Embed Size (px)
Citation preview

/ . Embryol. exp. Morph. Vol. 26, 3, pp. 543-570, 1971 5 4 3
Printed in Great Britain
Histological features of neural inductionin Xenopus laevis
by D. TARIN1
From the Department of Anatomy, University of Leeds
SUMMARY
It was first established by grafting experiments that neural induction occurs in Xenopuslaevis and that it is the mesoderm in the dorsal lip of the blastopore which normally exercisesthis function. The subsequent histological work provided the following information:
At stage 10i mesodermal invagination was already well under way, in advance of theformation of the archenteric cavity. This confirms the earlier observations of Nieuwkoop &Florschutz(1950).
The first evidence of neural induction, thickening of the mid-dorsal ectoderm combinedwith the development of an inner tier of columnar cells, occurred at stage 11£.
By stage .12 there was generalized thickening of the dorsal ectoderm and between stage 12£and .13 the brain and spinal cord regions of the neural plate became distinguishable.
The dorsal mesoderm segregated into notochord rudiment and two lateral masses at stage13 and the latter further subdivided into paraxial mesoderm and lateral plates by stage 14.
The margins of the neural plate were clearly distinguished from presumptive epidermis bystage 15 and the median neural groove was also well marked.
In the next two stages the folding of the neural plate in the line of this groove proceededrapidly. The dorsoventral enlargement of the somites and the relative shrinkage of the noto-chord were considered to contribute to the mechanism of neurulation.
Regionalization of the brain into prosencephalon, mesencephalon and rhombencephalonwas in progress at stages 18 and 19.
These results indicate that induction consists of an initial activation of dorsal ectoderm(generalized thickening) followed by gradual transformation of the neural plate to form thedifferent parts of the central nervous system (regionalization).
Intercellular metachromatic material was noted in various parts of the embryo. This wasmost plentiful between stage 10£ and stage .13 and thereafter gradually decreased. It was theonly feature which persisted long enough to represent a possible inductive agent.
At all stages the archenteron was lined with a continuous layer of endoderm. This indicatesthat the mode of formation of the gastro-intestinal tube in Xenopus is different to that inurodeles. It further implies that the mesoderm is not present on the blastular surface prior togastrulation but lies in deeper layers.
INTRODUCTION
This investigation was performed as an essential accompaniment to aprogramme incorporating electron microscopical, histochemical and trans-plantation techniques in the study of primary embryonic induction in Xenopuslaevis.
It was first necessary to establish that induction occurs in Xenopus and that it1 Author's address: Department of Anatomy, University of Leeds, Leeds 2, U.K.

544 D. TARIN
is effected by the mesodermal cells migrating through the dorsal lip of theblastopore. The process was next observed in progress by time-lapse cinemato-graphy (Tarin, Scott & Sharp, 1970; Sharp & Tarin, 1970) and these histologicalobservations were then performed to provide a basis for the interpretation ofresults obtained by other techniques, which will be presented in later publica-tions.
The work has been performed on Xenopus he vis because the eggs can beobtained at will in any season and this allows greater flexibility in the planningof experiments.
MATERIALS AND METHODS
Fertilized eggs were obtained by the injection of' Pregnyl' (Organon Labora-tories) according to the scheme described by Brown (1970). These were washedtwice in tap water and once in distilled water and finally placed in a dish con-taining 10% Niu-Twitty medium. The eggs were then reared and stagedaccording to the instructions of Nieuwkoop & Faber (1967). For histologicalstudies, six specimens at each of stages 10i, 11, H i , 12, 13, 14, 15, 16, 17, 18, 19(Nieuwkoop & Faber, 1967) inclusive, were selected with a binocular dissectingmicroscope and fixed in 4% glutaraldehyde in cacodylate buffer, pH 7-35. Theeggs were embedded in agar to permit subsequent orientation in the embeddingmedium by the technique described by Scott, Tarin & Sharp (1970). Next theywere washed overnight in cacodylate-sucrose mixture, post-fixed in osmiumtetroxide, dehydrated in a graded series of alcohols and embedded in Araldite.For each stage, three specimens were oriented to permit sectioning in a coronalplane and three in a sagittal plane. Thick sections (1-2 /im) were cut with anLKB Ultratome III and stained with 1 % toluidene blue. This study of neuralinduction was performed mainly on mid-sagittal and mid-coronal sections.
Grafting techniques (Fig. 1)
The operations were performed in Petri dishes half filled with blackened agarproduced by adding animal charcoal to molten agar and then allowing it to set.The agar was covered with full strength Niu-Twitty medium which was usedbecause the concentrated solution promotes more rapid healing than the 10%dilution used for standard rearing of embryos. Five embryos of stages 10^-11were used as donors for transplantation of the dorsal lip of the blastopore. Eachwas placed beside a recipient of similar age in an agar lined Petri dish and thejelly coats removed with forceps. The eggs were then placed in shallow groovesin the agar and the central portion of the dorsal lip of the donor excised with fineglass needles. This was then placed in a depression of similar size created oneach recipient by excising a piece of tissue in the region of the prospectivelateral or ventral lip. The graft was held in position while healing with micro-needles, controlled by a Leitz micromanipulator, for approximately half anhour. This procedure was repeated for each pair of eggs and the operated
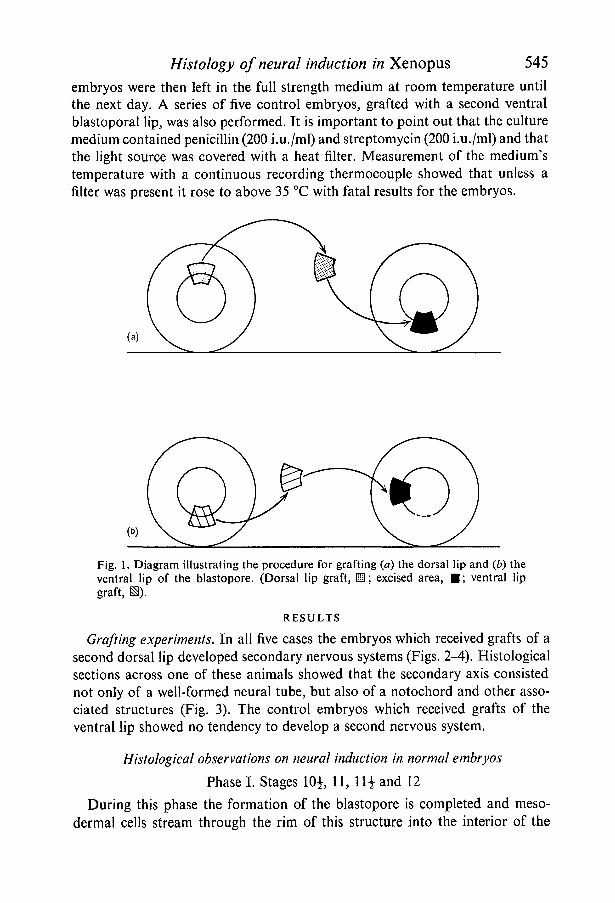
Histology of neural induction in Xenopus 545embryos were then left in the full strength medium at room temperature untilthe next day. A series of five control embryos, grafted with a second ventralblastoporal lip, was also performed. It is important to point out that the culturemedium contained penicillin (200 i.u./ml) and streptomycin (200 i.u./ml) and thatthe light source was covered with a heat filter. Measurement of the medium'stemperature with a continuous recording thermocouple showed that unless afilter was present it rose to above 35 °C with fatal results for the embryos.
Fig. 1. Diagram illustrating the procedure for grafting («) the dorsal lip and (6) theventral lip of the blastopore. (Dorsal lip graft, H; excised area, • ; ventral lipgraft, S).
RESULTS
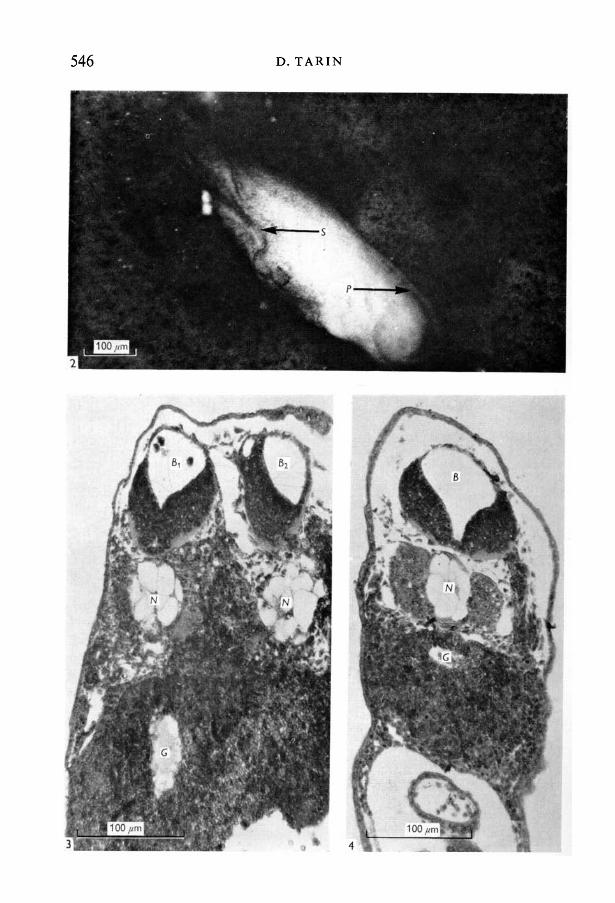
Grafting experiments. In all five cases the embryos which received grafts of asecond dorsal lip developed secondary nervous systems (Figs. 2-4). Histologicalsections across one of these animals showed that the secondary axis consistednot only of a well-formed neural tube, but also of a notochord and other asso-ciated structures (Fig. 3). The control embryos which received grafts of theventral lip showed no tendency to develop a second nervous system.
Histological observations on neural induction in normal embryos
Phase I. Stages 10*, 11, 1H and 12
During this phase the formation of the blastopore is completed and meso-dermal cells stream through the rim of this structure into the interior of the
546 D. TARIN

Histology of neural induction in Xenopus 547embryo. HistologicaUy it was evident that mesodermal imagination occurredearlier and faster in the dorsal than in the ventral region. Prior to stage 12coronal sections were difficult to interpret and yielded no really valuableinformation. The description of stages 10-£, 11, and 11 -̂ is therefore based onsagittal sections alone.
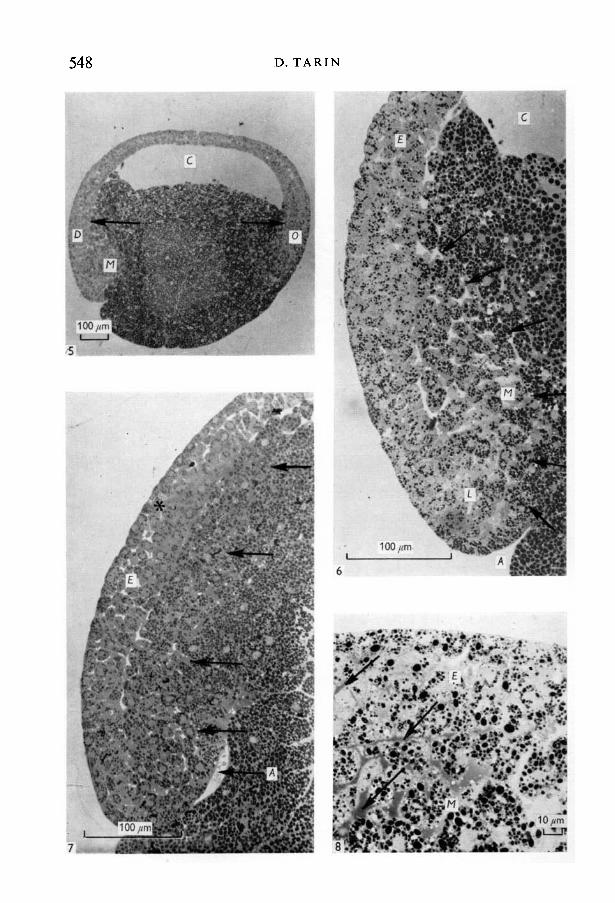
Stage 10\External appearance. Small shallow blastoporal groove and dorsal lip of
blastopore evident.Histological features. Although indentation of the surface to form the archen-
teron was only just beginning, dorsal mesodermal invagination was well underway (Figs. 5, 6). These cells were rounded and loosely grouped but formed aneasily recognized second sheet of tissue (Fig. 6) distinct from the surface layer.(For ease of description the latter will be referred to as the ectoderm although,at this stage, it cannot strictly be classified as such because it contained cellswhich later invaginated to form the endoderm.) The many large intercelluarspaces in the mesoderm were mostly filled with metachromatic material. Incontrast the dorsal ectoderm was composed of tightly packed polygonal cells,separated by small amounts of metachromatic material and it had a clearboundary with the underlying tissues. It was also noted that the ectoderm was
ABxB»CDEGHJKLx
L«M
KEY TO LABELLING OF FIGURES
ArchenteronBrain (host)Brain (induced)Blastocoele cavityDorsal aspect of embryoEctodermGut cavityEndodermSpinal cordParaxial mesodermDorsal lip of the blastoporeVentral lip of the blastoporeMesoderm
N0PQRSTUV
wXY
z
FIGURES
NotochordVentral aspect of embryoPrimary (host) nervous systemBlastoporeMetachromatic materialSecondary (induced) nervous systemLateral plate mesodermProsencephalonMesenchephalonRhombencephalonPrechordal plateYolk plugStomadeal membrane
2-4
Fig. 2. Xenopus laevis embryo with two nervous systems (x 180) - primary (P) andsecondary (S). The secondary neural axis resulted from grafting the embryo with anextra dorsal lip of the blastopore.Fig. 3. Histological section (x 400) across a similar embryo to that in Fig. 2 to showthe presence of two brains (BL and B»). Two notochords (M) are also visible. Note thatinduced nervous systems (B») are commonly not so perfectly formed as normal ones(Fig. 4).Fig. 4. Section across a normal embryo for comparison with Fig. 3 (x 400).
35 E M B 26
548 D. TARIN

Histology of neural induction in Xenopus 549thicker in the dorsal and ventral regions of the egg (Fig. 5) than over the blasto-coele cavity where its cells were somewhat more loosely arranged.
The presence of 'flask-shaped' cells described earlier by Holtfreter (1943) andby Perry & Waddington (1966) in the vicinity of the invaginating archenteronwas confirmed. Strong metachromasia was also observed in the intercellularspaces around these.
Stage 11
External inspection showed lateral extension of the blastoporal groove sothat the lateral lips of the blastopore were present.
Histological features. These were, in general, quite similar to those of stage-10-i-embryos. Some differences were noted, however. Thus, for instance, the archen-teron cavity was slightly deeper and the surface indentation in the ventral regionwas just beginning. The ventral mesoderm was also beginning to invaginate andthere was more metachromatic material in the intercellular spaces in both dorsaland ventral regions. In the dorsal lip, however, there was no real change in theextent of mesodermal movement or the arrangement of cells in either the meso-derm or the ectoderm.
Stage 11}
Externally the blastoporal groove formed a circle around the yolk plug. Thedorsal, ventral and lateral lips of the blastopore were distinguishable.
Histological sections showed that the archenteric and mesodermal invagina-tion was further advanced with the latter still being somewhat ahead (Fig. 7).The mesodermal cells were rounded and irregularly arranged but were moreclosely grouped in the dorsal than the ventral lip. The ectoderm was thickest inthe mid-dorsal area where an inner tier of low columnar cells could be distin-
FIGURES 5-8
Fig. 5. Sagittal section of stage-10£ embryo (x 110). The dorsal (Z>) and ventral (O)'ectoderm' is thicker than that over the blastocoele (C). Mesodermal invagination(A/) is well under way on the dorsal side.Fig. 6. Detail of the dorsal lip of stage-10£ embryo (x 400), sagittal section. This is thesame embryo as shown in Fig. 5. The mesodermal invagination (M) is far in advanceof the archenteron (A) and has formed a separate sheet of tissue under the ectoderm(£). The arrows mark the approximate boundary between this sheet and the endo-derm. Note that the mesodermal cells are rounded and loosely packed.Fig. 7. Dorsal lip of stage-11 \ embryo (x 360). The extent of the mesodermal invagi-nation and its boundary with the endoderm are marked with arrows. In the mid-dorsal region (asterisk) the ectodermal cells are tightly packed and the mesodermaland ectodermal layers are in close apposition.Fig. 8. Mid-dorsal region (x 660) stage 1 \\. The blastopore lies to the left but is notvisible in this picture. Note the presence of intercellular metachromatic material(arrows) in the mesoderm and to a lesser extent in the ectoderm and at the ecto-mesodermal boundary. Towards the right (rostrally) there is a decrease in thismaterial and the dorsal ectoderm is thinner.
35-2
550 D. TARIN
100/mi

Histology of neural induction in Xenopus 551guished. These cells were extremely tightly packed together and in the same areathe mesodermal cells were very closely applied to the deep surface of the ecto-dermal layer (Fig. 7). These new features were considered to be the first evidenceof nervous system induction.
Metachromatic material was abundant in the mesodermal layer between therim of the dorsal lip of the blastopore and the mid-dorsal region. More rostrallyit abruptly terminated as the ectoderm decreased in thickness (Fig. 8).
Stage 12
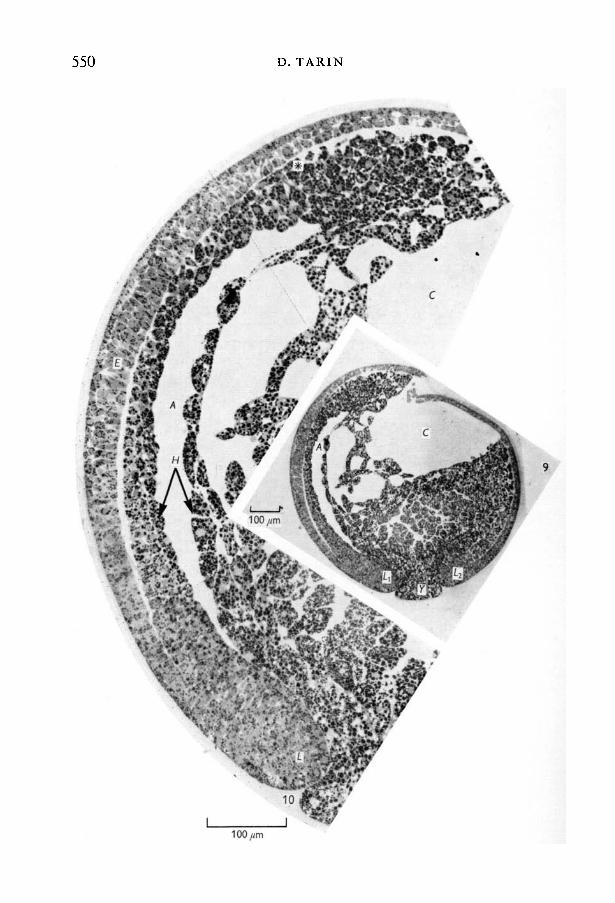
Externally the circular blastopore was contracting in size.Sagittal sections showed that the invagination of the archenteric cavity and
of the mesoderm was quite advanced (Figs. 9,10). It seemed that the mesodermalcells had probably almost reached the blastocoele, but in this area it wasdifficult to distinguish them from endodermal ones, because the size andappearance of the cells present were intermediate between the two types.
Comparison of the dorsal and ventral lips of the blastopore revealed thatthey differed in two main respects. First, the arrangement of the mesodermalcells in the dorsal lip was much tighter and more regular (Fig. 11). In fact, somein the caudal end of the advancing mesodermal sheet were columnar in shape.Secondly, metachromatic material was present both in the large inter-cellularspaces of the dorsal mesoderm and also in the narrow clefts between cells inthe outer (ectodermal) layer. In the ventral lip, however, it was present onlybetween the mesodermal cells.
The ectomesodermal junction was quite distinct in both dorsal and ventralregions and the degree of separation between the two layers varied both fromspecimen to specimen and in different areas of the same specimen. Dorsally,however (Fig. 10), the layers were usually closely apposed (never more than10 /mi apart and frequently less) but when the space was large it nearly alwayscontained clumps of metachromatic material. Towards the lip of the blastoporethe ectomesodermal boundary became indistinct about 8-10 cells from the actualrim. Further towards the rim, the cells became irregularly disposed although stillclosely grouped, and it was assumed that organisation into layers took place inthis region as the mesodermal cells passed through.
It was found that the cells of the 'ectodermal layer' are fairly uniform in
F I G U R E S 9, 10
Fig. 9. Sagittal section, stage-1 2 embryo (x 100). Survey picture showing the yolk plug(Y) protruding between the dorsal and ventral lips of the blastopore (LY and L2).
Fig. 10. Dorsal part of same (stage- .12) embryo as Fig. 9 ( x 300). Invagination of thearchenteron cavity (A) has progressed so that it now extends under most of thedorsal surface. Note the thickening of the ectoderm and the presence of columnarcells in this region. The rostral limit of mesodermal invagination is vague but itprobably corresponds with the position marked by the asterisk. The ectomesodermaljunction is clearly defined and the gap between the two layers varies in size.
552 D. TARIN
appearance and it was therefore not possible to distinguish the prospectivemesodermal elements on morphological criteria alone. Once they had passedinto the mesodermal layer however they were identifiable by such features asgrouping, position in the embryo, relative size, etc.
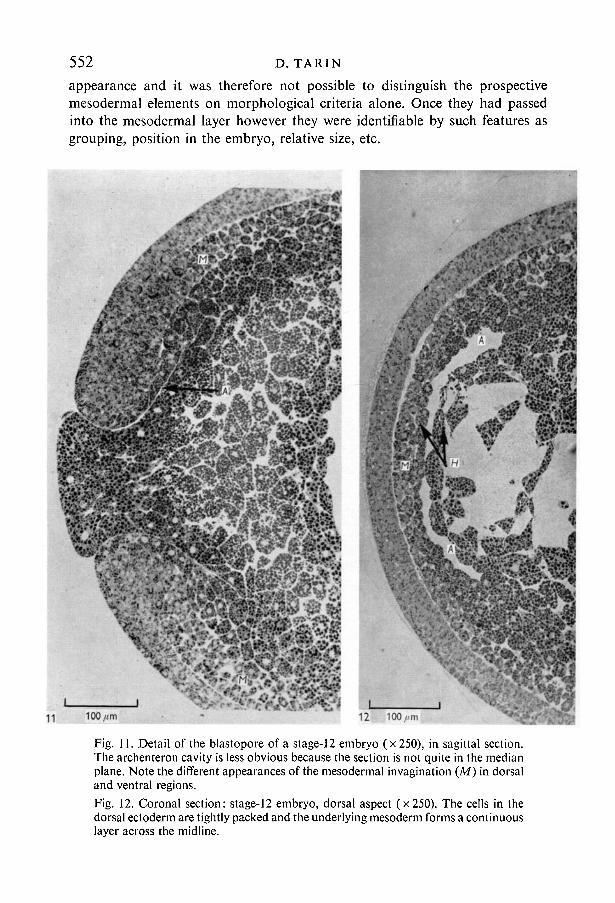
Fig. 11. Detail of the blastopore of a stage-12 embryo (x 250), in sagittal section.The archenteron cavity is less obvious because the section is not quite in the medianplane. Note the different appearances of the mesodermal invagination (M) in dorsaland ventral regions.Fig. 12. Coronal section: stage-12 embryo, dorsal aspect (x 250). The cells in thedorsal ectoderm are tightly packed and the underlying mesoderm forms a continuouslayer across the midline.
Histology of neural induction in Xenopus 553The dorsal ectodermal layer was thickest approximately half way between the
edge of the lip and the blastocoele cavity (Fig. 10). Here there were three tiersof cells, the inner one of which was clearly columnar. Proceeding towards theblastopore, although the ectoderm became thinner, the cells remained closelygrouped. Towards the blastocoele, however, the ectoderm reverted to a two-tiered arrangement in which the cells were more loosely packed. In some speci-mens it was noted that the ectodermal thickening and rearrangement apparently
13
Fig. 13. Sagittal section: stage-13 embryo (x!20), survey picture. Note that thearchenteron cavity (A) has expanded. The arrow heads indicate the gradual decreasein dorsal mesodermal thickness between the blastopore (Q) and the cephalic end.
extended further rostrally than the position reached by the invaginating meso-dermal cells. This might have been an illusion due to the difficulty in distinguish-ing mesodermal cells from endodermal ones at the apex of the advance.Alternatively, it might have indicated that the inductive effect can be exerted ata distance of ten cell diameters or more.
Mid-coronal sections (Fig. 12) confirmed that the dorsal ectoderm was in
554 D. TARIN
14 100//m

Histology of neural induction in Xenopus 555some places three-tiered with an inner layer of columnar cells. The dorsalmesoderm contained rounded cells which were more closely packed in themidline than elsewhere and the archenteron cavity was clearly lined with acontinuous layer of endoderm.
Summary of features of phase IWhile mesodermal invagination is well under way in the dorsal area at stage
10} (i.e. before archenteron formation begins) it is quite evident that the firstectodermal responses to inductive stimuli are not seen until stage 11} or 12.These consist of increase in the total thickness and number of layers of theectoderm and the appearance of the columnar cells in the inner layer. Somefeatures of mesodermal invagination differed in dorsal and ventral regions.
Phase II. Stages 13-17In this phase the neural plate appeared and then began to fold in the pro-
duction of the neural tube. Internal changes included a rapid expansion in thesize of the archenteron cavity to form the primitive gut and the segregation ofthe notochord from the two lateral sheets of mesoderm.
Stage 13Externally the blastopore was small and slit-shaped indicating imminent
closure and the dorsal ectoderm was less pigmented than other regions.Sagittal sections showed an enormously expanded archenteron and a cor-
respondingly decreased blastocoele (Fig. 13). As in the stage-12 embryos thearchenteron had a complete and continuous endodermal lining.
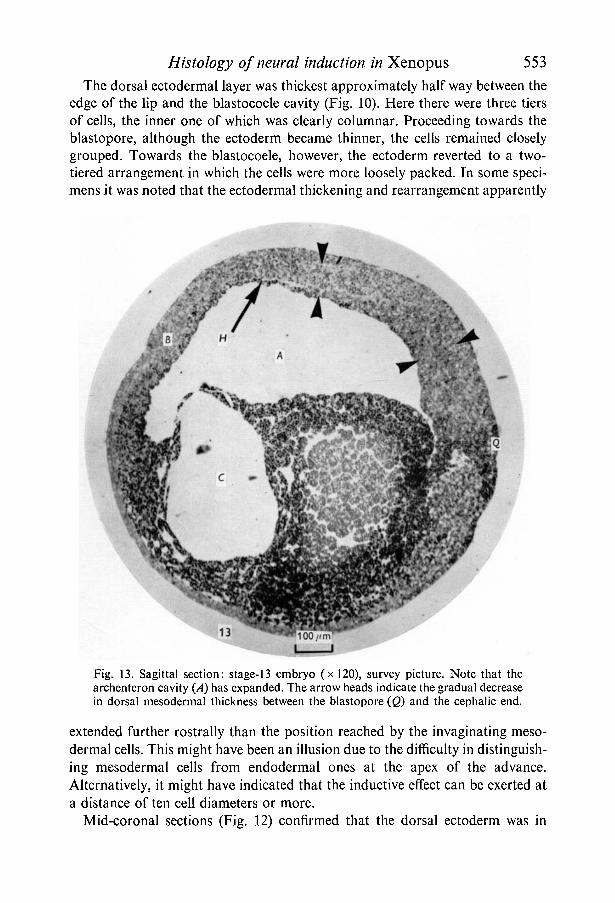
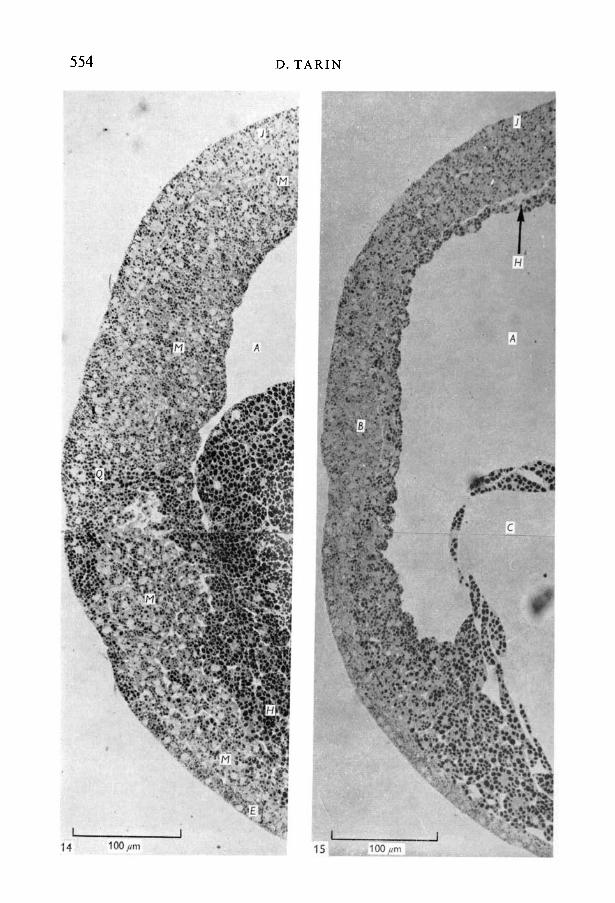
The closing blastopore formed the caudal end of the developing embryo andthe dorsal and ventral components of the continuing mesodermal invaginationin this region were very different. Dorsally, it extended the whole length of theembryo and gradually declined in thickness from caudal to cephalic end (Fig. 13).Starting in the dorsal lip of the blastopore the cells were closely grouped together(Fig. 14) and slightly more rostrally became arranged in columnar array toform a mesodermal sheet 5-6 columnar cells thick. This then narrowed until inthe head region a single layer of flattened cells (the prechordal plate) lay betweenthe brain and the endodermal lining of the archenteron (Fig. 15).
In contrast the mesodermal cells in the ventral lip of the blastopore were rounderand more irregularly arranged with large intercellular spaces (Fig. 14). The ventral
FIGURES 14, 15
Fig. .14. Detail of caudal end of stage-13 embryo, sagittal section (x400). Sameembryo as Fig. 13. The dorsal mesodermal layer is thicker, has more closely groupedcells and extends further rostrally than its ventral counterpart.Fig. 15. Detail of rostral end of stage-13 embryo, sagittal section (x400). Sameembryo as Figs. 13 and 14. The brain (B) and spinal cord (/) portions of the neuralplate can now be distinguished. The single layer of endodermal cells lining the roofof the archenteron is also easily seen.
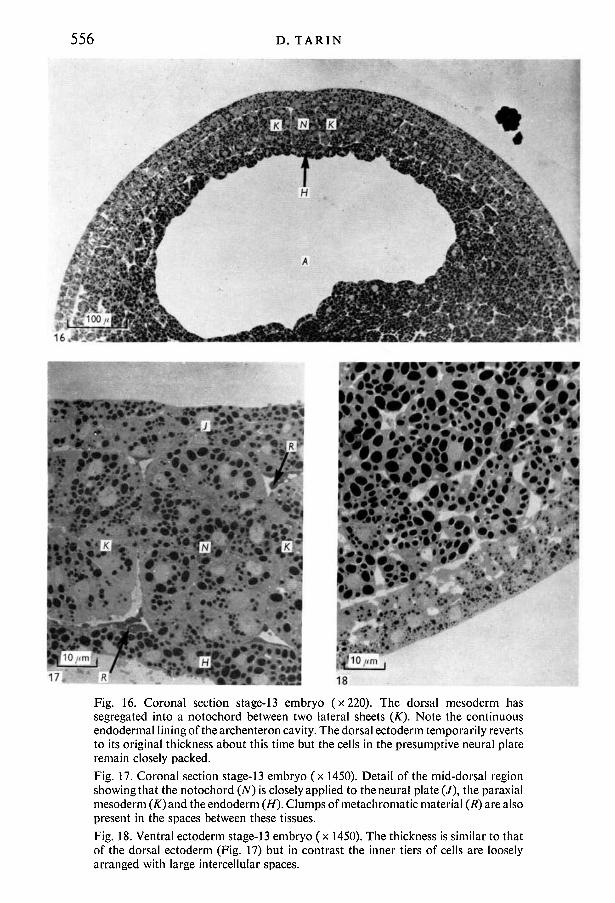
556
17/ 18Fig. 16. Coronal section stage-13 embryo (x220). The dorsal mesoderm hassegregated into a notochord between two lateral sheets (K). Note the continuousendodermal lining of the archenteron cavity. The dorsal ectoderm temporarily revertsto its original thickness about this time but the cells in the presumptive neural plateremain closely packed.Fig. 17. Coronal section stage-13 embryo (x 1450). Detail of the mid-dorsal regionshowing that the notochord (N) is closely applied to the neural plate (/), the paraxialmesoderm (̂ T)and the endoderm (H). Clumps of metachromatic material (R) are alsopresent in the spaces between these tissues.Fig. 18. Ventral ectoderm stage-13 embryo (x 1450). The thickness is similar to thatof the dorsal ectoderm (Fig. 17) but in contrast the inner tiers of cells are looselyarranged with large intercellular spaces.

Histology of neural induction in Xenopus 557part of the invaginating mesodermal sleeve did not extend for more than one-thirdof the length of the embryo.
The ectoderm also differed in dorsal and ventral regions of the stage 13embryos. Although of approximately similar thickness (except at the rostralend where the brain was forming) the dorsal ectodermal cells were closelypacked together and the inner tier was low columnar in shape. The ventralectoderm consisted of two tiers of cuboidal cells with small intercellular spacesin the inner one (compare Figs. 17 and 18). It will be remembered that thedorsal ectoderm thickened at stage 12 and this transient reversion to a thinnerstructure in which the cells remained columnar and closely attached was ofsome interest. It was seen only in the trunk region and for a brief period.
At the head end the dorsal ectoderm was four to six columnar cells thick andthis constituted the developing brain plate (Fig. 15). It had a prominent rostralmargin and tapered caudally to merge with the spinal cord. In this region theinner ectodermal cells were sometimes not as closely packed as in Fig. 15. Theoutermost, as in other regions, were not columnar and were always tightlyjoined together.
The ectomesodermal junction was well defined in both dorsal and ventralregions but had no special features in either. Once again the degree of separationbetween the two germ layers varied slightly. Metachromatic material wasprominent in the extracellular spaces of both dorsal and ventral lips of theblastopore. More rostrally, in the trunk region, there was none extracellularlybut some cytoplasmic metachromasia was observed. At the head end similarmaterial was once again present in quantity in the extracellular spaces of thebrain plate and underlying mesoderm and at the ectomesodermal junction.
Coronal sections showed that the dorsal mesoderm had segregated into threecomponents consisting of the notochord and two lateral sheets. There was littleevidence as yet of subdivision of the lateral masses to form somites (paraxialmesoderm) and lateral plate mesoderm. The notochord was in contact dorsallywith the neural plate and ventrally with the single layer of endodermal cellsforming the roof of the archenteron cavity (Figs. 16, 17).
Stage 14
External features. Early neural plate formation could be distinguished by aslight prominence of the dorsal surface. The caudal part was most evident andfeatured three faint grooves radiating rostrally from the blastopore. The brainplate was, however, not visible.
His to logically these embryos were similar to those of the previous stage andthe main changes affected the dorsal ectoderm, which was now clearly thickerthan elsewhere. This thickening constituted the neural plate which displayed awell-marked median (neural) groove, and a prominent rostral edge. The time-lapse cinematographic study mentioned earlier showed that at this period ofdevelopment a distinct ridge moved across the dorsal surface of the embryo
558 D. TARIN
20 100 //m

Histology of neural induction in Xenopus 559and in the wake of this the neural plate could be distinguished. Examination ofhistological sections of embryos from stages 13-16 showed that it was the pro-gressive rostral extension of this structure which was responsible for the movingridge phenomenon.
In most places the neural plate was composed of three tiers of closely packedcells, the inner one of which was columnar. Laterally the neural plate mergedwith the two-tiered non-neural ectoderm. The boundary between these was notdistinct but corresponded roughly with the junction of somitic and lateral platecomponents of the underlying mesoderm. A further point of difference betweenthe two types of ectoderm was that intracellular metachromasia was present inthe neural plate, whereas in the lateral ectoderm similar material was in theextracellular space. (The cephalo-caudal distribution of metachromasia was thesame as at stage 13.)
As with the previous stage the notochord was very close to the neural plate,endoderm and lateral sheets of mesoderm. The latter was now, however, furthersubdivided into paraxial and lateral plate sections. The paraxial mesoderm is theforerunner of the somites and coronal sections showed that the cells wereelongated and rearranged in radial fashion giving a rosette-like appearance (seefigures of later stages).
Stage 15
External features. The complete outline of the neural plate was clearly seenon the surface of the embryo. Most embryos were still round at this stage.
Once again histological sections showed only few changes since the previousstage. Sagittal sections showed that the brain plate was thicker and longer.Metachromatic material was still present although reduced. Most was intra-cellular although some was still observed at the ectomesodermal junction in thetrunk region. There was little in the vicinity of the brain plate.
Coronal sections showed that the neural plate was much thicker than the
FIGURES 19-21
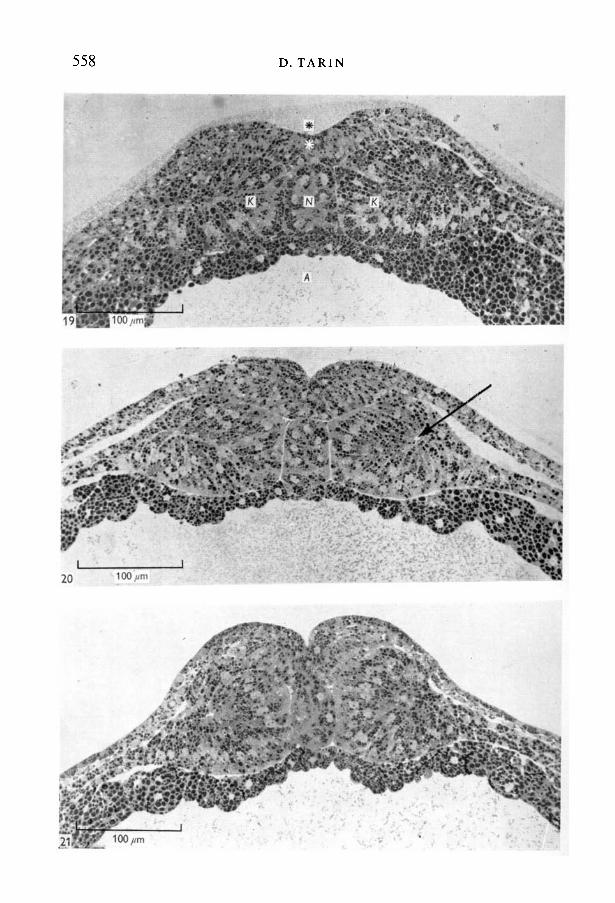
Fig. 19. Coronal section stage 15 (x400), dorsal aspect. The neural groove, blackasterisk, is evident and the neural plate, white asterisk, contains columnar cells whichslope inwards at the edges. The paraxial mesoderm (K) has segregated from thelateral plate mesoderm (T) and is now thicker than the notochord. Metachromaticgranules are present in the archenteron cavity and also in the space between theembryo and the vitelline membrane.Fig. 20. Coronal section stage 18 (x 400), dorsal aspect. The neural plate is muchthicker and the neural groove deeper than in the previous figure. The paraxialmesoderm is also thicker and contains an eccentrically placed myocoele cavity, arrow.Note the rosette-like arrangement of its columnar cells.Fig. 21. Coronal section stage 19 (x400), dorsal aspect showing further dorso-ventral enlargement of the somites, deepening of the neural groove and thickeningof the neural plate. Notice how the boundary between the somites and the neuralplate is more oblique than in previous stages.
560 D. TARIN
lateral ectoderm and the transition was moderately sharp (Fig. 19). The deeperpart of the neural plate contained columnar cells which at its edges slopedtowards the neural groove. It was also noted that clumps of metachromaticmaterial were present at the four corners of the notochord and that its shapehad become rounder. Accompanying this change was an apparent reduction inits dorso-ventral diameter relative to the somites which now projected above it.In fact the measurements recorded in Table 1 show that the notochord stayed
Table 1. Dimensions of the notochord and somites between stages 1.3 and 19
Stage
13
14
15
16
17
18
19
No. ofspecimens
2
2
2
2
1
2
2
NotochordA
Verticalmeasurement
Om)
111126
132126
120126
126120
117
120117
111126
Transversemeasurement
7896
9696
9090
12090
90
8784
81102
Somites.Vertical dimension(jim) only. Figureindicates mean of
each pair
108126
177180
162156
195171
192O
O O
OO
N
O
207204
The general trend is for the notochord to stay constant in size while the somitesgradually expand with particularly large increments between stages 13 and 14 andstages 18 and 19.
almost constant in size and that the somites enlarged vertically. Thus the neuralgroove dipped into a longitudinal trough formed between the somites andremained closely applied to the notochord which formed the floor of thisdepression.
Stages 16 and 17
During these stages the neural plate started to fold along a cephalo-caudalaxis to form the neural tube. Histological sections showed no dramatic internalchanges; only a gradual thickening of the neural plate coupled with a deepeningof the neural groove so that by the end of stage 17 the two sides of the neuralplate were approximating to one another. Under each half lay the masses ofparaxial mesoderm forming the presumptive somites and these graduallyincreased in size relative to the notochord which remained attached to the
Histology of neural induction in Xenopus 561ectodermal cells in the base of the neural groove. At the same time the embryostarted to elongate and the archenteron cavity increased in size.
Summary of Phase II
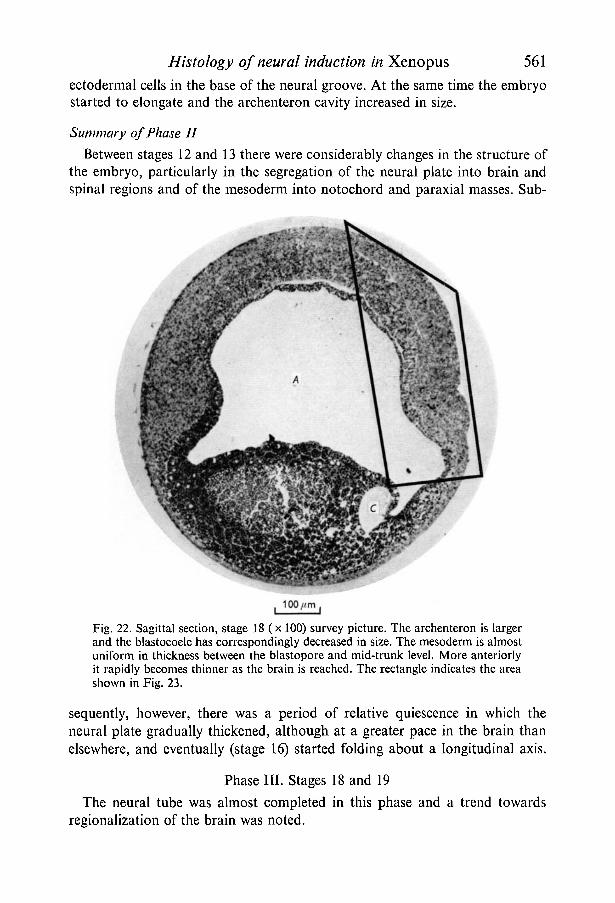
Between stages 12 and 13 there were considerably changes in the structure ofthe embryo, particularly in the segregation of the neural plate into brain andspinal regions and of the mesoderm into notochord and paraxial masses. Sub-
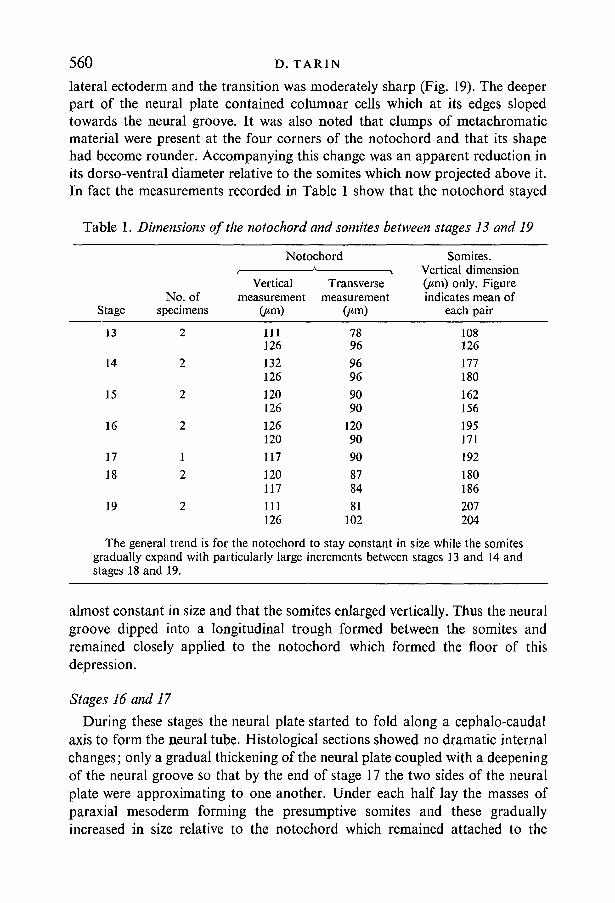
Fig. 22. Sagittal section, stage 18 (x 100) survey picture. The archenteron is largerand the blastocoele has correspondingly decreased in size. The mesoderm is almostuniform in thickness between the blastopore and mid-trunk level. More anteriorlyit rapidly becomes thinner as the brain is reached. The rectangle indicates the areashown in Fig. 23.
sequently, however, there was a period of relative quiescence in which theneural plate gradually thickened, although at a greater pace in the brain thanelsewhere, and eventually (stage 16) started folding about a longitudinal axis.
Phase III. Stages 18 and 19
The neural tube was almost completed in this phase and a trend towardsregionalization of the brain was noted.
562 D. TARIN
2 3 '

Histology of neural induction in Xenopus 563
Stage 18
In coronal sections the neural groove was narrow and deep (Fig. 20). Theneural plate consisted of a deeper part, two to three tiers of columnar cellsthick and a superficial one composed of a single tier of tightly apposed roundedcells. In the neural groove the cells of the superficial tier became columnar andoccupied most of the thickness of the neural plate as described by Schroeder(1970). The edges of the neural plate were now sharply defined and lay over themiddle of the paraxial mesoderm. In contrast to the underlying mesoderm theneural plate contained numerous mitotic figures.
The notochord and the paraxial mesoderm looked much the same as previouslywith the exception that a myocoele cavity could now be distinguished in thelatter. This contained metachromatic material and was eccentrically placed inthe presumptive somite mass so that the cells medial and inferior to it werecolumnar and those superior and lateral were small and rounded (Fig. 20).
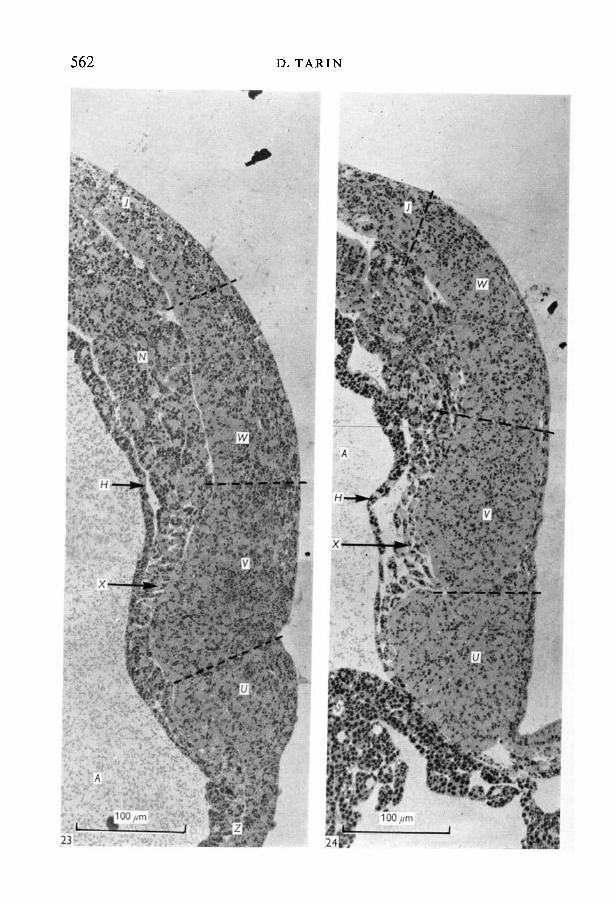
The first evidence of subdivision of the brain plate into three parts, separatedby grooves on the ventral aspect of the brain plate was observed in sagittalsections at this stage (Figs. 22, 23). The anterior, middle and posterior segmentsthus created represent the prosencephalon, mesencephalon and rhombenceph-alon respectively. In contrast to these developments the ventral ectoderm andmesoderm were substantially unchanged in thickness, cellular arrangement anddistribution since stage 13 or 14.
Deep to the brain lay the very thin portion of the mesodermal sheet known asthe prechordal plate (Fig. 23). Under the prosencephalon this consisted of asingle layer of flattened irregularly arranged cells. This increased to approxi-mately three layers of loosely packed cells in the mesencephalic region. Morecaudally the mesoderm was much thicker and the columnar cells were closelypacked. Moreover, the trunk and caudal sections of the dorsal mesoderm werenow of roughly uniform thickness (Fig. 22) thus differing from the situation inearlier stages (see stage 13).
The ectomesodermal junction was very tight in the posterior part of theembryo but more rostrally small spaces were occasionally present.
Metachromatic material, both extra- and intracellular, was still present at thisstage but was considerably reduced. In the ectoderm only minute traces remained
FIGURES 23, 24
Fig. 23. Detail of brain of stage-18 embryo (x 400). Same embryo as Fig. 22. Thebrain is divided into three regions (U, V, W) by ventral grooves. The rhombence-phalon (WO merges imperceptibly with the spinal cord (/) and the prosencephalon(£/) has a sharp boundary with the ectoderm of the stomadeal membrane (Z). Theprosencephalon (U) and mesencephalon (K) are underlain by a very thin sheet ofmesoderm - the prechordal plate (X).Fig. 24. Detail of brain of stage-19 embryo (x 400), sagittal section. The subdivision ofthe brain is more marked than at stage 18 and the rostral groove is much deeper.
36 EMB 26

564 D. TARIN
but in the mesoderm moderate quantities of extracellular metachromasia werestill visible in the prechordal plate and at the caudal end of the embryo. Coronalsections revealed that similar material was still present at the corners of thenotochord and at infrequent intervals around the somite blocks. A smallblastocoele cavity was still evident and this contained metachromatic granulesas did the capacious archenteron.
Stage 19
Externally the neural folds were in apposition along the length of the embryo.The protuberant spinal cord and flattened but laterally expanded brain wereeasily recognizable. Fusion of the neural folds was imminent.
Sagittal sections revealed that the regionalization of the brain was moreevident with a deep ventral groove separating the prosencephalon from themesencephalon (Fig. 24). Once again there were several mitoses in the ectodermbut apparently none in the mesoderm.
The thin prechordal plate was present only under the prosencephalon and theanterior part of the mesencephalon. The hind-brain overlay a thicker sheet ofmesoderm, the cells of which were not arranged so regularly as under the spinalcord.
Coronal sections (Fig. 21) showed a very deep neural groove, the neck ofwhich was constricted by the apposing neural folds in some parts of the embryo.The deeper portion, which is known to form the central canal of the spinal cord,was very small in comparison with the mass of nervous tissue around it and thefloor of the neural groove was lined with microvilli.
The somites were by this stage very thick in a dorsoventral plane and thenotochord and lateral mesoderm were relatively small. The notochord was almostround in cross-section and there were only traces of extracellular metachromaticmaterial associated with it. The archenteron cavity still contained abundantmetachromatic granules. Elsewhere in the embryo there was virtually no extra-cellular metachromatic material left. The intracellular variety was, however,still widely present.
DISCUSSION
The results of the grafting experiments described in the first part of this paperare similar to those obtained in chicks (Waddington & Schmidt, 1933), fish(Oppenheimer, 1936) and other amphibia (Spemann & Mangold, 1924). Theyshow that induction of the nervous system by mesoderm does occur in Xenopuslaevis and that, as in other amphibian species (Spemann, 1938) the portion ofthe mesoderm responsible for this invaginates through the dorsal lip of theblastopore.
The histological observations reveal a fairly clear-cut sequence of events inneural induction. Initial thickening occurs in the mid-dorsal ectoderm at aboutstage 1H and spreads over the rest of the dorsal surface by stage 12-12^. This is

Histology of neural induction in Xenopus 565rapidly followed by segregation of brain and spinal regions, clearly recognizableby stage 13.
There is next a period of relative quiescence during which the neural platecontinues to thicken and mechanical changes bring about folding around alongitudinal axis. Then, between stages 17 and 19 regionalization of the brainbegins with the segregation of its substance into three main masses separatedfrom each other by deep constrictions.
These observations show that the morphological events comprising neuralinduction are complex and gradual. Thus the nervous system does not appear abinitio as a preformed unit with inherent structural differences in various regionsbut, instead, as a diffuse thickening of the dorsal ectoderm (interpreted as'activation') which subsequently becomes regionally modified to form thedifferent parts of the C.N.S. (interpreted as 'transformation'). One thereforeexpects that the inductive processes responsible for these changes will prove tobe similarly complex and sequential rather than dependant on a single rapid event.This conclusion is similar to those reached by other investigators using graftingand other experimental manipulations (Nieuwkoop et al. 1952; Takaya, 1955;Eyal-Giladi, 1954), and supports in principle the activation-transformationhypothesis advanced by Nieuwkoop et al. (1952) and the similar scheme proposedby Sala (1955).
Although the present work has established when the thickening of the dorsalectoderm and its regionalization take place in Xenopus neural induction, thetiming of the stimuli which trigger these responses remains unknown. However,the work of Spemann (1938) and his associates shows that this is not of par-ticular importance because inductive stimuli are present for long after theneural tube is formed and it is the ability of the ectoderm to respond whichdecides when induction begins. Thus, we should consider (a) whether there areany features which persist for long periods during induction and might representthe agents responsible, and {b) whether there are any changes in the ectodermprior to induction which indicate when the cells become competent to respondto the inductive stimulus.
Consider first item (a): in a histological study it is of course only possible toidentify visible features, and the only one of these which persisted for long enoughto represent a possible inductive agent was the extracellular metachromaticmaterial. The exact chemical nature of this is at present uncertain, although thetypes of compounds possessing this staining property, which could be present atthis stage of development are quite limited. The most likely alternatives are eitherthe nucleic acids or some complex polysaccharide such as glycogen. Earlyinvestigators claimed both ribonucleic acid (Brachet, 1941, 1947) and glycogen(Woerdeman, 1933; Raven, 1933; see also review by Spemann, 1938) to bepresent in quantity in the dorsal lip of the blastopore but these reports did notspecifically discuss the extracellular distribution of such material. More recentworkers using the electron microscope also differ in their claims, for while Van
36-2

566 D. TARIN
Gansen & Schramm (1969) found only glycogen in the intercellular spaces of thegastrula of Xenopus laevis, Kelly (1970) contended that both RNA and glycogenare present in this site. The chemical nature of the intercellular material istherefore still not settled and its identification will require further work.
Any assessment of the role of the extracellular metachromatic material indevelopment must of course take into account its distribution in the variousparts of the embryo and the regional pattern of its disappearance. As it ispresent in both dorsal and ventral regions of the embryo from stage 11 onwards(see above) it seems unlikely that it is specifically involved in neural induction.(The counter-argument, that the ventral ectoderm might be simply unresponsiveto the inductive stimulus, is refuted by the production of secondary nervoussystems on the ventral aspect by transplantation of the dorsal lip of the blasto-pore - see above). On the other hand, it has to be conceded that while the sub-stances in dorsal and ventral lips of the blastopore are histochemically similar(Tarin, 1971a), these tests are relatively crude. For instance, they will notdistinguish one family of RNA or glycogen from another. Thus, in differentregions, there may be minute chemical differences in the molecular structure ofthe metachromatic material which are of great biological significance.
Consideration of the regional pattern of disappearance of this material isalso of some value in attempting to assess its function. It was noted above that itpersisted for longest in the dorsal and ventral lips of the blastopore and in theregion of the developing brain. This might indicate a role in cellular movementor in other activities involving high energy consumption. Further studies on thechemical composition and significance of this material are at present in progressand will be reported separately.
With regard to item (b) this light microscopical study did not result in the dis-covery of any changes which might assist in assessing when the ectoderm acquiresthe competence to respond to primary embryonic induction. Further work usinghistochemical and electron microscopical methods (Tarin, 1971 a, b), has alsofailed to reveal any obvious features of value for this purpose.
It seems therefore that a proper assessment of when reactivity to the activatingand regionalizing stimuli is acquired and lost will depend on grafting procedures.Yet these, too, have their technical limitations as evidenced by the conflictingresults of comparable experiments. Some of these discrepancies are probablydue to the difficulty of separating the ectodermal and mesodermal layers attheir natural interface. As noted in the results the relationship between thesetwo sheets of cells is usually very close although it is somewhat variable. Thus,unless future experiments are controlled by the histological assessment ofportions of the grafted material or the present methods are refined so thateither the ectoderm or the mesoderm is specifically marked in some way(Tarin, 1971 c) prior to the cutting of the graft, these discrepancies willcontinue.
The slight variability of the closeness of apposition of the ectodermal and

Histology of neural induction in Xenopus 567mesodermal layers in different embryos and in different parts of the same embryois compatible with the view that the inducing agent is a diffusible substance forit is known that neural induction can occur even across a Millipore filter provid-ing it is less than 25 pan. thick (Gallera, Nicolet & Bauman 1968), (i.e. the inter-acting tissues do not have to be in contact). Even in the most extreme cases theseparation did not exceed 10 /im in the specimens described above.
It seems established by the work of several investigators (Spemann, 1931;Holtfreter, 1933; Alderman, 1935; Deuchar, 1953; Takaya, 1955; see alsoreview by Saxen & Toivonen, 1962) that the segregation of the neural plate intodifferent regions (regionalization) is effected by the underlying mesoderm. Whatis not agreed, however, is whether the mesoderm itself is a mosaic of separateareas each responsible for the evocation of a different part of the central nervoussystem or whether regionalization is the result of variation in the length ofexposure to cumulative influences exerted by the mesoderm as it slides under theectoderm. As might be expected in such a situation there are various items ofexperimental evidence which are considered to support or oppose each of theseinterpretations. For instance, Waddington & Deuchar (1952) substituted theneural plate of late gastrulae with vitally stained ectoderm from early ones andfound that, in the embryos which survived, a whole normal neural axis wasformed from the marked transplant. This suggests that regionalization is notproduced by differential effects exerted during mesodermal imagination andtends to favour the alternative possibility of regional differences in the meso-derm. On the other hand, Eyal-Giladi (1954) removed portions of ectodermfrom the dorsal surface at different stages during gastrulation and transplantedthem to the ventral aspect of the embryos. She noted the development of pro-gressively more caudal neural structures in the grafts taken at successively laterstages of mesodermal invagination, and attributed this sequence to differencesin the length of exposure to cumulative effects exerted by the mesoderm slidingunderneath the dorsal surface.
The histological results presented above do not permit a choice between thesealternatives. However, the pertinence of the present paper to the problem isthat the further grafting experiments which will be needed to understand howregionalization occurs can be performed at suitable stages selected on the basisof the histological changes.
The mechanism of folding of the neural plate to form a neural tube is aproblem related to this study of neural induction. In contrast to the newt, wherethe neural plate is only one cell thick (Burnside & Jacobson, 1968), in Xenopusit is composed of several cell layers which all participate in the folding process.Thus it seems unlikely that the deformation in the shape of the surface ecto-dermal cells described by some investigators (Waddington & Perry, 1966;Baker & Schroeder, 1967; see also Wren & Wessels, 1969) is alone an adequatecausal factor in this species because it is difficult to see how this could achievemore than the production of a surface dimple or wrinkle. Although such a

568 D. TARIN
mechanism might initiate the formation of a neural groove it could be expectedto have little effect on the deeper layers of the ectoderm. It is therefore imaginedthat to accomplish folding of the full thickness of the neural plate other factorsmust be in operation. In fact, it seems highly likely that the formation of alongitudinal gutter, by the sinking of the notochord and the relatively fast dorsalexpansion of the somite masses between stages 13 and 19 makes a significantcontribution to this process. These histological observations confirm those ofSchroeder (1970) who drew similar conclusions and argued that this mechanismaugmented the effects of cellular rearrangement and alteration of cell shape.
The continuity of the endodermal lining of the archenteron cavity throughoutgastrulation in Xenopus contrasts markedly with the mode of formation of thegastro-intestinal tube in urodele amphibians. In such embryos the roof of thearchenteron cavity is initially formed by mesoderm which must segregate fromthe more laterally placed endoderm before the latter can grow medially to meetits neighbour under the mesoderm and thus fill the defect in the endodermaltube. The importance of this observation for future work on Xenopus is that itimplies that the fate map of this animal differs considerably from others inhaving no mesoderm represented on the surface immediately prior to gastrula-tion. In turn, this suggests that the mesoderm is derived from deeper layers ofthe lining of the blastula. These deductions are supported by the results ofpreliminary surface marking experiments using vital dyes (D. Tarin, unpublishedobservations), and exactly concur with the views of Nieuwkoop & Florschutz(1950).
A further point of agreement between this work and theirs is that, in thisanimal, mesodermal invagination occurs well before the invagination of thearchenteric cavity. These two studies are also complementary in many respectswhere they do not overlap and the special contributions of the present work areto provide a detailed account of the relationships between ectoderm and meso-derm during neural induction, the distribution and fate of metachromaticmaterial and the changes in the ectoderm during individuation of the nervoussystem. The analysis of these particular problems was facilitated by the use of1 [im thick sections of material embedded in Araldite, which can be recom-mended as providing clearer visualization of histological features than paraffinwax sections.
I am very grateful to Mrs E. K. Jones for the outstanding quality of the technical assistanceshe has provided throughout this work. I also wish to thank Professor R. L. Holmes forreading and criticizing the manuscript and the Nunield Foundation for providing financialsupport for this research programme.

Histology of neural induction in Xenopus 569
REFERENCES
ALDERMAN, A. L. (1935). The determination of the eye in the anuran Hyla regilla. J. exp. Zool.70, 205-232.
BAKER, P. C. & SCHROEDER, T. E. (1967). Cytoplasmic filaments and morphogenetic move-ments in the amphibian neural tube. Devi Biol. 15, 432-450.
BRACHET, J. (1941). La localisation des acides pentosenucleiques dans les tissus animaux etles oeufs amphibiens et des Oiseaux. Archs Biol. 53, 207-257.
BRACHET, J. (1947). Nucleic acids in the cell and the embryo. Symp. Soc. exp. Biol. 1, 207-224.BROWN, A. L. (1970). The South African Clawed Toad. London: Butterworths.BURNSIDE, M. B. & JACOBSON, A. G. (1968). Analysis of morphogenetic movements in the
neural plate of the newt Taricha torosa. Devi Biol. 18, 537-552.DEUCHAR, E. M. (1953). The regional properties of amphibian organiser tissue after dis-
aggregation of its cells in alkali. / . exp. Biol. 30, 18-43.EYAL-GILADI, H. (1954). Dynamic aspects of neural induction in amphibia. Archs Biol. 65,
179-259.GALLERA, J., NICOLET, G. & BAUMAN, M. (1968). Neural induction in birds through a milli-
pore filter: study by optical and electron microscopy. / . Embryol. exp. Morph. 19,439-450.HOLTFRETER, J. (1933). Organisierungstuffen nach regionaler Kombination von Entomeso-
derm mit Ektoderm. Biol. Zbl. 53, 404-431.HOLTFRETER, J. (1943). A study of the mechanics of gastrulation I. /. exp. Zool. 94, 261-318.KELLY, R. O. (1970). An electron microscope study of chordamesoderm-neurectoderm
association in gastrulae of a toad, Xenopus laevis. J. exp. Zool. 170, 153-180.NIEUWKOOP, P. D. et al. (1952). Activation and organisation of the central nervous system in
amphibians I, I! and III. /. exp. Zool. 120, 1-108.NIEUWKOOP, P. D. & FABER, J. (1967). Normal table o/Xenopus laevis (Daudin). Amsterdam:
North Holland Publishing Company.NIEUWKOOP, P. D. & FLORSCHUTZ, P. A. (1950). Quelques caracteres speciaux de la gastrula-
tion et de la neurulation de l'oeuf de Xenopus laevis, Daud. et de quelques autres Anoures.Archs Biol. 61, 113-150.
OPPENHEIMER, J. M. (1936). Transplantation experiments on developing teleosts (Fundulusand Perca). J. exp. Zool. 72, 409-437.
PERRY, M. M. & WADDINGTON, C. H. (1966). Ultrastructure of the blastopore cells in the newt./. Embryol. exp. Morph. 15, 317-330.
RAVEN, C. P. (1933). Experimentelle Untersuchunge iiber den Glycogenstoffswechsel desOrganisationszentrums in der Amphibien Gastrula. Proc. K. ned. Akad. Wet. Ser. C. 36,566-569.
SALA, M. (1955). Distribution of activating and transforming influences in the archenteronroof during the induction of the nervous system in amphibians. Proc. K. ned. Akad. Wet.Ser. C. 58, 635-647.
SAXEN, L. & TOIVONEN, S. (1962). Primary Embryonic Induction. London: Logos Press.SCHROEDER, T. E. (1970). Neurulation in Xenopus laevis. An analysis and model based upon
light and electron microscopy. /. Embryol. exp. Morph. 23, 427-462.SCOTT, K., TARIN, D. & SHARP, J. A. (1970). Orientation of spherical specimens for ultrathin
sectioning in selected planes by embedding in agar. / . Microsc. 91, 217-220.SHARP, J. A. & TARIN, D. (1970). Time lapse cinematographic study of gastrulation and
neurulation. in the amphibian embryo (Xenopus laevis): a 16 mm. film. /. Anat. 107, 388.SPEMANN, H. (1931). Uber den Anteil von Implantat und Wirtskeim an der Orientierung und
Beschaffenheit der induzierten Embryonalanlage. Wilhelm Roux Arch. EntwMech. Org.123,390-517.
SPEMANN, H. (1938). Embryonic Development and Induction. Yale University Press.SPEMANN, H. & MANGOLD, H. (1924). Uber Induktion von Embryonalanlagen durch Implan-
tation artfremder Organisatoren. Arch, mikrosk. Anat. EntwMech. 100, 599-638.TAKAYA, H. (1955). Formation of the brain from the prospective spinal cord of amphibian
embryos. Proc. imp. Acad., Japan 31, 360-385.

570 D. TARINTARIN, D., SCOTT, K. & SHARP, J. A. (1970). Gastrulation and neurulation in the amphibian
embryo {Xenopus laevis): a 16 mm. film listed in the British National film catalogue andBritish Universities film catalogue and available for study.
TARIN, D. (1971a). Histochemical features of neural induction in Xenopus laevis (in prepara-tion).
TARIN, D. (19716). Ultrastructural features of neural induction in Xenopus laevis. J. Anat. (Inthe Press.)
TARIN, D. (1971C). A technique for ensuring complete separation of mesoderm from ecto-derm during neural induction in Xenopus embryos, based on vital staining of the mesodermprior to invagination (in preparation).
VAN GANSEN, P. & SCHRAM, A. (1969). Etude des ribosomes et du glycogene des gastrules deXenopus laevis par cytochimie ultrastructurale. / . Embryol. exp. Morph. 22, 69-98.
WADDINGTON, C. H. & DEUCHAR, E. (1952). The effect of type of contact with the organiseron the nature of the resulting induction. / . exp. Biol. 29, 496-512.
WADDINGTON, C. H. & PERRY, M. M. (1966). A note on the mechanisms of cell deformationin the neural folds of the amphibian. Expl Cell Res. 41, 691-693.
WADDINGTON, C. H. & SCHMIDT, G. A. (1933). Induction by heteroplastic grafts of theprimitive streak in birds. Wilhelm Roux Arch. EntwMech. Org. 128, 522-563.
WOERDEMAN, M. W. (1933). Uber den Glycogenstofswechsel des Organisationszentrums inder Amphibiengastrulu. Proc. K. ned. Akad. Wet. Ser. C. 36, 189-193.
WREN, J. T. & WESSELS, N. K. (1969). An ultrastructural study of lens invagination in themouse. /. exp. Zool. Ill, 359-369.
{Manuscript received 5 April 1971)





























