Embed Size (px)
Citation preview

Proc. Nati. Acad. Sci. USAVol. 82, pp. 8827-8831, December 1985Physiological Sciences
Kinetic properties of a sex pheromone-degrading enzyme:The sensillar esterase of Antheraea polyphemus
(insect/sensory physiolog/pheromone reception)
RICHARD G. VOGT*, LYNN M. RIDDIFORD*, AND GLENN D. PRESTWICHt*Department of Zoology, University of Washington, Seattle, WA 98195; and tDepartment of Chemistry, State University of New York at Stony Brook,Stony Brook, NY 11794
Communicated by L. M. Beidler, August 12, 1985
ABSTRACT Behavioral and electrophysiological evidencehas suggested that sex pheromone is rapidly inactivated withinthe sensory hairs soon after initiation of the action-potentialspike. We report the isolation and characterization of asex-pheromone-degrading enzyme from the sensory hairs ofthesilkmoth Antheraeapolyphemus. In the presence of this enzymeat physiological concentration, the pheromone [(6E,11Z)-hexadecadienyl acetate] has an estimated half-life of 15 msec.Our findings suggest a molecular model for pheromone recep-tion in which a previously reported pheromone-binding proteinacts as a pheromone carrier, and an enzyme acts as a rapidpheromone inactivator, maintaining a low stimulus noise levelwithin the sensory hairs.
A male moth seeks out a sexually receptive female by flyingin a zig-zag manner along her pheromone plume (1-3). Duringthis pheromone-oriented flight, the male displays rapid be-havioral responses (0.5-1.0 sec) to abrupt changes in theambient pheromone concentration (4, 5). For these behav-ioral responses to occur, rapid fluctuations of externalpheromone levels must be biochemically recreated in theinternal environment surrounding the sensory neurons.Thus, stimulus molecules that have already entered thesensory hair must be rapidly inactivated so that the neuronsexperience only incoming information.
Electrophysiological recordings of the response of a singlesensillum to pheromone (6, 7) show high-frequency spikefiring during stimulus and a rapid response decay with ahalf-life ofabout 1 sec when the stimulus is removed (8). Yet,when intact antennae of the silkmoth Bombyx mori wereexposed to [3H]bombykol, the half-life of undegraded pher-omone was 4 min (9). This apparent discrepancy betweenphysiological and molecular half-life led to a suggestion thatthere must be a nonenzymatic "early inactivation" processwithin the sensory hairs to account for the rapid physiologicalinactivation of the pheromone (6).The identification of a male-specific sensillar esterase in
the wild silkmoth Antheraea polyphemus (10, 11) suggestedthat an enzymatic process inside the sensillum itself could beresponsible for this early inactivation. Here we show that thepurified sensillar esterase from the receptor lymph surround-ing the dendrite degrades the A. polyphemus pheromone witha half-life of 15 msec, a rate compatible with the electrophys-iological and behavioral response times. We therefore sug-gest that rapid stimulus inactivation is enzymatic in nature,involving degradation of the pheromone.
MATERIALS AND METHODSAnimals. A. polyphemus pupae were purchased (H. W.
Hartman), stored at 4°C, and reared to adults at 25°C on an
18-hr-light/6-hr-dark cycle. Antennal tissues used for en-zyme purification were taken from adult males no more than3 days old.Enzyme Purification. Sensillar esterase was purified from
centrifuged (12,000 x g, 5 min) tissue homogenates bynondenaturing polyacrylamide slab gel electrophoresis (12,13) (14-cm gel width, 10% separation gel, 5% stacking gel,40C, 15 mA constant current/mm ofgel thickness). Gels werestained for esterase activity (14), and the sensillar esteraseband (10, 11) was removed and homogenized by passagethrough a series of 18- to 26-gauge syringe needles at 00C,with buffer added as needed (10 mM Tris Cl/0.02% sodiumazide, pH 7.2; final buffer volume equal to 10 times the gelvolume). Esterase was eluted from gel particles over 24 hr at4°C on a rotary mixer. Gel particles were removed bycentrifugation (20,000 x g, 1 hr, 4°C), and enzyme solutionwas stored on ice and used within 3 days or frozen (-70°C)and recovered later after lyophilization; such treatmentsresulted in no loss in activity.For pheromone kinetic studies, the tissue source was
isolated sensory hairs, collected by a technique modifiedfrom that described by Klein and Kiel (15, 16). Antennalbranches were mixed with equal volumes of 0.1- and 0.5-mm-diameter glass beads (B. Braun, VWR Scientific) in alyophilization flask. Chilling the flask in liquid nitrogenfollowed by violent shaking caused the glass beads to breakoff the brittle sensory hairs at their bases. The lyophilizedcontents were poured down a trough of Mylar film, to whichthe hairs adhered and from which the rest was shaken away.Hairs were scraped from the film with the edge of a cover slipand stored in sealed microcentrifuge tubes at -70°C. Onehundred-fifty antennae routinely yielded -20 mg of lyophil-ized sensory hairs. For the kinetic analysis, sensillar esterasewas purified from 15 mg of sensory hairs. After dialysis andconcentration (Amicon no. 4201 "Centricon" filters), werecovered 5 ml of enzyme solution, which was stored on icein microcentrifuge tubes. By comparing the activity of thissolution with the activity of crude sensory-hair homogenate(0.2 mg of hair/ml, 0.13 mg of hair/antennal unit) andassuming 100% efficiency in sensory-hair isolation, we esti-mated this stock of purified sensillar esterase to represent6.67 antennal units of activity/ml.For naphthyl acetate studies, 350 whole antennae (1.37 g)
were homogenized in 70 ml of 10 mM Tris Cl (pH 7.2) andconcentrated by vacuum dialysis (11). Gel elution yielded 52ml of enzyme solution, which we assumed to represent 6.7antennal units of activity/ml.
Preparation of Pheromone and Its Analogs. Labeled pher-omone (17) and diazoacetates (17, 18) were synthesized andstored in hexane at -70°C. When purity as assessed by TLCwas <98%, compounds were repurified by flash chromatog-raphy (19, 20). In all cases, aqueous working solutions were
Abbreviation: NapOAc, naphthyl acetate.
8827
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.
Dow
nloa
ded
by g
uest
on
June
2, 2
020
8828 Physiological Sciences: Vogt et al.
prepared from ethanol stocks immediately prior to an assayto minimize rapid loss due to glass adsorption.The tritiated pheromone (6E,11Z)-[11,12-3H]hexadeca-
dienyl acetate (58 Ci/mmol; 1 Ci = 37 GBq) (refs. 17 and 33;Fig. 2, compound 1) was prepared by drying an aliquot ofhexane solution under nitrogen and redissolving the phero-mone at 10 ,uM in ethanol. A 1:100 aqueous assay solutionwas prepared by mixing 10 ,.d of ethanol stock into 1 ml of 10mM Tris Cl buffer (pH 7.2) on a Vortex.Unlabeled pheromone [provided by K.-E. Kaissling
(M.P.I.V., 8131 Seewiesen, F.R.G.), from material synthe-sized by H. J. Bestmann (Universitit Erlangen Nurnberg,F.R.G.), at 0.59 mg/ml in hexane] was diluted in ethanol toa concentration of 2 mM, and this was diluted serially 1:3 inethanol. These ethanol stocks were prepared each day, and1:100 aqueous assay solutions were prepared within 5 min ofeach assay.Hexane stocks of nonradiolabeled diazoacetate analogs
(Fig. 2, compounds 2-4) were diluted to 20 mM in ethanol.Aqueous assay solutions (1:100) were prepared from theethanol stocks.
1-Naphthyl acetate (1-NapOAc) and 2-naphthyl acetate(2-NapOAc) (Sigma) were dissolved to 0.1 M in ethanol.Aqueous stocks (1:100) were prepared in 1,4-piperazine-bis(ethanesulfonate) (Pipes) buffer (100 mM, pH as indicat-ed).
Esterase inhibitors were supplied by T. C. Sparks (Loui-siana State University, Baton Rouge, LA) and B. D. Ham-mock (University of California, Davis) in ethanol solutions inconcentrations ranging from 1 to 100 mM. Aqueous stocks(1:50) were prepared in Pipes buffer (100 mM, pH 7.2).Enzyme Characterization: Pheromone Kinetics. Enzyme
solution (60 ,ul) was incubated with 60 ,ul of 0.3 ,uM[3H]pheromone plus 60 ,u1 of unlabeled pheromone at variousconcentrations. Incubations were in 10 x 75 mm borosilicatetubes at 23°C on a rotary mixer. Four incubations were runtogether: three with enzyme and the fourth with buffer(control). At 1-min intervals, pheromone was added to eachtube, and reactions were quenched at 5 min by addition of 180,l of ethyl acetate. On quenching, the reaction mixtures weremixed immediately on a Vortex for 10 sec. After phaseseparation, one ethyl acetate aliquot (3 p.l) was assayed forradioactivity, and two aliquots (3 ,u) were chromatographedon thin-layer plates (Macherey & Nagel, Polygram SILG/UV254, 4 x 8cm, coated plastic, Brinkman 661120-4). TLCplates were developed in ethyl acetate/hexane, 1:3 (vol/vol).Plates were pre-loaded with unlabeled pheromone [(6E,11Z)-hexadecadienyl acetate] and pheromone product [(6E,11Z)-hexadecadienol] and, after development, these were madevisible over iodine vapors (Rfs 0.61 and 0.33, respectively).Spots were cut out and associated radioactivity was mea-sured in Aquasol in a Beckman LS230 scintillation counter.The sensillar esterase concentration was adjusted to 0.1
antennal unit per incubation mixture, to produce about 30%conversion over 5 min (0.1 p.M pheromone). Control incu-bations showed <0.4% background radioactivity at productRf. Substrate concentrations were determined as the sum ofunlabeled plus labeled pheromone. Lowest-concentrationincubations (sO.1 p.M used only serially diluted [3H]phero-mone, with buffer replacing unlabeled material. Velocitieswere determined by multiplying the product/(product +substrate) ratios (from TLC) by the substrate concentration.In these experiments, 43 incubations were sampled in dupli-cate, over pheromone concentrations ranging from 7 pum to 2nM. Thirty-three of these incubations, with pheromoneconcentrations ranging upward from 0.1 ,uM, were used toderive the kinetic constants in Table 1.Enzyme Characterization: 2-NapOAc and 1-NapOAc Kinet-
ics. Kinetic constants were determined spectrophoto-metrically (Varian Superscan II). For each assay, 0.1 ml of
enzyme solution (0.67 antennal units of activity) was addedto 0.9 ml of substrate solution in a quartz cuvette. Prior tomixing, solutions were equilibrated at the incubation tem-perature. Velocities were taken from slopes oflines on a chartrecorder at 327 nm (2-naphthol) and 321.5 nm (1-naphthol).Dilutions (1:2) were made from aqueous substrate stocks,ranging from 18 ,uM to 18 nM for 2-NapOAc and from 90 p.Mto 5.6 p.M for 1-NapOAc. At least two measurements weremade at each concentration.
Inhibition Studies. Esterase inhibitors. The ability of var-ious esterase inhibitors to interfere with 2-NapOAc degra-dation was studied using the spectrophotometric assay.Esterase (0.1 ml, 0.67 antennal unit) was preincubated withinhibitor (0.8 ml) for 10 min, followed by addition of 2-NapOAc (0.1 ml, 180 p.M). Assays were performed at decadedilution concentrations for each inhibitor, with at least twomeasurements per concentration. IC50s were determined asthe inhibitor concentration at which the rate of 2-NapOAcdegradation was half-maximal.
Diazoacetates. Diazoacetate competition with 2-NapOAcwas examined using the spectrophotometric assay. Crudesensory-hair homogenate (0.1 ml, 0.33 mg of hair/ml) wasincubated with 2-NapOAc (0.8 ml) plus diazoacetate (0.1 ml).This hair homogenate was first characterized with 2-NapOAcsubstrate; an apparent Km of -2 x 10-6 M was determinedfor these conditions. The 2-NapOAc concentration (5 ,uM)was chosen based on this.
Diazoacetate competition with [3H]pheromone was exam-ined using the same protocol described above for the pher-omone kinetic studies. Hair homogenate (60 ,ul, 1 mg ofhair/ml) was 'preincubated 10 min with diazoacetate (60 ,pl),followed by addition of [3H]pheromone (60 p.1). The reactionwas quenched after 10 additional minutes.
RESULTS AND DISCUSSIONEnzyme Characterization. The sensillar esterase was ini-
tially identified in polyacrylamide gels by use of an enzymestain that utilized the substrates 1-NapOAc and 2-NapOAc(11). In the present studies, the esterase kinetics weredetermined by TLC, using the tritiated natural pheromone,and spectrophotometrically, using 1- and 2-NapOAc.
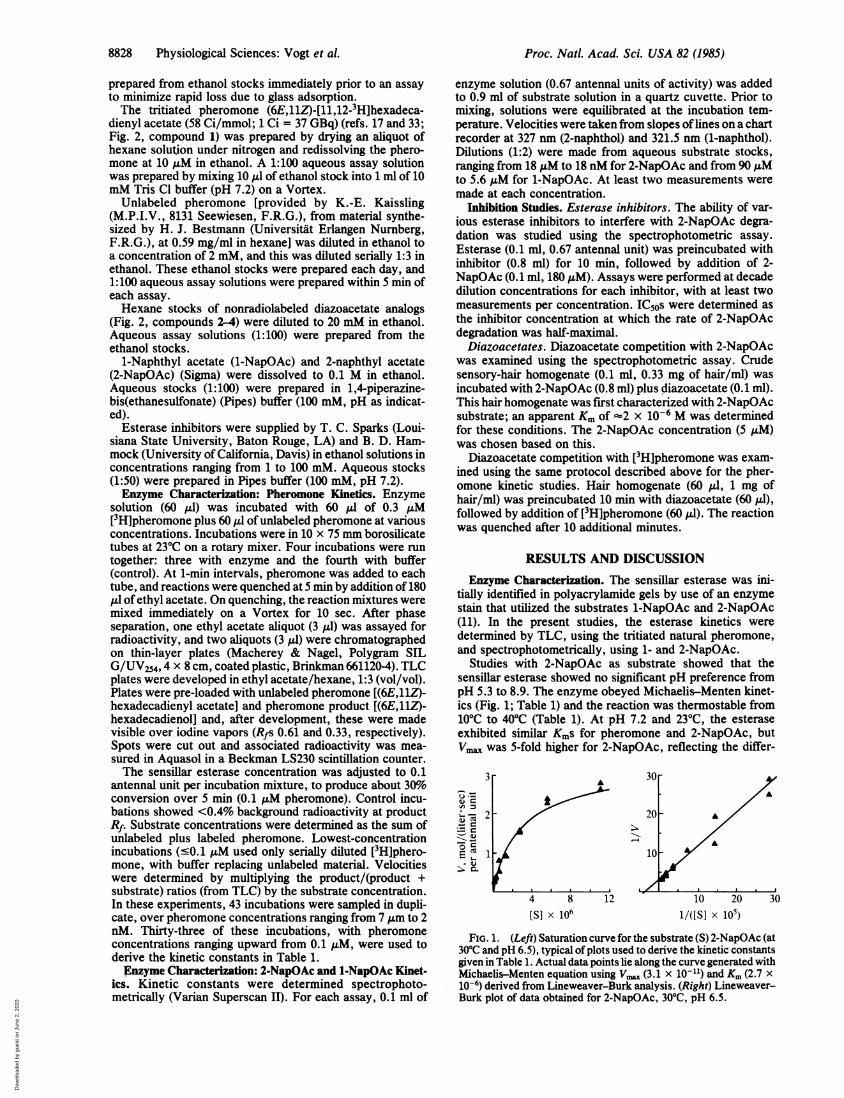
Studies with 2-NapOAc as substrate showed that thesensillar esterase showed no significant pH preference frompH 5.3 to 8.9. The enzyme obeyed Michaelis-Menten kinet-ics (Fig. 1; Table 1) and the reaction was thermostable from10°C to 40°C (Table 1). At pH 7.2 and 23°C, the esteraseexhibited similar Kms for pheromone and 2-NapOAc, butVmax was 5-fold higher for 2-NapOAc, reflecting the differ-
3- ~~A 30-
C~~~~~~~~~~~~~2 ~~~~~20- A
E 1Z 10-
4 8 12 10 20 30[S] x 106 1/([S] x 105)
FIG. 1. (Left) Saturation curve for the substrate (S) 2-NapOAc (at300C and pH 6.5), typical of plots used to derive the kinetic constantsgiven in Table 1. Actual data points lie along the curve generated withMichaelis-Menten equation using Vm.. (3.1 x 10-11) and Km (2.7 x10-6) derived from Lineweaver-Burk analysis. (Right) Lineweaver-Burk plot of data obtained for 2-NapOAc, 30'C, pH 6.5.
Proc. Natl. Acad. Sci. USA 82 (1985)
Dow
nloa
ded
by g
uest
on
June
2, 2
020
Proc. Natl. Acad. Sci. USA 82 (1985) 8829
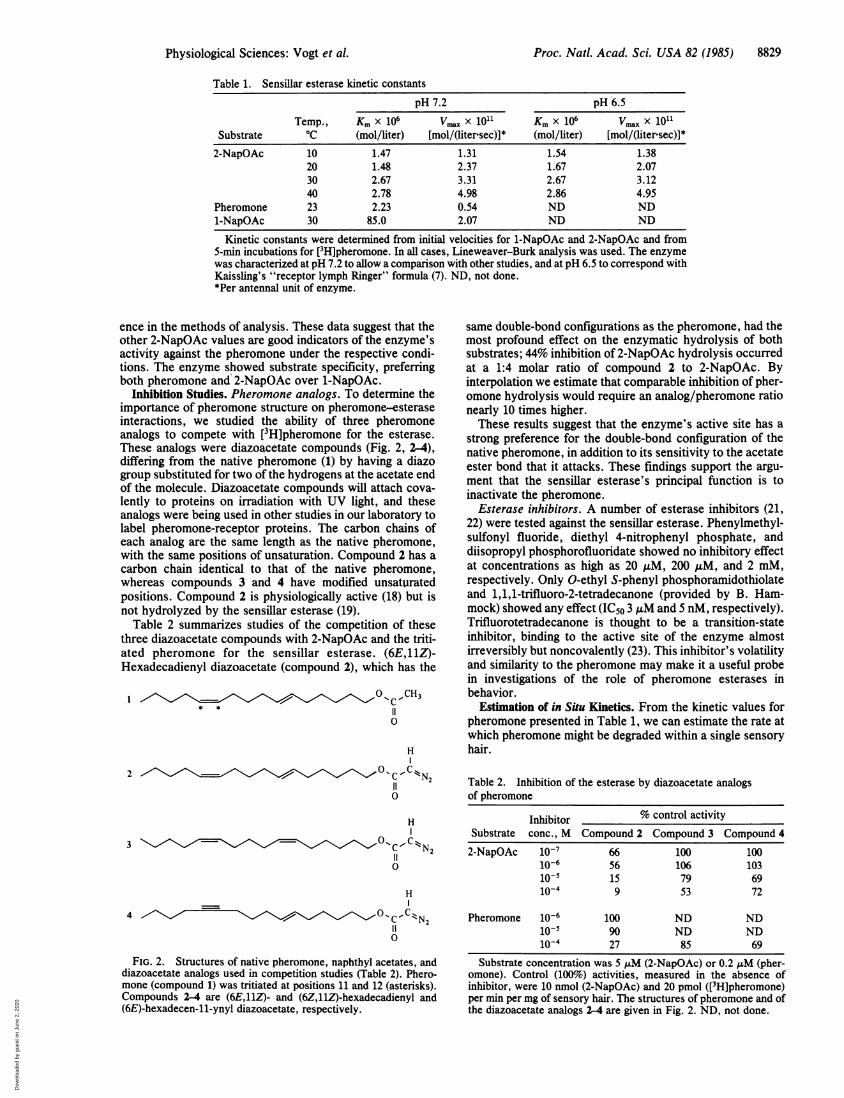
Table 1. Sensillar esterase kinetic constants
pH 7.2 pH 6.5
Temp., Km x 106 Vrax X 1011 Km x 106 Vmax X 1011Substrate 0C (mol/liter) [mol/(liter-sec)]* (mol/liter) [mol/(liter sec)]*2-NapOAc 10 1.47 1.31 1.54 1.38
20 1.48 2.37 1.67 2.0730 2.67 3.31 2.67 3.1240 2.78 4.98 2.86 4.95
Pheromone 23 2.23 0.54 ND ND1-NapOAc 30 85.0 2.07 ND ND
Kinetic constants were determined from initial velocities for 1-NapOAc and 2-NapOAc and from5-min incubations for [3H]pheromone. In all cases, Lineweaver-Burk analysis was used. The enzymewas characterized at pH 7.2 to allow a comparison with other studies, and at pH 6.5 to correspond withKaissling's "receptor lymph Ringer" formula (7). ND, not done.*Per antennal unit of enzyme.
ence in the methods of analysis. These data suggest that theother 2-NapOAc values are good indicators of the enzyme'sactivity against the pheromone under the respective condi-tions. The enzyme showed substrate specificity, preferringboth pheromone and 2-NapOAc over 1-NapOAc.
Inhibition Studies. Pheromone analogs. To determine theimportance of pheromone structure on pheromone-esteraseinteractions, we studied the ability of three pheromoneanalogs to compete with [3H]pheromone for the esterase.These analogs were diazoacetate compounds (Fig. 2, 2-4),differing from the native pheromone (1) by having a diazogroup substituted for two of the hydrogens at the acetate endof the molecule. Diazoacetate compounds will attach cova-lently to proteins on irradiation with UV light, and theseanalogs were being used in other studies in our laboratory tolabel pheromone-receptor proteins. The carbon chains ofeach analog are the same length as the native pheromone,with the same positions of unsaturation. Compound 2 has acarbon chain identical to that of the native pheromone,whereas compounds 3 and 4 have modified unsaturatedpositions. Compound 2 is physiologically active (18) but isnot hydrolyzed by the sensillar esterase (19).
Table 2 summarizes studies of the competition of thesethree diazoacetate compounds with 2-NapOAc and the triti-ated pheromone for the sensillar esterase. (6E,11Z)-Hexadecadienyl diazoacetate (compound 2), which has the
O CCH3* * 11
H
2 0, C-., N
H
II11
H
4 O1 .1-~ N
110
FIG. 2. Structures of native pheromone, naphthyl acetates, anddiazoacetate analogs used in competition studies (Table 2). Phero-mone (compound 1) was tritiated at positions 11 and 12 (asterisks).Compounds 2-4 are (6E,11Z)- and (6Z,11Z)-hexadecadienyl and(6E)-hexadecen-11-ynyl diazoacetate, respectively.
same double-bond configurations as the pheromone, had themost profound effect on the enzymatic hydrolysis of bothsubstrates; 44% inhibition of 2-NapOAc hydrolysis occurredat a 1:4 molar ratio of compound 2 to 2-NapOAc. Byinterpolation we estimate that comparable inhibition of pher-omone hydrolysis would require an analog/pheromone rationearly 10 times higher.These results suggest that the enzyme's active site has a
strong preference for the double-bond configuration of thenative pheromone, in addition to its sensitivity to the acetateester bond that it attacks. These findings support the argu-ment that the sensillar esterase's principal function is toinactivate the pheromone.
Esterase inhibitors. A number of esterase inhibitors (21,22) were tested against the sensillar esterase. Phenylmethyl-sulfonyl fluoride, diethyl 4-nitrophenyl phosphate, anddiisopropyl phosphorofluoridate showed no inhibitory effectat concentrations as high as 20 ,uM, 200 ,uM, and 2 mM,respectively. Only O-ethyl S-phenyl phosphoramidothiolateand 1,1,1-trifluoro-2-tetradecanone (provided by B. Ham-mock) showed any effect (IC50 3 ,M and 5 nM, respectively).Trifluorotetradecanone is thought to be a transition-stateinhibitor, binding to the active site of the enzyme almostirreversibly but noncovalently (23). This inhibitor's volatilityand similarity to the pheromone may make it a useful probein investigations of the role of pheromone esterases inbehavior.
Estimation of in Situ Kinetics. From the kinetic values forpheromone presented in Table 1, we can estimate the rate atwhich pheromone might be degraded within a single sensoryhair.
Table 2. Inhibition of the esterase by diazoacetate analogsof pheromone
Inhibitor % control activitySubstrate conc., M Compound 2 Compound 3 Compound 4
2-NapOAc 10-7 66 100 10010-6 56 106 10310-5 15 79 6910-4 9 53 72
Pheromone 10-6 100 ND ND10-5 90 ND ND10-4 27 85 69
Substrate concentration was S gM (2-NapOAc) or 0.2 ,uM (pher-omone). Control (100%) activities, measured in the absence ofinhibitor, were 10 nmol (2-NapOAc) and 20 pmol ([3H]pheromone)per min per mg of sensory hair. The structures of pheromone and ofthe diazoacetate analogs 2-4 are given in Fig. 2. ND, not done.
Physiological Sciences: Vogt et al.
Dow
nloa
ded
by g
uest
on
June
2, 2
020

8830 Physiological Sciences: Vogt et al.
Estimation ofpheromone concentrations. A single hair ofA. polyphemus is 300 um long (24), with a lumen diameter ofabout 2 Aum (25). Thus, the volume of the lumen is about 1 pl.A single pheromone molecule per hair, reported to besufficient to generate a spike (26), translates to an effectivepheromone concentration of about 1 pM. Physiologicalconcentrations extend upward from this value; 10 nM pher-omone corresponds to 104 molecules per hair lumen (Fig. 3,x axis).
Single-hair velocities. From the Km and Visa derived frompheromone degradation (Table 1), the Michaelis-Mentenequation {V = Vm,,,/(l + Km/[S])} can be used to estimate theactivity of the sensillar esterase at any substrate concentra-tion (Fig. 3, left axis). Using the pheromone Vm... (Table 1)divided by 55,000 sensory hairs per antenna (24), and mul-tiplying the calculated velocity by 6.02 x 1023 (Avogadro'snumber), we estimate that at the stimulus threshold concen-tration of 1 molecule per sensory hair (26) ([S] = 10-12 M), theenzyme content of a single hair can degrade pheromone at arate exceeding 27 molecules per second.Pheromone half-life determination. The half-life of pher-
omone in the presence of purified esterase is easily estimatedusing the equation t1/2 = (In 2)/(Vmax/Km) (27). This equationis valid only under conditions of first-order kinetics, wherethe pheromone concentration is well below the Km of theenzyme. Since the esterase's Km for pheromone (2.23 x 10-6M, Table 1) is 106 times higher than the physiologicalthreshold (26), we assume that the typical physiologicalsituation is one of first-order kinetics.By using a Vn, adjusted to a physiologically relevant value
based on the in situ enzyme concentration, we estimate thepheromone half-life within a sensory hair to be 15 msec. V..in Table 1 is reported for an antennal unit of enzyme in a
86661 10 3
20~~~~~
0
01~~~~~~~~~~~~~~~~~~~
-12 -11 -10 -9 -8 -7 -6 -5 -4(0) (1) (2) (3) (4) (5) (6) (7) (8)
Iog[pheromone]Iog(molecules per hair)
FIG. 3. Kinetics of sensillar esterase with the natural pheromone(230C, pH 7.2): the velocities at which a single hair's complement ofenzyme can degrade pheromone, and the expected half-life ofpheromone molecules in the presence of a physiological concentra-tion of enzyme. The abscissa represents the logarithms of both thepheromone concentration (mol/liter) and the corresponding numberof pheromone molecules per hair (see text). The left ordinaterepresents the logarithm of the velocity in molecules degraded perhair per second based on the Km and Vn, values for pheromone inTable 1 divided by 55,000 hairs per antenna (24). The right ordinaterepresents the logarithm of the pheromone half-life in msec. Thehalf-life plot was generated by digitally integrating the Michaelis-Menten equation over time, using the adjusted V., described in thetext. The triangles on the velocity curve represent pooled experi-mental data, with the number of assays per point indicated. In eachcase, the range of these data lay within the respective triangle.
1-liter volume. Therefore, we multiplied the pheromone Vm.[5.4 x 10-12 mol/(litersec), Table 1] by a dilution factor of11(5.5 x 10-) [(55,000 hairs per antenna) x (10-12 liter perhair) = 5.5 x 10-8 liter of receptor lymph per antenna)]. Thisyielded an adjusted Vm,, of 9.8 x 10-5 moles/(liter sec) at thephysiological enzyme concentration. Similar treatment ofthe2-NapOAc data (20'C, pH 7.2) suggests an even shorterhalf-life of 2.4 msec. These half-life values are biased on theslow side, because we assumed 100% efficiency in tissue andprotein isolation.The 15-msec degradative half-life holds for pheromone
concentrations ranging from 1 to 50,000 molecules per hair(Fig. 3), with the half-life increasing considerably at concen-trations >106 molecules per hair. The reported half-life ofthedecay in the electroantennogram response ofB. mori was onthe order of 1 sec at concentrations below 106 molecules perhair and much longer at higher concentrations (8). The shortenzymatic half-life compared with the longer electrophysio-logical half-life suggests that enzymatic degradation could beresponsible for rapid stimulus inactivation and that it is not arate-limiting step in the reception process. The increase in theelectrophysiological half-life occurred at a pheromone con-centration that would be approaching saturation for the A.polyphemus esterase, suggesting that enzymatic saturationmay have been responsible for this increase.
Intact antennae vs. sensory hair interior. There is anapparent contradiction between our proposed short phero-mone half-life within sensory hairs and the much longerhalf-life reported for pheromone applied to intact antennae.Previous studies using intact B. mori antennae suggested thatpheromone was degraded with a half-life of 4 min (9).Moreover, when between 1010 and 1012 molecules of [3H]-bombykol were applied to an antenna and the antenna wassubsequently washed first in pentane and then in chloroform/methanol, the pheromone appeared to pass from a pentane-extractable compartment into a chloroform/methanol-extractable compartment (28). This passage was interpretedas pheromone penetration toward the interior of the antenna,and occurred with a relatively long half-time of 1.6 min (28).Primarily undegraded pheromone was found in the pentane,whereas metabolites predominated in the chloroform/meth-anol (28). Thus, most of the pheromone seemed to bedegraded only after it had slowly penetrated the antenna.Similar findings have been published recently for A.polyphemus (29).
In light of our enzyme data, we feel these whole-antennaexperiments suggest that most of the pheromone moleculesthat contact a sensory hair never convey physiologicallyrelevant information. While a sufficient number of phero-mone molecules enter the hairs rapidly via the pore-tubulesto account for the rapid physiological responses, most of themolecules may enter the hair cuticle directly. The lattermolecules would then pass through the cuticle until theyslowly entered either the epithelial cells in the antennalbranch or the hair interior where they would become rapidlydegraded. This hypothesis would explain the comparabletime courses of both penetration and intact-antennal degra-dation (9, 28), as well as the disparity between the timecourses of the physiological response and intact-antennaldegradation (6).
Molecular Model for Pheromone Reception. We previouslyreported the presence of a soluble pheromone-binding pro-tein in the receptor lymph of A. polyphemus sensory hairs(10, 11). To be consistent with Kaissling's pheromone-reception model (6), we suggested that the binding proteinprovided a mechanism for Kaissling's "early inactivation"(6), and that the sensillar esterase might act to degradepheromone slowly after it was bound by the binding protein.The above kinetic data on the sensillar esterase requires
reevaluation of this model. The sensillar esterase can rapidly
Proc. Nati. Acad. Sci. USA 82 (1985)
Dow
nloa
ded
by g
uest
on
June
2, 2
020

Physiological Sciences: Vogt et al.
FIG. 4. In this model for pheromone reception, the sensillaresterase (SE) functions to inactivate pheromone (P) rapidly in orderto maintain a low stimulus-noise level within the sensory hair. Thepheromone binding protein (BP) functions in part as a carrier,transporting the pheromone across the receptor lymph space fromtubule to receptor. X-section, cross-section.
inactivate the pheromone. Therefore, the binding protein isnot necessary for pheromone inactivation but rather may
function as a carrier, transporting hydrophobic pheromonemolecules across the aqueous receptor lymph space frompore-tubule to dendrite (Fig. 4). In our studies, we routinelyobserve that pheromone solubility is enhanced in the pres-
ence of binding protein. In addition, binding protein concen-
trations >10 ,uM significantly interfered with pheromonedegradation by the esterase (30). These binding-proteinproperties are consistent with this protein having a role as a
pheromone carrier.The suggestion that the pheromone-binding protein acts as
a pheromone carrier runs counter to the notion that thepore-tubules act as a direct conduit for pheromone from hairsurface to membrane receptor via tubule-dendrite contacts.Although there are several reports of such contacts (25, 31),they were infrequently observed, evident in electron micro-graphs only when the dendrites were lying close to the innerhair wall (25). The appearance of tubule-dendrite contactsdepended on the histological fixation technique (31, 32), andthe observed contacts may have been mediated throughinteractions between surface coats of the tubules anddendritic membrane (25). If the contacts do exist in situ, it isnot known whether their function would be to providepheromone direct access to the dendritic membrane or to act
to stabilize the orientation of the tubules.If the binding protein does act as a carrier, how then does
the pheromone pass through the screen of apparently very
aggressive esterase, arriving at the membrane receptor in-tact? The answer may lie in the difference in concentrationbetween esterase and binding protein. The binding protein ispresent in concentrations on the order of 20 mM (11). Ourinability to stain the esterase with Coomassie blue afterelectrophoresis suggests that the enzyme is present at siO-times that concentration (unpublished data).The studies presented in this paper examined the rate at
which a purified enzyme degrades pheromone, under condi-tions where there were no competing pheromone-proteininteractions. Yet, within the sensory hair, there are at least
Proc. Natl. Acad. Sci. USA 82 (1985) 8831
two other pheromone-specific proteins, the binding proteinand the membrane receptors. The dynamic interactionsbetween the pheromone and these proteins translate theexternal world of stimulus fluctuations to the sensory neuron.We suggest that the kinetics of these biochemical interactionsare central in determining how efficiently a male moth canfunction behaviorally as he tracks pheromone plumes to asexually receptive female.
We thank Drs. B. D. Hammock, T. Sparks, and K.-E. Kaisslingfor generously supplying esterase inhibitors and unlabeled phero-mone and Drs. J. W. Truman, B. D. Hammock, and T. Hines formost helpful discussions and suggestions. R.G.V. was supported byNational Research Service Awards GM07270 and GM07108. Re-search was also supported by grants from the National ScienceFoundation [PCM-80-11152 (L.M.R.) and PCM-83-16931 (G.D.P.)],the Alfred P. Sloan Foundation (G.D.P.) and the Camille and HenryDreyfus Foundation (G.D.P.).
1. Kennedy, J. S. & Marsh, D. (1974) Science 184, 999-1001.2. Kennedy, J. S. (1978) Physiol. Entomol. 3, 91-98.3. David, C. T., Kennedy, J. S. & Ludlow, A. R. (1983) Nature
(London) 303, 804-806.4. Kramer, E. (1975) in Olfaction and Taste, eds. Denton, D. A. &
Coghlan, J. P. (Academic, New York), Vol. 5, pp. 329-335.5. Marsh, D., Kennedy, J. S. & Ludlow, A. R. (1981) Physiol.
Entomol. 6, 225.6. Kaissling, K.-E. (1974) in Biochemistry of Sensory Function, ed.
Jaenicke, L. (Springer-Verlag, Heidelberg), pp. 243-273.7. Kaissling, K.-E. & Thorson, J. (1980) in Receptors for
Neurotransmitters, Hormones and Pheromones in Insects, eds.Satelle, D. & Hall, L. (Elsevier, New York), pp. 261-282.
8. Kaissling, K.-E. (1977) in Chemical Control of Insect Behavior-Theory and Application, eds. Shorey, H. H. & Mckelvey, J. J.(Wiley, New York), pp. 45-65.
9. Kasang, G. & Kaissling, K.-E. (1972) in International Symposia onOlfaction and Taste, ed. Schneider, D. (Verlagsgesellschaft, Stutt-gart, F.R.G.), Vol. 4, pp. 200-206.
10. Vogt, R. G. & Riddiford, L. M. (1981a) in Regulation of InsectDevelopment and Behaviour, eds. Sehnal, F., Zabza, A., Menn,J. J. & Cymborowski, B. (Polytech. Univ. of Wroclaw Press,Wroclaw, Poland), pp. 955-967.
11. Vogt, R. G. & Riddiford, L. M. (1981b) Nature (London) 293,161-163.
12. Laemmli, U. K. (1970) Nature (London) 227, 680-685.13. Ames, G. F.-L. (1974) J. Biol. Chem. 249, 634-644.14. Shaw, C. R. & Prasad, R. (1970) Biochem. Genet. 4, 297-320.15. Klein, U. & Keil, T. A. (1984) Cell. Molec. Neurobiol. 4, 385-3%.16. Vogt, R. G. (1984) Dissertation (Univ. of Washington, Seattle).17. Prestwich, G. D., Golec, F. A., Jr., & Andersen, N. H. (1984) J.
Lab. Comp. Radiopharm. 21, 593-601.18. Ganjian, I., Pettei, M. J., Nakanishi, K. & Kaissling, K.-E. (1978)
Nature (London) 271, 157-158.19. Prestwich, G. D., Vogt, R. G. & Riddiford, L. M. (1986) J. Chem.
Ecol. 12(2), in press.20. Still, W. C., Kahn, M. & Mitra, A. (1978) J. Org. Chem. 43,
2923-2925.21. Dixon, M. & Webb, E. C. (1964) Enzymes (Academic, New York),
pp. 346-351.22. Sparks, T. C., Hammock, B. D. & Riddiford, L. M. (1983) Insect
Biochem. 13, 529-541.23. Hammock, B. D., Wing, K. D., McGlaughlin, J., Lovell, V. M. &
Sparks, T. C. (1982) Pestic. Biochem. Physiol. 17, 76-88.24. Boeckh, J., Kaissling, K.-E. & Schneider, D. (1960) Zool. Jahrb.
Abt. Anat. Ontog. Tiere 78, 559-584.25. Keil, T. A. (1984) Tissue Cell 16, 705-717.26. Schneider, D., Kasang, G. & Kaissling, K.-E. (1968) Naturwis-
senschaften 55, 395-3%.27. Segel, I. H. (1975) Enzyme Kinetics (Wiley, New York), p. 41.28. Kasang, G. (1974) J. Insect Physiol. 20, 2407-2422.29. Kanaujia, S. & Kaissling, K.-E. (1985) J. Insect Physiol. 31, 71-81.30. Vogt, R. G. & Riddiford, L. M. (1986) in Mechanisms of Percep-
tion and Orientation to Insect Olfactory Signals, eds. Payne, T.,Carde, R. & Boeckh, J. (Oxford Univ. Press, Oxford), in press.
31. Steinbrecht, R. A. & Muller, B. (1971) Z. Zellforsch. Mikrosk.Anat. 117, 570-575.
32. Steinbrecht, R. A. (1980) Tissue Cell 12, 73-100.33. Kochansky, J., Tette, J., Taschenberg, E. F., Carde, R. T., Kais-
sling, K.-E. & Roelofs, W. L. (1975) J. Insect Physiol. 21,1977-1983.
X-Section of anA _ ~~~~Olfactorya....**. *9 H ai r
I -:*B P =binding protin:.*SE = sensilla estera se )
~0 0 .0000
C 000000 0
n JI000P *
nactive
Dow
nloa
ded
by g
uest
on
June
2, 2
020























![[ pheromone ] 01](https://img.pdfslide.us/doc/110x75/568caab71a28ab186da2ad9b/-pheromone-01.jpg)





