mM boric acid (pH 8.3)–2 mMEDTA (TBE buffer) containing 1 mg of ethidium bromide per ml.
DNA binding assay for Ku. pBSK(1) was biotinylated at the HindIII site asdescribed earlier (23). After a second cut with PstI, the linear plasmid DNA wasgel purified. Then, 50 ng of DNA was bound to 5 ml of streptavidin-coatedparamagnetic particles (10 mg/ml; M-280 Dynabeads; Dynal) as described pre-viously (23) and incubated with 8 ml of egg extract at 15°C for 1 h. After theaddition of 10 ml of NTN (0.15 M NaCl, 50 mM Tris [pH 8], 0.1% Nonidet P-40),the beads were separated from the non-DNA binding fraction (the “superna-tant”) by magnetic separation. The beads were washed twice with 150 ml of NTN,and the proteins were eluted from the DNA beads by heating in SDS loadingbuffer (62.5 mM Tris-HCl [pH 6.8], 2% [wt/vol] SDS, 10% glycerol, 40 mM DTT,0.01% [wt/vol] bromophenol blue).
Preparation of antibodies against Xenopus DNA-PKcs. To generate antibodiesagainst Xenopus DNA-PK, the kinase domain of Xenopus DNA-PK (amino acidresidues 264 to 640; numbering as in reference 24) was expressed as a hexahis-tidine-tagged recombinant protein in E. coli BL21(DE3) by using the vectorpET-28a(1) (Novagen). The 46.9-kDa protein (his-DNPK) was purified onnickel-nitrilotriacetic acid-agarose under denaturing conditions and further pu-rified by preparative SDS–12% polyacrylamide gel electrophoresis for the pro-duction of polyclonal antibodies in rabbits (BioWorld, Dublin, Ohio). DNA-PK-specific antibodies (anti-xDNPKcs) were purified from the serum by affinitychromatography on CNBr-activated Sepharose 4B (Pharmacia) containing thecovalently bound recombinant antigen.
Western blot analysis. Proteins were electrophoresed on 5% (for assay ofDNA-PKcs) or 7.5% (for assay of Ku) discontinuous SDS-polyacrylamide gelsand electrotransferred to nitrocellulose filters in 25 mM Tris (pH 8.3), 192 mMglycine, 20% (vol/vol) methanol, and 0.02% SDS. Filters were probed with theprimary antibody in 0.5 M NaCl, 20 mM Tris (pH 7.5), 0.1% Tween 20, and 1%nonfat dry milk (Bio-Rad). Reactive proteins were detected with peroxidase-conjugated secondary antibody and enhanced chemiluminescence by using theSuperSignal Substrate (Pierce).
Immunoprecipitation. To label extract proteins with 32P, egg extract wasincubated in the presence of [g-32P]ATP, 6 mM MgCl2, and 50 ng of linear DNA.Labeled Ku proteins were immunoprecipitated by using protein A-Sepharose(Pharmacia) in a buffer containing 500 mM NaCl, 50 mM NaF, 20 mM sodiumphosphate (pH 8.0), and 0.2% Nonidet P-40 according to standard protocols.
Immunodepletion. A 10-ml portion bead volume of protein G-Sepharose (Sig-ma) was incubated with 2 ml of human serum or 7.5 to 15 mg of immunoglobulins(purified MAb) in DB/BSA buffer in a volume of 250 ml overnight at 6°C. Beadswere washed once in DB/BSA and twice in egg extraction buffer containing 0.5mM phenylmethylsulfonyl fluoride (250 ml each). After the washes, extra liquidwas removed from the beads, and 12 ml of egg extract was added. After end-over-end rotation at 6°C for 90 min, the samples were centrifuged and 9 ml ofdepleted extract (supernatant) was removed and used in NHEJ assays. For“double depletion”, the first supernatant was added to another 10 ml of IgG2b-
or N3H10-coated protein G-Sepharose, and the incubation was continued for 60min before the supernatant was used in NHEJ assays. To analyze the proteinsbound to protein G-Sepharose, beads were washed two times with 500 ml of NTNand resuspended in SDS-gel loading buffer.
RESULTS
Molecular and immunological characterization of XenopusDNA-PK. Before investigating whether DNA-PK was requiredduring NHEJ in the Xenopus egg extract, it was important todemonstrate the presence of DNA-PK in the extract. XenopusDNA-PK had not been characterized very well, and only apartial cDNA encoding a C-terminal segment of DNA-PKcshad been isolated (24). Western blot analysis showed thatantibodies directed against this recombinant C-terminal kinasedomain of Xenopus DNA-PKcs (anti-xDNPKcs) reacted with aprotein of very high molecular weight in the egg extract (Fig.1A, lanes 5 to 8). This same band was also detected with MAbsagainst human DNA-PKcs (clones 18-2 and 42-26 [6]), and itcomigrated with human DNA-PKcs, indicating that XenopusDNA-PKcs is similar in size to its human counterpart (Fig. 1B,
FIG. 1. Immunological characterization of DNA-PKcs and Ku in the Xeno-pus egg extract. (A) Various amounts (as indicated above the lanes) of recom-binant C-terminal domain of Xenopus DNA-PKcs (lanes 1 to 4, his-DNPK) oregg extract (lanes 5 to 8) were analyzed by Western blotting. Filters were probedwith polyclonal rabbit antibodies against his-DNPK. (B) Various amounts ofpurified human Ku (lanes 1 to 3), purified human DNA-PK (lane 7), and eggextract (lanes 4 to 6, 8 and 9) were analyzed by Western blotting. Filters wereprobed with anti-Ku70 MAb N3H10 (lanes 1 to 9, upper panel) or anti-DNA-PKcs MAb 42-26 (lanes 7 to 9, lower panel). The migration of prestained markerproteins and their approximate molecular weight in kilodaltons is indicated tothe left. The signals corresponding to Ku70 and DNA-PKcs are labeled to theright.
2586 LABHART MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 17
Oct
ober
202
1 by
114
.69.
161.
189.

lanes 7 to 9, lower panel). The band reactive with MAb 42-26could also be immunoprecipitated from Xenopus extract withanti-xDNPKcs (not shown), thus confirming that this band isthe Xenopus homologue of DNA-PKcs.
An MAb against human Ku70 (clone N3H10 [22]) reactedvery strongly with a Xenopus protein comigrating with purifiedhuman Ku (Fig. 1B, lanes 1 to 6), indicating that this bandrepresents the Xenopus homologue of Ku70. This interpreta-tion was supported by the finding that the same N3H10-reac-tive protein was immunoprecipitated from egg extracts withanti-Ku autoimmune sera but not with normal human serum(see Fig. 4, bottom, lanes 1 to 3). When anti-Ku autoantibodieswere used in immunoprecipitation experiments with 32P-la-beled extract proteins, two proteins of the predicted sizes forKu70 and Ku80 were immunoprecipitated, suggesting that Xe-nopus Ku is also a heterodimer (Fig. 2A). Finally, the N3H10-reactive protein binds to streptavidin beads containing biotin-ylated (linear) DNA but not to control beads without DNA, afinding consistent with the DNA end-binding properties of Kuprotein (Fig. 2C, lanes 1 and 2).
DNA-PK is very abundant in the egg extract. Two indepen-dent approaches were taken to estimate the amount ofDNA-PK in the egg extract. By using the recombinant kinasedomain of Xenopus DNA-PKcs as a standard in a Western blot
probed with anti-xDNPKcs, I found that the 470-kDa DNA-PKcs present in 10 ml of extract gave a signal similar to that of40 ng of the recombinant 47-kDa protein (Fig. 1A, comparelanes 1 to 4 to lanes 5 to 8), indicating that 1 ml of extractcontains 40 ng of DNA-PKcs. In the second approach, prepa-rations of purified human Ku and DNA-PK were used asstandards in Western blots probed with MAbs against thehuman proteins. This approach is based on the assumption thatthese MAbs bind to the Xenopus proteins with equal affinity asto their human counterparts, an assumption that, if incorrect,more likely would lead to an underestimate of the amount ofXenopus DNA-PK in the extract. These experiments showedthat the Xenopus Ku70 signal obtained with 1 ml of egg extractwas about as intense as the signal obtained with 30 to 40 ng ofpurified human Ku heterodimer (Fig. 1B, lanes 1 to 6) or with150 ng of purified human DNA-PK holoenzyme (lanes 7 to 9;upper panel). Furthermore, 1 ml of egg extract gave a DNA-PKcs signal of about the same intensity as 150 ng of humanDNA-PK (Fig. 1B, lanes 7 to 9; lower panel). These data alsoindicated that DNA-PKcs and Ku were present in aboutequimolar amounts in both the preparation of purified, activehuman DNA-PK and the Xenopus egg extract. Based on thesetwo estimates and by using a dilution factor of the proteinconcentration from the egg to the extract of 1.33, I concludethat one Xenopus egg (;1 ml) contains 70 to 200 ng ofDNA-PK or 0.7 3 1011 to 2 3 1011 molecules.
Assay for NHEJ in the Xenopus egg extract. The standardNHEJ substrate used in the present study was a linear plasmidDNA molecule bearing nonmatching XhoI and PstI ends (Fig.3, lane 1; see diagram in Fig. 5). Upon incubation in theextract, this substrate underwent both intramolecular NHEJ togive rise to monomeric covalently closed circles (CCC), as wellas intermolecular NHEJ to form multimeric forms (Fig. 3, lane2). Whereas the monomeric circular products can only beformed by joining an XhoI end to a PstI end in a head-to-tailconfiguration (H/T), the multimers generated from the XhoI/PstI substrate can be formed both by joining of mismatchedends (H/T), as well as by ligation of pairs of cohesive ends ashead-to-head (H/H) or tail-to-tail joints (T/T) (see Fig. 5).Sequence analysis of cloned circularized products showed thatthe majority of XhoI and PstI ends were joined without nucle-otide loss, i.e., by end-to-end joining of the 59- and 39-PSS andfilling in of the 8-nucleotide (nt) gap (25), which is in agree-ment with previous studies (32, 39). Likewise, digestion of therepair products with XhoI and PstI indicated that H/H and T/Tjoints were generated in an error-free manner (data notshown).
Autoantibodies against Ku inhibit NHEJ. To determinewhether DNA-PK was required for NHEJ in the egg extract, Iattempted to inhibit the reaction by the addition of antibodiesagainst DNA-PK components. Whereas even high amounts ofvarious antibodies against DNA-PKcs did not affect the NHEJreaction (data not shown; see Discussion), inhibitory effectswere readily detected with antibodies against the Ku compo-nent of DNA-PK. The present study therefore focuses entirelyon the role of Ku during NHEJ in the Xenopus egg extract.
Four different anti-Ku autoimmune sera were used. Asshown above in Fig. 2A, all four sera immunoprecipitated theKu heterodimer. However, to precipitate similar amounts ofKu, two to four times more aKu-1 and aKu-3 serum thanaKu-2 and aKu-4 serum was necessary, indicating that aKu-2and aKu-4 had a higher titer. To further characterize thesefour sera, different dilutions were used to probe Western blotsof Xenopus egg extract proteins. Sera aKu-1 and aKu-3 ap-peared to be very similar; they both did not react with thedenatured Ku proteins but at a dilution of 1:1,000 recognized
FIG. 2. Characterization of Xenopus Ku with human anti-Ku autoantibodies.(A) Immunoprecipitation of 32P-labeled Xenopus Ku from egg extracts. Threemicroliters of autoimmune sera aKu-1 (lane 2) and aKu-3 (lane 4), 1.5 ml ofaKu-2 (lane 3), or 2 ml of aKu-4 (lane 5) and NHS (lane 1) was used. Note thattwo proteins of the expected sizes for Xenopus Ku70 and Ku80 are precipitatedby the autoimmune sera but not by the NHS. (B) Western blot analysis of eggextract proteins. Filters were probed with the sera indicated above the lanes. Serawere used at the following dilutions: NHS, aKu-1, and aKu-3 at 1:1,000; aKu-4at 1:2,000; and aKu-2 at 1:4,000. The band recognized by aKu-2 and aKu-4 andcorresponding to Xenopus Ku70 is indicated. Note that aKu-1 and aKu-3 rec-ognize a protein of about 200 kDa. (C) Western blot analysis of unbound(supernatant, upper panel) and DNA-bound Ku70 (DNA beads, lower panel)after incubation of immobilized DNA in the egg extract. Lane 1 shows a controlwith paramagnetic particles without DNA. For lanes 3 and 4, the extract waspreincubated with 1 ml of autoimmune serum aKu-3 or 0.25 ml of serum aKu-4,respectively, amounts that strongly inhibit NHEJ (see Fig. 3, lanes 5 and 6), whilethe control reactions shown in lanes 1 and 2 contain NHS.
VOL. 19, 1999 Ku-DEPENDENT DNA END JOINING IN VITRO 2587
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 17
Oct
ober
202
1 by
114
.69.
161.
189.

an unidentified protein of about 200 kDa (Fig. 2B, lanes 2 and4). This band was detected with dilutions of these sera of up to1:4,000 (not shown). Sera aKu-2 and aKu-4, on the otherhand, both recognized a band comigrating with Ku70 (lanes 3and 5). While this band was the only reactive band when serumaKu-4 was used at a 1:2,000 dilution, aKu-2 also reacted withthree major additional proteins, up to a dilution of 1:8,000.These Western blot analyses thus indicate that aKu-1 andaKu-3 on the one hand, and aKu-2 and aKu-4 on the otherhand, belong to two different subclasses of anti-Ku autoim-mune sera (1, 34, 35).
When directly added to NHEJ reactions, all four anti-Kuautoimmune sera were found to inhibit end-joining reactionswith the XhoI/PstI substrate (Fig. 3, lanes 3 to 6). Consistentwith their higher titer, aKu-2 and aKu-4 strongly inhibitedNHEJ even when only 0.25 ml of serum was added to thereaction (lanes 4 and 6), while 1 ml of aKu-1 and aKu-3 had tobe added to see inhibition (lanes 3 and 5). The addition of 1 mlof normal human serum (NHS) did not inhibit the reaction(lane 2). To test whether the inhibition was due to anti-Kuantibodies or to antibodies with other specificities, which areoften present in autoimmune sera (33), I preincubated theautoantibodies with purified human Ku protein. Figure 3, lanes8 to 13, shows that the inhibition of NHEJ by sera aKu-3 andaKu-4 was indeed neutralized by this preincubation (lanes 11and 13), while preincubation of NHS with Ku protein had noeffect (lane 9). This result thus demonstrates that anti-Ku an-tibodies specifically inhibit NHEJ by binding to Ku. NHEJreactions with immobilized DNA showed that inhibitoryamounts of aKu-3 and aKu-4 serum did not significantly affectthe amount of Ku binding to DNA (Fig. 2C, lanes 3 and 4).Thus, inhibition of NHEJ by anti-Ku antibodies appears not tobe due to interference with the DNA-binding site of Ku.
Linear repair substrates bearing blunt ends (SmaI) or cohe-sive 4-nt 59-PSS (BamHI) were circularized and multimerized
in the extract with similar efficiency as the XhoI/PstI substrate(Fig. 3, lanes 15, 16, 19, and 20). Additional experimentsshowed that these NHEJ products could be completely di-gested with SmaI or BamHI, respectively, indicating that thejoining was error-free (data not shown). NHEJ of these sub-strates was also inhibited by the aKu-4 serum (lanes 17 and21), and the inhibition was relieved by preincubation of theaKu-4 serum with Ku protein (lanes 18 and 22). In the exper-iment with the SmaI and BamHI substrates, the NHEJ prod-ucts were analyzed under electrophoretic conditions that sep-arated the “open” (nicked or gapped) circles from the inputlinear substrates (Fig. 3, lanes 14 to 22). The results showedthat the formation of these incompletely repaired intermedi-ates was also inhibited by the aKu-4 serum, indicating that theinhibition by anti-Ku antibodies occurs at an early step of theNHEJ reaction.
NHEJ in Ku-depleted extracts. Since autoantibodies werefound to immunoprecipitate Ku from the extract (see Fig. 2A),I also used these antibodies to prepare Ku-depleted extracts.Western blot analysis showed that Ku protein could effectivelybe removed from the extract with protein G-Sepharose con-taining aKu-4 immunoglobulins (Fig. 4, bottom panel, lane 3).Ku remained in the extract when protein G-Sepharose con-taining NHS immunoglobulins were used (lanes 1 and 2). Thesame extracts were then tested for NHEJ activity (Fig. 4, toppanel). Unlike NHEJ reactions with crude, untreated extract(as in Fig. 3), mock-depleted extracts reproducibly showed veryefficient circularization but less multimerization (Fig. 4, lanes 1and 2). This phenomenon was not further examined, but it islikely a consequence of the depletion protocol. In Ku-depletedextracts the circularization of the XhoI/PstI substrate was re-duced to 15% of the level observed in control extracts treatedwith NHS (lane 3 [band labeled CCC]). Interestingly, however,there was an increase in linear dimer products, and the low
FIG. 3. Inhibition of NHEJ by anti-Ku autoantibodies. Lanes 1 to 7: Southern blot analysis of NHEJ reactions in the presence of 1 ml of NHS (lane 2), 1 ml of aKu-1serum (lane 3), 0.25 ml of aKu-2 serum (lane 4), 1 ml of aKu-3 serum (lane 5), 0.25 ml of aKu-4 serum (lane 6), or without addition (lane 7). Lane 1 shows 10 ng ofinput XhoI/PstI substrate. The bands corresponding to the substrate (S) and the main NHEJ products CCC and linear dimer (Lin2) are indicated. Lanes 8 to 13 aresimilar to the reactions shown in lanes 2, 5, and 6, except that the sera were preincubated with 210 ng of purified Ku (lanes 9, 11, and 13) or with the same volume(3 ml) of Ku buffer (lanes 8, 10, and 12). Note that preincubation of anti-Ku antibodies with Ku relieves the inhibition of NHEJ. Lanes 15 to 22 are similar to thereactions shown in lanes 8 to 11, except that the substrate was pBSK linearized with SmaI (lanes 16 to 18; input substrate is shown in lane 15) or pBSK linearized withBamHI (lanes 20 to 22; input substrate is shown in lane 19). Lane 14, circular pBSK. The DNA analyzed in lanes 14 to 22 was electrophoresed in TBE buffer to separatethe linear substrate (S) from the open circles (OC). The slightly different migration of purified linear substrate DNA (S; lanes 1, 15, and 19) and unreacted substratein NHEJ reactions (other lanes) is due to the presence in the latter samples of a large amount of RNA and possibly other impurities originating from the egg extract.
2588 LABHART MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 17
Oct
ober
202
1 by
114
.69.
161.
189.
level of higher multimers formed was barely affected (see be-low).
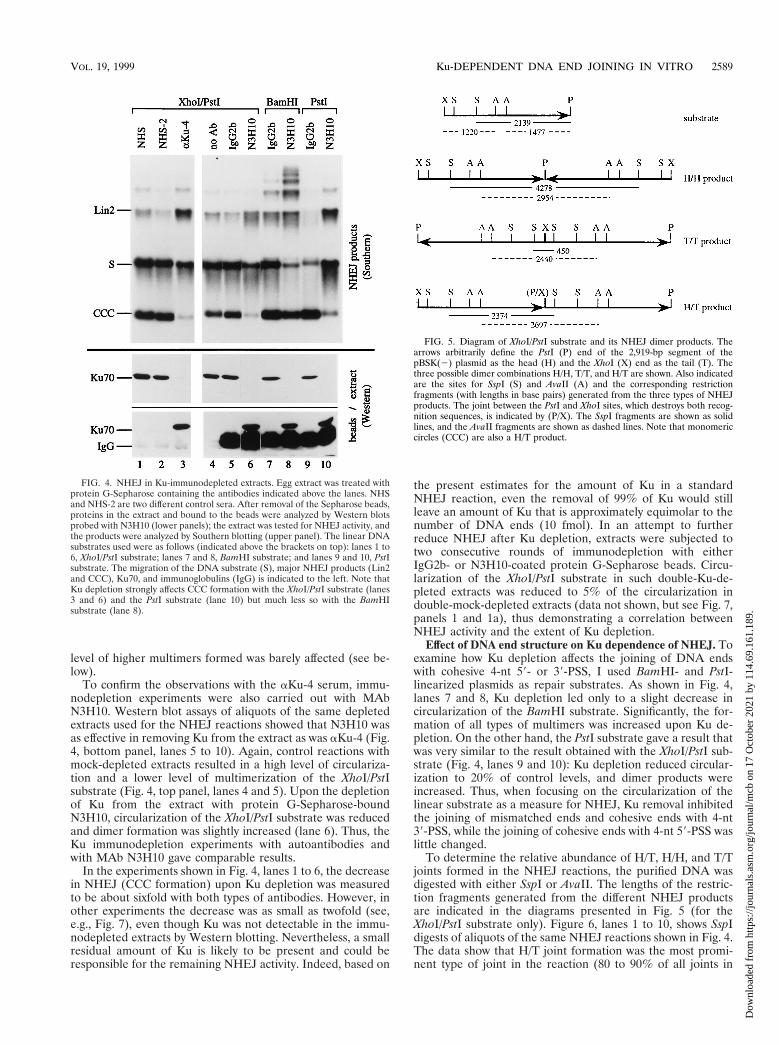
To confirm the observations with the aKu-4 serum, immu-nodepletion experiments were also carried out with MAbN3H10. Western blot assays of aliquots of the same depletedextracts used for the NHEJ reactions showed that N3H10 wasas effective in removing Ku from the extract as was aKu-4 (Fig.4, bottom panel, lanes 5 to 10). Again, control reactions withmock-depleted extracts resulted in a high level of circulariza-tion and a lower level of multimerization of the XhoI/PstIsubstrate (Fig. 4, top panel, lanes 4 and 5). Upon the depletionof Ku from the extract with protein G-Sepharose-boundN3H10, circularization of the XhoI/PstI substrate was reducedand dimer formation was slightly increased (lane 6). Thus, theKu immunodepletion experiments with autoantibodies andwith MAb N3H10 gave comparable results.
In the experiments shown in Fig. 4, lanes 1 to 6, the decreasein NHEJ (CCC formation) upon Ku depletion was measuredto be about sixfold with both types of antibodies. However, inother experiments the decrease was as small as twofold (see,e.g., Fig. 7), even though Ku was not detectable in the immu-nodepleted extracts by Western blotting. Nevertheless, a smallresidual amount of Ku is likely to be present and could beresponsible for the remaining NHEJ activity. Indeed, based on
the present estimates for the amount of Ku in a standardNHEJ reaction, even the removal of 99% of Ku would stillleave an amount of Ku that is approximately equimolar to thenumber of DNA ends (10 fmol). In an attempt to furtherreduce NHEJ after Ku depletion, extracts were subjected totwo consecutive rounds of immunodepletion with eitherIgG2b- or N3H10-coated protein G-Sepharose beads. Circu-larization of the XhoI/PstI substrate in such double-Ku-de-pleted extracts was reduced to 5% of the circularization indouble-mock-depleted extracts (data not shown, but see Fig. 7,panels 1 and 1a), thus demonstrating a correlation betweenNHEJ activity and the extent of Ku depletion.
Effect of DNA end structure on Ku dependence of NHEJ. Toexamine how Ku depletion affects the joining of DNA endswith cohesive 4-nt 59- or 39-PSS, I used BamHI- and PstI-linearized plasmids as repair substrates. As shown in Fig. 4,lanes 7 and 8, Ku depletion led only to a slight decrease incircularization of the BamHI substrate. Significantly, the for-mation of all types of multimers was increased upon Ku de-pletion. On the other hand, the PstI substrate gave a result thatwas very similar to the result obtained with the XhoI/PstI sub-strate (Fig. 4, lanes 9 and 10): Ku depletion reduced circular-ization to 20% of control levels, and dimer products wereincreased. Thus, when focusing on the circularization of thelinear substrate as a measure for NHEJ, Ku removal inhibitedthe joining of mismatched ends and cohesive ends with 4-nt39-PSS, while the joining of cohesive ends with 4-nt 59-PSS waslittle changed.
To determine the relative abundance of H/T, H/H, and T/Tjoints formed in the NHEJ reactions, the purified DNA wasdigested with either SspI or AvaII. The lengths of the restric-tion fragments generated from the different NHEJ productsare indicated in the diagrams presented in Fig. 5 (for theXhoI/PstI substrate only). Figure 6, lanes 1 to 10, shows SspIdigests of aliquots of the same NHEJ reactions shown in Fig. 4.The data show that H/T joint formation was the most promi-nent type of joint in the reaction (80 to 90% of all joints in
FIG. 4. NHEJ in Ku-immunodepleted extracts. Egg extract was treated withprotein G-Sepharose containing the antibodies indicated above the lanes. NHSand NHS-2 are two different control sera. After removal of the Sepharose beads,proteins in the extract and bound to the beads were analyzed by Western blotsprobed with N3H10 (lower panels); the extract was tested for NHEJ activity, andthe products were analyzed by Southern blotting (upper panel). The linear DNAsubstrates used were as follows (indicated above the brackets on top): lanes 1 to6, XhoI/PstI substrate; lanes 7 and 8, BamHI substrate; and lanes 9 and 10, PstIsubstrate. The migration of the DNA substrate (S), major NHEJ products (Lin2and CCC), Ku70, and immunoglobulins (IgG) is indicated to the left. Note thatKu depletion strongly affects CCC formation with the XhoI/PstI substrate (lanes3 and 6) and the PstI substrate (lane 10) but much less so with the BamHIsubstrate (lane 8).
FIG. 5. Diagram of XhoI/PstI substrate and its NHEJ dimer products. Thearrows arbitrarily define the PstI (P) end of the 2,919-bp segment of thepBSK(2) plasmid as the head (H) and the XhoI (X) end as the tail (T). Thethree possible dimer combinations H/H, T/T, and H/T are shown. Also indicatedare the sites for SspI (S) and AvaII (A) and the corresponding restrictionfragments (with lengths in base pairs) generated from the three types of NHEJproducts. The joint between the PstI and XhoI sites, which destroys both recog-nition sequences, is indicated by (P/X). The SspI fragments are shown as solidlines, and the AvaII fragments are shown as dashed lines. Note that monomericcircles (CCC) are also a H/T product.
VOL. 19, 1999 Ku-DEPENDENT DNA END JOINING IN VITRO 2589
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 17
Oct
ober
202
1 by
114
.69.
161.
189.

control reactions) and that circularization accounted for most,if not all, of the H/T joints. Accordingly, Ku-dependentchanges of the H/T bands in Fig. 6 paralleled the changesobserved with the undigested CCC products, i.e., H/T joiningwas decreased upon Ku depletion in reactions with the XhoI/PstI substrate (lanes 1 to 6, 12, and 13) and the PstI substrate(lanes 9 and 10) but was essentially unchanged in reactionswith the BamHI substrate (lanes 7, 8, 15, and 16).
On the other hand, H/H and T/T joint formations weremuch less frequent events (10 to 20% of all joints). Interest-ingly, their formation showed a Ku and substrate dependencethat was similar, but clearly not identical, to the formation ofthe H/T joints described above. With the XhoI/PstI substrate,Ku depletion led to a decrease in the PstI-PstI joints (H/H),while in the same reaction the XhoI-XhoI joints (T/T) wereincreased (Fig. 6, lanes 12 and 13; for H/H, see also lanes 1 to
6). In reactions with the BamHI substrate, both H/H and T/TBamHI-BamHI joints were increased upon Ku depletion (Fig.6, lanes 7 and 8 and lanes 15 and 16). However, in reactionswith the PstI substrate, the formation of PstI-PstI joints in H/Hconfiguration was unchanged (Fig. 6, lanes 9 and 10), whiletheir formation was decreased in T/T configuration (see Fig. 7,panel 4), indicating that not all types of joints involving cohe-sive 4-nt 39-PSS show the same Ku dependence. These resultsalso provide an explanation for the increased dimer formationseen with the XhoI/PstI and PstI substrates (Fig. 4): mathemat-ical models (25; not shown) predict that decreased probabilityfor H/H joining at one end of a linear repair substrate andincreased or unchanged probability for T/T joining at the otherend would indeed lead to increased probability for dimer for-mation.
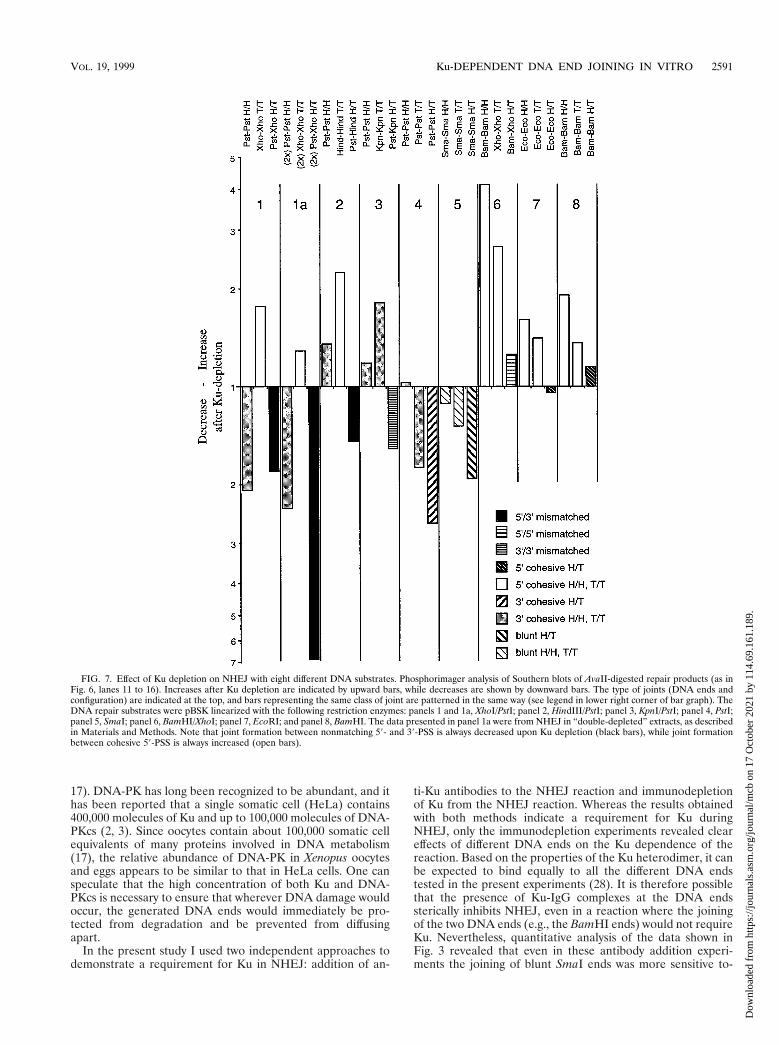
To further test the validity and generality of these findings,an independent experiment with the XhoI/PstI, BamHI, andPstI substrates, as well as five additional NHEJ substrates, wascarried out and its quantitative analysis is shown in Fig. 7. TheKu-dependent changes in H/T, H/H, and T/T joint formationwere determined by AvaII digestion of the repair products andphosphorimager analysis of the Southern blots. The results canbe summarized as follows: a decrease in NHEJ after Ku de-pletion was always observed with a nonmatching pair having a59-PSS and a 39-PSS (XhoI-PstI and HindIII-PstI; Fig. 7, blackbars) or two 39-PSS (KpnI-PstI) and with pairs of blunt ends(SmaI-SmaI) or cohesive ends with 39-PSS (PstI-PstI) in H/Tconfiguration (third bar in panels 1 to 5). On the other hand,an increase in NHEJ after Ku depletion was consistently seenwith pairs of cohesive ends bearing 59-PSS in H/H or T/Tconfiguration (BamHI-BamHI, EcoRI-EcoRI, XhoI-XhoI, andHindIII-HindIII; open bars), while Ku depletion had little ef-fect on joint formation between pairs of ends with 59-PSS inH/T configuration (BamHI-BamHI, EcoRI-EcoRI, andBamHI-XhoI; third bar in panels 6 to 8; it should be noted thatthe nonmatching XhoI and BamHI ends can anneal by forminga 2-bp overlap). Mixed results were again obtained with pairsof cohesive ends with 39-PSS in H/H or T/T configuration (darkgray bars in panels 1 to 4): while the formation of PstI-PstIjoints was reproducibly decreased in reactions with the XhoI/PstI substrate (see also Fig. 6), it showed minor increases withother substrates, and the formation of the single KpnI-KpnIjoint tested in these experiments was increased after Ku de-pletion. From the combined data it can be concluded that Kudependence of NHEJ is primarily a function of the structure ofthe two DNA ends, but that NHEJ between two ends in H/Tconfiguration is generally more Ku dependent than NHEJbetween the same two ends in H/H or T/T configuration, sug-gesting that the presence of extended inverted repeats flankingthe joining site can obviate the need for Ku.
DISCUSSION
NHEJ in an extract from activated Xenopus eggs had beencharacterized in detail with respect to the repair productsformed from defined exogenous linear DNA substrates (31,32). However, little is known about the protein factors in theextract that are involved in this process. Here I identify theDNA-PK component Ku as a protein factor that is required forthe joining of various types of DNA ends in this system.
Both DNA-PKcs and Ku are very abundant in the egg ex-tract. They are present in about equimolar amounts, and myresults indicate that one egg contains ca. 70 to 200 ng or 0.7 31011 to 2 3 1011 molecules of the DNA-PK complex. This is inthe same range as the chromatin components histones (140ng/oocyte) or nucleoplasmin (250 ng/oocyte) (see reference
FIG. 6. Restriction analysis of NHEJ products. Lanes 1 to 10 show SspIdigests of aliquots of the same NHEJ products as shown in Fig. 4. The sizes ofthe restriction fragments (for the XhoI/PstI substrate only) and the type of jointthat they represent are indicated (see Fig. 5). The BamHI and PstI substrateswere made from pBSK(1), giving rise to a larger fragment for H/T joints (2,828bp) than the XhoI/PstI substrate [made from pBSK(2)]. To make the changes inthe signals between the control lanes (IgG2b) and minus-Ku lanes (N3H10)more visible, the exposure shown for lanes 7 to 10 is twice as short as that shownfor lanes 4 to 6. Lanes 11 to 16, AvaII digests of independent NHEJ reactions inmock- and Ku-depleted extracts with the XhoI/PstI (lanes 12 and 13) or theBamHI substrate (lanes 15 and 16). Lane 11, AvaII digest of input substrate (S);lane 14, AvaII digest of XhoI/PstI substrate treated with T4 DNA ligase. TheAvaII fragments generated from the reactions with the BamHI substrate (lanes15 and 16) are slightly different from the sizes indicated to the left for theXhoI/PstI substrate. (The sizes are as follows: H/H, 2,930 bp; H/T, 2,736 bp; T/T,2,542 bp; and S, 1,465 and 1,271 bp.) Note that with the XhoI/PstI substrate theH/H band (PstI-PstI joints) decreases, whereas the T/T band (XhoI-XhoI joints)increases upon Ku depletion (lanes 12 and 13).
2590 LABHART MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 17
Oct
ober
202
1 by
114
.69.
161.
189.
17). DNA-PK has long been recognized to be abundant, and ithas been reported that a single somatic cell (HeLa) contains400,000 molecules of Ku and up to 100,000 molecules of DNA-PKcs (2, 3). Since oocytes contain about 100,000 somatic cellequivalents of many proteins involved in DNA metabolism(17), the relative abundance of DNA-PK in Xenopus oocytesand eggs appears to be similar to that in HeLa cells. One canspeculate that the high concentration of both Ku and DNA-PKcs is necessary to ensure that wherever DNA damage wouldoccur, the generated DNA ends would immediately be pro-tected from degradation and be prevented from diffusingapart.
In the present study I used two independent approaches todemonstrate a requirement for Ku in NHEJ: addition of an-
ti-Ku antibodies to the NHEJ reaction and immunodepletionof Ku from the NHEJ reaction. Whereas the results obtainedwith both methods indicate a requirement for Ku duringNHEJ, only the immunodepletion experiments revealed cleareffects of different DNA ends on the Ku dependence of thereaction. Based on the properties of the Ku heterodimer, it canbe expected to bind equally to all the different DNA endstested in the present experiments (28). It is therefore possiblethat the presence of Ku-IgG complexes at the DNA endssterically inhibits NHEJ, even in a reaction where the joiningof the two DNA ends (e.g., the BamHI ends) would not requireKu. Nevertheless, quantitative analysis of the data shown inFig. 3 revealed that even in these antibody addition experi-ments the joining of blunt SmaI ends was more sensitive to-
FIG. 7. Effect of Ku depletion on NHEJ with eight different DNA substrates. Phosphorimager analysis of Southern blots of AvaII-digested repair products (as inFig. 6, lanes 11 to 16). Increases after Ku depletion are indicated by upward bars, while decreases are shown by downward bars. The type of joints (DNA ends andconfiguration) are indicated at the top, and bars representing the same class of joint are patterned in the same way (see legend in lower right corner of bar graph). TheDNA repair substrates were pBSK linearized with the following restriction enzymes: panels 1 and 1a, XhoI/PstI; panel 2, HindIII/PstI; panel 3, KpnI/PstI; panel 4, PstI;panel 5, SmaI; panel 6, BamHI/XhoI; panel 7, EcoRI; and panel 8, BamHI. The data presented in panel 1a were from NHEJ in “double-depleted” extracts, as describedin Materials and Methods. Note that joint formation between nonmatching 59- and 39-PSS is always decreased upon Ku depletion (black bars), while joint formationbetween cohesive 59-PSS is always increased (open bars).
VOL. 19, 1999 Ku-DEPENDENT DNA END JOINING IN VITRO 2591
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 17
Oct
ober
202
1 by
114
.69.
161.
189.

ward anti-Ku autoantibodies (both circularization and dimer-ization were reduced to 10%) than the joining of the cohesiveBamHI ends (circularization and dimerization reduced to35%), and that with the XhoI/PstI substrate circularization wasconsistently at least twofold more inhibited by anti-Ku anti-bodies than was dimer formation, probably reflecting the pres-ence of XhoI-XhoI (T/T) products among the dimers.
These differences in the Ku dependence between NHEJreactions with different types of linear DNA substrates weremuch more evident in immunodepletion experiments, wherethe inhibition of NHEJ cannot be due to steric hindrance butmust reflect the lack of an essential component of the NHEJmachinery. While more work is needed to understand why thejoining of certain pairs of DNA ends requires Ku and others donot, it is intriguing that Ku is required for NHEJ between a59-PSS and a 39-PSS, as well as for NHEJ between two bluntends. In both of these cases, the alignment of the two DNAends cannot be achieved by base pairing, and Ku might benecessary to hold the two ends together. On the other hand,the joining of two ends bearing the same type of PSS is clearlyKu-dependent for 39-PSS but shows little Ku dependence for59-PSS. Thus, at least in the case of 59-PSS, the ability of twoends to align by base pairing seems to obviate the need for Ku.
The present results suggest the existence of an alternativeend-joining pathway, which becomes more dominant in theabsence of Ku. Thus “simple ligation” of ends with 4-nt 59-PSSin H/H or T/T configuration is stimulated upon Ku removal,leading to increased multimer formation with the BamHI sub-strate and to increased dimer formation with the XhoI/PstIsubstrate. And even for DNA substrates with other end struc-tures, the formation of the H/H and T/T products is less Kudependent than the corresponding H/T products (Fig. 7). Thepreference of this Ku-independent mechanism for H/H andT/T is reminiscent of an NHEJ activity described by Derby-shire et al. (10). This activity in nuclear extracts preferentiallyforms H/H and T/T joints after 39-exonucleolytic modificationof the DNA ends. However, BamHI digestion of the productsformed in Ku-depleted Xenopus egg extracts indicates that themajority of ends are still joined with high precision (data notshown). The present results thus suggest a mechanism duringwhich two ends in H/H or T/T configuration can anneal bymelting of the ends and the formation of a cruciform-likestructure between the extended homologous single strands,which then would promote the ligation of the two strandswithout nucleotide loss. Such a hypothetical mechanism againwould be consistent with the notion that in the egg extract Kuas an alignment factor is less important for the joining of DNAends that can interact by base pairing.
Genetic approaches have shown that in mammalian cells thecatalytic subunit of DNA-PK is required in addition to the Kuheterodimer for the repair of double-stranded DNA breaks (4,21). Furthermore, coding joint formation during V(D)J recom-bination was found to be inhibited by anti-DNA-PKcs antibod-ies (MAb 18-2) in a cell-free system (40). On the other hand,signal joint formation does not require DNA-PKcs (4, 27), andno DNA-PKcs homologue has been identified in yeast, sug-gesting that certain types of DNA end-joining reactions do notrequire DNA-PKcs. Attempts to inhibit NHEJ in the Xenopusegg extract with antibodies against DNA-PKcs, including MAb18-2 and the X. laevis-specific anti-xDNPKcs (used in Fig. 1A),were unsuccessful. While there are several possible explana-tions for these negative results, one could speculate that theerror-free NHEJ in the Xenopus egg extract is mechanisticallyrelated to signal joint formation and does not require DNA-PKcs. However, it has been reported that NHEJ in the Xeno-pus egg extract is sensitive toward wortmannin (15, 16), an
inhibitor of DNA-PK and phosphatidylinositol kinase-relatedkinases (19). When increasing concentrations of wortmanninhave been tested, there was a good correlation between theinhibition of NHEJ and the inhibition of DNA-PK activity,suggesting that the wortmannin-sensitive kinase that is re-quired at an early step during NHEJ is indeed DNA-PK. Icould confirm these findings by using the present assay systemfor NHEJ (25). However, my results show that, unlike Kudepletion, wortmannin inhibits NHEJ between pairs of cohe-sive 4-nt 59-PSS and 39-PSS to a similar degree. These obser-vations thus suggest that the kinase activity of DNA-PK andthe Ku heterodimer are required at different steps duringNHEJ.
Addition of purified human Ku to Ku-depleted Xenopus eggextracts did not restore NHEJ activity but instead further re-duced the residual NHEJ activity (25). Additional data showthat up to 50% of DNA-PKcs in the extract coimmunoprecipi-tates with Ku during the present Ku depletion protocol (25).However, purified DNA-PK was also unable to rescue NHEJin Ku-depleted extracts. These findings could indicate that Kufunction is species specific or that additional factors requiredfor NHEJ are bound to Ku or DNA-PKcs and have beenremoved from the extract during immunodepletion. To ad-dress these questions, to learn more about the mechanism andthe biochemistry of the NHEJ reaction, and to ultimately iden-tify all of the protein factors required for NHEJ in this system,it will be necessary to fractionate the egg extract and to recon-stitute NHEJ from purified components.
ACKNOWLEDGMENTS
This work was supported by grant MCB-9630773 from the NationalScience Foundation.
I thank W. S. Dynan, S. Yoo, and J. A. Hardin (Augusta, Ga.) forgenerously providing me with antibody and protein reagents.
REFERENCES
1. Abu-Elheiga, L., and M. Yaneva. 1992. Antigenic determinants of the 70-kDa subunit of the Ku autoantigen. Clin. Immunol. Immunopathol. 64:145–152.
2. Anderson, C. W., and T. H. Carter. 1996. The DNA-activated protein kinase:DNA-PK. Curr. Top. Microbiol. Immunol. 217:91–111.
3. Anderson, C. W., and S. P. Lees-Miller. 1992. The nuclear serine/threonineprotein kinase DNA-PK. Crit. Rev. Eukaryotic Gene Expr. 2:283–314.
4. Blunt, T., N. J. Finnie, G. E. Taccioli, G. C. M. Smith, J. Demengeot, T. M.Gottlieb, R. Mizuta, A. J. Varghese, F. W. Alt, P. A. Jeggo, and S. P. Jackson.1995. Defective DNA-dependent protein kinase activity is linked to V(D)Jrecombination and DNA repair defects associated with the murine scidmutation. Cell 80:813–823.
5. Boulton, S. J., and S. P. Jackson. 1996. Identification of a Saccharomycescerevisiae Ku80 homologue: roles in DNA double strand break rejoining andin telomeric maintenance. Nucleic Acids Res. 24:4639–4648.
6. Carter, T., I. Vancurova, I. Sun, W. Lou, and S. DeLeon. 1990. A DNA-activated protein kinase from HeLa cell nuclei. Mol. Cell. Biol. 10:6460–6471.
7. Chu, G. 1997. Double strand break repair. J. Biol. Chem. 272:24097–24100.8. Cortes, P., F. Weis-Garcia, Z. Misulovin, A. Nussenzweig, J.-S. Lai, G. Li,
M. C. Nussenzweig, and D. Baltimore. 1996. In vitro V(D)J recombination:signal joint formation. Proc. Natl. Acad. Sci. USA 93:14008–14013.
9. Daza, P., S. Reichenberger, B. Gottlich, M. Hagmann, E. Feldmann, and P.Pfeiffer. 1996. Mechanisms of nonhomologous DNA end joining in frogs,mice and men. Biol. Chem. Hoppe-Seyler 377:775–786.
10. Derbyshire, M. K., L. H. Epstein, C. S. H. Young, P. L. Munz, and R. Fishel.1994. Nonhomologous recombination in human cells. Mol. Cell. Biol. 14:156–169.
11. Dynan, W. S., and S. Yoo. 1998. Interaction of Ku protein and DNA-dependent protein kinase catalytic subunit with nucleic acids. Nucleic AcidsRes. 26:1551–1559.
12. Goedecke, W., W. Vielmetter, and P. Pfeiffer. 1992. Activation of a system forthe joining of nonhomologous DNA ends during Xenopus egg maturation.Mol. Cell. Biol. 12:811–816.
13. Gottlieb, T. M., and S. P. Jackson. 1993. The DNA-dependent proteinkinase: requirement for DNA ends and association with Ku antigen. Cell72:131–142.
2592 LABHART MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 17
Oct
ober
202
1 by
114
.69.
161.
189.

14. Gravel, S., M. Larrivee, P. Labrecque, and R. J. Wellinger. 1998. Yeast Kuas a regulator of chromosomal DNA end structure. Science 280:741–744.
15. Gu, X.-Y., R. A. O. Bennett, and L. F. Povirk. 1996. End-joining of freeradical-mediated DNA double-strand breaks in vitro is blocked by the kinaseinhibitor wortmannin at a step preceding removal of damaged 39 termini.J. Biol. Chem. 271:19660–19663.
16. Gu, X.-Y., M. A. Weinfeld, and L. F. Povirk. 1998. Implication of DNA-dependent protein kinase in an early, essential, local phosphorylation eventduring end-joining of DNA double-strand breaks in vitro. Biochemistry 37:9827–9835.
17. Gurdon, J. B., and M. P. Wickens. 1983. The use of Xenopus oocytes for theexpression of cloned genes. Methods Enzymol. 101:370–386.
18. Hagmann, M., K. Adlkofer, P. Pfeiffer, R. Bruggmann, O. Georgiev, D.Rungger, and W. Schaffner. 1996. Dramatic changes in the ratio of homol-ogous recombination to nonhomologous DNA-end joining in oocytes andearly embryos of Xenopus laevis. Biol. Chem. Hoppe-Seyler 377:239–250.
19. Hartley, K. O., D. Gell, G. C. M. Smith, H. Zhang, N. Divecha, M. A.Connelly, A. Admon, S. P. Lees-Miller, C. W. Anderson, and S. P. Jackson.1995. DNA-dependent protein kinase catalytic subunit: a relative of phos-phatidylinositol 3-kinase and the ataxia telangiectasia gene product. Cell82:849–856.
20. Jackson, S. P., and P. A. Jeggo. 1995. DNA double-strand break repair andV(D)J recombination: involvement of DNA-PK. Trends Biol. Sci. 20:412–415.
21. Kirchgessner, C. U., C. K. Patil, J. W. Evans, C. A. Cuomo, L. M. Fried, T.Carter, M. A. Oettinger, and J. M. Brown. 1995. DNA-dependent kinase(p350) as a candidate gene for the murine SCID defect. Science 267:1178–1183.
22. Knuth, M. W., S. I. Gunderson, N. E. Thompson, L. A. Strasheim, and R. R.Burgess. 1990. Purification and characterization of proximal sequence ele-ment-binding protein 1, a transcription activating protein related to Ku andTREF that binds the proximal sequence element of the human U1 promoter.J. Biol. Chem. 265:17911–17920.
23. Labhart, P. 1995. The Xenopus 9-bp ribosomal terminator (T3 box) is apause signal for the RNA polymerase I elongation complex. Nucleic AcidsRes. 23:2252–2258.
24. Labhart, P. 1997. mRNA encoding the catalytic subunit of DNA-dependentprotein kinase is widely expressed in Xenopus cells. Gene 203:235–240.
25. Labhart, P. Unpublished observations.26. Lehman, C. W., M. Clemens, D. K. Worthylake, J. K. Trautman, and D.
Carroll. 1993. Homologous and illegitimate recombination in developingXenopus oocytes and eggs. Mol. Cell. Biol. 13:6897–6906.
27. Lieber, M. R., J. E. Hesse, S. Lewis, G. C. Bosma, N. Rosenberg, K. Mizuu-chi, M. J. Bosma, and M. Gellert. 1988. The defect in murine severe com-
bined immune deficiency: joining of signal sequences but not coding seg-ments in V(D)J recombination. Cell 55:7–16.
28. Mimori, T., and J. A. Hardin. 1986. Mechanism of interaction between Kuprotein and DNA. J. Biol. Chem. 261:10375–10379.
29. Nicolas, A. L., and C. S. H. Young. 1994. Characterization of DNA endjoining in a mammalian cell nuclear extract: junction formation is accompa-nied by nucleotide loss, which is limited and uniform but not site specific.Mol. Cell. Biol. 14:170–180.
30. Pang, D., S. Yoo, W. S. Dynan, M. Jung, and A. Dritschilo. 1997. Ku proteinsjoin DNA fragments as shown by atomic force microscopy. Cancer Res.57:1412–1415.
31. Pfeiffer, P., S. Thode, J. Hancke, and W. Vielmetter. 1994. Mechanisms ofoverlap formation in nonhomologous DNA end joining. Mol. Cell. Biol.14:888–895.
32. Pfeiffer, P., and W. Vielmetter. 1988. Joining of nonhomologous DNA dou-ble strand breaks in vitro. Nucleic Acids Res. 16:907–924.
33. Reeves, W. H. 1985. Use of monoclonal antibodies for the characterization ofnovel DNA-binding proteins recognized by human autoimmune sera. J. Exp.Med. 161:18–39.
34. Reeves, W. H. 1992. Antibodies to the p70/p80 (Ku) antigens in systemiclupus erythematosus. Rheum. Dis. Clin. N. Am. 18:391–414.
35. Reeves, W. H., A. Pierani, C. H. Chou, T. Ng, C. Nicastri, R. G. Roeder, andZ. M. Sthoeger. 1991. Epitopes of the p70 and p80 (Ku) lupus autoantigens.J. Immunol. 146:2678–2686.
36. Schaal, H., P. Pfeiffer, M. Klein, P. Gehrmann, and A. Scheid. 1993. Use ofDNA end joining activity of a Xenopus laevis egg extract for construction ofdeletions and expression vectors for HIV-1 Tat and Rev proteins. Gene124:275–280.
37. Smider, V., W. K. Rathmell, M. R. Lieber, and G. Chu. 1994. Restoration ofx-ray resistance and V(D)J recombination in mutant cells by Ku cDNA.Science 266:288–291.
38. Taccioli, G. E., T. M. Gottlieb, T. Blunt, A. Priestley, J. Demengeot, R.Mizuta, A. R. Lehmann, F. W. Alt, S. P. Jackson, and P. A. Jeggo. 1994.Ku80: product of the XRCC5 gene and its role in DNA repair and V(D)Jrecombination. Science 265:1442–1445.
39. Thode, S., A. Schafer, P. Pfeiffer, and W. Vielmetter. 1990. A novel pathwayof DNA end-to-end joining. Cell 60:921–928.
40. Weis-Garcia, F., E. Besmer, D. J. Sawchuk, W. Yu, Y. Hu, S. Cassard, M. C.Nussenzweig, and P. Cortes. 1997. V(D)J recombination: in vitro codingjoint formation. Mol. Cell. Biol. 17:6379–6385.
41. Yaneva, M., T. Kowalewski, and M. R. Lieber. 1997. Interaction of DNA-dependent protein kinase with DNA and with Ku: biochemical and atomic-force microscopy studies. EMBO J. 16:5098–5112.
VOL. 19, 1999 Ku-DEPENDENT DNA END JOINING IN VITRO 2593
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 17
Oct
ober
202
1 by
114
.69.
161.
189.





























